
Revealing the effects of amino acid, organic acid, and phytohormones on the germination of tomato seeds under salinity stress
-
Faika Yarali Karakan
 ,
Riaz Ullah
,
Riaz Ullah

Abstract
Salinity accumulation poses a threat to the production and productivity of economically important crops such as tomatoes (Solanum lycopersicum L.). Currently, salt tolerance breeding programs have been limited by insufficient genetic and physiological knowledge of tolerance-related traits and a lack of an efficient selection domain. For that purpose, we aimed to determine the ability of tomato cultivars to tolerate salt based on seed traits by multiple biochemical pathways. First, we tested three tomato cultivars according to their response to different sodium chloride (NaCl) concentrations (0, 6.3, 9.8, 13.0, and 15.8 dS m−1) and then we analysed their amino acids, organic acids, and phytohormones. Considering the results of germination traits, it is possible to conclude that cultivar H-2274 was more tolerant to salt stress than others. As a result, multivariate discriminant analysis including principal component analysis and two-way hierarchical clustering analyses were constructed and demonstrated that tomato cultivars were separated from each other by the amino acid, organic acid, and phytohormone contents. Considering germination traits of tomato seeds, cv. ‘H-2274’ was more tolerant to salinity than others depending on high proline (29 pmol µl−1) and citric acid (568 ng µl−1) assays. Biochemical variability offers a valuable tool for investigating salt tolerance mechanisms in tomatoes, and it will be appreciated to find high-tolerant tomato cultivar(s) to saline conditions. Also, the findings of this study have significant potential for practical applications in agriculture, particularly in developing salt-tolerant tomato cultivars to enhance productivity in saline environments and address socio-economic challenges.
1 Introduction
Tomato (Solanum lycopersicum L.) is one of the most widely grown vegetables [1]. Tomatoes are widely consumed for their nutrition and health effects [2]. However, a variety of biotic or abiotic stress factors have a negative impact on tomato growth and development [3]. Among abiotic factors, salt stress is the major environmental stress, affecting about 23% of cultivated lands worldwide, especially in arid and semi-arid regions, and decreasing one-third of crop production [4,5]. Therefore, dealing with salinity is crucial in achieving sustainable agricultural practices and meeting current and future food demand globally. Many commercial tomato cultivars range from moderately sensitive to sensitive to salinity. Hereby, in the future decades, it is critical to choose or produce salinity-resistant cultivars.
Salinity stress, with sodium chloride (NaCl) being the predominant salt, has had a detrimental impact on tomato seed germination and early seedling growth, which are the most decisive stages in species establishment. In particular, salinity inhibits water uptake of plants by causing ionic toxicity and osmotic stress as well as reducing germination rates [3,5]. In addition, salinity affects other aspects of plant growth, such as biochemical metabolism. Hence, plants respond to salt through diverse strategies and approaches that modulate self-physiological and biochemical mechanisms, providing new insights into how plants adapt to saline conditions. Plants alter biochemical functions such as nitrogenous compounds, amino acids, and organic acids to help alleviate the osmotic stress induced by salinity [6,7,8].
Amino acids tend to accumulate in the tissues of plants when exposed to salt stress. For instance, proline enhances antioxidant machinery, maintains protein structures, and stabilizes cell membranes during salt stress [5]. Thus, stress proteins such as proline could be used as a selective parameter for salt resistance [7]. Organic acid metabolism is of primary importance at the cellular level for various biochemical pathways such as amino acid biosynthesis, energy production, and modifying plant adaptability to the environmental condition. When plants are subjected to salt stress, they synthesize short-chain organic acids to maintain cell turgor, allowing them to absorb excess sodium while producing organic acids [3,9]. Gong et al. [10] reported that NaCl increased oxalic, tartaric, and malic acids in tomato roots. In addition, determining the amount of amino acids and organic acids in seeds and revealing their effects on germination under salt stress may aid in the identification of cultivars or genotypes resistant to salt stress at the first selection stage of breeding studies. Similar to amino acids and organic acids, phytohormones play an important role in controlling plant responses to abiotic stress, allowing the plant to focus its own resources on stress resistance [11]. Plants upregulate the levels of endogenous phytohormones, such as abscisic acid (ABA), salicylic acid (SA), and ethylene under stress conditions [5,12]. In fact, ABA and ethylene are stress hormones with specific roles in regulating salt stress tolerance and resistance [6].
A number of studies have been conducted to investigate the effects of organic acids, amino acids, and phytohormones on seed germination and the growth of tomato plants or roots under salt stress. However, to the best of our knowledge, no studies have been conducted on the involvement of tomato seed biochemical components in germination parameters under salt stress. Focusing on this idea, to fill this gap, we aimed to determine the role of some biochemical components such as phytohormones, amino acids, and organic acids in tomato seed salinity resistance and also to investigate whether the biochemical content of seeds could be used to predict salinity resistance.
2 Materials and methods
2.1 Plant material
The seeds of three tomato cultivars (S. lycopersicum L. ‘Falcon,’ ‘Rio Grande,’ and cultivar H-2274) were selected in this study because of their strong plant structure [13]. These cultivars were obtained from various commercial seed companies in 2021.
2.2 Salt stress treatments
The seeds were sterilized with 5% sodium hypochlorite solution for 5 min, and then they were washed three times with distilled water [14]. A standard germination test was conducted using four replications of 50 seeds from each cultivar in petri dishes (with 9 cm diameter) and spread across two filter paper sheets moistened with 0 (control), 6.3, 9.8, 13.0, and 15.8 dS m−1 NaCl solutions. They were kept at 20°C in a growth chamber with a photoperiod of 9/15 h each day [15]. The petri dishes were placed in a complete randomized design with four replications. Visible-radicle protrusion (≥2 mm) was the criterion of germination. Germinated seeds were counted at 24-h intervals for 14 days. At the end of the standard germination test, the germination percentage in the first count (5th day), germination percentage (%), germination speed (%), and radicle length (cm) were determined. The germination percentage was calculated by dividing the number of germinated seeds in any Petri dishes by the total number of seeds, then multiplied by 100 [16,17]. Germination speed was calculated according to the equation (germination speed = germination percentage in 1st day/1 + … + germination percentage 10th day/10’) of Kaymak [18]. To assess the vigour, seeds were sown in plastic trays (20.5 × 11.0 × 6.2 cm) filled with sphagnum peat [19,20]. The plastic trays were placed in laboratory conditions for a period of 3 weeks. The trays were irrigated daily with five (0, 6.3, 9.8, 13.0, and 15.8 dS m−1) different NaCl solutions, which were derived from distilled water by the use of NaCl. In addition, small holes were drilled so that excess water can drain naturally from the bottom of the boxes. Counts started as seedlings began to emerge and continued until the end of the 3 weeks. Seedlings were considered to have emerged when their cotyledons were free of the soil surface.
2.3 Biochemical assessments
2.3.1 Amino acids
Amino acid content was analysed as described by Gunes et al. [21]. For extraction, 1 g seed samples were taken and placed into a tube; then, 0.1 N HCl solution was added to it, and then prepared samples homogenized with a homogenizer (IKA-Labortechnik, Ultra-turrax T25) for 1.5 min at 9,500 × g. The homogenate was incubated at 4°C for 12 h. Subsequently, the mixture was centrifuged for 50 min at 1,200 × g. The supernatant was separated and filtered, followed by a 0.22 μm membrane filter (Millipore, USA). The prepared filtrates were transferred to glass vials and used for high-pressure liquid chromatography (HPLC) analysis. The amino acid derivatives were determined using HPLC (Agilent-1200) equipped with a Zorbax Eclipse-AAA column (4.6 × 150 mm, 3.5 µm) and a UV visible detector. A (40 mM NaH2PO4 [pH 7.8]) and B (acetonitrile/methanol/water [45/45/10, v/v/v]) solutions were used gradient at a flow rate of 2 ml min−1 as mobile phase. Column temperature was set as 40°C, and readings were done at 254 nm. The amino acids were identified through comparison to standards. O-phthaldialdehyde, fluorenylmethyl-chloroformate, and 0.4 N borate were used for derivation processes in an autosampler. Arginine, histidine, isoleucine, leucine, lysine, methionine, phenylalanine, proline, threonine, and valine quantities from seed samples were determined as pmol µl−1 after a 26 min derivation process in HPLC.
2.3.2 Organic acids
To determine the organic acid content of tomato seed, a 1 g seed sample was homogenized with 10 ml of doubly distilled water using an IKA-Labortechnik, Ultra-turrax T25 homogenizer at 24,000 rpm for 20 s. After centrifugation at 1,200 rpm for 50 min, the supernatant was filtered with a 0.22 µm membrane filter, collected in glass vials, and then analysed with an Agilent-1200 HPLC equipped with a Zorbax Eclipse-AAA column (4.6 × 250 mm, 5 µm) and UV visible detector. The flow rate was 1 ml min−1, the column temperature was 25°C, and readings were taken at 220 nm. The results were calculated based on external citric acid, fumaric acid, malic acid, succinic acid, oxalic acid, and tartaric acid standards using 25 mM potassium phosphate (pH 2.5) as the mobile phase. Data were expressed as ng µl−1.
2.3.3 Phytohormones
The extraction and quantification were carried out following the method described by Gunes et al. [22] with partial modifications. The tomato tissue was ground in liquid nitrogen, and 80% methanol, which was held at −40°C, was added. Then, the mixture was homogenized with a homogenizer (Ultra-Turrax) for 10 min and then incubated in dark conditions for 24 h. Afterwards the solution was first filtered with filter paper and second filtered through a 0.45 m pore filter. The obtained samples were dried at 35°C using evaporator pumps. Subsequently, the dried samples were dissolved in 0.1 M KH2PO4 (pH 8.0). To separate fatty acids, the extracts were centrifuged at 5,000 × g at 4°C for 1 h. Then, 1 g of polyvinyl polypyrrolidone (PVPP) was added to the supernatant in order to separate phenolic and colour matters (Gunes et al., [22]). To separate PVPP from the supernatant, the supernatant was filtered through Whatman No. 1 filter paper, and the Sep-Pak C-18 (Waters) cartridge was used for further specific separation. After adsorption of phytohormones by cartridge, the solution was transferred to vials and directly injected into HPLC (Agilent 1200) equipped with C-18 column Zorbax Eclipse (4.6 × 150 mm, 3.5 µm), and a UV detector was used for separation and determinations. The mobile phase consisted of acetonitrile and water (13:87, v/v) at pH 4.98 with a flow rate of 1.2 ml min−1, and the column temperature was 25°C. The readings were done at 265 nm, gibberellic acid (GA3), SA, indole acetic acid (IAA), ABA, ethylene, and zeatin were determined and given as ng µl−1.
2.4 Statistical analysis
This experiment was designed as a randomized complete design, with each analysis replicated four times. All statistical analyses were performed using JMP pro version 14 (SAS Institute, NC, USA). Significant differences between NaCl treatments were evaluated by one-way analysis of variance (ANOVA). When significant effects were detected, the Tukey test was performed to compare the NaCl treatments within each cultivar at a significance level of 0.05. Principal component analysis (PCA) and hierarchical cluster analysis (HCA) were used to describe patterns of variation in the biochemical profile of cultivars.
3 Results and discussion
3.1 Effects of NaCl treatments on germination parameters of three tomato cultivars
In the experiment, different concentrations of NaCl showed a significant effect on the germination percentage in the first count, germination percentage and speed, vigour, and radicle length of the different tomato cultivars (Table 1 and Figure 1).
First count, germination percentage and speed, radicle length, and vigour in tomato cultivars under different NaCl treatments
| NaCl (dS m−1) | Falcon | H-2274 | Rio Grande | |
|---|---|---|---|---|
| First count (%) | Control | 70.66 ± 3.05bc* | 94.66 ± 1.15a | 79.33 ± 4.61b |
| 6.3 | 69.33 ± 5.77bc | 93.33 ± 5.03a | 76.66 ± 1.15b | |
| 9.8 | 69.33 ± 5.77bc | 61.33 ± 3.05cd | 54.66 ± 2.30d | |
| 13.0 | 25.33 ± 8.32e | 8.00 ± 4.00f | 8.66 ± 2.30f | |
| 15.8 | 6.00 ± 5.29f | 2.66 ± 1.15f | 2.00 ± 0.00f | |
| Germination percentage (%) | Control | 84.00 ± 3.46ab | 100.00 ± 0.00a | 84.66 ± 2.30ab |
| 6.3 | 82.00 ± 2.00b | 98.66 ± 2.30a | 81.33 ± 1.15b | |
| 9.8 | 76.00 ± 4.00b | 91.33 ± 4.16ab | 80.66 ± 4.16b | |
| 13.0 | 54.00 ± 12.16c | 50.66 ± 1.15c | 39.33 ± 4.16c | |
| 15.8 | 12.66 ± 10.26d | 11.33 ± 7.57d | 8.00 ± 5.29d | |
| Germination speed (%) | Control | 35.67 ± 3.21a | 34.63 ± 1.90a | 22.76 ± 2.43cd |
| 6.3 | 28.14 ± 2.68bc | 33.28 ± 4.24ab | 21.15 ± 0.52d | |
| 9.8 | 22.41 ± 1.51cd | 22.92 ± 1.14cd | 16.87 ± 1.31d | |
| 13.0 | 9.90 ± 2.22e | 9.68 ± 0.54e | 5.92 ± 0.55ef | |
| 15.8 | 3.07 ± 2.60f | 1.80 ± 1.24f | 1.26 ± 0.67f | |
| Radicle length (cm) | Control | 14.38 ± 0.89a | 12.50 ± 0.00ab | 12.61 ± 1.56ab |
| 6.3 | 13.93 ± 1.11a | 11.53 ± 1.07ab | 11.51 ± 2.12ab | |
| 9.8 | 10.19 ± 1.58b | 10.08 ± 0.83b | 6.06 ± 1.46c | |
| 13.0 | 4.44 ± 0.78cd | 4.96 ± 0.57c | 1.49 ± 0.86de | |
| 15.8 | 1.21 ± 0.41de | 1.69 ± 0.32de | 0.46 ± 0.10e | |
| Vigour (%) | Control | 62.00 ± 0.00bcd | 95.33 ± 1.15a | 70.00 ± 3.46b |
| 6.3 | 59.33 ± 1.15cd | 93.33 ± 3.05a | 66.00 ± 0.00bc | |
| 9.8 | 58.67 ± 2.31cd | 91.33 ± 2.31a | 62.00 ± 0.00bcd | |
| 13.0 | 54.00 ± 0.00d | 71.00 ± 6.08b | 40.67 ± 5.77e | |
| 15.8 | 19.33 ± 8.08f | 31.33 ± 2.31e | 7.33 ± 2.31g |
*: Values within rows followed by different letters differ significantly by Tukey’s multiple range tests at 5%.
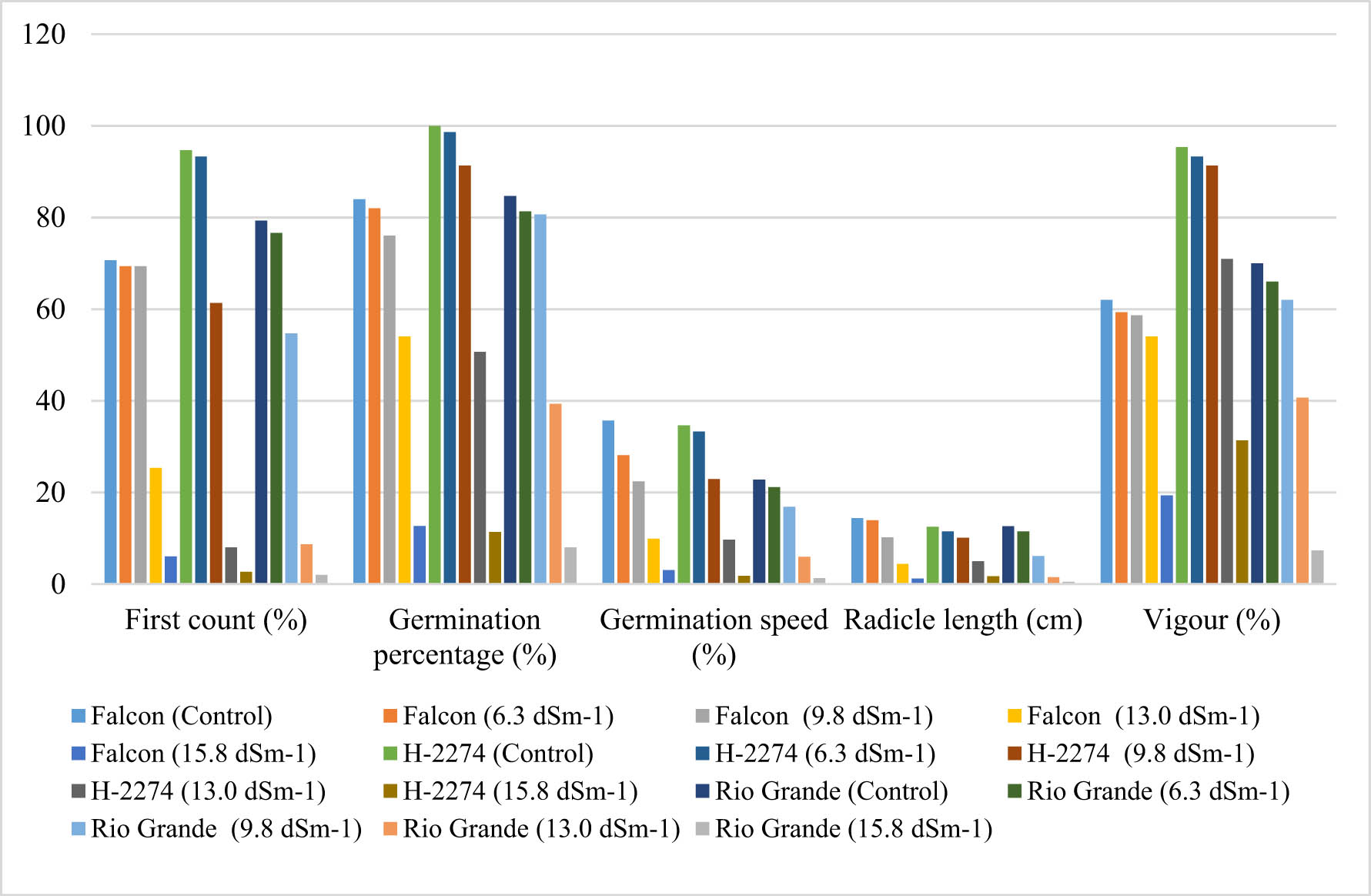
First count, germination percentage and speed, radicle length, and vigour in tomato cultivars under different NaCl treatments.
The first count of seeds ranged from 2.00 to 94.76%, with the germination percentage ranging from 8.00 to 100% among the cultivars. For each cultivar, as the salt concentration increased, the germination percentage and speed, vigour, and radicle length decreased. It has been reported that the most harmful effects of saline conditions are seen in the germination phase by inhibiting germination significantly [23,24]. As expected, the highest germination percentage and speed, vigour, and radicle length were obtained from the control, which was non-treated with salt. These results are in conformity with those reported by Sardoei and Mohammadi [14], Kumar et al. [23], and Seth and Kendurkar [25]. The germination percentage and speed, vigour, and radicle length of tomato cultivars in the control were 100.00, 35.67, 95.33, and 14.38% on average, respectively. Similarly, Kumar et al. [23] stated that the germination percentage was lowest (34.31%) in the cross EC-620428 × EC-620557, genotype NS-516 exhibited 87.93%, 85.66%, 85.54%, 82.87%, and EC-620428 × EC-520078 exhibited minimum 32.82, 26.24, 26.24, and 19.82% of germination percentage values at 50, 75, and 100 mM concentration of NaCl, respectively. When taking into account the NaCl concentrations, the highest germination percentage (54.00%) was determined in the cv. ‘H-2274,’ which tolerated salinity up to higher NaCl concentration (13.0 dS m−1), followed by the cv. ‘Falcon’ (50.66%), and the cv. ‘Rio Grande’ (39.33%). Considering the results of germination percentage, it could be concluded that the cv. ‘H-2274’ was more tolerant to salt stress than others based on the reduction in germination percentage at the highest NaCl concentration (15.8 dS m−1). It is worthy to mention that the cv. ‘Rio Grande’ represented more sensitivity by reducing drastically (90.56%) to saline conditions, especially in the highest concentrations (15.8 dS m−1). Consistent with our results, previous reports have stated that the salt tolerance of tomato cultivars varies depending on species manifested by physiological adaptations of the plants [1,6].
Apart from the harmful effects of salinity on germination percentage, salinity prolongs the complete germination elapsed time [26]. In this study, the germination speed of the studied tomato cultivars was affected by NaCl concentrations presented in Table 1. The obtained germination percentage results revealed that the cultivars ‘Falcon,’ ‘H-2274,’ and ‘Rio Grande’ exhibited reduction up to 72.00%, 72.04%, and 73.99%, respectively. A similar observation reported by Singh et al. [27] showed that 3% NaCl-treated tomato seeds required 100% more time to germinate than those treated with 1% NaCl. Adilu and Gebre [28] also found that the germination speed of tomato seeds was significantly affected by the combined effects of variety and salinity, and 4 dS m−1 NaCl treatment had an adverse effect on the germination speed of tomato cultivars.
Regarding vigour, data presented in Table 1 demonstrated that with each cultivar, increasing NaCl concentration led to progressively decreased vigour. Our results are in conformity with previous studies in tomato lines [29]. It is obvious that salinity stress caused a significant decrease in vigour, and a considerable decrease was recorded in the cv. ‘Rio Grande’ (90%) among cultivars at the highest NaCl concentration (15.8 dS m−1). While the cv. ‘Rio Grande’ was susceptible to salinity levels, and the cultivar ‘H-2274’ had higher tolerance to salinity compared to the other cultivar.
With respect to the radicle length, salinity significantly decreased the seedling growth, leading to a reduction in radicle length. Similar results were also observed in tomato and spinach [30]. At 15.8 dS m−1, the radicle length of the seedlings was reduced in cv. ‘Rio Grande’ (96.35%), and in the cv. ‘Falcon’ (91.58%) preceded by cv. ‘H-2274’ (86.48%). This is consistent with the findings by Seth and Kendurkar [25], which showed that salinity leads to a 77.64% reduction in root length at a higher level (100 mM NaCl).
3.2 Relationship between amino acids in tomato seeds and salt tolerance
The amino acid composition of three tomato cultivars was determined as depicted in Figures 1 and 2. A total of 10 amino acids including arginine, histidine, isoleucine, leucine, lysine, methionine, phenylalanine, proline, threonine, and valine have been identified in tomato cultivars (Figure 2). There were significant differences in amino acid concentration among cultivars. Arginine, threonine, and isoleucine were found in high amounts in tomato cultivars, accounting for more than 50.84, 55.46, and 56.71% of the total studied amino acid contents in cultivars ‘Falcon,’ ‘H-2274,’ and ‘Rio Grande,’ respectively. Other amino acids, histidine, leucine, lysine, methionine, phenylalanine, proline, and valine, were found in small amounts. The dendrogram and biplot graph also created and revealed that there was a clear separation among the cultivars (Figure 3). Figure 3 shows cluster I comprised cv. ‘Falcon’ was grouped individually, and the other cluster (cluster II) consisted of two cultivars (‘H-2274’ and ‘Rio Grande’), which were divided into two subgroups. In the case of PCA, it was calculated by using the scatter biplot method that PC1 (69.5%) and PC2 (30.5%) contents were constituted to be 100% of the total variation. In addition, the relationships between cultivars and amino acid content were computed by principal components biplot analysis and given in Figure 3.
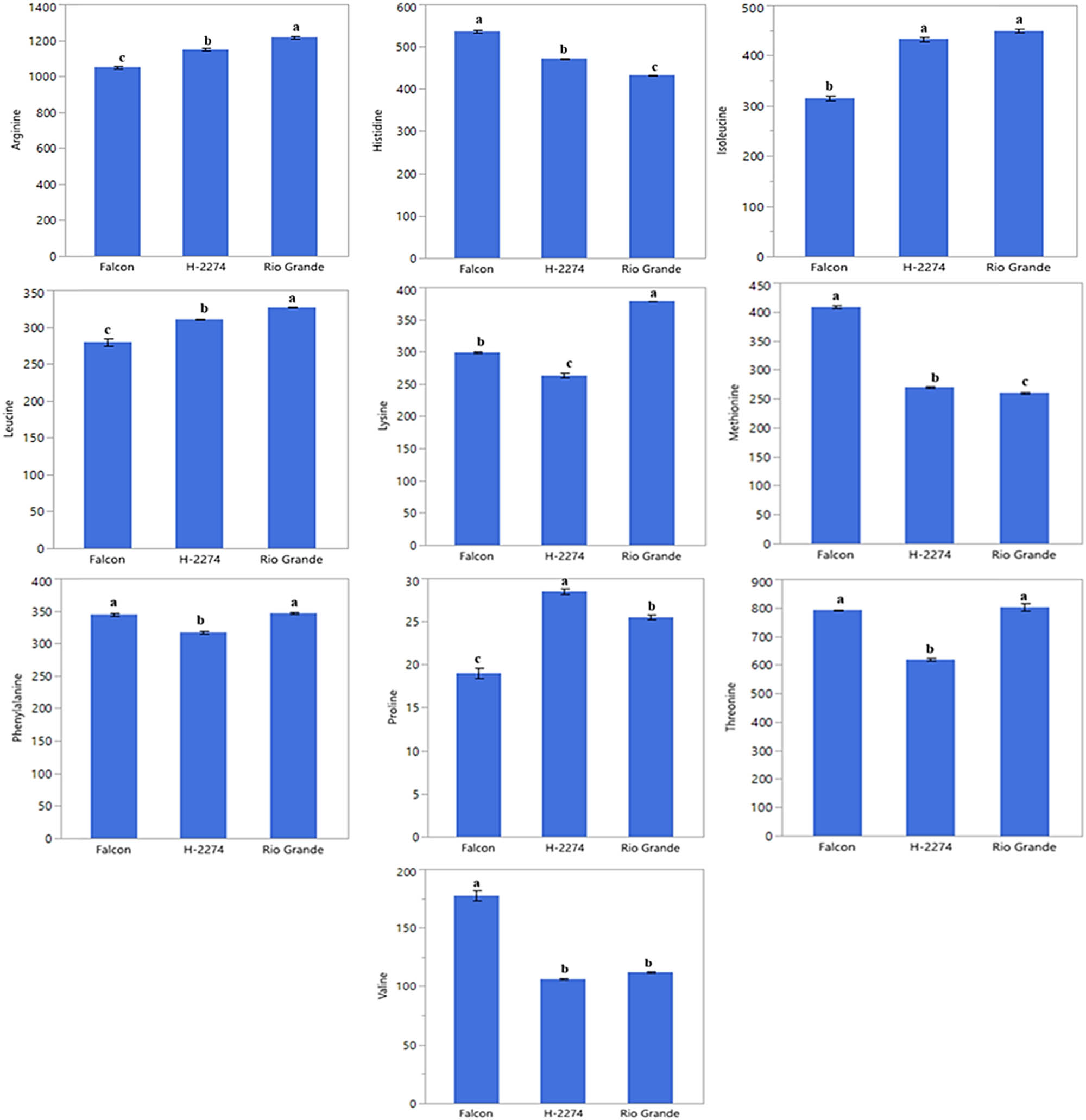
Detected amino acids in tomato cultivars (pmol µl−1) (the error bars represent confidence intervals; different letters indicate the significant results are those for which the p-value was lower than 0.05).
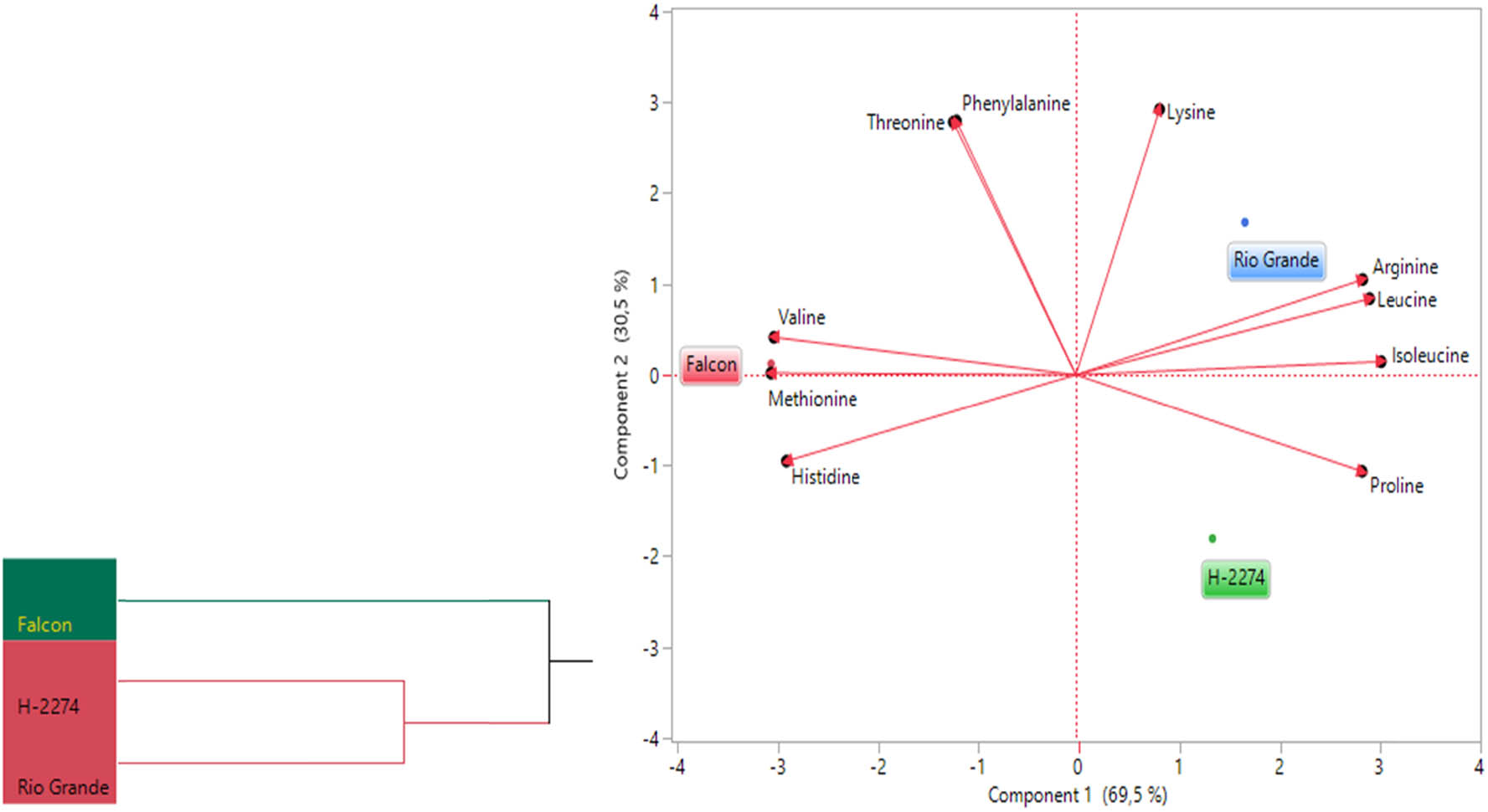
Dendogram obtained by two-way hierarchical clustering analysis (HCA) shows the distribution of tomato varieties according to amino acid content (left). The biplot plot graph obtained by principal component analysis shows the relationships between tomato cultivars and amino acids (right).
When the relationships among cultivars and amino acids are interpreted by means of a biplot graph, it can be clearly correlated relationship between them. In terms of PCA results, arginine, leucine, isoleucine, and proline were positively related to each other. Likewise, valine, methionine, and histidine were positively related to each other. However, these two groups demonstrated that the correlation relationship with these properties is negative and has a low degree of overlap. Conversely, the cultivar ‘Falcon’ presents higher values in histidine, methionine, and valine than the rest of the cultivar. In addition, cv. ‘Rio Grande’ is found to be rich in arginine, leucine, and lysine; conversely, the cv. ‘H-2274’ has a high value only in proline (Figures 3 and 8).
According to experience, to find any cultivar of species tolerant to saline conditions, the results of amino acid assays of these cultivars can be considered. Hence, as aforementioned in the germination experiments, the cultivar H-2274 was salt tolerant, the cv. ‘Falcon’ and cv. ‘Rio Grande’ were found as sensitive. Depending on the high proline content in the cv. ‘H-2274’ seeds, it can be an inference that it has a high tolerance to salinity. In this respect, previous studies have stated that proline is an important amino acid known to accumulate in plants when exposed to salinity, and it has been used as a tool to measure the saline stress [31]. Similar conclusions were drawn in tomato cultivars by Torre-González et al. [8] and Shin et al. [32], indicating that the amount of proline in the stem and leaf of tomato cultivars increased under saline conditions. Similarly, Abouelsaad et al. [33] reported that the level of proline in leaf tissues of tomatoes was exposed to NaCl increased by 4.71 fold. Similar correlations have been described by Gharsallah et al. [5], who reported an increase of proline concentration in the leaves of the tomato genotype ‘San Miguel,’ which is known to have high salt tolerance with increasing salt stress. Our results corroborate these findings related to the high salt-tolerant cultivars (cv. ‘Rio Grande’ and cv. ‘H-2274’), which have high arginine and proline content. As outlined by Zushi and Matsuzoeba [9], proline, phenylalanine, histidine, threonine, and arginine were higher in the cultivar ‘House Momotaro’ treated with the 100 mM salt, whereas proline and phenylalanine were higher in the cultivar ‘Mini Carol’ treated at the same concentration. Zhang et al. [3], in experiments involving phenylalanine, glutathione, cysteine, methionine, arginine, proline, alanine, aspartate, glutamate, glycine, serine, and threonine role in tomato, reported significantly enriched in the adaption to salt stress. According to these authors, arginine is a vital amino acid to occupy a precursor to the synthesis of other amino acids, and proline plays an important role during the salt stress response.
3.3 Relationship between organic acids in tomato seeds and salt tolerance
The analyses of organic acid content from seeds of three tomato cultivars identified six organic acids consisting of citric acid, fumaric acid, malic acid, succinic acid, oxalic acid, and tartaric acid by HPLC (Figure 4). We next determined similarities and dissimilarities between cultivars and organic acids, as graphed in Figure 5. The organic acids assay revealed that it ranged from 1,192 to 1,470 ng µl−1, from 458 to 568 ng µl−1, from 125 to 177 ng µl−1 with succinic, citric, and tartaric acid being the major constituents in tomato seeds. As for cultivars, the succinic acid values were 1,470, 1,389, and 1,192 ng µl−1 in the cv. ‘Rio Grande,’ ‘H-2274,’ and ‘Falcon,’ respectively. Citric acid (568 ng µl−1), succinic acid (1,470 ng µl−1), and tartaric acid (177 ng µl−1) were most widely detected in the ‘H-2274’ cultivar. The content of malic acid and fumaric acids was significantly higher in the ‘Falcon’ than in other cultivars. The cultivar ‘Rio Grande’ showed the highest oxalic acid content (Figures 5 and 8). The dendrogram and biplot graph presented two major clusters, and each cluster showed the existence of diversity and similarity in the cultivars based on the organic acids. As seen in Figure 5, cluster I comprised the ‘Falcon’ and the other cluster (cluster II) consisted of two cultivars (‘H-2274’ and ‘Rio Grande’), which were divided into two subgroups. In addition, the relationships between the cultivars and organic acid contents are given in Figure 5. According to the relations, a positive correlation was found among succinic acid, citric acid, and tartaric acid. Also, malic acid and fumaric acid were positively correlated with each other. It was no coincidence; Chen et al. [34] explored the mutual conversion process between fumaric acid and malic acid under different conditions. Additionally, malic acid, fumaric acid, and oxalic acid showed a negative and low correlation.
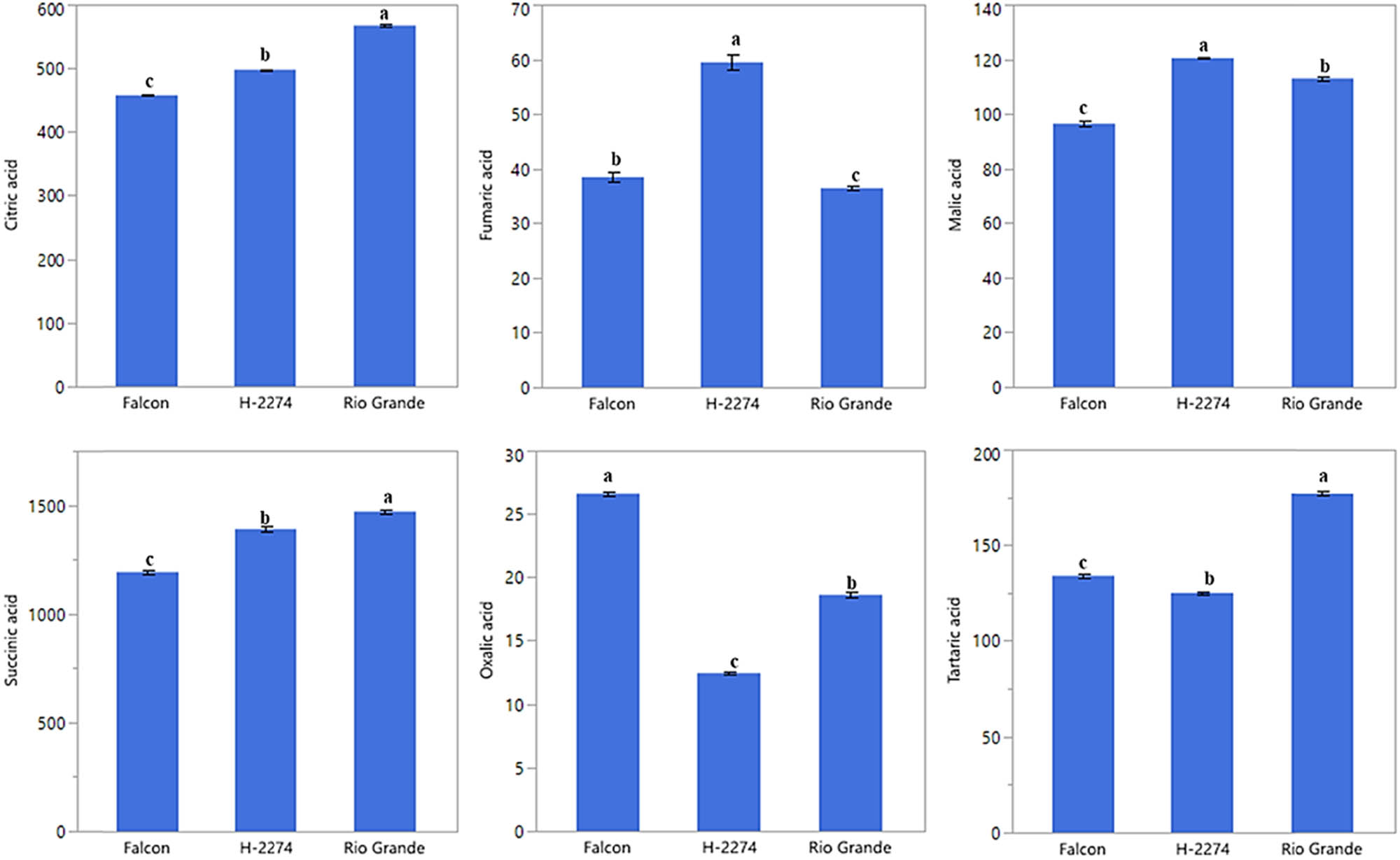
Detected organic acids in tomato cultivar seeds (ng µl−1) (the error bars represent confidence intervals; different letters indicate the significant results are those for which the p-value was lower than 0.05).
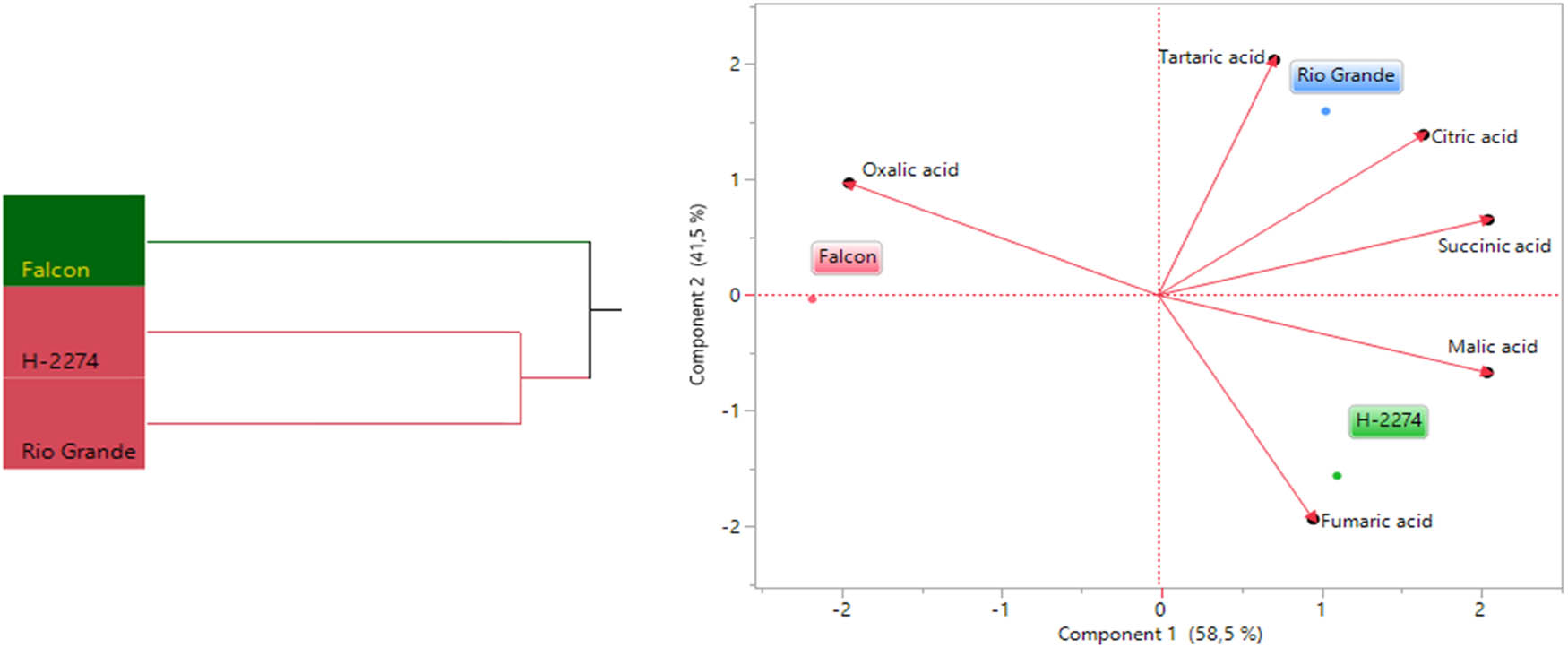
Dendogram obtained by two-way HCA shows the distribution of tomato varieties according to organic acid content (left). The biplot plot graph obtained by principal component analysis shows the relationships between tomato cultivars and organic acids (right).
Tolerance to salinity is a composite phenomenon involving a lot of metabolic pathway modulation [35]. Our study involving organic acid content in tomato seeds contributed important evidence in understanding the role of their tolerance to salt stress. A very common phenomenon of organic acids including oxalic acid, tartaric acid, malic acid, citric acid, and succinic acid is an accumulation of these, an inevitable consequence of salt stress in tomatoes, as mentioned in previous reports [10]. Comparing organic acid content, Zushi and Matsuzoe [9] determined the higher citric acid content in the salt-stressed fruit, but the malic acid content remained unchanged. Therefore, it could be said that the high amount of citric acid in the seed provides an advantage in salt response. When taking into account of aforesaid approaches, from the present study, it was noted that the cultivar ‘H-2274,’ having higher citric acid and malic acid had high salt tolerance.
3.4 Relationship between phytohormones in tomato seeds and salt tolerance
A total of six phytohormones consisting of ABA, ethylene, GA3, IAA, SA, and zeatin were identified in the seed of three tomato cultivars (Figure 6). Like amino acid and organic acid contents, these phytohormones are differentially altered in tomato seeds. GA3, SA, and zeatin were present in high quantities. The values of GA3, SA, and zeatin were 1,535, 1,609, and 1,870 ng µl−1; 79.15, 85.94, 85.94, and 92.89 ng µl−1; 49.07, 39.88, and 31.13 ng µl−1 in ‘Falcon,’ ‘H-2274,’ and ‘Rio Grande,’ respectively. The highest levels of ABA (0.89 ng µl−1), ethylene (0.67 ng µl−1), IAA (15.46 ng µl−1), and zeatin (49.07 ng µl−1) were found in the cultivar ‘Falcon’; the cultivar ‘Rio Grande’ had the higher (GA3) and SA with a mean of 1,870, 92.89 ng µl−1, respectively. To obtain the detailed separation of the detected phytohormones in this study, multivariate discriminant analysis PCA and HCA were also constructed. Based on the existence of diversity and similarity in the cultivars, we found two major cluster groups, cluster I comprised cv. ‘Falcon,’ and cluster II consisted of the two cultivars (‘H-2274’ and ‘Rio Grande’), which were divided into two subgroups (Figure 7). We further determined by using the scatter biplot method that PC1 (96.5%) and PC2 (3.52%) contents were constituted to be 100% of the total variation. In addition, the relationships between cultivars and phytohormones in tomato seeds and the distribution of phytohormones are given separately based on tomato cultivars (Figure 7). According to the visual classification of the phytohormones, there was a strict correlation among ABA, IAA, ethylene, and zeatin. Such a similar correlation was observed in GA3 and SA. However, both two groups showed negative and weak correlations with each other. Our results indicated that ABA, IAA, ethylene, and zeatin were the major phytohormones for the cv. ‘Falcon,’ whereas the cv. ‘Rio Grande’ was rich in SA, and GA3 (Figures 7 and 8). Among studied cultivars, the ‘Falcon’ can be differentiated from the ‘Rio Grande’ and ‘H-2274’ depending on the major abundance of phytohormones.
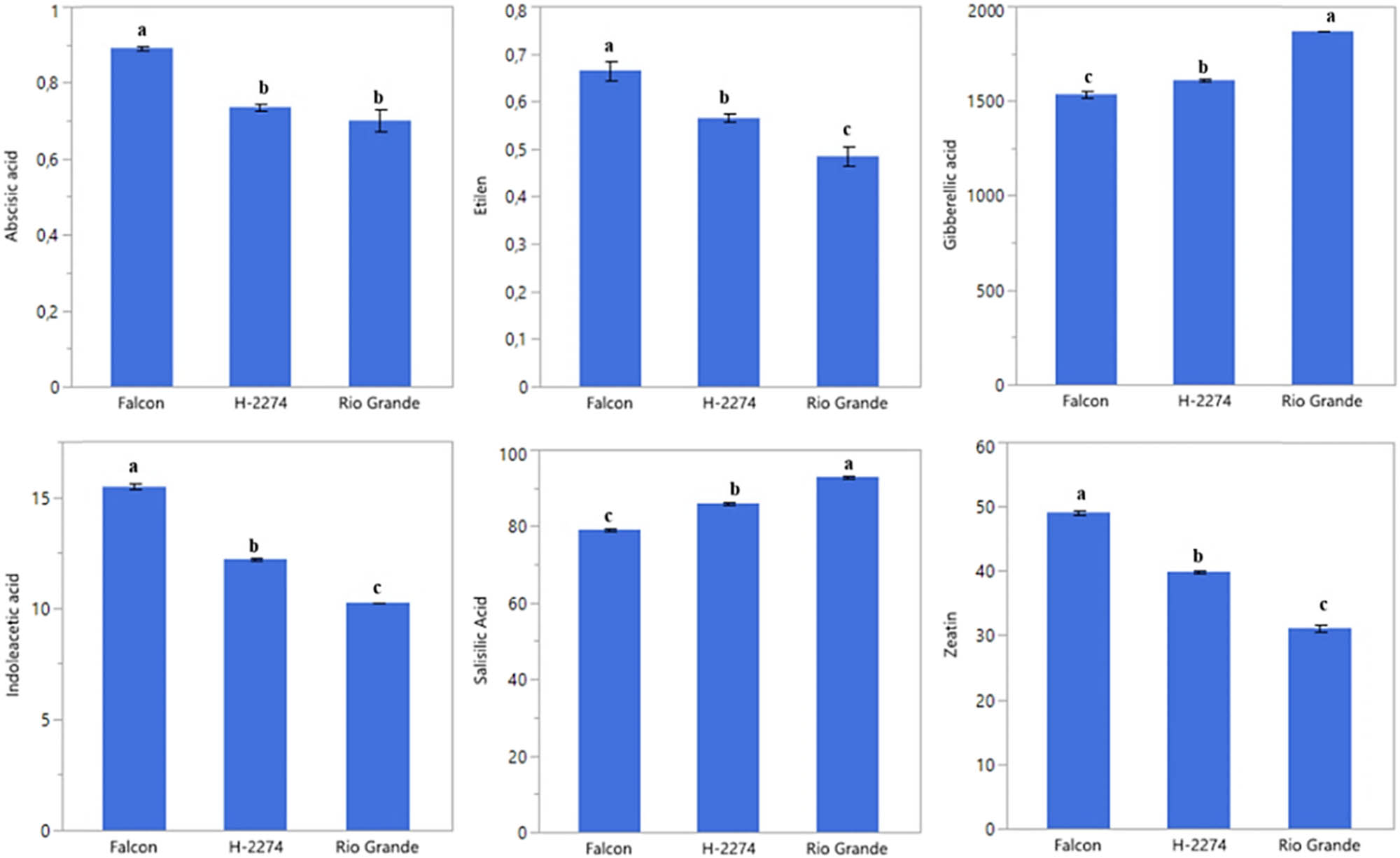
Detected phytohormones in the seed of tomato cultivars (ng µl−1) (the error bars represent confidence intervals; different letters indicate the significant results are those for which the p-value was lower than 0.05).
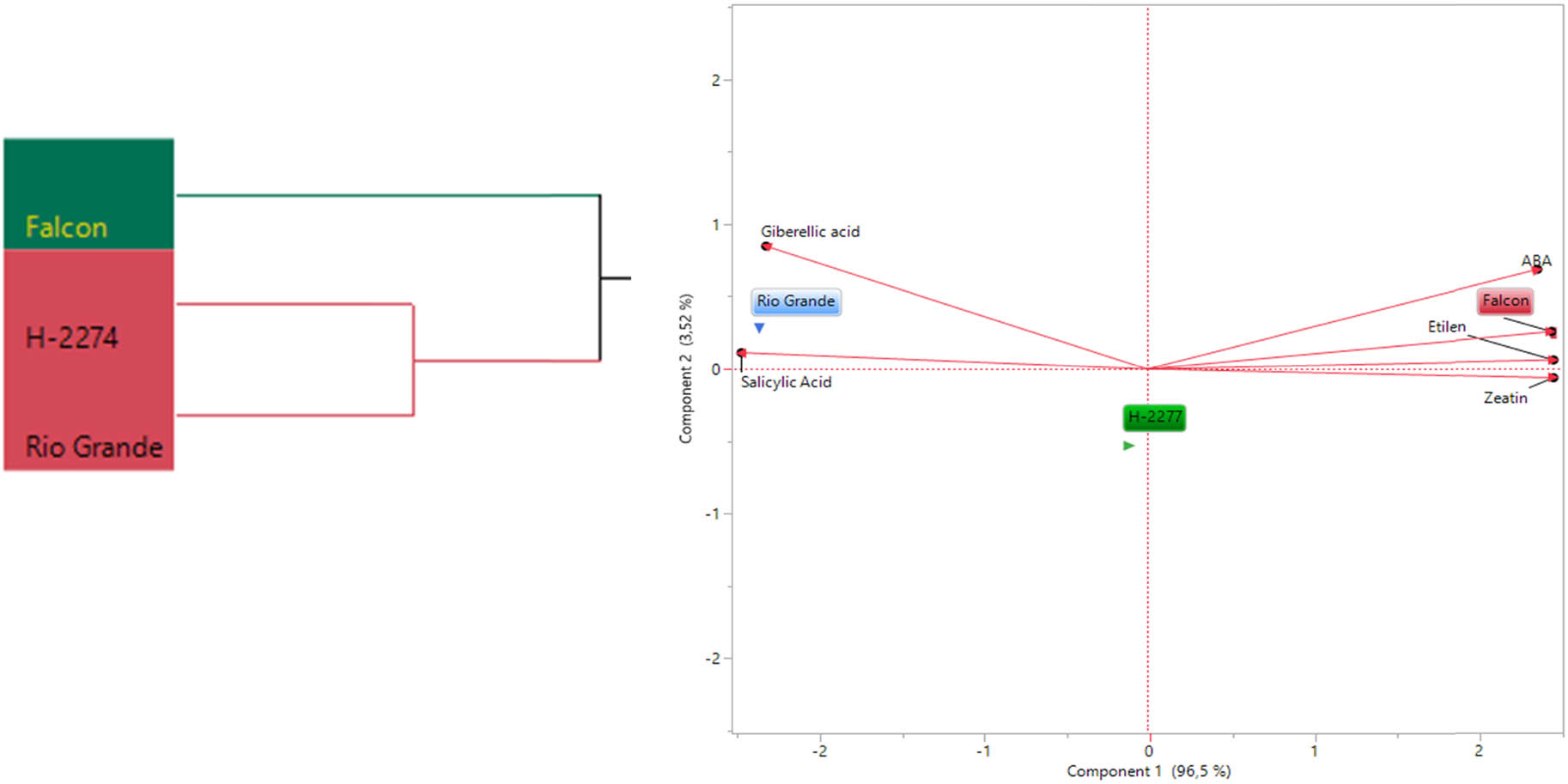
The dendogram obtained by two-way (HCA shows the distribution of tomato varieties according to phytohormones content (left). The biplot plot graph obtained by principal component analysis shows the relationships between tomato cultivars and phytohormones content (right).

The heat map of the principal component analysis was conducted to demonstrate the similarities and dissimilarities among the biochemical composition of the seed of three tomato cultivars in terms of the content of common detected amino acids, organic acids, and phytohormones (in the heat map, rows and columns are grouped by correlation distance and average linkage).
The role of phytohormones under saline conditions is crucial in regulating physiological responses, which will finally lead to adaptation to an unfavourable environment [36]. Based on these facts, previous works have conjectured that endogenous phytohormones were important in response to salt stress [11,37]. For example, ABA has a mediator role in responses to salt stress [12,38,39], and IAA controls almost all aspects of plant life, from seed germination to flowering stages [4]. Amjad et al. [6] reported that the salt-tolerant genotype had a significantly higher amount of ABA and ethylene than the salt-sensitive genotype under saline conditions. A similar result has also been reported by Babu et al. [4] found the amount of IAA and ABA increased as salt treatment increased. Interestingly, in our experiment, it was observed that the most abundant ABA, IAA, ethylene, and zeatin were found in cv. ‘Falcon’ while this cultivar was not resistant to saline conditions considering germination traits under salt stress. These inconsistent results could be explained by the global effect of the different metabolites in the seed of the salt tolerance of the cultivars. Furthermore, the germination percentage and germination speed are affected by the amount of GA3 and ethylene in the seed, especially in saline conditions. That is to say, the germination percentage of the seed is decreased when the seed has low GA3 content [27].
4 Conclusions
The selection of salt-tolerant cultivars could be a wise solution to minimize the harmful effects of salinity. Biochemical variability offers a valuable tool for investigating salt tolerance mechanisms in tomatoes, and it will be appreciated to find high-tolerant tomato cultivar(s) to saline conditions. Screening of the complex correlation of biochemical content is the most critical requirement to fill the gap in knowledge about the relationships between biochemical profile and seed germination characteristics. In the study, multivariate discriminant analyses were constructed and demonstrated that tomato cultivars were separated from each other by the amino acid, organic acid and phytohormone contents. Considering germination traits of tomato seeds, cv. ‘H-2274’ was more tolerant to salinity than others, depending on high proline and citric acid assays. Based on our results, the biochemical parameters of the seed species could be used for assuming tolerance to saline conditions, although it is not clear that tolerance is in critical stages of plant growth.
Acknowledgments
The authors wish to thank the Researchers Supporting Project number (RSP2024R45) at King Saud University Riyadh, Saudi Arabia, for financial support.
-
Funding information: This article was funded by project number (RSP2024R45) from King Saud University in Riyadh, Saudi Arabia, for financial support.
-
Author contributions: Conceptualization, H.Ç.K.; writing – original draft preparation, F.Y.K.; A.A.; S.E. and S.A.; data curation, H.Ç.K.; F.Y.K. and S.A.; validation, H.Ç.K.; F.Y. and S.A.; visualization, H.Ç.K.; F.Y. and S.A.; writing – review and editing, A.A.; F.Y.K.; S.A. and S.E.; investigation, H.Ç.K.; methodology, H.Ç.K.; and F.Y.K.; supervision, H.Ç.K. and S.E.; resource, H.Ç.K.; F.Y.K. and S.A.; software, A.A.; F.Y.K. and S.A.; formal analysis H.Ç.K.; F.Y.K.; S.A. and S.E; All authors have read and agreed to the published version of the manuscript.
-
Conflict of interest: Authors state no conflict of interest.
-
Data availability statement: The datasets generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
References
[1] Sane AK, Diallo B, Kane A, Sagna M, Sane D, Sy MO. In vitro germination and early vegetative growth of five tomato (Solanum lycopersicum L.) varieties under salt stress conditions. Am J Plant Sci. 2021;12:796–817.Search in Google Scholar
[2] Salehi B, Sharifi-Rad R, Sharopov F, Namiesnik J, Roointan A, Kamle M, et al. Beneficial effects and potential risks of tomato consumption for human health: An overview. Nutrition. 2019;62:201–8.Search in Google Scholar
[3] Zhang Z, Mao C, Shi Z, Kou X. The amino acid metabolic and carbohydrate metabolic pathway play important roles during salt-stress response in tomato. Front Plant Sci. 2017;8:1231.Search in Google Scholar
[4] Babu MA, Singh D, Gothandam KM. The Effect of salinity on growth, hormones and mineral elements in leaf and fruit of tomato cultivar PKM1. J Anim Plant Sci. 2012;22(1):159–64.Search in Google Scholar
[5] Gharsallah C, Fakhfakh H, Grubb D, Gorsane F. Effect of salt stress on ion concentration, proline content, antioxidant enzyme activities and gene expression in tomato cultivars. AOB Plant. 2016;8:plw055.Search in Google Scholar
[6] Amjad M, Akhtar J, Anwar-ul-Haq M, Yang A, Akhtar SS, Jacobsen SE. Integrating role of ethylene and ABA in tomato plants adaptation to salt stress. Sci Hortic. 2014;172:109–16.Search in Google Scholar
[7] Assouguem A, Joutei AB, Lahlali R, Kara M, Bari A, Ali EA, et al. Evaluation of the impact of two citrus plants on the variation of Panonychus citri (Acari: Tetranychidae) and beneficial phytoseiid mites. Open Life Sci. 2024;19(1):1–10.Search in Google Scholar
[8] Torre-González AD, Montesinos-Pereira D, Blasco B, Ruiz JM. Influence of the proline metabolism and glycine betaine on tolerance to salt stress in tomato (Solanum lycopersicum L.) commercial genotypes. J Plant Physiol. 2018;231:329–36.Search in Google Scholar
[9] Zushi K, Matsuzoe N. Metabolic profile of organoleptic and health-promoting qualities in two tomato cultivars subjected to salt stress and their interactions using correlation network analysis. Sci Hortic. 2015;184:8–17.Search in Google Scholar
[10] Gong B, Wen D, Bloszies S, Li X, Wie M, Yang F, et al. Comparative effects of NaCl and NaHCO3 stresses on respiratory metabolism, antioxidant system, nutritional status, and organic acid metabolism in tomato roots. Acta Physiol Plant. 2014;36:2167–81.Search in Google Scholar
[11] Ma Y, Dias MC, Freitas H. Drought and salinity stress responses and microbe-ınduced tolerance in plants. Front Plant Sci. 2020;11:591911.Search in Google Scholar
[12] Hu E, Liu M, Zhou R, Jiang F, Sun M, Wen J, et al. Relationship between melatonin and abscisic acid in response to salt stress of tomato. Sci Hortic. 2021;285:110176.Search in Google Scholar
[13] Demirkaya M. Improvement in tolerance to salt stress during tomato cultivation. Turkish J Biol. 2014;38:193–9.Search in Google Scholar
[14] Sardoei AS, Mohammadi GA. Study of salinity effect on germination of tomato (Lycopersicon esculentum L.) genotypes. Eur J Exp Biol. 2014;4(1):283–7.Search in Google Scholar
[15] ISTA. International rules for seed testing. The International Seed Testing Association (ISTA), 2018, 298p Secretariat Zürichstrasse 50, CH-8303 Bassersdorf, Switzerland.Search in Google Scholar
[16] Kaymak HÇ, Güvenç İ, Yaralı F, Dönmez MF. The effects of bio-priming with PGPR on germination of radish (Raphanus sativus L.) seeds under saline conditions. Turkish J Agric For. 2009;33(2):173–179.Search in Google Scholar
[17] Kaymak HÇ. Seed fatty acid profiles: Potential relations between seed germination under temperature stress in selected vegetable species. Acta Sci Pol Hortorum Cultus. 2014;13(2):119–33.Search in Google Scholar
[18] Kaymak HÇ. The relationships between seed fatty acids profile and seed germination in cucurbit species. Žemdirbystė–Agriculture. 2012;99(3):299–304.Search in Google Scholar
[19] Kaymak HÇ. Potential effect of seed fatty acid profile of pepper (Capsicum annuum L.) cultivars on germination at various temperatures. Žemdirbystė–Agriculture. 2014;101(3):321–6.Search in Google Scholar
[20] Kaymak HÇ, Akan S, Yarali Karakan F. Pathway among fatty acid profile, seed germination, and vigor of watermelon cultivars. Emirates J Food Agric. 2022;34(6):494–501.Search in Google Scholar
[21] Gunes A, Tarhan L, Ozdemir BS, Turan M, Sahin F. Effects of bacterial inoculated tuff material on yield and physiological parameters of grape (Vitis vinifera) plant. Erwerbs-Obstbau. 2021;63:43–51, 1.Search in Google Scholar
[22] Gunes A, Kordali S, Turan M, Bozhuyuk AU. Determination of antioxidant enzyme activity and phenolic contents of some species of the Asteraceae family from medicanal plants. Ind Crop Prod. 2019;137:208–13.Search in Google Scholar
[23] Kumar PA, Reddy NN, Lakshmi NJ. Screening tomato genotypes for salt tolerance. Int J Curr Microbiol App Sci. 2017;6(11):1037–49.Search in Google Scholar
[24] Tahir M, Zafar MM, Imran A, Hafeez MA, Rasheed MS, Mustafa HSB, et al. Response of tomato genotypes against salinity stress at germination and seedling stage. Nat Sci. 2018;16(4):10–7.Search in Google Scholar
[25] Seth R, Kendurkar SV. In vitro screening: An effective method for evaluation of commercial cultivars of tomato towards salinity stress. Int J Curr Microbiol App Sci. 2015;4(1):725–30.Search in Google Scholar
[26] Tanveer K, Gilani S, Hussain Z, Ishaq R, Adeel M, Ilyas N. Effect of salt stress on tomato plant and the role of calcium. J Plant Nutr. 2020;43(1):28–35.Search in Google Scholar
[27] Singh J, Divakar Sastry EV, Singh V. Effect of salinity on tomato (Lycopersicon esculentum Mill.) during seed germination stage. Physiol Mol Biol Plant. 2012;18:45–50.Search in Google Scholar
[28] Adilu GS, Gebre YG. Effect of salinity on seed germination of some tomato (Lycopersicon esculentum Mill.) varieties. J Aridland Agric. 2021;7:76–82.Search in Google Scholar
[29] Rofekuggaman M, Kubra K, Mahmood S. Effect of different salt concentrations (NaCl) on seed germination and seedling growth of tomato cv. BINA tomato-10. Asian Plant Res J. 2020;5(3):38–44.Search in Google Scholar
[30] Chakma P, Hossain MM, Rabbani MG. Effects of salinity stress on seed germination and seedling growth of tomato. J Bangladesh Agric Univ. 2019;17(4):490–9.Search in Google Scholar
[31] Alhasnawi AN. Role of proline in plant stress tolerance: A mini review. Res Crop. 2019;20(1):223–9.Search in Google Scholar
[32] Shin YK, Bhandari SR, Cho MC, Lee JG. Evaluation of chlorophyll fuorescence parameters and proline content in tomato seedlings grown under diferent salt stress conditions. Hortic Environ Biotechnol. 2020;61:433–43.Search in Google Scholar
[33] Abouelsaad I, Weihrauch D, Renault S. Effects of salt stress on the expression of key genes related to nitrogen assimilation and transport in the roots of the cultivated tomato and its wild salt-tolerant relative. Sci Hortic. 2016;211:70–8.Search in Google Scholar
[34] Chen W, Chen X, Yi S. Kinetic study on the preparation of fumaric acid from maleic acid by noncatalytic isomerization. ACS Omega. 2019;4(5):8274–81.Search in Google Scholar
[35] Das P, Manna I, Sil P, Bandyopadhyay M, Biswas AK. Exogenous silicon alters organic acid production and enzymatic activity of TCA cycle in two NaCl stressed indica rice cultivars. Plant Physiol Biochem. 2019;136:76–91.Search in Google Scholar
[36] Ryu H, Cho YG. Plant hormones in salt stress tolerance. J Plant Biol. 2015;58:147–55.Search in Google Scholar
[37] Fahad S, Hussain S, Matloob A, Khan FA, Khaliq A, Saud S, et al. Phytohormones and plant responses to salinity stress: A review. Plant Growth Regul. 2015;75:391–404.Search in Google Scholar
[38] Assouguem A, Lahlali R, Joutei AB, Kara M, Bari A, Aberkani K, et al. Assessing Panonychus ulmi (Acari: Tetranychidae) infestations and their key predators on Malus domestica borkh in varied ecological settings. Agronomy. 2024;14(3):457.Search in Google Scholar
[39] Yang AR, Yang T, Zhang H, Qi Y, Xing Y, Zhang N, et al. Hormone profiling and transcription analysis reveal a major role of ABA in tomato salt tolerance. Plant Physiol Biochem. 2014;77:23–34.Search in Google Scholar
© 2024 the author(s), published by De Gruyter
This work is licensed under the Creative Commons Attribution 4.0 International License.
Articles in the same Issue
- Biomedical Sciences
- Constitutive and evoked release of ATP in adult mouse olfactory epithelium
- LARP1 knockdown inhibits cultured gastric carcinoma cell cycle progression and metastatic behavior
- PEGylated porcine–human recombinant uricase: A novel fusion protein with improved efficacy and safety for the treatment of hyperuricemia and renal complications
- Research progress on ocular complications caused by type 2 diabetes mellitus and the function of tears and blepharons
- The role and mechanism of esketamine in preventing and treating remifentanil-induced hyperalgesia based on the NMDA receptor–CaMKII pathway
- Brucella infection combined with Nocardia infection: A case report and literature review
- Detection of serum interleukin-18 level and neutrophil/lymphocyte ratio in patients with antineutrophil cytoplasmic antibody-associated vasculitis and its clinical significance
- Ang-1, Ang-2, and Tie2 are diagnostic biomarkers for Henoch-Schönlein purpura and pediatric-onset systemic lupus erythematous
- PTTG1 induces pancreatic cancer cell proliferation and promotes aerobic glycolysis by regulating c-myc
- Role of serum B-cell-activating factor and interleukin-17 as biomarkers in the classification of interstitial pneumonia with autoimmune features
- Effectiveness and safety of a mumps containing vaccine in preventing laboratory-confirmed mumps cases from 2002 to 2017: A meta-analysis
- Low levels of sex hormone-binding globulin predict an increased breast cancer risk and its underlying molecular mechanisms
- A case of Trousseau syndrome: Screening, detection and complication
- Application of the integrated airway humidification device enhances the humidification effect of the rabbit tracheotomy model
- Preparation of Cu2+/TA/HAP composite coating with anti-bacterial and osteogenic potential on 3D-printed porous Ti alloy scaffolds for orthopedic applications
- Aquaporin-8 promotes human dermal fibroblasts to counteract hydrogen peroxide-induced oxidative damage: A novel target for management of skin aging
- Current research and evidence gaps on placental development in iron deficiency anemia
- Single-nucleotide polymorphism rs2910829 in PDE4D is related to stroke susceptibility in Chinese populations: The results of a meta-analysis
- Pheochromocytoma-induced myocardial infarction: A case report
- Kaempferol regulates apoptosis and migration of neural stem cells to attenuate cerebral infarction by O‐GlcNAcylation of β-catenin
- Sirtuin 5 regulates acute myeloid leukemia cell viability and apoptosis by succinylation modification of glycine decarboxylase
- Apigenin 7-glucoside impedes hypoxia-induced malignant phenotypes of cervical cancer cells in a p16-dependent manner
- KAT2A changes the function of endometrial stromal cells via regulating the succinylation of ENO1
- Current state of research on copper complexes in the treatment of breast cancer
- Exploring antioxidant strategies in the pathogenesis of ALS
- Helicobacter pylori causes gastric dysbacteriosis in chronic gastritis patients
- IL-33/soluble ST2 axis is associated with radiation-induced cardiac injury
- The predictive value of serum NLR, SII, and OPNI for lymph node metastasis in breast cancer patients with internal mammary lymph nodes after thoracoscopic surgery
- Carrying SNP rs17506395 (T > G) in TP63 gene and CCR5Δ32 mutation associated with the occurrence of breast cancer in Burkina Faso
- P2X7 receptor: A receptor closely linked with sepsis-associated encephalopathy
- Probiotics for inflammatory bowel disease: Is there sufficient evidence?
- Identification of KDM4C as a gene conferring drug resistance in multiple myeloma
- Microbial perspective on the skin–gut axis and atopic dermatitis
- Thymosin α1 combined with XELOX improves immune function and reduces serum tumor markers in colorectal cancer patients after radical surgery
- Highly specific vaginal microbiome signature for gynecological cancers
- Sample size estimation for AQP4-IgG seropositive optic neuritis: Retinal damage detection by optical coherence tomography
- The effects of SDF-1 combined application with VEGF on femoral distraction osteogenesis in rats
- Fabrication and characterization of gold nanoparticles using alginate: In vitro and in vivo assessment of its administration effects with swimming exercise on diabetic rats
- Mitigating digestive disorders: Action mechanisms of Mediterranean herbal active compounds
- Distribution of CYP2D6 and CYP2C19 gene polymorphisms in Han and Uygur populations with breast cancer in Xinjiang, China
- VSP-2 attenuates secretion of inflammatory cytokines induced by LPS in BV2 cells by mediating the PPARγ/NF-κB signaling pathway
- Factors influencing spontaneous hypothermia after emergency trauma and the construction of a predictive model
- Long-term administration of morphine specifically alters the level of protein expression in different brain regions and affects the redox state
- Application of metagenomic next-generation sequencing technology in the etiological diagnosis of peritoneal dialysis-associated peritonitis
- Clinical diagnosis, prevention, and treatment of neurodyspepsia syndrome using intelligent medicine
- Case report: Successful bronchoscopic interventional treatment of endobronchial leiomyomas
- Preliminary investigation into the genetic etiology of short stature in children through whole exon sequencing of the core family
- Cystic adenomyoma of the uterus: Case report and literature review
- Mesoporous silica nanoparticles as a drug delivery mechanism
- Dynamic changes in autophagy activity in different degrees of pulmonary fibrosis in mice
- Vitamin D deficiency and inflammatory markers in type 2 diabetes: Big data insights
- Lactate-induced IGF1R protein lactylation promotes proliferation and metabolic reprogramming of lung cancer cells
- Meta-analysis on the efficacy of allogeneic hematopoietic stem cell transplantation to treat malignant lymphoma
- Mitochondrial DNA drives neuroinflammation through the cGAS-IFN signaling pathway in the spinal cord of neuropathic pain mice
- Application value of artificial intelligence algorithm-based magnetic resonance multi-sequence imaging in staging diagnosis of cervical cancer
- Embedded monitoring system and teaching of artificial intelligence online drug component recognition
- Investigation into the association of FNDC1 and ADAMTS12 gene expression with plumage coloration in Muscovy ducks
- Yak meat content in feed and its impact on the growth of rats
- A rare case of Richter transformation with breast involvement: A case report and literature review
- First report of Nocardia wallacei infection in an immunocompetent patient in Zhejiang province
- Rhodococcus equi and Brucella pulmonary mass in immunocompetent: A case report and literature review
- Downregulation of RIP3 ameliorates the left ventricular mechanics and function after myocardial infarction via modulating NF-κB/NLRP3 pathway
- Evaluation of the role of some non-enzymatic antioxidants among Iraqi patients with non-alcoholic fatty liver disease
- The role of Phafin proteins in cell signaling pathways and diseases
- Ten-year anemia as initial manifestation of Castleman disease in the abdominal cavity: A case report
- Coexistence of hereditary spherocytosis with SPTB P.Trp1150 gene variant and Gilbert syndrome: A case report and literature review
- Utilization of convolutional neural networks to analyze microscopic images for high-throughput screening of mesenchymal stem cells
- Exploratory evaluation supported by experimental and modeling approaches of Inula viscosa root extract as a potent corrosion inhibitor for mild steel in a 1 M HCl solution
- Imaging manifestations of ductal adenoma of the breast: A case report
- Gut microbiota and sleep: Interaction mechanisms and therapeutic prospects
- Isomangiferin promotes the migration and osteogenic differentiation of rat bone marrow mesenchymal stem cells
- Prognostic value and microenvironmental crosstalk of exosome-related signatures in human epidermal growth factor receptor 2 positive breast cancer
- Circular RNAs as potential biomarkers for male severe sepsis
- Knockdown of Stanniocalcin-1 inhibits growth and glycolysis in oral squamous cell carcinoma cells
- The expression and biological role of complement C1s in esophageal squamous cell carcinoma
- A novel GNAS mutation in pseudohypoparathyroidism type 1a with articular flexion deformity: A case report
- Predictive value of serum magnesium levels for prognosis in patients with non-small cell lung cancer undergoing EGFR-TKI therapy
- HSPB1 alleviates acute-on-chronic liver failure via the P53/Bax pathway
- IgG4-related disease complicated by PLA2R-associated membranous nephropathy: A case report
- Baculovirus-mediated endostatin and angiostatin activation of autophagy through the AMPK/AKT/mTOR pathway inhibits angiogenesis in hepatocellular carcinoma
- Metformin mitigates osteoarthritis progression by modulating the PI3K/AKT/mTOR signaling pathway and enhancing chondrocyte autophagy
- Evaluation of the activity of antimicrobial peptides against bacterial vaginosis
- Atypical presentation of γ/δ mycosis fungoides with an unusual phenotype and SOCS1 mutation
- Analysis of the microecological mechanism of diabetic kidney disease based on the theory of “gut–kidney axis”: A systematic review
- Omega-3 fatty acids prevent gestational diabetes mellitus via modulation of lipid metabolism
- Refractory hypertension complicated with Turner syndrome: A case report
- Interaction of ncRNAs and the PI3K/AKT/mTOR pathway: Implications for osteosarcoma
- Association of low attenuation area scores with pulmonary function and clinical prognosis in patients with chronic obstructive pulmonary disease
- Long non-coding RNAs in bone formation: Key regulators and therapeutic prospects
- The deubiquitinating enzyme USP35 regulates the stability of NRF2 protein
- Neutrophil-to-lymphocyte ratio and platelet-to-lymphocyte ratio as potential diagnostic markers for rebleeding in patients with esophagogastric variceal bleeding
- G protein-coupled receptor 1 participating in the mechanism of mediating gestational diabetes mellitus by phosphorylating the AKT pathway
- LL37-mtDNA regulates viability, apoptosis, inflammation, and autophagy in lipopolysaccharide-treated RLE-6TN cells by targeting Hsp90aa1
- The analgesic effect of paeoniflorin: A focused review
- Chemical composition’s effect on Solanum nigrum Linn.’s antioxidant capacity and erythrocyte protection: Bioactive components and molecular docking analysis
- Knockdown of HCK promotes HREC cell viability and inner blood–retinal barrier integrity by regulating the AMPK signaling pathway
- The role of rapamycin in the PINK1/Parkin signaling pathway in mitophagy in podocytes
- Laryngeal non-Hodgkin lymphoma: Report of four cases and review of the literature
- Clinical value of macrogenome next-generation sequencing on infections
- Overview of dendritic cells and related pathways in autoimmune uveitis
- TAK-242 alleviates diabetic cardiomyopathy via inhibiting pyroptosis and TLR4/CaMKII/NLRP3 pathway
- Hypomethylation in promoters of PGC-1α involved in exercise-driven skeletal muscular alterations in old age
- Profile and antimicrobial susceptibility patterns of bacteria isolated from effluents of Kolladiba and Debark hospitals
- The expression and clinical significance of syncytin-1 in serum exosomes of hepatocellular carcinoma patients
- A histomorphometric study to evaluate the therapeutic effects of biosynthesized silver nanoparticles on the kidneys infected with Plasmodium chabaudi
- PGRMC1 and PAQR4 are promising molecular targets for a rare subtype of ovarian cancer
- Analysis of MDA, SOD, TAOC, MNCV, SNCV, and TSS scores in patients with diabetes peripheral neuropathy
- SLIT3 deficiency promotes non-small cell lung cancer progression by modulating UBE2C/WNT signaling
- The relationship between TMCO1 and CALR in the pathological characteristics of prostate cancer and its effect on the metastasis of prostate cancer cells
- Heterogeneous nuclear ribonucleoprotein K is a potential target for enhancing the chemosensitivity of nasopharyngeal carcinoma
- PHB2 alleviates retinal pigment epithelium cell fibrosis by suppressing the AGE–RAGE pathway
- Anti-γ-aminobutyric acid-B receptor autoimmune encephalitis with syncope as the initial symptom: Case report and literature review
- Comparative analysis of chloroplast genome of Lonicera japonica cv. Damaohua
- Human umbilical cord mesenchymal stem cells regulate glutathione metabolism depending on the ERK–Nrf2–HO-1 signal pathway to repair phosphoramide mustard-induced ovarian cancer cells
- Electroacupuncture on GB acupoints improves osteoporosis via the estradiol–PI3K–Akt signaling pathway
- Renalase protects against podocyte injury by inhibiting oxidative stress and apoptosis in diabetic nephropathy
- Review: Dicranostigma leptopodum: A peculiar plant of Papaveraceae
- Combination effect of flavonoids attenuates lung cancer cell proliferation by inhibiting the STAT3 and FAK signaling pathway
- Renal microangiopathy and immune complex glomerulonephritis induced by anti-tumour agents: A case report
- Correlation analysis of AVPR1a and AVPR2 with abnormal water and sodium and potassium metabolism in rats
- Gastrointestinal health anti-diarrheal mixture relieves spleen deficiency-induced diarrhea through regulating gut microbiota
- Myriad factors and pathways influencing tumor radiotherapy resistance
- Exploring the effects of culture conditions on Yapsin (YPS) gene expression in Nakaseomyces glabratus
- Screening of prognostic core genes based on cell–cell interaction in the peripheral blood of patients with sepsis
- Coagulation factor II thrombin receptor as a promising biomarker in breast cancer management
- Ileocecal mucinous carcinoma misdiagnosed as incarcerated hernia: A case report
- Methyltransferase like 13 promotes malignant behaviors of bladder cancer cells through targeting PI3K/ATK signaling pathway
- The debate between electricity and heat, efficacy and safety of irreversible electroporation and radiofrequency ablation in the treatment of liver cancer: A meta-analysis
- ZAG promotes colorectal cancer cell proliferation and epithelial–mesenchymal transition by promoting lipid synthesis
- Baicalein inhibits NLRP3 inflammasome activation and mitigates placental inflammation and oxidative stress in gestational diabetes mellitus
- Impact of SWCNT-conjugated senna leaf extract on breast cancer cells: A potential apoptotic therapeutic strategy
- MFAP5 inhibits the malignant progression of endometrial cancer cells in vitro
- Major ozonated autohemotherapy promoted functional recovery following spinal cord injury in adult rats via the inhibition of oxidative stress and inflammation
- Axodendritic targeting of TAU and MAP2 and microtubule polarization in iPSC-derived versus SH-SY5Y-derived human neurons
- Differential expression of phosphoinositide 3-kinase/protein kinase B and Toll-like receptor/nuclear factor kappa B signaling pathways in experimental obesity Wistar rat model
- The therapeutic potential of targeting Oncostatin M and the interleukin-6 family in retinal diseases: A comprehensive review
- BA inhibits LPS-stimulated inflammatory response and apoptosis in human middle ear epithelial cells by regulating the Nf-Kb/Iκbα axis
- Role of circRMRP and circRPL27 in chronic obstructive pulmonary disease
- Investigating the role of hyperexpressed HCN1 in inducing myocardial infarction through activation of the NF-κB signaling pathway
- Characterization of phenolic compounds and evaluation of anti-diabetic potential in Cannabis sativa L. seeds: In vivo, in vitro, and in silico studies
- Quantitative immunohistochemistry analysis of breast Ki67 based on artificial intelligence
- Ecology and Environmental Science
- Screening of different growth conditions of Bacillus subtilis isolated from membrane-less microbial fuel cell toward antimicrobial activity profiling
- Degradation of a mixture of 13 polycyclic aromatic hydrocarbons by commercial effective microorganisms
- Evaluation of the impact of two citrus plants on the variation of Panonychus citri (Acari: Tetranychidae) and beneficial phytoseiid mites
- Prediction of present and future distribution areas of Juniperus drupacea Labill and determination of ethnobotany properties in Antalya Province, Türkiye
- Population genetics of Todarodes pacificus (Cephalopoda: Ommastrephidae) in the northwest Pacific Ocean via GBS sequencing
- A comparative analysis of dendrometric, macromorphological, and micromorphological characteristics of Pistacia atlantica subsp. atlantica and Pistacia terebinthus in the middle Atlas region of Morocco
- Macrofungal sporocarp community in the lichen Scots pine forests
- Assessing the proximate compositions of indigenous forage species in Yemen’s pastoral rangelands
- Food Science
- Gut microbiota changes associated with low-carbohydrate diet intervention for obesity
- Reexamination of Aspergillus cristatus phylogeny in dark tea: Characteristics of the mitochondrial genome
- Differences in the flavonoid composition of the leaves, fruits, and branches of mulberry are distinguished based on a plant metabolomics approach
- Investigating the impact of wet rendering (solventless method) on PUFA-rich oil from catfish (Clarias magur) viscera
- Non-linear associations between cardiovascular metabolic indices and metabolic-associated fatty liver disease: A cross-sectional study in the US population (2017–2020)
- Knockdown of USP7 alleviates atherosclerosis in ApoE-deficient mice by regulating EZH2 expression
- Utility of dairy microbiome as a tool for authentication and traceability
- Agriculture
- Enhancing faba bean (Vicia faba L.) productivity through establishing the area-specific fertilizer rate recommendation in southwest Ethiopia
- Impact of novel herbicide based on synthetic auxins and ALS inhibitor on weed control
- Perspectives of pteridophytes microbiome for bioremediation in agricultural applications
- Fertilizer application parameters for drip-irrigated peanut based on the fertilizer effect function established from a “3414” field trial
- Improving the productivity and profitability of maize (Zea mays L.) using optimum blended inorganic fertilization
- Application of leaf multispectral analyzer in comparison to hyperspectral device to assess the diversity of spectral reflectance indices in wheat genotypes
- Animal Sciences
- Knockdown of ANP32E inhibits colorectal cancer cell growth and glycolysis by regulating the AKT/mTOR pathway
- Development of a detection chip for major pathogenic drug-resistant genes and drug targets in bovine respiratory system diseases
- Exploration of the genetic influence of MYOT and MB genes on the plumage coloration of Muscovy ducks
- Transcriptome analysis of adipose tissue in grazing cattle: Identifying key regulators of fat metabolism
- Comparison of nutritional value of the wild and cultivated spiny loaches at three growth stages
- Transcriptomic analysis of liver immune response in Chinese spiny frog (Quasipaa spinosa) infected with Proteus mirabilis
- Disruption of BCAA degradation is a critical characteristic of diabetic cardiomyopathy revealed by integrated transcriptome and metabolome analysis
- Plant Sciences
- Effect of long-term in-row branch covering on soil microorganisms in pear orchards
- Photosynthetic physiological characteristics, growth performance, and element concentrations reveal the calcicole–calcifuge behaviors of three Camellia species
- Transcriptome analysis reveals the mechanism of NaHCO3 promoting tobacco leaf maturation
- Bioinformatics, expression analysis, and functional verification of allene oxide synthase gene HvnAOS1 and HvnAOS2 in qingke
- Water, nitrogen, and phosphorus coupling improves gray jujube fruit quality and yield
- Improving grape fruit quality through soil conditioner: Insights from RNA-seq analysis of Cabernet Sauvignon roots
- Role of Embinin in the reabsorption of nucleus pulposus in lumbar disc herniation: Promotion of nucleus pulposus neovascularization and apoptosis of nucleus pulposus cells
- Revealing the effects of amino acid, organic acid, and phytohormones on the germination of tomato seeds under salinity stress
- Combined effects of nitrogen fertilizer and biochar on the growth, yield, and quality of pepper
- Comprehensive phytochemical and toxicological analysis of Chenopodium ambrosioides (L.) fractions
- Impact of “3414” fertilization on the yield and quality of greenhouse tomatoes
- Exploring the coupling mode of water and fertilizer for improving growth, fruit quality, and yield of the pear in the arid region
- Metagenomic analysis of endophytic bacteria in seed potato (Solanum tuberosum)
- Antibacterial, antifungal, and phytochemical properties of Salsola kali ethanolic extract
- Exploring the hepatoprotective properties of citronellol: In vitro and in silico studies on ethanol-induced damage in HepG2 cells
- Enhanced osmotic dehydration of watermelon rind using honey–sucrose solutions: A study on pre-treatment efficacy and mass transfer kinetics
- Effects of exogenous 2,4-epibrassinolide on photosynthetic traits of 53 cowpea varieties under NaCl stress
- Comparative transcriptome analysis of maize (Zea mays L.) seedlings in response to copper stress
- An optimization method for measuring the stomata in cassava (Manihot esculenta Crantz) under multiple abiotic stresses
- Fosinopril inhibits Ang II-induced VSMC proliferation, phenotype transformation, migration, and oxidative stress through the TGF-β1/Smad signaling pathway
- Antioxidant and antimicrobial activities of Salsola imbricata methanolic extract and its phytochemical characterization
- Bioengineering and Biotechnology
- Absorbable calcium and phosphorus bioactive membranes promote bone marrow mesenchymal stem cells osteogenic differentiation for bone regeneration
- New advances in protein engineering for industrial applications: Key takeaways
- An overview of the production and use of Bacillus thuringiensis toxin
- Research progress of nanoparticles in diagnosis and treatment of hepatocellular carcinoma
- Bioelectrochemical biosensors for water quality assessment and wastewater monitoring
- PEI/MMNs@LNA-542 nanoparticles alleviate ICU-acquired weakness through targeted autophagy inhibition and mitochondrial protection
- Unleashing of cytotoxic effects of thymoquinone-bovine serum albumin nanoparticles on A549 lung cancer cells
- Erratum
- Erratum to “Investigating the association between dietary patterns and glycemic control among children and adolescents with T1DM”
- Erratum to “Activation of hypermethylated P2RY1 mitigates gastric cancer by promoting apoptosis and inhibiting proliferation”
- Retraction
- Retraction to “MiR-223-3p regulates cell viability, migration, invasion, and apoptosis of non-small cell lung cancer cells by targeting RHOB”
- Retraction to “A data mining technique for detecting malignant mesothelioma cancer using multiple regression analysis”
- Special Issue on Advances in Neurodegenerative Disease Research and Treatment
- Transplantation of human neural stem cell prevents symptomatic motor behavior disability in a rat model of Parkinson’s disease
- Special Issue on Multi-omics
- Inflammasome complex genes with clinical relevance suggest potential as therapeutic targets for anti-tumor drugs in clear cell renal cell carcinoma
- Gastroesophageal varices in primary biliary cholangitis with anti-centromere antibody positivity: Early onset?
Articles in the same Issue
- Biomedical Sciences
- Constitutive and evoked release of ATP in adult mouse olfactory epithelium
- LARP1 knockdown inhibits cultured gastric carcinoma cell cycle progression and metastatic behavior
- PEGylated porcine–human recombinant uricase: A novel fusion protein with improved efficacy and safety for the treatment of hyperuricemia and renal complications
- Research progress on ocular complications caused by type 2 diabetes mellitus and the function of tears and blepharons
- The role and mechanism of esketamine in preventing and treating remifentanil-induced hyperalgesia based on the NMDA receptor–CaMKII pathway
- Brucella infection combined with Nocardia infection: A case report and literature review
- Detection of serum interleukin-18 level and neutrophil/lymphocyte ratio in patients with antineutrophil cytoplasmic antibody-associated vasculitis and its clinical significance
- Ang-1, Ang-2, and Tie2 are diagnostic biomarkers for Henoch-Schönlein purpura and pediatric-onset systemic lupus erythematous
- PTTG1 induces pancreatic cancer cell proliferation and promotes aerobic glycolysis by regulating c-myc
- Role of serum B-cell-activating factor and interleukin-17 as biomarkers in the classification of interstitial pneumonia with autoimmune features
- Effectiveness and safety of a mumps containing vaccine in preventing laboratory-confirmed mumps cases from 2002 to 2017: A meta-analysis
- Low levels of sex hormone-binding globulin predict an increased breast cancer risk and its underlying molecular mechanisms
- A case of Trousseau syndrome: Screening, detection and complication
- Application of the integrated airway humidification device enhances the humidification effect of the rabbit tracheotomy model
- Preparation of Cu2+/TA/HAP composite coating with anti-bacterial and osteogenic potential on 3D-printed porous Ti alloy scaffolds for orthopedic applications
- Aquaporin-8 promotes human dermal fibroblasts to counteract hydrogen peroxide-induced oxidative damage: A novel target for management of skin aging
- Current research and evidence gaps on placental development in iron deficiency anemia
- Single-nucleotide polymorphism rs2910829 in PDE4D is related to stroke susceptibility in Chinese populations: The results of a meta-analysis
- Pheochromocytoma-induced myocardial infarction: A case report
- Kaempferol regulates apoptosis and migration of neural stem cells to attenuate cerebral infarction by O‐GlcNAcylation of β-catenin
- Sirtuin 5 regulates acute myeloid leukemia cell viability and apoptosis by succinylation modification of glycine decarboxylase
- Apigenin 7-glucoside impedes hypoxia-induced malignant phenotypes of cervical cancer cells in a p16-dependent manner
- KAT2A changes the function of endometrial stromal cells via regulating the succinylation of ENO1
- Current state of research on copper complexes in the treatment of breast cancer
- Exploring antioxidant strategies in the pathogenesis of ALS
- Helicobacter pylori causes gastric dysbacteriosis in chronic gastritis patients
- IL-33/soluble ST2 axis is associated with radiation-induced cardiac injury
- The predictive value of serum NLR, SII, and OPNI for lymph node metastasis in breast cancer patients with internal mammary lymph nodes after thoracoscopic surgery
- Carrying SNP rs17506395 (T > G) in TP63 gene and CCR5Δ32 mutation associated with the occurrence of breast cancer in Burkina Faso
- P2X7 receptor: A receptor closely linked with sepsis-associated encephalopathy
- Probiotics for inflammatory bowel disease: Is there sufficient evidence?
- Identification of KDM4C as a gene conferring drug resistance in multiple myeloma
- Microbial perspective on the skin–gut axis and atopic dermatitis
- Thymosin α1 combined with XELOX improves immune function and reduces serum tumor markers in colorectal cancer patients after radical surgery
- Highly specific vaginal microbiome signature for gynecological cancers
- Sample size estimation for AQP4-IgG seropositive optic neuritis: Retinal damage detection by optical coherence tomography
- The effects of SDF-1 combined application with VEGF on femoral distraction osteogenesis in rats
- Fabrication and characterization of gold nanoparticles using alginate: In vitro and in vivo assessment of its administration effects with swimming exercise on diabetic rats
- Mitigating digestive disorders: Action mechanisms of Mediterranean herbal active compounds
- Distribution of CYP2D6 and CYP2C19 gene polymorphisms in Han and Uygur populations with breast cancer in Xinjiang, China
- VSP-2 attenuates secretion of inflammatory cytokines induced by LPS in BV2 cells by mediating the PPARγ/NF-κB signaling pathway
- Factors influencing spontaneous hypothermia after emergency trauma and the construction of a predictive model
- Long-term administration of morphine specifically alters the level of protein expression in different brain regions and affects the redox state
- Application of metagenomic next-generation sequencing technology in the etiological diagnosis of peritoneal dialysis-associated peritonitis
- Clinical diagnosis, prevention, and treatment of neurodyspepsia syndrome using intelligent medicine
- Case report: Successful bronchoscopic interventional treatment of endobronchial leiomyomas
- Preliminary investigation into the genetic etiology of short stature in children through whole exon sequencing of the core family
- Cystic adenomyoma of the uterus: Case report and literature review
- Mesoporous silica nanoparticles as a drug delivery mechanism
- Dynamic changes in autophagy activity in different degrees of pulmonary fibrosis in mice
- Vitamin D deficiency and inflammatory markers in type 2 diabetes: Big data insights
- Lactate-induced IGF1R protein lactylation promotes proliferation and metabolic reprogramming of lung cancer cells
- Meta-analysis on the efficacy of allogeneic hematopoietic stem cell transplantation to treat malignant lymphoma
- Mitochondrial DNA drives neuroinflammation through the cGAS-IFN signaling pathway in the spinal cord of neuropathic pain mice
- Application value of artificial intelligence algorithm-based magnetic resonance multi-sequence imaging in staging diagnosis of cervical cancer
- Embedded monitoring system and teaching of artificial intelligence online drug component recognition
- Investigation into the association of FNDC1 and ADAMTS12 gene expression with plumage coloration in Muscovy ducks
- Yak meat content in feed and its impact on the growth of rats
- A rare case of Richter transformation with breast involvement: A case report and literature review
- First report of Nocardia wallacei infection in an immunocompetent patient in Zhejiang province
- Rhodococcus equi and Brucella pulmonary mass in immunocompetent: A case report and literature review
- Downregulation of RIP3 ameliorates the left ventricular mechanics and function after myocardial infarction via modulating NF-κB/NLRP3 pathway
- Evaluation of the role of some non-enzymatic antioxidants among Iraqi patients with non-alcoholic fatty liver disease
- The role of Phafin proteins in cell signaling pathways and diseases
- Ten-year anemia as initial manifestation of Castleman disease in the abdominal cavity: A case report
- Coexistence of hereditary spherocytosis with SPTB P.Trp1150 gene variant and Gilbert syndrome: A case report and literature review
- Utilization of convolutional neural networks to analyze microscopic images for high-throughput screening of mesenchymal stem cells
- Exploratory evaluation supported by experimental and modeling approaches of Inula viscosa root extract as a potent corrosion inhibitor for mild steel in a 1 M HCl solution
- Imaging manifestations of ductal adenoma of the breast: A case report
- Gut microbiota and sleep: Interaction mechanisms and therapeutic prospects
- Isomangiferin promotes the migration and osteogenic differentiation of rat bone marrow mesenchymal stem cells
- Prognostic value and microenvironmental crosstalk of exosome-related signatures in human epidermal growth factor receptor 2 positive breast cancer
- Circular RNAs as potential biomarkers for male severe sepsis
- Knockdown of Stanniocalcin-1 inhibits growth and glycolysis in oral squamous cell carcinoma cells
- The expression and biological role of complement C1s in esophageal squamous cell carcinoma
- A novel GNAS mutation in pseudohypoparathyroidism type 1a with articular flexion deformity: A case report
- Predictive value of serum magnesium levels for prognosis in patients with non-small cell lung cancer undergoing EGFR-TKI therapy
- HSPB1 alleviates acute-on-chronic liver failure via the P53/Bax pathway
- IgG4-related disease complicated by PLA2R-associated membranous nephropathy: A case report
- Baculovirus-mediated endostatin and angiostatin activation of autophagy through the AMPK/AKT/mTOR pathway inhibits angiogenesis in hepatocellular carcinoma
- Metformin mitigates osteoarthritis progression by modulating the PI3K/AKT/mTOR signaling pathway and enhancing chondrocyte autophagy
- Evaluation of the activity of antimicrobial peptides against bacterial vaginosis
- Atypical presentation of γ/δ mycosis fungoides with an unusual phenotype and SOCS1 mutation
- Analysis of the microecological mechanism of diabetic kidney disease based on the theory of “gut–kidney axis”: A systematic review
- Omega-3 fatty acids prevent gestational diabetes mellitus via modulation of lipid metabolism
- Refractory hypertension complicated with Turner syndrome: A case report
- Interaction of ncRNAs and the PI3K/AKT/mTOR pathway: Implications for osteosarcoma
- Association of low attenuation area scores with pulmonary function and clinical prognosis in patients with chronic obstructive pulmonary disease
- Long non-coding RNAs in bone formation: Key regulators and therapeutic prospects
- The deubiquitinating enzyme USP35 regulates the stability of NRF2 protein
- Neutrophil-to-lymphocyte ratio and platelet-to-lymphocyte ratio as potential diagnostic markers for rebleeding in patients with esophagogastric variceal bleeding
- G protein-coupled receptor 1 participating in the mechanism of mediating gestational diabetes mellitus by phosphorylating the AKT pathway
- LL37-mtDNA regulates viability, apoptosis, inflammation, and autophagy in lipopolysaccharide-treated RLE-6TN cells by targeting Hsp90aa1
- The analgesic effect of paeoniflorin: A focused review
- Chemical composition’s effect on Solanum nigrum Linn.’s antioxidant capacity and erythrocyte protection: Bioactive components and molecular docking analysis
- Knockdown of HCK promotes HREC cell viability and inner blood–retinal barrier integrity by regulating the AMPK signaling pathway
- The role of rapamycin in the PINK1/Parkin signaling pathway in mitophagy in podocytes
- Laryngeal non-Hodgkin lymphoma: Report of four cases and review of the literature
- Clinical value of macrogenome next-generation sequencing on infections
- Overview of dendritic cells and related pathways in autoimmune uveitis
- TAK-242 alleviates diabetic cardiomyopathy via inhibiting pyroptosis and TLR4/CaMKII/NLRP3 pathway
- Hypomethylation in promoters of PGC-1α involved in exercise-driven skeletal muscular alterations in old age
- Profile and antimicrobial susceptibility patterns of bacteria isolated from effluents of Kolladiba and Debark hospitals
- The expression and clinical significance of syncytin-1 in serum exosomes of hepatocellular carcinoma patients
- A histomorphometric study to evaluate the therapeutic effects of biosynthesized silver nanoparticles on the kidneys infected with Plasmodium chabaudi
- PGRMC1 and PAQR4 are promising molecular targets for a rare subtype of ovarian cancer
- Analysis of MDA, SOD, TAOC, MNCV, SNCV, and TSS scores in patients with diabetes peripheral neuropathy
- SLIT3 deficiency promotes non-small cell lung cancer progression by modulating UBE2C/WNT signaling
- The relationship between TMCO1 and CALR in the pathological characteristics of prostate cancer and its effect on the metastasis of prostate cancer cells
- Heterogeneous nuclear ribonucleoprotein K is a potential target for enhancing the chemosensitivity of nasopharyngeal carcinoma
- PHB2 alleviates retinal pigment epithelium cell fibrosis by suppressing the AGE–RAGE pathway
- Anti-γ-aminobutyric acid-B receptor autoimmune encephalitis with syncope as the initial symptom: Case report and literature review
- Comparative analysis of chloroplast genome of Lonicera japonica cv. Damaohua
- Human umbilical cord mesenchymal stem cells regulate glutathione metabolism depending on the ERK–Nrf2–HO-1 signal pathway to repair phosphoramide mustard-induced ovarian cancer cells
- Electroacupuncture on GB acupoints improves osteoporosis via the estradiol–PI3K–Akt signaling pathway
- Renalase protects against podocyte injury by inhibiting oxidative stress and apoptosis in diabetic nephropathy
- Review: Dicranostigma leptopodum: A peculiar plant of Papaveraceae
- Combination effect of flavonoids attenuates lung cancer cell proliferation by inhibiting the STAT3 and FAK signaling pathway
- Renal microangiopathy and immune complex glomerulonephritis induced by anti-tumour agents: A case report
- Correlation analysis of AVPR1a and AVPR2 with abnormal water and sodium and potassium metabolism in rats
- Gastrointestinal health anti-diarrheal mixture relieves spleen deficiency-induced diarrhea through regulating gut microbiota
- Myriad factors and pathways influencing tumor radiotherapy resistance
- Exploring the effects of culture conditions on Yapsin (YPS) gene expression in Nakaseomyces glabratus
- Screening of prognostic core genes based on cell–cell interaction in the peripheral blood of patients with sepsis
- Coagulation factor II thrombin receptor as a promising biomarker in breast cancer management
- Ileocecal mucinous carcinoma misdiagnosed as incarcerated hernia: A case report
- Methyltransferase like 13 promotes malignant behaviors of bladder cancer cells through targeting PI3K/ATK signaling pathway
- The debate between electricity and heat, efficacy and safety of irreversible electroporation and radiofrequency ablation in the treatment of liver cancer: A meta-analysis
- ZAG promotes colorectal cancer cell proliferation and epithelial–mesenchymal transition by promoting lipid synthesis
- Baicalein inhibits NLRP3 inflammasome activation and mitigates placental inflammation and oxidative stress in gestational diabetes mellitus
- Impact of SWCNT-conjugated senna leaf extract on breast cancer cells: A potential apoptotic therapeutic strategy
- MFAP5 inhibits the malignant progression of endometrial cancer cells in vitro
- Major ozonated autohemotherapy promoted functional recovery following spinal cord injury in adult rats via the inhibition of oxidative stress and inflammation
- Axodendritic targeting of TAU and MAP2 and microtubule polarization in iPSC-derived versus SH-SY5Y-derived human neurons
- Differential expression of phosphoinositide 3-kinase/protein kinase B and Toll-like receptor/nuclear factor kappa B signaling pathways in experimental obesity Wistar rat model
- The therapeutic potential of targeting Oncostatin M and the interleukin-6 family in retinal diseases: A comprehensive review
- BA inhibits LPS-stimulated inflammatory response and apoptosis in human middle ear epithelial cells by regulating the Nf-Kb/Iκbα axis
- Role of circRMRP and circRPL27 in chronic obstructive pulmonary disease
- Investigating the role of hyperexpressed HCN1 in inducing myocardial infarction through activation of the NF-κB signaling pathway
- Characterization of phenolic compounds and evaluation of anti-diabetic potential in Cannabis sativa L. seeds: In vivo, in vitro, and in silico studies
- Quantitative immunohistochemistry analysis of breast Ki67 based on artificial intelligence
- Ecology and Environmental Science
- Screening of different growth conditions of Bacillus subtilis isolated from membrane-less microbial fuel cell toward antimicrobial activity profiling
- Degradation of a mixture of 13 polycyclic aromatic hydrocarbons by commercial effective microorganisms
- Evaluation of the impact of two citrus plants on the variation of Panonychus citri (Acari: Tetranychidae) and beneficial phytoseiid mites
- Prediction of present and future distribution areas of Juniperus drupacea Labill and determination of ethnobotany properties in Antalya Province, Türkiye
- Population genetics of Todarodes pacificus (Cephalopoda: Ommastrephidae) in the northwest Pacific Ocean via GBS sequencing
- A comparative analysis of dendrometric, macromorphological, and micromorphological characteristics of Pistacia atlantica subsp. atlantica and Pistacia terebinthus in the middle Atlas region of Morocco
- Macrofungal sporocarp community in the lichen Scots pine forests
- Assessing the proximate compositions of indigenous forage species in Yemen’s pastoral rangelands
- Food Science
- Gut microbiota changes associated with low-carbohydrate diet intervention for obesity
- Reexamination of Aspergillus cristatus phylogeny in dark tea: Characteristics of the mitochondrial genome
- Differences in the flavonoid composition of the leaves, fruits, and branches of mulberry are distinguished based on a plant metabolomics approach
- Investigating the impact of wet rendering (solventless method) on PUFA-rich oil from catfish (Clarias magur) viscera
- Non-linear associations between cardiovascular metabolic indices and metabolic-associated fatty liver disease: A cross-sectional study in the US population (2017–2020)
- Knockdown of USP7 alleviates atherosclerosis in ApoE-deficient mice by regulating EZH2 expression
- Utility of dairy microbiome as a tool for authentication and traceability
- Agriculture
- Enhancing faba bean (Vicia faba L.) productivity through establishing the area-specific fertilizer rate recommendation in southwest Ethiopia
- Impact of novel herbicide based on synthetic auxins and ALS inhibitor on weed control
- Perspectives of pteridophytes microbiome for bioremediation in agricultural applications
- Fertilizer application parameters for drip-irrigated peanut based on the fertilizer effect function established from a “3414” field trial
- Improving the productivity and profitability of maize (Zea mays L.) using optimum blended inorganic fertilization
- Application of leaf multispectral analyzer in comparison to hyperspectral device to assess the diversity of spectral reflectance indices in wheat genotypes
- Animal Sciences
- Knockdown of ANP32E inhibits colorectal cancer cell growth and glycolysis by regulating the AKT/mTOR pathway
- Development of a detection chip for major pathogenic drug-resistant genes and drug targets in bovine respiratory system diseases
- Exploration of the genetic influence of MYOT and MB genes on the plumage coloration of Muscovy ducks
- Transcriptome analysis of adipose tissue in grazing cattle: Identifying key regulators of fat metabolism
- Comparison of nutritional value of the wild and cultivated spiny loaches at three growth stages
- Transcriptomic analysis of liver immune response in Chinese spiny frog (Quasipaa spinosa) infected with Proteus mirabilis
- Disruption of BCAA degradation is a critical characteristic of diabetic cardiomyopathy revealed by integrated transcriptome and metabolome analysis
- Plant Sciences
- Effect of long-term in-row branch covering on soil microorganisms in pear orchards
- Photosynthetic physiological characteristics, growth performance, and element concentrations reveal the calcicole–calcifuge behaviors of three Camellia species
- Transcriptome analysis reveals the mechanism of NaHCO3 promoting tobacco leaf maturation
- Bioinformatics, expression analysis, and functional verification of allene oxide synthase gene HvnAOS1 and HvnAOS2 in qingke
- Water, nitrogen, and phosphorus coupling improves gray jujube fruit quality and yield
- Improving grape fruit quality through soil conditioner: Insights from RNA-seq analysis of Cabernet Sauvignon roots
- Role of Embinin in the reabsorption of nucleus pulposus in lumbar disc herniation: Promotion of nucleus pulposus neovascularization and apoptosis of nucleus pulposus cells
- Revealing the effects of amino acid, organic acid, and phytohormones on the germination of tomato seeds under salinity stress
- Combined effects of nitrogen fertilizer and biochar on the growth, yield, and quality of pepper
- Comprehensive phytochemical and toxicological analysis of Chenopodium ambrosioides (L.) fractions
- Impact of “3414” fertilization on the yield and quality of greenhouse tomatoes
- Exploring the coupling mode of water and fertilizer for improving growth, fruit quality, and yield of the pear in the arid region
- Metagenomic analysis of endophytic bacteria in seed potato (Solanum tuberosum)
- Antibacterial, antifungal, and phytochemical properties of Salsola kali ethanolic extract
- Exploring the hepatoprotective properties of citronellol: In vitro and in silico studies on ethanol-induced damage in HepG2 cells
- Enhanced osmotic dehydration of watermelon rind using honey–sucrose solutions: A study on pre-treatment efficacy and mass transfer kinetics
- Effects of exogenous 2,4-epibrassinolide on photosynthetic traits of 53 cowpea varieties under NaCl stress
- Comparative transcriptome analysis of maize (Zea mays L.) seedlings in response to copper stress
- An optimization method for measuring the stomata in cassava (Manihot esculenta Crantz) under multiple abiotic stresses
- Fosinopril inhibits Ang II-induced VSMC proliferation, phenotype transformation, migration, and oxidative stress through the TGF-β1/Smad signaling pathway
- Antioxidant and antimicrobial activities of Salsola imbricata methanolic extract and its phytochemical characterization
- Bioengineering and Biotechnology
- Absorbable calcium and phosphorus bioactive membranes promote bone marrow mesenchymal stem cells osteogenic differentiation for bone regeneration
- New advances in protein engineering for industrial applications: Key takeaways
- An overview of the production and use of Bacillus thuringiensis toxin
- Research progress of nanoparticles in diagnosis and treatment of hepatocellular carcinoma
- Bioelectrochemical biosensors for water quality assessment and wastewater monitoring
- PEI/MMNs@LNA-542 nanoparticles alleviate ICU-acquired weakness through targeted autophagy inhibition and mitochondrial protection
- Unleashing of cytotoxic effects of thymoquinone-bovine serum albumin nanoparticles on A549 lung cancer cells
- Erratum
- Erratum to “Investigating the association between dietary patterns and glycemic control among children and adolescents with T1DM”
- Erratum to “Activation of hypermethylated P2RY1 mitigates gastric cancer by promoting apoptosis and inhibiting proliferation”
- Retraction
- Retraction to “MiR-223-3p regulates cell viability, migration, invasion, and apoptosis of non-small cell lung cancer cells by targeting RHOB”
- Retraction to “A data mining technique for detecting malignant mesothelioma cancer using multiple regression analysis”
- Special Issue on Advances in Neurodegenerative Disease Research and Treatment
- Transplantation of human neural stem cell prevents symptomatic motor behavior disability in a rat model of Parkinson’s disease
- Special Issue on Multi-omics
- Inflammasome complex genes with clinical relevance suggest potential as therapeutic targets for anti-tumor drugs in clear cell renal cell carcinoma
- Gastroesophageal varices in primary biliary cholangitis with anti-centromere antibody positivity: Early onset?