NAT10 promotes radiotherapy resistance in non-small cell lung cancer by regulating KPNB1-mediated PD-L1 nuclear translocation
-
Dagao Zhu
 ,
Mingliang Lu
,
Mingliang Lu

Abstract
Radiotherapy (RT) resistance in non-small cell lung cancer (NSCLC) is a significant contributor to tumor recurrence. NAT10, an enzyme that catalyzes ac4C RNA modification, has an unclear role in RT resistance. This study aimed to explore the function of NAT10 in RT resistance in NSCLC. RT-resistant NSCLC cell lines (PC9R and A549R) were established through repeated irradiation. The impact of NAT10 on cellular immunity was evaluated by measuring immune cell populations, cytotoxicity levels, and markers of cell dysfunction. Results demonstrated elevated levels of ac4C and NAT10 in RT-resistant cells. Knockdown of NAT10 suppressed cell proliferation and enhanced immune function in PC9R and A549R cells by upregulating TNF-α and IFN-γ while downregulating PD-1 and TIM-3. Mechanistically, RT resistance in NSCLC was mediated by NAT10-dependent ac4C modification of KPNB1. Furthermore, KPNB1 facilitated PD-L1 nuclear translocation, promoting immune escape in RT-resistant NSCLC cells. Overexpression of KPNB1 enhanced cell proliferation but impaired immune function in RT-resistant NSCLC cells. In conclusion, this study demonstrates that NAT10 upregulates KPNB1 expression through ac4C modification, thereby promoting RT resistance in NSCLC via PD-L1 nuclear translocation. These findings reveal a novel mechanism underlying RT resistance in NSCLC.
Graphical abstract
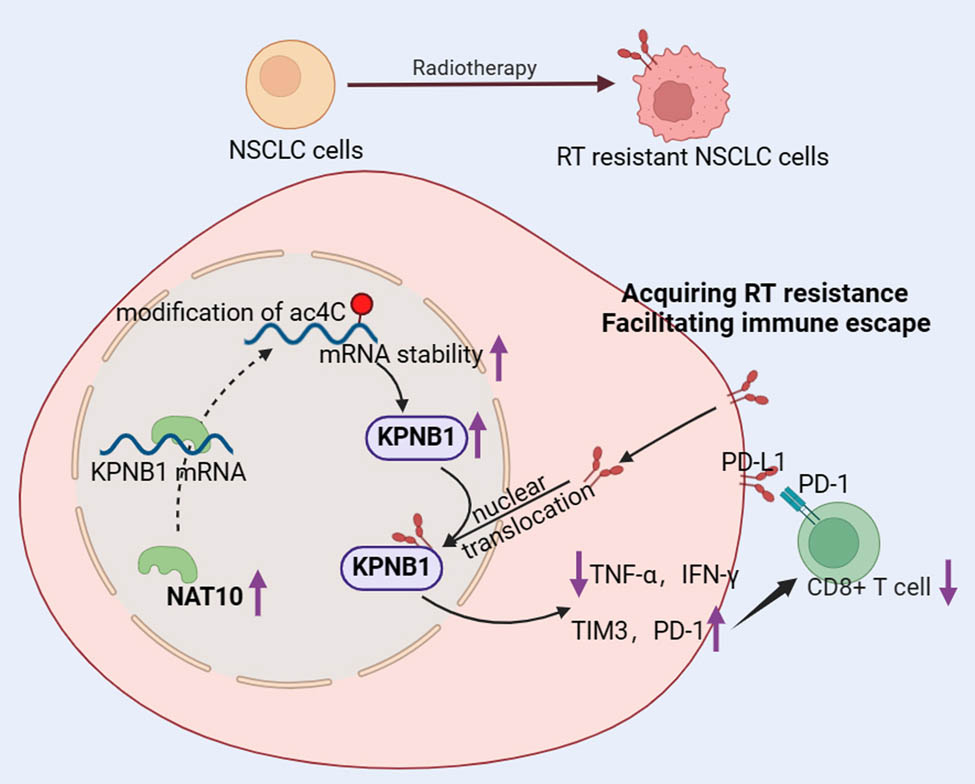
1 Introduction
Non-small cell lung cancer (NSCLC) constitutes approximately 80–85% of all lung cancer cases and remains one of the leading causes of cancer-related mortality worldwide. Based on histopathological characteristics, NSCLC is primarily classified into two major subtypes: lung adenocarcinoma (LUAD) and lung squamous cell carcinoma (LUSC) [1]. Current therapeutic strategies for NSCLC include surgery, chemotherapy, radiotherapy (RT), targeted therapy, immunotherapy, or a combination of these modalities, which have collectively contributed to improved survival rates among patients [2–4]. Among these, RT serves as a cornerstone treatment for NSCLC. Its mechanism of action involves inducing lethal DNA damage either directly in irradiated cells or indirectly through the generation of reactive oxygen species [5]. Notably, approximately 77% of NSCLC patients have evidence-based indications for RT, and this treatment has been demonstrated to significantly enhance clinical outcomes and overall survival rates [6,7]. Despite these advancements, a subset of patients experiences localized tumor recurrence following standard clinical doses of radiation, which may be attributed to the development of RT resistance in tumor cells [8]. This resistance poses a significant challenge to the efficacy of RT and underscores the need to elucidate its underlying mechanisms. Understanding the molecular basis of RT resistance is crucial for developing strategies to overcome this limitation, thereby improving therapeutic outcomes and prolonging survival in NSCLC patients.
RT resistance represents a major contributor to tumor treatment failure and metastasis [9]. The mechanisms underlying RT resistance are highly complex and remain incompletely understood. Current evidence suggests that RT resistance is influenced by a multitude of factors, including genetic alterations, the tumor microenvironment, and metabolic reprogramming such as aerobic glycolysis [10,11]. In NSCLC specifically, RT resistance has been linked to epigenetic modifications, dysregulation of non-coding RNAs, and imbalances in ferroptosis [12–14]. Notably, emerging research highlights the critical role of immune dysfunction in mediating RT resistance. For instance, RT resistance has been shown to impair the expansion of tumor-antigen-specific T cells and suppress the activation of immune priming mechanisms [15]. Additionally, RT resistance promotes the upregulation of immune checkpoint molecules, such as CD47 and PD-L1, which facilitate tumor metastasis and immune evasion [16]. Furthermore, studies have demonstrated that immunosuppressive effects, including PD-L1 upregulation and enhanced glucose metabolism in neutrophils, play pivotal roles in driving RT resistance in NSCLC [17,18]. Despite these advances, the mechanisms of RT resistance in NSCLC remain poorly characterized, and there is an urgent need to identify novel therapeutic targets to sensitize tumors to radiation and overcome resistance.
NAT10, a crucial member of the Gcn5-related N-acetyltransferase family, is the only known “writer” protein responsible for ac4C RNA modification, catalyzing the addition of ac4C to mRNA [19]. This NAT10-mediated ac4C modification plays a pivotal role in enhancing mRNA stability and increasing translation efficiency. Through its acetyltransferase activity, NAT10 regulates various cellular processes, including apoptosis, autophagy, and cell proliferation [20]. Accumulating evidence has highlighted the significance of NAT10 in cancer biology, where it promotes tumor initiation, metastasis, and progression by catalyzing ac4C modifications on mRNA, thereby enhancing translational efficiency and driving the expression of oncogenic genes [21–23]. In NSCLC specifically, NAT10 has been shown to be upregulated and is associated with poor patient prognosis [24,25]. Despite these findings, the role of NAT10 in mediating RT resistance in NSCLC remains poorly understood and warrants further investigation.
Despite these findings, the role of NAT10 in mediating RT resistance in NSCLC remains poorly understood and warrants further investigation. Elucidating the mechanisms by which NAT10 contributes to RT resistance could provide critical insights into overcoming therapeutic challenges and improving outcomes for NSCLC patients.
2 Methods
2.1 Cell culture and irradiation treatment
The LUAD cell line PC9 and the human NSCLC cell line A549 were obtained from the American Type Culture Collection (Manassas, VA, USA). Cells were cultured in RPMI-1640 medium (Gibco, Grand Island, NY, USA) supplemented with 10% FBS (Gibco) and 1% penicillin/streptomycin (Gibco) at 37°C in a humidified incubator with 5% CO₂. Radiation-resistant cell lines were generated following a previously described method [26]. Briefly, PC9 and A549 cells were seeded into 60 mm dishes and irradiated using a 137Cs source when they reached approximately 60% confluency. Cells were exposed to 2 Gy of radiation once daily until a cumulative dose of 100 Gy was achieved. The resulting radiation-resistant cell lines were designated as PC9R and A549R. To assess radiation resistance, a subset of PC9R and A549R cells was further irradiated with 6 Gy and incubated for 72 h. This dose was administrated to parental NSCLC cells, and results confirmed that 6 Gy irradiation significantly inhibited cell viability and proliferation while promoting apoptosis in parental PC9 and A549 cells (Figure S1a–e).
2.2 Cell transfection
Short hairpin RNAs targeting NAT10 (shNAT10) and shKPNB1, along with a negative control shRNA (shNC), were purchased from GenePharma (Shanghai, China). Additionally, KPNB1 overexpression plasmids (pcDNA3.1-KPNB1) and the corresponding empty vector control (pcDNA3.1-vector) were obtained from the same supplier. Transfection was performed using Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s protocol. Briefly, A549R and PC9R cells in the logarithmic growth phase were transfected, and cells were harvested 48 h post-transfection for subsequent analysis.
2.3 Cell viability assay
Cell viability was assessed using a Cell Counting Kit-8 (CCK-8; Yeasen, Shanghai, China) according to the manufacturer’s instructions. Briefly, cells were seeded into 96-well plates at an appropriate density and cultured under standard conditions. After the designated treatment, 10 µL of CCK-8 solution was added to each well, and the plates were incubated at 37°C for 2 h. The absorbance of each well was measured at 450 nm using a microplate reader (BioTek, Winooski, VT, USA).
2.4 Colony formation assay
The colony formation assay was performed to assess cell proliferation capacity. Cells were seeded into six-well plates at a density of 500 cells per well and cultured at 37°C in a humidified incubator with 5% CO₂. After 14 days of incubation, cells were fixed with 4% paraformaldehyde for 15 min and then stained with 0.2% crystal violet solution for 30 min at room temperature. Colonies containing more than 50 cells were counted manually under a microscope.
2.5 Detection of apoptosis
Apoptosis in PC9R and A549R cells was assessed using an Annexin V-FITC/PI Apoptosis Detection Kit (Beyotime, Shanghai, China) according to the manufacturer’s instructions. Briefly, cells were harvested, washed twice with PBS, and centrifuged at 1,000g for 5 min to remove the supernatant. The cell pellet was resuspended in Annexin V-FITC binding buffer and stained with Annexin V-FITC and PI for 20 min at room temperature in the dark. Apoptotic cells were analyzed using a flow cytometer (BD Biosciences, San Jose, CA, USA). The percentages of early apoptotic (Annexin V-FITC-positive/PI-negative) and late apoptotic (Annexin V-FITC-positive/PI-positive) cells were quantified.
2.6 Dot blot assay
The levels of ac4C modification in PC9 and A549 cells were measured using a dot blot assay. Total RNA was extracted from PC9 and A549 cells using Trizol reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions. The isolated RNA was denatured by heating at 65°C for 5 min and then spotted onto a Hybond-N + membrane (GE Healthcare, Chicago, IL, USA). The RNA was immobilized on the membrane by UV cross-linking for 5 min. Subsequently, the membrane was blocked with 5% skim milk in Tris-buffered saline with Tween-20 for 1 h at room temperature to prevent nonspecific binding. The membrane was then incubated with a primary anti-ac4C antibody overnight at 4°C, followed by incubation with a horseradish peroxidase (HRP)-conjugated secondary antibody for 1 h at room temperature. Finally, the signal was detected using an enhanced chemiluminescence (ECL) kit (Thermo Scientific, Waltham, MA, USA) and visualized using a chemiluminescence imaging system.
2.7 Quantitative real-time PCR (qPCR)
RNA isolation was conducted using Trizol reagent and reverse transcribed into cDNA using a HiScript II first strand cDNA synthesis kit (Vazyme, Nanjing, China). qPCR was performed using SYBR green reagent and the mixture was prepared followed by the manufacturer’s protocol. The relative mRNA expression was calculated using the 2−∆∆Ct method with GAPDH serving as the internal reference. The qPCR primers used in this study were as follows: NAT10, 5′-ATAGCAGCCACAAACATTCGC-3′(forward) and 5′-ACACACATGCCGAAGGTATTG-3′ (reverse); TNF-α, 5′-CCTCTCTCTAATCAGCCCTCTG-3′(forward) and 5′-GAGGACCTGGGAGTAGATGAG-3′(reverse); IFN-γ, 5′-TCGGTAACTGACTTGAATGTCCA-3′(forward) and 5′-TCGCTTCCCTGTTTTAGCTGC-3′(reverse); PD-1, 5′-CCAGGATGGTTCTTAGACTCCC-3′(forward) and 5′-TTTAGCACGAAGCTCTCCGAT-3′(reverse); TIM3, 5′-CTGCTGCTACTACTTACAAGGTC-3′(forward) and 5′-GCAGGGCAGATAGGCATTCT-3′(reverse); KPNB1, 5′-CCACTTTCCTTGTGGAACTGT-3′(forward) and 5′-CTCTGCTGATATTGTGCCTTGA-3′ (reverse); NCL, 5′-GGTGGTCGTTTCCCCAACAAA-3′(forward) and 5′-GCCAGGTGTGGTAACTGCT-3′ (reverse).
2.8 Immunofluorescence (IF) staining
IF staining was performed using an IF staining kit (Beyotime, Shanghai, China) according to the manufacturer’s protocol. Briefly, cells were fixed with 4% paraformaldehyde (included in the kit) overnight at 4°C. After fixation, cells were washed three times with PBS containing 0.1% Tween-20 (PBST) to remove residual fixative. Cells were then blocked with blocking buffer (provided in the kit) for 1 h at room temperature to prevent nonspecific binding. Subsequently, cells were incubated overnight at 4°C with primary antibodies against NAT10, KPNB1, or PD-L1. After washing with PBST, cells were incubated with fluorescence-conjugated secondary antibodies for 1 h at room temperature in the dark. Finally, cells were counterstained with 4′,6-diamidino-2-phenylindole to visualize nuclei and imaged using a fluorescence microscope (Nikon, Tokyo, Japan).
2.9 Co-culture of T cells and tumor cells
Peripheral blood mononuclear cells (PBMCs) were obtained from BLUEFBIO (Shanghai, China). T cells were isolated using the Dynabeads® Untouched™ Human T Cells Kit (Thermo Scientific, Waltham, MA, USA) according to the manufacturer’s instructions. The co-culture of T cells and tumor cells was performed as previously described [27]. Briefly, PBMCs were seeded into six-well plates at a density of 2 × 106 cells per well and stimulated with cell lysates derived from PC9R or A549R cells, along with anti-CD3ε and anti-CD28 antibodies, to activate T cells. Activated T cells were then co-cultured with PC9R or A549R cells at a 10:1 ratio (T cells:tumor cells) for 16 h. Cells were incubated with PE-conjugated anti-CD4 and APC-conjugated anti-CD8 antibodies for 30 min at 4°C. The proportions of CD4+ and CD8+ T cells were analyzed by flow cytometry. Additionally, the expression levels of TNF-α, IFN-γ, PD-1, and TIM-3 in the co-culture system were quantified using qPCR.
2.10 Bioinformatic analysis
Genes associated with NAT10 in LUAD and LUSC were screened using the LinkedOmics database (https://www.linkedomics.org/). Genes meeting the criteria of p < 0.05 and |r| > 0.4 (r > 0.4 or r < −0.4) in both LUAD and LUSC were selected and intersected. Pathway enrichment analysis was subsequently performed on these overlapping genes using FunRich software. Additionally, the correlations between KPNB1 and NAT10, as well as between NCL and NAT10, were analyzed using the LinkedOmics database.
2.11 Methylated RNA immunoprecipitation (MeRIP)
The ac4C modification level on KPNB1 in PC9R and A549R cells was assessed using a GenSeq ac4C RIP kit (Cloudseq, Shanghai, China) according to the manufacturer’s instructions. Briefly, RNA was fragmented using a fragmentation buffer on a PCR instrument, followed by incubation at 70°C for 6 min. Protein G magnetic (PGM) beads were pre-incubated with anti-ac4C antibody for 1 h at room temperature. The fragmented RNA was then incubated with the antibody-coated PGM beads for 1 h on a rotator at 4°C. After incubation, the beads were washed and purified following the manufacturer’s protocol. The ac4C modification level of KPNB1 was subsequently quantified by qPCR.
2.12 RIP
The interaction between NAT10 and KPNB1 was investigated using an RIP assay kit (Beyotime, China). Briefly, protein A/G agarose beads were incubated with anti-KPNB1 antibody or anti-IgG (negative control) for 30 min at room temperature. Cells were lysed using a specific lysis buffer for 15 min on ice, followed by centrifugation at 14,000g for 10 min at 4°C to collect the supernatant. The antibody-coated protein A/G agarose beads were then incubated with the supernatant for 4 h at 4°C on a shaker. After incubation, the samples were eluted and purified according to the manufacturer’s protocol. The expression of KPNB1 was subsequently quantified by qPCR.
2.13 RNA stability assay
To assess the stability of KPNB1 mRNA in PC9R and A549R cells, cells were treated with 5 μg/mL actinomycin D (Merck, Darmstadt, Germany) to inhibit transcription. The expression levels of KPNB1 were measured by qPCR at baseline (0 h) and at 4, 8, and 12 h post-treatment.
2.14 Dual luciferase reports
Wild-type (wt) and mutant (mut) sequences containing potential KPNB1 binding sites were cloned into the pGL3 vector to construct luciferase reporter plasmids. Transfection was performed in PC9R and A549R cells when they reached approximately 80% confluency, using Lipofectamine 2000 reagent (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s instructions. After 48 h of transfection, luciferase activity was measured using a dual-luciferase reporter assay system (Promega, San Luis Obispo, CA, USA).
2.15 Co-immunoprecipitation (co-IP)
The interaction between KPNB1 and PD-L1 was examined using a Pierce Classic Magnetic IP/Co-IP kit (Thermo Scientific, USA). Cells were lysed with IP lysis buffer on ice for 5 min, followed by centrifugation to collect the supernatant. The lysate was incubated overnight at 4°C with anti-KPNB1, anti-PD-L1, or anti-IgG (negative control) antibodies. Subsequently, the samples were incubated with protein A/G magnetic beads for 1 h at room temperature. After washing, the immunoprecipitated complexes were eluted according to the manufacturer’s protocol. The protein levels of KPNB1 and PD-L1 were analyzed by western blot.
2.16 Western blot
Total protein was extracted from PC9R and A549R cells using RIPA lysis buffer (Beyotime, China). Nuclear protein isolation was performed using a nuclear protein extraction kit (Beyotime, China) according to the manufacturer’s instructions. Protein samples were separated on 10% SDS-PAGE gels and transferred to PVDF membranes. The membranes were blocked with 5% skimmed milk for 1 h at room temperature and then incubated with primary antibodies overnight at 4°C. The primary antibodies used in this study included anti-KPNB1 (1:1,000, ab313370; Abcam, Cambridge, MA, USA), anti-PD-L1 (1:1,000, ab213524; Abcam), anti-GAPDH (1:10,000, ab181602; Abcam), and anti-Lamin B1 (1:10,000, ab133741; Abcam). After incubation, the membranes were washed and incubated with HRP-conjugated secondary antibodies (1:10,000, ab6721; Abcam) for 2 h at room temperature. Protein bands were visualized using an ECL reagent.
2.17 Statistical analysis
All experiments were independently repeated at least three times, and the data are presented as mean ± SD. Statistical analyses were performed using GraphPad Prism 7.0 software (GraphPad Software, San Diego, CA, USA). Differences between two groups were assessed using an unpaired Student’s t-test, while comparisons among multiple groups were analyzed by one-way analysis of variance. A p-value of less than 0.05 (p < 0.05) was considered statistically significant.
3 Results
3.1 RT leads to therapeutic resistance in NSCLC cells
To determine whether RT contributes to therapeutic resistance in NSCLC cells, radiation-resistant PC9R and A549R cells were subjected to additional irradiation. Therapeutic resistance was assessed by evaluating cell viability, proliferation, and apoptosis. The results demonstrated no statistically significant differences in cell viability (Figure 1a), proliferation (Figure 1b and d), or apoptosis (Figure 1c and e) between RT-treated and untreated PC9R and A549R cells. These findings suggest that RT induces therapeutic resistance in NSCLC cells.

RT led to therapeutic resistance in NSCLC cells. (a) Cell viability of PC9R and A549R cells was detected using a CCK8 kit. (b) and (d) Cell proliferation ability of PC9R and A549R cells was evaluated by colony formation assay. (c) and (e) Cell apoptosis of PC9R and A549R cells was detected by flow cytometry.
3.2 ac4C modification and NAT10 expression are enhanced in radiation-resistant NSCLC cells
To explore the role of ac4C modification in RT resistance in NSCLC, we compared the levels of ac4C modification and NAT10 expression between parental (PC9 and A549) and radiation-resistant (PC9R and A549R) cells. The results revealed a significant increase in both ac4C modification levels and NAT10 expression in PC9R and A549R cells compared to their parental counterparts (Figure 2a and b). Furthermore, IF staining demonstrated enhanced fluorescence intensity of NAT10 in PC9R and A549R cells (Figure 2c). These findings indicate that ac4C modification and NAT10 expression are upregulated in radiation-resistant NSCLC cells.

ac4C modification and NAT10 expression were enhanced in radiation-resistant NSCLC cells. (a) ac4C levels in PC9 and A549 cells were detected by dot blot assay. (b) NAT10 expression in PC9 and A549 cells was measured by qPCR. (c) IF staining showed the expression of NAT10 in PC9R and A549R cells.
3.3 NAT10 knockdown improves RT resistance in radiation-resistant NSCLC cells
To investigate the functional role of NAT10 in RT resistance in NSCLC cells, rescue experiments were conducted. NAT10 expression in radiation-resistant NSCLC cells was significantly reduced following transfection with shNAT10 (Figure 3a). NAT10 knockdown markedly inhibited the proliferation of PC9R and A549R cells (Figure 3b) and promoted apoptosis in these cells (Figure 3c). Flow cytometry analysis revealed that NAT10 knockdown significantly increased the proportion of CD8+ T cells but had no effect on CD4+ T cell populations, suggesting that the attenuation of RT resistance by NAT10 knockdown is mediated through CD8+ T cells (Figure 3c). Furthermore, NAT10 knockdown elevated the levels of TNF-α and IFN-γ (Figure 3d and e) while reducing the expression of PD-1 and TIM-3 in the co-culture system (Figure 3f and g). Similar effects were observed in parental PC9 and A549 cells, where NAT10 knockdown increased CD8+ T cell populations and levels of TNF-α and IFN-γ but decreased PD-1 and TIM-3 expression (Figure S2a–c). Collectively, these findings demonstrate that NAT10 knockdown suppresses cell proliferation, enhances immune function, and attenuates RT resistance in radiation-resistant NSCLC cells.

NAT10 knockdown improved RT resistance in radiation-resistant NSCLC cells. (a) qPCR was performed to measure the expression of NAT10 in PC9R and A549R cells. (b) Cell proliferation ability of PC9R and A549R cells was evaluated by colony formation assay. (c) Apoptosis of PC9R and A549R cells was evaluated by flow cytometry. (d) The amount of CD4+ and CD8+ T cells was detected by flow cytometry. (e)–(h) The levels of TNF-α, IFN-γ, PD-1, and TIM3 were measured by qPCR.
3.4 NAT10 knockdown inhibits KPNB1 expression by decreasing ac4C modification on KPNB1 in radiation-resistant NSCLC cells
To elucidate the mechanism by which NAT10 mediates RT resistance in NSCLC cells, we analyzed genes associated with NAT10 in LUAD and LUSC using the LinkedOmics database (Figure 4a). Pathway enrichment analysis of the intersecting genes from LUAD and LUSC revealed the IFN-γ pathway as the most significantly enriched pathway (Figure 4b). KPNB1 and NCL, both enriched in the IFN-γ pathway, showed a positive correlation with NAT10 in LUAD and LUSC (Figure 4c and d). To determine which gene interacts with NAT10 in radiation-resistant NSCLC cells, we assessed the expression of KPNB1 and NCL. NAT10 knockdown significantly reduced KPNB1 expression in PC9R and A549R cells but had no effect on NCL expression (Figure 4e and f). Furthermore, NAT10 knockdown decreased ac4C modification levels in PC9R and A549R cells (Figure 4g). RIP confirmed the interaction between NAT10 and KPNB1 (Figure 4h). NAT10 knockdown also accelerated the degradation of KPNB1 mRNA, indicating reduced stability of KPNB1 transcripts (Figure 4i). Dual-luciferase reporter assays demonstrated that NAT10 knockdown significantly reduced the luciferase activity of wt KPNB1 but had no effect on mut KPNB1 in radiation-resistant NSCLC cells (Figure 4j). Collectively, these findings demonstrate that NAT10 knockdown suppresses KPNB1 expression by reducing ac4C modification on KPNB1 mRNA in radiation-resistant NSCLC cells.

NAT10 knockdown inhibited KPNB1 expression by decreasing ac4C modification on KPNB1 in radiation-resistant NSCLC cells. (a) Genes related to NAT10 in LUAD and LUSC were screened using the LinkedOmics database. (b) Pathway enrichment analysis was conducted using Funrich software. (c) and (d) Correlation between genes related to NAT10 and NAT10 was analyzed by the LinkedOmics database. (e) and (f) Expression of KPNB1 and NCL in PC9R and A549R cells was measured by qPCR. (g) The ac4C levels on KPNB1 in PC9R and A549R cells were detected by MeRIP. (h) Interaction between NAT10 and KPNB1 was evaluated by RIP. (i) Stability of KPNB1 mRNA in PC9R and A549R cells was evaluated by qPCR before and 4, 8, and 12 h after 5 μg/mL actinomycin D treatment. (j) Luciferase activity of wt-KPNB1 and mut-KPNB1 in PC9R and A549R cells was measured by dual luciferase report.
3.5 KPNB1 promotes radiation-resistant NSCLC cells immune escape by promotion of PD-L1 nuclear translocation
Our previous findings indicated that NAT10 knockdown downregulated nuclear PD-L1 protein levels in PC9R and A549R cells (Figure S3). Given that PD-L1 is a critical mediator of immune escape and its nuclear translocation enhances immune evasion in tumor cells, we investigated whether KPNB1, a nuclear transport factor, directly regulates PD-L1 nuclear translocation in radiation-resistant NSCLC cells. Results showed that both KPNB1 and PD-L1 expression levels were significantly higher in PC9R and A549R cells compared to their parental counterparts (Figure 5a and b). Co-IP confirmed the physical interaction between KPNB1 and PD-L1 in PC9R and A549R cells (Figure 5c and d). IF staining further demonstrated co-localization of KPNB1 and PD-L1 in these cells (Figure 5e). To assess the functional role of KPNB1 in PD-L1 nuclear translocation, we examined the effects of KPNB1 modulation. KPNB1 knockdown significantly reduced nuclear PD-L1 protein levels in PC9R and A549R cells (Figure 5d), whereas KPNB1 overexpression increased nuclear PD-L1 levels (Figure 5e), indicating that KPNB1 facilitates PD-L1 nuclear entry in radiation-resistant NSCLC cells. Furthermore, overexpression of nuclear PD-L1 in PC9R and A549R cells significantly decreased the proportion of CD8+ T cells, suggesting enhanced immune escape (Figure S4a–c). Together, these findings demonstrate that KPNB1 promotes immune escape in radiation-resistant NSCLC cells by facilitating PD-L1 nuclear translocation.

KPNB1 promoted radiation-resistant NSCLC cells immune escape by promotion of PD-L1 nuclear translocation. (a) and (b) Expression of KPNB1 and PD-L1 in parental and resistant PC9 and A549 cells was measured by qPCR. (c) and (d) The interaction between KPNB1 and PD-L1 was detected by Co-IP. (e) The location of KPNB1 and PD-L1 in PC9R and A549R cells was identified by IF staining. (f) and (g) The protein levels of KPNB1 in cells and PD-L1 in nuclear in PC9R and A549R cells were detected by western blot.
3.6 KPNB1 overexpression restores RT resistance in NSCLC cells improved by NAT10 knockdown
To investigate the role of KPNB1 in RT resistance in NSCLC cells, we examined the effects of NAT10 knockdown and KPNB1 overexpression. NAT10 knockdown significantly reduced the mRNA expression of both NAT10 and KPNB1 in PC9R and A549R cells, while KPNB1 overexpression markedly increased KPNB1 mRNA levels without affecting NAT10 expression (Figure 6a and b). At the protein level, NAT10 knockdown downregulated KPNB1 and PD-L1 expression in PC9R and A549R cells, whereas KPNB1 overexpression partially restored PD-L1 protein levels without altering NAT10 expression (Figure 6c). Functionally, KPNB1 overexpression rescued the inhibition of cell proliferation induced by NAT10 knockdown in PC9R and A549R cells (Figure 6d). Additionally, the enhanced apoptosis caused by NAT10 knockdown was significantly suppressed by KPNB1 overexpression (Figure 6e). Flow cytometry analysis revealed that the increase in CD8+ T cells resulting from NAT10 knockdown was reversed by KPNB1 overexpression (Figure 6f). Furthermore, KPNB1 overexpression reduced the levels of TNF-α and IFN-γ, which were elevated by NAT10 knockdown (Figure 6g and h), while restoring the levels of PD-1 and TIM-3 that were suppressed by NAT10 knockdown (Figure 6i and j). Collectively, these findings demonstrate that KPNB1 overexpression restores RT resistance in NSCLC cells by reversing the enhanced immune function and cellular effects induced by NAT10 knockdown.

KPNB1 overexpression restored RT resistance in NSCLC cells improved by NAT10 knockdown. (a) and (b) Expression of NAT10 and KPNB1 of PC9R and A549R cells was measured by qPCR. (c) The protein levels of NAT10, KPNB1, and PD-L1 of PC9R and A549R cells were detected by western blot. (d) Cell proliferation ability of PC9R and A549R cells was evaluated by colony formation assay. (e) Apoptosis of PC9R and A549R cells was evaluated by flow cytometry. (f) The amount of CD4+ and CD8+ T cells was detected by flow cytometry. (g)–(j) The levels of TNF-α, IFN-γ, PD-1, and TIM3 were measured by qPCR.
4 Discussion
Lung cancer remains the second leading cause of cancer-related mortality worldwide, with NSCLC accounting for approximately 80% of cases. Despite advances in treatment, the prognosis for NSCLC patients remains poor, characterized by a significantly low 5-year survival rate [28]. RT is a cornerstone in the treatment of various cancers, including NSCLC; however, a growing body of evidence indicates that patients often develop RT resistance over time, leading to treatment failure, metastasis, cancer recurrence, and poor prognosis. This resistance represents a major challenge in cancer therapy [29]. The development of RT resistance is closely linked to RT-induced immune dysfunction, including immunogenic cell death and depletion of T cells, which collectively maintain an immunosuppressive tumor microenvironment conducive to tumor progression [29,30]. In NSCLC, strategies targeting PD-L1 and preventing CD8+ T cell depletion have shown promise in overcoming RT resistance and enhancing anti-tumor immunity [17,31]. Therefore, elucidating the mechanisms underlying RT resistance, particularly those involving immune regulation, is critical for improving anti-tumor immunity and patient outcomes. In this study, we demonstrated for the first time that NAT10 expression is elevated in radiation-resistant NSCLC cells. Mechanistically, NAT10 enhances KPNB1 expression through increased ac4C modification, thereby promoting immune escape in NSCLC cells by facilitating PD-L1 nuclear translocation. This process ultimately contributes to RT resistance in NSCLC.
NAT10 is the sole known enzyme responsible for catalyzing ac4C production in eukaryotic RNA. By mediating ac4C modification, NAT10 regulates RNA stability and translation efficiency, playing a critical role in the pathogenesis of various diseases. Accumulating evidence indicates that upregulated NAT10 expression is associated with cancer development and metastasis [21–23]. Specifically, NAT10 is significantly overexpressed in patients with NSCLC and has been shown to promote the proliferation, migration, and metastasis of NSCLC cells [25]. Previous studies have reported that NAT10 regulates NSCLC progression through mechanisms involving epithelial–mesenchymal transition, cell cycle modulation, and immune cell infiltration [24,25,32]. NAT10 also plays a pivotal role in cancer immune regulation. For instance, Li et al. [33] demonstrated that NAT10 enhances YWHAH expression through ac4C modification, leading to CD8+ T cell depletion in colorectal cancer. Additionally, NAT10 promotes PD-L1 transcription in various cancers by facilitating NPM1 acetylation, which correlates with poor patient prognosis [34]. Notably, radiation has been shown to enhance NAT10-mediated acetylation of MORC2 in breast cancer, suggesting that NAT10 may serve as a potential therapeutic target to overcome RT resistance [35]. However, the mechanisms by which NAT10 mediates RT resistance in NSCLC remain poorly understood. In this study, we demonstrated that NAT10 is upregulated in radiation-resistant NSCLC cells, where it enhances ac4C modification and mRNA stability of KPNB1. This process promotes PD-L1 nuclear translocation and immune escape in radiation-resistant NSCLC cells. These findings establish NAT10 as a key driver of RT resistance in NSCLC.
KPNB1, a nuclear transport receptor, plays a critical role in mediating nuclear import processes [36]. It facilitates the nuclear translocation of numerous key tumor-related mediators, thereby driving tumor progression [37]. For instance, Li et al. [38] demonstrated that KPNB1 regulates the downstream expression of NLGN3 by mediating the nuclear import of YBX1, promoting the growth of glioma cells. Similarly, Du et al. [39] showed that KPNB1 facilitates the nuclear translocation of PD-L1 in NSCLC cells, enhancing their proliferation. Additionally, KPNB1 has been implicated in RT resistance. Hazawa et al. [40] reported that inhibiting KPNB1 enhances radiation-induced apoptosis and radiosensitivity while reducing PD-L1 expression on the cell surface in human head and neck squamous cell carcinoma cells.
In this study, we provide the first evidence of KPNB1’s role in RT resistance in NSCLC. We demonstrated that KPNB1 interacts with PD-L1 in radiation-resistant NSCLC cells and promotes PD-L1 nuclear translocation, thereby contributing to radiation resistance in NSCLC.
In summary, this study demonstrates that NAT10 acts as a key promoter of RT resistance in NSCLC. NAT10 enhances KPNB1 expression through ac4C modification, thereby facilitating PD-L1 nuclear translocation and promoting immune escape in radiation-resistant NSCLC cells. These findings reveal a novel mechanism underlying RT resistance in NSCLC and suggest that targeting NAT10 or its downstream pathways may provide a promising therapeutic strategy to overcome RT resistance in clinical settings.
-
Funding information: This study was supported by Natural Key Project of Bengbu Medical University in 2023 (2023byzd024).
-
Author contributions: All authors contributed to the study conception and design. Material preparation, data collection, and analysis were performed by Dagao Zhu, Mingliang Lu, and Hongmin Cheng. The first draft of the manuscript was written by Dagao Zhu and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
-
Conflict of interest: Authors state no conflict of interest.
-
Data availability statement: The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
[1] Chen P, Liu Y, Wen Y, Zhou C. Non-small cell lung cancer in China. Cancer Commun (Lond). 2022;42(10):937–70.10.1002/cac2.12359Search in Google Scholar PubMed PubMed Central
[2] Chaft JE, Rimner A, Weder W, Azzoli CG, Kris MG, Cascone T. Evolution of systemic therapy for stages I–III non-metastatic non-small-cell lung cancer. Nat Rev Clin Oncol. 2021;18(9):547–57.10.1038/s41571-021-00501-4Search in Google Scholar PubMed PubMed Central
[3] Miao D, Zhao J, Han Y, Zhou J, Li X, Zhang T, et al. Management of locally advanced non‐small cell lung cancer: state of the art and future directions. Cancer Commun (London, Engl). 2024;44(1):23–46.10.1002/cac2.12505Search in Google Scholar PubMed PubMed Central
[4] de Scordilli M, Michelotti A, Bertoli E, De Carlo E, Del CA, Bearz A. Targeted therapy and immunotherapy in early-stage non-small cell lung cancer: current evidence and ongoing trials. Int J Mol Sci. 2022;23(13):7222.10.3390/ijms23137222Search in Google Scholar PubMed PubMed Central
[5] Yang C, Liang Y, Liu N, Sun M. Role of the cGAS-STING pathway in radiotherapy for non-small cell lung cancer. Radiat Oncol (London, Engl). 2023;18(1):1–145.10.1186/s13014-023-02335-zSearch in Google Scholar PubMed PubMed Central
[6] Li Y, Juergens RA, Finley C, Swaminath A. Current and future treatment options in the management of stage III NSCLC. J Thorac Oncol. 2023;18(11):1478–91.10.1016/j.jtho.2023.08.011Search in Google Scholar PubMed
[7] Chen ZY, Duan XT, Qiao SM, Zhu XX. Radiotherapy combined with PD-1/PD-L1 inhibitors in NSCLC brain metastases treatment: the mechanisms, advances, opportunities, and challenges. Cancer Med. 2023;12(2):995–1006.10.1002/cam4.5016Search in Google Scholar PubMed PubMed Central
[8] Iyengar P, Zhang-Velten E, Court L, Westover K, Yan Y, Lin MH, et al. Accelerated hypofractionated image-guided vs conventional radiotherapy for patients with stage II/III non-small cell lung cancer and poor performance status: a randomized clinical trial. JAMA Oncol. 2021;7(10):1497–505.10.1001/jamaoncol.2021.3186Search in Google Scholar PubMed PubMed Central
[9] An L, Li M, Jia Q. Mechanisms of radiotherapy resistance and radiosensitization strategies for esophageal squamous cell carcinoma. Mol Cancer. 2023;22(1):140.10.1186/s12943-023-01839-2Search in Google Scholar PubMed PubMed Central
[10] Xu K, Guo H, Xia A, Wang Z, Wang S, Wang Q. Non-coding RNAs in radiotherapy resistance: roles and therapeutic implications in gastrointestinal cancer. Biomed Pharmacother. 2023;161:114485.10.1016/j.biopha.2023.114485Search in Google Scholar PubMed
[11] Liu Y, Feng Z, Zhang P, Chen H, Zhu S, Wang X. Advances in the study of aerobic glycolytic effects in resistance to radiotherapy in malignant tumors. PeerJ (San Francisco, CA). 2023;11:e14930.10.7717/peerj.14930Search in Google Scholar PubMed PubMed Central
[12] Zhai G, Li J, Zheng J, An P, Chen X, Wang X, et al. hTERT promoter methylation promotes small cell lung cancer progression and radiotherapy resistance. J Radiat Res. 2020;61(5):674–83.10.1093/jrr/rraa052Search in Google Scholar PubMed PubMed Central
[13] Zhang N, Liu X, Huang L, Zeng J, Ma C, Han L, et al. LINC00921 reduces lung cancer radiosensitivity by destabilizing NUDT21 and driving aberrant MED23 alternative polyadenylation. Cell Rep. 2023;42(12):113479.10.1016/j.celrep.2023.113479Search in Google Scholar PubMed
[14] Zhang L, Xu Y, Cheng Z, Zhao J, Wang M, Sun Y, et al. The EGR1/miR-139/NRF2 axis orchestrates radiosensitivity of non-small-cell lung cancer via ferroptosis. Cancer Lett. 2024;595:217000.10.1016/j.canlet.2024.217000Search in Google Scholar PubMed
[15] Lander VE, Belle JI, Kingston NL, Herndon JM, Hogg GD, Liu X, et al. Stromal reprogramming by FAK inhibition overcomes radiation resistance to allow for immune priming and response to checkpoint blockade. Cancer Discov. 2022;12(12):2774–99.10.1158/2159-8290.CD-22-0192Search in Google Scholar PubMed PubMed Central
[16] Zhang P, Rashidi A, Zhao J, Silvers C, Wang H, Castro B, et al. STING agonist-loaded, CD47/PD-L1-targeting nanoparticles potentiate antitumor immunity and radiotherapy for glioblastoma. Nat Commun. 2023;14(1):1610.10.1038/s41467-023-37328-9Search in Google Scholar PubMed PubMed Central
[17] Wang J, Xu Y, Rao X, Zhang R, Tang J, Zhang D, et al. BRD4-IRF1 axis regulates chemoradiotherapy-induced PD-L1 expression and immune evasion in non-small cell lung cancer. Clin Transl Med. 2022;12(1):e718.10.1002/ctm2.718Search in Google Scholar PubMed PubMed Central
[18] Ancey PB, Contat C, Boivin G, Sabatino S, Pascual J, Zangger N, et al. GLUT1 expression in tumor-associated neutrophils promotes lung cancer growth and resistance to radiotherapy. Cancer Res. 2021;81(9):2345–57.10.1158/0008-5472.CAN-20-2870Search in Google Scholar PubMed PubMed Central
[19] Xie L, Zhong X, Cao W, Liu J, Zu X, Chen L. Mechanisms of NAT10 as ac4C writer in diseases. Mol Ther Nucleic Acids. 2023;32:359–68.10.1016/j.omtn.2023.03.023Search in Google Scholar PubMed PubMed Central
[20] Zhang S, Liu Y, Ma X, Gao X, Ru Y, Hu X, et al. Recent advances in the potential role of RNA N4-acetylcytidine in cancer progression. Cell Commun Signal. 2024;22(1):49.10.1186/s12964-023-01417-5Search in Google Scholar PubMed PubMed Central
[21] Liao L, He Y, Li SJ, Yu XM, Liu ZC, Liang YY, et al. Lysine 2-hydroxyisobutyrylation of NAT10 promotes cancer metastasis in an ac4C-dependent manner. Cell Res. 2023;33(5):355–71.10.1038/s41422-023-00793-4Search in Google Scholar PubMed PubMed Central
[22] Zhang Y, Jing Y, Wang Y, Tang J, Zhu X, Jin W, et al. NAT10 promotes gastric cancer metastasis via N4-acetylated COL5A1. Signal Transduct Target Ther. 2021;6(1):173.10.1038/s41392-021-00489-4Search in Google Scholar PubMed PubMed Central
[23] Jin C, Wang T, Zhang D, Yang P, Zhang C, Peng W, et al. Acetyltransferase NAT10 regulates the Wnt/β-catenin signaling pathway to promote colorectal cancer progression via ac4C acetylation of KIF23 mRNA. J Exp Clin Cancer Res. 2022;41(1):1–345.10.1186/s13046-022-02551-7Search in Google Scholar PubMed PubMed Central
[24] Wang Z, Huang Y, Lu W, Liu J, Li X, Zhu S, et al. c-myc-mediated upregulation of NAT10 facilitates tumor development via cell cycle regulation in non-small cell lung cancer. Med Oncol (Northwood, London, Engl). 2022;39(10):140.10.1007/s12032-022-01736-6Search in Google Scholar PubMed
[25] Guo Q, Yu W, Tan J, Zhang J, Chen J, Rao S, et al. Remodelin delays non-small cell lung cancer progression by inhibiting NAT10 via the EMT pathway. Cancer Med. 2024;13(11):e7283.10.1002/cam4.7283Search in Google Scholar PubMed PubMed Central
[26] Kim TW, Hong D, Hong SH. PB01 suppresses radio-resistance by regulating ATR signaling in human non-small-cell lung cancer cells. Sci Rep-UK. 2021;11(1):12093.10.1038/s41598-021-91716-zSearch in Google Scholar PubMed PubMed Central
[27] Xu S, Tao Z, Hai B, Liang H, Shi Y, Wang T, et al. miR-424(322) reverses chemoresistance via T-cell immune response activation by blocking the PD-L1 immune checkpoint. Nat Commun. 2016;7:11406.10.1038/ncomms11406Search in Google Scholar PubMed PubMed Central
[28] Qiu H, Cao S, Xu R. Cancer incidence, mortality, and burden in China: a time-trend analysis and comparison with the United States and United Kingdom based on the global epidemiological data released in 2020. Cancer Commun (London, Engl). 2021;41(10):1037–48.10.1002/cac2.12197Search in Google Scholar PubMed PubMed Central
[29] Wu Y, Song Y, Wang R, Wang T. Molecular mechanisms of tumor resistance to radiotherapy. Mol Cancer. 2023;22(1):96.10.1186/s12943-023-01801-2Search in Google Scholar PubMed PubMed Central
[30] Huang Z, Wang Y, Yao D, Wu J, Hu Y, Yuan A. Nanoscale coordination polymers induce immunogenic cell death by amplifying radiation therapy mediated oxidative stress. Nat Commun. 2021;12(1):145.10.1038/s41467-020-20243-8Search in Google Scholar PubMed PubMed Central
[31] Zhang Y, Hu J, Ji K, Jiang S, Dong Y, Sun L, et al. CD39 inhibition and VISTA blockade may overcome radiotherapy resistance by targeting exhausted CD8+ T cells and immunosuppressive myeloid cells. Cell Rep Med. 2023;4(8):101151.10.1016/j.xcrm.2023.101151Search in Google Scholar PubMed PubMed Central
[32] Wang Z, Luo J, Huang H, Wang L, Lv T, Wang Z, et al. NAT10-mediated upregulation of GAS5 facilitates immune cell infiltration in non-small cell lung cancer via the MYBBP1A-p53/IRF1/type I interferon signaling axis. Cell Death Discov. 2024;10(1):240.10.1038/s41420-024-01997-2Search in Google Scholar PubMed PubMed Central
[33] Li Q, Yuan Z, Wang Y, Zhai P, Wang J, Zhang C, et al. Unveiling YWHAH: A potential therapeutic target for overcoming CD8+ T cell exhaustion in colorectal cancer. Int Immunopharmacol. 2024;135:112317.10.1016/j.intimp.2024.112317Search in Google Scholar PubMed
[34] Qin G, Bai F, Hu H, Zhang J, Zhan W, Wu Z, et al. Targeting the NAT10/NPM1 axis abrogates PD-L1 expression and improves the response to immune checkpoint blockade therapy. Mol Med. 2024;30(1):13.10.1186/s10020-024-00780-4Search in Google Scholar PubMed PubMed Central
[35] Liu HY, Liu YY, Yang F, Zhang L, Zhang FL, Hu X, et al. Acetylation of MORC2 by NAT10 regulates cell-cycle checkpoint control and resistance to DNA-damaging chemotherapy and radiotherapy in breast cancer. Nucleic Acids Res. 2020;48(7):3638–56.10.1093/nar/gkaa130Search in Google Scholar PubMed PubMed Central
[36] Newell S, van der Watt PJ, Leaner VD. Therapeutic targeting of nuclear export and import receptors in cancer and their potential in combination chemotherapy. IUBMB Life. 2024;76(1):4–25.10.1002/iub.2773Search in Google Scholar PubMed PubMed Central
[37] Shi Q, Lin M, Cheng X, Zhang Z, Deng S, Lang K, et al. KPNB1-mediated nuclear import in cancer. EUR J Pharmacol. 2023;955:175925.10.1016/j.ejphar.2023.175925Search in Google Scholar PubMed
[38] Li J, Zhang B, Feng Z, An D, Zhou Z, Wan C, et al. Stabilization of KPNB1 by deubiquitinase USP7 promotes glioblastoma progression through the YBX1-NLGN3 axis. J Exp Clin Cancer Res. 2024;43(1):28.10.1186/s13046-024-02954-8Search in Google Scholar PubMed PubMed Central
[39] Du W, Zhu J, Zeng Y, Liu T, Zhang Y, Cai T, et al. KPNB1-mediated nuclear translocation of PD-L1 promotes non-small cell lung cancer cell proliferation via the Gas6/MerTK signaling pathway. Cell Death Differ. 2021;28(4):1284–300.10.1038/s41418-020-00651-5Search in Google Scholar PubMed PubMed Central
[40] Hazawa M, Yoshino H, Nakagawa Y, Shimizume R, Nitta K, Sato Y, et al. Karyopherin-β1 regulates radioresistance and radiation-increased programmed death-ligand 1 expression in human head and neck squamous cell carcinoma cell lines. Cancers. 2020;12(4):908.10.3390/cancers12040908Search in Google Scholar PubMed PubMed Central
© 2025 the author(s), published by De Gruyter
This work is licensed under the Creative Commons Attribution 4.0 International License.
Articles in the same Issue
- Biomedical Sciences
- Mechanism of triptolide regulating proliferation and apoptosis of hepatoma cells by inhibiting JAK/STAT pathway
- Maslinic acid improves mitochondrial function and inhibits oxidative stress and autophagy in human gastric smooth muscle cells
- Comparative analysis of inflammatory biomarkers for the diagnosis of neonatal sepsis: IL-6, IL-8, SAA, CRP, and PCT
- Post-pandemic insights on COVID-19 and premature ovarian insufficiency
- Proteome differences of dental stem cells between permanent and deciduous teeth by data-independent acquisition proteomics
- Optimizing a modified cetyltrimethylammonium bromide protocol for fungal DNA extraction: Insights from multilocus gene amplification
- Preliminary analysis of the role of small hepatitis B surface proteins mutations in the pathogenesis of occult hepatitis B infection via the endoplasmic reticulum stress-induced UPR-ERAD pathway
- Efficacy of alginate-coated gold nanoparticles against antibiotics-resistant Staphylococcus and Streptococcus pathogens of acne origins
- Battling COVID-19 leveraging nanobiotechnology: Gold and silver nanoparticle–B-escin conjugates as SARS-CoV-2 inhibitors
- Neurodegenerative diseases and neuroinflammation-induced apoptosis
- Impact of fracture fixation surgery on cognitive function and the gut microbiota in mice with a history of stroke
- COLEC10: A potential tumor suppressor and prognostic biomarker in hepatocellular carcinoma through modulation of EMT and PI3K-AKT pathways
- High-temperature requirement serine protease A2 inhibitor UCF-101 ameliorates damaged neurons in traumatic brain-injured rats by the AMPK/NF-κB pathway
- SIK1 inhibits IL-1β-stimulated cartilage apoptosis and inflammation in vitro through the CRTC2/CREB1 signaling
- Rutin–chitooligosaccharide complex: Comprehensive evaluation of its anti-inflammatory and analgesic properties in vitro and in vivo
- Knockdown of Aurora kinase B alleviates high glucose-triggered trophoblast cells damage and inflammation during gestational diabetes
- Calcium-sensing receptors promoted Homer1 expression and osteogenic differentiation in bone marrow mesenchymal stem cells
- ABI3BP can inhibit the proliferation, invasion, and epithelial–mesenchymal transition of non-small-cell lung cancer cells
- Changes in blood glucose and metabolism in hyperuricemia mice
- Rapid detection of the GJB2 c.235delC mutation based on CRISPR-Cas13a combined with lateral flow dipstick
- IL-11 promotes Ang II-induced autophagy inhibition and mitochondrial dysfunction in atrial fibroblasts
- Short-chain fatty acid attenuates intestinal inflammation by regulation of gut microbial composition in antibiotic-associated diarrhea
- Application of metagenomic next-generation sequencing in the diagnosis of pathogens in patients with diabetes complicated by community-acquired pneumonia
- NAT10 promotes radiotherapy resistance in non-small cell lung cancer by regulating KPNB1-mediated PD-L1 nuclear translocation
- Phytol-mixed micelles alleviate dexamethasone-induced osteoporosis in zebrafish: Activation of the MMP3–OPN–MAPK pathway-mediating bone remodeling
- Association between TGF-β1 and β-catenin expression in the vaginal wall of patients with pelvic organ prolapse
- Primary pleomorphic liposarcoma involving bilateral ovaries: Case report and literature review
- Effects of de novo donor-specific Class I and II antibodies on graft outcomes after liver transplantation: A pilot cohort study
- Sleep architecture in Alzheimer’s disease continuum: The deep sleep question
- Ephedra fragilis plant extract: A groundbreaking corrosion inhibitor for mild steel in acidic environments – electrochemical, EDX, DFT, and Monte Carlo studies
- Langerhans cell histiocytosis in an adult patient with upper jaw and pulmonary involvement: A case report
- Inhibition of mast cell activation by Jaranol-targeted Pirin ameliorates allergic responses in mouse allergic rhinitis
- Aeromonas veronii-induced septic arthritis of the hip in a child with acute lymphoblastic leukemia
- Clusterin activates the heat shock response via the PI3K/Akt pathway to protect cardiomyocytes from high-temperature-induced apoptosis
- Research progress on fecal microbiota transplantation in tumor prevention and treatment
- Low-pressure exposure influences the development of HAPE
- Stigmasterol alleviates endplate chondrocyte degeneration through inducing mitophagy by enhancing PINK1 mRNA acetylation via the ESR1/NAT10 axis
- AKAP12, mediated by transcription factor 21, inhibits cell proliferation, metastasis, and glycolysis in lung squamous cell carcinoma
- Association between PAX9 or MSX1 gene polymorphism and tooth agenesis risk: A meta-analysis
- A case of bloodstream infection caused by Neisseria gonorrhoeae
- Case of nasopharyngeal tuberculosis complicated with cervical lymph node and pulmonary tuberculosis
- p-Cymene inhibits pro-fibrotic and inflammatory mediators to prevent hepatic dysfunction
- GFPT2 promotes paclitaxel resistance in epithelial ovarian cancer cells via activating NF-κB signaling pathway
- Transfer RNA-derived fragment tRF-36 modulates varicose vein progression via human vascular smooth muscle cell Notch signaling
- RTA-408 attenuates the hepatic ischemia reperfusion injury in mice possibly by activating the Nrf2/HO-1 signaling pathway
- Decreased serum TIMP4 levels in patients with rheumatoid arthritis
- Sirt1 protects lupus nephritis by inhibiting the NLRP3 signaling pathway in human glomerular mesangial cells
- Sodium butyrate aids brain injury repair in neonatal rats
- Interaction of MTHFR polymorphism with PAX1 methylation in cervical cancer
- Convallatoxin inhibits proliferation and angiogenesis of glioma cells via regulating JAK/STAT3 pathway
- The effect of the PKR inhibitor, 2-aminopurine, on the replication of influenza A virus, and segment 8 mRNA splicing
- Effects of Ire1 gene on virulence and pathogenicity of Candida albicans
- Small cell lung cancer with small intestinal metastasis: Case report and literature review
- GRB14: A prognostic biomarker driving tumor progression in gastric cancer through the PI3K/AKT signaling pathway by interacting with COBLL1
- 15-Lipoxygenase-2 deficiency induces foam cell formation that can be restored by salidroside through the inhibition of arachidonic acid effects
- FTO alleviated the diabetic nephropathy progression by regulating the N6-methyladenosine levels of DACT1
- Clinical relevance of inflammatory markers in the evaluation of severity of ulcerative colitis: A retrospective study
- Zinc valproic acid complex promotes osteoblast differentiation and exhibits anti-osteoporotic potential
- Primary pulmonary synovial sarcoma in the bronchial cavity: A case report
- Metagenomic next-generation sequencing of alveolar lavage fluid improves the detection of pulmonary infection
- Uterine tumor resembling ovarian sex cord tumor with extensive rhabdoid differentiation: A case report
- Genomic analysis of a novel ST11(PR34365) Clostridioides difficile strain isolated from the human fecal of a CDI patient in Guizhou, China
- Effects of tiered cardiac rehabilitation on CRP, TNF-α, and physical endurance in older adults with coronary heart disease
- Changes in T-lymphocyte subpopulations in patients with colorectal cancer before and after acupoint catgut embedding acupuncture observation
- Modulating the tumor microenvironment: The role of traditional Chinese medicine in improving lung cancer treatment
- Alterations of metabolites related to microbiota–gut–brain axis in plasma of colon cancer, esophageal cancer, stomach cancer, and lung cancer patients
- Research on individualized drug sensitivity detection technology based on bio-3D printing technology for precision treatment of gastrointestinal stromal tumors
- CEBPB promotes ulcerative colitis-associated colorectal cancer by stimulating tumor growth and activating the NF-κB/STAT3 signaling pathway
- Oncolytic bacteria: A revolutionary approach to cancer therapy
- A de novo meningioma with rapid growth: A possible malignancy imposter?
- Diagnosis of secondary tuberculosis infection in an asymptomatic elderly with cancer using next-generation sequencing: Case report
- Hesperidin and its zinc(ii) complex enhance osteoblast differentiation and bone formation: In vitro and in vivo evaluations
- Research progress on the regulation of autophagy in cardiovascular diseases by chemokines
- Anti-arthritic, immunomodulatory, and inflammatory regulation by the benzimidazole derivative BMZ-AD: Insights from an FCA-induced rat model
- Immunoassay for pyruvate kinase M1/2 as an Alzheimer’s biomarker in CSF
- The role of HDAC11 in age-related hearing loss: Mechanisms and therapeutic implications
- Evaluation and application analysis of animal models of PIPNP based on data mining
- Therapeutic approaches for liver fibrosis/cirrhosis by targeting pyroptosis
- Fabrication of zinc oxide nanoparticles using Ruellia tuberosa leaf extract induces apoptosis through P53 and STAT3 signalling pathways in prostate cancer cells
- Haplo-hematopoietic stem cell transplantation and immunoradiotherapy for severe aplastic anemia complicated with nasopharyngeal carcinoma: A case report
- Modulation of the KEAP1-NRF2 pathway by Erianin: A novel approach to reduce psoriasiform inflammation and inflammatory signaling
- The expression of epidermal growth factor receptor 2 and its relationship with tumor-infiltrating lymphocytes and clinical pathological features in breast cancer patients
- Innovations in MALDI-TOF Mass Spectrometry: Bridging modern diagnostics and historical insights
- BAP1 complexes with YY1 and RBBP7 and its downstream targets in ccRCC cells
- Hypereosinophilic syndrome with elevated IgG4 and T-cell clonality: A report of two cases
- Electroacupuncture alleviates sciatic nerve injury in sciatica rats by regulating BDNF and NGF levels, myelin sheath degradation, and autophagy
- Polydatin prevents cholesterol gallstone formation by regulating cholesterol metabolism via PPAR-γ signaling
- RNF144A and RNF144B: Important molecules for health
- Analysis of the detection rate and related factors of thyroid nodules in the healthy population
- Artesunate inhibits hepatocellular carcinoma cell migration and invasion through OGA-mediated O-GlcNAcylation of ZEB1
- Endovascular management of post-pancreatectomy hemorrhage caused by a hepatic artery pseudoaneurysm: Case report and review of the literature
- Efficacy and safety of anti-PD-1/PD-L1 antibodies in patients with relapsed refractory diffuse large B-cell lymphoma: A meta-analysis
- SATB2 promotes humeral fracture healing in rats by activating the PI3K/AKT pathway
- Overexpression of the ferroptosis-related gene, NFS1, corresponds to gastric cancer growth and tumor immune infiltration
- Understanding risk factors and prognosis in diabetic foot ulcers
- Atractylenolide I alleviates the experimental allergic response in mice by suppressing TLR4/NF-kB/NLRP3 signalling
- FBXO31 inhibits the stemness characteristics of CD147 (+) melanoma stem cells
- Immune molecule diagnostics in colorectal cancer: CCL2 and CXCL11
- Inhibiting CXCR6 promotes senescence of activated hepatic stellate cells with limited proinflammatory SASP to attenuate hepatic fibrosis
- Cadmium toxicity, health risk and its remediation using low-cost biochar adsorbents
- Pulmonary cryptococcosis with headache as the first presentation: A case report
- Solitary pulmonary metastasis with cystic airspaces in colon cancer: A rare case report
- RUNX1 promotes denervation-induced muscle atrophy by activating the JUNB/NF-κB pathway and driving M1 macrophage polarization
- Morphometric analysis and immunobiological investigation of Indigofera oblongifolia on the infected lung with Plasmodium chabaudi
- The NuA4/TIP60 histone-modifying complex and Hr78 modulate the Lobe2 mutant eye phenotype
- Experimental study on salmon demineralized bone matrix loaded with recombinant human bone morphogenetic protein-2: In vitro and in vivo study
- A case of IgA nephropathy treated with a combination of telitacicept and half-dose glucocorticoids
- Analgesic and toxicological evaluation of cannabidiol-rich Moroccan Cannabis sativa L. (Khardala variety) extract: Evidence from an in vivo and in silico study
- Wound healing and signaling pathways
- Combination of immunotherapy and whole-brain radiotherapy on prognosis of patients with multiple brain metastases: A retrospective cohort study
- To explore the relationship between endometrial hyperemia and polycystic ovary syndrome
- Research progress on the impact of curcumin on immune responses in breast cancer
- Biogenic Cu/Ni nanotherapeutics from Descurainia sophia (L.) Webb ex Prantl seeds for the treatment of lung cancer
- Dapagliflozin attenuates atrial fibrosis via the HMGB1/RAGE pathway in atrial fibrillation rats
- Glycitein alleviates inflammation and apoptosis in keratinocytes via ROS-associated PI3K–Akt signalling pathway
- ADH5 inhibits proliferation but promotes EMT in non-small cell lung cancer cell through activating Smad2/Smad3
- Apoptotic efficacies of AgNPs formulated by Syzygium aromaticum leaf extract on 32D-FLT3-ITD human leukemia cell line with PI3K/AKT/mTOR signaling pathway
- Novel cuproptosis-related genes C1QBP and PFKP identified as prognostic and therapeutic targets in lung adenocarcinoma
- Bee venom promotes exosome secretion and alters miRNA cargo in T cells
- Treatment of pure red cell aplasia in a chronic kidney disease patient with roxadustat: A case report
- Comparative bioinformatics analysis of the Wnt pathway in breast cancer: Selection of novel biomarker panels associated with ER status
- Kynurenine facilitates renal cell carcinoma progression by suppressing M2 macrophage pyroptosis through inhibition of CASP1 cleavage
- RFX5 promotes the growth, motility, and inhibits apoptosis of gastric adenocarcinoma cells through the SIRT1/AMPK axis
- ALKBH5 exacerbates early cardiac damage after radiotherapy for breast cancer via m6A demethylation of TLR4
- Phytochemicals of Roman chamomile: Antioxidant, anti-aging, and whitening activities of distillation residues
- Circadian gene Cry1 inhibits the tumorigenicity of hepatocellular carcinoma by the BAX/BCL2-mediated apoptosis pathway
- The TNFR-RIPK1/RIPK3 signalling pathway mediates the effect of lanthanum on necroptosis of nerve cells
- Longitudinal monitoring of autoantibody dynamics in patients with early-stage non-small-cell lung cancer undergoing surgery
- The potential role of rutin, a flavonoid, in the management of cancer through modulation of cell signaling pathways
- Construction of pectinase gene engineering microbe and its application in tobacco sheets
- Construction of a microbial abundance prognostic scoring model based on intratumoral microbial data for predicting the prognosis of lung squamous cell carcinoma
- Sepsis complicated by haemophagocytic lymphohistiocytosis triggered by methicillin-resistant Staphylococcus aureus and human herpesvirus 8 in an immunocompromised elderly patient: A case report
- Sarcopenia in liver transplantation: A comprehensive bibliometric study of current research trends and future directions
- Advances in cancer immunotherapy and future directions in personalized medicine
- Can coronavirus disease 2019 affect male fertility or cause spontaneous abortion? A two-sample Mendelian randomization analysis
- Heat stroke associated with novel leukaemia inhibitory factor receptor gene variant in a Chinese infant
- PSME2 exacerbates ulcerative colitis by disrupting intestinal barrier function and promoting autophagy-dependent inflammation
- Hyperosmolar hyperglycemic state with severe hypernatremia coexisting with central diabetes insipidus: A case report and literature review
- Efficacy and mechanism of escin in improving the tissue microenvironment of blood vessel walls via anti-inflammatory and anticoagulant effects: Implications for clinical practice
- Merkel cell carcinoma: Clinicopathological analysis of three patients and literature review
- Genetic variants in VWF exon 26 and their implications for type 1 Von Willebrand disease in a Saudi Arabian population
- Lipoxin A4 improves myocardial ischemia/reperfusion injury through the Notch1-Nrf2 signaling pathway
- High levels of EPHB2 expression predict a poor prognosis and promote tumor progression in endometrial cancer
- Knockdown of SHP-2 delays renal tubular epithelial cell injury in diabetic nephropathy by inhibiting NLRP3 inflammasome-mediated pyroptosis
- Exploring the toxicity mechanisms and detoxification methods of Rhizoma Paridis
- Concomitant gastric carcinoma and primary hepatic angiosarcoma in a patient: A case report
- Ecology and Environmental Science
- Optimization and comparative study of Bacillus consortia for cellulolytic potential and cellulase enzyme activity
- The complete mitochondrial genome analysis of Haemaphysalis hystricis Supino, 1897 (Ixodida: Ixodidae) and its phylogenetic implications
- Epidemiological characteristics and risk factors analysis of multidrug-resistant tuberculosis among tuberculosis population in Huzhou City, Eastern China
- Indices of human impacts on landscapes: How do they reflect the proportions of natural habitats?
- Genetic analysis of the Siberian flying squirrel population in the northern Changbai Mountains, Northeast China: Insights into population status and conservation
- Diversity and environmental drivers of Suillus communities in Pinus sylvestris var. mongolica forests of Inner Mongolia
- Global assessment of the fate of nitrogen deposition in forest ecosystems: Insights from 15N tracer studies
- Fungal and bacterial pathogenic co-infections mainly lead to the assembly of microbial community in tobacco stems
- Influencing of coal industry related airborne particulate matter on ocular surface tear film injury and inflammatory factor expression in Sprague-Dawley rats
- Temperature-dependent development, predation, and life table of Sphaerophoria macrogaster (Thomson) (Diptera: Syrphidae) feeding on Myzus persicae (Sulzer) (Homoptera: Aphididae)
- Eleonora’s falcon trophic interactions with insects within its breeding range: A systematic review
- Agriculture
- Integrated analysis of transcriptome, sRNAome, and degradome involved in the drought-response of maize Zhengdan958
- Variation in flower frost tolerance among seven apple cultivars and transcriptome response patterns in two contrastingly frost-tolerant selected cultivars
- Heritability of durable resistance to stripe rust in bread wheat (Triticum aestivum L.)
- Molecular mechanism of follicular development in laying hens based on the regulation of water metabolism
- Animal Science
- Effect of sex ratio on the life history traits of an important invasive species, Spodoptera frugiperda
- Plant Sciences
- Hairpin in a haystack: In silico identification and characterization of plant-conserved microRNA in Rafflesiaceae
- Widely targeted metabolomics of different tissues in Rubus corchorifolius
- The complete chloroplast genome of Gerbera piloselloides (L.) Cass., 1820 (Carduoideae, Asteraceae) and its phylogenetic analysis
- Field trial to correlate mineral solubilization activity of Pseudomonas aeruginosa and biochemical content of groundnut plants
- Correlation analysis between semen routine parameters and sperm DNA fragmentation index in patients with semen non-liquefaction: A retrospective study
- Plasticity of the anatomical traits of Rhododendron L. (Ericaceae) leaves and its implications in adaptation to the plateau environment
- Effects of Piriformospora indica and arbuscular mycorrhizal fungus on growth and physiology of Moringa oleifera under low-temperature stress
- Effects of different sources of potassium fertiliser on yield, fruit quality and nutrient absorption in “Harward” kiwifruit (Actinidia deliciosa)
- Comparative efficiency and residue levels of spraying programs against powdery mildew in grape varieties
- The DREB7 transcription factor enhances salt tolerance in soybean plants under salt stress
- Using plant electrical signals of water hyacinth (Eichhornia crassipes) for water pollution monitoring
- Food Science
- Phytochemical analysis of Stachys iva: Discovering the optimal extract conditions and its bioactive compounds
- Review on role of honey in disease prevention and treatment through modulation of biological activities
- Computational analysis of polymorphic residues in maltose and maltotriose transporters of a wild Saccharomyces cerevisiae strain
- Optimization of phenolic compound extraction from Tunisian squash by-products: A sustainable approach for antioxidant and antibacterial applications
- Liupao tea aqueous extract alleviates dextran sulfate sodium-induced ulcerative colitis in rats by modulating the gut microbiota
- Toxicological qualities and detoxification trends of fruit by-products for valorization: A review
- Polyphenolic spectrum of cornelian cherry fruits and their health-promoting effect
- Optimizing the encapsulation of the refined extract of squash peels for functional food applications: A sustainable approach to reduce food waste
- Advancements in curcuminoid formulations: An update on bioavailability enhancement strategies curcuminoid bioavailability and formulations
- Impact of saline sprouting on antioxidant properties and bioactive compounds in chia seeds
- The dilemma of food genetics and improvement
- Bioengineering and Biotechnology
- Impact of hyaluronic acid-modified hafnium metalorganic frameworks containing rhynchophylline on Alzheimer’s disease
- Emerging patterns in nanoparticle-based therapeutic approaches for rheumatoid arthritis: A comprehensive bibliometric and visual analysis spanning two decades
- Application of CRISPR/Cas gene editing for infectious disease control in poultry
- Preparation of hafnium nitride-coated titanium implants by magnetron sputtering technology and evaluation of their antibacterial properties and biocompatibility
- Preparation and characterization of lemongrass oil nanoemulsion: Antimicrobial, antibiofilm, antioxidant, and anticancer activities
- Corrigendum
- Corrigendum to “Utilization of convolutional neural networks to analyze microscopic images for high-throughput screening of mesenchymal stem cells”
- Corrigendum to “Effects of Ire1 gene on virulence and pathogenicity of Candida albicans”
- Retraction
- Retraction of “Down-regulation of miR-539 indicates poor prognosis in patients with pancreatic cancer”
Articles in the same Issue
- Biomedical Sciences
- Mechanism of triptolide regulating proliferation and apoptosis of hepatoma cells by inhibiting JAK/STAT pathway
- Maslinic acid improves mitochondrial function and inhibits oxidative stress and autophagy in human gastric smooth muscle cells
- Comparative analysis of inflammatory biomarkers for the diagnosis of neonatal sepsis: IL-6, IL-8, SAA, CRP, and PCT
- Post-pandemic insights on COVID-19 and premature ovarian insufficiency
- Proteome differences of dental stem cells between permanent and deciduous teeth by data-independent acquisition proteomics
- Optimizing a modified cetyltrimethylammonium bromide protocol for fungal DNA extraction: Insights from multilocus gene amplification
- Preliminary analysis of the role of small hepatitis B surface proteins mutations in the pathogenesis of occult hepatitis B infection via the endoplasmic reticulum stress-induced UPR-ERAD pathway
- Efficacy of alginate-coated gold nanoparticles against antibiotics-resistant Staphylococcus and Streptococcus pathogens of acne origins
- Battling COVID-19 leveraging nanobiotechnology: Gold and silver nanoparticle–B-escin conjugates as SARS-CoV-2 inhibitors
- Neurodegenerative diseases and neuroinflammation-induced apoptosis
- Impact of fracture fixation surgery on cognitive function and the gut microbiota in mice with a history of stroke
- COLEC10: A potential tumor suppressor and prognostic biomarker in hepatocellular carcinoma through modulation of EMT and PI3K-AKT pathways
- High-temperature requirement serine protease A2 inhibitor UCF-101 ameliorates damaged neurons in traumatic brain-injured rats by the AMPK/NF-κB pathway
- SIK1 inhibits IL-1β-stimulated cartilage apoptosis and inflammation in vitro through the CRTC2/CREB1 signaling
- Rutin–chitooligosaccharide complex: Comprehensive evaluation of its anti-inflammatory and analgesic properties in vitro and in vivo
- Knockdown of Aurora kinase B alleviates high glucose-triggered trophoblast cells damage and inflammation during gestational diabetes
- Calcium-sensing receptors promoted Homer1 expression and osteogenic differentiation in bone marrow mesenchymal stem cells
- ABI3BP can inhibit the proliferation, invasion, and epithelial–mesenchymal transition of non-small-cell lung cancer cells
- Changes in blood glucose and metabolism in hyperuricemia mice
- Rapid detection of the GJB2 c.235delC mutation based on CRISPR-Cas13a combined with lateral flow dipstick
- IL-11 promotes Ang II-induced autophagy inhibition and mitochondrial dysfunction in atrial fibroblasts
- Short-chain fatty acid attenuates intestinal inflammation by regulation of gut microbial composition in antibiotic-associated diarrhea
- Application of metagenomic next-generation sequencing in the diagnosis of pathogens in patients with diabetes complicated by community-acquired pneumonia
- NAT10 promotes radiotherapy resistance in non-small cell lung cancer by regulating KPNB1-mediated PD-L1 nuclear translocation
- Phytol-mixed micelles alleviate dexamethasone-induced osteoporosis in zebrafish: Activation of the MMP3–OPN–MAPK pathway-mediating bone remodeling
- Association between TGF-β1 and β-catenin expression in the vaginal wall of patients with pelvic organ prolapse
- Primary pleomorphic liposarcoma involving bilateral ovaries: Case report and literature review
- Effects of de novo donor-specific Class I and II antibodies on graft outcomes after liver transplantation: A pilot cohort study
- Sleep architecture in Alzheimer’s disease continuum: The deep sleep question
- Ephedra fragilis plant extract: A groundbreaking corrosion inhibitor for mild steel in acidic environments – electrochemical, EDX, DFT, and Monte Carlo studies
- Langerhans cell histiocytosis in an adult patient with upper jaw and pulmonary involvement: A case report
- Inhibition of mast cell activation by Jaranol-targeted Pirin ameliorates allergic responses in mouse allergic rhinitis
- Aeromonas veronii-induced septic arthritis of the hip in a child with acute lymphoblastic leukemia
- Clusterin activates the heat shock response via the PI3K/Akt pathway to protect cardiomyocytes from high-temperature-induced apoptosis
- Research progress on fecal microbiota transplantation in tumor prevention and treatment
- Low-pressure exposure influences the development of HAPE
- Stigmasterol alleviates endplate chondrocyte degeneration through inducing mitophagy by enhancing PINK1 mRNA acetylation via the ESR1/NAT10 axis
- AKAP12, mediated by transcription factor 21, inhibits cell proliferation, metastasis, and glycolysis in lung squamous cell carcinoma
- Association between PAX9 or MSX1 gene polymorphism and tooth agenesis risk: A meta-analysis
- A case of bloodstream infection caused by Neisseria gonorrhoeae
- Case of nasopharyngeal tuberculosis complicated with cervical lymph node and pulmonary tuberculosis
- p-Cymene inhibits pro-fibrotic and inflammatory mediators to prevent hepatic dysfunction
- GFPT2 promotes paclitaxel resistance in epithelial ovarian cancer cells via activating NF-κB signaling pathway
- Transfer RNA-derived fragment tRF-36 modulates varicose vein progression via human vascular smooth muscle cell Notch signaling
- RTA-408 attenuates the hepatic ischemia reperfusion injury in mice possibly by activating the Nrf2/HO-1 signaling pathway
- Decreased serum TIMP4 levels in patients with rheumatoid arthritis
- Sirt1 protects lupus nephritis by inhibiting the NLRP3 signaling pathway in human glomerular mesangial cells
- Sodium butyrate aids brain injury repair in neonatal rats
- Interaction of MTHFR polymorphism with PAX1 methylation in cervical cancer
- Convallatoxin inhibits proliferation and angiogenesis of glioma cells via regulating JAK/STAT3 pathway
- The effect of the PKR inhibitor, 2-aminopurine, on the replication of influenza A virus, and segment 8 mRNA splicing
- Effects of Ire1 gene on virulence and pathogenicity of Candida albicans
- Small cell lung cancer with small intestinal metastasis: Case report and literature review
- GRB14: A prognostic biomarker driving tumor progression in gastric cancer through the PI3K/AKT signaling pathway by interacting with COBLL1
- 15-Lipoxygenase-2 deficiency induces foam cell formation that can be restored by salidroside through the inhibition of arachidonic acid effects
- FTO alleviated the diabetic nephropathy progression by regulating the N6-methyladenosine levels of DACT1
- Clinical relevance of inflammatory markers in the evaluation of severity of ulcerative colitis: A retrospective study
- Zinc valproic acid complex promotes osteoblast differentiation and exhibits anti-osteoporotic potential
- Primary pulmonary synovial sarcoma in the bronchial cavity: A case report
- Metagenomic next-generation sequencing of alveolar lavage fluid improves the detection of pulmonary infection
- Uterine tumor resembling ovarian sex cord tumor with extensive rhabdoid differentiation: A case report
- Genomic analysis of a novel ST11(PR34365) Clostridioides difficile strain isolated from the human fecal of a CDI patient in Guizhou, China
- Effects of tiered cardiac rehabilitation on CRP, TNF-α, and physical endurance in older adults with coronary heart disease
- Changes in T-lymphocyte subpopulations in patients with colorectal cancer before and after acupoint catgut embedding acupuncture observation
- Modulating the tumor microenvironment: The role of traditional Chinese medicine in improving lung cancer treatment
- Alterations of metabolites related to microbiota–gut–brain axis in plasma of colon cancer, esophageal cancer, stomach cancer, and lung cancer patients
- Research on individualized drug sensitivity detection technology based on bio-3D printing technology for precision treatment of gastrointestinal stromal tumors
- CEBPB promotes ulcerative colitis-associated colorectal cancer by stimulating tumor growth and activating the NF-κB/STAT3 signaling pathway
- Oncolytic bacteria: A revolutionary approach to cancer therapy
- A de novo meningioma with rapid growth: A possible malignancy imposter?
- Diagnosis of secondary tuberculosis infection in an asymptomatic elderly with cancer using next-generation sequencing: Case report
- Hesperidin and its zinc(ii) complex enhance osteoblast differentiation and bone formation: In vitro and in vivo evaluations
- Research progress on the regulation of autophagy in cardiovascular diseases by chemokines
- Anti-arthritic, immunomodulatory, and inflammatory regulation by the benzimidazole derivative BMZ-AD: Insights from an FCA-induced rat model
- Immunoassay for pyruvate kinase M1/2 as an Alzheimer’s biomarker in CSF
- The role of HDAC11 in age-related hearing loss: Mechanisms and therapeutic implications
- Evaluation and application analysis of animal models of PIPNP based on data mining
- Therapeutic approaches for liver fibrosis/cirrhosis by targeting pyroptosis
- Fabrication of zinc oxide nanoparticles using Ruellia tuberosa leaf extract induces apoptosis through P53 and STAT3 signalling pathways in prostate cancer cells
- Haplo-hematopoietic stem cell transplantation and immunoradiotherapy for severe aplastic anemia complicated with nasopharyngeal carcinoma: A case report
- Modulation of the KEAP1-NRF2 pathway by Erianin: A novel approach to reduce psoriasiform inflammation and inflammatory signaling
- The expression of epidermal growth factor receptor 2 and its relationship with tumor-infiltrating lymphocytes and clinical pathological features in breast cancer patients
- Innovations in MALDI-TOF Mass Spectrometry: Bridging modern diagnostics and historical insights
- BAP1 complexes with YY1 and RBBP7 and its downstream targets in ccRCC cells
- Hypereosinophilic syndrome with elevated IgG4 and T-cell clonality: A report of two cases
- Electroacupuncture alleviates sciatic nerve injury in sciatica rats by regulating BDNF and NGF levels, myelin sheath degradation, and autophagy
- Polydatin prevents cholesterol gallstone formation by regulating cholesterol metabolism via PPAR-γ signaling
- RNF144A and RNF144B: Important molecules for health
- Analysis of the detection rate and related factors of thyroid nodules in the healthy population
- Artesunate inhibits hepatocellular carcinoma cell migration and invasion through OGA-mediated O-GlcNAcylation of ZEB1
- Endovascular management of post-pancreatectomy hemorrhage caused by a hepatic artery pseudoaneurysm: Case report and review of the literature
- Efficacy and safety of anti-PD-1/PD-L1 antibodies in patients with relapsed refractory diffuse large B-cell lymphoma: A meta-analysis
- SATB2 promotes humeral fracture healing in rats by activating the PI3K/AKT pathway
- Overexpression of the ferroptosis-related gene, NFS1, corresponds to gastric cancer growth and tumor immune infiltration
- Understanding risk factors and prognosis in diabetic foot ulcers
- Atractylenolide I alleviates the experimental allergic response in mice by suppressing TLR4/NF-kB/NLRP3 signalling
- FBXO31 inhibits the stemness characteristics of CD147 (+) melanoma stem cells
- Immune molecule diagnostics in colorectal cancer: CCL2 and CXCL11
- Inhibiting CXCR6 promotes senescence of activated hepatic stellate cells with limited proinflammatory SASP to attenuate hepatic fibrosis
- Cadmium toxicity, health risk and its remediation using low-cost biochar adsorbents
- Pulmonary cryptococcosis with headache as the first presentation: A case report
- Solitary pulmonary metastasis with cystic airspaces in colon cancer: A rare case report
- RUNX1 promotes denervation-induced muscle atrophy by activating the JUNB/NF-κB pathway and driving M1 macrophage polarization
- Morphometric analysis and immunobiological investigation of Indigofera oblongifolia on the infected lung with Plasmodium chabaudi
- The NuA4/TIP60 histone-modifying complex and Hr78 modulate the Lobe2 mutant eye phenotype
- Experimental study on salmon demineralized bone matrix loaded with recombinant human bone morphogenetic protein-2: In vitro and in vivo study
- A case of IgA nephropathy treated with a combination of telitacicept and half-dose glucocorticoids
- Analgesic and toxicological evaluation of cannabidiol-rich Moroccan Cannabis sativa L. (Khardala variety) extract: Evidence from an in vivo and in silico study
- Wound healing and signaling pathways
- Combination of immunotherapy and whole-brain radiotherapy on prognosis of patients with multiple brain metastases: A retrospective cohort study
- To explore the relationship between endometrial hyperemia and polycystic ovary syndrome
- Research progress on the impact of curcumin on immune responses in breast cancer
- Biogenic Cu/Ni nanotherapeutics from Descurainia sophia (L.) Webb ex Prantl seeds for the treatment of lung cancer
- Dapagliflozin attenuates atrial fibrosis via the HMGB1/RAGE pathway in atrial fibrillation rats
- Glycitein alleviates inflammation and apoptosis in keratinocytes via ROS-associated PI3K–Akt signalling pathway
- ADH5 inhibits proliferation but promotes EMT in non-small cell lung cancer cell through activating Smad2/Smad3
- Apoptotic efficacies of AgNPs formulated by Syzygium aromaticum leaf extract on 32D-FLT3-ITD human leukemia cell line with PI3K/AKT/mTOR signaling pathway
- Novel cuproptosis-related genes C1QBP and PFKP identified as prognostic and therapeutic targets in lung adenocarcinoma
- Bee venom promotes exosome secretion and alters miRNA cargo in T cells
- Treatment of pure red cell aplasia in a chronic kidney disease patient with roxadustat: A case report
- Comparative bioinformatics analysis of the Wnt pathway in breast cancer: Selection of novel biomarker panels associated with ER status
- Kynurenine facilitates renal cell carcinoma progression by suppressing M2 macrophage pyroptosis through inhibition of CASP1 cleavage
- RFX5 promotes the growth, motility, and inhibits apoptosis of gastric adenocarcinoma cells through the SIRT1/AMPK axis
- ALKBH5 exacerbates early cardiac damage after radiotherapy for breast cancer via m6A demethylation of TLR4
- Phytochemicals of Roman chamomile: Antioxidant, anti-aging, and whitening activities of distillation residues
- Circadian gene Cry1 inhibits the tumorigenicity of hepatocellular carcinoma by the BAX/BCL2-mediated apoptosis pathway
- The TNFR-RIPK1/RIPK3 signalling pathway mediates the effect of lanthanum on necroptosis of nerve cells
- Longitudinal monitoring of autoantibody dynamics in patients with early-stage non-small-cell lung cancer undergoing surgery
- The potential role of rutin, a flavonoid, in the management of cancer through modulation of cell signaling pathways
- Construction of pectinase gene engineering microbe and its application in tobacco sheets
- Construction of a microbial abundance prognostic scoring model based on intratumoral microbial data for predicting the prognosis of lung squamous cell carcinoma
- Sepsis complicated by haemophagocytic lymphohistiocytosis triggered by methicillin-resistant Staphylococcus aureus and human herpesvirus 8 in an immunocompromised elderly patient: A case report
- Sarcopenia in liver transplantation: A comprehensive bibliometric study of current research trends and future directions
- Advances in cancer immunotherapy and future directions in personalized medicine
- Can coronavirus disease 2019 affect male fertility or cause spontaneous abortion? A two-sample Mendelian randomization analysis
- Heat stroke associated with novel leukaemia inhibitory factor receptor gene variant in a Chinese infant
- PSME2 exacerbates ulcerative colitis by disrupting intestinal barrier function and promoting autophagy-dependent inflammation
- Hyperosmolar hyperglycemic state with severe hypernatremia coexisting with central diabetes insipidus: A case report and literature review
- Efficacy and mechanism of escin in improving the tissue microenvironment of blood vessel walls via anti-inflammatory and anticoagulant effects: Implications for clinical practice
- Merkel cell carcinoma: Clinicopathological analysis of three patients and literature review
- Genetic variants in VWF exon 26 and their implications for type 1 Von Willebrand disease in a Saudi Arabian population
- Lipoxin A4 improves myocardial ischemia/reperfusion injury through the Notch1-Nrf2 signaling pathway
- High levels of EPHB2 expression predict a poor prognosis and promote tumor progression in endometrial cancer
- Knockdown of SHP-2 delays renal tubular epithelial cell injury in diabetic nephropathy by inhibiting NLRP3 inflammasome-mediated pyroptosis
- Exploring the toxicity mechanisms and detoxification methods of Rhizoma Paridis
- Concomitant gastric carcinoma and primary hepatic angiosarcoma in a patient: A case report
- Ecology and Environmental Science
- Optimization and comparative study of Bacillus consortia for cellulolytic potential and cellulase enzyme activity
- The complete mitochondrial genome analysis of Haemaphysalis hystricis Supino, 1897 (Ixodida: Ixodidae) and its phylogenetic implications
- Epidemiological characteristics and risk factors analysis of multidrug-resistant tuberculosis among tuberculosis population in Huzhou City, Eastern China
- Indices of human impacts on landscapes: How do they reflect the proportions of natural habitats?
- Genetic analysis of the Siberian flying squirrel population in the northern Changbai Mountains, Northeast China: Insights into population status and conservation
- Diversity and environmental drivers of Suillus communities in Pinus sylvestris var. mongolica forests of Inner Mongolia
- Global assessment of the fate of nitrogen deposition in forest ecosystems: Insights from 15N tracer studies
- Fungal and bacterial pathogenic co-infections mainly lead to the assembly of microbial community in tobacco stems
- Influencing of coal industry related airborne particulate matter on ocular surface tear film injury and inflammatory factor expression in Sprague-Dawley rats
- Temperature-dependent development, predation, and life table of Sphaerophoria macrogaster (Thomson) (Diptera: Syrphidae) feeding on Myzus persicae (Sulzer) (Homoptera: Aphididae)
- Eleonora’s falcon trophic interactions with insects within its breeding range: A systematic review
- Agriculture
- Integrated analysis of transcriptome, sRNAome, and degradome involved in the drought-response of maize Zhengdan958
- Variation in flower frost tolerance among seven apple cultivars and transcriptome response patterns in two contrastingly frost-tolerant selected cultivars
- Heritability of durable resistance to stripe rust in bread wheat (Triticum aestivum L.)
- Molecular mechanism of follicular development in laying hens based on the regulation of water metabolism
- Animal Science
- Effect of sex ratio on the life history traits of an important invasive species, Spodoptera frugiperda
- Plant Sciences
- Hairpin in a haystack: In silico identification and characterization of plant-conserved microRNA in Rafflesiaceae
- Widely targeted metabolomics of different tissues in Rubus corchorifolius
- The complete chloroplast genome of Gerbera piloselloides (L.) Cass., 1820 (Carduoideae, Asteraceae) and its phylogenetic analysis
- Field trial to correlate mineral solubilization activity of Pseudomonas aeruginosa and biochemical content of groundnut plants
- Correlation analysis between semen routine parameters and sperm DNA fragmentation index in patients with semen non-liquefaction: A retrospective study
- Plasticity of the anatomical traits of Rhododendron L. (Ericaceae) leaves and its implications in adaptation to the plateau environment
- Effects of Piriformospora indica and arbuscular mycorrhizal fungus on growth and physiology of Moringa oleifera under low-temperature stress
- Effects of different sources of potassium fertiliser on yield, fruit quality and nutrient absorption in “Harward” kiwifruit (Actinidia deliciosa)
- Comparative efficiency and residue levels of spraying programs against powdery mildew in grape varieties
- The DREB7 transcription factor enhances salt tolerance in soybean plants under salt stress
- Using plant electrical signals of water hyacinth (Eichhornia crassipes) for water pollution monitoring
- Food Science
- Phytochemical analysis of Stachys iva: Discovering the optimal extract conditions and its bioactive compounds
- Review on role of honey in disease prevention and treatment through modulation of biological activities
- Computational analysis of polymorphic residues in maltose and maltotriose transporters of a wild Saccharomyces cerevisiae strain
- Optimization of phenolic compound extraction from Tunisian squash by-products: A sustainable approach for antioxidant and antibacterial applications
- Liupao tea aqueous extract alleviates dextran sulfate sodium-induced ulcerative colitis in rats by modulating the gut microbiota
- Toxicological qualities and detoxification trends of fruit by-products for valorization: A review
- Polyphenolic spectrum of cornelian cherry fruits and their health-promoting effect
- Optimizing the encapsulation of the refined extract of squash peels for functional food applications: A sustainable approach to reduce food waste
- Advancements in curcuminoid formulations: An update on bioavailability enhancement strategies curcuminoid bioavailability and formulations
- Impact of saline sprouting on antioxidant properties and bioactive compounds in chia seeds
- The dilemma of food genetics and improvement
- Bioengineering and Biotechnology
- Impact of hyaluronic acid-modified hafnium metalorganic frameworks containing rhynchophylline on Alzheimer’s disease
- Emerging patterns in nanoparticle-based therapeutic approaches for rheumatoid arthritis: A comprehensive bibliometric and visual analysis spanning two decades
- Application of CRISPR/Cas gene editing for infectious disease control in poultry
- Preparation of hafnium nitride-coated titanium implants by magnetron sputtering technology and evaluation of their antibacterial properties and biocompatibility
- Preparation and characterization of lemongrass oil nanoemulsion: Antimicrobial, antibiofilm, antioxidant, and anticancer activities
- Corrigendum
- Corrigendum to “Utilization of convolutional neural networks to analyze microscopic images for high-throughput screening of mesenchymal stem cells”
- Corrigendum to “Effects of Ire1 gene on virulence and pathogenicity of Candida albicans”
- Retraction
- Retraction of “Down-regulation of miR-539 indicates poor prognosis in patients with pancreatic cancer”