Circ-UBR4 regulates the proliferation, migration, inflammation, and apoptosis in ox-LDL-induced vascular smooth muscle cells via miR-515-5p/IGF2 axis
-
Liuliu Feng


Abstract
The aim of our study is to disclose the role and underlying molecular mechanisms of circular RNA ubiquitin protein ligase E3 component n-recognin 4 (circ-UBR4) in atherosclerosis (AS). Our data showed that circ-UBR4 expression was upregulated in AS patients and oxidized low-density lipoprotein (ox-LDL)-induced vascular smooth muscle cells (VSMCs) compared with healthy volunteer and untreated VSMCs. In addition, ox-LDL stimulated proliferation, migration, and inflammation but decreased apoptosis in VSMCs, which were overturned by the inhibition of circ-UBR4. miR-515-5p was sponged by circ-UBR4, and its inhibitor reversed the inhibitory effect of circ-UBR4 knockdown on proliferation, migration, and inflammation in ox-LDL-induced VSMCs. Insulin-like growth factor2 (IGF2) was a functional target of miR-515-5p, and overexpression of IGF2 reversed the suppressive effect of miR-515-5p on ox-LDL-stimulated VSMCs proliferation, migration, and inflammation. Collectively, circ-UBR4 knockdown decreased proliferation, migration, and inflammation but stimulated apoptosis in ox-LDL-induced VSMCs by targeting the miR-515-5p/IGF2 axis.
1 Introduction
Atherosclerosis (AS), an inflammatory disease, is a crucial pathogenic factor of most cardiovascular and cerebrovascular diseases, characterized by lipid metabolism disorder and chronic inflammation [1,2]. AS is a multistep disease and can be induced by various risk factors [3]. The dysfunction and inflammation of vascular smooth muscle cells (VSMCs) are closely related to AS progression [4], and oxidized low-density lipoprotein (ox-LDL)-induced VSMCs’ injury are often used to construct AS models in vitro [5,6]. Therefore, elucidation of the molecular mechanism affecting ox-LDL induced VSMCs injury is expected to provide potential molecular targets for the treatment of AS.
Circular RNAs (circRNAs) are novel group of endogenous non-coding RNAs with a circular structure generated by back-splicing [7]. Previous research confirmed that circRNAs played significant functions in AS progression and served as independent markers for AS diagnosis [8]. For example, the inhibition of circCHFR impeded the cell growth and mobility of VSMCs through microRNA (miRNA)-mRNA network [9]. Hsa_circ_0010283 is derived from the ubiquitin protein ligase E3 component n-recognin 4 (UBR4) gene, also named as circ-UBR4. Circ-UBR4 was initially manifested to be highly expressed in ox-LDL-treated VSMCs by circRNA microarray analysis [9], and the functional effects of circ-UBR4 on ox-LDL-treated VSMC activities were partly elucidated [10,11]. However, the regulatory mechanisms of circ-UBR4 in AS progression are complex and have not been fully uncovered. Therefore, circ-UBR4 was selected as a target in our study to explore its role and molecular mechanism in ox-LDL-induced VSMCs injury, providing more evidence for it to be a potential target in AS therapy.
miRNAs are non-coding and single-strand RNAs (18–25 nucleotides) and play vital roles in multiple pathological processes [12]. Mechanistically, miRNAs have been confirmed to function inhibitory effects on gene expression by targeting 3′untranslational region (UTR) of their target genes [13]. Abundant reports discovered that miR-515-5p acted as a tumor-suppressor in numerous tumors [14,15,16]. We guessed that miR-515-5p also played significant roles in AS progression.
Insulin-like growth factor2 (IGF2) is a member of insulin-like growth factor (IGF) family [17]. The potential association between IGF family and hematological malignancies was revealed in a previous report [18]. Interestingly, Sun et al. also revealed that ox-LDL-induced upregulation of IGF2 could activate downstream IGF-related pathways, thereby enhancing the proliferation of VSMCs [19]. Therefore, the roles and functional effects of IGF2 were explored in AS progression.
In this research, ox-LDL-stimulated VSMCs were used as cell models of AS. Through bioinformatics analysis, we found that circ-UBR4 had complementary binding sites for miR-515-5p, and miR-515-5p could target IGF2. Therefore, we determined the role of circ-UBR4 and investigated the interplays among circ-UBR4, miR-515-5p, and IGF2 in ox-LDL-induced VSMCs.
2 Materials and methods
2.1 Serum samples
The serum samples were collected from 31 AS patients and 25 healthy normal controls in Shanghai Shidong hospital. The written informed consent was acquired from each patient and volunteer, and our study was approved by the Ethics Committee of Shanghai Shidong hospital.
2.2 Cell culture
T/G HA-VSMCs (derived from Aorta) were purchased from ATCC (catalogue: PCS-100-012; Manassas, VA, USA) and cultured in Dulbecco’s modified eagle medium (GIBCO BRL, Grand Island, NY, USA) containing 10% (v/v) fetal bovine serum (FBS; Thermo Fisher Scientific, Carlsbad, CA, USA) at 37°C with 5% CO2. The medium was replaced for every 2 or 3 days. Cells between passages 3 and 6 were employed in this study. To establish cell models of AS, VSMCs were treated with ox-LDL (Thermo Fisher Scientific) with different concentrations for 24 h or 50 μg/mL for different times. In function experiments, VSMCs were treated with 50 μg/mL of ox-LDL for 24 h after transfection for 24 h.
2.3 Cell transfection
Short interfering RNA (siRNA) targeting circ-UBR4 (si-circ-UBR4), control (si-NC), circ-UBR4 overexpression vector (circ-UBR4), empty vector (pCD5-ciR), IGF2-overexpression vector (IGF2), and empty vector (pcDNA) were designed and optimized by GenePharma (Shanghai, China). MiR-515-5p mimic (miR-515-5p), miR-NC, miR-515-5p inhibitor (anti-miR-515-5p), and anti-miR-NC were synthesized by Sangon (Shanghai, China). For cell transfection, VSMCs were seeded into 24-well plates (5 × 104 cells/well) and then transfected with plasmids (2 μg), miRNA oligonucleotides (50 nM), or siRNA (50 nM) by Lipofectamine 2000 reagent (Thermo Fisher Scientific) referring to the recommended protocol.
2.4 Real-time quantitative PCR (RT-qPCR)
Total RNAs were isolated by TRIzol reagent (Invitrogen, Carlsbad, CA, USA) in compliance with the manufacturer’s direction. After quantifying under the Nanodrop 2000c (Thermo Fisher Scientific), the extracted RNA was used to transcribe into complementary DNA (cDNA) by First-Strand cDNA Synthesis SuperMix (CapitalBio, Beijing, China). RT-qPCR reactions were performed by SYBR Green Real-Time PCR Master Mix (Thermo Fisher Scientific) under the Roche LightCycler (Roche, Basel, Switzerland). The comparative threshold cycle (Ct) method was carried out to evaluate the relative expression of target RNAs, with glyceraldehyde-3-phosphate dehydrogenase (GAPDH) or nuclear RNA U6 as the control. The primer sequences were as follows: circ-UBR4, 5′-AGTGTGGTTACAGCCAGCTC-3′ (forward) and 5′-ACCATAACTACCAGCGGCAC-3′ (reverse); UBR4, 5′-CCCCGGAACCAACTTCAGTC-3′ (forward) and 5′-TTGGCGGATTTCATCATTGCT-3′ (reverse); miR-515-5p, 5′-GCCGAGTTCTCCAAAAGAAAGC-3′ (forward) and 5′-CAGTGCAGGGTCCGAGGTAT-3′ (reverse); IGF2, 5′-ACGAAATATCCCGCCTCATTTAC-3′ (forward) and 5′-GCAGTTTCCGAGTCAGTGTTCA-3′ (reverse); U6, 5′-CTCGCTTCGGCAGCACA-3′ (forward) and 5′-AACGCTTCACGAATTTGCGT-3′ (reverse); and GAPDH, 5′-TGAACCATGAGAAGTATGAC-3′ (forward) and 5′-TCTTACTCCTTGGAGGCCA-3′ (reverse).
2.5 RNase R treatment and nuclear-cytoplasmic fractionation
Purified RNAs were incubated with RNase R (Epicentre Technologies, Madison, USA), followed by purification with Trizol (Invitrogen).
In addition, Cytoplasmic and Nuclear RNA Purification Kit was used for nuclear-cytoplasmic fractionation assay. Briefly, ice-cold lysis buffer J was added into culture plate to lyse VSMCs. After centrifuging, supernatant was collected as cytoplasmic RNA. The precipitate containing the nuclear RNA was incubated with buffer SK. Finally, cytoplasmic RNA and nuclear RNA were eluted with elution buffer E.
2.6 MTT assay
VSMCs were planted into 96-well plates (5,000 cells per well) and cultured at 37°C with 5% CO2. After incubation for 24 h, 20 μL of MTT solution (5 mg/mL; Thermo Fisher Scientific) was added into each well at 37°C, and cells were allowed to incubate for another 4 h. The formatted crystal formazan was dissolved by dimethyl sulfoxide (Thermo Fisher Scientific). Microplate reader was used to assess optical density value at 490 nm.
2.7 EdU assay
Cell proliferation was also evaluated by EdU incorporation assay using an EdU Apollo DNA in vitro kit (RiboBio, Guangzhou, China). Briefly, cells after transfection were incubated in 96-well plates (5,000 cells per well) for 24 h at 37°C and next subjected with 100 μL of 50 μM EdU per well. After culturing cells with EdU for 12 h, cells were fixed and then counterstained with DAPI. The EdU staining was observed via a fluorescence microscopy (Mshot, Guangdong, China).
2.8 Flow cytometry assay
Apoptosis of VSMCs was assessed by Annexin V-FITC Apoptosis Detection Kit (BD Pharmingen, Franklin Lakes, NJ, USA). After incubation for 24 h, transfected VSMCs were harvested as a single cell suspension (1 × 106/mL) by trypsin digestion. The staining buffer containing Annexin V-FITC and propidium iodide was added to incubate VSMCs at 4°C for 30 min. The Flow Cytometer (Beckman Coulter, Miami, FL, USA) was used for apoptosis assay.
2.9 Transwell assay
For in vitro cell migration assay, 24-well transwell chamber (BD Pharmingen, San Jose, CA, USA) was used. VSMCs resuspended in 200 µL medium without FBS (5 × 104 cells/well) were seeded into the upper compartment, while complete medium was used as nutrients to induce cell migration. After 24 h, the remaining culture medium and the cells that did not migrate were removed carefully, while the migrated cells were fixed and then stained with 0.1% crystal violet (Thermo Fisher Scientific). The migrated cells were imaged under a microscope (100× amplification; Mshot). The number of migrated cells was counted in five randomly selected regions.
2.10 Enzyme-linked immunosorbent assay (ELISA)
Cell culture supernatants were harvested by centrifugation at 1,000 × g for 10 min. The ELISA kit (Invitrogen; #BMS223INST; #BMS213HS; and #KAC1211) was used to detect the levels of tumor necrosis factor-α (TNF-α), Interleukin-6 (IL-6), and Interleukin-1β (IL-1β) according to the commodity instruction. The concentrations of TNF-α, IL-6, and IL-1β were obtained based on standard curve.
2.11 Western blot assay
Briefly, a protein extraction kit (Applygen Technologies, Beijing, China) was used to extract protein from VSMCs. Protein concentration was assessed by bicinchoninic acid protein assay (Applygen Technologies). 30 μg of protein was fractionated by sodium dodecyl sulfate-polyacrylamide gels and electroblotted onto nitrocellulose membranes (Bio-Rad, Hercules, CA, USA). The membranes were incubated in 4% skim milk solution and then reacted with antibodies at 4°C overnight, including anti-β-actin (ab8226, Abcam, Cambridge, MA, USA), anti-CyclinD1 (ab16633, Abcam), anti-matrix metallopeptidase 9 (MMP9; ab76003, Abcam), anti-BCL2-Associated X (Bax; ab325033, Abcam), anti-B-cell lymphoma-2 (Bcl-2; ab196495, Abcam), and anti-IGF2 (ab177467, Abcam). After washing with Tris-buffered saline with Tween 20, the membranes were incubated with HRP-conjugated secondary antibodies (Abcam). Finally, the Alpha Innotech Imaging System (ProteinSimple, Santa Clara, CA, USA) was used to visualize protein signal. The band density was analyzed using Image J software (NIH, Bethesda, MA, USA).
2.12 Dual-luciferase reporter assay
The downstream targets of circ-UBR4 and miR-515-5p were predicted by circRNA interactome (https://circinteractome.irp.nia.nih.gov/) and Starbase (http://starbase.sysu.edu.cn/), respectively. The circ-UBR4 sequences containing miR-515-5p binding sites were inserted into pmirGLO luciferase vector (GeneCreat, Wuhan, China), named as WT-circ-UBR4, with MUT-circ-UBR4 as control. Similarly, the wild- and mutant-type of IGF2 luciferase reporter vector (WT-IGF2 3′UTR and MUT-IGF2 3′UTR) were synthesized by GeneCreat. VSMCs were transfected with luciferase reporter vectors in the presence or absence of miR-515-5p by Lipofectamine 2000 (Thermo Fisher Scientific). Relative luciferase activity was assessed under the VICTOR2 fluorometry (PerkinElmer, Waltham, MA, USA) at 48 h post-transfection.
2.13 RNA immunoprecipitation (RIP) assay
RIP assay was carried out using Imprint® RIP kit (Sigma, Louis, MO, USA) according to manufacturer’s instruction. For RIP assay, VSMCs were lysed in RIP lysis buffer, followed by incubation with magnetic beads embracing Ago2 (Millipore, Billerica, MA, USA) or IgG (Millipore) antibodies for 24 h at 4°C. The level of RNA enriched by RIP was assessed by RT-qPCR.
2.14 Pull-down assay
miR-515-5p probe and miR-NC probe were labeled with biotin by RiboBio (Bio-miR-515-5p and Bio-miR-NC). VSMCs were transfected with Bio-miR-515-5p and Bio-miR-NC and subsequently lysed by lysis reagent (Thermo Fisher Scientific). Cell lysates were cultured with Streptavidin-Dynabeads (Thermo Fisher Scientific). RNA complexes pulled down by Bio-miR-515-5p and Bio-miR-NC were captured by beads. RNA samples were washed from beads and analyzed by RT-qPCR.
2.15 Statistical analysis
For each experiment, we set three duplications in adjacent three wells of 96- or 24-well plates. A total of three independent experiments were performed at few days interval using the same batch of frozen cells at passages 3–6. All quantitative data were displayed as mean value ± standard deviation. SPSS 21.0 software (IBM, Somers, NY, USA) was used to process data and analyze differences. The statistical differences in different groups were analyzed by Student’s t-test or analysis of variance (followed by Tukey’s post-hoc test). P-value less than 0.05 was considered to be statistically significant.
3 Results
3.1 circ-UBR4 was overexpressed in ox-LDL-stimulated VSMCs
Schematic diagram in Figure 1a manifested the information of circ-UBR4, showing that circ-UBR4 was produced from UBR4 gene (NM_020765), with 3358 bp in length. We detected circ-UBR4 expression in the serum of AS patients or healthy normal controls and confirmed that circ-UBR4 was overexpressed in AS patients (Figure A1). In order to analyze the potential role of circ-UBR4 in AS progression, we used ox-LDL-stimulated VSMCs as cell models of AS in vitro. As presented in Figure 1b, circ-UBR4 was significantly and dose-dependently increased in ox-LDL-stimulated VSMCs. Also, treatment with 50 μg/mL of ox-LDL significantly enhanced the expression of circ-UBR4 in VSMCs in a time-dependent manner (Figure 1c). RT-qPCR assay suggested the predominant cytoplasmic distribution of circ-UBR4 in VSMCs (Figure 1d). Furthermore, circ-UBR4 could resist the digestion of RNase R when compared with linear-UBR4 (Figure 1e). Therefore, the function of circ-UBR4 was explored in ox-LDL-stimulated VSMCs.

The expression level of circ-UBR4 in ox-LDL-stimulated VSMCs. (a) The information of circ-UBR4 structure and formation. (b) and (c) The relative expression level of circ-UBR4 was evaluated by RT-qPCR in ox-LDL-stimulated VSMCs. (d) The expression levels of circ-UBR4, GAPDH, and U6 were assessed by RT-qPCR in cytoplasmic and nuclear fraction RNAs. (e) RT-qPCR was used to show the expression of linear-UBR4 and circ-UBR4 in VSMCs after treatment with RNase R. ***P < 0.001.
3.2 Knockdown of circ-UBR4 reversed ox-LDL-stimulated VSMCs functions
As circ-UBR4 was overexpressed in ox-LDL-stimulated VSMCs, we next investigated the functional effects of circ-UBR4 inhibition in ox-LDL-stimulated VSMCs. Treatment with 50 μg/mL of ox-LDL enhanced the expression of circ-UBR4 in VSMCs, which was abolished by transfection with si-circ-UBR4 (Figure 2a). The results of MTT assay revealed that cell viability increased by ox-LDL was obviously inhibited by transfection with si-circ-UBR4 (Figure 2b). Similarly, knockdown of circ-UBR4 suppressed the inflammation in ox-LDL-stimulated VSMCs by reducing TNF-α, IL-6, and IL-1β levels (Figure 2c–e). Besides, ox-LDL-aggravated EdU incorporation and the number of migrated cells in VSMCs were largely repressed by circ-UBR4 downregulation (Figure 2f and g). Treatment with ox-LDL could inhibit cell apoptosis, while this effect was reversed by the knockdown of circ-UBR4 (Figure 2h). The results of western blot assay indicated that MMP9, CyclinD1, and Bcl-2 were upregulated, while Bax was downregulated in ox-LDL-stimulated VSMCs. However, the alterations induced by ox-LDL were all reversed by knockdown of circ-UBR4 (Figure 2i). Therefore, knockdown of circ-UBR4 inhibited cell proliferation, migration, and inflammation but promoted apoptosis in ox-LDL-stimulated VSMCs.
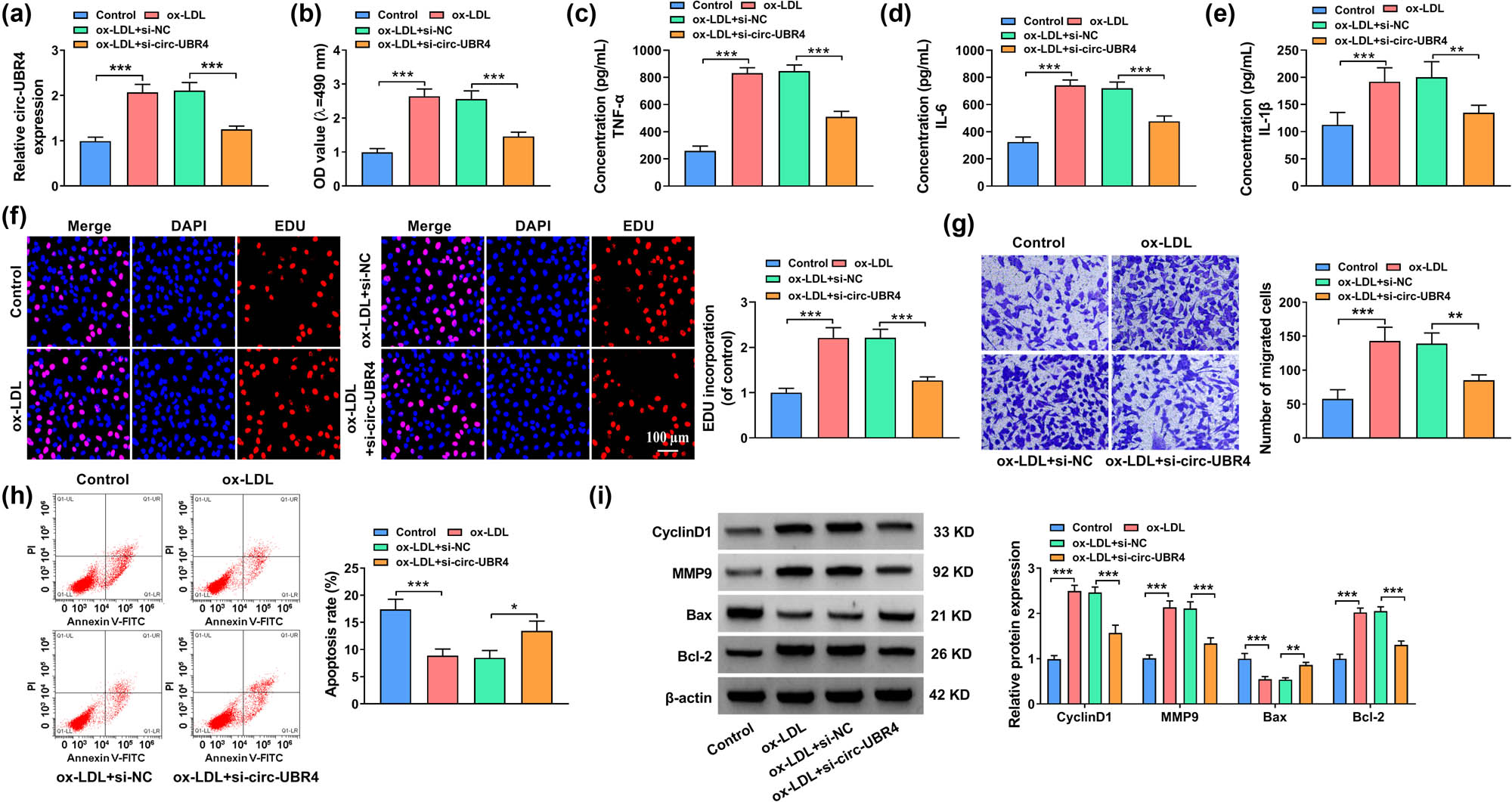
Inhibition of circ-UBR4 regulated proliferation, apoptosis, migration, and inflammation of ox-LDL-stimulated VSMCs. (a)–(i) VSMCs were divided into four groups: Control, ox-LDL, ox-LDL + si-NC, and ox-LDL + si-circ-UBR4. Un-treated VSMCs were used as Control. (a) The expression of circ-UBR4 was determined by RT-qPCR. (b) MTT assay was used for examining cell viability. (c)–(e) The levels of TNF-α, IL-6, and IL-1β were measured by ELISA. (f) EdU assay was used to examine cell proliferation. (g) Transwell assay was used to assess VSMCs migration. (h) The apoptosis of VSMCs was assessed by flow cytometry. (i) The protein expression levels of MMP9, CyclinD1, Bax, and Bcl-2 were quantified by western blot assay. *P < 0.05, **P < 0.01, ***P < 0.001.
3.3 miR-515-5p was a direct target of circ-UBR4
The circRNA interactome software predicted that circ-UBR4 had more targeted miRNAs. Through literature research, four miRNAs (miR-144-3p, miR-326, miR-370-3p, and miR-515-5p) with low expression in AS and having an inhibition on proliferation and metastasis of VSMCs were selected for RT-qPCR analysis. The results show that circ-UBR4 knockdown could affect the expression of multiple miRNAs, but si-circ-UBR4 had the most obvious promotion effect on miR-515-5p expression (Figure A2). Therefore, miR-515-5p was selected as the target of circ-UBR4 for this study. The possible complementary sequences between circ-UBR4 and miR-515-5p are presented in Figure 3a. The upregulation of miR-515-5p led to great loss of luciferase activity in WT-circ-UBR4 group, while luciferase activity in MUT-circ-UBR4 group was not affected by miR-515-5p overexpression (Figure 3b). In RIP assay, the immunopurification of Ago2 can be performed to confirm the interaction between miRNA and target genes by detecting RNA enrichment. The results of RIP assay revealed that circ-UBR4 and miR-515-5p were all enriched by Ago2 (Figure 3c). Additionally, pull-down assay showed that high abundance of circ-UBR4 could be pulled down by Bio-miR-515-5p probe (Figure 3d). The above data confirmed the interaction between circ-UBR4 and miR-515-5p. MiR-515-5p expression was inhibited by ox-LDL in VSMCs in a dose-dependent manner and time-dependent manner (Figure 3e–f).
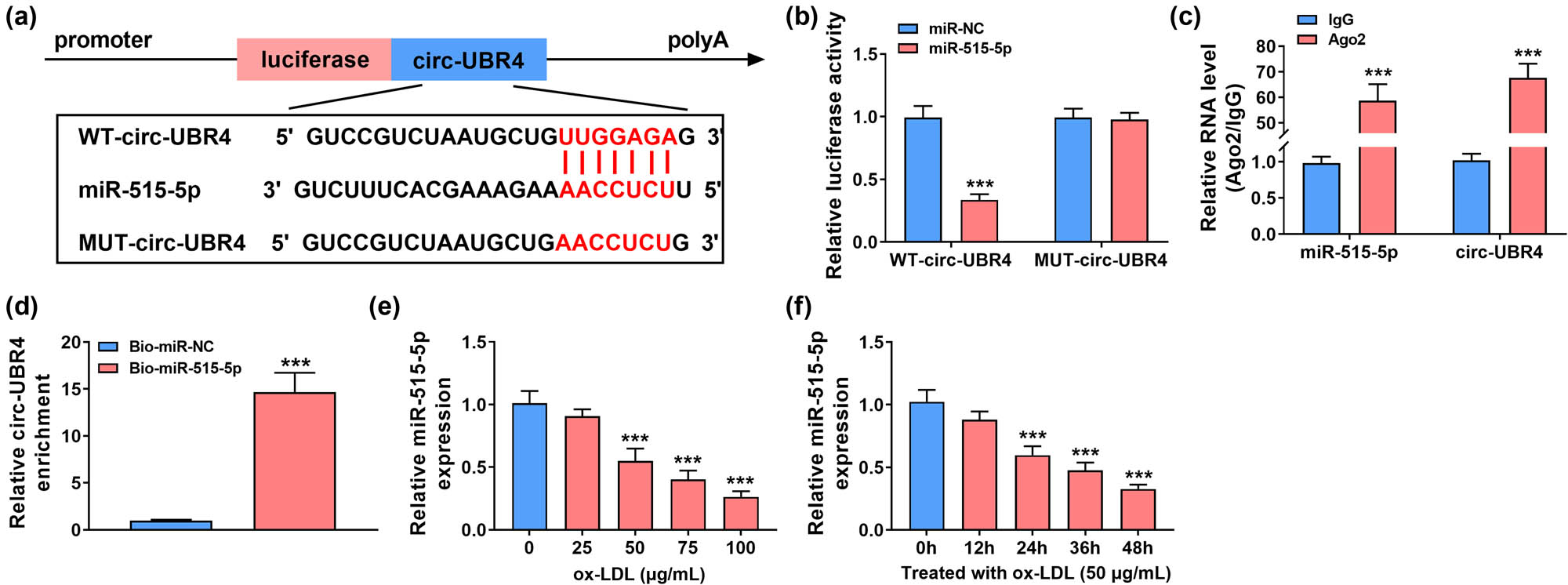
Circ-UBR4 regulated miR-515-5p expression in VSMCs. (a) Schematic diagram presented the complementary sequences between circ-UBR4 and miR-515-5p. (b)–(d) The potential association between circ-UBR4 and miR-515-5p was analyzed by dual-luciferase reporter, RIP, and pull-down assays. (e and f) RT-qPCR assay was performed to determine expression level of miR-515-5p in ox-LDL-stimulated VSMCs. ***P < 0.001.
3.4 miR-515-5p inhibitor reversed the effect of circ-UBR4 knockdown on ox-LDL-induced VSMCs functions
The association between circ-UBR4 and miR-515-5p was investigated in ox-LDL-induced VSMCs. Transfection with anti-miR-515-5p abolished the upregulation of miR-515-5p expression in si-circ-UBR4-transfected VSMCs (Figure 4a). Inhibition of miR-515-5p could rescue the inhibition effect of si-circ-UBR4 on the viability, inflammation factor levels, proliferation, and migration of ox-LDL-treated VSMCs (Figure 4b–g). In addition, the downregulation of miR-515-5p protected VSMCs from si-circ-UBR4-induced apoptosis in ox-LDL-treated VSMCs (Figure 4h). MiR-515-5p knockdown also restored the expression of CyclinD1, MMP9, and Bcl-2 inhibited by circ-UBR4 silencing, and repressed the expression of Bax promoted by circ-UBR4 silencing in ox-LDL-treated VSMCs (Figure 4i). Therefore, the knockdown of miR-515-5p abolished circ-UBR4 inhibition-induced effects on ox-LDL-treated VSMCs. We measured the expression levels of both circ-UBR4 and miR-515-5p in the same ox-LDL-treated cells, and confirmed that circ-UBR4 was increased and miR-515-5p was decreased in ox-LDL-induced VSMCs (Figure A3a). In VSMCs transfected with circ-UBR4 overexpression vector, we found that circ-UBR4 was markedly promoted and miR-515-5p was significantly inhibited (Figure A3b). In addition, we examined cell proliferation, inflammation, migration, and apoptosis in ox-LDL-induced VSMCs-transfected circ-UBR4 overexpression vector. The circ-UBR4 expression was remarkably enhanced after the transfection of circ-UBR4 overexpression vector in ox-LDL-induced VSMCs (Figure A3c). Then, function experiments showed that circ-UBR4 overexpression promoted cell viability, the levels of inflammation factors (TNF-α, IL-6, and IL-1β), EdU incorporation, and the number of migrated cells, while suppressed apoptosis rate in ox-LDL-induced VSMCs (Figure A3d–j). These data confirmed that circ-UBR4 inhibited miR-515-5p expression to promote cell proliferation, migration, and inflammation, and inhibit apoptosis.
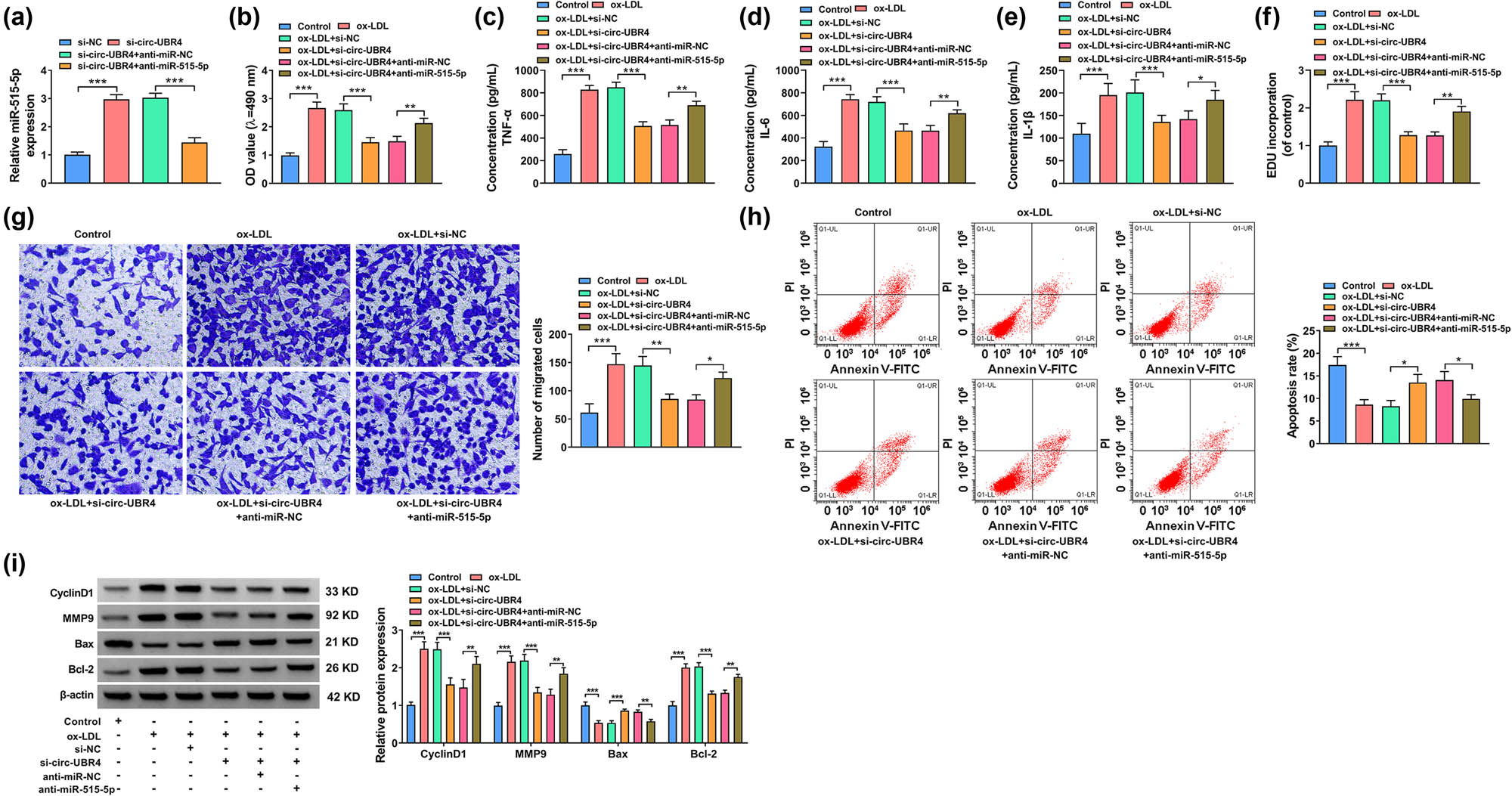
Knockdown of circ-UBR4-mediated effects in ox-LDL-stimulated VSMCs were abolished by silencing miR-515-5p. (a) RT-qPCR was performed to assess miR-515-5p level in VSMCs transfected with si-NC, si-circ-UBR4, si-circ-UBR4 + anti-miR-NC, or si-circ-UBR4 + anti-miR-515-5p. (b)–(i) VSMCs were divided into six groups: Control, ox-LDL, ox-LDL + si-NC, ox-LDL + si-circ-UBR4, ox-LDL + si-circ-UBR4 + anti-miR-NC, and ox-LDL + si-circ-UBR4 + anti-miR-515-5p. Untreated VSMCs were used as Control. (b) MTT assay was conducted to analyze cell viability. (c)–(e) The inflammation was assessed by measuring the levels of TNF-α, IL-6, and IL-1β by ELISA. (f) EdU assay was conducted to analyze cell proliferation. (g) The apoptosis of VSMCs was examined by flow cytometry assay. (h) The transwell was used to analyze VSMCs migration. (i) Western blot assay was performed to test protein expression levels of MMP9, CyclinD1, Bax, and Bcl-2 in VSMCs. *P < 0.05, **P < 0.01, ***P < 0.001.
3.5 IGF2 was a functional gene of miR-515-5p
The online software Starbase was used to predict the target genes of miR-515-5p. We found that miR-515-5p had putative binding regions in 3′UTR of IGF2 mRNA (Figure 5a). Besides, the overexpression of miR-515-5p reduced the luciferase activity of WT-IGF2 3′UTR group but not MUT-IGF2 3′UTR group (Figure 5b). RIP assay also suggested that miR-515-5p and IGF2 were enriched in Ago2-immunoprecipitated complex when compared with control group, revealing the association between miR-515-5p and IGF2 (Figure 5c). Treatment with ox-LDL increased the mRNA and protein expression levels of IGF2 in VSMCs in dose-dependent manner and time-dependent manner (Figure 5d–g). Transfection with miR-515-5p mimic increased the expression of miR-515-5p in VSMCs, while miR-515-5p decreased in VSMCs after transfection with anti-miR-515-5p (Figure 5h). More importantly, IGF2 was substantially increased in the presence of anti-miR-515-5p in VSMCs but decreased in miR-515-5p-transfected cells, suggesting that IGF2 was negatively regulated by miR-515-5p (Figure 5i and j). These results together suggested that IGF2 was a direct target of miR-515-5p.
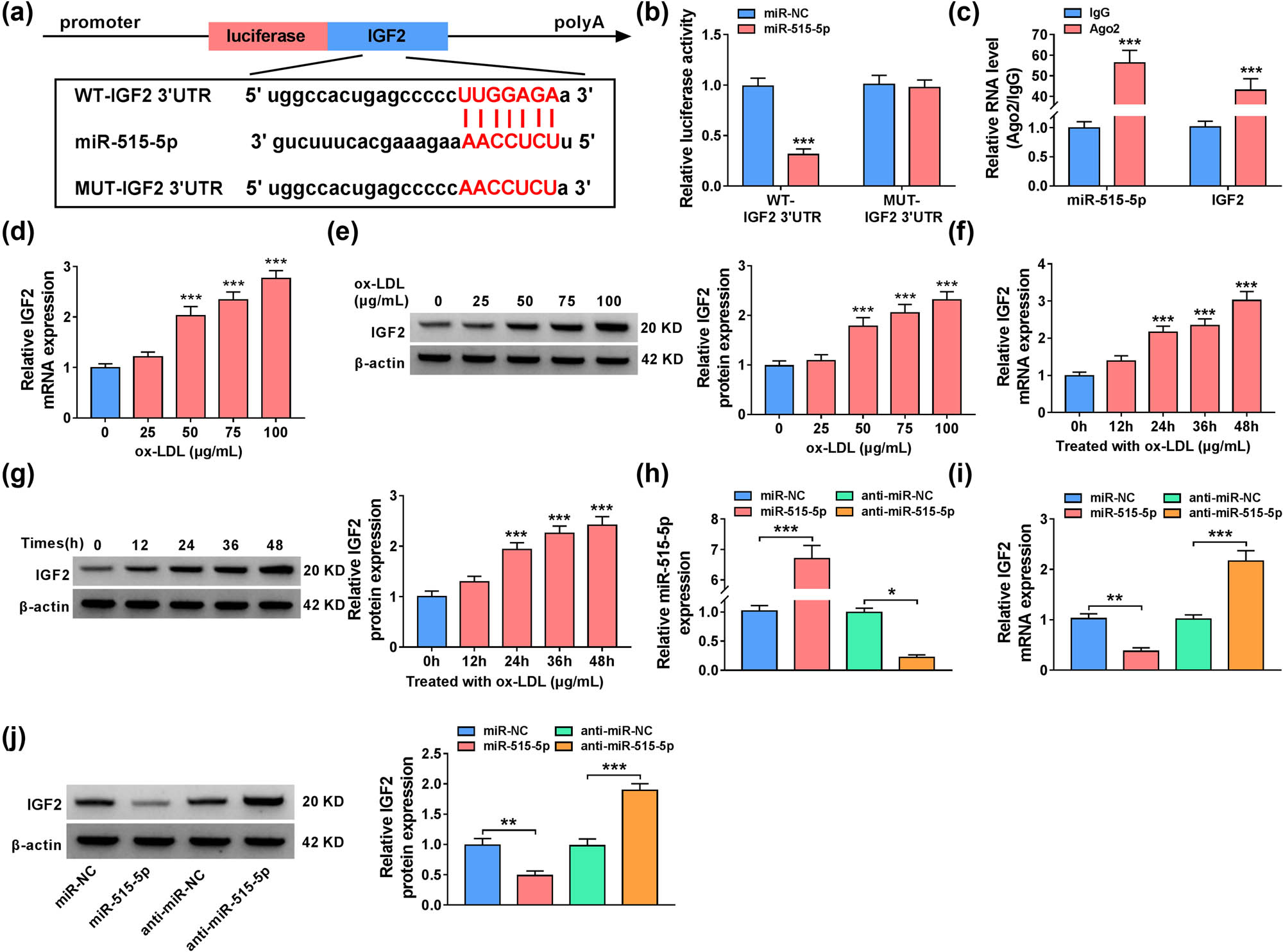
MiR-515-5p regulated the expression of IGF2 in VSMCs. (a) The binding regions between miR-515-5p and IGF2 were shown. (b) and (c) Dual-luciferase reporter and RIP assays were used to confirm the association between miR-515-5p and IGF2. (d)–(g) RT-qPCR and western blot assays were used to examine IGF2 levels in ox-LDL-stimulated VSMCs. (h) The expression level of miR-515-5p was assessed by RT-qPCR in VSMCs transfected with miR-NC, miR-515-5p, anti-miR-NC, or anti-miR-515-5p. (i) and (j) The mRNA and protein expression levels of IGF2 were measured by RT-qPCR and western blot assays in VSMCs transfected with miR-NC, miR-515-5p, anti-miR-NC, or anti-miR-515-5p. *P < 0.05, **P < 0.01, ***P < 0.001.
3.6 Overexpression of IGF2 reversed miR-515-5p-induced effects on ox-LDL-stimulated VSMCs functions
The expression level of IGF2 was inhibited by the overexpression of miR-515-5p in ox-LDL-stimulated VSMCs, which was overturned by transfection with IGF2 (Figure 6a and b). The upregulation of miR-515-5p inhibited the viability, inflammation, proliferation, and migration in ox-LDL-stimulated VSMCs, while these effects were counteracted by the overexpression of IGF2 (Figure 6c–h). MiR-515-5p enrichment-triggered apoptosis of ox-LDL-treated VSMCs was largely inhibited by IGF2 overexpression (Figure 6i). Also, miR-515-5p decreased the Cyclin D1, MMP9, and Bcl-2 protein expression, while it enhanced the Bax protein expression in ox-LDL-stimulated VSMCs. However, these effects were reversed by significantly decreased IGF2 overexpression (Figure 6j). Above all, miR-515-5p regulated proliferation, migration, inflammation, and apoptosis in ox-LDL-stimulated VSMCs by targeting IGF2.
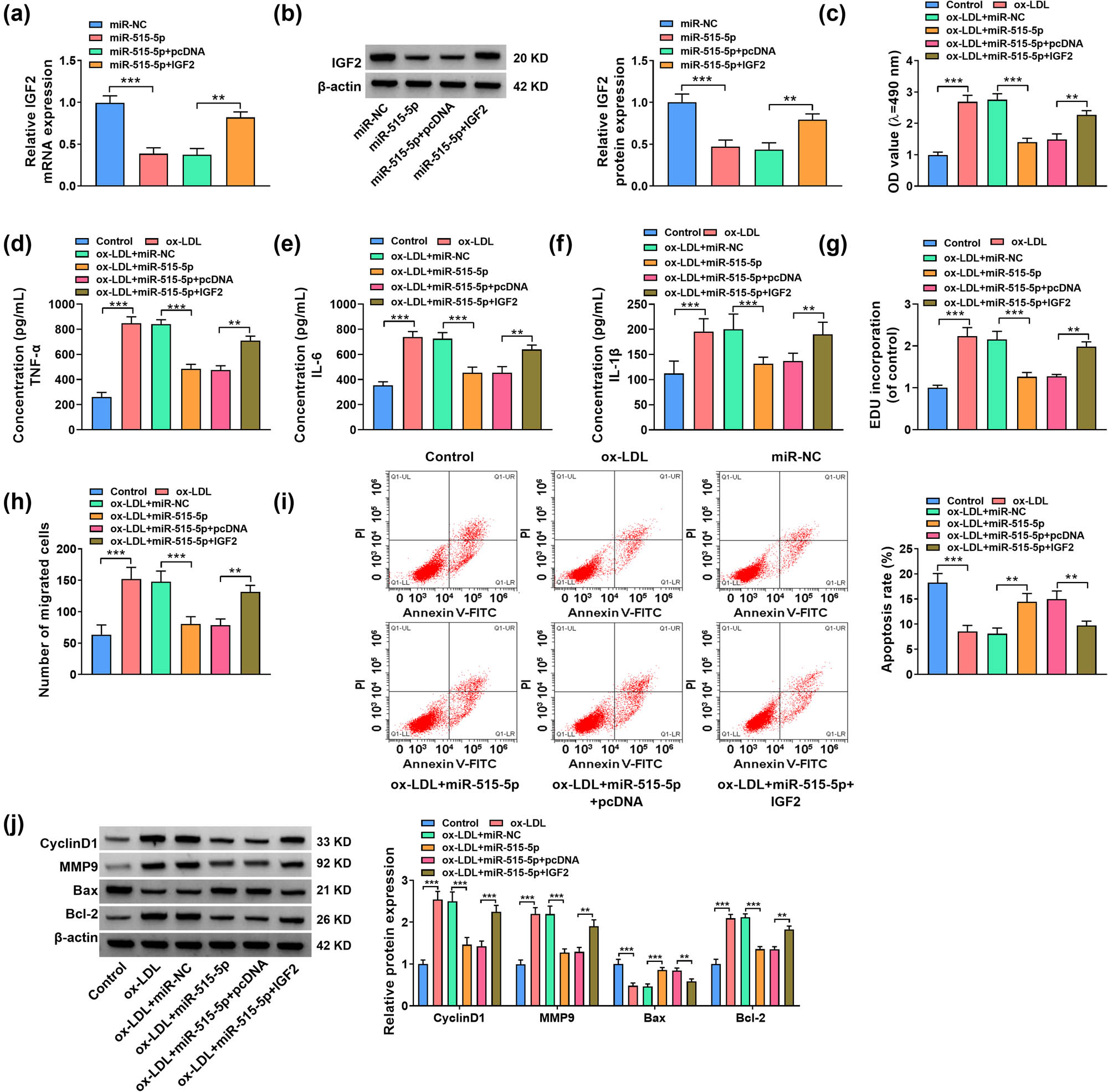
The miR-515-5p/IGF2 axis regulated proliferation, migration, inflammation, and apoptosis of ox-LDL-stimulated VSMCs. (a) and (b) The expression of IGF2 was assessed by RT-qPCR and western blot assays in VSMCs transfected with miR-NC, miR-515-5p, miR-515-5p + pcDNA, or miR-515-5p + IGF2. (c)–(j) ox-LDL-stimulated VSMCs were transfected with miR-NC, miR-515-5p, miR-515-5p + pcDNA, or miR-515-5p + IGF2, with untreated VSMCs as Control. (c) The cell viability of VSMCs was assessed by MTT assay. (d)–(f) ELISA was used to determine the levels of TNF-α, IL-6, and IL-1β in VSMCs. (g) The cell proliferation of VSMCs was assessed by EdU assay. (h) The migration of VSMCs was examined by transwell assay. (i) The apoptosis of VSMCs was examined by flow cytometry. (j) The protein expression levels of CyclinD1, MMP9, Bax, and Bcl-2 were assessed by western blot. *P < 0.05, **P < 0.01, ***P < 0.001.
3.7 circ-UBR4 regulated IGF2 by targeting miR-515-5p
As shown in Figure 7a and b, circ-UBR4 knockdown decreased the expression of IGF2 in ox-LDL-stimulated VSMCs, which was recused by the silencing of miR-515-5p, at both mRNA and protein levels. To sum up, circ-UBR4 regulated IGF2 expression by targeting miR-515-5p, thereby promoting cell proliferation, inflammation, and migration in ox-LDL-stimulated VSMCs (Figure 7c).
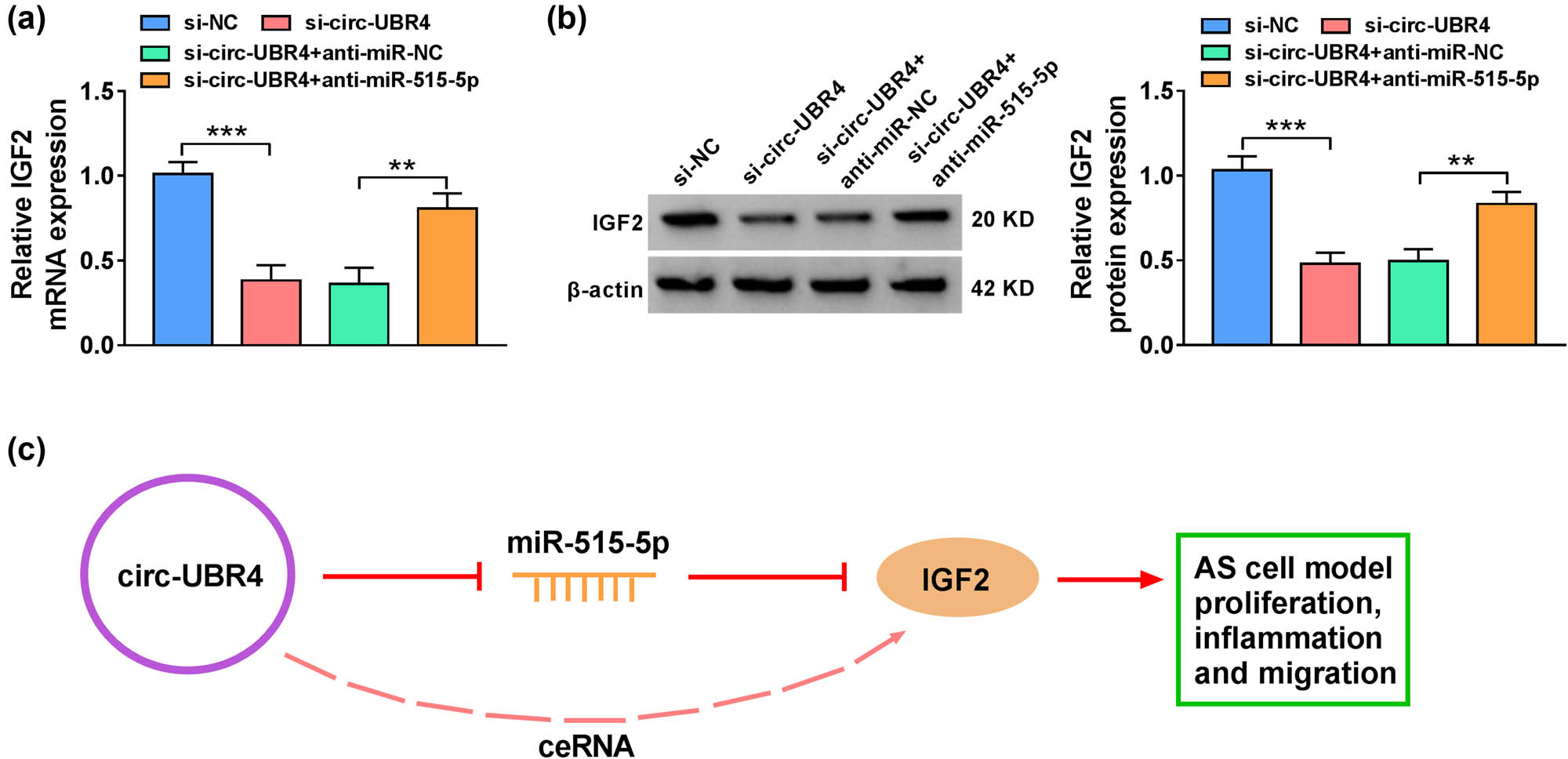
Circ-UBR4 governed the miR-515-5p/IGF2 axis. (a) and (b) The mRNA and protein expression levels of IGF2 were measured by RT-qPCR and western blot assays in VSMCs transfected with si-NC, si-circ-UBR4, si-circ-UBR4 + anti-miR-NC, or si-circ-UBR4 + anti-miR-515-5p. (c) A model for circ-UBR4/miR-515-5p/IGF2 axis was displayed. **P < 0.01, ***P < 0.001.
4 Discussion
AS is a common risk factor for cardiovascular and cerebrovascular diseases all over the world [20]. Although majority of basic and clinical research focusing on AS, the pathogenesis mechanism of AS is not fully addressed. Our data suggested that the suppression of circ-UBR4 decreased proliferation, migration, and inflammation, while it increased the apoptosis in ox-LDL-stimulated VSMCs by targeting the miR-515-5p/IGF2 axis.
As we all know, ox-LDL could stimulate migration and proliferation of VSMCs [21]. Under pathological conditions, migrated and proliferated VSMCs in intimal layer of artery were found to participate in the early AS formation [22]. Besides, it was identified that ox-LDL stimulated cell adhesion molecules expression and thus induced a series of pathological changes, including inflammatory reactions and injury [23]. Therefore, ox-LDL-induced VSMCs were widely used as cell models of AS. Not surprisingly, inflammatory reaction was also associated with the pathogenesis of AS [24]. The pro-inflammatory cytokines, including TNF-α and IL-6, could induce dysfunction of VSMCs and promote AS progression [25]. We also confirmed that the productions of TNF-α, IL-6, and IL-1β were increased in ox-LDL-induced VSMCs.
Recently, circRNAs function as competitive endogenous RNAs to sponge miRNAs and then suppress their functions, which has been confirmed in the development of AS [26]. For example, Guo et al. reported a series of differentially expressed circRNAs in ox-LDL-induced VSMCs, including circ-UBR4 [27]. Recently, Ding et al. reported that the circ-UBR4/miR-370-3p/high mobility group box 1 networks mediated cell proliferation of ox-LDL-treated VSMCs, suggesting the important roles of circ-UBR4 in AS [10]. Our results suggested that miR-515-5p was involved in the regulatory mechanism of circ-UBR4 in AS progression.
The previous research confirmed that miR-515-5p was implicated in the development of human diseases by targeting key mRNAs, such as chromobox homolog 4 [28], IL-6 [29], and YES proto-oncogene 1 [30]. In addition, miR-515-5p was confirmed to play an important role in AS progression, and long noncoding RNA LOXL1 antisense RNA 1 could sponge miR-515-5p to facilitate the development of AS [31]. Similar to the above conclusion, we found that miR-515-5p regulated proliferation, migration, inflammation, and apoptosis of ox-LDL-treated VSMCs by targeting IGF2.
Zaina et al. provided the atherogenic activity of IGF2 in mice lacking apolipoprotein E [32]. Mechanistically, IGF2 was also involved in proliferation and apoptosis of ox-LDL-induced VSMC, and it was reported as a functional target of miR-148b [33], miR-424-5p [34], and miR-210-3p [35]. Besides, the pro-inflammation characteristic of IGF2 also was confirmed in ox-LDL-stimulate THP-1 macrophages [36]. Therefore, IGF2 served as an active participant in AS progression.
Taken together, circ-UBR4 regulated the proliferation, migration, inflammation, and apoptosis of ox-LDL-induced VSMCs via miR-515-5p/IGF2 axis. Our study enriched the role of circ-UBR4 in ox-LDL-induced VSMCs and thus provided a new perspective to understand AS pathogenesis. However, some limitations existed in our present study. We only determined the role of circ-UBR4 in cell models of AS in vitro, and animal models of AS were not provided in our present study. Therefore, VSMCs from various resources and animal models should be used to further validate our present findings in future work.
5 Conclusion
In summary, treatment with ox-LDL increased the expression of circ-UBR4 in VSMCs. We also demonstrated that the downregulation of circ-UBR4 could effectively repress proliferation, migration, and inflammation but increase apoptosis of ox-LDL-induced VSMCs via miR-515-5p/IGF2 axis, hinting that circ-UBR4 might be a new diagnostic marker for AS patients.
-
Funding information: This work was supported by Key Discipline in Yangpu District, Shanghai (YP19ZA09).
-
Author contributions: Conceptualization and methodology: Tianhua Liu and Jun Shi; formal analysis and data curation: Yu Wang, Yuya Yang, and Wenyin Xiao; validation and investigation: Wenyin Xiao and Yanyan Bai; writing – original draft preparation and writing – review and editing: Liuliu Feng, Tianhua Liu, and Jun Shi. All authors have accepted responsibility for the entire content of this manuscript and approved its submission.
-
Conflict of interest: The authors declare that they have no competing interest.
-
Data availability statement: The analyzed datasets generated during the present study are available from the corresponding author on reasonable request.
Appendix
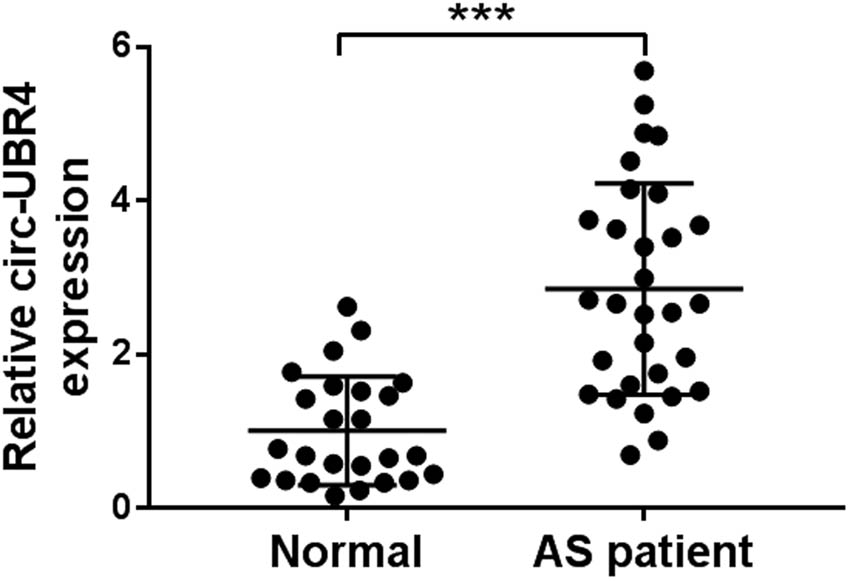
The expression of circ-UBR4 in the serum of AS patients or healthy normal controls. RT-qPCR was used to detect the expression of circ-UBR4 in the serum of AS patients (n = 31) or healthy normal controls (n = 25). ***P < 0.001.
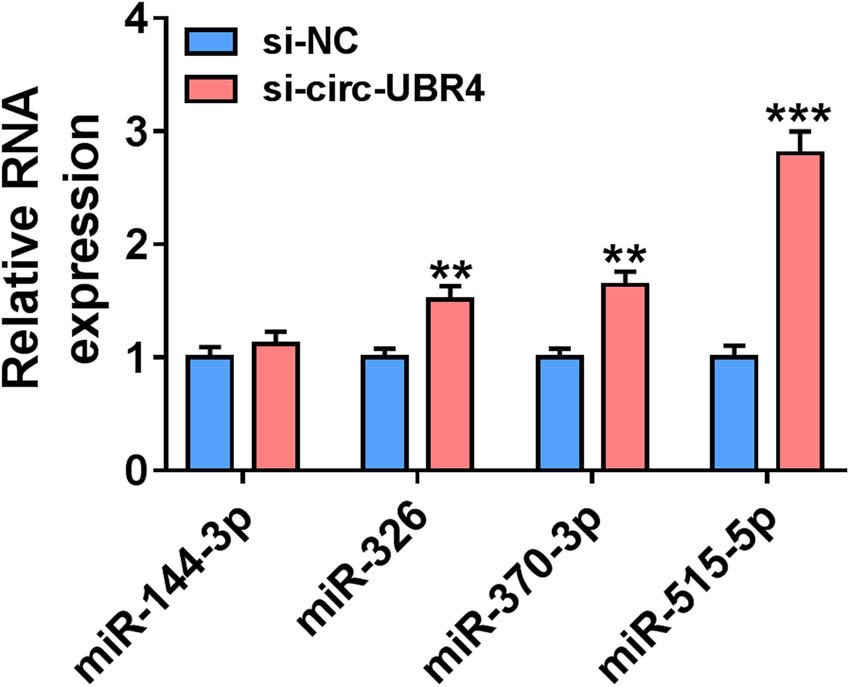
The selection of targeted miRNA of circ-UBR4. The expression of 4 miRNAs (miR-144-3p, miR-326, miR-370-3p and miR-515-5p) was detected by RT-qPCR in VSMCs transfected with si-circ-UBR4 or si-NC. **P < 0.01, ***P < 0.001.
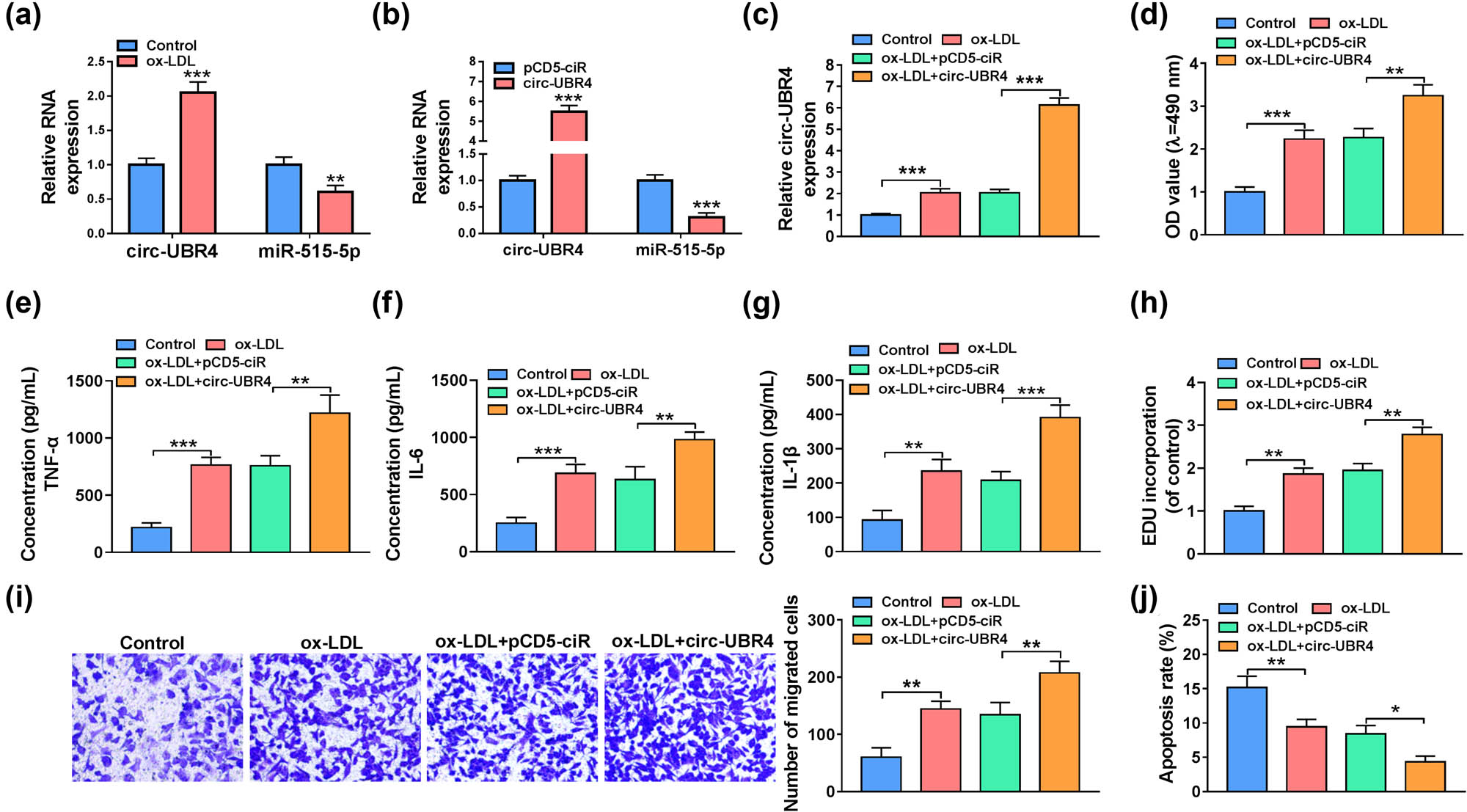
Effects of circ-UBR4 on miR-515-5p expression and ox-LDL-stimulated VSMCs function. (a) The expression levels of circ-UBR4 and miR-515-5p were assessed by RT-qPCR in VSMCs treated with or without ox-LDL. (b) The expression levels of circ-UBR4 and miR-515-5p were assessed by RT-qPCR in VSMSc transfected with pCD5-ciR or circ-UBR4 overexpression vector. (c–j) Ox-LDL-stimulated VSMCs were transfected with pCD5-ciR or circ-UBR4 overexpression vector, with un-treated cells as Control. (c) The circ-UBR4 expression was detected by RT-qPCR. (d) Cell viability was assessed by MTT assay. (e–g) ELISA was used to determine the levels of TNF-α, IL-6, and IL-1β. (h) Cell proliferation was assessed by EdU assay. (i) VSMCs migration was examined by transwell assay. (j) Cell apoptosis was examined by flow cytometry. *P < 0.05, **P < 0.01, ***P < 0.001.
References
[1] Torres N, Guevara-Cruz M, Velazquez-Villegas LA, Tovar AR. Nutrition and atherosclerosis. Arch Med Res. 2015;46(5):408–26.10.1016/j.arcmed.2015.05.010Suche in Google Scholar PubMed
[2] Schaftenaar F, Frodermann V, Kuiper J, Lutgens E. Atherosclerosis: the interplay between lipids and immune cells. Curr OpLipidol. 2016;27(3):209–15.10.1097/MOL.0000000000000302Suche in Google Scholar PubMed
[3] Libby P, Loscalzo J, Ridker PM, Farkouh ME, Hsue PY, Fuster V, et al. Inflammation, immunity, and infection in atherothrombosis: JACC review topic of the week. J Am Coll Cardiol. 2018;72(17):2071–81.10.1016/j.jacc.2018.08.1043Suche in Google Scholar PubMed PubMed Central
[4] Lim S, Park S. Role of vascular smooth muscle cell in the inflammation of atherosclerosis. BMB Rep. 2014;47(1):1–7.10.5483/BMBRep.2014.47.1.285Suche in Google Scholar
[5] Fan M, Huang Y, Li K, Yang X, Bai J, Si Q, et al. ox-LDL regulates proliferation and apoptosis in VSMCs by controlling the miR-183-5p/FOXO1. Genes Genom. 2022;44(6):671–81.10.1007/s13258-022-01236-xSuche in Google Scholar PubMed
[6] Zhang L, Cheng H, Yue Y, Li S, Zhang D, He R. H19 knockdown suppresses proliferation and induces apoptosis by regulating miR-148b/WNT/beta-catenin in ox-LDL-stimulated vascular smooth muscle cells. J Biomed Sci. 2018;25(1):11.10.1186/s12929-018-0418-4Suche in Google Scholar PubMed PubMed Central
[7] Salzman J. Circular RNA expression: Its potential regulation and function. Trends Genet. 2016;32(5):309–16.10.1016/j.tig.2016.03.002Suche in Google Scholar PubMed PubMed Central
[8] Cao Q, Guo Z, Du S, Ling H, Song C. Circular RNAs in the pathogenesis of atherosclerosis. Life Sci. 2020;255:117837.10.1016/j.lfs.2020.117837Suche in Google Scholar PubMed
[9] Yang L, Yang F, Zhao H, Wang M, Zhang Y. Circular RNA circCHFR facilitates the proliferation and migration of vascular smooth muscle via miR-370/FOXO1/Cyclin D1 pathway. Mol Therapy Nucleic acids. 2019;16:434–1.10.1016/j.omtn.2019.02.028Suche in Google Scholar PubMed PubMed Central
[10] Ding P, Ding Y, Tian Y, Lei X. Circular RNA circ_0010283 regulates the viability and migration of oxidized low-density lipoprotein-induced vascular smooth muscle cells via an miR3703p/HMGB1 axis in atherosclerosis. Int J Mol Med. 2020;46(4):1399–408.10.3892/ijmm.2020.4703Suche in Google Scholar PubMed PubMed Central
[11] Feng Z, Zhu Y, Zhang J, Yang W, Chen Z, Li B. Hsa-circ_0010283 regulates oxidized low-density lipoprotein-induced proliferation and migration of vascular smooth muscle cells by targeting the miR-133a-3p/pregnancy-associated plasma protein A axis. Circ J off J Jap Circ Soc. 2020;84(12):2259–69.10.1253/circj.CJ-20-0345Suche in Google Scholar PubMed
[12] Vishnoi A, Rani S. miRNA biogenesis and regulation of diseases: an overview. Methods Mol Biol. 2017;1509:1–10.10.1007/978-1-4939-6524-3_1Suche in Google Scholar PubMed
[13] Liu B, Li J, Cairns MJ. Identifying miRNAs, targets and functions. Brief Bioinform. 2014;15(1):1–19.10.1093/bib/bbs075Suche in Google Scholar PubMed PubMed Central
[14] Pardo OE, Castellano L, Munro CE, Hu Y, Mauri F, Krell J, et al. miR-515-5p controls cancer cell migration through MARK4 regulation. EMBO Rep. 2016;17(4):570–84.10.15252/embr.201540970Suche in Google Scholar PubMed PubMed Central
[15] Zhang X, Zhou J, Xue D, Li Z, Liu Y, Dong L. miR-515-5p acts as a tumor suppressor via targeting TRIP13 in prostate cancer. Int J Biol Macromol. 2019;129:227–32.10.1016/j.ijbiomac.2019.01.127Suche in Google Scholar PubMed
[16] Qiao K, Ning S, Wan L, Wu H, Wang Q, Zhang X, et al. LINC00673 is activated by YY1 and promotes the proliferation of breast cancer cells via the miR-515-5p/MARK4/Hippo signaling pathway. J Exp Clin Cancer Res. 2019;38(1):418.10.1186/s13046-019-1421-7Suche in Google Scholar PubMed PubMed Central
[17] Nordin M, Bergman D, Halje M, Engstrom W, Ward A. Epigenetic regulation of the IGF2/H19 gene cluster. Cell Prolif. 2014;47(3):189–99.10.1111/cpr.12106Suche in Google Scholar PubMed PubMed Central
[18] Vishwamitra D, George SK, Shi P, Kaseb AO, Amin HM. Type I insulin-like growth factor receptor signaling in hematological malignancies. Oncotarget. 2017;8(1):1814–44.10.18632/oncotarget.12123Suche in Google Scholar PubMed PubMed Central
[19] Sun Y, Chen D, Cao L, Zhang R, Zhou J, Chen H, et al. MiR-490-3p modulates the proliferation of vascular smooth muscle cells induced by ox-LDL through targeting PAPP-A. Cardiovasc Res. 2013;100(2):272–9.10.1093/cvr/cvt172Suche in Google Scholar PubMed
[20] Barquera S, Pedroza-Tobias A, Medina C, Hernandez-Barrera L, Bibbins-Domingo K, Lozano R, et al. Global overview of the epidemiology of atherosclerotic cardiovascular disease. Arch Med Res. 2015;46(5):328–38.10.1016/j.arcmed.2015.06.006Suche in Google Scholar PubMed
[21] Kattoor AJ, Kanuri SH, Mehta JL. Role of ox-LDL and LOX-1 in atherogenesis. Curr Medicin Chem. 2019;26(9):1693–700.10.2174/0929867325666180508100950Suche in Google Scholar PubMed
[22] Bennett MR, Sinha S, Owens GK. Vascular smooth muscle cells in atherosclerosis. Circ Res. 2016;118(4):692–702.10.1161/CIRCRESAHA.115.306361Suche in Google Scholar PubMed PubMed Central
[23] Rhoads JP, Major AS. How oxidized low-density lipoprotein activates inflammatory responses. Crit Rev Immunology. 2018;38(4):333–42.10.1615/CritRevImmunol.2018026483Suche in Google Scholar PubMed PubMed Central
[24] Fredman G, Tabas I. Boosting inflammation resolution in atherosclerosis: the next frontier for therapy. Am J Pathol. 2017;187(6):1211–21.10.1016/j.ajpath.2017.01.018Suche in Google Scholar PubMed PubMed Central
[25] Back M, Hansson GK. Anti-inflammatory therapies for atherosclerosis. Nat Rev Cardiol. 2015;12(4):199–211.10.1038/nrcardio.2015.5Suche in Google Scholar PubMed
[26] Zhang F, Zhang R, Zhang X, Wu Y, Li X, Zhang S, et al. Comprehensive analysis of circRNA expression pattern and circRNA-miRNA-mRNA network in the pathogenesis of atherosclerosis in rabbits. Aging (Albany NY). 2018;10(9):2266–83.10.18632/aging.101541Suche in Google Scholar PubMed PubMed Central
[27] Guo M, Yan R, Ji Q, Yao H, Sun M, Duan L, et al. IFN regulatory factor-1 induced macrophage pyroptosis by modulating m6A modification of circ_0029589 in patients with acute coronary syndrome. Int Immunopharmacol. 2020;86:106800.10.1016/j.intimp.2020.106800Suche in Google Scholar PubMed
[28] Huang FJ, Dang JQ, Zhang S, Cheng ZY. Circular RNA hsa_circ_0008039 promotes proliferation, migration and invasion of breast cancer cells through upregulating CBX4 via sponging miR-515-5p. Eur Rev Med Pharmacol Sci. 2020;24(4):1887–98.Suche in Google Scholar
[29] Ni JS, Zheng H, Ou YL, Tao YP, Wang ZG, Song LH, et al. miR-515-5p suppresses HCC migration and invasion via targeting IL6/JAK/STAT3 pathway. Surg Oncol. 2020;34:113–20.10.1016/j.suronc.2020.03.003Suche in Google Scholar PubMed
[30] Zhang Y, Shi Z, Li Z, Wang X, Zheng P, Li H. Circ_0057553/miR-515-5p regulates prostate cancer cell proliferation, apoptosis, migration, invasion and aerobic glycolysis by targeting YES1. Onco Targets Ther. 2020;13:11289–99.10.2147/OTT.S272294Suche in Google Scholar PubMed PubMed Central
[31] Xie Q, Li F, Shen K, Luo C, Song G. LOXL1-AS1/miR-515-5p/STAT3 positive feedback loop facilitates cell proliferation and migration in atherosclerosis. J Cardiovasc Pharmacol. 2020;76(2):151–8.10.1097/FJC.0000000000000853Suche in Google Scholar PubMed
[32] Zaina S, Pettersson L, Ahren B, Branen L, Hassan AB, Lindholm M, et al. Insulin-like growth factor II plays a central role in atherosclerosis in a mouse model. J Biol Chem. 2002;277(6):4505–11.10.1074/jbc.M108061200Suche in Google Scholar PubMed
[33] Wu X, Zheng X, Cheng J, Zhang K, Ma C. LncRNA TUG1 regulates proliferation and apoptosis by regulating miR-148b/IGF2 axis in ox-LDL-stimulated VSMC and HUVEC. Life Sci. 2020;243:117287.10.1016/j.lfs.2020.117287Suche in Google Scholar PubMed
[34] Yu H, Zhao L, Zhao Y, Fei J, Zhang W. Circular RNA circ_0029589 regulates proliferation, migration, invasion, and apoptosis in ox-LDL-stimulated VSMCs by regulating miR-424-5p/IGF2 axis. Vasc Pharmacol. 2020;135:106782.10.1016/j.vph.2020.106782Suche in Google Scholar PubMed
[35] Qiao XR, Wang L, Liu M, Tian Y, Chen T. MiR-210-3p attenuates lipid accumulation and inflammation in atherosclerosis by repressing IGF2. Biosci Biotechnol Biochem. 2020;84(2):321–9.10.1080/09168451.2019.1685370Suche in Google Scholar PubMed
[36] Li SF, Hu YW, Zhao JY, Ma X, Wu SG, Lu JB, et al. Ox-LDL upregulates CRP expression through the IGF2 pathway in THP-1 macrophages. Inflammation. 2015;38(2):576–83.10.1007/s10753-014-9964-4Suche in Google Scholar PubMed
© 2023 the author(s), published by De Gruyter
This work is licensed under the Creative Commons Attribution 4.0 International License.
Artikel in diesem Heft
- Research Articles
- Exosomes derived from mesenchymal stem cells overexpressing miR-210 inhibits neuronal inflammation and contribute to neurite outgrowth through modulating microglia polarization
- Current situation of acute ST-segment elevation myocardial infarction in a county hospital chest pain center during an epidemic of novel coronavirus pneumonia
- circ-IARS depletion inhibits the progression of non-small-cell lung cancer by circ-IARS/miR-1252-5p/HDGF ceRNA pathway
- circRNA ITGA7 restrains growth and enhances radiosensitivity by up-regulating SMAD4 in colorectal carcinoma
- WDR79 promotes aerobic glycolysis of pancreatic ductal adenocarcinoma (PDAC) by the suppression of SIRT4
- Up-regulation of collagen type V alpha 2 (COL5A2) promotes malignant phenotypes in gastric cancer cell via inducing epithelial–mesenchymal transition (EMT)
- Inhibition of TERC inhibits neural apoptosis and inflammation in spinal cord injury through Akt activation and p-38 inhibition via the miR-34a-5p/XBP-1 axis
- 3D-printed polyether-ether-ketone/n-TiO2 composite enhances the cytocompatibility and osteogenic differentiation of MC3T3-E1 cells by downregulating miR-154-5p
- Propofol-mediated circ_0000735 downregulation restrains tumor growth by decreasing integrin-β1 expression in non-small cell lung cancer
- PVT1/miR-16/CCND1 axis regulates gastric cancer progression
- Silencing of circ_002136 sensitizes gastric cancer to paclitaxel by targeting the miR-16-5p/HMGA1 axis
- Short-term outcomes after simultaneous gastrectomy plus cholecystectomy in gastric cancer: A pooling up analysis
- SCARA5 inhibits oral squamous cell carcinoma via inactivating the STAT3 and PI3K/AKT signaling pathways
- Molecular mechanism by which the Notch signaling pathway regulates autophagy in a rat model of pulmonary fibrosis in pigeon breeder’s lung
- lncRNA TPT1-AS1 promotes cell migration and invasion in esophageal squamous-cell carcinomas by regulating the miR-26a/HMGA1 axis
- SIRT1/APE1 promotes the viability of gastric cancer cells by inhibiting p53 to suppress ferroptosis
- Glycoprotein non-metastatic melanoma B interacts with epidermal growth factor receptor to regulate neural stem cell survival and differentiation
- Treatments for brain metastases from EGFR/ALK-negative/unselected NSCLC: A network meta-analysis
- Association of osteoporosis and skeletal muscle loss with serum type I collagen carboxyl-terminal peptide β glypeptide: A cross-sectional study in elder Chinese population
- circ_0000376 knockdown suppresses non-small cell lung cancer cell tumor properties by the miR-545-3p/PDPK1 pathway
- Delivery in a vertical birth chair supported by freedom of movement during labor: A randomized control trial
- UBE2J1 knockdown promotes cell apoptosis in endometrial cancer via regulating PI3K/AKT and MDM2/p53 signaling
- Metabolic resuscitation therapy in critically ill patients with sepsis and septic shock: A pilot prospective randomized controlled trial
- Lycopene ameliorates locomotor activity and urinary frequency induced by pelvic venous congestion in rats
- UHRF1-induced connexin26 methylation is involved in hearing damage triggered by intermittent hypoxia in neonatal rats
- LINC00511 promotes melanoma progression by targeting miR-610/NUCB2
- Ultra-high-performance liquid chromatography-tandem mass spectrometry analysis of serum metabolomic characteristics in people with different vitamin D levels
- Role of Jumonji domain-containing protein D3 and its inhibitor GSK-J4 in Hashimoto’s thyroiditis
- circ_0014736 induces GPR4 to regulate the biological behaviors of human placental trophoblast cells through miR-942-5p in preeclampsia
- Monitoring of sirolimus in the whole blood samples from pediatric patients with lymphatic anomalies
- Effects of osteogenic growth peptide C-terminal pentapeptide and its analogue on bone remodeling in an osteoporosis rat model
- A novel autophagy-related long non-coding RNAs signature predicting progression-free interval and I-131 therapy benefits in papillary thyroid carcinoma
- WGCNA-based identification of potential targets and pathways in response to treatment in locally advanced breast cancer patients
- Radiomics model using preoperative computed tomography angiography images to differentiate new from old emboli of acute lower limb arterial embolism
- Dysregulated lncRNAs are involved in the progress of myocardial infarction by constructing regulatory networks
- Single-arm trial to evaluate the efficacy and safety of baclofen in treatment of intractable hiccup caused by malignant tumor chemotherapy
- Genetic polymorphisms of MRPS30-DT and NINJ2 may influence lung cancer risk
- Efficacy of immune checkpoint inhibitors in patients with KRAS-mutant advanced non-small cell lung cancer: A retrospective analysis
- Pyroptosis-based risk score predicts prognosis and drug sensitivity in lung adenocarcinoma
- Upregulation of lncRNA LANCL1-AS1 inhibits the progression of non-small-cell lung cancer via the miR-3680-3p/GMFG axis
- CircRANBP17 modulated KDM1A to regulate neuroblastoma progression by sponging miR-27b-3p
- Exosomal miR-93-5p regulated the progression of osteoarthritis by targeting ADAMTS9
- Downregulation of RBM17 enhances cisplatin sensitivity and inhibits cell invasion in human hypopharyngeal cancer cells
- HDAC5-mediated PRAME regulates the proliferation, migration, invasion, and EMT of laryngeal squamous cell carcinoma via the PI3K/AKT/mTOR signaling pathway
- The association between sleep duration, quality, and nonalcoholic fatty liver disease: A cross-sectional study
- Myostatin silencing inhibits podocyte apoptosis in membranous nephropathy through Smad3/PKA/NOX4 signaling pathway
- A novel long noncoding RNA AC125257.1 facilitates colorectal cancer progression by targeting miR-133a-3p/CASC5 axis
- Impact of omicron wave and associated control measures in Shanghai on health management and psychosocial well-being of patients with chronic conditions
- Clinicopathological characteristics and prognosis of young patients aged ≤45 years old with non-small cell lung cancer
- TMT-based comprehensive proteomic profiling identifies serum prognostic signatures of acute myeloid leukemia
- The dose limits of teeth protection for patients with nasopharyngeal carcinoma undergoing radiotherapy based on the early oral health-related quality of life
- miR-30b-5p targeting GRIN2A inhibits hippocampal damage in epilepsy
- Long non-coding RNA AL137789.1 promoted malignant biological behaviors and immune escape of pancreatic carcinoma cells
- IRF6 and FGF1 polymorphisms in non-syndromic cleft lip with or without cleft palate in the Polish population
- Comprehensive analysis of the role of SFXN family in breast cancer
- Efficacy of bronchoscopic intratumoral injection of endostar and cisplatin in lung squamous cell carcinoma patients underwent conventional chemoradiotherapy
- Silencing of long noncoding RNA MIAT inhibits the viability and proliferation of breast cancer cells by promoting miR-378a-5p expression
- AG1024, an IGF-1 receptor inhibitor, ameliorates renal injury in rats with diabetic nephropathy via the SOCS/JAK2/STAT pathway
- Downregulation of KIAA1199 alleviated the activation, proliferation, and migration of hepatic stellate cells by the inhibition of epithelial–mesenchymal transition
- Exendin-4 regulates the MAPK and WNT signaling pathways to alleviate the osteogenic inhibition of periodontal ligament stem cells in a high glucose environment
- Inhibition of glycolysis represses the growth and alleviates the endoplasmic reticulum stress of breast cancer cells by regulating TMTC3
- The function of lncRNA EMX2OS/miR-653-5p and its regulatory mechanism in lung adenocarcinoma
- Tectorigenin alleviates the apoptosis and inflammation in spinal cord injury cell model through inhibiting insulin-like growth factor-binding protein 6
- Ultrasound examination supporting CT or MRI in the evaluation of cervical lymphadenopathy in patients with irradiation-treated head and neck cancer
- F-box and WD repeat domain containing 7 inhibits the activation of hepatic stellate cells by degrading delta-like ligand 1 to block Notch signaling pathway
- Knockdown of circ_0005615 enhances the radiosensitivity of colorectal cancer by regulating the miR-665/NOTCH1 axis
- Long noncoding RNA Mhrt alleviates angiotensin II-induced cardiac hypertrophy phenotypes by mediating the miR-765/Wnt family member 7B pathway
- Effect of miR-499-5p/SOX6 axis on atrial fibrosis in rats with atrial fibrillation
- Cholesterol induces inflammation and reduces glucose utilization
- circ_0004904 regulates the trophoblast cell in preeclampsia via miR-19b-3p/ARRDC3 axis
- NECAB3 promotes the migration and invasion of liver cancer cells through HIF-1α/RIT1 signaling pathway
- The poor performance of cardiovascular risk scores in identifying patients with idiopathic inflammatory myopathies at high cardiovascular risk
- miR-2053 inhibits the growth of ovarian cancer cells by downregulating SOX4
- Nucleophosmin 1 associating with engulfment and cell motility protein 1 regulates hepatocellular carcinoma cell chemotaxis and metastasis
- α-Hederin regulates macrophage polarization to relieve sepsis-induced lung and liver injuries in mice
- Changes of microbiota level in urinary tract infections: A meta-analysis
- Identification of key enzalutamide-resistance-related genes in castration-resistant prostate cancer and verification of RAD51 functions
- Falls during oxaliplatin-based chemotherapy for gastrointestinal malignancies – (lessons learned from) a prospective study
- Outcomes of low-risk birth care during the Covid-19 pandemic: A cohort study from a tertiary care center in Lithuania
- Vitamin D protects intestines from liver cirrhosis-induced inflammation and oxidative stress by inhibiting the TLR4/MyD88/NF-κB signaling pathway
- Integrated transcriptome analysis identifies APPL1/RPS6KB2/GALK1 as immune-related metastasis factors in breast cancer
- Genomic analysis of immunogenic cell death-related subtypes for predicting prognosis and immunotherapy outcomes in glioblastoma multiforme
- Circular RNA Circ_0038467 promotes the maturation of miRNA-203 to increase lipopolysaccharide-induced apoptosis of chondrocytes
- An economic evaluation of fine-needle cytology as the primary diagnostic tool in the diagnosis of lymphadenopathy
- Midazolam impedes lung carcinoma cell proliferation and migration via EGFR/MEK/ERK signaling pathway
- Network pharmacology combined with molecular docking and experimental validation to reveal the pharmacological mechanism of naringin against renal fibrosis
- PTPN12 down-regulated by miR-146b-3p gene affects the malignant progression of laryngeal squamous cell carcinoma
- miR-141-3p accelerates ovarian cancer progression and promotes M2-like macrophage polarization by targeting the Keap1-Nrf2 pathway
- lncRNA OIP5-AS1 attenuates the osteoarthritis progression in IL-1β-stimulated chondrocytes
- Overexpression of LINC00607 inhibits cell growth and aggressiveness by regulating the miR-1289/EFNA5 axis in non-small-cell lung cancer
- Subjective well-being in informal caregivers during the COVID-19 pandemic
- Nrf2 protects against myocardial ischemia-reperfusion injury in diabetic rats by inhibiting Drp1-mediated mitochondrial fission
- Unfolded protein response inhibits KAT2B/MLKL-mediated necroptosis of hepatocytes by promoting BMI1 level to ubiquitinate KAT2B
- Bladder cancer screening: The new selection and prediction model
- circNFATC3 facilitated the progression of oral squamous cell carcinoma via the miR-520h/LDHA axis
- Prone position effect in intensive care patients with SARS-COV-2 pneumonia
- Clinical observation on the efficacy of Tongdu Tuina manipulation in the treatment of primary enuresis in children
- Dihydroartemisinin ameliorates cerebral I/R injury in rats via regulating VWF and autophagy-mediated SIRT1/FOXO1 pathway
- Knockdown of circ_0113656 assuages oxidized low-density lipoprotein-induced vascular smooth muscle cell injury through the miR-188-3p/IGF2 pathway
- Low Ang-(1–7) and high des-Arg9 bradykinin serum levels are correlated with cardiovascular risk factors in patients with COVID-19
- Effect of maternal age and body mass index on induction of labor with oral misoprostol for premature rupture of membrane at term: A retrospective cross-sectional study
- Potential protective effects of Huanglian Jiedu Decoction against COVID-19-associated acute kidney injury: A network-based pharmacological and molecular docking study
- Clinical significance of serum MBD3 detection in girls with central precocious puberty
- Clinical features of varicella-zoster virus caused neurological diseases detected by metagenomic next-generation sequencing
- Collagen treatment of complex anorectal fistula: 3 years follow-up
- LncRNA CASC15 inhibition relieves renal fibrosis in diabetic nephropathy through down-regulating SP-A by sponging to miR-424
- Efficacy analysis of empirical bismuth quadruple therapy, high-dose dual therapy, and resistance gene-based triple therapy as a first-line Helicobacter pylori eradication regimen – An open-label, randomized trial
- SMOC2 plays a role in heart failure via regulating TGF-β1/Smad3 pathway-mediated autophagy
- A prospective cohort study of the impact of chronic disease on fall injuries in middle-aged and older adults
- circRNA THBS1 silencing inhibits the malignant biological behavior of cervical cancer cells via the regulation of miR-543/HMGB2 axis
- hsa_circ_0000285 sponging miR-582-3p promotes neuroblastoma progression by regulating the Wnt/β-catenin signaling pathway
- Long non-coding RNA GNAS-AS1 knockdown inhibits proliferation and epithelial–mesenchymal transition of lung adenocarcinoma cells via the microRNA-433-3p/Rab3A axis
- lncRNA UCA1 regulates miR-132/Lrrfip1 axis to promote vascular smooth muscle cell proliferation
- Twenty-four-color full spectrum flow cytometry panel for minimal residual disease detection in acute myeloid leukemia
- Hsa-miR-223-3p participates in the process of anthracycline-induced cardiomyocyte damage by regulating NFIA gene
- Anti-inflammatory effect of ApoE23 on Salmonella typhimurium-induced sepsis in mice
- Analysis of somatic mutations and key driving factors of cervical cancer progression
- Hsa_circ_0028007 regulates the progression of nasopharyngeal carcinoma through the miR-1179/SQLE axis
- Variations in sexual function after laparoendoscopic single-site hysterectomy in women with benign gynecologic diseases
- Effects of pharmacological delay with roxadustat on multi-territory perforator flap survival in rats
- Analysis of heroin effects on calcium channels in rat cardiomyocytes based on transcriptomics and metabolomics
- Risk factors of recurrent bacterial vaginosis among women of reproductive age: A cross-sectional study
- Alkbh5 plays indispensable roles in maintaining self-renewal of hematopoietic stem cells
- Study to compare the effect of casirivimab and imdevimab, remdesivir, and favipiravir on progression and multi-organ function of hospitalized COVID-19 patients
- Correlation between microvessel maturity and ISUP grades assessed using contrast-enhanced transrectal ultrasonography in prostate cancer
- The protective effect of caffeic acid phenethyl ester in the nephrotoxicity induced by α-cypermethrin
- Norepinephrine alleviates cyclosporin A-induced nephrotoxicity by enhancing the expression of SFRP1
- Effect of RUNX1/FOXP3 axis on apoptosis of T and B lymphocytes and immunosuppression in sepsis
- The function of Foxp1 represses β-adrenergic receptor transcription in the occurrence and development of bladder cancer through STAT3 activity
- Risk model and validation of carbapenem-resistant Klebsiella pneumoniae infection in patients with cerebrovascular disease in the ICU
- Calycosin protects against chronic prostatitis in rats via inhibition of the p38MAPK/NF-κB pathway
- Pan-cancer analysis of the PDE4DIP gene with potential prognostic and immunotherapeutic values in multiple cancers including acute myeloid leukemia
- The safety and immunogenicity to inactivated COVID-19 vaccine in patients with hyperlipemia
- Circ-UBR4 regulates the proliferation, migration, inflammation, and apoptosis in ox-LDL-induced vascular smooth muscle cells via miR-515-5p/IGF2 axis
- Clinical characteristics of current COVID-19 rehabilitation outpatients in China
- Luteolin alleviates ulcerative colitis in rats via regulating immune response, oxidative stress, and metabolic profiling
- miR-199a-5p inhibits aortic valve calcification by targeting ATF6 and GRP78 in valve interstitial cells
- The application of iliac fascia space block combined with esketamine intravenous general anesthesia in PFNA surgery of the elderly: A prospective, single-center, controlled trial
- Elevated blood acetoacetate levels reduce major adverse cardiac and cerebrovascular events risk in acute myocardial infarction
- The effects of progesterone on the healing of obstetric anal sphincter damage in female rats
- Identification of cuproptosis-related genes for predicting the development of prostate cancer
- Lumican silencing ameliorates β-glycerophosphate-mediated vascular smooth muscle cell calcification by attenuating the inhibition of APOB on KIF2C activity
- Targeting PTBP1 blocks glutamine metabolism to improve the cisplatin sensitivity of hepatocarcinoma cells through modulating the mRNA stability of glutaminase
- A single center prospective study: Influences of different hip flexion angles on the measurement of lumbar spine bone mineral density by dual energy X-ray absorptiometry
- Clinical analysis of AN69ST membrane continuous venous hemofiltration in the treatment of severe sepsis
- Antibiotics therapy combined with probiotics administered intravaginally for the treatment of bacterial vaginosis: A systematic review and meta-analysis
- Construction of a ceRNA network to reveal a vascular invasion associated prognostic model in hepatocellular carcinoma
- A pan-cancer analysis of STAT3 expression and genetic alterations in human tumors
- A prognostic signature based on seven T-cell-related cell clustering genes in bladder urothelial carcinoma
- Pepsin concentration in oral lavage fluid of rabbit reflux model constructed by dilating the lower esophageal sphincter
- The antihypertensive felodipine shows synergistic activity with immune checkpoint blockade and inhibits tumor growth via NFAT1 in LUSC
- Tanshinone IIA attenuates valvular interstitial cells’ calcification induced by oxidized low density lipoprotein via reducing endoplasmic reticulum stress
- AS-IV enhances the antitumor effects of propofol in NSCLC cells by inhibiting autophagy
- Establishment of two oxaliplatin-resistant gallbladder cancer cell lines and comprehensive analysis of dysregulated genes
- Trial protocol: Feasibility of neuromodulation with connectivity-guided intermittent theta-burst stimulation for improving cognition in multiple sclerosis
- LncRNA LINC00592 mediates the promoter methylation of WIF1 to promote the development of bladder cancer
- Factors associated with gastrointestinal dysmotility in critically ill patients
- Mechanisms by which spinal cord stimulation intervenes in atrial fibrillation: The involvement of the endothelin-1 and nerve growth factor/p75NTR pathways
- Analysis of two-gene signatures and related drugs in small-cell lung cancer by bioinformatics
- Silencing USP19 alleviates cigarette smoke extract-induced mitochondrial dysfunction in BEAS-2B cells by targeting FUNDC1
- Menstrual irregularities associated with COVID-19 vaccines among women in Saudi Arabia: A survey during 2022
- Ferroptosis involves in Schwann cell death in diabetic peripheral neuropathy
- The effect of AQP4 on tau protein aggregation in neurodegeneration and persistent neuroinflammation after cerebral microinfarcts
- Activation of UBEC2 by transcription factor MYBL2 affects DNA damage and promotes gastric cancer progression and cisplatin resistance
- Analysis of clinical characteristics in proximal and distal reflux monitoring among patients with gastroesophageal reflux disease
- Exosomal circ-0020887 and circ-0009590 as novel biomarkers for the diagnosis and prediction of short-term adverse cardiovascular outcomes in STEMI patients
- Upregulated microRNA-429 confers endometrial stromal cell dysfunction by targeting HIF1AN and regulating the HIF1A/VEGF pathway
- Bibliometrics and knowledge map analysis of ultrasound-guided regional anesthesia
- Knockdown of NUPR1 inhibits angiogenesis in lung cancer through IRE1/XBP1 and PERK/eIF2α/ATF4 signaling pathways
- D-dimer trends predict COVID-19 patient’s prognosis: A retrospective chart review study
- WTAP affects intracranial aneurysm progression by regulating m6A methylation modification
- Using of endoscopic polypectomy in patients with diagnosed malignant colorectal polyp – The cross-sectional clinical study
- Anti-S100A4 antibody administration alleviates bronchial epithelial–mesenchymal transition in asthmatic mice
- Prognostic evaluation of system immune-inflammatory index and prognostic nutritional index in double expressor diffuse large B-cell lymphoma
- Prevalence and antibiogram of bacteria causing urinary tract infection among patients with chronic kidney disease
- Reactive oxygen species within the vaginal space: An additional promoter of cervical intraepithelial neoplasia and uterine cervical cancer development?
- Identification of disulfidptosis-related genes and immune infiltration in lower-grade glioma
- A new technique for uterine-preserving pelvic organ prolapse surgery: Laparoscopic rectus abdominis hysteropexy for uterine prolapse by comparing with traditional techniques
- Self-isolation of an Italian long-term care facility during COVID-19 pandemic: A comparison study on care-related infectious episodes
- A comparative study on the overlapping effects of clinically applicable therapeutic interventions in patients with central nervous system damage
- Low intensity extracorporeal shockwave therapy for chronic pelvic pain syndrome: Long-term follow-up
- The diagnostic accuracy of touch imprint cytology for sentinel lymph node metastases of breast cancer: An up-to-date meta-analysis of 4,073 patients
- Mortality associated with Sjögren’s syndrome in the United States in the 1999–2020 period: A multiple cause-of-death study
- CircMMP11 as a prognostic biomarker mediates miR-361-3p/HMGB1 axis to accelerate malignant progression of hepatocellular carcinoma
- Analysis of the clinical characteristics and prognosis of adult de novo acute myeloid leukemia (none APL) with PTPN11 mutations
- KMT2A maintains stemness of gastric cancer cells through regulating Wnt/β-catenin signaling-activated transcriptional factor KLF11
- Evaluation of placental oxygenation by near-infrared spectroscopy in relation to ultrasound maturation grade in physiological term pregnancies
- The role of ultrasonographic findings for PIK3CA-mutated, hormone receptor-positive, human epidermal growth factor receptor-2-negative breast cancer
- Construction of immunogenic cell death-related molecular subtypes and prognostic signature in colorectal cancer
- Long-term prognostic value of high-sensitivity cardiac troponin-I in patients with idiopathic dilated cardiomyopathy
- Establishing a novel Fanconi anemia signaling pathway-associated prognostic model and tumor clustering for pediatric acute myeloid leukemia patients
- Integrative bioinformatics analysis reveals STAT2 as a novel biomarker of inflammation-related cardiac dysfunction in atrial fibrillation
- Adipose-derived stem cells repair radiation-induced chronic lung injury via inhibiting TGF-β1/Smad 3 signaling pathway
- Real-world practice of idiopathic pulmonary fibrosis: Results from a 2000–2016 cohort
- lncRNA LENGA sponges miR-378 to promote myocardial fibrosis in atrial fibrillation
- Diagnostic value of urinary Tamm-Horsfall protein and 24 h urine osmolality for recurrent calcium oxalate stones of the upper urinary tract: Cross-sectional study
- The value of color Doppler ultrasonography combined with serum tumor markers in differential diagnosis of gastric stromal tumor and gastric cancer
- The spike protein of SARS-CoV-2 induces inflammation and EMT of lung epithelial cells and fibroblasts through the upregulation of GADD45A
- Mycophenolate mofetil versus cyclophosphamide plus in patients with connective tissue disease-associated interstitial lung disease: Efficacy and safety analysis
- MiR-1278 targets CALD1 and suppresses the progression of gastric cancer via the MAPK pathway
- Metabolomic analysis of serum short-chain fatty acid concentrations in a mouse of MPTP-induced Parkinson’s disease after dietary supplementation with branched-chain amino acids
- Cimifugin inhibits adipogenesis and TNF-α-induced insulin resistance in 3T3-L1 cells
- Predictors of gastrointestinal complaints in patients on metformin therapy
- Prescribing patterns in patients with chronic obstructive pulmonary disease and atrial fibrillation
- A retrospective analysis of the effect of latent tuberculosis infection on clinical pregnancy outcomes of in vitro fertilization–fresh embryo transferred in infertile women
- Appropriateness and clinical outcomes of short sustained low-efficiency dialysis: A national experience
- miR-29 regulates metabolism by inhibiting JNK-1 expression in non-obese patients with type 2 diabetes mellitus and NAFLD
- Clinical features and management of lymphoepithelial cyst
- Serum VEGF, high-sensitivity CRP, and cystatin-C assist in the diagnosis of type 2 diabetic retinopathy complicated with hyperuricemia
- ENPP1 ameliorates vascular calcification via inhibiting the osteogenic transformation of VSMCs and generating PPi
- Significance of monitoring the levels of thyroid hormone antibodies and glucose and lipid metabolism antibodies in patients suffer from type 2 diabetes
- The causal relationship between immune cells and different kidney diseases: A Mendelian randomization study
- Interleukin 33, soluble suppression of tumorigenicity 2, interleukin 27, and galectin 3 as predictors for outcome in patients admitted to intensive care units
- Identification of diagnostic immune-related gene biomarkers for predicting heart failure after acute myocardial infarction
- Long-term administration of probiotics prevents gastrointestinal mucosal barrier dysfunction in septic mice partly by upregulating the 5-HT degradation pathway
- miR-192 inhibits the activation of hepatic stellate cells by targeting Rictor
- Diagnostic and prognostic value of MR-pro ADM, procalcitonin, and copeptin in sepsis
- Review Articles
- Prenatal diagnosis of fetal defects and its implications on the delivery mode
- Electromagnetic fields exposure on fetal and childhood abnormalities: Systematic review and meta-analysis
- Characteristics of antibiotic resistance mechanisms and genes of Klebsiella pneumoniae
- Saddle pulmonary embolism in the setting of COVID-19 infection: A systematic review of case reports and case series
- Vitamin C and epigenetics: A short physiological overview
- Ebselen: A promising therapy protecting cardiomyocytes from excess iron in iron-overloaded thalassemia patients
- Aspirin versus LMWH for VTE prophylaxis after orthopedic surgery
- Mechanism of rhubarb in the treatment of hyperlipidemia: A recent review
- Surgical management and outcomes of traumatic global brachial plexus injury: A concise review and our center approach
- The progress of autoimmune hepatitis research and future challenges
- METTL16 in human diseases: What should we do next?
- New insights into the prevention of ureteral stents encrustation
- VISTA as a prospective immune checkpoint in gynecological malignant tumors: A review of the literature
- Case Reports
- Mycobacterium xenopi infection of the kidney and lymph nodes: A case report
- Genetic mutation of SLC6A20 (c.1072T > C) in a family with nephrolithiasis: A case report
- Chronic hepatitis B complicated with secondary hemochromatosis was cured clinically: A case report
- Liver abscess complicated with multiple organ invasive infection caused by hematogenous disseminated hypervirulent Klebsiella pneumoniae: A case report
- Urokinase-based lock solutions for catheter salvage: A case of an upcoming kidney transplant recipient
- Two case reports of maturity-onset diabetes of the young type 3 caused by the hepatocyte nuclear factor 1α gene mutation
- Immune checkpoint inhibitor-related pancreatitis: What is known and what is not
- Does total hip arthroplasty result in intercostal nerve injury? A case report and literature review
- Clinicopathological characteristics and diagnosis of hepatic sinusoidal obstruction syndrome caused by Tusanqi – Case report and literature review
- Synchronous triple primary gastrointestinal malignant tumors treated with laparoscopic surgery: A case report
- CT-guided percutaneous microwave ablation combined with bone cement injection for the treatment of transverse metastases: A case report
- Malignant hyperthermia: Report on a successful rescue of a case with the highest temperature of 44.2°C
- Anesthetic management of fetal pulmonary valvuloplasty: A case report
- Rapid Communication
- Impact of COVID-19 lockdown on glycemic levels during pregnancy: A retrospective analysis
- Erratum
- Erratum to “Inhibition of miR-21 improves pulmonary vascular responses in bronchopulmonary dysplasia by targeting the DDAH1/ADMA/NO pathway”
- Erratum to: “Fer exacerbates renal fibrosis and can be targeted by miR-29c-3p”
- Retraction
- Retraction of “Study to compare the effect of casirivimab and imdevimab, remdesivir, and favipiravir on progression and multi-organ function of hospitalized COVID-19 patients”
- Retraction of “circ_0062491 alleviates periodontitis via the miR-142-5p/IGF1 axis”
- Retraction of “miR-223-3p alleviates TGF-β-induced epithelial-mesenchymal transition and extracellular matrix deposition by targeting SP3 in endometrial epithelial cells”
- Retraction of “SLCO4A1-AS1 mediates pancreatic cancer development via miR-4673/KIF21B axis”
- Retraction of “circRNA_0001679/miR-338-3p/DUSP16 axis aggravates acute lung injury”
- Retraction of “lncRNA ACTA2-AS1 inhibits malignant phenotypes of gastric cancer cells”
- Special issue Linking Pathobiological Mechanisms to Clinical Application for cardiovascular diseases
- Effect of cardiac rehabilitation therapy on depressed patients with cardiac insufficiency after cardiac surgery
- Special issue The evolving saga of RNAs from bench to bedside - Part I
- FBLIM1 mRNA is a novel prognostic biomarker and is associated with immune infiltrates in glioma
- Special Issue Computational Intelligence Methodologies Meets Recurrent Cancers - Part III
- Development of a machine learning-based signature utilizing inflammatory response genes for predicting prognosis and immune microenvironment in ovarian cancer
Artikel in diesem Heft
- Research Articles
- Exosomes derived from mesenchymal stem cells overexpressing miR-210 inhibits neuronal inflammation and contribute to neurite outgrowth through modulating microglia polarization
- Current situation of acute ST-segment elevation myocardial infarction in a county hospital chest pain center during an epidemic of novel coronavirus pneumonia
- circ-IARS depletion inhibits the progression of non-small-cell lung cancer by circ-IARS/miR-1252-5p/HDGF ceRNA pathway
- circRNA ITGA7 restrains growth and enhances radiosensitivity by up-regulating SMAD4 in colorectal carcinoma
- WDR79 promotes aerobic glycolysis of pancreatic ductal adenocarcinoma (PDAC) by the suppression of SIRT4
- Up-regulation of collagen type V alpha 2 (COL5A2) promotes malignant phenotypes in gastric cancer cell via inducing epithelial–mesenchymal transition (EMT)
- Inhibition of TERC inhibits neural apoptosis and inflammation in spinal cord injury through Akt activation and p-38 inhibition via the miR-34a-5p/XBP-1 axis
- 3D-printed polyether-ether-ketone/n-TiO2 composite enhances the cytocompatibility and osteogenic differentiation of MC3T3-E1 cells by downregulating miR-154-5p
- Propofol-mediated circ_0000735 downregulation restrains tumor growth by decreasing integrin-β1 expression in non-small cell lung cancer
- PVT1/miR-16/CCND1 axis regulates gastric cancer progression
- Silencing of circ_002136 sensitizes gastric cancer to paclitaxel by targeting the miR-16-5p/HMGA1 axis
- Short-term outcomes after simultaneous gastrectomy plus cholecystectomy in gastric cancer: A pooling up analysis
- SCARA5 inhibits oral squamous cell carcinoma via inactivating the STAT3 and PI3K/AKT signaling pathways
- Molecular mechanism by which the Notch signaling pathway regulates autophagy in a rat model of pulmonary fibrosis in pigeon breeder’s lung
- lncRNA TPT1-AS1 promotes cell migration and invasion in esophageal squamous-cell carcinomas by regulating the miR-26a/HMGA1 axis
- SIRT1/APE1 promotes the viability of gastric cancer cells by inhibiting p53 to suppress ferroptosis
- Glycoprotein non-metastatic melanoma B interacts with epidermal growth factor receptor to regulate neural stem cell survival and differentiation
- Treatments for brain metastases from EGFR/ALK-negative/unselected NSCLC: A network meta-analysis
- Association of osteoporosis and skeletal muscle loss with serum type I collagen carboxyl-terminal peptide β glypeptide: A cross-sectional study in elder Chinese population
- circ_0000376 knockdown suppresses non-small cell lung cancer cell tumor properties by the miR-545-3p/PDPK1 pathway
- Delivery in a vertical birth chair supported by freedom of movement during labor: A randomized control trial
- UBE2J1 knockdown promotes cell apoptosis in endometrial cancer via regulating PI3K/AKT and MDM2/p53 signaling
- Metabolic resuscitation therapy in critically ill patients with sepsis and septic shock: A pilot prospective randomized controlled trial
- Lycopene ameliorates locomotor activity and urinary frequency induced by pelvic venous congestion in rats
- UHRF1-induced connexin26 methylation is involved in hearing damage triggered by intermittent hypoxia in neonatal rats
- LINC00511 promotes melanoma progression by targeting miR-610/NUCB2
- Ultra-high-performance liquid chromatography-tandem mass spectrometry analysis of serum metabolomic characteristics in people with different vitamin D levels
- Role of Jumonji domain-containing protein D3 and its inhibitor GSK-J4 in Hashimoto’s thyroiditis
- circ_0014736 induces GPR4 to regulate the biological behaviors of human placental trophoblast cells through miR-942-5p in preeclampsia
- Monitoring of sirolimus in the whole blood samples from pediatric patients with lymphatic anomalies
- Effects of osteogenic growth peptide C-terminal pentapeptide and its analogue on bone remodeling in an osteoporosis rat model
- A novel autophagy-related long non-coding RNAs signature predicting progression-free interval and I-131 therapy benefits in papillary thyroid carcinoma
- WGCNA-based identification of potential targets and pathways in response to treatment in locally advanced breast cancer patients
- Radiomics model using preoperative computed tomography angiography images to differentiate new from old emboli of acute lower limb arterial embolism
- Dysregulated lncRNAs are involved in the progress of myocardial infarction by constructing regulatory networks
- Single-arm trial to evaluate the efficacy and safety of baclofen in treatment of intractable hiccup caused by malignant tumor chemotherapy
- Genetic polymorphisms of MRPS30-DT and NINJ2 may influence lung cancer risk
- Efficacy of immune checkpoint inhibitors in patients with KRAS-mutant advanced non-small cell lung cancer: A retrospective analysis
- Pyroptosis-based risk score predicts prognosis and drug sensitivity in lung adenocarcinoma
- Upregulation of lncRNA LANCL1-AS1 inhibits the progression of non-small-cell lung cancer via the miR-3680-3p/GMFG axis
- CircRANBP17 modulated KDM1A to regulate neuroblastoma progression by sponging miR-27b-3p
- Exosomal miR-93-5p regulated the progression of osteoarthritis by targeting ADAMTS9
- Downregulation of RBM17 enhances cisplatin sensitivity and inhibits cell invasion in human hypopharyngeal cancer cells
- HDAC5-mediated PRAME regulates the proliferation, migration, invasion, and EMT of laryngeal squamous cell carcinoma via the PI3K/AKT/mTOR signaling pathway
- The association between sleep duration, quality, and nonalcoholic fatty liver disease: A cross-sectional study
- Myostatin silencing inhibits podocyte apoptosis in membranous nephropathy through Smad3/PKA/NOX4 signaling pathway
- A novel long noncoding RNA AC125257.1 facilitates colorectal cancer progression by targeting miR-133a-3p/CASC5 axis
- Impact of omicron wave and associated control measures in Shanghai on health management and psychosocial well-being of patients with chronic conditions
- Clinicopathological characteristics and prognosis of young patients aged ≤45 years old with non-small cell lung cancer
- TMT-based comprehensive proteomic profiling identifies serum prognostic signatures of acute myeloid leukemia
- The dose limits of teeth protection for patients with nasopharyngeal carcinoma undergoing radiotherapy based on the early oral health-related quality of life
- miR-30b-5p targeting GRIN2A inhibits hippocampal damage in epilepsy
- Long non-coding RNA AL137789.1 promoted malignant biological behaviors and immune escape of pancreatic carcinoma cells
- IRF6 and FGF1 polymorphisms in non-syndromic cleft lip with or without cleft palate in the Polish population
- Comprehensive analysis of the role of SFXN family in breast cancer
- Efficacy of bronchoscopic intratumoral injection of endostar and cisplatin in lung squamous cell carcinoma patients underwent conventional chemoradiotherapy
- Silencing of long noncoding RNA MIAT inhibits the viability and proliferation of breast cancer cells by promoting miR-378a-5p expression
- AG1024, an IGF-1 receptor inhibitor, ameliorates renal injury in rats with diabetic nephropathy via the SOCS/JAK2/STAT pathway
- Downregulation of KIAA1199 alleviated the activation, proliferation, and migration of hepatic stellate cells by the inhibition of epithelial–mesenchymal transition
- Exendin-4 regulates the MAPK and WNT signaling pathways to alleviate the osteogenic inhibition of periodontal ligament stem cells in a high glucose environment
- Inhibition of glycolysis represses the growth and alleviates the endoplasmic reticulum stress of breast cancer cells by regulating TMTC3
- The function of lncRNA EMX2OS/miR-653-5p and its regulatory mechanism in lung adenocarcinoma
- Tectorigenin alleviates the apoptosis and inflammation in spinal cord injury cell model through inhibiting insulin-like growth factor-binding protein 6
- Ultrasound examination supporting CT or MRI in the evaluation of cervical lymphadenopathy in patients with irradiation-treated head and neck cancer
- F-box and WD repeat domain containing 7 inhibits the activation of hepatic stellate cells by degrading delta-like ligand 1 to block Notch signaling pathway
- Knockdown of circ_0005615 enhances the radiosensitivity of colorectal cancer by regulating the miR-665/NOTCH1 axis
- Long noncoding RNA Mhrt alleviates angiotensin II-induced cardiac hypertrophy phenotypes by mediating the miR-765/Wnt family member 7B pathway
- Effect of miR-499-5p/SOX6 axis on atrial fibrosis in rats with atrial fibrillation
- Cholesterol induces inflammation and reduces glucose utilization
- circ_0004904 regulates the trophoblast cell in preeclampsia via miR-19b-3p/ARRDC3 axis
- NECAB3 promotes the migration and invasion of liver cancer cells through HIF-1α/RIT1 signaling pathway
- The poor performance of cardiovascular risk scores in identifying patients with idiopathic inflammatory myopathies at high cardiovascular risk
- miR-2053 inhibits the growth of ovarian cancer cells by downregulating SOX4
- Nucleophosmin 1 associating with engulfment and cell motility protein 1 regulates hepatocellular carcinoma cell chemotaxis and metastasis
- α-Hederin regulates macrophage polarization to relieve sepsis-induced lung and liver injuries in mice
- Changes of microbiota level in urinary tract infections: A meta-analysis
- Identification of key enzalutamide-resistance-related genes in castration-resistant prostate cancer and verification of RAD51 functions
- Falls during oxaliplatin-based chemotherapy for gastrointestinal malignancies – (lessons learned from) a prospective study
- Outcomes of low-risk birth care during the Covid-19 pandemic: A cohort study from a tertiary care center in Lithuania
- Vitamin D protects intestines from liver cirrhosis-induced inflammation and oxidative stress by inhibiting the TLR4/MyD88/NF-κB signaling pathway
- Integrated transcriptome analysis identifies APPL1/RPS6KB2/GALK1 as immune-related metastasis factors in breast cancer
- Genomic analysis of immunogenic cell death-related subtypes for predicting prognosis and immunotherapy outcomes in glioblastoma multiforme
- Circular RNA Circ_0038467 promotes the maturation of miRNA-203 to increase lipopolysaccharide-induced apoptosis of chondrocytes
- An economic evaluation of fine-needle cytology as the primary diagnostic tool in the diagnosis of lymphadenopathy
- Midazolam impedes lung carcinoma cell proliferation and migration via EGFR/MEK/ERK signaling pathway
- Network pharmacology combined with molecular docking and experimental validation to reveal the pharmacological mechanism of naringin against renal fibrosis
- PTPN12 down-regulated by miR-146b-3p gene affects the malignant progression of laryngeal squamous cell carcinoma
- miR-141-3p accelerates ovarian cancer progression and promotes M2-like macrophage polarization by targeting the Keap1-Nrf2 pathway
- lncRNA OIP5-AS1 attenuates the osteoarthritis progression in IL-1β-stimulated chondrocytes
- Overexpression of LINC00607 inhibits cell growth and aggressiveness by regulating the miR-1289/EFNA5 axis in non-small-cell lung cancer
- Subjective well-being in informal caregivers during the COVID-19 pandemic
- Nrf2 protects against myocardial ischemia-reperfusion injury in diabetic rats by inhibiting Drp1-mediated mitochondrial fission
- Unfolded protein response inhibits KAT2B/MLKL-mediated necroptosis of hepatocytes by promoting BMI1 level to ubiquitinate KAT2B
- Bladder cancer screening: The new selection and prediction model
- circNFATC3 facilitated the progression of oral squamous cell carcinoma via the miR-520h/LDHA axis
- Prone position effect in intensive care patients with SARS-COV-2 pneumonia
- Clinical observation on the efficacy of Tongdu Tuina manipulation in the treatment of primary enuresis in children
- Dihydroartemisinin ameliorates cerebral I/R injury in rats via regulating VWF and autophagy-mediated SIRT1/FOXO1 pathway
- Knockdown of circ_0113656 assuages oxidized low-density lipoprotein-induced vascular smooth muscle cell injury through the miR-188-3p/IGF2 pathway
- Low Ang-(1–7) and high des-Arg9 bradykinin serum levels are correlated with cardiovascular risk factors in patients with COVID-19
- Effect of maternal age and body mass index on induction of labor with oral misoprostol for premature rupture of membrane at term: A retrospective cross-sectional study
- Potential protective effects of Huanglian Jiedu Decoction against COVID-19-associated acute kidney injury: A network-based pharmacological and molecular docking study
- Clinical significance of serum MBD3 detection in girls with central precocious puberty
- Clinical features of varicella-zoster virus caused neurological diseases detected by metagenomic next-generation sequencing
- Collagen treatment of complex anorectal fistula: 3 years follow-up
- LncRNA CASC15 inhibition relieves renal fibrosis in diabetic nephropathy through down-regulating SP-A by sponging to miR-424
- Efficacy analysis of empirical bismuth quadruple therapy, high-dose dual therapy, and resistance gene-based triple therapy as a first-line Helicobacter pylori eradication regimen – An open-label, randomized trial
- SMOC2 plays a role in heart failure via regulating TGF-β1/Smad3 pathway-mediated autophagy
- A prospective cohort study of the impact of chronic disease on fall injuries in middle-aged and older adults
- circRNA THBS1 silencing inhibits the malignant biological behavior of cervical cancer cells via the regulation of miR-543/HMGB2 axis
- hsa_circ_0000285 sponging miR-582-3p promotes neuroblastoma progression by regulating the Wnt/β-catenin signaling pathway
- Long non-coding RNA GNAS-AS1 knockdown inhibits proliferation and epithelial–mesenchymal transition of lung adenocarcinoma cells via the microRNA-433-3p/Rab3A axis
- lncRNA UCA1 regulates miR-132/Lrrfip1 axis to promote vascular smooth muscle cell proliferation
- Twenty-four-color full spectrum flow cytometry panel for minimal residual disease detection in acute myeloid leukemia
- Hsa-miR-223-3p participates in the process of anthracycline-induced cardiomyocyte damage by regulating NFIA gene
- Anti-inflammatory effect of ApoE23 on Salmonella typhimurium-induced sepsis in mice
- Analysis of somatic mutations and key driving factors of cervical cancer progression
- Hsa_circ_0028007 regulates the progression of nasopharyngeal carcinoma through the miR-1179/SQLE axis
- Variations in sexual function after laparoendoscopic single-site hysterectomy in women with benign gynecologic diseases
- Effects of pharmacological delay with roxadustat on multi-territory perforator flap survival in rats
- Analysis of heroin effects on calcium channels in rat cardiomyocytes based on transcriptomics and metabolomics
- Risk factors of recurrent bacterial vaginosis among women of reproductive age: A cross-sectional study
- Alkbh5 plays indispensable roles in maintaining self-renewal of hematopoietic stem cells
- Study to compare the effect of casirivimab and imdevimab, remdesivir, and favipiravir on progression and multi-organ function of hospitalized COVID-19 patients
- Correlation between microvessel maturity and ISUP grades assessed using contrast-enhanced transrectal ultrasonography in prostate cancer
- The protective effect of caffeic acid phenethyl ester in the nephrotoxicity induced by α-cypermethrin
- Norepinephrine alleviates cyclosporin A-induced nephrotoxicity by enhancing the expression of SFRP1
- Effect of RUNX1/FOXP3 axis on apoptosis of T and B lymphocytes and immunosuppression in sepsis
- The function of Foxp1 represses β-adrenergic receptor transcription in the occurrence and development of bladder cancer through STAT3 activity
- Risk model and validation of carbapenem-resistant Klebsiella pneumoniae infection in patients with cerebrovascular disease in the ICU
- Calycosin protects against chronic prostatitis in rats via inhibition of the p38MAPK/NF-κB pathway
- Pan-cancer analysis of the PDE4DIP gene with potential prognostic and immunotherapeutic values in multiple cancers including acute myeloid leukemia
- The safety and immunogenicity to inactivated COVID-19 vaccine in patients with hyperlipemia
- Circ-UBR4 regulates the proliferation, migration, inflammation, and apoptosis in ox-LDL-induced vascular smooth muscle cells via miR-515-5p/IGF2 axis
- Clinical characteristics of current COVID-19 rehabilitation outpatients in China
- Luteolin alleviates ulcerative colitis in rats via regulating immune response, oxidative stress, and metabolic profiling
- miR-199a-5p inhibits aortic valve calcification by targeting ATF6 and GRP78 in valve interstitial cells
- The application of iliac fascia space block combined with esketamine intravenous general anesthesia in PFNA surgery of the elderly: A prospective, single-center, controlled trial
- Elevated blood acetoacetate levels reduce major adverse cardiac and cerebrovascular events risk in acute myocardial infarction
- The effects of progesterone on the healing of obstetric anal sphincter damage in female rats
- Identification of cuproptosis-related genes for predicting the development of prostate cancer
- Lumican silencing ameliorates β-glycerophosphate-mediated vascular smooth muscle cell calcification by attenuating the inhibition of APOB on KIF2C activity
- Targeting PTBP1 blocks glutamine metabolism to improve the cisplatin sensitivity of hepatocarcinoma cells through modulating the mRNA stability of glutaminase
- A single center prospective study: Influences of different hip flexion angles on the measurement of lumbar spine bone mineral density by dual energy X-ray absorptiometry
- Clinical analysis of AN69ST membrane continuous venous hemofiltration in the treatment of severe sepsis
- Antibiotics therapy combined with probiotics administered intravaginally for the treatment of bacterial vaginosis: A systematic review and meta-analysis
- Construction of a ceRNA network to reveal a vascular invasion associated prognostic model in hepatocellular carcinoma
- A pan-cancer analysis of STAT3 expression and genetic alterations in human tumors
- A prognostic signature based on seven T-cell-related cell clustering genes in bladder urothelial carcinoma
- Pepsin concentration in oral lavage fluid of rabbit reflux model constructed by dilating the lower esophageal sphincter
- The antihypertensive felodipine shows synergistic activity with immune checkpoint blockade and inhibits tumor growth via NFAT1 in LUSC
- Tanshinone IIA attenuates valvular interstitial cells’ calcification induced by oxidized low density lipoprotein via reducing endoplasmic reticulum stress
- AS-IV enhances the antitumor effects of propofol in NSCLC cells by inhibiting autophagy
- Establishment of two oxaliplatin-resistant gallbladder cancer cell lines and comprehensive analysis of dysregulated genes
- Trial protocol: Feasibility of neuromodulation with connectivity-guided intermittent theta-burst stimulation for improving cognition in multiple sclerosis
- LncRNA LINC00592 mediates the promoter methylation of WIF1 to promote the development of bladder cancer
- Factors associated with gastrointestinal dysmotility in critically ill patients
- Mechanisms by which spinal cord stimulation intervenes in atrial fibrillation: The involvement of the endothelin-1 and nerve growth factor/p75NTR pathways
- Analysis of two-gene signatures and related drugs in small-cell lung cancer by bioinformatics
- Silencing USP19 alleviates cigarette smoke extract-induced mitochondrial dysfunction in BEAS-2B cells by targeting FUNDC1
- Menstrual irregularities associated with COVID-19 vaccines among women in Saudi Arabia: A survey during 2022
- Ferroptosis involves in Schwann cell death in diabetic peripheral neuropathy
- The effect of AQP4 on tau protein aggregation in neurodegeneration and persistent neuroinflammation after cerebral microinfarcts
- Activation of UBEC2 by transcription factor MYBL2 affects DNA damage and promotes gastric cancer progression and cisplatin resistance
- Analysis of clinical characteristics in proximal and distal reflux monitoring among patients with gastroesophageal reflux disease
- Exosomal circ-0020887 and circ-0009590 as novel biomarkers for the diagnosis and prediction of short-term adverse cardiovascular outcomes in STEMI patients
- Upregulated microRNA-429 confers endometrial stromal cell dysfunction by targeting HIF1AN and regulating the HIF1A/VEGF pathway
- Bibliometrics and knowledge map analysis of ultrasound-guided regional anesthesia
- Knockdown of NUPR1 inhibits angiogenesis in lung cancer through IRE1/XBP1 and PERK/eIF2α/ATF4 signaling pathways
- D-dimer trends predict COVID-19 patient’s prognosis: A retrospective chart review study
- WTAP affects intracranial aneurysm progression by regulating m6A methylation modification
- Using of endoscopic polypectomy in patients with diagnosed malignant colorectal polyp – The cross-sectional clinical study
- Anti-S100A4 antibody administration alleviates bronchial epithelial–mesenchymal transition in asthmatic mice
- Prognostic evaluation of system immune-inflammatory index and prognostic nutritional index in double expressor diffuse large B-cell lymphoma
- Prevalence and antibiogram of bacteria causing urinary tract infection among patients with chronic kidney disease
- Reactive oxygen species within the vaginal space: An additional promoter of cervical intraepithelial neoplasia and uterine cervical cancer development?
- Identification of disulfidptosis-related genes and immune infiltration in lower-grade glioma
- A new technique for uterine-preserving pelvic organ prolapse surgery: Laparoscopic rectus abdominis hysteropexy for uterine prolapse by comparing with traditional techniques
- Self-isolation of an Italian long-term care facility during COVID-19 pandemic: A comparison study on care-related infectious episodes
- A comparative study on the overlapping effects of clinically applicable therapeutic interventions in patients with central nervous system damage
- Low intensity extracorporeal shockwave therapy for chronic pelvic pain syndrome: Long-term follow-up
- The diagnostic accuracy of touch imprint cytology for sentinel lymph node metastases of breast cancer: An up-to-date meta-analysis of 4,073 patients
- Mortality associated with Sjögren’s syndrome in the United States in the 1999–2020 period: A multiple cause-of-death study
- CircMMP11 as a prognostic biomarker mediates miR-361-3p/HMGB1 axis to accelerate malignant progression of hepatocellular carcinoma
- Analysis of the clinical characteristics and prognosis of adult de novo acute myeloid leukemia (none APL) with PTPN11 mutations
- KMT2A maintains stemness of gastric cancer cells through regulating Wnt/β-catenin signaling-activated transcriptional factor KLF11
- Evaluation of placental oxygenation by near-infrared spectroscopy in relation to ultrasound maturation grade in physiological term pregnancies
- The role of ultrasonographic findings for PIK3CA-mutated, hormone receptor-positive, human epidermal growth factor receptor-2-negative breast cancer
- Construction of immunogenic cell death-related molecular subtypes and prognostic signature in colorectal cancer
- Long-term prognostic value of high-sensitivity cardiac troponin-I in patients with idiopathic dilated cardiomyopathy
- Establishing a novel Fanconi anemia signaling pathway-associated prognostic model and tumor clustering for pediatric acute myeloid leukemia patients
- Integrative bioinformatics analysis reveals STAT2 as a novel biomarker of inflammation-related cardiac dysfunction in atrial fibrillation
- Adipose-derived stem cells repair radiation-induced chronic lung injury via inhibiting TGF-β1/Smad 3 signaling pathway
- Real-world practice of idiopathic pulmonary fibrosis: Results from a 2000–2016 cohort
- lncRNA LENGA sponges miR-378 to promote myocardial fibrosis in atrial fibrillation
- Diagnostic value of urinary Tamm-Horsfall protein and 24 h urine osmolality for recurrent calcium oxalate stones of the upper urinary tract: Cross-sectional study
- The value of color Doppler ultrasonography combined with serum tumor markers in differential diagnosis of gastric stromal tumor and gastric cancer
- The spike protein of SARS-CoV-2 induces inflammation and EMT of lung epithelial cells and fibroblasts through the upregulation of GADD45A
- Mycophenolate mofetil versus cyclophosphamide plus in patients with connective tissue disease-associated interstitial lung disease: Efficacy and safety analysis
- MiR-1278 targets CALD1 and suppresses the progression of gastric cancer via the MAPK pathway
- Metabolomic analysis of serum short-chain fatty acid concentrations in a mouse of MPTP-induced Parkinson’s disease after dietary supplementation with branched-chain amino acids
- Cimifugin inhibits adipogenesis and TNF-α-induced insulin resistance in 3T3-L1 cells
- Predictors of gastrointestinal complaints in patients on metformin therapy
- Prescribing patterns in patients with chronic obstructive pulmonary disease and atrial fibrillation
- A retrospective analysis of the effect of latent tuberculosis infection on clinical pregnancy outcomes of in vitro fertilization–fresh embryo transferred in infertile women
- Appropriateness and clinical outcomes of short sustained low-efficiency dialysis: A national experience
- miR-29 regulates metabolism by inhibiting JNK-1 expression in non-obese patients with type 2 diabetes mellitus and NAFLD
- Clinical features and management of lymphoepithelial cyst
- Serum VEGF, high-sensitivity CRP, and cystatin-C assist in the diagnosis of type 2 diabetic retinopathy complicated with hyperuricemia
- ENPP1 ameliorates vascular calcification via inhibiting the osteogenic transformation of VSMCs and generating PPi
- Significance of monitoring the levels of thyroid hormone antibodies and glucose and lipid metabolism antibodies in patients suffer from type 2 diabetes
- The causal relationship between immune cells and different kidney diseases: A Mendelian randomization study
- Interleukin 33, soluble suppression of tumorigenicity 2, interleukin 27, and galectin 3 as predictors for outcome in patients admitted to intensive care units
- Identification of diagnostic immune-related gene biomarkers for predicting heart failure after acute myocardial infarction
- Long-term administration of probiotics prevents gastrointestinal mucosal barrier dysfunction in septic mice partly by upregulating the 5-HT degradation pathway
- miR-192 inhibits the activation of hepatic stellate cells by targeting Rictor
- Diagnostic and prognostic value of MR-pro ADM, procalcitonin, and copeptin in sepsis
- Review Articles
- Prenatal diagnosis of fetal defects and its implications on the delivery mode
- Electromagnetic fields exposure on fetal and childhood abnormalities: Systematic review and meta-analysis
- Characteristics of antibiotic resistance mechanisms and genes of Klebsiella pneumoniae
- Saddle pulmonary embolism in the setting of COVID-19 infection: A systematic review of case reports and case series
- Vitamin C and epigenetics: A short physiological overview
- Ebselen: A promising therapy protecting cardiomyocytes from excess iron in iron-overloaded thalassemia patients
- Aspirin versus LMWH for VTE prophylaxis after orthopedic surgery
- Mechanism of rhubarb in the treatment of hyperlipidemia: A recent review
- Surgical management and outcomes of traumatic global brachial plexus injury: A concise review and our center approach
- The progress of autoimmune hepatitis research and future challenges
- METTL16 in human diseases: What should we do next?
- New insights into the prevention of ureteral stents encrustation
- VISTA as a prospective immune checkpoint in gynecological malignant tumors: A review of the literature
- Case Reports
- Mycobacterium xenopi infection of the kidney and lymph nodes: A case report
- Genetic mutation of SLC6A20 (c.1072T > C) in a family with nephrolithiasis: A case report
- Chronic hepatitis B complicated with secondary hemochromatosis was cured clinically: A case report
- Liver abscess complicated with multiple organ invasive infection caused by hematogenous disseminated hypervirulent Klebsiella pneumoniae: A case report
- Urokinase-based lock solutions for catheter salvage: A case of an upcoming kidney transplant recipient
- Two case reports of maturity-onset diabetes of the young type 3 caused by the hepatocyte nuclear factor 1α gene mutation
- Immune checkpoint inhibitor-related pancreatitis: What is known and what is not
- Does total hip arthroplasty result in intercostal nerve injury? A case report and literature review
- Clinicopathological characteristics and diagnosis of hepatic sinusoidal obstruction syndrome caused by Tusanqi – Case report and literature review
- Synchronous triple primary gastrointestinal malignant tumors treated with laparoscopic surgery: A case report
- CT-guided percutaneous microwave ablation combined with bone cement injection for the treatment of transverse metastases: A case report
- Malignant hyperthermia: Report on a successful rescue of a case with the highest temperature of 44.2°C
- Anesthetic management of fetal pulmonary valvuloplasty: A case report
- Rapid Communication
- Impact of COVID-19 lockdown on glycemic levels during pregnancy: A retrospective analysis
- Erratum
- Erratum to “Inhibition of miR-21 improves pulmonary vascular responses in bronchopulmonary dysplasia by targeting the DDAH1/ADMA/NO pathway”
- Erratum to: “Fer exacerbates renal fibrosis and can be targeted by miR-29c-3p”
- Retraction
- Retraction of “Study to compare the effect of casirivimab and imdevimab, remdesivir, and favipiravir on progression and multi-organ function of hospitalized COVID-19 patients”
- Retraction of “circ_0062491 alleviates periodontitis via the miR-142-5p/IGF1 axis”
- Retraction of “miR-223-3p alleviates TGF-β-induced epithelial-mesenchymal transition and extracellular matrix deposition by targeting SP3 in endometrial epithelial cells”
- Retraction of “SLCO4A1-AS1 mediates pancreatic cancer development via miR-4673/KIF21B axis”
- Retraction of “circRNA_0001679/miR-338-3p/DUSP16 axis aggravates acute lung injury”
- Retraction of “lncRNA ACTA2-AS1 inhibits malignant phenotypes of gastric cancer cells”
- Special issue Linking Pathobiological Mechanisms to Clinical Application for cardiovascular diseases
- Effect of cardiac rehabilitation therapy on depressed patients with cardiac insufficiency after cardiac surgery
- Special issue The evolving saga of RNAs from bench to bedside - Part I
- FBLIM1 mRNA is a novel prognostic biomarker and is associated with immune infiltrates in glioma
- Special Issue Computational Intelligence Methodologies Meets Recurrent Cancers - Part III
- Development of a machine learning-based signature utilizing inflammatory response genes for predicting prognosis and immune microenvironment in ovarian cancer