
Single-cell RNA sequencing data analysis of the inner ear in gentamicin-treated mice via intraperitoneal injection
-
Xiaolin Bao


Abstract
Objective
This study aims to delineate the mechanisms through which intraperitoneal injection of gentamicin (GEN) influences the inner ear cells of mice by employing single-cell RNA sequencing (scRNA-seq) technology.
Methods
Eight-week-old Kunming mice were randomly assigned to three groups: a normal control group, a GEN group, and a GEN + dexamethasone (DEX) group. The mice received continuous intraperitoneal injections of the corresponding drugs for 10 days. Auditory brainstem response (ABR) was assessed to evaluate hearing threshold shifts, and cochlear tissues were harvested for scRNA-seq. The Seurat analysis workflow was employed for data quality control, dimensionality reduction clustering, and differential gene expression analysis.
Results
ABR results demonstrated a significant elevation in hearing thresholds in the GEN group, whereas thresholds in the DEX group showed improvement but remained elevated compared to the NOR group (P < 0.05). Single-cell sequencing revealed notable alterations in the populations of outer hair cells, supporting cells, and immune cells in the GEN group. Analysis of differentially expressed genes identified significant downregulation of cell-specific genes Gbp6, Ppfia4 in hair cells of the GEN group, alongside upregulation of inflammation-related genes Nnat, Gh, indicating that hair cell damage and enhanced immune responses may be pivotal mechanisms underlying GEN-induced ototoxicity.
Conclusion
Utilizing scRNA-seq technology, this study uncovers substantial transcriptional changes induced by GEN in cochlear hair cells, supporting cells, and immune cells in mice, highlighting the role of inflammation and oxidative stress, TNF signaling pathways in its ototoxicity. DEX partially ameliorates hair cell damage.
1 Introduction
The clinical application of ototoxic drugs may result in irreversible damage to the inner ear’s structure and function, manifesting as auditory and vestibular symptoms including hearing loss, tinnitus, and vertigo, which significantly impair patients’ quality of life [1]. Presently, the diagnosis of drug-induced hearing loss primarily depends on medical history, clinical presentations, and auditory function assessments; however, the absence of specific biomarkers poses substantial limitations on early detection and intervention [2,3].
Aminoglycosides, loop diuretics, antitumor agents (e.g., cisplatin), and salicylates are among the most common ototoxic drugs used clinically. Among them, aminoglycosides such as gentamicin (GEN) can induce cochlear hair cell damage, resulting in permanent hearing loss [4]. The pathogenic mechanisms are multifaceted, involving genetic predisposition, gene mutations, mitochondrial dysfunction, oxidative stress, and inflammatory responses [5–7]. Nonetheless, current research predominantly focuses on the whole tissue level, with limited analysis of the inner ear’s diverse cell types at a refined level. Specifically, the cell-specific transcriptomic changes induced by ototoxic drugs at single-cell resolution remain poorly understood, representing a considerable research gap [8,9].
GEN was chosen for this study due to its extensive clinical use and high ototoxicity risk among aminoglycosides. Widely prescribed worldwide for its broad-spectrum efficacy, stability, and low cost, GEN is especially prevalent in developing countries [10]. However, its clinical utility is hindered by a 10–25% incidence of irreversible hearing loss, even at therapeutic doses. GEN accumulates in cochlear tissues, particularly the stria vascularis (SV) and hair cells, where it disrupts mitochondrial function and induces oxidative stress. Its ototoxic effects are further amplified in pediatric patients, individuals with mitochondrial mutations, and those receiving concurrent ototoxic agents like loop diuretics [10–12]. Given its high-risk profile, GEN serves as an ideal model for studying aminoglycoside-induced hearing loss.
The advent of single-cell RNA sequencing (scRNA-seq) technology has provided a novel approach to unravel the cytological and molecular mechanisms underlying drug-induced hearing loss [13,14]. By extracting mouse inner ear tissue for scRNA-seq, coupled with cell clustering analysis, cell type identification, differential gene expression (DEG) analysis, and functional enrichment analysis, the susceptible cell types affected by GEN can be precisely identified, and their gene expression change profiles, along with key damage pathways, can be uncovered [15,16]. And it allows for the identification of rare or previously uncharacterized cell types, such as specific subpopulations of hair cells, supporting cells, or immune cells, which are often masked in bulk RNA-seq [17]. However, scRNA-seq is technically challenging when applied to small, calcified tissues like the cochlea due to difficulties in achieving high-quality single-cell suspensions. Transcript dropout, low capture efficiency, and batch effects can also affect interpretation. Despite these challenges, scRNA-seq remains a powerful tool for elucidating the complex cellular landscape of the inner ear, and is especially relevant in the context of ototoxicity and regeneration research, where cellular heterogeneity plays a key role [18]. This paves the way for screening potential biomarkers, offering a scientific foundation for the early diagnosis and monitoring of drug-induced hearing loss [19,20].
This study addresses three key questions using scRNA-seq: (1) How does GEN alter gene expression in specific cochlear cell types (e.g., outer hair cells [OHC], supporting cells, immune cells)? (2) Which molecular pathways (e.g., inflammation, oxidative stress, apoptosis) are primarily involved? (3) Can dexamethasone (DEX) mitigate these effects by modulating gene expression or inflammatory responses? These findings aim to clarify GEN’s ototoxic mechanisms and guide strategies for clinical risk reduction.
2 Materials and methods
2.1 Experimental animals and grouping
The study was approved by the Ethics Committee of Tianjin TEDA Hospital and conducted in strict accordance with ethical guidelines for animal experiments. Forty-five 8-week-old Kunming mice (KM, Mus musculus), regardless of sex, were housed in a specific-pathogen-free barrier conditions with controlled temperature (22 ± 2°C) and humidity (50 ± 5%), under a 12:12 h light cycle (light/dark), and provided with free access to food and water.
GEN is associated with high rates of ototoxicity. To counter the irreversible hearing loss, DEX was included in this study due to its well-documented protective mechanisms. DEX exerts anti-inflammatory effects by suppressing pro-inflammatory cytokines (e.g., tumor necrosis factor-alpha [TNF-α], IL-1β) and modulating nuclear factor-kappa B signaling, a key pathway involved in GEN-induced inflammation. In GEN-exposed cochlear explants, DEX has been shown to reduce TNF-α-mediated apoptosis by downregulating caspase-3 activation and preserving OHC viability [21,22]. Additionally, DEX exhibits anti-apoptotic properties by inhibiting the mitochondrial apoptotic pathway, specifically suppressing Bax overexpression, preventing cytochrome c release, and reducing caspase-3 activation. These effects collectively prevent GEN-triggered hair cell death, supporting DEX as a potential otoprotective agent [22]. Given these dual protective mechanisms, DEX serves as an ideal intervention for assessing its ability to mitigate GEN-induced ototoxicity.
The mice were randomly divided into three groups (n = 15):
GEN + DEX group: Intraperitoneal injection of GEN at 200 mg/kg plus DEX sodium phosphate at 5 mg/kg daily for 10 consecutive days.
Normal control group (NOR group): Intraperitoneal injection of 0.2 mL sterilized water daily for 10 consecutive days.
During the experiment, the general behavior of the mice (e.g., activity level, gait, and posture) was monitored daily, and body weight changes were recorded. Neurological reflexes were evaluated using the air righting reflex and tail suspension test, while auditory brainstem response (ABR) thresholds and rotarod performance were assessed before and after the experiment to evaluate vestibular function.
At the conclusion of the experiment, ten mice per group (totaling 30 mice, 30 cochleae in total) were used for scRNA-seq. Each sequencing sample was obtained from an individual mouse cochlea to ensure independent biological replicates.
The cochleae of the remaining five mice per group (totaling 15 mice) were used for histological and imaging analyses, including pathological sectioning, scanning electron microscopy (SEM), and histopathological examination of the cochlea (HE staining). Each experiment was performed on separate biological samples to ensure independent observations.
2.2 ABR testing in mice
ABR measurements were performed using the Tucker-Davis Technologies (TDT) auditory system (TDT, Alachua, FL, USA) and Biosig software. All experiments were carried out in a sound-attenuating chamber, and mice were anesthetized with 1% pentobarbital sodium (0.04 mL/g, intraperitoneal injection) prior to testing.
ABR signals were recorded using needle electrodes:
Recording electrode: placed subcutaneously at the midpoint of the line connecting the anterior edges of the two ears.
Reference electrode: placed subcutaneously behind the test ear.
Ground electrode: placed subcutaneously behind the contralateral ear.
The loudspeaker was placed approximately 1 cm from the entrance of the external auditory canal. The stimulus was a click sound (primary frequency: 12 kHz) with a band-pass filter ranging from 30 to 3,000 Hz, and signals were averaged over 512 repetitions with a scan time of 10 ms.
The steps of ABR testing are as follows:
The initial sound intensity was set at 90 dB sound pressure level (SPL), decreasing in 10 dB increments until no repeatable ABR waveform was detected.
The intensity was then increased in 5 dB increments until a repeatable ABR waveform reappeared.
The lowest intensity at which wave II could be identified as the auditory threshold.
The latencies of waves I and III, as well as the inter-peak intervals, were measured at 90 dB SPL.
2.3 Extraction of mouse cochlea
At the end of the experiments, mice were euthanized by carbon dioxide inhalation. Both ears were fixed, and the skull was excised using surgical scissors, following these steps to extract the cochlea:
The skin was cut along the midline sagittal suture to expose the skull.
The surrounding both ear canals were cut, and the occipital foramen was dissected open to separate the skull into left and right halves.
Brain tissue on one side was removed, and the temporal bone was rapidly identified via the superior semicircular canal or uvula cerebelli, and then dissected out with micro-tweezers.
Protocol for cell isolation, the cochlear tissues were transferred into a digestion enzyme solution containing 0.1–0.3% collagenase IV (w/v) and 0.05% trypsin (w/v) dissolved in Hanks’ Balanced Salt Solution, followed by incubation at 37°C for 15–30 min. Gentle pipetting was performed every 5 min to facilitate tissue dissociation. To terminate digestion, the reaction was quenched by adding culture medium supplemented with 10% fetal bovine serum. The cell suspension was filtered through a 40 μm cell strainer to remove undigested tissue debris. The filtrate was centrifuged at 300 × g for 5 min at 4°C, and the supernatant was discarded. The cell pellet was washed twice with phosphate-buffered saline. These details ensure reproducibility and clarify our workflow.
For single-cell sequencing samples, the temporal bone was maintained on ice throughout the procedure. Fascia, nerves, and muscle tissues were removed, and the bone was immediately transferred to a 4°C storage solution for prompt scRNA-seq.
For pathological examination, the cochlea was fixed in 4% paraformaldehyde according to the following steps:
The cochlea was exposed under a dissecting microscope, and surrounding tissue was cleared.
The round window and oval window were opened to avoid damage to the basilar membrane.
4% paraformaldehyde was slowly injected through an apical opening until it exited from the round window.
The cochlea was fixed in the refrigerator at 4°C for subsequent SEM analysis.
2.4 scRNA-seq
scRNA-seq in this study was performed with the assistance of Shanghai GeneChem Co., Ltd, utilizing the BD Rhapsody microplate sequencing technology. The experimental procedures included the following steps:
Library preparation was carried out using the BD Rhapsody single-cell whole transcriptome amplification technology.
cDNA sequencing was performed using Illumina 150 bp paired-end (PE150) sequencing.
Raw data were processed using the BD Rhapsody Analysis Pipeline to obtain single-cell transcriptomic sequencing data.
scRNA-seq was conducted using cochlear tissues from ten mice per group, ensuring independent biological replicates. Each scRNA-seq experiment was performed on a separate cochlea rather than pooled samples to maintain independent observations.
Cell-type-specific marker genes were identified using the Seurat package by applying the FindAllMarkers function. This function utilizes the Wilcoxon rank-sum test to detect DEGs between each cell cluster and all other clusters, selecting genes with a minimum log-fold change threshold and statistical significance (adjusted P-value <0.05). Only genes exhibiting a cluster-specific expression pattern and meeting the predefined selection criteria were retained as marker genes.
2.5 Data statistics and analysis
Data analysis was performed using R software (R 4.2.0) with the Seurat package for single-cell clustering analysis, cell type identification, and DEGs analysis.
The statistical analysis procedures included the following steps:
Cell clustering analysis involved t-distributed stochastic neighbour embedding (t-SNE) or uniform manifold approximation and projection (UMAP) dimensionality reduction methods for visualization.
DEGs analysis was conducted using the Wilcoxon rank-sum test to identify DEGs (P < 0.05).
Gene ontology (GO)/kyoto encyclopedia of genes and genomes (KEGG) enrichment analysis was performed using the ClusterProfiler package to analyze significantly enriched biological pathways.
ABR data analysis was conducted with SPSS 26.0 software. Data are expressed as mean ± standard deviation (
-
Ethical approval: The study was approved by the Ethics Committee of Tianjin TEDA Hospital and conducted in strict accordance with ethical guidelines for animal experiments.
3 Results
3.1 ABR testing results in mice
ABR testing was employed to evaluate the auditory threshold changes in mice. A minimal Wave II threshold of ≤25 dB SPL was defined as the criterion for normal hearing, with thresholds >25 dB SPL considered indicative of hearing impairment.
NOR group: All auditory thresholds were below 25 dB SPL, with a mean threshold of 22 dB SPL, indicating normal hearing.
GEN + DEX group: After 10 days, five mice (33.3%) retained normal hearing with an average auditory threshold of 30.5 dB SPL, which was significantly elevated compared to the NOR group (P < 0.05).
GEN group: Only two mice (13.3%) retained normal hearing, with an average auditory threshold of 38.5 dB SPL, demonstrating significant hearing loss (P < 0.01, compared to the NOR group). (Supplementary ABR-figure).
Statistical analysis revealed that the auditory thresholds in the GEN group were significantly higher than those in the NOR and DEX groups (P < 0.01), suggesting marked ototoxic damage induced by GEN, while DEX could partially mitigate hearing loss (P < 0.05).
The results of body weight changes across experimental groups and neurological reflex test results (air righting reflex and tail suspension test) are shown in Supplementary 1. The representative images and analysis of histopathological examination of the cochlea (HE staining), and representative images and analysis of SEM findings are shown in Supplementary HE stain-figure and SEM-figure.
3.2 scRNA-seq results
In the present study, scRNA-seq was performed on inner ear tissues from the three groups of mice using the BD Rhapsody platform. A total of 7,206 cells from the NOR group, 6,201 cells from the GEN group, and 6,990 cells from the DEX group were detected. Following data quality control (QC) and batch effect removal, the samples were included in the analysis.
3.2.1 Data pre-processing – QC and data normalization
Setting of data QC criteria:
QC was applied to the raw sequencing data to exclude low-quality cells (with insufficient gene counts), potentially damaged cells (with high mitochondrial gene content), and outlier cells (with abnormal gene counts). The filtering criteria were as follows, based on previous studies [6,16,25,26]:
500 < Number of genes (nFeature_RNA) <6,000
Number of transcripts (nCount_RNA) <25,000
Mitochondrial gene percentage (mito%) <20%
Ribosomal gene proportion >1% (indicating cell activity)
Hemoglobin gene proportion <1% (excluding the impact of blood contamination)
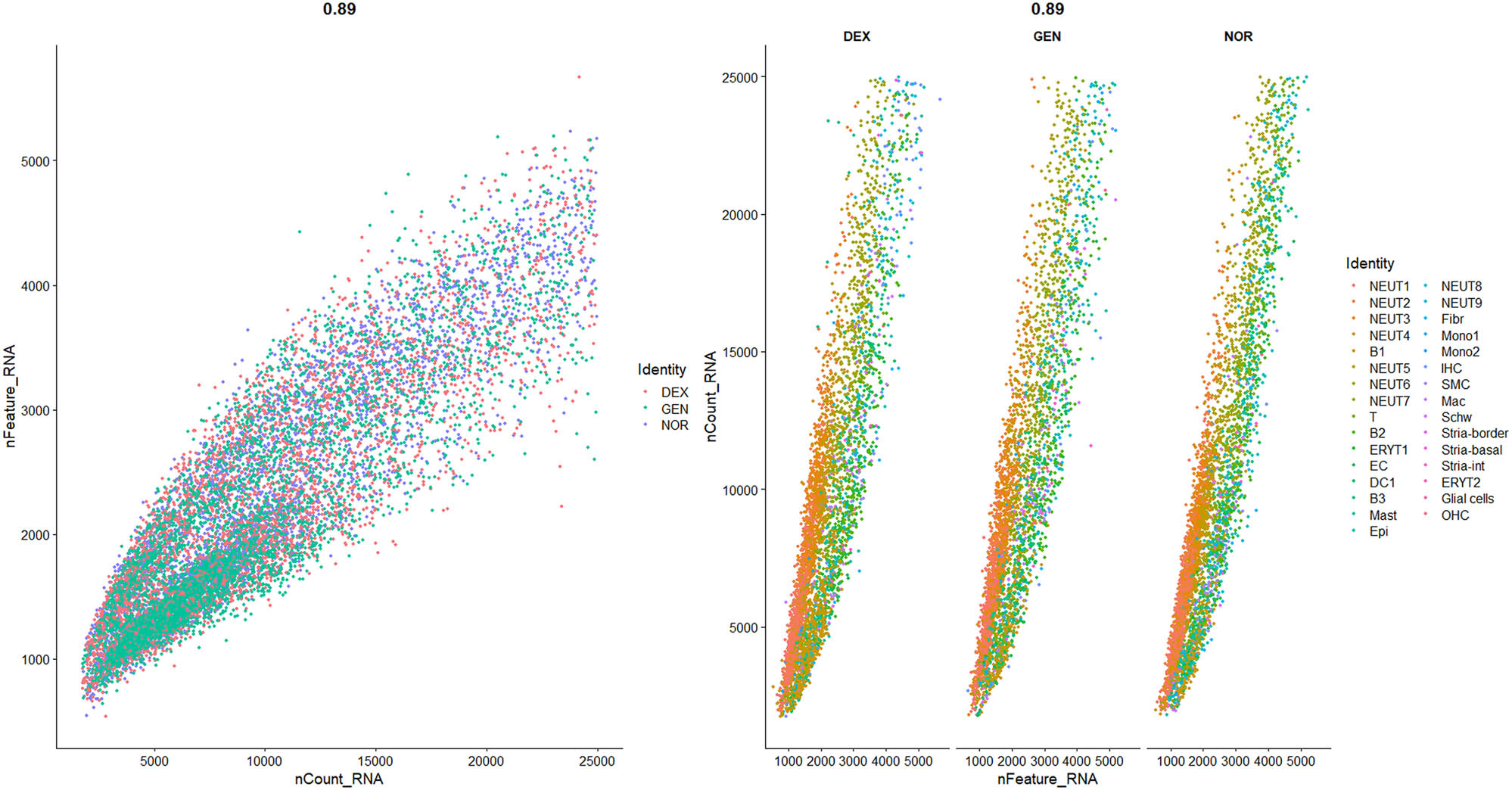
The data distribution before and after filtering is shown in Figures 1–3, highlighting a significant enhancement in data quality post-filtering. PostQC, hemoglobin gene transcripts constituted <0.5% of total reads, confirming minimal blood contamination.
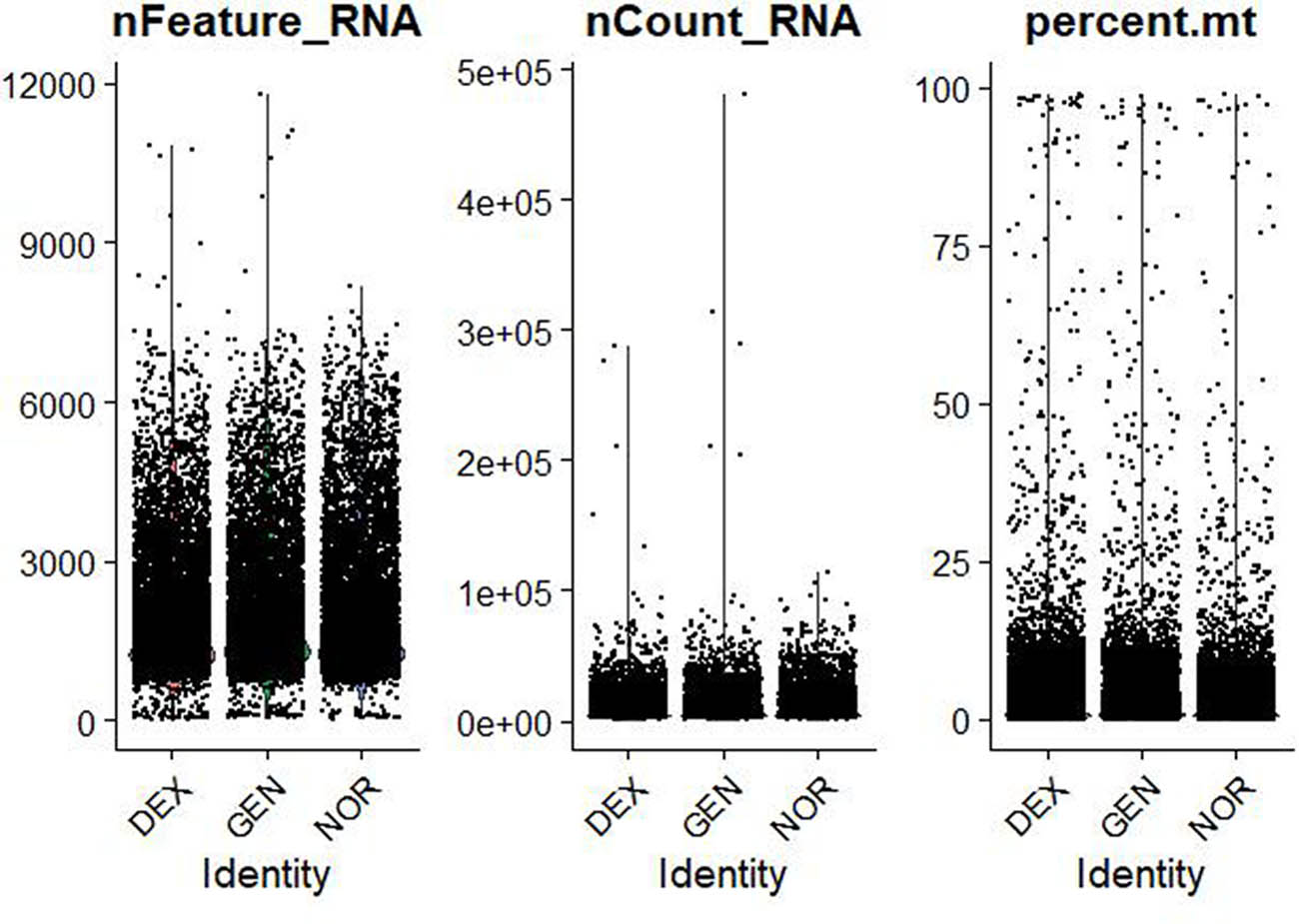
Violin plots of raw single‐cell QC metrics across DEX, GEN, and NOR groups prior to filtering. Violin plots of raw QC metrics (nFeature_RNA, nCount_RNA, percent.mt, percent_ribo, percent_Hb) across DEX, GEN, and NOR groups before filtering. These distributions illustrate the presence of empty droplets (<500 genes, <1,000 UMIs), doublets/high-content cells (>8,000 genes, >50,000 UMIs), high mitochondrial content (up to 60%), and blood contamination (percent_Hb > 1%).
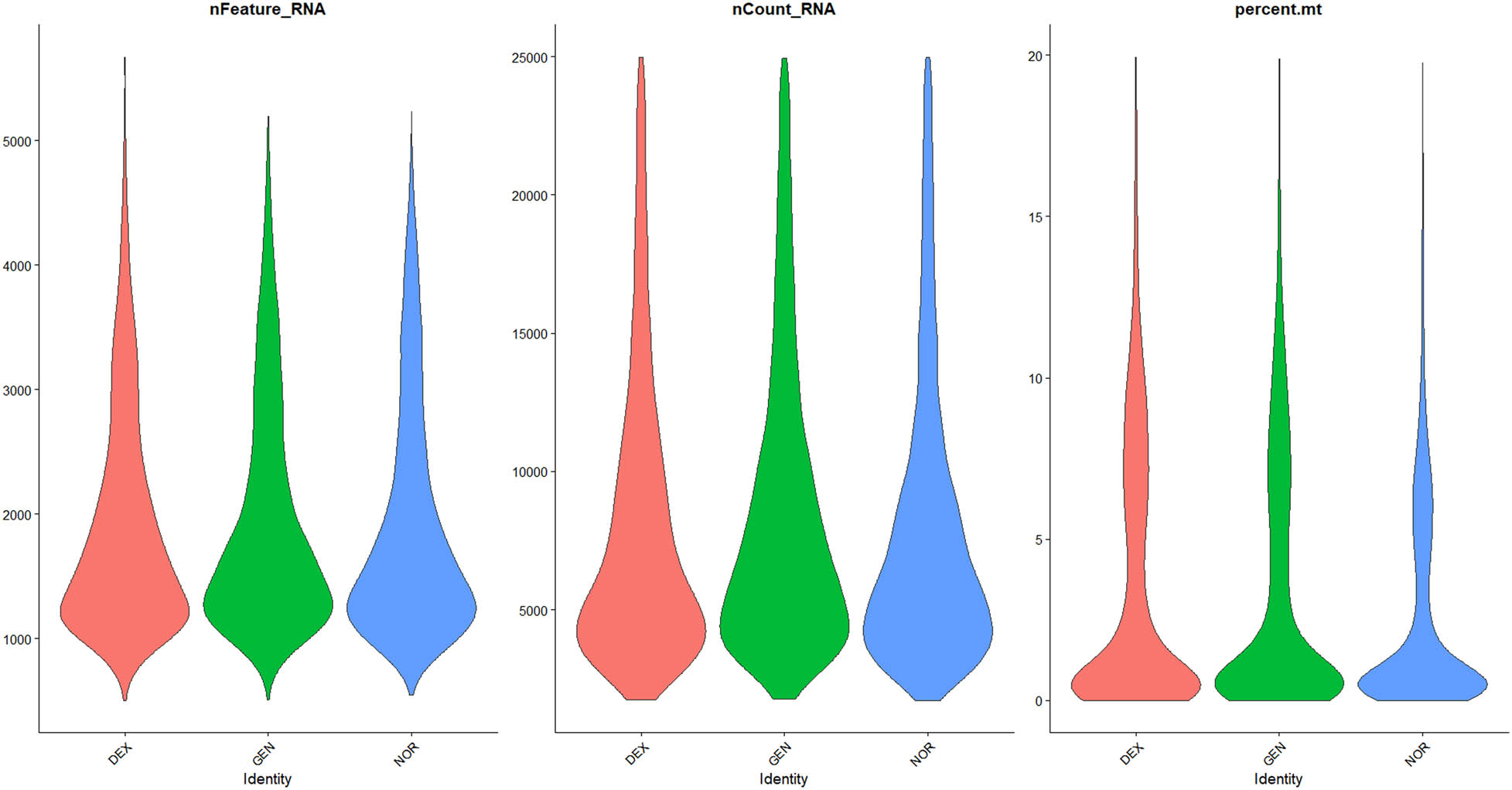
Violin plots of filtered single‐cell QC metrics across DEX, GEN, and NOR groups after applying QC thresholds. Violin plots of QC metrics after applying cutoffs (500 < nFeature_RNA < 6,000; nCount_RNA < 25,000; percent.mt < 20%; percent_ribo > 1%; percent_Hb < 1%). The post-filter distributions collapse tightly within these thresholds, confirming removal of low-quality, apoptotic, and blood-contaminated cells while retaining transcriptionally active single cells for downstream analysis.
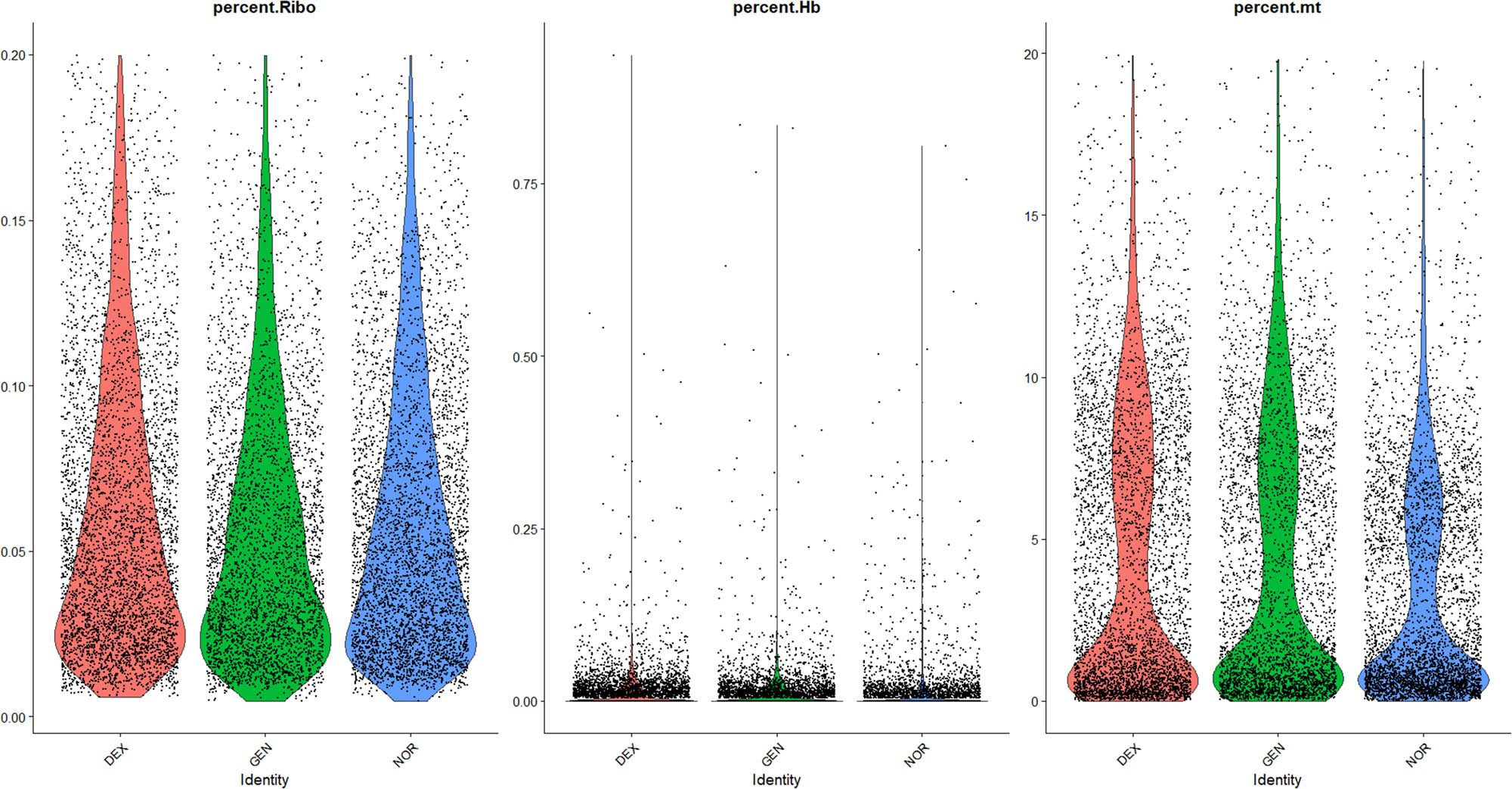
Comparative analysis of ribosomal genes, erythrocyte genes, and mitochondrial genes.
3.2.2 Cell cycle analysis
Assessment of cell cycle states:
Given the potential influence of cell cycle status on gene expression differences, cell cycle analysis was performed on data from all three groups. Results indicated that the majority of inner ear cells in mice resided primarily in the S and G2M phases, with cell cycle scores centralized around zero, suggesting no significant intergroup differences in cell cycle status, thereby not impeding subsequent analyses (Figure 4).
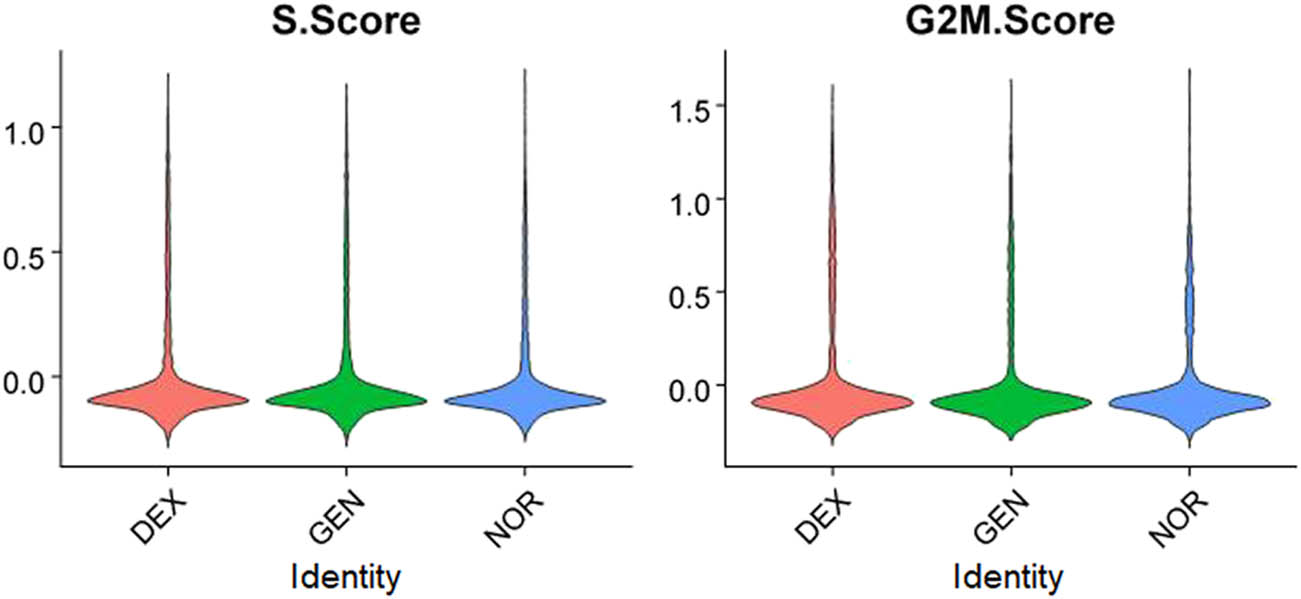
Cell cycle distribution of inner ear cells in different groups.
3.2.3 Cell clustering analysis
3.2.3.1 Feature gene selection and dimensionality reduction clustering
High-variable genes were selected for clustering analysis based on Seurat’s highly variable gene selection. Principal component analysis (PCA) identified the top 20 principal components, followed by t-SNE and UMAP dimensionality reduction for visualization of clustering (Figure 5).
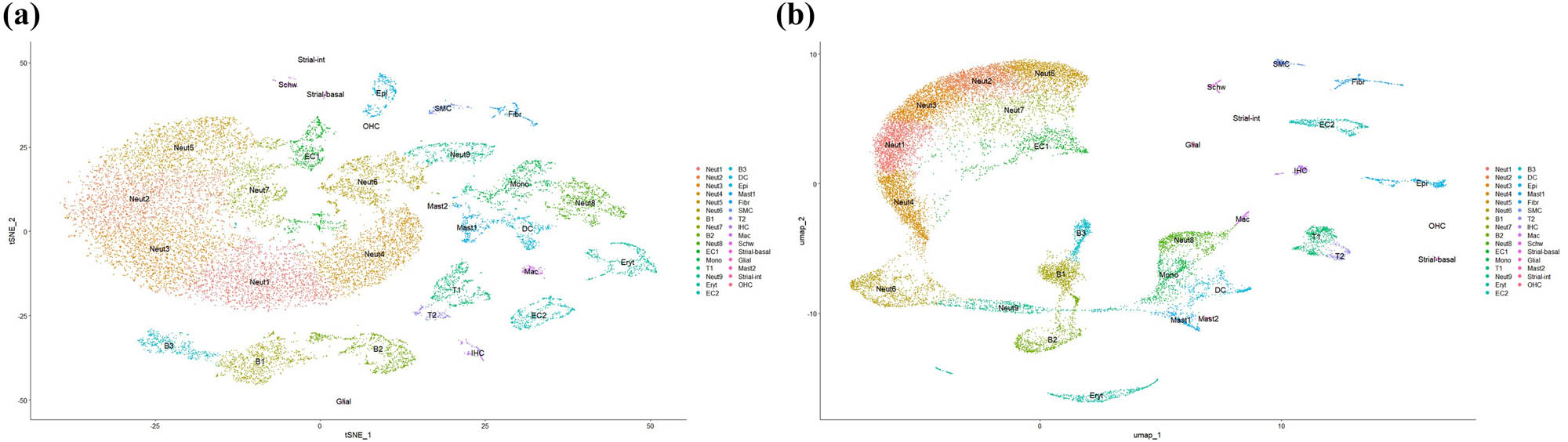
UMAP and t-SNE visualizations of Harmony‐corrected single‐cell data demonstrating batch integration across DEX, GEN, and NOR samples. (a) UMAP visualization of Harmony-corrected single-cell data across DEX, GEN, and NOR samples. (b) t-SNE visualization of the same integrated dataset, highlighting batch alignment.
3.2.3.2 Cell population classification
Clustering analysis identified a total of 31 cell populations, encompassing hair cells (OHC, inner hair cells), supporting cells (pillar cells, Deiters’ cells), immune cells (B cells, T cells, dendritic cells, macrophages), strial vascular cells (basal cells, marginal cells, intermediate cells), glial cells, Schwann cells, epithelial cells, among others.
3.2.4 Cell type annotation – DEG analysis
3.2.4.1 Cell type identification
The characteristic genes for each cell cluster were queried using a single-cell transcriptome database, and combined with known cell-specific marker genes to determine cell types (Table 1).
Cell types and proportions and Chi-square statistic in the three groups
| Cell type | NOR (%) | DEX (%) | GEN (%) | Chi-squared | P_val | P < 0.01 |
|---|---|---|---|---|---|---|
| B cell | 8.00 | 21.84 | 12.34 | 368.19 | 2.2 × 10−16 | Yes |
| Dendritic cell | 2.89 | 2.70 | 2.55 | 4.3898 | 0.1114 | No |
| Endothelial cell | 3.03 | 2.19 | 3.98 | 12.382 | 0.002048 | Yes |
| Epithelial cell | 2.78 | 1.82 | 1.51 | 25.799 | 0.0000025 | Yes |
| Erythrocyte | 0.13 | 0.18 | 0.33 | 2.8889 | 0.2359 | No |
| Fibroblast | 1.94 | 1.19 | 1.66 | 8.7246 | 0.01275 | No |
| Glial cell | 0.18 | 0.24 | 0.56 | 7.9512 | 0.01877 | No |
| Hair cell | 0.15 | 1.40 | 0.77 | 48.851 | 2.466 × 10−11 | Yes |
| Macrophagecyte | 0.75 | 0.94 | 1.02 | 1.0769 | 0.5836 | No |
| Mastocyte | 2.50 | 1.56 | 2.58 | 9.8526 | 0.007253 | Yes |
| Monocyte | 0.86 | 0.55 | 0.97 | 3.5882 | 0.1663 | No |
| Neutrophil | 68.98 | 60.69 | 63.65 | 71.474 | 3.018 × 10−16 | Yes |
| Schwann cell | 0.53 | 0.59 | 0.66 | 0.18182 | 0.9131 | No |
| Smooth muscle cell | 1.46 | 0.61 | 0.97 | 17.636 | 0.000148 | Yes |
| Strial vascular | 0.53 | 0.83 | 0.64 | 4.2069 | 0.122 | No |
| T cell | 5.28 | 2.66 | 5.80 | 43.107 | 4.359 × 10−16 | Yes |
| Total | 100.00 | 100.00 | 100.00 | 60.406 | 7.637 × 10−14 | Yes |
3.2.4.2 Cell type-specific gene expression
A heatmap of cell type-specific genes was generated (Figure 6) to further validate the accuracy of cell annotation. For example:
OHC: highly express “Nnat”, “Dlk1”.
B cells: highly express “Fcrla”, “Vpreb3”.
T cells: highly express “Ccl5”, “Gzma”.
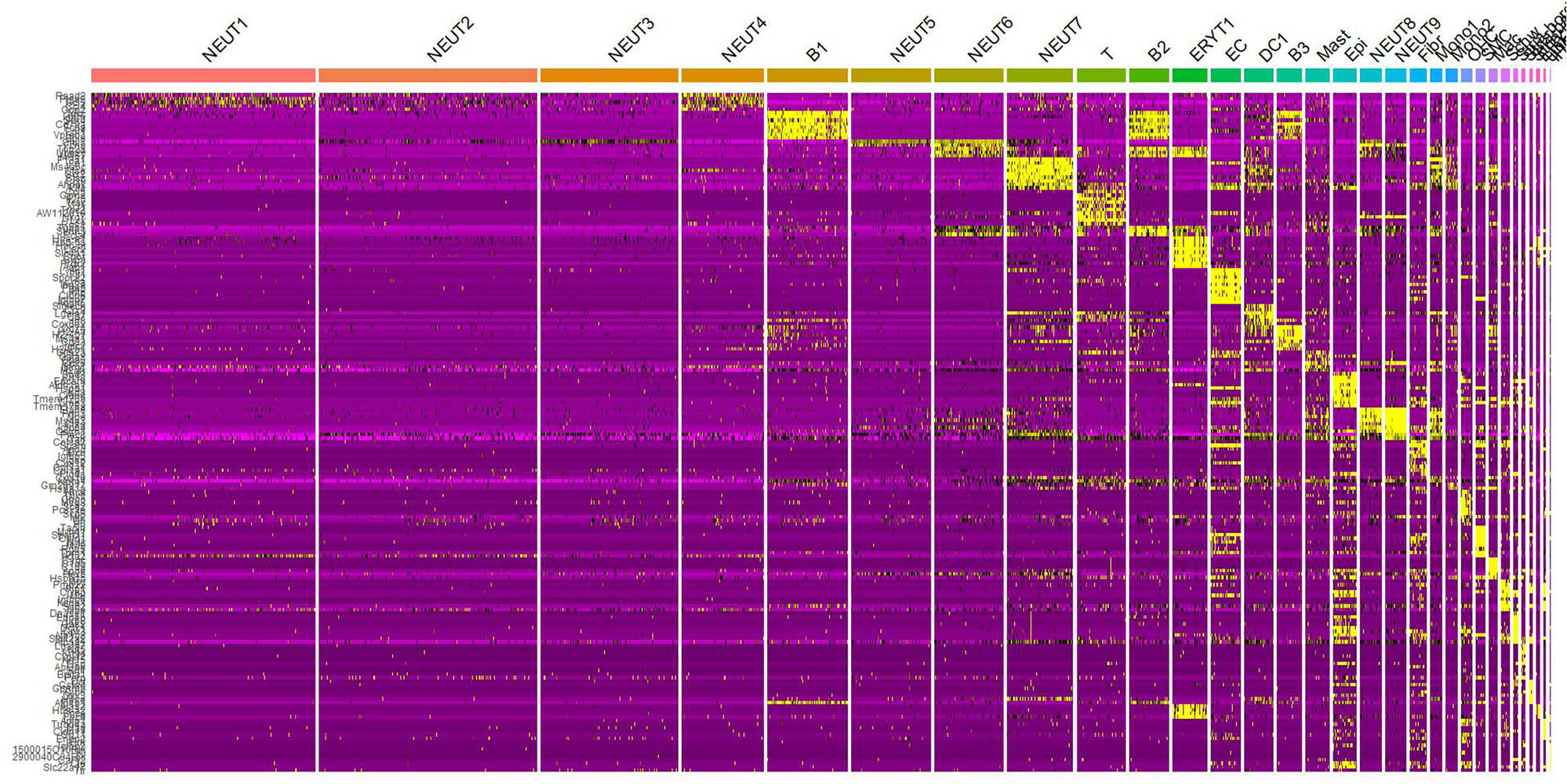
Heatmap of cell type-specific gene expression.
3.2.4.3 DEG analysis
The Wilcoxon rank-sum test was used to identify DEGs (P < 0.05), and a volcano plot depicting the top ten most upregulated genes was generated (Figure 7). Results indicate that, compared to the NOR group, expression of hair cell-specific genes (such as “Gbp6” and “Igfbpl1”) in the GEN group was significantly downregulated, whereas inflammation-related genes (such as “Gh” and “Nnat”) were markedly upregulated. The gene distribution visualizations of “Gbp6”, “Igfbpl1”, “Gh”, and “Nnat” are showed in Supplementary 3.
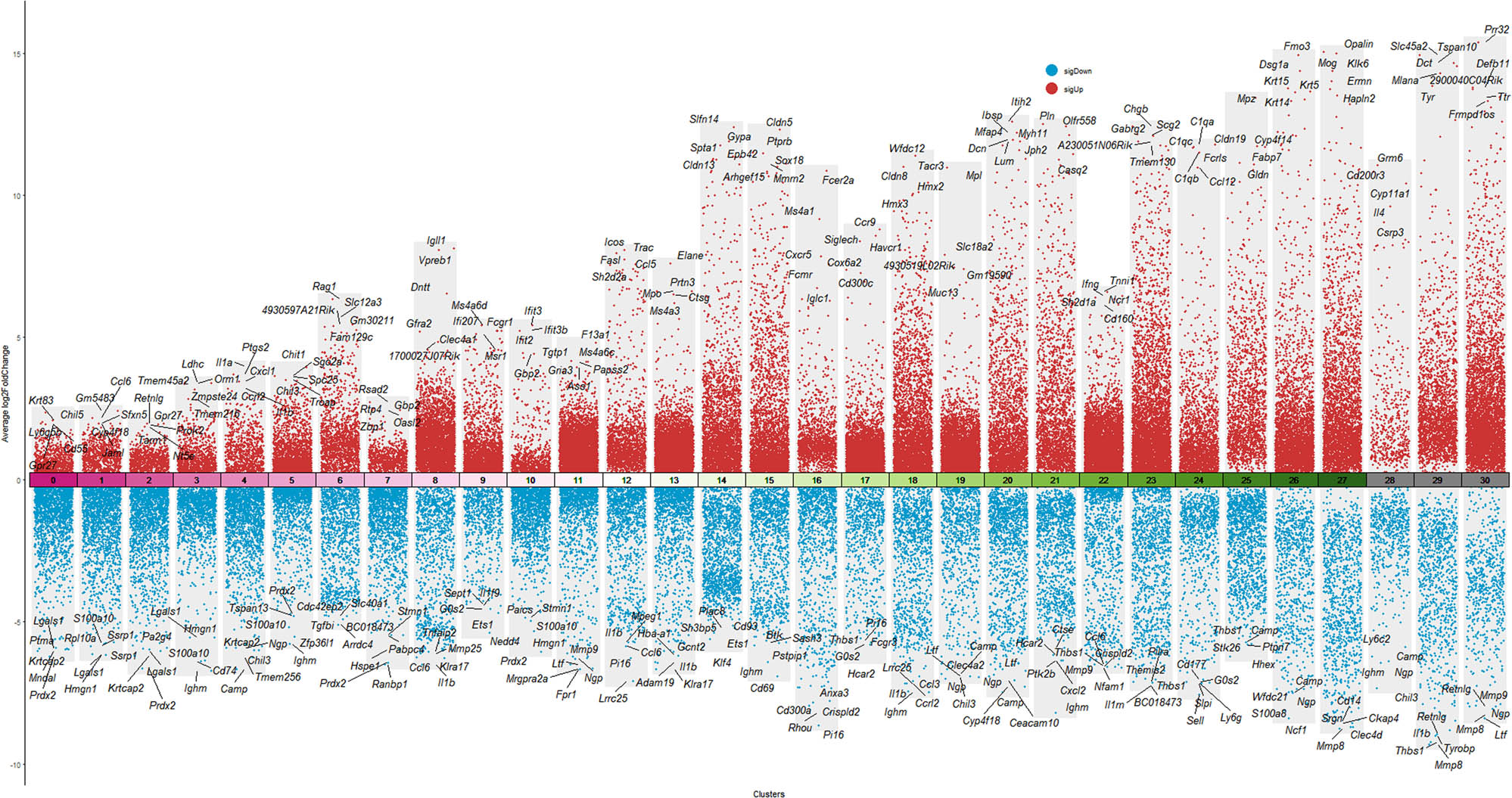
Volcano plot of DEGs between GEN and NOR groups.
3.2.4.4 Batch effect removal and data integration
① Batch effect removal
To mitigate technical variability arising from differences in sampling time, operators, and reagent lots, we applied the Harmony algorithm to our merged single‐cell dataset for batch‐effect correction. Following integration, we re-computed PCA and performed UMAP on the corrected embeddings. As shown in Figure 8, Harmony successfully collapsed the three original acquisition batches (DEX, GEN, NOR) into a unified transcriptional landscape: cells from all batches now co‐localize within shared UMAP clusters, and previously batch‐segregated subpopulations merge seamlessly while preserving well‐defined hair cell, supporting cell, and immune cell clusters. This result confirms that biological variation, rather than technical artifacts, drives the observed cellular heterogeneity and validates the use of the integrated dataset for downstream differential expression and trajectory analyses.
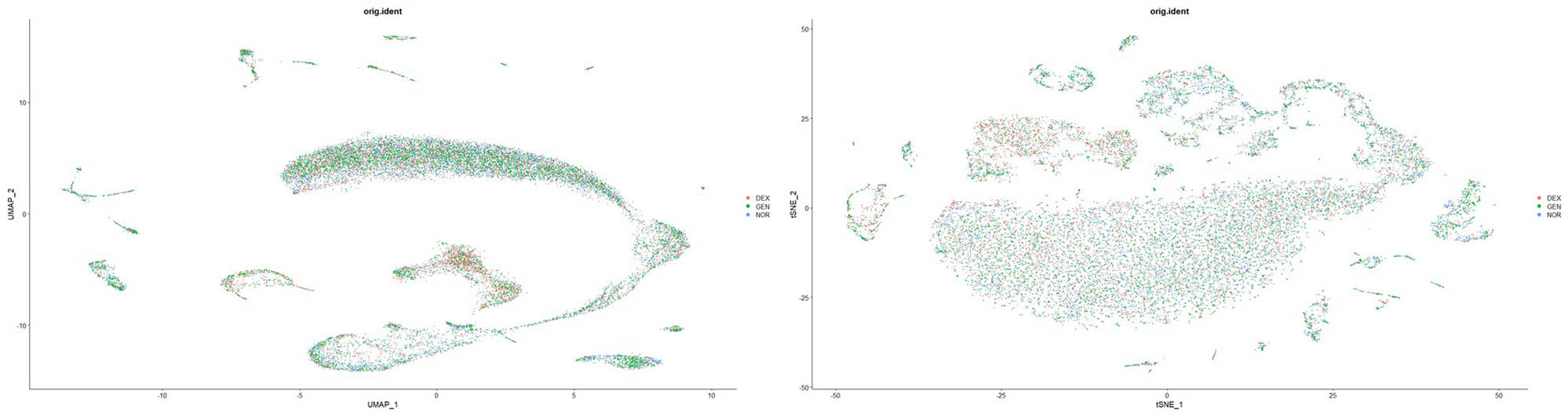
Batch effect removal and data integration results.
② Cell composition analysis
The proportion of each cell type is detailed in Table 1. Comparisons among the three groups revealed:
A significant decrease in the proportion of hair cells in the GEN group (P < 0.01), while increases were observed in B cells and Schwann cells (P < 0.05), suggesting an enhanced inflammatory response.
In the DEX group, the proportion of hair cells increased relative to the GEN group (P < 0.05), indicating a protective effect of DEX on hair cells.
3.2.5 Distribution characteristics of mitochondrial-related genes
3.2.5.1 Mitochondrial gene analysis
Based on data from the mouse mitochondrial gene database, a total of 1,140 mitochondrial genes were identified. Genes related to the oxidative phosphorylation (OXPHOS) pathway (ND1, ND2, CYTB) were significantly downregulated in the GEN group (P < 0.01), suggesting mitochondrial dysfunction as a critical mechanism of GEN-induced ototoxicity (Figure 9).
Expression of mitochondrial-related genes in different groups.
3.2.6 Functional enrichment analysis
GO enrichment analysis of the DEGs revealed that genes upregulated in the GEN group were primarily enriched in pathways related to “inflammatory response”, “apoptosis”, and “oxidative stress” (P < 0.01, Figure 9).
KEGG analysis demonstrated that GEN group DEGs were significantly enriched in the p53 signaling pathway, TNF signaling pathway, and MAPK signaling pathway, all closely related to hair cell damage and inflammatory response (P < 0.05, Figure 10, Supplementary 2).
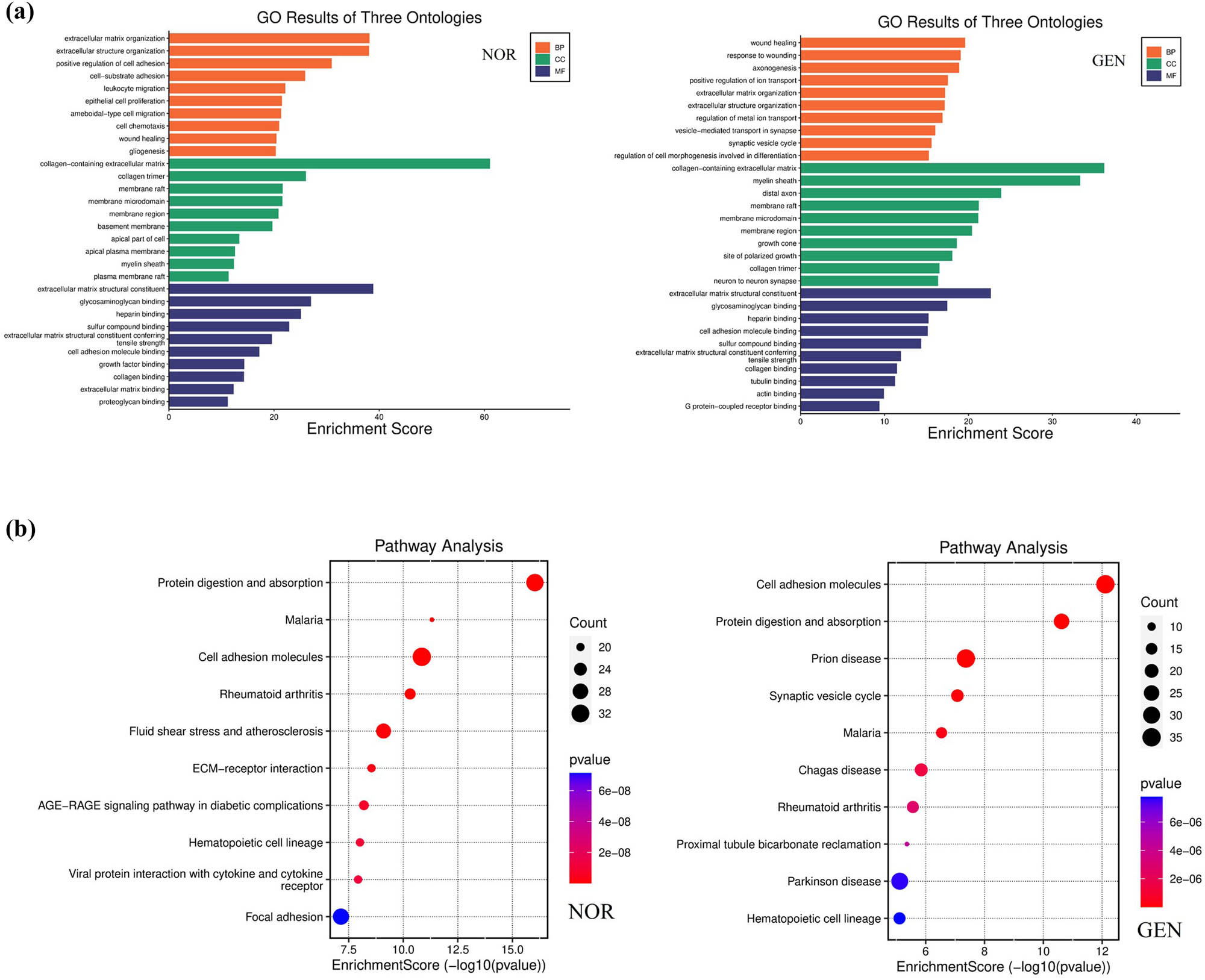
GO and KEGG enrichment analysis of DEGs. (a) GO enrichment analysis of DEGs between GEN and NOR groups. (b) KEGG pathway enrichment analysis of the same DEG set.
3.2.7 Distribution characteristics of nuclear genes
Following batch effect correction using Harmony and cell clustering annotation, 31 cell clusters were identified. DEGs for each cell cluster were selected and visualized with Dotplot to preliminarily identify highly specific marker genes in each cluster and Prestin (Slc26a5) and Myo7a acted as canonical hair cell markers. The results are as follows (Table 2):
Top ten DEGs in each cell cluster following integration of single-cell data from all three groups
| Gene | p_val | avg_log2FC | pct.1 | pct.2 | p_val_adj | Cluster |
|---|---|---|---|---|---|---|
| Il1b | 0 | 2.296307 | 0.874 | 0.306 | 0 | NEUT1 |
| Ccl6 | 0 | 2.230189 | 0.943 | 0.357 | 0 | NEUT1 |
| Cxcl2 | 0 | 2.137592 | 0.976 | 0.551 | 0 | NEUT1 |
| Ccrl2 | 0 | 2.130405 | 0.92 | 0.516 | 0 | NEUT1 |
| Clec4d | 0 | 2.074705 | 0.994 | 0.505 | 0 | NEUT1 |
| Hcar2 | 0 | 2.073142 | 0.937 | 0.465 | 0 | NEUT1 |
| Nlrp3 | 0 | 1.974674 | 0.946 | 0.465 | 0 | NEUT1 |
| Acod1 | 0 | 1.941379 | 0.945 | 0.532 | 0 | NEUT1 |
| Il1r2 | 0 | 1.901822 | 0.957 | 0.515 | 0 | NEUT1 |
| Csf3r | 0 | 1.899401 | 0.977 | 0.583 | 0 | NEUT1 |
| Retnlg | 0 | 2.079243 | 0.999 | 0.711 | 0 | NEUT2 |
| Mmp8 | 0 | 1.922687 | 0.995 | 0.514 | 0 | NEUT2 |
| Fpr1 | 0 | 1.508132 | 0.907 | 0.39 | 0 | NEUT2 |
| S100a6 | 0 | 1.455033 | 0.979 | 0.676 | 0 | NEUT2 |
| Il1f9 | 0 | 1.407325 | 0.929 | 0.438 | 0 | NEUT2 |
| Mmp9 | 0 | 1.353328 | 0.996 | 0.519 | 0 | NEUT2 |
| Prr13 | 0 | 1.297979 | 0.989 | 0.764 | 0 | NEUT2 |
| Adam8 | 0 | 1.297021 | 0.99 | 0.578 | 0 | NEUT2 |
| Thbs1 | 0 | 1.231989 | 0.738 | 0.42 | 0 | NEUT2 |
| R3hdm4 | 0 | 1.21724 | 0.982 | 0.774 | 0 | NEUT2 |
| Ltf | 0 | 2.369141 | 0.986 | 0.443 | 0 | NEUT3 |
| Ngp | 0 | 2.02572 | 0.999 | 0.725 | 0 | NEUT3 |
| Ifitm6 | 0 | 1.926356 | 0.995 | 0.516 | 0 | NEUT3 |
| Lcn2 | 0 | 1.822955 | 1 | 0.695 | 0 | NEUT3 |
| Anxa1 | 0 | 1.805717 | 1 | 0.706 | 0 | NEUT3 |
| Camp | 0 | 1.759623 | 0.986 | 0.395 | 0 | NEUT3 |
| Wfdc21 | 0 | 1.666808 | 0.999 | 0.559 | 0 | NEUT3 |
| Plbd1 | 0 | 1.590771 | 0.972 | 0.421 | 0 | NEUT3 |
| Cybb | 0 | 1.582672 | 0.992 | 0.617 | 0 | NEUT3 |
| Ly6g | 0 | 1.580378 | 0.977 | 0.425 | 0 | NEUT3 |
| Rsad2 | 0 | 2.767022 | 0.575 | 0.172 | 0 | NEUT4 |
| Il1b | 0 | 2.566372 | 0.938 | 0.365 | 0 | NEUT4 |
| Ccrl2 | 0 | 2.562171 | 0.961 | 0.558 | 0 | NEUT4 |
| Ptgs2 | 0 | 2.456352 | 0.639 | 0.182 | 0 | NEUT4 |
| Ccl3 | 0 | 2.412587 | 0.831 | 0.434 | 0 | NEUT4 |
| Cxcl2 | 0 | 2.222273 | 0.972 | 0.598 | 0 | NEUT4 |
| Cd274 | 0 | 2.147376 | 0.616 | 0.185 | 0 | NEUT4 |
| Csf1 | 1.74 × 10−249 | 2.254707 | 0.577 | 0.22 | 4.33 × 10−245 | NEUT4 |
| Ccl4 | 4.85 × 10−214 | 2.276061 | 0.511 | 0.185 | 1.21 × 10−209 | NEUT4 |
| Gbp2 | 1.53 × 10−100 | 2.634624 | 0.419 | 0.218 | 3.81 × 10−96 | NEUT4 |
| Igkc | 0 | 4.413135 | 0.889 | 0.174 | 0 | B1 |
| Ighm | 0 | 4.053927 | 0.982 | 0.214 | 0 | B1 |
| Sox4 | 0 | 3.720767 | 0.842 | 0.127 | 0 | B1 |
| Cd79a | 0 | 3.564249 | 0.981 | 0.068 | 0 | B1 |
| Fcrla | 0 | 3.533266 | 0.814 | 0.047 | 0 | B1 |
| Ebf1 | 0 | 3.497047 | 0.935 | 0.093 | 0 | B1 |
| Vpreb3 | 0 | 3.475994 | 0.932 | 0.05 | 0 | B1 |
| Spib | 0 | 3.323718 | 0.854 | 0.064 | 0 | B1 |
| Cecr2 | 0 | 3.288302 | 0.894 | 0.063 | 0 | B1 |
| Fam129c | 0 | 3.243563 | 0.843 | 0.057 | 0 | B1 |
| Chil3 | 0 | 2.477715 | 0.989 | 0.446 | 0 | NEUT5 |
| Camp | 0 | 2.461097 | 0.988 | 0.422 | 0 | NEUT5 |
| Ltf | 0 | 2.376249 | 0.985 | 0.467 | 0 | NEUT5 |
| Ngp | 0 | 2.301347 | 0.994 | 0.737 | 0 | NEUT5 |
| Zmpste24 | 0 | 1.966458 | 0.813 | 0.282 | 0 | NEUT5 |
| Lcn2 | 0 | 1.958909 | 0.997 | 0.709 | 0 | NEUT5 |
| Ifitm6 | 0 | 1.90725 | 0.983 | 0.538 | 0 | NEUT5 |
| Cybb | 0 | 1.869416 | 0.991 | 0.634 | 0 | NEUT5 |
| Anxa1 | 0 | 1.690295 | 0.997 | 0.719 | 0 | NEUT5 |
| Orm1 | 0 | 1.688015 | 0.749 | 0.099 | 0 | NEUT5 |
| Chil3 | 0 | 3.356882 | 0.998 | 0.45 | 0 | NEUT6 |
| Camp | 0 | 2.526071 | 0.997 | 0.426 | 0 | NEUT6 |
| Fcnb | 0 | 2.270154 | 0.832 | 0.113 | 0 | NEUT6 |
| Hmgn2 | 0 | 2.210258 | 0.993 | 0.645 | 0 | NEUT6 |
| Ngp | 0 | 2.075144 | 0.998 | 0.739 | 0 | NEUT6 |
| Top2a | 0 | 2.022558 | 0.939 | 0.187 | 0 | NEUT6 |
| Ube2c | 0 | 2.012327 | 0.76 | 0.136 | 0 | NEUT6 |
| Smc4 | 0 | 1.900779 | 0.991 | 0.41 | 0 | NEUT6 |
| Mki67 | 0 | 1.859998 | 0.975 | 0.2 | 0 | NEUT6 |
| Hmgb2 | 0 | 1.831134 | 1 | 0.906 | 0 | NEUT6 |
| F13a1 | 0 | 3.848398 | 0.965 | 0.052 | 0 | NEUT7 |
| Fn1 | 0 | 3.575091 | 0.932 | 0.06 | 0 | NEUT7 |
| Ms4a6c | 0 | 3.2431 | 0.953 | 0.052 | 0 | NEUT7 |
| Ctss | 0 | 3.069778 | 0.998 | 0.181 | 0 | NEUT7 |
| Ccr2 | 0 | 3.014537 | 0.975 | 0.053 | 0 | NEUT7 |
| Ctsc | 0 | 2.95775 | 0.991 | 0.237 | 0 | NEUT7 |
| Ccl9 | 0 | 2.94072 | 0.955 | 0.066 | 0 | NEUT7 |
| Ahnak | 0 | 2.632812 | 0.985 | 0.22 | 0 | NEUT7 |
| Psap | 0 | 2.597219 | 1 | 0.924 | 0 | NEUT7 |
| Klf4 | 0 | 2.522328 | 0.929 | 0.238 | 0 | NEUT7 |
| Ccl5 | 0 | 5.335997 | 0.645 | 0.022 | 0 | T |
| Gzma | 0 | 3.902751 | 0.448 | 0.004 | 0 | T |
| Il2rb | 0 | 3.702139 | 0.833 | 0.004 | 0 | T |
| Trac | 0 | 3.575134 | 0.606 | 0.005 | 0 | T |
| Xcl1 | 0 | 3.492293 | 0.516 | 0.003 | 0 | T |
| Trbc2 | 0 | 3.233065 | 0.703 | 0.006 | 0 | T |
| AW112010 | 0 | 3.049239 | 0.743 | 0.123 | 0 | T |
| Nkg7 | 0 | 2.791016 | 0.691 | 0.079 | 0 | T |
| Lck | 0 | 2.761573 | 0.89 | 0.028 | 0 | T |
| Rgs1 | 0 | 2.74656 | 0.708 | 0.061 | 0 | T |
| Top2a | 0 | 3.014256 | 0.939 | 0.204 | 0 | B2 |
| Ezh2 | 0 | 2.872195 | 0.995 | 0.3 | 0 | B2 |
| Vpreb3 | 0 | 2.830962 | 0.992 | 0.076 | 0 | B2 |
| Ptma | 0 | 2.761883 | 1 | 0.585 | 0 | B2 |
| Hmgb1 | 0 | 2.664208 | 1 | 0.6 | 0 | B2 |
| Tubb5 | 0 | 2.630771 | 0.992 | 0.358 | 0 | B2 |
| Stmn1 | 0 | 2.600264 | 0.983 | 0.234 | 0 | B2 |
| Pclaf | 0 | 2.593159 | 0.92 | 0.163 | 0 | B2 |
| Ighm | 0 | 2.564379 | 0.993 | 0.237 | 0 | B2 |
| H2afv | 0 | 2.434125 | 0.983 | 0.325 | 0 | B2 |
| Hba-a1 | 0 | 6.761932 | 0.96 | 0.166 | 0 | ERYT1 |
| Hbb-bs | 0 | 6.581102 | 0.947 | 0.084 | 0 | ERYT1 |
| Hbb-bt | 0 | 6.126738 | 0.918 | 0.037 | 0 | ERYT1 |
| Car2 | 0 | 5.755228 | 0.991 | 0.1 | 0 | ERYT1 |
| Slc4a1 | 0 | 5.243651 | 0.879 | 0.013 | 0 | ERYT1 |
| Cpox | 0 | 4.634532 | 0.935 | 0.147 | 0 | ERYT1 |
| Gypa | 0 | 4.32368 | 0.901 | 0.005 | 0 | ERYT1 |
| Prdx2 | 0 | 4.211731 | 0.972 | 0.232 | 0 | ERYT1 |
| Aqp1 | 0 | 3.916893 | 0.911 | 0.021 | 0 | ERYT1 |
| Mki67 | 0 | 3.868076 | 0.975 | 0.22 | 0 | ERYT1 |
| Ly6c1 | 0 | 5.497781 | 0.967 | 0.067 | 0 | EC |
| Flt1 | 0 | 5.244363 | 0.98 | 0.016 | 0 | EC |
| Spock2 | 0 | 5.181551 | 0.792 | 0.031 | 0 | EC |
| Ly6a | 0 | 4.560383 | 0.967 | 0.068 | 0 | EC |
| Itm2a | 0 | 4.519889 | 0.767 | 0.052 | 0 | EC |
| Ptprb | 0 | 4.503494 | 0.971 | 0.004 | 0 | EC |
| Pltp | 0 | 4.399358 | 0.9 | 0.042 | 0 | EC |
| Cldn5 | 0 | 4.363167 | 0.865 | 0.002 | 0 | EC |
| Igfbp7 | 0 | 4.323402 | 0.958 | 0.031 | 0 | EC |
| Adgrf5 | 0 | 4.276998 | 0.984 | 0.005 | 0 | EC |
| Siglech | 0 | 3.664939 | 0.609 | 0.006 | 0 | DC1 |
| Ccr9 | 0 | 3.04166 | 0.405 | 0.007 | 0 | DC1 |
| Lgals1 | 0 | 2.909442 | 0.982 | 0.218 | 0 | DC1 |
| Cd7 | 0 | 2.5661 | 0.657 | 0.022 | 0 | DC1 |
| Irf8 | 0 | 2.498057 | 0.863 | 0.153 | 0 | DC1 |
| Cox6a2 | 0 | 2.293865 | 0.469 | 0.009 | 0 | DC1 |
| Cd74 | 6.72 × 10−265 | 2.685859 | 0.879 | 0.219 | 1.67 × 10−260 | DC1 |
| Tcf4 | 3.61 × 10−256 | 2.57813 | 0.86 | 0.228 | 9.00 × 10−252 | DC1 |
| H2-Aa | 4.45 × 10−184 | 2.793063 | 0.515 | 0.097 | 1.11 × 10−179 | DC1 |
| H2-Ab1 | 1.37 × 10−164 | 2. 561331 | 0.481 | 0.093 | 3.41 × 10−160 | DC1 |
| Cd74 | 0 | 4.631161 | 0.974 | 0.219 | 0 | B3 |
| H2-Aa | 0 | 4.368422 | 0.774 | 0.093 | 0 | B3 |
| Igkc | 0 | 4.182451 | 0.955 | 0.202 | 0 | B3 |
| H2-Ab1 | 0 | 4.098241 | 0.826 | 0.087 | 0 | B3 |
| Ms4a1 | 0 | 3.857937 | 0.866 | 0.009 | 0 | B3 |
| Cd79a | 0 | 3.618734 | 0.992 | 0.105 | 0 | B3 |
| Cd83 | 0 | 3.183551 | 0.795 | 0.104 | 0 | B3 |
| Iglc2 | 0 | 3.138845 | 0.713 | 0.013 | 0 | B3 |
| Ighm | 1.28 × 10−297 | 3.286376 | 0.966 | 0.246 | 3.19 × 10−293 | B3 |
| H2-Eb1 | 1.83 × 10−297 | 3.199098 | 0.574 | 0.072 | 4.56 × 10−293 | B3 |
| Ctla2a | 0 | 3.431689 | 0.732 | 0.043 | 0 | Mast |
| Gata2 | 0 | 2.792429 | 0.668 | 0.037 | 0 | Mast |
| Cpa3 | 0 | 2.784117 | 0.343 | 0.002 | 0 | Mast |
| Rgs1 | 0 | 2.456825 | 0.67 | 0.074 | 0 | Mast |
| Cdk6 | 6.59 × 10−303 | 2.111122 | 0.824 | 0.158 | 1.64 × 10−298 | Mast |
| Ifitm1 | 2.69 × 10−200 | 2.154224 | 0.803 | 0.195 | 6.70 × 10−196 | Mast |
| Rpl32 | 8.27 × 10−199 | 2.063971 | 0.989 | 0.545 | 2.06 × 10−194 | Mast |
| Rps4x | 1.23 × 10−183 | 2.053586 | 0.995 | 0.65 | 3.06 × 10−179 | Mast |
| H2afy | 1.44 × 10−113 | 2.090347 | 0.943 | 0.615 | 3.60 × 10−109 | Mast |
| Ccl4 | 1.55 × 10−47 | 2.254853 | 0.511 | 0.199 | 3.85 × 10−43 | Mast |
| Sod3 | 0 | 5.568798 | 0.77 | 0.025 | 0 | Epi |
| Krt18 | 0 | 4.645166 | 0.943 | 0.025 | 0 | Epi |
| Epcam | 0 | 4.588636 | 0.986 | 0.038 | 0 | Epi |
| Aldh1a1 | 0 | 4.416025 | 0.842 | 0.032 | 0 | Epi |
| Hspb1 | 0 | 4.318796 | 0.92 | 0.074 | 0 | Epi |
| Krt8 | 0 | 4.196297 | 0.876 | 0.019 | 0 | Epi |
| Cldn4 | 0 | 4.112089 | 0.819 | 0.009 | 0 | Epi |
| Tmem176b | 0 | 4.064797 | 0.94 | 0.119 | 0 | Epi |
| Timp3 | 0 | 3.824602 | 0.871 | 0.069 | 0 | Epi |
| Tmem176a | 0 | 3.822391 | 0.902 | 0.095 | 0 | Epi |
| Fcnb | 0 | 4.308985 | 0.979 | 0.136 | 0 | NEUT8 |
| Elane | 0 | 4.034507 | 0.991 | 0.128 | 0 | NEUT8 |
| Prtn3 | 0 | 4.015441 | 0.997 | 0.131 | 0 | NEUT8 |
| Mpo | 0 | 3.745653 | 0.967 | 0.103 | 0 | NEUT8 |
| Ms4a3 | 0 | 2.589234 | 0.985 | 0.075 | 0 | NEUT8 |
| Ctsg | 0 | 2.297662 | 0.86 | 0.068 | 0 | NEUT8 |
| Igfbp4 | 0 | 2.199301 | 0.985 | 0.185 | 0 | NEUT8 |
| Gstm1 | 1.31 × 10−272 | 2.26708 | 0.976 | 0.233 | 3.26 × 10−268 | NEUT8 |
| Lta4h | 2.20 × 10−219 | 2.179699 | 0.985 | 0.397 | 5.47 × 10−215 | NEUT8 |
| Serpinb1a | 1.29 × 10−204 | 2.343175 | 0.967 | 0.384 | 3.20 × 10−200 | NEUT8 |
| Mpo | 0 | 5.599578 | 1 | 0.103 | 0 | NEUT9 |
| Elane | 0 | 5.508864 | 0.991 | 0.128 | 0 | NEUT9 |
| Prtn3 | 0 | 5.064998 | 0.997 | 0.131 | 0 | NEUT9 |
| Ctsg | 0 | 4.421696 | 0.997 | 0.066 | 0 | NEUT9 |
| Gstm1 | 0 | 3.344881 | 0.997 | 0.233 | 0 | NEUT9 |
| Ms4a3 | 0 | 3.313892 | 0.997 | 0.075 | 0 | NEUT9 |
| Nkg7 | 0 | 2.83007 | 0.994 | 0.087 | 0 | NEUT9 |
| Fcnb | 6.02 × 10−254 | 2.89561 | 0.784 | 0.139 | 1.50 × 10−249 | NEUT9 |
| Plac8 | 6.13 × 10−209 | 2.799115 | 1 | 0.487 | 1.53 × 10−204 | NEUT9 |
| Calr | 3.32 × 10−202 | 2.740057 | 1 | 0.607 | 8.27 × 10−198 | NEUT9 |
| Col1a2 | 0 | 7.019246 | 0.874 | 0.05 | 0 | Fibr |
| Sparc | 0 | 5.429979 | 0.976 | 0.086 | 0 | Fibr |
| Apod | 0 | 5.414049 | 0.417 | 0.017 | 0 | Fibr |
| Dcn | 0 | 5.353722 | 0.76 | 0.005 | 0 | Fibr |
| Igfbp5 | 0 | 5.224145 | 0.728 | 0.023 | 0 | Fibr |
| Ibsp | 0 | 4.879163 | 0.331 | 0.003 | 0 | Fibr |
| Cxcl12 | 0 | 4.216762 | 0.492 | 0.024 | 0 | Fibr |
| Col3a1 | 0 | 3.903568 | 0.583 | 0.005 | 0 | Fibr |
| Col1a1 | 2.31 × 10−284 | 7.387203 | 0.689 | 0.084 | 5.76 × 10−280 | Fibr |
| Ptgds | 2.91 × 10−37 | 4.747365 | 0.276 | 0.074 | 7.25 × 10−33 | Fibr |
| Mpo | 0 | 3.564739 | 0.989 | 0.109 | 0 | Mono1 |
| F13a1 | 0 | 2.742456 | 0.995 | 0.089 | 0 | Mono1 |
| Ctsg | 0 | 2.494044 | 1 | 0.072 | 0 | Mono1 |
| Ms4a6c | 0 | 2.284719 | 0.995 | 0.088 | 0 | Mono1 |
| Prtn3 | 8.53 × 10−282 | 3.03184 | 1 | 0.137 | 2.12 × 10−277 | Mono1 |
| Cxcl10 | 5.68 × 10−194 | 2.58553 | 0.934 | 0.16 | 1.41 × 10−189 | Mono1 |
| Lgals1 | 1.57 × 10−188 | 2.709256 | 1 | 0.228 | 3.91 × 10−184 | Mono1 |
| Plac8 | 1.16 × 10−115 | 2.598258 | 1 | 0.49 | 2.88 × 10−111 | Mono1 |
| H2afy | 1.35 × 10−113 | 2.518226 | 1 | 0.617 | 3.37 × 10−109 | Mono1 |
| Npm1 | 4.53 × 10−110 | 2.273215 | 1 | 0.511 | 1.13 × 10−105 | Mono1 |
| Ahnak | 6.02 × 10−77 | 2.320335 | 0.797 | 0.253 | 1.50 × 10−72 | Mono2 |
| Gm26917 | 2.23 × 10−66 | 4.469736 | 0.571 | 0.16 | 5.56 × 10−62 | Mono2 |
| mt-Nd1 | 1.16 × 10−56 | 2.309783 | 0.983 | 0.747 | 2.88 × 10−52 | Mono2 |
| mt-Atp6 | 4.78 × 10−53 | 2.272857 | 1 | 0.907 | 1.19 × 10−48 | Mono2 |
| mt-Co3 | 1.37 × 10−52 | 2.383037 | 0.989 | 0.867 | 3.41 × 10−48 | Mono2 |
| Gm47283 | 3.48 × 10−49 | 2.464701 | 0.78 | 0.377 | 8.66 × 10−45 | Mono2 |
| Hspa1a | 4.86 × 10−42 | 2.564324 | 0.52 | 0.169 | 1.21 × 10−37 | Mono2 |
| Cd74 | 5.99 × 10−37 | 2.841195 | 0.576 | 0.23 | 1.49 × 10−32 | Mono2 |
| H2-Aa | 1.46 × 10−26 | 2.417142 | 0.339 | 0.104 | 3.64 × 10−22 | Mono2 |
| H2-Ab1 | 8.68 × 10−22 | 3.144168 | 0.299 | 0.099 | 2.16 × 10−17 | Mono2 |
| Nnat | 0 | 6.004872 | 0.738 | 0.015 | 0 | OHC |
| Dlk1 | 0 | 5.616719 | 0.64 | 0.019 | 0 | OHC |
| Chgb | 0 | 5.293739 | 0.849 | 0.004 | 0 | OHC |
| Meg3 | 0 | 4.986895 | 0.89 | 0.042 | 0 | OHC |
| Scg2 | 0 | 4.377325 | 0.837 | 0.003 | 0 | OHC |
| Pcsk1n | 0 | 4.219726 | 0.919 | 0.008 | 0 | OHC |
| Scg5 | 0 | 3.727801 | 0.93 | 0.016 | 0 | OHC |
| Mt2 | 1.66 × 10−87 | 4.415268 | 0.814 | 0.272 | 4.14 × 10−83 | OHC |
| Gh | 3.68 × 10−86 | 8.454414 | 0.692 | 0.166 | 9.16 × 10−82 | OHC |
| Prl | 1.16 × 10−85 | 8.370354 | 0.424 | 0.063 | 2.90 × 10−81 | OHC |
| Tagln | 0 | 5.881273 | 0.993 | 0.012 | 0 | SMC |
| Myh11 | 0 | 5.459393 | 1 | 0.003 | 0 | SMC |
| Sparcl1 | 0 | 5.442591 | 1 | 0.048 | 0 | SMC |
| Cald1 | 0 | 5.404352 | 1 | 0.044 | 0 | SMC |
| Mylk | 0 | 5.299315 | 1 | 0.032 | 0 | SMC |
| Myl9 | 0 | 5.18155 | 1 | 0.01 | 0 | SMC |
| Rgs5 | 0 | 5.142747 | 0.646 | 0.008 | 0 | SMC |
| Igfbp7 | 0 | 4.757058 | 1 | 0.045 | 0 | SMC |
| Tpm1 | 3.01 × 10−236 | 5.111376 | 1 | 0.153 | 7.50 × 10−232 | SMC |
| Acta2 | 2.69 × 10−185 | 5.673155 | 0.993 | 0.206 | 6.71 × 10−181 | SMC |
| C1qc | 0 | 5.054586 | 0.97 | 0.003 | 0 | Mac |
| C1qb | 0 | 4.399197 | 0.985 | 0.002 | 0 | Mac |
| C1qa | 0 | 4.208365 | 0.978 | 0.001 | 0 | Mac |
| Apoe | 1.76 × 10−201 | 5.181583 | 0.888 | 0.113 | 4.39 × 10−197 | Mac |
| Ctss | 3.54 × 10−160 | 4.210803 | 1 | 0.216 | 8.82 × 10−156 | Mac |
| Hexb | 9.40 × 10−114 | 4.133809 | 0.94 | 0.275 | 2.34 × 10−109 | Mac |
| Hspa1a | 1.60 × 10−98 | 4.162597 | 0.776 | 0.168 | 3.98 × 10−94 | Mac |
| Cd74 | 3.72 × 10−94 | 4.039485 | 0.873 | 0.229 | 9.26 × 10−90 | Mac |
| Hspa1b | 1.65 × 10−93 | 4.112077 | 0.799 | 0.183 | 4.10 × 10−89 | Mac |
| Ccl4 | 3.32 × 10−89 | 4.251913 | 0.799 | 0.2 | 8.26 × 10−85 | Mac |
| Mpz | 0 | 7.79169 | 0.878 | 0.01 | 0 | Schw |
| Pmp22 | 0 | 5.492076 | 0.962 | 0.039 | 0 | Schw |
| Plp1 | 0 | 4.780818 | 0.992 | 0.022 | 0 | Schw |
| Cryab | 0 | 3.844024 | 0.939 | 0.045 | 0 | Schw |
| Mal | 0 | 3.824259 | 0.931 | 0.039 | 0 | Schw |
| Igfbp6 | 0 | 3.815037 | 0.45 | 0.011 | 0 | Schw |
| Kcna1 | 0 | 3.745437 | 0.847 | 0.004 | 0 | Schw |
| Scd2 | 8.83 × 10−194 | 4.816866 | 1 | 0.168 | 2.20 × 10−189 | Schw |
| Mbp | 1.11 × 10−109 | 4.330123 | 0.954 | 0.304 | 2.77 × 10−105 | Schw |
| Apoe | 2.18 × 10−24 | 4.129665 | 0.389 | 0.117 | 5.42 × 10−20 | Schw |
| Dnase1 | 0 | 5.292244 | 0.357 | 0.003 | 0 | Stria-border |
| Enpep | 0 | 4.539279 | 0.986 | 0.031 | 0 | Stria-border |
| Lrp2 | 0 | 4.342953 | 0.986 | 0.007 | 0 | Stria-border |
| Gas2 | 0 | 4.168965 | 0.957 | 0.024 | 0 | Stria-border |
| Dclk1 | 0 | 4.027127 | 0.971 | 0.029 | 0 | Stria-border |
| Gpx3 | 5.84 × 10−255 | 4.562391 | 0.986 | 0.058 | 1.46 × 10−250 | Stria-border |
| Atp1b2 | 2.67 × 10−229 | 6.197312 | 1 | 0.068 | 6.64 × 10−225 | Stria-border |
| Ptgds | 2.88 × 10−213 | 6.358935 | 1 | 0.073 | 7.16 × 10−209 | Stria-border |
| Slc12a2 | 4.65 × 10−204 | 5.600001 | 1 | 0.077 | 1.16 × 10−199 | Stria-border |
| Atp1a1 | 5.64 × 10−65 | 4.404918 | 1 | 0.34 | 1.41 × 10−60 | Stria-border |
| Lgals7 | 0 | 4.824722 | 0.606 | 0.007 | 0 | Stria-basal |
| Krt5 | 0 | 4.822037 | 0.697 | 0.002 | 0 | Stria-basal |
| Krt14 | 0 | 4.793011 | 0.667 | 0.002 | 0 | Stria-basal |
| Cyp2f2 | 0 | 4.789308 | 0.47 | 0.003 | 0 | Stria-basal |
| Krt15 | 0 | 3.727416 | 0.621 | 0.001 | 0 | Stria-basal |
| Dsp | 0 | 3.701699 | 0.955 | 0.03 | 0 | Stria-basal |
| Abi3bp | 1.97 × 10−292 | 3.756986 | 0.439 | 0.008 | 4.90 × 10−288 | Stria-basal |
| Krt1 | 2.09 × 10−235 | 4.324499 | 0.288 | 0.004 | 5.21 × 10−231 | Stria-basal |
| Bpifa1 | 2.58 × 10−166 | 7.675366 | 0.409 | 0.013 | 6.42 × 10−162 | Stria-basal |
| Perp | 2.48 × 10−143 | 3.96785 | 0.955 | 0.095 | 6.18 × 10−139 | Stria-basal |
| Dct | 0 | 7.754018 | 0.964 | 0.005 | 0 | Stria-int |
| Gsta4 | 0 | 5.431319 | 1 | 0.036 | 0 | Stria-int |
| Gpnmb | 0 | 4.870221 | 0.964 | 0.007 | 0 | Stria-int |
| Gjb6 | 0 | 4.782277 | 0.964 | 0.006 | 0 | Stria-int |
| Dkk3 | 0 | 4.764847 | 0.964 | 0.023 | 0 | Stria-int |
| Ptgds | 5.15 × 10−171 | 5.846893 | 1 | 0.074 | 1.28 × 10−166 | Stria-int |
| Hpse | 4.20 × 10−154 | 4.975566 | 0.964 | 0.077 | 1.05 × 10−149 | Stria-int |
| Slc12a2 | 4.50 × 10−144 | 4.876733 | 0.946 | 0.078 | 1.12 × 10−139 | Stria-int |
| Atp1b1 | 9.40 × 10−77 | 6.169884 | 1 | 0.189 | 2.34 × 10−72 | Stria-int |
| Atp1a1 | 1.30 × 10−51 | 4.6746 | 1 | 0.34 | 3.23 × 10−47 | Stria-int |
| Alas2 | 3.80 × 10−178 | 4.639409 | 0.692 | 0.028 | 9.46 × 10−174 | ERYT2 |
| Hba-a2 | 8.36 × 10−177 | 5.912645 | 0.654 | 0.026 | 2.08 × 10−172 | ERYT2 |
| Hbb-bt | 2.01 × 10−150 | 6.383028 | 0.885 | 0.058 | 5.01 × 10−146 | ERYT2 |
| Snca | 3.82 × 10−130 | 3.519117 | 0.577 | 0.027 | 9.51 × 10−126 | ERYT2 |
| Hbb-bs | 1.14 × 10−96 | 6.471828 | 0.923 | 0.104 | 2.84 × 10−92 | ERYT2 |
| Hba-a1 | 3.76 × 10−68 | 6.565205 | 0.981 | 0.184 | 9.36 × 10−64 | ERYT2 |
| Bpgm | 3.23 × 10−62 | 3.488751 | 0.538 | 0.051 | 8.05 × 10−58 | ERYT2 |
| Fech | 4.01 × 10−31 | 4.045983 | 0.615 | 0.134 | 1.00 × 10−26 | ERYT2 |
| Tent5c | 2.97 × 10−19 | 3.63628 | 0.654 | 0.236 | 7.39 × 10−15 | ERYT2 |
| Mkrn1 | 6.39 × 10−14 | 3.270593 | 0.75 | 0.463 | 1.59 × 10−09 | ERYT2 |
| Plp1 | 0 | 8.631235 | 1 | 0.026 | 0 | Glial cell |
| Aplp1 | 0 | 6.191495 | 1 | 0.025 | 0 | Glial cell |
| Tubb4a | 0 | 6.185396 | 1 | 0.025 | 0 | Glial cell |
| Mag | 0 | 5.740784 | 1 | 0.014 | 0 | Glial cell |
| Cldn11 | 0 | 5.729523 | 1 | 0.01 | 0 | Glial cell |
| Mal | 4.01 × 10−230 | 5.340918 | 1 | 0.043 | 9.99 × 10−226 | Glial cell |
| Syt11 | 1.05 × 10−212 | 5.388431 | 1 | 0.047 | 2.62 × 10−208 | Glial cell |
| Enpp2 | 1.86 × 10−211 | 5.084028 | 1 | 0.047 | 4.62 × 10−207 | Glial cell |
| Scd2 | 8.20 × 10−69 | 5.230188 | 1 | 0.171 | 2.04 × 10−64 | Glial cell |
| Mbp | 1.58 × 10−45 | 5.137117 | 1 | 0.306 | 3.94 × 10−41 | Glial cell |
| Prlr | 0 | 4.387577 | 0.955 | 0.008 | 0 | IHC |
| Igfbp2 | 0 | 4.374309 | 1 | 0.012 | 0 | IHC |
| 1500015O10Rik | 0 | 4.269537 | 1 | 0.007 | 0 | IHC |
| Kl | 0 | 3.985654 | 1 | 0.004 | 0 | IHC |
| 2900040C04Rik | 0 | 3.664745 | 1 | 0.001 | 0 | IHC |
| Car12 | 6.04 × 10−308 | 3.42191 | 1 | 0.015 | 1.50 × 10−303 | IHC |
| Clu | 1.19 × 10−106 | 4.345548 | 1 | 0.046 | 2.96 × 10−102 | IHC |
| Enpp2 | 5.49 × 10−105 | 8.203794 | 1 | 0.048 | 1.37 × 10−100 | IHC |
| Slc22a17 | 3.77 × 10−99 | 4.114721 | 1 | 0.05 | 9.40 × 10−95 | IHC |
| Ttr | 4.09 × 10−96 | 9.836147 | 1 | 0.053 | 1.02 × 10−91 | IHC |
| Myo7a | 5.91 × 10−57 | 0.821842364 | 0.818 | 0.053 | 1.47 × 10−52 | IHC |
| Slc26a5 | 1.02 × 10−24 | 1.895397361 | 0.955 | 0.196 | 2.53 × 10−20 | IHC |
B cells: “Fcrla”. “Vpreb3”. “Cecr2”. “Fam129c”. “Cd79a”. Dendritic cells: “Siglech”. “Ccr9”. “Cd7”. “Cox6a2”. “Ly86”. Endothelial cells: “Ly6c1”. “Flt1”. “Ptprb”. “Cldn5”. “Adgrf5”. Epithelial cells: “Sod3”. “Krt18”. “Epcam”. “Aldh1a1”. “Hspb1”. Red blood cells: “Hba-a1”. “Hbb-bs”. “Slc4a1”. “Gypa”. “Aqp1”. Fibroblasts: “Col1a1”. “Col1a2”. “Apod”. “Dcn”. “Igfbp5”. Glial Cells: “Mag”. “Cldn11”. “Mobp”. “Ermn”. “Efnb3”. Inner hair cells: “Ttr”. “Enpp2”. “Prlr”. “Igfbp2”. “Clu”. Macrophages: “C1qc”. “C1qb”. “C1qa”. “Hspa1a”. “Hspa1b”. Mast cells: “Cd34”. “Slc18a2”. “Rps20”. “Rpl5”. “Eef1a1”. Monocytes: “Ass1”. “Pld4”. “Itga5”. “Ccdc88a”. Neutrophils: “Rsad2”. “Gbp2”. “Il1b”. “Ccrl2”. “Chil3”. Outer hair cells: “Nnat”. “Dlk1”. “Chgb”. “Meg3”. “Scg2”. Schwann cells: “Mpz”. “Art3”. “Slc6a15”. “Ncmap”. Smooth muscle cells: “Tagln”. “Myh11”. “Rgs 5”. “Mustn1”. “Notch3”. Strial basal cells: “Bpifa1”. “Lgals7”. “Krt5”. “Krt14”. “Cyp2f2”. Strial marginal cells: “Ptgds”. “Atp1b2”. “Slc12a2”. “Dnase1”. “Gpx3”. Strial intermediate cells: “Dct”. “Hpse”. “Gpnmb”. “Dkk3”. “Slc45a2”. T Cells: “Ccl5”. “Gzma”. “Il2rb”. “Trac”. “Xcl1”.
3.3 QC and sample filtering
To ensure high data quality and minimize contamination from blood-derived cells, we implemented a multi-step QC pipeline based on both global metrics and cell-type markers. First, we computed the following per-cell metrics: number of detected genes (nFeature_RNA), total UMI counts (nCount_RNA), mitochondrial transcript fraction (percent.mt), ribosomal transcript fraction (percent_ribo), and hemoglobin transcript fraction (percent_Hb). Cells satisfying 500 < nFeature_RNA < 6 000, nCount_RNA < 25,000, percent.mt < 20%, percent_ribo > 1%, and percent_Hb < 1% were retained for downstream analysis.
Next, we quantified expression of canonical neutrophil markers (e.g., Ly6g, Mpo) and confirmed that contaminating neutrophil-like cells accounted for <1% of all barcodes post-filtering.
Finally, batch effects arising from three experimental groups (NOR, GEN, DEX) were corrected using the Harmony algorithm, ensuring that downstream clustering reflects true biological variance rather than technical confounders.
4 Discussion
The study employed scRNA-seq technology to systematically analyze the impact of GEN intraperitoneal injection on the inner ear cells of mice, revealing characteristic changes across different cell types involved in ototoxic damage. The inner ear is a critical organ for auditory and vestibular functions, involving the coordination of various cell types such as hair cells, supporting cells, and strial vascular cells to maintain auditory function stability [27,28]. However, aminoglycoside antibiotics, such as GEN, can cause irreversible cochlear hair cell damage, leading to permanent hearing loss and vestibular dysfunction, whose underlying molecular mechanisms remain to be explored [29,30]. Traditional RNA sequencing methods, due to their reliance on tissue homogenization, cannot resolve the transcriptional features of specific cell types [4], whereas scRNA-seq technology overcomes this limitation, allowing researchers to reveal the gene expression patterns and drug responses of cochlear cells at single-cell resolution [31,32].
The study discovered, through ABR testing, that the auditory threshold in the GEN group was significantly higher than in the NOR group, while DEX group intervention partially improved the threshold, indicating that GEN significantly damages auditory function, and glucocorticoid drugs might have a protective effect on the cochlea. Further single-cell sequencing analysis revealed significant changes in the composition of cochlear hair cells, supporting cells, and immune cells in mice from the GEN group. Compared to the NOR group, the proportion of hair cells significantly decreased, whereas the proportion of T cells, B cells, macrophages, and Schwann cells significantly increased, suggesting that hair cell damage might be accompanied by activation of the inflammatory response. Furthermore, DEG analysis showed that genes specific to hair cells such as Nnat and Dlk1 were significantly downregulated in the GEN group, while genes related to the complement system, such as C1qa, C1qb, and C1qc, were notably upregulated, indicating a possible enhancement of local immune response accompanying hair cell damage.
This study applied scRNA-seq using the 10x Genomics platform with Seurat v4 for data processing, focusing on rigorous QC, UMAP clustering, and pathway analysis using clusterProfiler. This approach differs from previous study [33], which integrated snRNA-seq and scRNA-seq with Cell Ranger and performed trajectory analysis (Monocle, RNA velocity). Yan et al. used the Fluidigm C1 system for precise single-cell capture and performed pathway analysis with Metascape and transcription factor analysis via Cytoscape [34]. Compared to these studies, our method avoids complex integration and trajectory analysis but ensures robust cell-type classification and DEG validation using complementary techniques. This streamlined approach is suitable for capturing cell-type-specific responses in our acute injury model. Although minor blood cell contamination can occur during cochlear dissociation, our rigorous QC thresholds and marker-based exclusion reduced neutrophil-derived barcodes to negligible levels (<1%). We therefore conclude that residual neutrophil content does not compromise the integrity of our single-cell transcriptomic analyses.
Studies have shown that aminoglycoside antibiotics can bind to mitochondrial rRNA, inhibiting electron transport chain (ETC) complex activity, leading to increased reactive oxygen species levels, thereby causing mitochondrial oxidative damage [35]. The study found that genes related to OXPHOS, such as ND1, ND2, and CYTB, were significantly downregulated in the cochlea of GEN group mice, suggesting that mitochondrial dysfunction could be a crucial factor in hair cell death. Additionally, KEGG pathway analysis revealed significant activation of the p53 signaling pathway, TNF signaling pathway, and MAPK signaling pathway in the cochlea of GEN group mice. The p53 signaling pathway plays a vital role in apoptosis, and its upregulation can induce an imbalanced Bax/Bcl-2 ratio, activating the mitochondrial-dependent apoptotic pathway, ultimately leading to hair cell death [36,37]. Concurrently, activation of the TNF signaling pathway may exacerbate inflammatory responses, further deteriorating the cochlear microenvironment, accelerating hair cell loss [38,39].
The study’s scRNA-seq results also revealed GEN-induced immune-inflammatory responses. The proportions of macrophages, B cells, and T cells were significantly increased in the GEN group mice, with notable upregulation of C1q complement system genes (C1qa, C1qb, C1qc), suggesting that GEN might enhance local immune responses by activating the complement system and releasing inflammatory factors [40,41]. Previous studies have indicated that excessive activation of the complement system may disrupt the cochlear microenvironment, exacerbating hair cell apoptosis and worsening hearing loss [42].
Our findings align with and expand upon previous scRNA-seq studies investigating ototoxicity, providing further insight into the cellular and molecular mechanisms involved. For example, a study on the SV of the cochlea identified downregulation of mitochondrial ETC pathways as a common feature across various forms of sensorineural hearing loss, implicating mitochondrial dysfunction as a central contributor to SV impairment [43]. Another study using single-cell transcriptomics revealed the complex cellular landscape of the middle ear, highlighting the roles of innate immune responses, particularly among resident monocytes/macrophages, in mediating inflammation and tissue repair [44]. In the context of drug-induced ototoxicity, scRNA-seq analysis of cisplatin-treated SV identified transcriptional alterations affecting genes critical to EP generation and suggested potential therapeutic targets, including Alcam, Atp1b2, and Spp1, which could mitigate cisplatin-induced damage [45]. These findings collectively emphasize that while mitochondrial dysfunction and immune regulation are central to ototoxicity, this study provides a more nuanced view by directly analyzing the transcriptional responses of cochlear cells exposed to different treatments.
Moreover, the study found that after DEX intervention, the expression levels of some inflammation-related genes decreased, with the proportion of hair cells partially restored, indicating that glucocorticoids might have a protective role against GEN ototoxicity by suppressing inflammatory responses. An in vitro study using 3-day-old rat organ of Corti explants exposed to TNFα revealed that DXM effectively mitigated TNFα-induced hair cell loss. Specifically, DXM treatment counteracted TNFα-induced upregulation of the pro-apoptotic gene Bax and restored the expression of anti-apoptotic genes Bcl-2 and Bcl-xl [46]. These protective effects were also observed in DXM-eluting biopolymers (SIBS), which prevented hair cell death and maintained the anti- and pro-apoptotic gene expression profile similar to that of DXM-treated explants [47]. Furthermore, recent studies using an inner ear simulating system demonstrated that PSD-NPs loaded with DXM, particularly under a magnetic field, provided enhanced otoprotection compared to DXM alone, significantly reducing hearing loss through anti-apoptotic pathways [48]. However, the protective effect of DEX remains limited, with its precise mechanisms requiring further investigation.
Integrating AlphaFold with our scRNA-seq data can enhance understanding of GEN-induced ototoxicity. AlphaFold’s AI-driven structure prediction can reveal 3D configurations of key proteins, clarify protein–protein interactions, and identify GEN’s binding sites. It also helps explore how DEX modulates protective proteins, uncovering its anti-inflammatory and anti-apoptotic effects. By bridging transcriptomics and protein function, AlphaFold aids in discovering novel therapeutic targets and refining drug designs. This approach offers a high-precision framework for mitigating aminoglycoside-induced hearing loss [49].
Despite the insights provided by this study into the key cell types and molecular mechanisms underlying GEN ototoxicity, certain limitations remain. First, while single-cell sequencing offers high-resolution cellular transcriptome information, experimental procedures such as tissue dissociation and cell capture may result in the loss or bias of some cellular information. For instance, the inability to detect Prestin protein mRNA in this study may be related to its limited molecular structure and quantity. Moreover, this study primarily examines the short-term ototoxic effects following intraperitoneal injection of GEN; future research is required to explore its long-term mechanisms, including variability in ototoxicity across different doses and modes of drug administration. Another limitation of this study is the lack of frequency-specific ABR threshold data (e.g., 8–32 kHz), which would have offered more detailed insights into frequency-dependent hearing loss. Future studies should incorporate full-spectrum ABR evaluations to better characterize cochlear function across the tonotopic axis. In addition, due to pooling of ten cochleae per group into single libraries, differential abundance analyses should be interpreted with caution. Future studies will employ multiple biological replicates to validate these findings. We acknowledged the short-term nature of our exposure model and proposed long-term studies in future work to better reflect chronic clinical scenarios. Finally, although this study identifies numerous DEGs and signaling pathways through bioinformatic analysis, further experimental verification is necessary, such as evaluating the expression levels of key genes using techniques like western blot, qRT-PCR, and immunofluorescence staining, alongside cellular functional assays to explore their specific roles in GEN-induced ototoxicity.
In addition, our single-cell data provide gene-level insights into GEN ototoxicity. Future studies could leverage AI-driven structure prediction tools such as AlphaFold to map DEGs (e.g., complement gene C1q) onto 3D protein structures for drug–target interaction simulations [50]. For instance, GEN may disrupt C1q oligomerization, leading to complement activation, while direct binding to mitochondrial ETC proteins could underlie OXPHOS gene inhibition. Combining computational docking, molecular dynamics, and cryo-EM validation will facilitate structure-based design of otoprotective agents, accelerating translation from mechanistic discovery to clinical intervention [51].
5 Conclusion
To conclude, this study systematically elucidates the effects of GEN on different cell types in the mouse inner ear using scRNA-seq technology, revealing that hair cell damage, mitochondrial dysfunction, and immune-inflammatory responses may be pivotal mechanisms in its ototoxic pathway. DEX intervention is shown to partially alleviate hair cell damage, yet its mechanism of action and long-term protective effects warrant further investigation. The findings present potential biomarkers for early detection and targeted therapy of ototoxic damage and provide theoretical support for future ototoxicity prevention and treatment strategies.
Acknowledgments
Not applicable.
-
Funding information: No funding was used in this study.
-
Author contributions: Conception and design of the research: Guo Y.F., Bao X.L. Acquisition of data: Bao X.L., Wang Y., Liu W. Analysis and interpretation of the data: Bao X.L., Tang H.L., Liu W. Statistical analysis: Bao X.L., Wang Y., Liu W. Obtaining financing: None. Writing of the manuscript: Bao X.L., Tang H.L. Critical revision of the manuscript for intellectual content: Guo Y.F., Bao X.L.
-
Conflict of interest: The authors declare no competing interests.
-
Data availability statement: The datasets used and analyzed during the current study are available from the corresponding author on reasonable request.
References
[1] Kros CJ, Steyger PS. Aminoglycoside- and cisplatin-induced ototoxicity: mechanisms and otoprotective strategies. Cold Spring Harb Perspect Med. 2019;9(11):a033548. 10.1101/cshperspect.a033548.Search in Google Scholar PubMed PubMed Central
[2] Wu PZ, Liberman LD, Bennett K, de Gruttola V, O’Malley JT, Liberman MC. Primary neural degeneration in the human cochlea: evidence for hidden hearing loss. Neuroscience. 2019;407:8–20. 10.1016/j.neuroscience.2018.07.053.Search in Google Scholar PubMed PubMed Central
[3] Huth ME, Ricci AJ, Cheng AG. Mechanisms of aminoglycoside ototoxicity and targets of hair cell protection. Int J Otolaryngol. 2011;2011:937861. 10.1155/2011/937861.Search in Google Scholar PubMed PubMed Central
[4] Tan WJT, Song L. Role of mitochondrial dysfunction and oxidative stress in sensorineural hearing loss. Hear Res. 2023;434:108783. 10.1016/j.heares.2023.108783.Search in Google Scholar PubMed
[5] Li H, Steyger PS. Synergistic ototoxicity due to noise and chemical exposure: a systematic review. Front Neurol. 2021;12:654206. 10.3389/fneur.2021.654206.Search in Google Scholar
[6] Jean P, Wong Jun Tai F, Singh-Estivalet A, Lelli A, Scandola C, Megharba S, et al. Single-cell transcriptomic profiling of the mouse cochlea: an atlas for hearing research. Proc Natl Acad Sci U S A. 2023;120(26):e2221744120. 10.1073/pnas.2221744120.Search in Google Scholar PubMed PubMed Central
[7] Fischenich A, Hots J, Verhey JL, Oberfeld D. The effect of silent gaps on temporal weights in loudness judgments. Hear Res. 2020;395:108028. 10.1016/j.heares.2020.108028.Search in Google Scholar PubMed
[8] Kolla L, Kelly MC, Mann ZF, Anaya-Rocha A, Ellis K, Lemons A, et al. Characterization of the development of the mouse cochlear epithelium at the single cell level. Nat Commun. 2020;11(1):2389. 10.1038/s41467-020-16113-y.Search in Google Scholar PubMed PubMed Central
[9] Wan G, Gómez-Casati ME, Gigliello AR, Liberman MC, Corfas G. Neurotrophin-3 regulates ribbon synapse density in the cochlea and induces synapse regeneration after acoustic trauma. Elife. 2014;3:e03564. 10.7554/eLife.03564.Search in Google Scholar PubMed PubMed Central
[10] Ghelfi E, Grondin Y, Millet EJ, Bartos A, Bortoni M, Oliveira Gomes Dos Santos C, et al. In vitro gentamicin exposure alters caveolae protein profile in cochlear spiral ligament pericytes. Proteome Sci. 2018;16:7. 10.1186/s12953-018-0132-x.Search in Google Scholar PubMed PubMed Central
[11] Gonçalves AC, Towers ER, Haq N, Porco Jr JA, Pelletier J, Dawson SJ, et al. Drug-induced stress granule formation protects sensory hair cells in mouse cochlear explants during ototoxicity. Sci Rep. 2019;9(1):12501. 10.1038/s41598-019-48393-w.Search in Google Scholar PubMed PubMed Central
[12] Ting HY, Yi HY, Shing L. EGb 761 (Ginkgo biloba) protects cochlear hair cells against ototoxicity induced by gentamicin via reducing reactive oxygen species and nitric oxide-related apoptosis. J Nutr Biochem. 2011;22(9):886–94. 10.1016/j.jnutbio.2010.08.009.Search in Google Scholar PubMed
[13] Tona Y, Inaoka T, Ito J, Kawano S, Nakagawa T. Development of an electrode for the artificial cochlear sensory epithelium. Hear Res. 2015;330(Pt A):106–12. 10.1016/j.heares.2015.08.007.Search in Google Scholar PubMed
[14] Ximerakis M, Lipnick SL, Innes BT, Simmons SK, Adiconis X, Dionne D, et al. Single-cell transcriptomic profiling of the aging mouse brain. Nat Neurosci. 2019;122(10):1696–708. 10.1038/s41593-019-0491-3.Search in Google Scholar PubMed
[15] Milon B, Shulman ED, So KS, Cederroth CR, Lipford EL, Sperber M, et al. A cell-type-specific atlas of the inner ear transcriptional response to acoustic trauma. Cell Rep. 2021;36(3):109758. 10.1016/j.celrep.2021.109758.Search in Google Scholar PubMed PubMed Central
[16] Li F, Yan K, Wu L, Zheng Z, Du Y, Liu Z, et al. Single-cell RNA-seq reveals cellular heterogeneity of mouse carotid artery under disturbed flow. Development. 2020;7(1):180. 10.1038/s41420-021-00567-0.Search in Google Scholar PubMed PubMed Central
[17] Li Y, Huang Z, Xu L, Fan Y, Ping J, Li G, et al. UDA-seq: universal droplet microfluidics-based combinatorial indexing for massive-scale multimodal single-cell sequencing. Nat Methods. 2025;22:1199–212. 10.1038/s41592-024-02586-y. Search in Google Scholar PubMed PubMed Central
[18] Eva H, Qiaolin D. Single-cell RNA sequencing: technical advancements and biological applications. Mol Asp Med. 2018;59:36–46. 10.1016/j.mam.2017.07.003.Search in Google Scholar PubMed
[19] Koh JY, Affortit C, Ranum PT, West C, Walls WD, Yoshimura H, et al. Single-cell RNA-sequencing of stria vascularis cells in the adult Slc26a4-/- mouseC. BMC Med Genomics. 2020;16(1):133. 10.1186/s12920-023-01549-0.Search in Google Scholar PubMed PubMed Central
[20] Kubota M, Scheibinger M, Jan TA, Heller S. Greater epithelial ridge cells are the principal organoid-forming progenitors of the mouse cochlea. Cell Rep. 2021;34(3):108646. 10.1016/j.celrep.2020.108646.Search in Google Scholar PubMed PubMed Central
[21] Bas E, Bohorquez J, Goncalves S, Perez E, Dinh CT, Garnham C, et al. Electrode array-eluted dexamethasone protects against electrode insertion trauma induced hearing and hair cell losses, damage to neural elements, increases in impedance and fibrosis: a dose response study. Hear Res. 2016;337:12–24. 10.1016/j.heares.2016.02.003.Search in Google Scholar PubMed
[22] Lee JH, Oh SH, Kim TH, Go YY, Song JJ. Anti-apoptotic effect of dexamethasone in an ototoxicity model. Biomater Res. 2017;21:4. 10.1186/s40824-017-0090-x.Search in Google Scholar PubMed PubMed Central
[23] Chen L, Xiong S, Liu Y, Shang X. Effect of different gentamicin dose on the plasticity of the ribbon synapses in cochlear inner hair cells of C57BL/6J mice. Mol Neurobiol. 2012;46(2):487–94. 10.1007/s12035-012-8312-7.Search in Google Scholar PubMed
[24] Li G, Gao Y, Wu H, Zhao T. Gentamicin administration leads to synaptic dysfunction in inner hair cells. Toxicol Lett. 2024;391:86–99. 10.1016/j.toxlet.2023.12.007.Search in Google Scholar PubMed
[25] Satija R, Farrell JA, Gennert D, Schier AF, Regev A. Spatial reconstruction of single-cell gene expression data. Nat Biotechnol. 2015;33(5):495–502. 10.1038/nbt.3192.Search in Google Scholar PubMed PubMed Central
[26] Butler A, Hoffman P, Smibert P, Papalexi E, Satija R. Integrating single-cell transcriptomic data across different conditions, technologies, and species. Nat Biotechnol. 2018;36(5):411–20. 10.1038/nbt.4096.Search in Google Scholar PubMed PubMed Central
[27] Breglio AM, Rusheen AE, Shide ED, Fernandez KA, Spielbauer KK, McLachlin KM, et al. Cisplatin is retained in the cochlea indefinitely following chemotherapy. Nat Commun. 2017;8:1654. 10.1038/s41467-017-01837-1.Search in Google Scholar PubMed PubMed Central
[28] Mittal R, Bencie N, Liu G, Eshraghi N, Nisenbaum E, Blanton SH, et al. Recent advancements in understanding the role of epigenetics in the auditory system. Gene. 2020;761:144996. 10.1016/j.gene.2020.144996.Search in Google Scholar PubMed PubMed Central
[29] Huang Y, Xu W, Zhou R. NLRP3 inflammasome activation and cell death. Front Immunol. 2021;18(9):2114–27. 10.1038/s41423-021-00740-6.Search in Google Scholar PubMed PubMed Central
[30] Hu BH, Cai Q, Manohar S, Jiang H, Ding D, Coling DE, et al. Differential expression of apoptosis-related genes in the cochlea of noise-exposed rats. Neuroscience. 2009;161(3):915–25. 10.1016/j.neuroscience.2009.03.072.Search in Google Scholar PubMed PubMed Central
[31] Kang C, Li X, Liu P, Liu Y, Niu Y, Zeng X, et al. Tolerogenic dendritic cells and TLR4/IRAK4/NF-κB signaling pathway in allergic rhinitis. J Neuroinflammation. 2023;14:1276512. 10.3389/fimmu.2023.1276512.Search in Google Scholar PubMed PubMed Central
[32] Pan H, Shen Z, Mukhopadhyay P, Wang H, Pacher P, Qin X, et al. Anaphylatoxin C5a contributes to the pathogenesis of cisplatin-induced nephrotoxicity. Am J Physiol Ren Physiol. 2009;296(3):F496–504. 10.1152/ajprenal.90443.2008.Search in Google Scholar PubMed PubMed Central
[33] Liu Q, Long Q, Zhao J, Wu W, Lin Z, Sun W, et al. Cold-induced reprogramming of subcutaneous white adipose tissue assessed by single-cell and single-nucleus RNA sequencing. Research. 2023;6:0182. 10.34133/research.0182.Search in Google Scholar PubMed PubMed Central
[34] Yan R, Zhang H, Ma Y, Lin R, Zhou B, Zhang T, et al. Discovery of Muscle-tendon progenitor subpopulation in human myotendinous junction at single-cell resolution. Research. 2022;28:9760390. 10.34133/2022/9760390.Search in Google Scholar PubMed PubMed Central
[35] O’Reilly M, Young L, Kirkwood NK, Richardson GP, Kros CJ, Moore AL. Gentamicin affects the bioenergetics of isolated mitochondria and collapses the mitochondrial membrane potential in cochlear sensory hair cells. Front Cell Neurosci. 2019;13:416. 10.3389/fncel.2019.00416.Search in Google Scholar PubMed PubMed Central
[36] Niu P, Sun Y, Wang S, Li G, Tang X, Sun J, et al. Puerarin alleviates the ototoxicity of gentamicin by inhibiting the mitochondria‑dependent apoptosis pathway. Mol Med Rep. 2021;24(6):851. 10.3892/mmr.2021.12491.Search in Google Scholar PubMed PubMed Central
[37] Yang Q, Zhou Y, Yin H, Li H, Zhou M, Sun G, et al. PINK1 protects against gentamicin-induced sensory hair cell damage: possible relation to induction of autophagy and inhibition of p53 signal pathway. Front Mol Neurosci. 2018;11:403. 10.3389/fnmol.2018.00403.Search in Google Scholar PubMed PubMed Central
[38] Zhang Y, Chen D, Zhao L, Li W, Ni Y, Chen Y, et al. Nfatc4 deficiency attenuates ototoxicity by suppressing Tnf-mediated hair cell apoptosis in the mouse cochlea. Front Immunol. 2019;10:1660. 10.3389/fimmu.2019.0166.Search in Google Scholar
[39] Chen D, Jia G, Zhang Y, Mao H, Zhao L, Li W, et al. Sox2 overexpression alleviates noise-induced hearing loss by inhibiting inflammation-related hair cell apoptosis. J Neuroinflammation. 2022;19(1):59. 10.1186/s12974-022-02414-0.Search in Google Scholar PubMed PubMed Central
[40] Sun C, Wang X, Chen D, Lin X, Yu D, Wu H. Dexamethasone loaded nanoparticles exert protective effects against Cisplatin-induced hearing loss by systemic administration. Neurosci Lett. 2016;619:142–8. 10.1016/j.neulet.2016.03.012.Search in Google Scholar PubMed
[41] Chattaraj A, Syed MP, Low CA, Owonikoko TK. Cisplatin-induced ototoxicity: a concise review of the burden, prevention, and interception strategies. JCO Oncol Pract. 2023;19(5):278–83. 10.1200/OP.22.00710.Search in Google Scholar PubMed PubMed Central
[42] Schiel V, Bhattacharya R, Gupta A, Eftekharian K, Xia A, Santa Maria PL. Targeting the NLRP3 inflammasome in cochlear macrophages protects against hearing loss in chronic suppurative otitis media. J Neuroinflammation. 2024;21(1):223. 10.1186/s12974-024-03212-6.Search in Google Scholar PubMed PubMed Central
[43] Gu X, Jiang K, Chen R, Chen Z, Wu X, Xiang H, et al. Identification of common stria vascularis cellular alteration in sensorineural hearing loss based on ScRNA-seq. BMC Genomics. 2024;25(1):213. 10.1186/s12864-024-10122-7.Search in Google Scholar PubMed PubMed Central
[44] Ryan AF, Nasamran CA, Pak K, Draf C, Fisch KM, Webster N, et al. Single-cell transcriptomes reveal a complex cellular landscape in the middle ear and differential capacities for acute response to infection. Front Genet. 2020;11:358. 10.3389/fgene.2020.00358.Search in Google Scholar PubMed PubMed Central
[45] Taukulis IA, Olszewski RT, Korrapati S, Fernandez KA, Boger ET, Fitzgerald TS, et al. Single-cell RNA-Seq of cisplatin-treated adult stria vascularis identifies cell type-specific regulatory networks and novel therapeutic gene targets. Front Mol Neurosci. 2021;14:718241. 10.3389/fnmol.2021.718241.Search in Google Scholar PubMed PubMed Central
[46] Dinh CT, Haake S, Chen S, Hoang K, Nong E, Eshraghi AA, et al. Dexamethasone protects organ of corti explants against tumor necrosis factor-alpha-induced loss of auditory hair cells and alters the expression levels of apoptosis-related genes. Neuroscience. 2008;157(2):405–13. 10.1016/j.neuroscience.2008.09.012.Search in Google Scholar PubMed
[47] Dinh C, Hoang K, Haake S, Chen S, Angeli S, Nong E, et al. Biopolymer-released dexamethasone prevents tumor necrosis factor alpha-induced loss of auditory hair cells in vitro: implications toward the development of a drug-eluting cochlear implant electrode array. Otol Neurotol. 2008;29(7):1012–9. 10.1097/MAO.0b013e3181859a1f.Search in Google Scholar PubMed
[48] Park JE, Kim WC, Kim SK, Ahn Y, Ha SM, Kim G, et al. Protection of hearing loss in ototoxic mouse model through SPIONs and dexamethasone-loaded PLGA nanoparticle delivery by magnetic attraction. Int J Nanomed. 2022;17:6317–34. 10.2147/IJN.S380810.Search in Google Scholar PubMed PubMed Central
[49] Guo SB, Meng Y, Lin L, Zhou ZZ, Li HL, Tian XP, et al. Artificial intelligence alphafold model for molecular biology and drug discovery: a machine-learning-driven informatics investigation. Mol Cancer. 2024;23(1):223. 10.1186/s12943-024-02140-6.Search in Google Scholar PubMed PubMed Central
[50] Xu K, Feng H, Zhang H, He C, Kang H, Yuan T, et al. Structure-guided discovery of highly efficient cytidine deaminases with sequence-context independence. Nat Biomed Eng. 2025;9(1):93–108. 10.1038/s41551-024-01220-8.Search in Google Scholar PubMed PubMed Central
[51] Krokidis MG, Koumadorakis DE, Lazaros K, Ivantsik O, Exarchos TP, Vrahatis AG, et al. AlphaFold3: an overview of applications and performance insights. Int J Mol Sci. 2025;26(8):3671. 10.3390/ijms26083671.Search in Google Scholar PubMed PubMed Central
© 2025 the author(s), published by De Gruyter
This work is licensed under the Creative Commons Attribution 4.0 International License.
Articles in the same Issue
- Research Articles
- Network pharmacological analysis and in vitro testing of the rutin effects on triple-negative breast cancer
- Impact of diabetes on long-term survival in elderly liver cancer patients: A retrospective study
- Knockdown of CCNB1 alleviates high glucose-triggered trophoblast dysfunction during gestational diabetes via Wnt/β-catenin signaling pathway
- Risk factors for severe adverse drug reactions in hospitalized patients
- Analysis of the effect of ALA-PDT on macrophages in footpad model of mice infected with Fonsecaea monophora based on single-cell sequencing
- Development and validation of headspace gas chromatography with a flame ionization detector method for the determination of ethanol in the vitreous humor
- CMSP exerts anti-tumor effects on small cell lung cancer cells by inducing mitochondrial dysfunction and ferroptosis
- Predictive value of plasma sB7-H3 and YKL-40 in pediatric refractory Mycoplasma pneumoniae pneumonia
- Antiangiogenic potential of Elaeagnus umbellata extracts and molecular docking study by targeting VEGFR-2 pathway
- Comparison of the effectiveness of nurse-led preoperative counseling and postoperative follow-up care vs standard care for patients with gastric cancer
- Comparing the therapeutic efficacy of endoscopic minimally invasive surgery and traditional surgery for early-stage breast cancer: A meta-analysis
- Adhered macrophages as an additional marker of cardiomyocyte injury in biopsies of patients with dilated cardiomyopathy
- Association between statin administration and outcome in patients with sepsis: A retrospective study
- Exploration of the association between estimated glucose disposal rate and osteoarthritis in middle-aged and older adults: An analysis of NHANES data from 2011 to 2018
- A comparative analysis of the binary and multiclass classified chest X-ray images of pneumonia and COVID-19 with ML and DL models
- Lysophosphatidic acid 2 alleviates deep vein thrombosis via protective endothelial barrier function
- Transcription factor A, mitochondrial promotes lymph node metastasis and lymphangiogenesis in epithelial ovarian carcinoma
- Serum PM20D1 levels are associated with nutritional status and inflammatory factors in gastric cancer patients undergoing early enteral nutrition
- Hydromorphone reduced the incidence of emergence agitation after adenotonsillectomy in children with obstructive sleep apnea: A randomized, double-blind study
- Vitamin D replacement therapy may regulate sleep habits in patients with restless leg syndrome
- The first-line antihypertensive nitrendipine potentiated the therapeutic effect of oxaliplatin by downregulating CACNA1D in colorectal cancer
- Health literacy and health-related quality of life: The mediating role of irrational happiness
- Modulatory effects of Lycium barbarum polysaccharide on bone cell dynamics in osteoporosis
- Mechanism research on inhibition of gastric cancer in vitro by the extract of Pinellia ternata based on network pharmacology and cellular metabolomics
- Examination of the causal role of immune cells in non-alcoholic fatty liver disease by a bidirectional Mendelian randomization study
- Clinical analysis of ten cases of HIV infection combined with acute leukemia
- Investigating the cardioprotective potential of quercetin against tacrolimus-induced cardiotoxicity in Wistar rats: A mechanistic insights
- Clinical observation of probiotics combined with mesalazine and Yiyi Baitouweng Decoction retention enema in treating mild-to-moderate ulcerative colitis
- Diagnostic value of ratio of blood inflammation to coagulation markers in periprosthetic joint infection
- Sex-specific associations of sex hormone binding globulin and risk of bladder cancer
- Core muscle strength and stability-oriented breathing training reduces inter-recti distance in postpartum women
- The ERAS nursing care strategy for patients undergoing transsphenoidal endoscopic pituitary tumor resection: A randomized blinded controlled trial
- The serum IL-17A levels in patients with traumatic bowel rupture post-surgery and its predictive value for patient prognosis
- Impact of Kolb’s experiential learning theory-based nursing on caregiver burden and psychological state of caregivers of dementia patients
- Analysis of serum NLR combined with intraoperative margin condition to predict the prognosis of cervical HSIL patients undergoing LEEP surgery
- Commiphora gileadensis ameliorate infertility and erectile dysfunction in diabetic male mice
- The correlation between epithelial–mesenchymal transition classification and MMP2 expression of circulating tumor cells and prognosis of advanced or metastatic nasopharyngeal carcinoma
- Tetrahydropalmatine improves mitochondrial function in vascular smooth muscle cells of atherosclerosis in vitro by inhibiting Ras homolog gene family A/Rho-associated protein kinase-1 signaling pathway
- A cross-sectional study: Relationship between serum oxidative stress levels and arteriovenous fistula maturation in maintenance dialysis patients
- A comparative analysis of the impact of repeated administration of flavan 3-ol on brown, subcutaneous, and visceral adipose tissue
- Identifying early screening factors for depression in middle-aged and older adults: A cohort study
- Perform tumor-specific survival analysis for Merkel cell carcinoma patients undergoing surgical resection based on the SEER database by constructing a nomogram chart
- Unveiling the role of CXCL10 in pancreatic cancer progression: A novel prognostic indicator
- High-dose preoperative intraperitoneal erythropoietin and intravenous methylprednisolone in acute traumatic spinal cord injuries following decompression surgeries
- RAB39B: A novel biomarker for acute myeloid leukemia identified via multi-omics and functional validation
- Impact of peripheral conditioning on reperfusion injury following primary percutaneous coronary intervention in diabetic and non-diabetic STEMI patients
- Clinical efficacy of azacitidine in the treatment of middle- and high-risk myelodysplastic syndrome in middle-aged and elderly patients: A retrospective study
- The effect of ambulatory blood pressure load on mitral regurgitation in continuous ambulatory peritoneal dialysis patients
- Expression and clinical significance of ITGA3 in breast cancer
- Single-nucleus RNA sequencing reveals ARHGAP28 expression of podocytes as a biomarker in human diabetic nephropathy
- rSIG combined with NLR in the prognostic assessment of patients with multiple injuries
- Toxic metals and metalloids in collagen supplements of fish and jellyfish origin: Risk assessment for daily intake
- Exploring causal relationship between 41 inflammatory cytokines and marginal zone lymphoma: A bidirectional Mendelian randomization study
- Gender beliefs and legitimization of dating violence in adolescents
- Effect of serum IL-6, CRP, and MMP-9 levels on the efficacy of modified preperitoneal Kugel repair in patients with inguinal hernia
- Effect of smoking and smoking cessation on hematological parameters in polycythemic patients
- Pathogen surveillance and risk factors for pulmonary infection in patients with lung cancer: A retrospective single-center study
- Necroptosis of hippocampal neurons in paclitaxel chemotherapy-induced cognitive impairment mediates microglial activation via TLR4/MyD88 signaling pathway
- Celastrol suppresses neovascularization in rat aortic vascular endothelial cells stimulated by inflammatory tenocytes via modulating the NLRP3 pathway
- Cord-lamina angle and foraminal diameter as key predictors of C5 palsy after anterior cervical decompression and fusion surgery
- GATA1: A key biomarker for predicting the prognosis of patients with diffuse large B-cell lymphoma
- Influencing factors of false lumen thrombosis in type B aortic dissection: A single-center retrospective study
- MZB1 regulates the immune microenvironment and inhibits ovarian cancer cell migration
- Integrating experimental and network pharmacology to explore the pharmacological mechanisms of Dioscin against glioblastoma
- Trends in research on preterm birth in twin pregnancy based on bibliometrics
- Four-week IgE/baseline IgE ratio combined with tryptase predicts clinical outcome in omalizumab-treated children with moderate-to-severe asthma
- Single-cell transcriptomic analysis identifies a stress response Schwann cell subtype
- Acute pancreatitis risk in the diagnosis and management of inflammatory bowel disease: A critical focus
- Effect of subclinical esketamine on NLRP3 and cognitive dysfunction in elderly ischemic stroke patients
- Interleukin-37 mediates the anti-oral tumor activity in oral cancer through STAT3
- CA199 and CEA expression levels, and minimally invasive postoperative prognosis analysis in esophageal squamous carcinoma patients
- Efficacy of a novel drainage catheter in the treatment of CSF leak after posterior spine surgery: A retrospective cohort study
- Comprehensive biomedicine assessment of Apteranthes tuberculata extracts: Phytochemical analysis and multifaceted pharmacological evaluation in animal models
- Relation of time in range to severity of coronary artery disease in patients with type 2 diabetes: A cross-sectional study
- Dopamine attenuates ethanol-induced neuronal apoptosis by stimulating electrical activity in the developing rat retina
- Correlation between albumin levels during the third trimester and the risk of postpartum levator ani muscle rupture
- Factors associated with maternal attention and distraction during breastfeeding and childcare: A cross-sectional study in the west of Iran
- Mechanisms of hesperetin in treating metabolic dysfunction-associated steatosis liver disease via network pharmacology and in vitro experiments
- The law on oncological oblivion in the Italian and European context: How to best uphold the cancer patients’ rights to privacy and self-determination?
- The prognostic value of the neutrophil-to-lymphocyte ratio, platelet-to-lymphocyte ratio, and prognostic nutritional index for survival in patients with colorectal cancer
- Factors affecting the measurements of peripheral oxygen saturation values in healthy young adults
- Comparison and correlations between findings of hysteroscopy and vaginal color Doppler ultrasonography for detection of uterine abnormalities in patients with recurrent implantation failure
- The effects of different types of RAGT on balance function in stroke patients with low levels of independent walking in a convalescent rehabilitation hospital
- Causal relationship between asthma and ankylosing spondylitis: A bidirectional two-sample univariable and multivariable Mendelian randomization study
- Correlations of health literacy with individuals’ understanding and use of medications in Southern Taiwan
- Correlation of serum calprotectin with outcome of acute cerebral infarction
- Comparison of computed tomography and guided bronchoscopy in the diagnosis of pulmonary nodules: A systematic review and meta-analysis
- Curdione protects vascular endothelial cells and atherosclerosis via the regulation of DNMT1-mediated ERBB4 promoter methylation
- The identification of novel missense variant in ChAT gene in a patient with gestational diabetes denotes plausible genetic association
- Molecular genotyping of multi-system rare blood types in foreign blood donors based on DNA sequencing and its clinical significance
- Exploring the role of succinyl carnitine in the association between CD39⁺ CD4⁺ T cell and ulcerative colitis: A Mendelian randomization study
- Dexmedetomidine suppresses microglial activation in postoperative cognitive dysfunction via the mmu-miRNA-125/TRAF6 signaling axis
- Analysis of serum metabolomics in patients with different types of chronic heart failure
- Diagnostic value of hematological parameters in the early diagnosis of acute cholecystitis
- Pachymaran alleviates fat accumulation, hepatocyte degeneration, and injury in mice with nonalcoholic fatty liver disease
- Decrease in CD4 and CD8 lymphocytes are predictors of severe clinical picture and unfavorable outcome of the disease in patients with COVID-19
- METTL3 blocked the progression of diabetic retinopathy through m6A-modified SOX2
- The predictive significance of anti-RO-52 antibody in patients with interstitial pneumonia after treatment of malignant tumors
- Exploring cerebrospinal fluid metabolites, cognitive function, and brain atrophy: Insights from Mendelian randomization
- Development and validation of potential molecular subtypes and signatures of ocular sarcoidosis based on autophagy-related gene analysis
- Widespread venous thrombosis: Unveiling a complex case of Behçet’s disease with a literature perspective
- Uterine fibroid embolization: An analysis of clinical outcomes and impact on patients’ quality of life
- Discovery of lipid metabolism-related diagnostic biomarkers and construction of diagnostic model in steroid-induced osteonecrosis of femoral head
- Serum-derived exomiR-188-3p is a promising novel biomarker for early-stage ovarian cancer
- Enhancing chronic back pain management: A comparative study of ultrasound–MRI fusion guidance for paravertebral nerve block
- Peptide CCAT1-70aa promotes hepatocellular carcinoma proliferation and invasion via the MAPK/ERK pathway
- Electroacupuncture-induced reduction of myocardial ischemia–reperfusion injury via FTO-dependent m6A methylation modulation
- Hemorrhoids and cardiovascular disease: A bidirectional Mendelian randomization study
- Cell-free adipose extract inhibits hypertrophic scar formation through collagen remodeling and antiangiogenesis
- HALP score in Demodex blepharitis: A case–control study
- Assessment of SOX2 performance as a marker for circulating cancer stem-like cells (CCSCs) identification in advanced breast cancer patients using CytoTrack system
- Risk and prognosis for brain metastasis in primary metastatic cervical cancer patients: A population-based study
- Comparison of the two intestinal anastomosis methods in pediatric patients
- Factors influencing hematological toxicity and adverse effects of perioperative hyperthermic intraperitoneal vs intraperitoneal chemotherapy in gastrointestinal cancer
- Endotoxin tolerance inhibits NLRP3 inflammasome activation in macrophages of septic mice by restoring autophagic flux through TRIM26
- Lateral transperitoneal laparoscopic adrenalectomy: A single-centre experience of 21 procedures
- Petunidin attenuates lipopolysaccharide-induced retinal microglia inflammatory response in diabetic retinopathy by targeting OGT/NF-κB/LCN2 axis
- Procalcitonin and C-reactive protein as biomarkers for diagnosing and assessing the severity of acute cholecystitis
- Factors determining the number of sessions in successful extracorporeal shock wave lithotripsy patients
- Development of a nomogram for predicting cancer-specific survival in patients with renal pelvic cancer following surgery
- Inhibition of ATG7 promotes orthodontic tooth movement by regulating the RANKL/OPG ratio under compression force
- A machine learning-based prognostic model integrating mRNA stemness index, hypoxia, and glycolysis‑related biomarkers for colorectal cancer
- Glutathione attenuates sepsis-associated encephalopathy via dual modulation of NF-κB and PKA/CREB pathways
- FAHD1 prevents neuronal ferroptosis by modulating R-loop and the cGAS–STING pathway
- Association of placenta weight and morphology with term low birth weight: A case–control study
- Investigation of the pathogenic variants induced Sjogren’s syndrome in Turkish population
- Nucleotide metabolic abnormalities in post-COVID-19 condition and type 2 diabetes mellitus patients and their association with endocrine dysfunction
- TGF-β–Smad2/3 signaling in high-altitude pulmonary hypertension in rats: Role and mechanisms via macrophage M2 polarization
- Ultrasound-guided unilateral versus bilateral erector spinae plane block for postoperative analgesia of patients undergoing laparoscopic cholecystectomy
- Profiling gut microbiome dynamics in subacute thyroiditis: Implications for pathogenesis, diagnosis, and treatment
- Delta neutrophil index, CRP/albumin ratio, procalcitonin, immature granulocytes, and HALP score in acute appendicitis: Best performing biomarker?
- Anticancer activity mechanism of novelly synthesized and characterized benzofuran ring-linked 3-nitrophenyl chalcone derivative on colon cancer cells
- H2valdien3 arrests the cell cycle and induces apoptosis of gastric cancer
- Prognostic relevance of PRSS2 and its immune correlates in papillary thyroid carcinoma
- Association of SGLT2 inhibition with psychiatric disorders: A Mendelian randomization study
- Motivational interviewing for alcohol use reduction in Thai patients
- Luteolin alleviates oxygen-glucose deprivation/reoxygenation-induced neuron injury by regulating NLRP3/IL-1β signaling
- Polyphyllin II inhibits thyroid cancer cell growth by simultaneously inhibiting glycolysis and oxidative phosphorylation
- Relationship between the expression of copper death promoting factor SLC31A1 in papillary thyroid carcinoma and clinicopathological indicators and prognosis
- CSF2 polarized neutrophils and invaded renal cancer cells in vitro influence
- Proton pump inhibitors-induced thrombocytopenia: A systematic literature analysis of case reports
- The current status and influence factors of research ability among community nurses: A sequential qualitative–quantitative study
- OKAIN: A comprehensive oncology knowledge base for the interpretation of clinically actionable alterations
- The relationship between serum CA50, CA242, and SAA levels and clinical pathological characteristics and prognosis in patients with pancreatic cancer
- Identification and external validation of a prognostic signature based on hypoxia–glycolysis-related genes for kidney renal clear cell carcinoma
- Engineered RBC-derived nanovesicles functionalized with tumor-targeting ligands: A comparative study on breast cancer targeting efficiency and biocompatibility
- Relationship of resting echocardiography combined with serum micronutrients to the severity of low-gradient severe aortic stenosis
- Effect of vibration on pain during subcutaneous heparin injection: A randomized, single-blind, placebo-controlled trial
- The diagnostic performance of machine learning-based FFRCT for coronary artery disease: A meta-analysis
- Comparing biofeedback device vs diaphragmatic breathing for bloating relief: A randomized controlled trial
- Serum uric acid to albumin ratio and C-reactive protein as predictive biomarkers for chronic total occlusion and coronary collateral circulation quality
- Multiple organ scoring systems for predicting in-hospital mortality of sepsis patients in the intensive care unit
- Single-cell RNA sequencing data analysis of the inner ear in gentamicin-treated mice via intraperitoneal injection
- Review Articles
- The effects of enhanced external counter-pulsation on post-acute sequelae of COVID-19: A narrative review
- Diabetes-related cognitive impairment: Mechanisms, symptoms, and treatments
- Microscopic changes and gross morphology of placenta in women affected by gestational diabetes mellitus in dietary treatment: A systematic review
- Review of mechanisms and frontier applications in IL-17A-induced hypertension
- Research progress on the correlation between islet amyloid peptides and type 2 diabetes mellitus
- The safety and efficacy of BCG combined with mitomycin C compared with BCG monotherapy in patients with non-muscle-invasive bladder cancer: A systematic review and meta-analysis
- The application of augmented reality in robotic general surgery: A mini-review
- The effect of Greek mountain tea extract and wheat germ extract on peripheral blood flow and eicosanoid metabolism in mammals
- Neurogasobiology of migraine: Carbon monoxide, hydrogen sulfide, and nitric oxide as emerging pathophysiological trinacrium relevant to nociception regulation
- Plant polyphenols, terpenes, and terpenoids in oral health
- Laboratory medicine between technological innovation, rights safeguarding, and patient safety: A bioethical perspective
- End-of-life in cancer patients: Medicolegal implications and ethical challenges in Europe
- The maternal factors during pregnancy for intrauterine growth retardation: An umbrella review
- Intra-abdominal hypertension/abdominal compartment syndrome of pediatric patients in critical care settings
- PI3K/Akt pathway and neuroinflammation in sepsis-associated encephalopathy
- Screening of Group B Streptococcus in pregnancy: A systematic review for the laboratory detection
- Giant borderline ovarian tumours – review of the literature
- Leveraging artificial intelligence for collaborative care planning: Innovations and impacts in shared decision-making – A systematic review
- Cholera epidemiology analysis through the experience of the 1973 Naples epidemic
- Risk factors of frailty/sarcopenia in community older adults: Meta-analysis
- Supplement strategies for infertility in overweight women: Evidence and legal insights
- Scurvy, a not obsolete disorder: Clinical report in eight young children and literature review
- A meta-analysis of the effects of DBS on cognitive function in patients with advanced PD
- Protective role of selenium in sepsis: Mechanisms and potential therapeutic strategies
- Strategies for hyperkalemia management in dialysis patients: A systematic review
- C-reactive protein-to-albumin ratio in peripheral artery disease
- Case Reports
- Delayed graft function after renal transplantation
- Semaglutide treatment for type 2 diabetes in a patient with chronic myeloid leukemia: A case report and review of the literature
- Diverse electrophysiological demyelinating features in a late-onset glycogen storage disease type IIIa case
- Giant right atrial hemangioma presenting with ascites: A case report
- Laser excision of a large granular cell tumor of the vocal cord with subglottic extension: A case report
- EsoFLIP-assisted dilation for dysphagia in systemic sclerosis: Highlighting the role of multimodal esophageal evaluation
- Molecular hydrogen-rhodiola as an adjuvant therapy for ischemic stroke in internal carotid artery occlusion: A case report
- Coronary artery anomalies: A case of the “malignant” left coronary artery and its surgical management
- Rapid Communication
- Biological properties of valve materials using RGD and EC
-
A single oral administration of flavanols enhances short
-term memory in mice along with increased brain-derived neurotrophic factor - Letter to the Editor
- Role of enhanced external counterpulsation in long COVID
- Expression of Concern
- Expression of concern “A ceRNA network mediated by LINC00475 in papillary thyroid carcinoma”
- Expression of concern “Notoginsenoside R1 alleviates spinal cord injury through the miR-301a/KLF7 axis to activate Wnt/β-catenin pathway”
- Expression of concern “circ_0020123 promotes cell proliferation and migration in lung adenocarcinoma via PDZD8”
- Corrigendum
- Corrigendum to “Empagliflozin improves aortic injury in obese mice by regulating fatty acid metabolism”
- Corrigendum to “Comparing the therapeutic efficacy of endoscopic minimally invasive surgery and traditional surgery for early-stage breast cancer: A meta-analysis”
- Corrigendum to “The progress of autoimmune hepatitis research and future challenges”
- Retraction
- Retraction of “miR-654-5p promotes gastric cancer progression via the GPRIN1/NF-κB pathway”
- Retraction of: “LncRNA CASC15 inhibition relieves renal fibrosis in diabetic nephropathy through downregulating SP-A by sponging to miR-424”
- Retraction of: “SCARA5 inhibits oral squamous cell carcinoma via inactivating the STAT3 and PI3K/AKT signaling pathways”
- Special Issue Advancements in oncology: bridging clinical and experimental research - Part II
- Unveiling novel biomarkers for platinum chemoresistance in ovarian cancer
- Lathyrol affects the expression of AR and PSA and inhibits the malignant behavior of RCC cells
- The era of increasing cancer survivorship: Trends in fertility preservation, medico-legal implications, and ethical challenges
- Bone scintigraphy and positron emission tomography in the early diagnosis of MRONJ
- Meta-analysis of clinical efficacy and safety of immunotherapy combined with chemotherapy in non-small cell lung cancer
- Special Issue Computational Intelligence Methodologies Meets Recurrent Cancers - Part IV
- Exploration of mRNA-modifying METTL3 oncogene as momentous prognostic biomarker responsible for colorectal cancer development
- Special Issue The evolving saga of RNAs from bench to bedside - Part III
- Interaction and verification of ferroptosis-related RNAs Rela and Stat3 in promoting sepsis-associated acute kidney injury
- The mRNA MOXD1: Link to oxidative stress and prognostic significance in gastric cancer
- Special Issue Exploring the biological mechanism of human diseases based on MultiOmics Technology - Part II
- Dynamic changes in lactate-related genes in microglia and their role in immune cell interactions after ischemic stroke
- A prognostic model correlated with fatty acid metabolism in Ewing’s sarcoma based on bioinformatics analysis
- Red cell distribution width predicts early kidney injury: A NHANES cross-sectional study
- Special Issue Diabetes mellitus: pathophysiology, complications & treatment
- Nutritional risk assessment and nutritional support in children with congenital diabetes during surgery
- Correlation of the differential expressions of RANK, RANKL, and OPG with obesity in the elderly population in Xinjiang
- A discussion on the application of fluorescence micro-optical sectioning tomography in the research of cognitive dysfunction in diabetes
- A review of brain research on T2DM-related cognitive dysfunction
- Metformin and estrogen modulation in LABC with T2DM: A 36-month randomized trial
- Special Issue Innovative Biomarker Discovery and Precision Medicine in Cancer Diagnostics
- CircASH1L-mediated tumor progression in triple-negative breast cancer: PI3K/AKT pathway mechanisms
Articles in the same Issue
- Research Articles
- Network pharmacological analysis and in vitro testing of the rutin effects on triple-negative breast cancer
- Impact of diabetes on long-term survival in elderly liver cancer patients: A retrospective study
- Knockdown of CCNB1 alleviates high glucose-triggered trophoblast dysfunction during gestational diabetes via Wnt/β-catenin signaling pathway
- Risk factors for severe adverse drug reactions in hospitalized patients
- Analysis of the effect of ALA-PDT on macrophages in footpad model of mice infected with Fonsecaea monophora based on single-cell sequencing
- Development and validation of headspace gas chromatography with a flame ionization detector method for the determination of ethanol in the vitreous humor
- CMSP exerts anti-tumor effects on small cell lung cancer cells by inducing mitochondrial dysfunction and ferroptosis
- Predictive value of plasma sB7-H3 and YKL-40 in pediatric refractory Mycoplasma pneumoniae pneumonia
- Antiangiogenic potential of Elaeagnus umbellata extracts and molecular docking study by targeting VEGFR-2 pathway
- Comparison of the effectiveness of nurse-led preoperative counseling and postoperative follow-up care vs standard care for patients with gastric cancer
- Comparing the therapeutic efficacy of endoscopic minimally invasive surgery and traditional surgery for early-stage breast cancer: A meta-analysis
- Adhered macrophages as an additional marker of cardiomyocyte injury in biopsies of patients with dilated cardiomyopathy
- Association between statin administration and outcome in patients with sepsis: A retrospective study
- Exploration of the association between estimated glucose disposal rate and osteoarthritis in middle-aged and older adults: An analysis of NHANES data from 2011 to 2018
- A comparative analysis of the binary and multiclass classified chest X-ray images of pneumonia and COVID-19 with ML and DL models
- Lysophosphatidic acid 2 alleviates deep vein thrombosis via protective endothelial barrier function
- Transcription factor A, mitochondrial promotes lymph node metastasis and lymphangiogenesis in epithelial ovarian carcinoma
- Serum PM20D1 levels are associated with nutritional status and inflammatory factors in gastric cancer patients undergoing early enteral nutrition
- Hydromorphone reduced the incidence of emergence agitation after adenotonsillectomy in children with obstructive sleep apnea: A randomized, double-blind study
- Vitamin D replacement therapy may regulate sleep habits in patients with restless leg syndrome
- The first-line antihypertensive nitrendipine potentiated the therapeutic effect of oxaliplatin by downregulating CACNA1D in colorectal cancer
- Health literacy and health-related quality of life: The mediating role of irrational happiness
- Modulatory effects of Lycium barbarum polysaccharide on bone cell dynamics in osteoporosis
- Mechanism research on inhibition of gastric cancer in vitro by the extract of Pinellia ternata based on network pharmacology and cellular metabolomics
- Examination of the causal role of immune cells in non-alcoholic fatty liver disease by a bidirectional Mendelian randomization study
- Clinical analysis of ten cases of HIV infection combined with acute leukemia
- Investigating the cardioprotective potential of quercetin against tacrolimus-induced cardiotoxicity in Wistar rats: A mechanistic insights
- Clinical observation of probiotics combined with mesalazine and Yiyi Baitouweng Decoction retention enema in treating mild-to-moderate ulcerative colitis
- Diagnostic value of ratio of blood inflammation to coagulation markers in periprosthetic joint infection
- Sex-specific associations of sex hormone binding globulin and risk of bladder cancer
- Core muscle strength and stability-oriented breathing training reduces inter-recti distance in postpartum women
- The ERAS nursing care strategy for patients undergoing transsphenoidal endoscopic pituitary tumor resection: A randomized blinded controlled trial
- The serum IL-17A levels in patients with traumatic bowel rupture post-surgery and its predictive value for patient prognosis
- Impact of Kolb’s experiential learning theory-based nursing on caregiver burden and psychological state of caregivers of dementia patients
- Analysis of serum NLR combined with intraoperative margin condition to predict the prognosis of cervical HSIL patients undergoing LEEP surgery
- Commiphora gileadensis ameliorate infertility and erectile dysfunction in diabetic male mice
- The correlation between epithelial–mesenchymal transition classification and MMP2 expression of circulating tumor cells and prognosis of advanced or metastatic nasopharyngeal carcinoma
- Tetrahydropalmatine improves mitochondrial function in vascular smooth muscle cells of atherosclerosis in vitro by inhibiting Ras homolog gene family A/Rho-associated protein kinase-1 signaling pathway
- A cross-sectional study: Relationship between serum oxidative stress levels and arteriovenous fistula maturation in maintenance dialysis patients
- A comparative analysis of the impact of repeated administration of flavan 3-ol on brown, subcutaneous, and visceral adipose tissue
- Identifying early screening factors for depression in middle-aged and older adults: A cohort study
- Perform tumor-specific survival analysis for Merkel cell carcinoma patients undergoing surgical resection based on the SEER database by constructing a nomogram chart
- Unveiling the role of CXCL10 in pancreatic cancer progression: A novel prognostic indicator
- High-dose preoperative intraperitoneal erythropoietin and intravenous methylprednisolone in acute traumatic spinal cord injuries following decompression surgeries
- RAB39B: A novel biomarker for acute myeloid leukemia identified via multi-omics and functional validation
- Impact of peripheral conditioning on reperfusion injury following primary percutaneous coronary intervention in diabetic and non-diabetic STEMI patients
- Clinical efficacy of azacitidine in the treatment of middle- and high-risk myelodysplastic syndrome in middle-aged and elderly patients: A retrospective study
- The effect of ambulatory blood pressure load on mitral regurgitation in continuous ambulatory peritoneal dialysis patients
- Expression and clinical significance of ITGA3 in breast cancer
- Single-nucleus RNA sequencing reveals ARHGAP28 expression of podocytes as a biomarker in human diabetic nephropathy
- rSIG combined with NLR in the prognostic assessment of patients with multiple injuries
- Toxic metals and metalloids in collagen supplements of fish and jellyfish origin: Risk assessment for daily intake
- Exploring causal relationship between 41 inflammatory cytokines and marginal zone lymphoma: A bidirectional Mendelian randomization study
- Gender beliefs and legitimization of dating violence in adolescents
- Effect of serum IL-6, CRP, and MMP-9 levels on the efficacy of modified preperitoneal Kugel repair in patients with inguinal hernia
- Effect of smoking and smoking cessation on hematological parameters in polycythemic patients
- Pathogen surveillance and risk factors for pulmonary infection in patients with lung cancer: A retrospective single-center study
- Necroptosis of hippocampal neurons in paclitaxel chemotherapy-induced cognitive impairment mediates microglial activation via TLR4/MyD88 signaling pathway
- Celastrol suppresses neovascularization in rat aortic vascular endothelial cells stimulated by inflammatory tenocytes via modulating the NLRP3 pathway
- Cord-lamina angle and foraminal diameter as key predictors of C5 palsy after anterior cervical decompression and fusion surgery
- GATA1: A key biomarker for predicting the prognosis of patients with diffuse large B-cell lymphoma
- Influencing factors of false lumen thrombosis in type B aortic dissection: A single-center retrospective study
- MZB1 regulates the immune microenvironment and inhibits ovarian cancer cell migration
- Integrating experimental and network pharmacology to explore the pharmacological mechanisms of Dioscin against glioblastoma
- Trends in research on preterm birth in twin pregnancy based on bibliometrics
- Four-week IgE/baseline IgE ratio combined with tryptase predicts clinical outcome in omalizumab-treated children with moderate-to-severe asthma
- Single-cell transcriptomic analysis identifies a stress response Schwann cell subtype
- Acute pancreatitis risk in the diagnosis and management of inflammatory bowel disease: A critical focus
- Effect of subclinical esketamine on NLRP3 and cognitive dysfunction in elderly ischemic stroke patients
- Interleukin-37 mediates the anti-oral tumor activity in oral cancer through STAT3
- CA199 and CEA expression levels, and minimally invasive postoperative prognosis analysis in esophageal squamous carcinoma patients
- Efficacy of a novel drainage catheter in the treatment of CSF leak after posterior spine surgery: A retrospective cohort study
- Comprehensive biomedicine assessment of Apteranthes tuberculata extracts: Phytochemical analysis and multifaceted pharmacological evaluation in animal models
- Relation of time in range to severity of coronary artery disease in patients with type 2 diabetes: A cross-sectional study
- Dopamine attenuates ethanol-induced neuronal apoptosis by stimulating electrical activity in the developing rat retina
- Correlation between albumin levels during the third trimester and the risk of postpartum levator ani muscle rupture
- Factors associated with maternal attention and distraction during breastfeeding and childcare: A cross-sectional study in the west of Iran
- Mechanisms of hesperetin in treating metabolic dysfunction-associated steatosis liver disease via network pharmacology and in vitro experiments
- The law on oncological oblivion in the Italian and European context: How to best uphold the cancer patients’ rights to privacy and self-determination?
- The prognostic value of the neutrophil-to-lymphocyte ratio, platelet-to-lymphocyte ratio, and prognostic nutritional index for survival in patients with colorectal cancer
- Factors affecting the measurements of peripheral oxygen saturation values in healthy young adults
- Comparison and correlations between findings of hysteroscopy and vaginal color Doppler ultrasonography for detection of uterine abnormalities in patients with recurrent implantation failure
- The effects of different types of RAGT on balance function in stroke patients with low levels of independent walking in a convalescent rehabilitation hospital
- Causal relationship between asthma and ankylosing spondylitis: A bidirectional two-sample univariable and multivariable Mendelian randomization study
- Correlations of health literacy with individuals’ understanding and use of medications in Southern Taiwan
- Correlation of serum calprotectin with outcome of acute cerebral infarction
- Comparison of computed tomography and guided bronchoscopy in the diagnosis of pulmonary nodules: A systematic review and meta-analysis
- Curdione protects vascular endothelial cells and atherosclerosis via the regulation of DNMT1-mediated ERBB4 promoter methylation
- The identification of novel missense variant in ChAT gene in a patient with gestational diabetes denotes plausible genetic association
- Molecular genotyping of multi-system rare blood types in foreign blood donors based on DNA sequencing and its clinical significance
- Exploring the role of succinyl carnitine in the association between CD39⁺ CD4⁺ T cell and ulcerative colitis: A Mendelian randomization study
- Dexmedetomidine suppresses microglial activation in postoperative cognitive dysfunction via the mmu-miRNA-125/TRAF6 signaling axis
- Analysis of serum metabolomics in patients with different types of chronic heart failure
- Diagnostic value of hematological parameters in the early diagnosis of acute cholecystitis
- Pachymaran alleviates fat accumulation, hepatocyte degeneration, and injury in mice with nonalcoholic fatty liver disease
- Decrease in CD4 and CD8 lymphocytes are predictors of severe clinical picture and unfavorable outcome of the disease in patients with COVID-19
- METTL3 blocked the progression of diabetic retinopathy through m6A-modified SOX2
- The predictive significance of anti-RO-52 antibody in patients with interstitial pneumonia after treatment of malignant tumors
- Exploring cerebrospinal fluid metabolites, cognitive function, and brain atrophy: Insights from Mendelian randomization
- Development and validation of potential molecular subtypes and signatures of ocular sarcoidosis based on autophagy-related gene analysis
- Widespread venous thrombosis: Unveiling a complex case of Behçet’s disease with a literature perspective
- Uterine fibroid embolization: An analysis of clinical outcomes and impact on patients’ quality of life
- Discovery of lipid metabolism-related diagnostic biomarkers and construction of diagnostic model in steroid-induced osteonecrosis of femoral head
- Serum-derived exomiR-188-3p is a promising novel biomarker for early-stage ovarian cancer
- Enhancing chronic back pain management: A comparative study of ultrasound–MRI fusion guidance for paravertebral nerve block
- Peptide CCAT1-70aa promotes hepatocellular carcinoma proliferation and invasion via the MAPK/ERK pathway
- Electroacupuncture-induced reduction of myocardial ischemia–reperfusion injury via FTO-dependent m6A methylation modulation
- Hemorrhoids and cardiovascular disease: A bidirectional Mendelian randomization study
- Cell-free adipose extract inhibits hypertrophic scar formation through collagen remodeling and antiangiogenesis
- HALP score in Demodex blepharitis: A case–control study
- Assessment of SOX2 performance as a marker for circulating cancer stem-like cells (CCSCs) identification in advanced breast cancer patients using CytoTrack system
- Risk and prognosis for brain metastasis in primary metastatic cervical cancer patients: A population-based study
- Comparison of the two intestinal anastomosis methods in pediatric patients
- Factors influencing hematological toxicity and adverse effects of perioperative hyperthermic intraperitoneal vs intraperitoneal chemotherapy in gastrointestinal cancer
- Endotoxin tolerance inhibits NLRP3 inflammasome activation in macrophages of septic mice by restoring autophagic flux through TRIM26
- Lateral transperitoneal laparoscopic adrenalectomy: A single-centre experience of 21 procedures
- Petunidin attenuates lipopolysaccharide-induced retinal microglia inflammatory response in diabetic retinopathy by targeting OGT/NF-κB/LCN2 axis
- Procalcitonin and C-reactive protein as biomarkers for diagnosing and assessing the severity of acute cholecystitis
- Factors determining the number of sessions in successful extracorporeal shock wave lithotripsy patients
- Development of a nomogram for predicting cancer-specific survival in patients with renal pelvic cancer following surgery
- Inhibition of ATG7 promotes orthodontic tooth movement by regulating the RANKL/OPG ratio under compression force
- A machine learning-based prognostic model integrating mRNA stemness index, hypoxia, and glycolysis‑related biomarkers for colorectal cancer
- Glutathione attenuates sepsis-associated encephalopathy via dual modulation of NF-κB and PKA/CREB pathways
- FAHD1 prevents neuronal ferroptosis by modulating R-loop and the cGAS–STING pathway
- Association of placenta weight and morphology with term low birth weight: A case–control study
- Investigation of the pathogenic variants induced Sjogren’s syndrome in Turkish population
- Nucleotide metabolic abnormalities in post-COVID-19 condition and type 2 diabetes mellitus patients and their association with endocrine dysfunction
- TGF-β–Smad2/3 signaling in high-altitude pulmonary hypertension in rats: Role and mechanisms via macrophage M2 polarization
- Ultrasound-guided unilateral versus bilateral erector spinae plane block for postoperative analgesia of patients undergoing laparoscopic cholecystectomy
- Profiling gut microbiome dynamics in subacute thyroiditis: Implications for pathogenesis, diagnosis, and treatment
- Delta neutrophil index, CRP/albumin ratio, procalcitonin, immature granulocytes, and HALP score in acute appendicitis: Best performing biomarker?
- Anticancer activity mechanism of novelly synthesized and characterized benzofuran ring-linked 3-nitrophenyl chalcone derivative on colon cancer cells
- H2valdien3 arrests the cell cycle and induces apoptosis of gastric cancer
- Prognostic relevance of PRSS2 and its immune correlates in papillary thyroid carcinoma
- Association of SGLT2 inhibition with psychiatric disorders: A Mendelian randomization study
- Motivational interviewing for alcohol use reduction in Thai patients
- Luteolin alleviates oxygen-glucose deprivation/reoxygenation-induced neuron injury by regulating NLRP3/IL-1β signaling
- Polyphyllin II inhibits thyroid cancer cell growth by simultaneously inhibiting glycolysis and oxidative phosphorylation
- Relationship between the expression of copper death promoting factor SLC31A1 in papillary thyroid carcinoma and clinicopathological indicators and prognosis
- CSF2 polarized neutrophils and invaded renal cancer cells in vitro influence
- Proton pump inhibitors-induced thrombocytopenia: A systematic literature analysis of case reports
- The current status and influence factors of research ability among community nurses: A sequential qualitative–quantitative study
- OKAIN: A comprehensive oncology knowledge base for the interpretation of clinically actionable alterations
- The relationship between serum CA50, CA242, and SAA levels and clinical pathological characteristics and prognosis in patients with pancreatic cancer
- Identification and external validation of a prognostic signature based on hypoxia–glycolysis-related genes for kidney renal clear cell carcinoma
- Engineered RBC-derived nanovesicles functionalized with tumor-targeting ligands: A comparative study on breast cancer targeting efficiency and biocompatibility
- Relationship of resting echocardiography combined with serum micronutrients to the severity of low-gradient severe aortic stenosis
- Effect of vibration on pain during subcutaneous heparin injection: A randomized, single-blind, placebo-controlled trial
- The diagnostic performance of machine learning-based FFRCT for coronary artery disease: A meta-analysis
- Comparing biofeedback device vs diaphragmatic breathing for bloating relief: A randomized controlled trial
- Serum uric acid to albumin ratio and C-reactive protein as predictive biomarkers for chronic total occlusion and coronary collateral circulation quality
- Multiple organ scoring systems for predicting in-hospital mortality of sepsis patients in the intensive care unit
- Single-cell RNA sequencing data analysis of the inner ear in gentamicin-treated mice via intraperitoneal injection
- Review Articles
- The effects of enhanced external counter-pulsation on post-acute sequelae of COVID-19: A narrative review
- Diabetes-related cognitive impairment: Mechanisms, symptoms, and treatments
- Microscopic changes and gross morphology of placenta in women affected by gestational diabetes mellitus in dietary treatment: A systematic review
- Review of mechanisms and frontier applications in IL-17A-induced hypertension
- Research progress on the correlation between islet amyloid peptides and type 2 diabetes mellitus
- The safety and efficacy of BCG combined with mitomycin C compared with BCG monotherapy in patients with non-muscle-invasive bladder cancer: A systematic review and meta-analysis
- The application of augmented reality in robotic general surgery: A mini-review
- The effect of Greek mountain tea extract and wheat germ extract on peripheral blood flow and eicosanoid metabolism in mammals
- Neurogasobiology of migraine: Carbon monoxide, hydrogen sulfide, and nitric oxide as emerging pathophysiological trinacrium relevant to nociception regulation
- Plant polyphenols, terpenes, and terpenoids in oral health
- Laboratory medicine between technological innovation, rights safeguarding, and patient safety: A bioethical perspective
- End-of-life in cancer patients: Medicolegal implications and ethical challenges in Europe
- The maternal factors during pregnancy for intrauterine growth retardation: An umbrella review
- Intra-abdominal hypertension/abdominal compartment syndrome of pediatric patients in critical care settings
- PI3K/Akt pathway and neuroinflammation in sepsis-associated encephalopathy
- Screening of Group B Streptococcus in pregnancy: A systematic review for the laboratory detection
- Giant borderline ovarian tumours – review of the literature
- Leveraging artificial intelligence for collaborative care planning: Innovations and impacts in shared decision-making – A systematic review
- Cholera epidemiology analysis through the experience of the 1973 Naples epidemic
- Risk factors of frailty/sarcopenia in community older adults: Meta-analysis
- Supplement strategies for infertility in overweight women: Evidence and legal insights
- Scurvy, a not obsolete disorder: Clinical report in eight young children and literature review
- A meta-analysis of the effects of DBS on cognitive function in patients with advanced PD
- Protective role of selenium in sepsis: Mechanisms and potential therapeutic strategies
- Strategies for hyperkalemia management in dialysis patients: A systematic review
- C-reactive protein-to-albumin ratio in peripheral artery disease
- Case Reports
- Delayed graft function after renal transplantation
- Semaglutide treatment for type 2 diabetes in a patient with chronic myeloid leukemia: A case report and review of the literature
- Diverse electrophysiological demyelinating features in a late-onset glycogen storage disease type IIIa case
- Giant right atrial hemangioma presenting with ascites: A case report
- Laser excision of a large granular cell tumor of the vocal cord with subglottic extension: A case report
- EsoFLIP-assisted dilation for dysphagia in systemic sclerosis: Highlighting the role of multimodal esophageal evaluation
- Molecular hydrogen-rhodiola as an adjuvant therapy for ischemic stroke in internal carotid artery occlusion: A case report
- Coronary artery anomalies: A case of the “malignant” left coronary artery and its surgical management
- Rapid Communication
- Biological properties of valve materials using RGD and EC
-
A single oral administration of flavanols enhances short
-term memory in mice along with increased brain-derived neurotrophic factor - Letter to the Editor
- Role of enhanced external counterpulsation in long COVID
- Expression of Concern
- Expression of concern “A ceRNA network mediated by LINC00475 in papillary thyroid carcinoma”
- Expression of concern “Notoginsenoside R1 alleviates spinal cord injury through the miR-301a/KLF7 axis to activate Wnt/β-catenin pathway”
- Expression of concern “circ_0020123 promotes cell proliferation and migration in lung adenocarcinoma via PDZD8”
- Corrigendum
- Corrigendum to “Empagliflozin improves aortic injury in obese mice by regulating fatty acid metabolism”
- Corrigendum to “Comparing the therapeutic efficacy of endoscopic minimally invasive surgery and traditional surgery for early-stage breast cancer: A meta-analysis”
- Corrigendum to “The progress of autoimmune hepatitis research and future challenges”
- Retraction
- Retraction of “miR-654-5p promotes gastric cancer progression via the GPRIN1/NF-κB pathway”
- Retraction of: “LncRNA CASC15 inhibition relieves renal fibrosis in diabetic nephropathy through downregulating SP-A by sponging to miR-424”
- Retraction of: “SCARA5 inhibits oral squamous cell carcinoma via inactivating the STAT3 and PI3K/AKT signaling pathways”
- Special Issue Advancements in oncology: bridging clinical and experimental research - Part II
- Unveiling novel biomarkers for platinum chemoresistance in ovarian cancer
- Lathyrol affects the expression of AR and PSA and inhibits the malignant behavior of RCC cells
- The era of increasing cancer survivorship: Trends in fertility preservation, medico-legal implications, and ethical challenges
- Bone scintigraphy and positron emission tomography in the early diagnosis of MRONJ
- Meta-analysis of clinical efficacy and safety of immunotherapy combined with chemotherapy in non-small cell lung cancer
- Special Issue Computational Intelligence Methodologies Meets Recurrent Cancers - Part IV
- Exploration of mRNA-modifying METTL3 oncogene as momentous prognostic biomarker responsible for colorectal cancer development
- Special Issue The evolving saga of RNAs from bench to bedside - Part III
- Interaction and verification of ferroptosis-related RNAs Rela and Stat3 in promoting sepsis-associated acute kidney injury
- The mRNA MOXD1: Link to oxidative stress and prognostic significance in gastric cancer
- Special Issue Exploring the biological mechanism of human diseases based on MultiOmics Technology - Part II
- Dynamic changes in lactate-related genes in microglia and their role in immune cell interactions after ischemic stroke
- A prognostic model correlated with fatty acid metabolism in Ewing’s sarcoma based on bioinformatics analysis
- Red cell distribution width predicts early kidney injury: A NHANES cross-sectional study
- Special Issue Diabetes mellitus: pathophysiology, complications & treatment
- Nutritional risk assessment and nutritional support in children with congenital diabetes during surgery
- Correlation of the differential expressions of RANK, RANKL, and OPG with obesity in the elderly population in Xinjiang
- A discussion on the application of fluorescence micro-optical sectioning tomography in the research of cognitive dysfunction in diabetes
- A review of brain research on T2DM-related cognitive dysfunction
- Metformin and estrogen modulation in LABC with T2DM: A 36-month randomized trial
- Special Issue Innovative Biomarker Discovery and Precision Medicine in Cancer Diagnostics
- CircASH1L-mediated tumor progression in triple-negative breast cancer: PI3K/AKT pathway mechanisms