METTL3 attenuates ferroptosis sensitivity in lung cancer via modulating TFRC
-
Peng Zhang


Abstracts
Overexpression of methyltransferase-like 3 (METTL3) is significantly correlated with the malignancy of lung cancer (LC). In the present study, we demonstrated that METTL3 had higher levels in LC tissues relative to normal tissues. METTL3 showed superior sensitivity and specificity for diagnosis and identification of LC functions. In addition, silencing METTL3 resulted in enhanced ferroptosis sensitivity, whereas overexpression of METTL3 exhibited the opposite effect. Inhibition of METTL3 impeded LC growth in cell-derived xenografts. Further exploratory studies found that METTL3 stimulated the low expression of transferrin receptor (TFRC), which was critical for ferroptosis sensitization in LC cells induced by silenced METTL3, as silencing of TFRC caused a decrease in negative regulators of ferroptosis (FTH1 and FTL) in METTL3 knockdown A549 and PC9 cells. Finally, we confirmed that METTL3 attenuation effectively maintained the stability of TFRC mRNA. In conclusion, we reported a novel mechanism of METTL3 desensitization to ferroptosis via regulating TFRC, and an appropriate reduction of METTL3 might sensitize cancer cells to ferroptosis-based therapy.
1 Introduction
Lung cancer (LC) accounts for 11.4% of malignancies and has leapt to the top of the list of cancer “super killers.” In particular, the prevalence of LC varies considerably by country and gender [1,2]. Men are more likely to develop LC than women, and the incidence and mortality ratios are 2:1 for both sexes. The occurrence of LC is age-related, mostly in the middle-aged and elderly population (45–65 years old), but in recent years there has been a trend toward younger age [3]. Indeed, LC exhibits rapid proliferation, metastasis, and intertumoral heterogeneity, resulting in poor patient survival due to frequent tumor recurrence and metastasis, discouraging overall survival [4]. Therefore, a more accurate understanding of the molecular mechanisms of LC, identification of new diagnostic molecular markers and new therapeutic modalities could help to detect LC at an early stage and develop appropriate treatment options for tumor progression.
Multicellular organisms possess the ability to remove damaged or redundant cells to maintain normal development of the organism and prevent the development of hyperproliferative diseases [5]. Thus, regulated cell death (RCD), such as apoptosis and necrosis, occurs in normal physiological programs [6]. However, either excessive or failing RCD can be a causative agent of cancer, especially in the lung [7]. In contrast to the known forms of cell death, ferroptosis was thought to be a new form of iron-dependent RCD, whose key feature is the lethal accumulation of iron-dependent reactive oxygen species (ROS) and lipid peroxides [8]. Several known molecules (e.g., glutathione peroxidase 4, p53, heat shock protein β-1, and nuclear factor E2-related factor 2) target iron metabolism or lipid peroxidation by specific mechanisms to alter ferroptosis sensitivity [9]. Furthermore, enhanced intracellular iron load is required to maintain ferroptosis [10]. More direct evidence came from the fact that suppression of the iron metabolism transcription factor IREB2 increased the expression of iron metabolism-related proteins (FTH1 and FTL), and that raised iron storage capacity effectively aggravated ferroptosis resistance [11]. In recent years, another new key player in the ferroptosis pathway, methyltransferase 3 (METTL3), has emerged. In-depth studies have revealed that METTL3 strengthened the expression of SLC7A11, which is a crucial negative regulator of ferroptosis, by means of its N6-methyladenosine (m6A) activity in hepatoblastoma and LC. Thus, in turn, it led to enhanced resistance of tumor cells to ferroptosis and induced malignant progression of cancer [12,13]. The hindrance of ferroptosis by METTL3 appeared to be dependent on the m6A pathway, both in tumors and in other diseases, such as brain hemorrhage [14] and diabetes mellitus [15]. However, regulatory mechanism of METTL3 on ferroptosis is complex and multifactorial and still requires in-depth investigation.
In this study, we reported heightened expression of METTL3, a common tumorigenic event in LC. Besides, we found an important role of METTL3 in mediating cellular ferroptosis sensitivity, i.e., ablation of METTL3 caused greatly enhanced LC cellular ferroptosis sensitivity. Also, we identified the negative regulation of transferrin receptor (TFRC) by METTL3 and the desensitization of cells to ferroptosis induced by TFRC attenuation. These results suggested that METTL3 combined with TFRC to induce ferroptosis could be used as a promising cancer therapy to address LC.
2 Materials and methods
2.1 LC tissue samples and cell cultures
A total of 40 pairs of tumor samples (LC specimens and paired adjacent normal tissue specimens) removed during surgery were obtained from Shanghai Tenth People’s Hospital, School of Clinical Medicine of Nanjing Medical University for this study. Patients with a history of anti-tumor therapy and those with other diseases were not included in this study. The protocol for the use and collection of human tissues was reviewed and approved by the Shanghai Tenth People’s Hospital, School of Clinical Medicine of Nanjing Medical University Ethics Committee. All participants were carefully informed and signed a written consent form before being subjected to the study. All experiments were performed in strict accordance with relevant guidelines and regulations and in compliance with the Declaration of Helsinki.
We obtained the normal human bronchial epithelial cell line NL20 and four human typical LC cell lines (A549, H1299, PC9, H1975, and HCC827) from the Cell Bank of the Chinese Academy of Sciences (Shanghai, China) for this study. All LC cell lines and NL20 cells were preserved in penicillin/streptomycin solution (1%, Gibco, USA) and 10% FBS (Sagecreation, China) enhanced with commercially available 1640 medium (Gibco, USA) according to the manufacturer’s requirements. All culture flasks were placed in humid air (culture environment: 37°C, 5% CO2) to maintain cell growth.
2.2 Plasmid construction and cell transfection
Specific small interfering RNAs (si-NC: CATTGACTAAGCCGATAGCCCGACTAGCT, si-TFRC 1#: GCGGTTCTTGGTACCAGCAACTTCA, si-TFRC 2#: CAGCCCACTGTTGTATACGCTTATT, si-METTL3 1#: GGAGGAGTGCATGAAAGCCAGTGAT, and si-METTL3 2#: AGCTGCACTTCAGACGAATTATCAA) targeting specific regions of METTL3 and TFRC were constructed and obtained by Obio Technology (Shanghai, China), and next, the sequences with the highest knock-down efficiency were selected for subsequent studies. Overexpression of intracellular METTL3 was achieved by ordering an overexpression plasmid (GV144 and oe-METTL3) from GeneChem (Shanghai, China), and an empty plasmid vector (oe-NC) served as a control. Transfection of METTL3 overexpressing plasmids and siRNAs targeting specific genes was achieved in A549 and PC9 cells with the aid of Lipofectamine 3000 (Invitrogen, USA) according to the manufacturer’s requirements.
2.3 In vitro cell viability assay
The viability values of two LC cell lines (A549 and PC9) after different treatments were determined using the Cell Counting Kit-8 (CCK-8, RiboBio, China) method. The prepared suspensions of multiple groups of target cells were sequentially injected into 96-well plates (5 × 103 cells/well). At the specified time points (0, 24, 48, 96 h), 10 μL of CCK-8 reaction reagent (ApexBio, USA) was added to the cell cultures to continue maintenance for 2 h (37°C, dark environment). Finally, the optical density (OD) values of each well were recorded at 450 nm using an automatic microplate reader (Tecan M200 PRO, Belgium).
2.4 4′-6-Diamidino-2-phenylindole (DAPI)/propidium iodide (PI) double staining
A549 and PC9 cells were treated with si-NC or si-METTL3 in 12-well plates (2 × 105 cells/well). To detect cell death, cells were stained with PI for 5 min in the dark and then restained with DAPI. Cells were washed three times in PBS to remove excess dye and then photographed with a fluorescence microscope (Olympus, BX53). PI staining was repeated in triplicate to obtain accurate results.
2.5 RNA stability assay
A549 cells and PC9 cells were treated with siRNA against METTL3. After 48 h, METTL3 knockdown or non-knockdown cells were mixed with actinomycin D (final concentration 5 μg/mL, APExBIO) to block the synthesis of new RNA in the cells. Next, RNA was extracted and collected at the indicated time points (1, 6, 12, 18, and 24 h) to perform RT-qPCR for stability assessment of TFRC mRNA.
2.6 Determination of intracellular lipid ROS, malondialdehyde (MDA), and glutathione (GSH) content
The levels of lipid ROS in the target cells were examined with the aid of a fluorescent probe C11-BODIPY 581/591 (5 μmmol, Thermo Fisher), which was sensitive to lipid peroxidation. A549 and PC9 cells were treated as required, followed by digesting (0.25% trypsin), washing, and resuspending cells in fresh medium supplemented with 10% FBS. Cells were stained with C11-BODIPY for 30 min under the prescribed conditions (protected from light, 37°C). Finally, the stained cells were collected for flow cytometry (BD Accuri C6) to collect the mean fluorescence intensity of lipid ROS-positive LC cells.
LC intracellular MDA levels were determined using a MDA assay kit (Abcam) according to the manufacturer’s standard protocol. Importantly, the complete MDA level was normalized to the total protein content of the sample and recorded as nmol/mg.
Total GSH content in tumor cells (50,000/group) was calculated by ordering a commercial GSH assay kit from Beyotime Biotechnology. The concentration of GSH was normalized according to the total protein content of the treated samples and denoted as nmol/mg.
2.7 Analysis of intracellular unstable iron content
Supernatants collected from LC cells of groups si-NC, oe-NC, si-METTL3, oe-METTL3, si-METTL3 + si-NC, and si-METTL3 + si-TFRC were analyzed for unstable iron (Fe2+) levels. Briefly, Iron Detection Kit was purchased from Abcam, and 5 µL of iron buffer was added to each sample well and incubated at 37°C for 30 min. Iron probe of 100 µL was continued to be added to the mixture awaiting detection, mixed well and maintained for 60 min (37°C, protected from light). Immediately after completion of the reaction, the OD at 593 nm was evaluated on a micro-plate reader.
2.8 Immunohistochemistry (IHC)
The strong and weak distribution of METTL3 expression in LC tumor tissues was identified by IHC. Tumor samples were fixed in tissue fixative (4% paraformaldehyde) and prepared into 4 μm paraffin sections with the addition of 3% H2O2 to block endogenous peroxidase activity. Antigen retrieval was done in 10 mM citrate solution. Sections were soaked with diluted primary antibody targeting METTL3 (1:100, Abcam, UK) in a humidified refrigerator at 4°C for 12 h. Sections were then rinsed with PBS and maintained for a further 60 min at 37°C with secondary antibody (Dako). Sections were developed with diaminobenzidine (DAB, Gene Tech) after re-rinsing with PBS, and the degree of METTL3 enrichment in the tumor tissue was determined by the intensity of staining.
2.9 M6A-RNA immunoprecipitation (Me-RIP) assay
A549 and PC9 cells transfected with si-NC or si-METTL3 were lysed, and the clarified lysates were subsequently incubated with appropriate amounts of the corresponding antibodies (anti-m6A, 1:500, Abcam, USA) as well as human IgG-conjugated magnetic beads in RIP buffer until overnight. The resulting magnetic bead–antibody complexes were resuspended and digested with proteinase K buffer to isolate and extract the immunoprecipitated target RNA. Purified RNA was quantified by RT-qPCR to verify the relative levels of TFRC.
2.10 RT-qPCR
Total RNA was isolated from LC tissue specimens, A549 and PC9 cells using RNA extraction reagent (TRIzol, Invitrogen, USA). The purity and concentration of the obtained total RNA were determined, and then the generation of cDNA templates was initiated using the HiFiScript cDNA synthesis kit (CWBIO, China). Next, PCR kits with specific primers (SYBR Green Master Mix, Applied Biosystems) and QuantStudio 3D system (Thermo Fisher, USA) were employed to complete the remaining RT-qPCR steps. The levels of both METTL3 and TFRC were fully normalized to β-actin. The relative expression levels of target genes in treated cells were calculated by the 2–∆∆Ct method. The primer sequences of the specific genes used were:
METTL3-F: 5′-AAGCTGCACTTCAGACGAAT-3′
METTL3-R: 5′-GGAATCACCTCCGACACTC-3′
TFRC-F: 5′-ACCTGTCCAGACAATCTCCAG-3′
TFRC-R: 5′-TGTTTTCCAGTCAGAGGGACA-3′
β-Actin-F: 5′-TGAGAGGGAAATCGTGCGTGAC-3′
β-Actin-R: 5′-AAGAAGGAAGGCTGGAAAAGAG-3′
2.11 Western blot
A549 and PC9 cells were harvested and lysed in lysis buffer (Beyotime) supplemented with a 1% protease inhibitor (Roche, USA) mixture and whole proteins were extracted. Protein concentrations were estimated using the BCA Protein Assay Kit (Pierce, USA). Equal amounts of denatured protein samples were resolved on 10–12% SDS-PAGE and transferred to PVDF (Millipore, USA) membranes. Membranes were sealed with 5% skim milk for 2 h and then incubated overnight with diluted primary antibodies specific for METTL3 (1:200, Abcam), TFRC (1:1,000, Abcam), FTH1 (1:500, ABclonal), FTL (1:250, Abcam), and β-actin (1:1,000, CST) at 4°C. The membranes were continued to be incubated with secondary antibodies for 1.5 h before visualization of protein blot images using the ECL kit (Millipore, USA).
2.12 Animal studies
Stable knockdown of METTL3 was achieved by transfection of A549 cells with shRNA specifically targeting METTL3. Approximately 5 × 106 A549 cells (0.1 mL) were injected subcutaneously into the axillae of male athymic BALB/C nude mice (4–6 weeks old, 18–22 g, 3 per group). The length (L) and width (W) of the tumors were measured weekly using calipers, and the tumor volume was calculated according to the formula V = (W 2 × L)/2. All mice were sacrificed 4 weeks later, and tumor tissues were isolated and examined for weight.
2.13 Statistical analysis
The experiments used were performed at least three times independently and the data obtained are reported as mean ± SD. Statistical differences between two or more consecutive data groups were compared using Student’s t-test and one-way analysis of variance. Pearson linear correlation analysis was applied to calculate the correlation between METTL3 and TFRC expression. Graphs were plotted by GraphPad Prism 8.0 software. P values less than 0.05 were determined to be statistically significant.
3 Results
3.1 LC is associated with up-regulation of METTL3
Considering the important role of METTL3 in controlling the growth, survival, and invasion of LC cells (A549, H1299) [16], we were motivated to explore additional mechanisms of METTL3 in LC. In the current study, we described and analyzed METTL3 expression levels in a cohort of LC patients and compared them with normal paracancer controls. Tissues from patients with untreated primary LC showed a strong up-regulation of METTL3 (Figure 1a). To further investigate the clinical significance of METTL3 in LC, we preprocessed the clinical data from archived LC patients and plotted ROC curves. As shown in Figure 1b, METTL3 presented sensitivity and specificity in distinguishing between healthy individuals and patients with an AUC of 0.9169 (P < 0.0001), indicating the strong detection ability of METTL3 in the early diagnosis of LC. In addition, evaluation of stored tissue samples by IHC staining showed a substantial increase in staining for METTL3 in lung tumors, which was consistent with previous observations of mRNA in tissues (Figure 1c). Indeed, relatively high levels of METTL3 were detected in five LC cell lines (A549, H1299, PC9, H1975, and HCC827). Among them, we noted that METTL3 was most significantly upregulated in A549 and PC9 cells compared to other types of LC cells, and we used these two cells as subjects for our next study (Figure 1d). In conclusion, these data suggested that METTL3 was indeed increased in LC and its presence might serve as a potential marker for early diagnosis in LC patients.
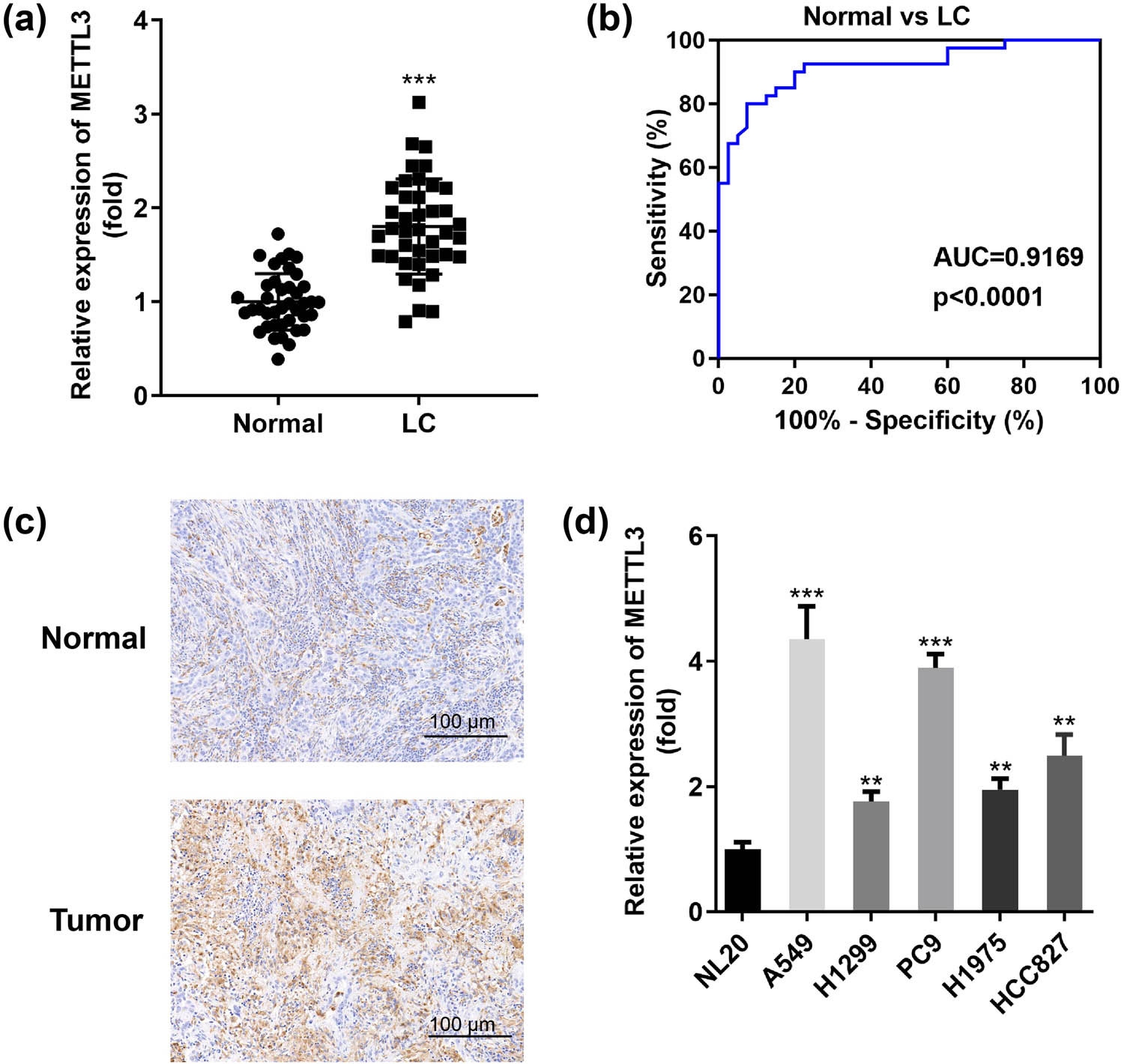
METTL3 exists in LC tumors in an overexpression pattern and is a promising biomarker for LC patients. (a) RT-qPCR assay was applied to show METTL3 expression in 40 pairs of collected clinical LC tumor tissues and paired normal tissues (Normal). (b) ROC curves were established to assess the potential diagnostic value of METTL3 based on its expression in tumor samples. (c) Typical IHC images of METTL3 staining in paired LC tumor tissues. Scale bar, 100 μm. (d) Differences in METTL3 expression levels between human bronchial epithelial cells (NL20) and representative LC cells (A549, H1299, PC9, H1975, and HCC827). **P < 0.01, ***P < 0.001.
3.2 METTL3 ablation activates ferroptosis in LC cells
To continue to systematically investigate the potential role of METTL3 in LC, we successfully constructed knockdown and overexpression of METTL3 at the cellular level by transfecting two human LC cell lines, A549 and PC9, with two independent siRNAs (named si-METTL3 1# and si-METTL3 2#) and METTL3 overexpression plasmids (named oe-METTL3), respectively (Figure 2a and b). As expected, ablation of METTL3 inhibited the cell viability of A549 and PC9, while enhanced METTL3 promoted LC cell growth in vitro (Figure 2c). Subsequently, we performed DAPI/PI double staining to evaluate cell death. DAPI stained nuclei (bright blue fluorescence) and PI stained dead cells (red fluorescence). The results showed that METTL3 knockdown caused an increased rate of PI-positive cells, suggesting increased cell death. In contrast, PI-positive cells were decreased by METTL3 overexpression (Figure 2d and e). The above data linked METTL3 to cell death. Ferroptosis was a newly identified form of cell death, and METTL3 has recently been found to cause altered ferroptosis sensitivity in LC cells [12]. Therefore, we sought to explore in A549 and PC9 cells whether METTL3 levels determine the degree of sensitivity of tumor cells to ferroptosis. Redox-active iron enrichment, GSH depletion, and lipid ROS accumulation were recognized as central elements of ferroptosis. Therefore, we monitored the levels of Fe2+, GSH, oxidative stress markers MDA, and lipid ROS in target cells. Our results presented that silencing METTL3 resulted in a decrease in GSH concentration and an increase in Fe2+ level, MDA content, and lipid ROS level in A549 and PC9 cells. Importantly, the opposite phenomenon was presented in A549 and PC9 cells overexpressing METTL3 (Figure 2f–i). Furthermore, we found that METTL3 silencing significantly hindered the expression of negative regulators of ferroptosis (FTH1 and FTL) in cells, and conversely, the levels of FTH1 and FTL proteins increased in METTL3 overexpressing cells (Figure 2j). The above convincing data strongly demonstrated that METTL3 assumed an important role in ferroptosis in LC cells, and that inhibition of METTL3 induced stronger ferroptosis sensitivity in LC cells.
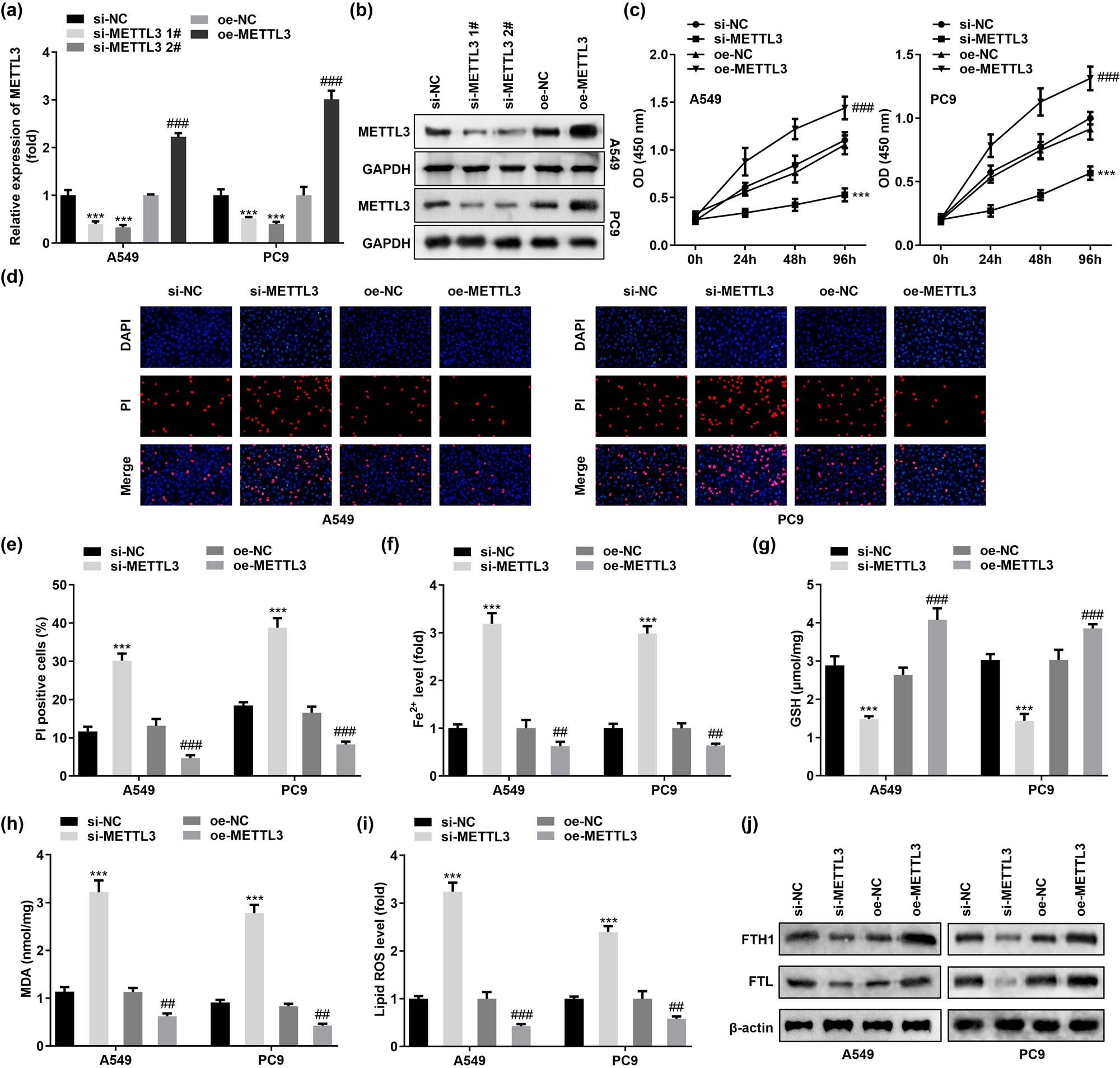
METTL3 silencing is critical for the induction of ferroptosis sensitivity in LC cells. (a and b) Knockdown or overexpression efficiency of METTL3 was verified by RT-qPCR and western blot after introduction of METTL3 siRNAs (si-METTL3 1# and si-METTL3 2#) and METTL3 overexpression plasmid (oe-METTL3) in A549 and PC9 cells, respectively. Among them, si-METTL3 2# showed stronger knockdown efficiency. (c) CCK-8 assay was conducted to obtain data on the change in cell viability after overexpression or knockdown of METTL3 in A549 and PC9 cells, respectively. (d and e) DAPI/PI staining was performed to detect cell death. (f–i) Determination of Fe2+, GSH, MDA, and lipid ROS in METTL3 knockdown or overexpression cells. (j) Expression of FTH1 and FTL proteins, the negative regulators of ferroptosis, was detected by western blot. ## P < 0.01, ### P < 0.001, ***P < 0.001.
Next, we performed subcutaneous xenograft experiments to assess the physiological relevance of METTL3 to LC cell growth in vivo. Consistent with the in vitro results, tumor growth was slowed and xenograft weight was reduced when implanted with METTL3 knockdown A549 cells relative to control (sh-NC) cells (Figure 3a–c). Collectively, these loss/gain-of-function experiments confirmed that METTL3 promoted LC cell growth and proliferation both in vivo and in vitro.
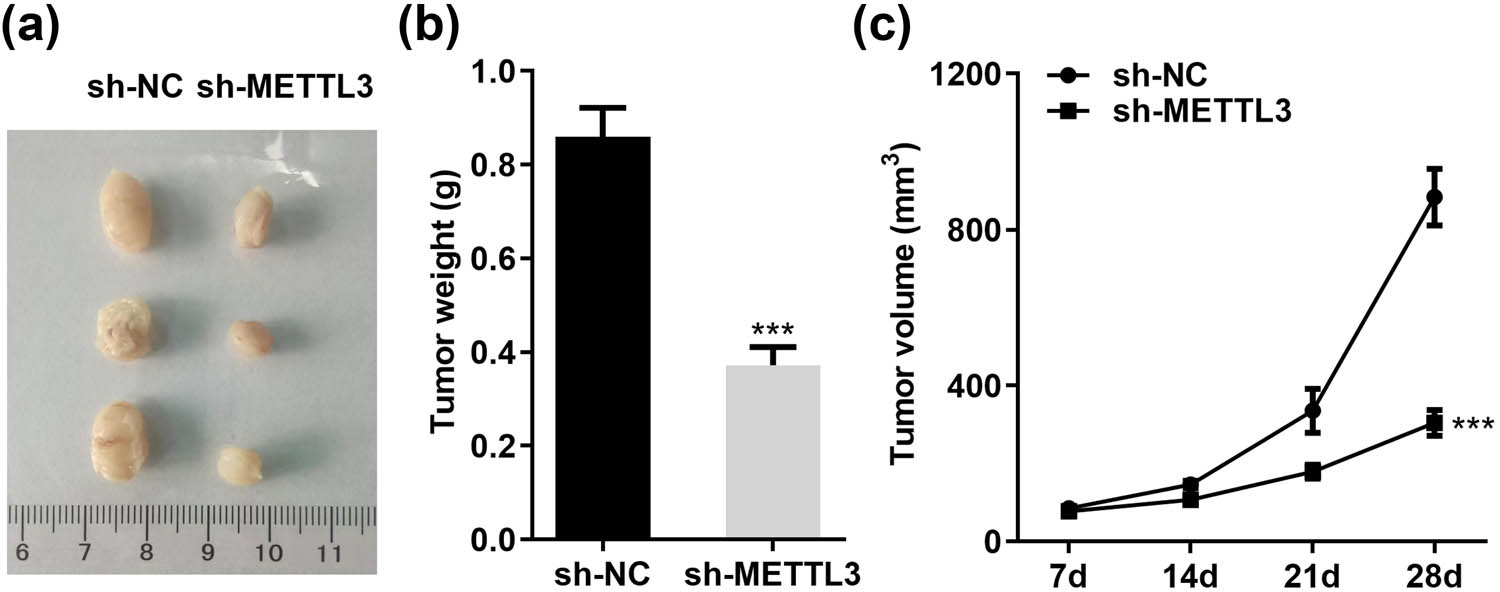
METTL3 accelerates the growth of LC cells in vivo. (a) Pictures of xenografts isolated from nude mice after sacrifice. (b and c) Tumor weights (b) and growth curves (c) of subcutaneous xenograft models injected with stably knockdown METTL3 (sh-METTL3) and negative control (sh-NC) A549 cells. ***P < 0.001.
3.3 METTL3 triggers TFRC low expression in LC cells
Ferroptosis is closely related to intracellular iron homeostasis, and TFRC is one of the essential molecules for increasing intracellular iron content as well as maintaining intracellular iron homeostasis [17]. Thus, we analyzed TFRC levels in LC specimens and normal paracancerous tissues from 40 patients. RT-qPCR results determined that TFRC was generally downregulated in LC tumor tissues (Figure 4a). The data from our cohort also showed the potential value of TFRC expression levels for the diagnosis of LC. As illustrated in Figure 4b, TFRC was sensitive and specific in distinguishing between LC patients and healthy controls with an AUC of 0.7631. Considering that TFRC was one of the important genes for intracellular iron uptake, we analyzed the correlation between the expression of TFRC and METTL3 in tissues based on the obtained data. Figure 4c indicates a significant negative correlation between TFRC and METTL3 in LC. Does the expression level of METTL3 actually determine the expression of TFRC in A549 and PC9 cells? To answer this question, we first analyzed TFRC expression in A549 and PC9 as well as NL20 cells. As expected, TFRC expression was remarkably lower in A549 and PC9 cells than in normal bronchial epithelial cells NL20 (Figure 4d). Furthermore, the expression of both TFRC mRNA and protein was enhanced in the absence of METTL3 in A549 and PC9 cells. In contrast, the introduction of METTL3 overexpression vector successfully decreased intracellular TFRC mRNA and protein levels (Figure 4e and f). Therefore, it was reasonable to speculate that METTL3 affected the sensitivity of LC cells to ferroptosis by reducing intracellular TFRC levels.
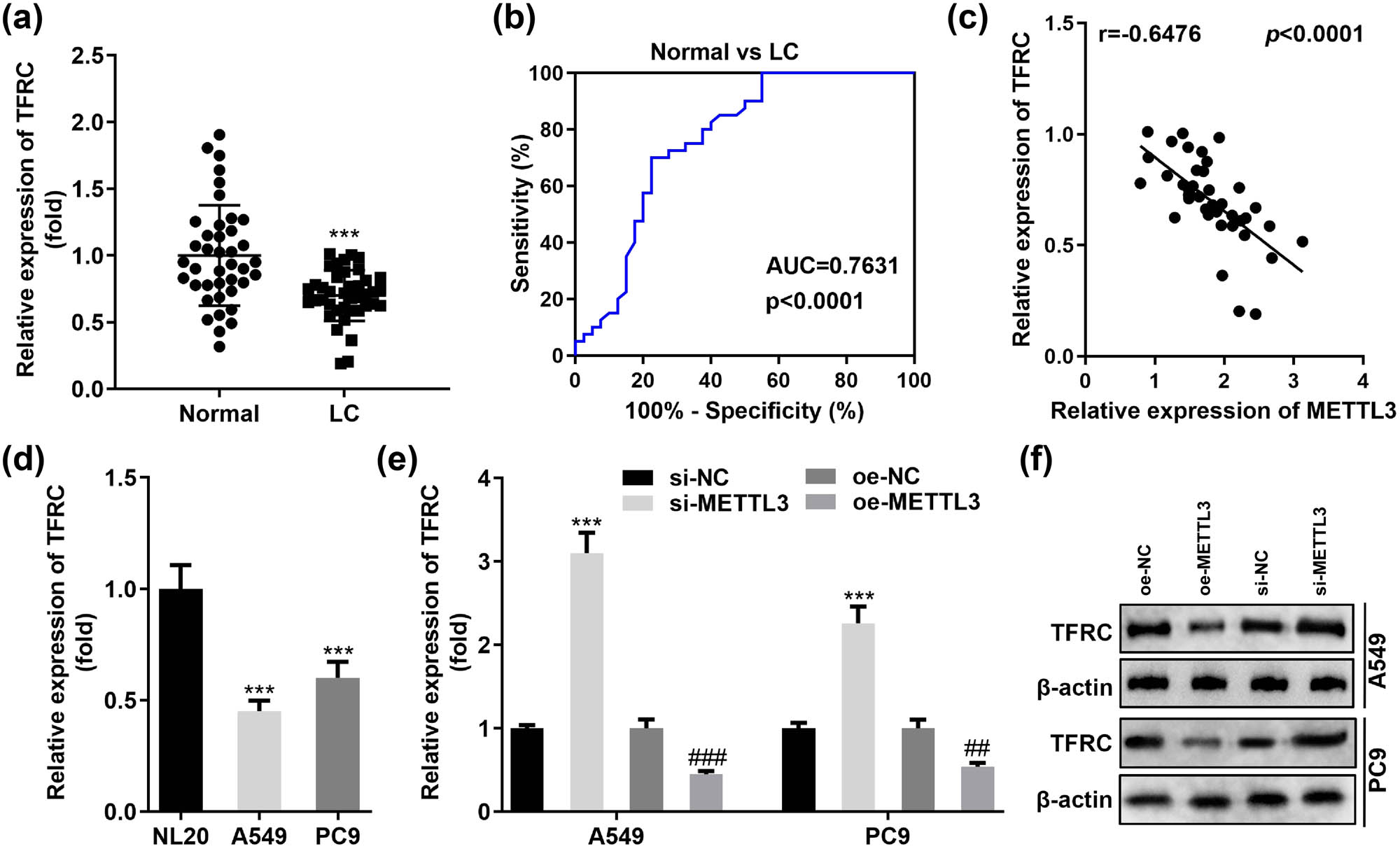
METTL3 negatively regulates TFRC levels in LC. (a) TFRC was significantly reduced in 40 primary LC tissues using normal tissues adjacent to the cancer as controls. (b) ROC curves were established to examine the practical value of TFRC in clinical practice. (c) Correlation between METTL3 and TFRC was analyzed based on RT-qPCR data. (d) Restricted TFRC levels were found in two representative LC cell lines (A549 and PC9). (e and f) Relative levels of TFRC mRNA and protein were detected by RT-qPCR and western blotting in A549 and PC9 cells transfected with si-METTL3 and oe-METTL3, respectively. ## P < 0.01, ### P < 0.001, ***P < 0.001.
3.4 TFRC is critical for ferroptosis sensitivity in METTL3-silenced LC cells
To confirm that METTL3-blocked TFRC expression inhibited intracellular iron uptake and thus caused ferroptosis desensitization in LC cells, we first designed two siRNAs (named si-TFRC 1# and si-TFRC 2#) to deplete TFRC in A549 and PC9 cells (Figure 5a) and verified their knockdown efficiency, respectively (Figure 5b). In addition, we added si-TFRC in METTL3-silenced A549 and PC9 cells and monitored the cellular activity. The results clearly demonstrated that the decrease in cellular activity or cell death caused by METTL3 attenuation could be rescued by si-TFRC treatment (Figure 5c). In addition, si-TFRC treatment also rescued A549 and PC9 cell death induced by METTL3 deletion (Figure 5d and e). Ferroptosis is the result of lethal accumulation of unstable iron in cells, and indeed, METTL3 silencing still increased intracellular Fe2+ levels even though TFRC was reduced in A549 and PC9 cells. Notably, ablation of TFRC weakened this effect (Figure 5f). Similarly, we also monitored the role of TFRC in regulating GSH concentration, MDA accumulation, and lipid ROS levels. The data showed that lower levels of TFRC caused an increase in GSH concentration, a decrease in MDA accumulation, and lipid ROS levels in A549 and PC9 cells after si-METTL3 treatment (Figure 5g–i). In addition, the opposite results of FTH1 and FTL induced by METTL3 silencing were also attenuated after knocking down TFRC in A549 and PC9 cells (Figure 5j). Combining the above data, we concluded that the induction of ferroptosis in LC cells by METTL3 silencing was dependent on the level of intracellular TFRC.
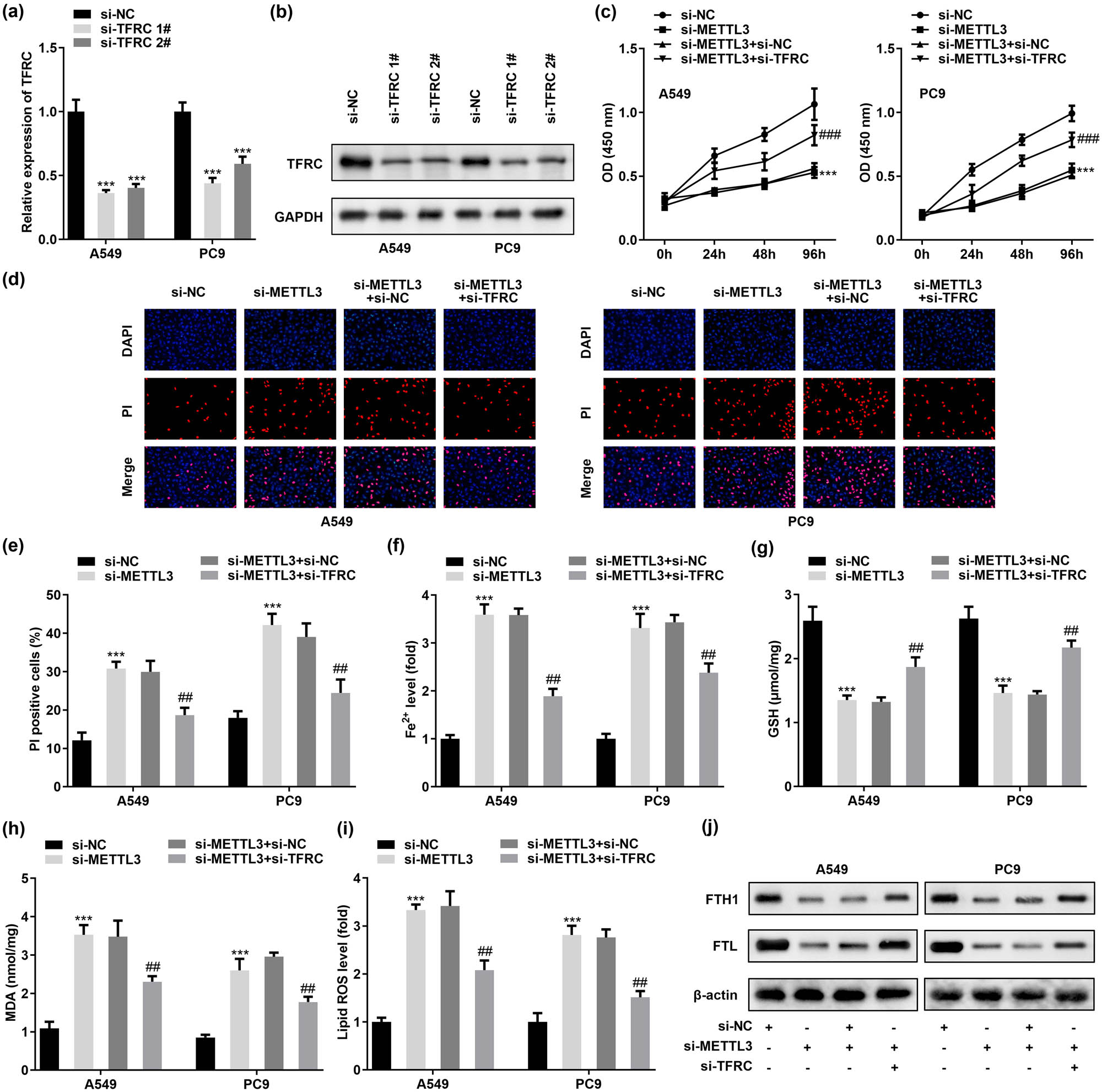
METTL3 knockdown-induced ferroptosis is limited by the decreased TFRC expression levels in LC cells. A549 and PC9 cells were transfected with si-NC, si-METTL3, si-METTL3 + si-NC, and si-METTL3 + si-TFRC, respectively. (a and b) RT-qPCR and western blot was employed to verify the knockdown efficiency of si-TFRC 1# and si-TFRC 2# in A549 and PC9 cells. (b) CCK-8 assay was performed to detect the effect of TFRC silencing on the viability of METTL3-knockdown A549 and PC9 cells. (c and d) DAPI/PI staining was used to verify the effect of TFRC silencing on the death of METTL3-knockdown A549 and PC9 cells. (f–i) Concentrations of Fe2+, GSH, MDA, and lipid ROS in A549 and PC9 cells were evaluated after co-transfection of si-TFRC and si-METTL3. (j) Protein expression of FTH1 and FTL in A549 and PC9 cells co-transfected with si-TFRC and si-METTL3 was measured by western blotting. ## P < 0.01, ### P < 0.001, ***P < 0.001.
3.5 METTL3-mediated m6A modification reduces TFRC stability
Previously, we demonstrated that TFRC levels could be upregulated or reduced in response to METTL3. To further dissect the mechanism by which METTL3 regulates TFRC expression, we performed Me-RIP assays in si-METTL3-treated A549 and PC9 cells. As shown in Figure 6a, METTL3 silencing greatly reduced TFRC enrichment in anti-m6A. Regulation of mRNA stability is one of the major functions of m6A methylation. Therefore, LC cells were treated with actinomycin D to examine the regulation of TFRC mRNA stability by METTL3. The results presented that METTL3 knockdown effectively prolonged the half-life of TFRC proteins in A549 and PC9 cells (Figure 6b and c). Our study further constructed a correlation between m6A methylation and TFRC expression and suggested that METTL3 might hinder TFRC expression in cells by reducing its protein stability in an m6A-dependent manner.
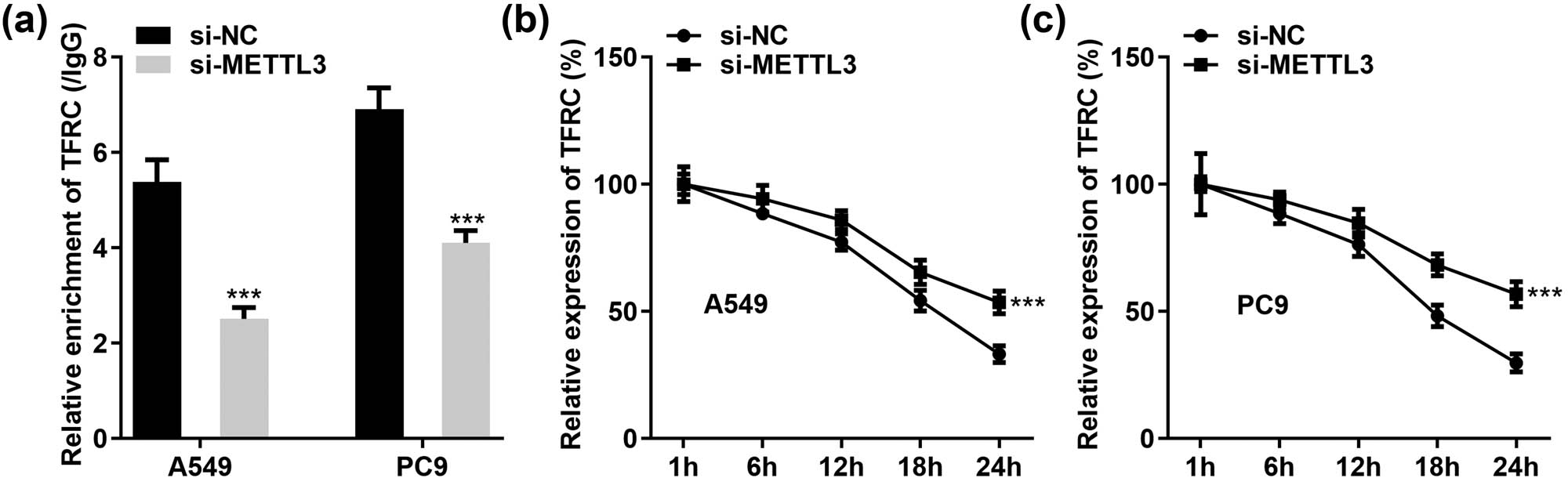
METTL3 silencing enhances the stability of TFRC mRNA. (a) Relative enrichment of TFRC in anti-m6A was reduced after treatment with si-NC or si-METTL3 in A549 and PC9 cells. (b and c) Half-life of TFRC mRNA in both cells (A549 and PC9) was prolonged after METTL3 silencing. ***P < 0.001.
4 Discussion
Ferroptosis, a novel form of cell death, is driven by the massive accumulation of iron and lethal ROS. It is a central force regulating the growth and proliferation of cancer cells, including LC [18], gastric cancer [19], hepatocellular carcinoma [20], and colorectal cancer [21]. Besides, exploratory studies have identified that triggering ferroptosis might be a possible therapeutic mechanism to eradicate malignant cells [22]. Emerging evidence suggested that METTL3 made a partially important contribution to ferroptosis in LC and hepatoblastoma [12,13]. In most cases, METTL3 exerted oncogenic or tumor suppressive functions through deposition of m6A. In highly lethal colorectal cancers, METTL3 was sufficient to stabilize HK2 and GLUT1 expression in an m6A-IGF2BP2/3 manner to accelerate tumor progression [23]. In gastric cancer, the enhancement of ZMYM1 mRNA stability by METTL3 was similarly dependent on its m6A catalytic activity, and enrichment of ZMYM1 could initiate the EMT procedure [24]. However, this is not always the case; for example, METTL3 mediated tumor inhibition by enhancing p53 protein stability in human LC, which was independent of m6A catalytic activity or downstream protein reader [25]. A similar mechanism was seen in the work of Hua et al. The induction of AXL translation by METTL3 in ovarian cancer was similarly independent of its catalytic activity [26]. Given the different mechanisms of METTL3 action in different diseases, we suspected that it might be a valuable biomarker for identifying cancer-impeded ferroptosis and LC. The mechanisms of METTL3 in ferroptosis of LC have rarely been reported. For example, the enrichment of METTL3 in lung adenocarcinoma activated a decrease in SLC7A11 m6A levels, which in turn impeded cell ferroptosis and accelerated tumor progression [12,27]. Our results on METTL3 were consistent with earlier work showing that METTL3 had relatively enriched levels in LC and that loss of METTL3 function increased cell ferroptosis sensitivity rather than resistance. Functional inactivation of METTL3 triggered higher levels of Fe2+, MDA, and lipid ROS, consistent with the general profile of ferroptosis. The upregulation of lipid ROS levels by definition comes from intracellular GSH depletion [28], and our data also showed that reduced GSH triggered ferroptosis after loss of METTL3 function. Studies have showed that FTH1 and FTL decrease free iron levels during ferroptosis, while inhibition of FTH1 and FTL effectively elevates ferroptosis rate, suggesting a negative correlation between FTH1 and FTL and ferroptosis [29,30]. In our study, loss of METTL3 stimulated A549 and PC9 cells to reduce FTH1 and FTL proteins, triggering ferroptosis.
We also found strong sensitivity and specificity of METTL3 and TFRC levels for the diagnosis of LC, and a negative correlation between the two. A previous study noted that elevated levels of TFRC expression in cancer cells contributed to increased iron uptake, and that degradation of TFRC was essential to reduce essential elements of ferroptosis (unstable iron) [31]. Logically, increasing TFRC expression in cancer cells was expected to cause an increase in iron loading and promote ferroptosis in malignant cells. To establish and obtain plausible results in A549 and PC9 cells, we tested the effect of co-silencing METTL3 and TFRC in these two cell lines. As expected, after the depletion of METTL3, TFRC silencing made cells resistant to the increase of unstable iron, with correspondingly elevated cell activity and highlighted levels of lipids ROS and MDA. In other words, TFRC downregulation might be a key event in the ferroptotic processes. A study on hepatocellular carcinoma reported that intracellular degradation of TFRC impeded the sensitivity of tumor cells to ferroptosis [20,32]. Furthermore, a study on intracerebral hemorrhage indicated that METTL3 induced the development of ferroptosis by regulating the m6A level of TFRC mRNA [33]. This prompted us to perform a study of TFRC m6A levels and mRNA stability. We found that in the absence of METTL3, m6A levels of TFRC were decreased, while mRNA stability was effectively improved. We hypothesized that METTL3 and its mediated stability of TFRC determined the resistance of LC cells to ferroptosis, and that this effect is m6A-dependent. The selective mechanism of METTL3 on its direct regulatory targets needs to be further elucidated in the future.
5 Conclusion
In conclusion, we demonstrated that METTL3 was overexpressed in primary LC and exhibited specificity and sensitivity in diagnosing LC. METTL3 silencing significantly increased the sensitivity of LC A549 and PC9 cells to ferroptosis, whereas overexpression of METTL3 presented the opposite effect. Further studies showed that METTL3-mediated ferroptosis resulted from its inverse regulation of TFRC expression. In summary, we suggested that an appropriate reduction of METTL3 might be beneficial for LC patients, as it was an effective strategy to sensitize tumors to ferroptosis.
Acknowledgement
Not applicable.
-
Funding information: This study was approved by Shanghai Municipal Commission of Health and Family Planning (No. 201640203), Shanghai Science and Technology Committee (No. 16140901600), and Talent Climbing Project from Shanghai Tenth People’s Hospital (No. 2018SYPDRC065).
-
Author contributions: Peng Zhang designed the study, performed the experiments, and drafted the manuscript. Su Wang collected data, processed statistical data, and performed the experiments. Yuanyuan Chen analyzed and interpreted the data. Qingbo Yang and Jian Zhou partly contributed to the experiments and data analysis. Wangfu Zang designed, supervised the study, and revised the manuscript. All authors read and approved the final version of the manuscript.
-
Conflict of interest: The authors declare that they have no conflicts of interest to report regarding the present study.
-
Data availability statement: The data that support the findings of this study are available from the corresponding author upon reasonable request.
References
[1] Sung H, Ferlay J, Siegel RL, Laversanne M, Soerjomataram I, Jemal A, et al. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2021;71(3):209–49.10.3322/caac.21660Suche in Google Scholar PubMed
[2] Ferlay J, Colombet M, Soerjomataram I, Parkin DM, Piñeros M, Znaor A, et al. Cancer statistics for the year 2020: an overview. Int J Cancer. 2021;149(4):778–89.10.1002/ijc.33588Suche in Google Scholar PubMed
[3] Nicolau JS, Lopez RVM, de Moraes Luizaga CT, Ribeiro KB, Roela RA, Maistro S, et al. Survival analysis of young adults from a Brazilian cohort of non-small cell lung cancer patients. Ecancermedicalscience. 2021;15:1279.10.3332/ecancer.2021.1279Suche in Google Scholar PubMed PubMed Central
[4] Ha D, Mazzone PJ, Ries AL, Malhotra A, Fuster M. The utility of exercise testing in patients with lung cancer. J Thorac Oncol. 2016;11(9):1397–410.10.1016/j.jtho.2016.04.021Suche in Google Scholar PubMed PubMed Central
[5] Sauler M, Bazan IS, Lee PJ. Cell death in the lung: the apoptosis-necroptosis axis. Annu Rev Physiol. 2019;81:375–402.10.1146/annurev-physiol-020518-114320Suche in Google Scholar PubMed PubMed Central
[6] Fuchs Y, Steller H. Programmed cell death in animal development and disease. Cell. 2011;147(4):742–58.10.1016/j.cell.2011.10.033Suche in Google Scholar PubMed PubMed Central
[7] Wu S, Zhu C, Tang D, Dou QP, Shen J, Chen X. The role of ferroptosis in lung cancer. Biomark Res. 2021;9(1):82.10.1186/s40364-021-00338-0Suche in Google Scholar PubMed PubMed Central
[8] Chen X, Li J, Kang R, Klionsky DJ, Tang D. Ferroptosis: machinery and regulation. Autophagy. 2021;17(9):2054–81.10.1080/15548627.2020.1810918Suche in Google Scholar PubMed PubMed Central
[9] Xie Y, Hou W, Song X, Yu Y, Huang J, Sun X, et al. Ferroptosis: process and function. Cell Death Differ. 2016;23(3):369–79.10.1038/cdd.2015.158Suche in Google Scholar PubMed PubMed Central
[10] Latunde-Dada GO. Ferroptosis: role of lipid peroxidation, iron and ferritinophagy. Biochim Biophys Acta Gen Subj. 2017;1861(8):1893–900.10.1016/j.bbagen.2017.05.019Suche in Google Scholar PubMed
[11] Tang D, Chen X, Kang R, Kroemer G. Ferroptosis: molecular mechanisms and health implications. Cell Res. 2021;31(2):107–25.10.1038/s41422-020-00441-1Suche in Google Scholar PubMed PubMed Central
[12] Xu Y, Lv D, Yan C, Su H, Zhang X, Shi Y, et al. METTL3 promotes lung adenocarcinoma tumor growth and inhibits ferroptosis by stabilizing SLC7A11 m(6)A modification. Cancer Cell Int. 2022;22(1):11.10.1186/s12935-021-02433-6Suche in Google Scholar PubMed PubMed Central
[13] Liu L, He J, Sun G, Huang N, Bian Z, Xu C, et al. The N6-methyladenosine modification enhances ferroptosis resistance through inhibiting SLC7A11 mRNA deadenylation in hepatoblastoma. Clin Transl Med. 2022;12(5):e778.10.1002/ctm2.778Suche in Google Scholar PubMed PubMed Central
[14] Zhang L, Wang X, Che W, Yi Y, Zhou S, Feng Y. Methyltransferase-like 3 silenced inhibited the ferroptosis development via regulating the glutathione peroxidase 4 levels in the intracerebral hemorrhage progression. Bioengineered. 2022;13(6):14215–26.10.1080/21655979.2022.2084494Suche in Google Scholar PubMed PubMed Central
[15] Lin Y, Shen X, Ke Y, Lan C, Chen X, Liang B, et al. Activation of osteoblast ferroptosis via the METTL3/ASK1-p38 signaling pathway in high glucose and high fat (HGHF)-induced diabetic bone loss. Faseb J. 2022;36(3):e22147.10.1096/fj.202101610RSuche in Google Scholar PubMed
[16] Ding Y, Qi N, Wang K, Huang Y, Liao J, Wang H, et al. FTO facilitates lung adenocarcinoma cell progression by activating cell migration through mRNA demethylation. Onco Targets Ther. 2020;13:1461–70.10.2147/OTT.S231914Suche in Google Scholar PubMed PubMed Central
[17] Bogdan AR, Miyazawa M, Hashimoto K, Tsuji Y. Regulators of iron homeostasis: new players in metabolism, cell death, and disease. Trends Biochem Sci. 2016;41(3):274–86.10.1016/j.tibs.2015.11.012Suche in Google Scholar PubMed PubMed Central
[18] Xu W, Deng H, Hu S, Zhang Y, Zheng L, Liu M, et al. Role of ferroptosis in lung diseases. J Inflamm Res. 2021;14:2079–90.10.2147/JIR.S307081Suche in Google Scholar PubMed PubMed Central
[19] Lee JY, Nam M, Son HY, Hyun K, Jang SY, Kim JW, et al. Polyunsaturated fatty acid biosynthesis pathway determines ferroptosis sensitivity in gastric cancer. Proc Natl Acad Sci U S A. 2020;117(51):32433–42.10.1073/pnas.2006828117Suche in Google Scholar PubMed PubMed Central
[20] Ajoolabady A, Tang D, Kroemer G, Ren J. Ferroptosis in hepatocellular carcinoma: mechanisms and targeted therapy. Br J Cancer. 2023;128(2):190–205.10.1038/s41416-022-01998-xSuche in Google Scholar PubMed PubMed Central
[21] Liu Y, Guo F, Guo W, Wang Y, Song W, Fu T. Ferroptosis-related genes are potential prognostic molecular markers for patients with colorectal cancer. Clin Exp Med. 2021;21(3):467–77.10.1007/s10238-021-00697-wSuche in Google Scholar PubMed
[22] Chen X, Kang R, Kroemer G, Tang D. Broadening horizons: the role of ferroptosis in cancer. Nat Rev Clin Oncol. 2021;18(5):280–96.10.1038/s41571-020-00462-0Suche in Google Scholar PubMed
[23] Shen C, Xuan B, Yan T, Ma Y, Xu P, Tian X, et al. m(6)A-dependent glycolysis enhances colorectal cancer progression. Mol Cancer. 2020;19(1):72.10.1186/s12943-020-01190-wSuche in Google Scholar PubMed PubMed Central
[24] Yue B, Song C, Yang L, Cui R, Cheng X, Zhang Z, et al. METTL3-mediated N6-methyladenosine modification is critical for epithelial–mesenchymal transition and metastasis of gastric cancer. Mol Cancer. 2019;18(1):142.10.1186/s12943-019-1065-4Suche in Google Scholar PubMed PubMed Central
[25] Raj N, Wang M, Seoane JA, Zhao RL, Kaiser AM, Moonie NA, et al. The Mettl3 epitranscriptomic writer amplifies p53 stress responses. Mol Cell. 2022;82(13):2370–84.10.1016/j.molcel.2022.04.010Suche in Google Scholar PubMed PubMed Central
[26] Hua W, Zhao Y, Jin X, Yu D, He J, Xie D, et al. METTL3 promotes ovarian carcinoma growth and invasion through the regulation of AXL translation and epithelial to mesenchymal transition. Gynecol Oncol. 2018;151(2):356–65.10.1016/j.ygyno.2018.09.015Suche in Google Scholar PubMed
[27] Ma C, Ma RJ, Hu K, Zheng QM, Wang YP, Zhang N, et al. The molecular mechanism of METTL3 promoting the malignant progression of lung cancer. Cancer Cell Int. 2022;22(1):133.10.1186/s12935-022-02539-5Suche in Google Scholar PubMed PubMed Central
[28] Su LJ, Zhang JH, Gomez H, Murugan R, Hong X, Xu D, et al. Reactive oxygen species-induced lipid peroxidation in apoptosis, autophagy, and ferroptosis. Oxid Med Cell Longev. 2019;2019:5080843.10.1155/2019/5080843Suche in Google Scholar PubMed PubMed Central
[29] Wang Y, Tang M. PM2.5 induces ferroptosis in human endothelial cells through iron overload and redox imbalance. Environ Pollut. 2019;254(Pt A):112937.10.1016/j.envpol.2019.07.105Suche in Google Scholar PubMed
[30] Tang M, Chen Z, Wu D, Chen L. Ferritinophagy/ferroptosis: iron-related newcomers in human diseases. J Cell Physiol. 2018;233(12):9179–90.10.1002/jcp.26954Suche in Google Scholar PubMed
[31] Ye Z, Liu W, Zhuo Q, Hu Q, Liu M, Sun Q, et al. Ferroptosis: final destination for cancer? Cell Prolif. 2020;53(3):e12761.10.1111/cpr.12761Suche in Google Scholar PubMed PubMed Central
[32] Guo S, Chen Y, Xue X, Yang Y, Wang Y, Qiu S, et al. TRIB2 desensitizes ferroptosis via βTrCP-mediated TFRC ubiquitiantion in liver cancer cells. Cell Death Discov. 2021;7(1):196.10.1038/s41420-021-00574-1Suche in Google Scholar PubMed PubMed Central
[33] Zhang L, Wang X, Che W, Zhou S, Feng Y. METTL3 silenced inhibited the ferroptosis development via regulating the TFRC levels in the Intracerebral hemorrhage progression. Brain Res. 2023;1811:148373.10.1016/j.brainres.2023.148373Suche in Google Scholar PubMed
© 2024 the author(s), published by De Gruyter
This work is licensed under the Creative Commons Attribution 4.0 International License.
Artikel in diesem Heft
- Research Articles
- EDNRB inhibits the growth and migration of prostate cancer cells by activating the cGMP-PKG pathway
- STK11 (LKB1) mutation suppresses ferroptosis in lung adenocarcinoma by facilitating monounsaturated fatty acid synthesis
- Association of SOX6 gene polymorphisms with Kashin-Beck disease risk in the Chinese Han population
- The pyroptosis-related signature predicts prognosis and influences the tumor immune microenvironment in dedifferentiated liposarcoma
- METTL3 attenuates ferroptosis sensitivity in lung cancer via modulating TFRC
- Identification and validation of molecular subtypes and prognostic signature for stage I and stage II gastric cancer based on neutrophil extracellular traps
- Novel lumbar plexus block versus femoral nerve block for analgesia and motor recovery after total knee arthroplasty
- Correlation between ABCB1 and OLIG2 polymorphisms and the severity and prognosis of patients with cerebral infarction
- Study on the radiotherapy effect and serum neutral granulocyte lymphocyte ratio and inflammatory factor expression of nasopharyngeal carcinoma
- Transcriptome analysis of effects of Tecrl deficiency on cardiometabolic and calcium regulation in cardiac tissue
- Aflatoxin B1 induces infertility, fetal deformities, and potential therapies
- Serum levels of HMW adiponectin and its receptors are associated with cytokine levels and clinical characteristics in chronic obstructive pulmonary disease
- METTL3-mediated methylation of CYP2C19 mRNA may aggravate clopidogrel resistance in ischemic stroke patients
- Understand how machine learning impact lung cancer research from 2010 to 2021: A bibliometric analysis
- Pressure ulcers in German hospitals: Analysis of reimbursement and length of stay
- Metformin plus L-carnitine enhances brown/beige adipose tissue activity via Nrf2/HO-1 signaling to reduce lipid accumulation and inflammation in murine obesity
- Downregulation of carbonic anhydrase IX expression in mouse xenograft nasopharyngeal carcinoma model via doxorubicin nanobubble combined with ultrasound
- Feasibility of 3-dimensional printed models in simulated training and teaching of transcatheter aortic valve replacement
- miR-335-3p improves type II diabetes mellitus by IGF-1 regulating macrophage polarization
- The analyses of human MCPH1 DNA repair machinery and genetic variations
- Activation of Piezo1 increases the sensitivity of breast cancer to hyperthermia therapy
- Comprehensive analysis based on the disulfidptosis-related genes identifies hub genes and immune infiltration for pancreatic adenocarcinoma
- Changes of serum CA125 and PGE2 before and after high-intensity focused ultrasound combined with GnRH-a in treatment of patients with adenomyosis
- The clinical value of the hepatic venous pressure gradient in patients undergoing hepatic resection for hepatocellular carcinoma with or without liver cirrhosis
- Development and validation of a novel model to predict pulmonary embolism in cardiology suspected patients: A 10-year retrospective analysis
- Downregulation of lncRNA XLOC_032768 in diabetic patients predicts the occurrence of diabetic nephropathy
- Circ_0051428 targeting miR-885-3p/MMP2 axis enhances the malignancy of cervical cancer
- Effectiveness of ginkgo diterpene lactone meglumine on cognitive function in patients with acute ischemic stroke
- The construction of a novel prognostic prediction model for glioma based on GWAS-identified prognostic-related risk loci
- Evaluating the impact of childhood BMI on the risk of coronavirus disease 2019: A Mendelian randomization study
- Lactate dehydrogenase to albumin ratio is associated with in-hospital mortality in patients with acute heart failure: Data from the MIMIC-III database
- CD36-mediated podocyte lipotoxicity promotes foot process effacement
- Efficacy of etonogestrel subcutaneous implants versus the levonorgestrel-releasing intrauterine system in the conservative treatment of adenomyosis
- FLRT2 mediates chondrogenesis of nasal septal cartilage and mandibular condyle cartilage
- Challenges in treating primary immune thrombocytopenia patients undergoing COVID-19 vaccination: A retrospective study
- Let-7 family regulates HaCaT cell proliferation and apoptosis via the ΔNp63/PI3K/AKT pathway
- Phospholipid transfer protein ameliorates sepsis-induced cardiac dysfunction through NLRP3 inflammasome inhibition
- Postoperative cognitive dysfunction in elderly patients with colorectal cancer: A randomized controlled study comparing goal-directed and conventional fluid therapy
- Long-pulsed ultrasound-mediated microbubble thrombolysis in a rat model of microvascular obstruction
- High SEC61A1 expression predicts poor outcome of acute myeloid leukemia
- Comparison of polymerase chain reaction and next-generation sequencing with conventional urine culture for the diagnosis of urinary tract infections: A meta-analysis
- Secreted frizzled-related protein 5 protects against renal fibrosis by inhibiting Wnt/β-catenin pathway
- Pan-cancer and single-cell analysis of actin cytoskeleton genes related to disulfidptosis
- Overexpression of miR-532-5p restrains oxidative stress response of chondrocytes in nontraumatic osteonecrosis of the femoral head by inhibiting ABL1
- Autologous liver transplantation for unresectable hepatobiliary malignancies in enhanced recovery after surgery model
- Clinical analysis of incomplete rupture of the uterus secondary to previous cesarean section
- Abnormal sleep duration is associated with sarcopenia in older Chinese people: A large retrospective cross-sectional study
- No genetic causality between obesity and benign paroxysmal vertigo: A two-sample Mendelian randomization study
- Identification and validation of autophagy-related genes in SSc
- Long non-coding RNA SRA1 suppresses radiotherapy resistance in esophageal squamous cell carcinoma by modulating glycolytic reprogramming
- Evaluation of quality of life in patients with schizophrenia: An inpatient social welfare institution-based cross-sectional study
- The possible role of oxidative stress marker glutathione in the assessment of cognitive impairment in multiple sclerosis
- Compilation of a self-management assessment scale for postoperative patients with aortic dissection
- Left atrial appendage closure in conjunction with radiofrequency ablation: Effects on left atrial functioning in patients with paroxysmal atrial fibrillation
- Effect of anterior femoral cortical notch grade on postoperative function and complications during TKA surgery: A multicenter, retrospective study
- Clinical characteristics and assessment of risk factors in patients with influenza A-induced severe pneumonia after the prevalence of SARS-CoV-2
- Analgesia nociception index is an indicator of laparoscopic trocar insertion-induced transient nociceptive stimuli
- High STAT4 expression correlates with poor prognosis in acute myeloid leukemia and facilitates disease progression by upregulating VEGFA expression
- Factors influencing cardiovascular system-related post-COVID-19 sequelae: A single-center cohort study
- HOXD10 regulates intestinal permeability and inhibits inflammation of dextran sulfate sodium-induced ulcerative colitis through the inactivation of the Rho/ROCK/MMPs axis
- Mesenchymal stem cell-derived exosomal miR-26a induces ferroptosis, suppresses hepatic stellate cell activation, and ameliorates liver fibrosis by modulating SLC7A11
- Endovascular thrombectomy versus intravenous thrombolysis for primary distal, medium vessel occlusion in acute ischemic stroke
- ANO6 (TMEM16F) inhibits gastrointestinal stromal tumor growth and induces ferroptosis
- Prognostic value of EIF5A2 in solid tumors: A meta-analysis and bioinformatics analysis
- The role of enhanced expression of Cx43 in patients with ulcerative colitis
- Choosing a COVID-19 vaccination site might be driven by anxiety and body vigilance
- Role of ICAM-1 in triple-negative breast cancer
- Cost-effectiveness of ambroxol in the treatment of Gaucher disease type 2
- HLA-DRB5 promotes immune thrombocytopenia via activating CD8+ T cells
- Efficacy and factors of myofascial release therapy combined with electrical and magnetic stimulation in the treatment of chronic pelvic pain syndrome
- Efficacy of tacrolimus monotherapy in primary membranous nephropathy
- Mechanisms of Tripterygium wilfordii Hook F on treating rheumatoid arthritis explored by network pharmacology analysis and molecular docking
- FBXO45 levels regulated ferroptosis renal tubular epithelial cells in a model of diabetic nephropathy by PLK1
- Optimizing anesthesia strategies to NSCLC patients in VATS procedures: Insights from drug requirements and patient recovery patterns
- Alpha-lipoic acid upregulates the PPARγ/NRF2/GPX4 signal pathway to inhibit ferroptosis in the pathogenesis of unexplained recurrent pregnancy loss
- Correlation between fat-soluble vitamin levels and inflammatory factors in paediatric community-acquired pneumonia: A prospective study
- CD1d affects the proliferation, migration, and apoptosis of human papillary thyroid carcinoma TPC-1 cells via regulating MAPK/NF-κB signaling pathway
- miR-let-7a inhibits sympathetic nerve remodeling after myocardial infarction by downregulating the expression of nerve growth factor
- Immune response analysis of solid organ transplantation recipients inoculated with inactivated COVID-19 vaccine: A retrospective analysis
- The H2Valdien derivatives regulate the epithelial–mesenchymal transition of hepatoma carcinoma cells through the Hedgehog signaling pathway
- Clinical efficacy of dexamethasone combined with isoniazid in the treatment of tuberculous meningitis and its effect on peripheral blood T cell subsets
- Comparison of short-segment and long-segment fixation in treatment of degenerative scoliosis and analysis of factors associated with adjacent spondylolisthesis
- Lycopene inhibits pyroptosis of endothelial progenitor cells induced by ox-LDL through the AMPK/mTOR/NLRP3 pathway
- Methylation regulation for FUNDC1 stability in childhood leukemia was up-regulated and facilitates metastasis and reduces ferroptosis of leukemia through mitochondrial damage by FBXL2
- Correlation of single-fiber electromyography studies and functional status in patients with amyotrophic lateral sclerosis
- Risk factors of postoperative airway obstruction complications in children with oral floor mass
- Expression levels and clinical significance of serum miR-19a/CCL20 in patients with acute cerebral infarction
- Physical activity and mental health trends in Korean adolescents: Analyzing the impact of the COVID-19 pandemic from 2018 to 2022
- Evaluating anemia in HIV-infected patients using chest CT
- Ponticulus posticus and skeletal malocclusion: A pilot study in a Southern Italian pre-orthodontic court
- Causal association of circulating immune cells and lymphoma: A Mendelian randomization study
- Assessment of the renal function and fibrosis indexes of conventional western medicine with Chinese medicine for dredging collaterals on treating renal fibrosis: A systematic review and meta-analysis
- Comprehensive landscape of integrator complex subunits and their association with prognosis and tumor microenvironment in gastric cancer
- New target-HMGCR inhibitors for the treatment of primary sclerosing cholangitis: A drug Mendelian randomization study
- Population pharmacokinetics of meropenem in critically ill patients
- Comparison of the ability of newly inflammatory markers to predict complicated appendicitis
- Comparative morphology of the cruciate ligaments: A radiological study
- Immune landscape of hepatocellular carcinoma: The central role of TP53-inducible glycolysis and apoptosis regulator
- Serum SIRT3 levels in epilepsy patients and its association with clinical outcomes and severity: A prospective observational study
- SHP-1 mediates cigarette smoke extract-induced epithelial–mesenchymal transformation and inflammation in 16HBE cells
- Acute hyper-hypoxia accelerates the development of depression in mice via the IL-6/PGC1α/MFN2 signaling pathway
- The GJB3 correlates with the prognosis, immune cell infiltration, and therapeutic responses in lung adenocarcinoma
- Physical fitness and blood parameters outcomes of breast cancer survivor in a low-intensity circuit resistance exercise program
- Exploring anesthetic-induced gene expression changes and immune cell dynamics in atrial tissue post-coronary artery bypass graft surgery
- Empagliflozin improves aortic injury in obese mice by regulating fatty acid metabolism
- Analysis of the risk factors of the radiation-induced encephalopathy in nasopharyngeal carcinoma: A retrospective cohort study
- Reproductive outcomes in women with BRCA 1/2 germline mutations: A retrospective observational study and literature review
- Evaluation of upper airway ultrasonographic measurements in predicting difficult intubation: A cross-section of the Turkish population
- Prognostic and diagnostic value of circulating IGFBP2 in pancreatic cancer
- Postural stability after operative reconstruction of the AFTL in chronic ankle instability comparing three different surgical techniques
- Research trends related to emergence agitation in the post-anaesthesia care unit from 2001 to 2023: A bibliometric analysis
- Frequency and clinicopathological correlation of gastrointestinal polyps: A six-year single center experience
- ACSL4 mediates inflammatory bowel disease and contributes to LPS-induced intestinal epithelial cell dysfunction by activating ferroptosis and inflammation
- Affibody-based molecular probe 99mTc-(HE)3ZHER2:V2 for non-invasive HER2 detection in ovarian and breast cancer xenografts
- Effectiveness of nutritional support for clinical outcomes in gastric cancer patients: A meta-analysis of randomized controlled trials
- The relationship between IFN-γ, IL-10, IL-6 cytokines, and severity of the condition with serum zinc and Fe in children infected with Mycoplasma pneumoniae
- Paraquat disrupts the blood–brain barrier by increasing IL-6 expression and oxidative stress through the activation of PI3K/AKT signaling pathway
- Sleep quality associate with the increased prevalence of cognitive impairment in coronary artery disease patients: A retrospective case–control study
- Dioscin protects against chronic prostatitis through the TLR4/NF-κB pathway
- Association of polymorphisms in FBN1, MYH11, and TGF-β signaling-related genes with susceptibility of sporadic thoracic aortic aneurysm and dissection in the Zhejiang Han population
- Application value of multi-parameter magnetic resonance image-transrectal ultrasound cognitive fusion in prostate biopsy
- Laboratory variables‐based artificial neural network models for predicting fatty liver disease: A retrospective study
- Decreased BIRC5-206 promotes epithelial–mesenchymal transition in nasopharyngeal carcinoma through sponging miR-145-5p
- Sepsis induces the cardiomyocyte apoptosis and cardiac dysfunction through activation of YAP1/Serpine1/caspase-3 pathway
- Assessment of iron metabolism and iron deficiency in incident patients on incident continuous ambulatory peritoneal dialysis
- Tibial periosteum flap combined with autologous bone grafting in the treatment of Gustilo-IIIB/IIIC open tibial fractures
- The application of intravenous general anesthesia under nasopharyngeal airway assisted ventilation undergoing ureteroscopic holmium laser lithotripsy: A prospective, single-center, controlled trial
- Long intergenic noncoding RNA for IGF2BP2 stability suppresses gastric cancer cell apoptosis by inhibiting the maturation of microRNA-34a
- Role of FOXM1 and AURKB in regulating keratinocyte function in psoriasis
- Parental control attitudes over their pre-school children’s diet
- The role of auto-HSCT in extranodal natural killer/T cell lymphoma
- Significance of negative cervical cytology and positive HPV in the diagnosis of cervical lesions by colposcopy
- Echinacoside inhibits PASMCs calcium overload to prevent hypoxic pulmonary artery remodeling by regulating TRPC1/4/6 and calmodulin
- ADAR1 plays a protective role in proximal tubular cells under high glucose conditions by attenuating the PI3K/AKT/mTOR signaling pathway
- The risk of cancer among insulin glargine users in Lithuania: A retrospective population-based study
- The unusual location of primary hydatid cyst: A case series study
- Intraoperative changes in electrophysiological monitoring can be used to predict clinical outcomes in patients with spinal cavernous malformation
- Obesity and risk of placenta accreta spectrum: A meta-analysis
- Shikonin alleviates asthma phenotypes in mice via an airway epithelial STAT3-dependent mechanism
- NSUN6 and HTR7 disturbed the stability of carotid atherosclerotic plaques by regulating the immune responses of macrophages
- The effect of COVID-19 lockdown on admission rates in Maternity Hospital
- Temporal muscle thickness is not a prognostic predictor in patients with high-grade glioma, an experience at two centers in China
- Luteolin alleviates cerebral ischemia/reperfusion injury by regulating cell pyroptosis
- Therapeutic role of respiratory exercise in patients with tuberculous pleurisy
- Effects of CFTR-ENaC on spinal cord edema after spinal cord injury
- Irisin-regulated lncRNAs and their potential regulatory functions in chondrogenic differentiation of human mesenchymal stem cells
- DMD mutations in pediatric patients with phenotypes of Duchenne/Becker muscular dystrophy
- Combination of C-reactive protein and fibrinogen-to-albumin ratio as a novel predictor of all-cause mortality in heart failure patients
- Significant role and the underly mechanism of cullin-1 in chronic obstructive pulmonary disease
- Ferroptosis-related prognostic model of mantle cell lymphoma
- Observation of choking reaction and other related indexes in elderly painless fiberoptic bronchoscopy with transnasal high-flow humidification oxygen therapy
- A bibliometric analysis of Prader-Willi syndrome from 2002 to 2022
- The causal effects of childhood sunburn occasions on melanoma: A univariable and multivariable Mendelian randomization study
- Oxidative stress regulates glycogen synthase kinase-3 in lymphocytes of diabetes mellitus patients complicated with cerebral infarction
- Role of COX6C and NDUFB3 in septic shock and stroke
- Trends in disease burden of type 2 diabetes, stroke, and hypertensive heart disease attributable to high BMI in China: 1990–2019
- Purinergic P2X7 receptor mediates hyperoxia-induced injury in pulmonary microvascular endothelial cells via NLRP3-mediated pyroptotic pathway
- Investigating the role of oviductal mucosa–endometrial co-culture in modulating factors relevant to embryo implantation
- Analgesic effect of external oblique intercostal block in laparoscopic cholecystectomy: A retrospective study
- Elevated serum miR-142-5p correlates with ischemic lesions and both NSE and S100β in ischemic stroke patients
- Correlation between the mechanism of arteriopathy in IgA nephropathy and blood stasis syndrome: A cohort study
- Risk factors for progressive kyphosis after percutaneous kyphoplasty in osteoporotic vertebral compression fracture
- Predictive role of neuron-specific enolase and S100-β in early neurological deterioration and unfavorable prognosis in patients with ischemic stroke
- The potential risk factors of postoperative cognitive dysfunction for endovascular therapy in acute ischemic stroke with general anesthesia
- Fluoxetine inhibited RANKL-induced osteoclastic differentiation in vitro
- Detection of serum FOXM1 and IGF2 in patients with ARDS and their correlation with disease and prognosis
- Rhein promotes skin wound healing by activating the PI3K/AKT signaling pathway
- Differences in mortality risk by levels of physical activity among persons with disabilities in South Korea
- Review Articles
- Cutaneous signs of selected cardiovascular disorders: A narrative review
- XRCC1 and hOGG1 polymorphisms and endometrial carcinoma: A meta-analysis
- A narrative review on adverse drug reactions of COVID-19 treatments on the kidney
- Emerging role and function of SPDL1 in human health and diseases
- Adverse reactions of piperacillin: A literature review of case reports
- Molecular mechanism and intervention measures of microvascular complications in diabetes
- Regulation of mesenchymal stem cell differentiation by autophagy
- Molecular landscape of borderline ovarian tumours: A systematic review
- Advances in synthetic lethality modalities for glioblastoma multiforme
- Investigating hormesis, aging, and neurodegeneration: From bench to clinics
- Frankincense: A neuronutrient to approach Parkinson’s disease treatment
- Sox9: A potential regulator of cancer stem cells in osteosarcoma
- Early detection of cardiovascular risk markers through non-invasive ultrasound methodologies in periodontitis patients
- Advanced neuroimaging and criminal interrogation in lie detection
- Maternal factors for neural tube defects in offspring: An umbrella review
- The chemoprotective hormetic effects of rosmarinic acid
- CBD’s potential impact on Parkinson’s disease: An updated overview
- Progress in cytokine research for ARDS: A comprehensive review
- Utilizing reactive oxygen species-scavenging nanoparticles for targeting oxidative stress in the treatment of ischemic stroke: A review
- NRXN1-related disorders, attempt to better define clinical assessment
- Lidocaine infusion for the treatment of complex regional pain syndrome: Case series and literature review
- Trends and future directions of autophagy in osteosarcoma: A bibliometric analysis
- Iron in ventricular remodeling and aneurysms post-myocardial infarction
- Case Reports
- Sirolimus potentiated angioedema: A case report and review of the literature
- Identification of mixed anaerobic infections after inguinal hernia repair based on metagenomic next-generation sequencing: A case report
- Successful treatment with bortezomib in combination with dexamethasone in a middle-aged male with idiopathic multicentric Castleman’s disease: A case report
- Complete heart block associated with hepatitis A infection in a female child with fatal outcome
- Elevation of D-dimer in eosinophilic gastrointestinal diseases in the absence of venous thrombosis: A case series and literature review
- Four years of natural progressive course: A rare case report of juvenile Xp11.2 translocations renal cell carcinoma with TFE3 gene fusion
- Advancing prenatal diagnosis: Echocardiographic detection of Scimitar syndrome in China – A case series
- Outcomes and complications of hemodialysis in patients with renal cancer following bilateral nephrectomy
- Anti-HMGCR myopathy mimicking facioscapulohumeral muscular dystrophy
- Recurrent opportunistic infections in a HIV-negative patient with combined C6 and NFKB1 mutations: A case report, pedigree analysis, and literature review
- Letter to the Editor
- Letter to the Editor: Total parenteral nutrition-induced Wernicke’s encephalopathy after oncologic gastrointestinal surgery
- Erratum
- Erratum to “Bladder-embedded ectopic intrauterine device with calculus”
- Retraction
- Retraction of “XRCC1 and hOGG1 polymorphisms and endometrial carcinoma: A meta-analysis”
- Corrigendum
- Corrigendum to “Investigating hormesis, aging, and neurodegeneration: From bench to clinics”
- Corrigendum to “Frankincense: A neuronutrient to approach Parkinson’s disease treatment”
- Special Issue The evolving saga of RNAs from bench to bedside - Part II
- Machine-learning-based prediction of a diagnostic model using autophagy-related genes based on RNA sequencing for patients with papillary thyroid carcinoma
- Unlocking the future of hepatocellular carcinoma treatment: A comprehensive analysis of disulfidptosis-related lncRNAs for prognosis and drug screening
- Elevated mRNA level indicates FSIP1 promotes EMT and gastric cancer progression by regulating fibroblasts in tumor microenvironment
- Special Issue Advancements in oncology: bridging clinical and experimental research - Part I
- Ultrasound-guided transperineal vs transrectal prostate biopsy: A meta-analysis of diagnostic accuracy and complication rates
- Assessment of diagnostic value of unilateral systematic biopsy combined with targeted biopsy in detecting clinically significant prostate cancer
- SENP7 inhibits glioblastoma metastasis and invasion by dissociating SUMO2/3 binding to specific target proteins
- MARK1 suppress malignant progression of hepatocellular carcinoma and improves sorafenib resistance through negatively regulating POTEE
- Analysis of postoperative complications in bladder cancer patients
- Carboplatin combined with arsenic trioxide versus carboplatin combined with docetaxel treatment for LACC: A randomized, open-label, phase II clinical study
- Special Issue Exploring the biological mechanism of human diseases based on MultiOmics Technology - Part I
- Comprehensive pan-cancer investigation of carnosine dipeptidase 1 and its prospective prognostic significance in hepatocellular carcinoma
- Identification of signatures associated with microsatellite instability and immune characteristics to predict the prognostic risk of colon cancer
- Single-cell analysis identified key macrophage subpopulations associated with atherosclerosis
Artikel in diesem Heft
- Research Articles
- EDNRB inhibits the growth and migration of prostate cancer cells by activating the cGMP-PKG pathway
- STK11 (LKB1) mutation suppresses ferroptosis in lung adenocarcinoma by facilitating monounsaturated fatty acid synthesis
- Association of SOX6 gene polymorphisms with Kashin-Beck disease risk in the Chinese Han population
- The pyroptosis-related signature predicts prognosis and influences the tumor immune microenvironment in dedifferentiated liposarcoma
- METTL3 attenuates ferroptosis sensitivity in lung cancer via modulating TFRC
- Identification and validation of molecular subtypes and prognostic signature for stage I and stage II gastric cancer based on neutrophil extracellular traps
- Novel lumbar plexus block versus femoral nerve block for analgesia and motor recovery after total knee arthroplasty
- Correlation between ABCB1 and OLIG2 polymorphisms and the severity and prognosis of patients with cerebral infarction
- Study on the radiotherapy effect and serum neutral granulocyte lymphocyte ratio and inflammatory factor expression of nasopharyngeal carcinoma
- Transcriptome analysis of effects of Tecrl deficiency on cardiometabolic and calcium regulation in cardiac tissue
- Aflatoxin B1 induces infertility, fetal deformities, and potential therapies
- Serum levels of HMW adiponectin and its receptors are associated with cytokine levels and clinical characteristics in chronic obstructive pulmonary disease
- METTL3-mediated methylation of CYP2C19 mRNA may aggravate clopidogrel resistance in ischemic stroke patients
- Understand how machine learning impact lung cancer research from 2010 to 2021: A bibliometric analysis
- Pressure ulcers in German hospitals: Analysis of reimbursement and length of stay
- Metformin plus L-carnitine enhances brown/beige adipose tissue activity via Nrf2/HO-1 signaling to reduce lipid accumulation and inflammation in murine obesity
- Downregulation of carbonic anhydrase IX expression in mouse xenograft nasopharyngeal carcinoma model via doxorubicin nanobubble combined with ultrasound
- Feasibility of 3-dimensional printed models in simulated training and teaching of transcatheter aortic valve replacement
- miR-335-3p improves type II diabetes mellitus by IGF-1 regulating macrophage polarization
- The analyses of human MCPH1 DNA repair machinery and genetic variations
- Activation of Piezo1 increases the sensitivity of breast cancer to hyperthermia therapy
- Comprehensive analysis based on the disulfidptosis-related genes identifies hub genes and immune infiltration for pancreatic adenocarcinoma
- Changes of serum CA125 and PGE2 before and after high-intensity focused ultrasound combined with GnRH-a in treatment of patients with adenomyosis
- The clinical value of the hepatic venous pressure gradient in patients undergoing hepatic resection for hepatocellular carcinoma with or without liver cirrhosis
- Development and validation of a novel model to predict pulmonary embolism in cardiology suspected patients: A 10-year retrospective analysis
- Downregulation of lncRNA XLOC_032768 in diabetic patients predicts the occurrence of diabetic nephropathy
- Circ_0051428 targeting miR-885-3p/MMP2 axis enhances the malignancy of cervical cancer
- Effectiveness of ginkgo diterpene lactone meglumine on cognitive function in patients with acute ischemic stroke
- The construction of a novel prognostic prediction model for glioma based on GWAS-identified prognostic-related risk loci
- Evaluating the impact of childhood BMI on the risk of coronavirus disease 2019: A Mendelian randomization study
- Lactate dehydrogenase to albumin ratio is associated with in-hospital mortality in patients with acute heart failure: Data from the MIMIC-III database
- CD36-mediated podocyte lipotoxicity promotes foot process effacement
- Efficacy of etonogestrel subcutaneous implants versus the levonorgestrel-releasing intrauterine system in the conservative treatment of adenomyosis
- FLRT2 mediates chondrogenesis of nasal septal cartilage and mandibular condyle cartilage
- Challenges in treating primary immune thrombocytopenia patients undergoing COVID-19 vaccination: A retrospective study
- Let-7 family regulates HaCaT cell proliferation and apoptosis via the ΔNp63/PI3K/AKT pathway
- Phospholipid transfer protein ameliorates sepsis-induced cardiac dysfunction through NLRP3 inflammasome inhibition
- Postoperative cognitive dysfunction in elderly patients with colorectal cancer: A randomized controlled study comparing goal-directed and conventional fluid therapy
- Long-pulsed ultrasound-mediated microbubble thrombolysis in a rat model of microvascular obstruction
- High SEC61A1 expression predicts poor outcome of acute myeloid leukemia
- Comparison of polymerase chain reaction and next-generation sequencing with conventional urine culture for the diagnosis of urinary tract infections: A meta-analysis
- Secreted frizzled-related protein 5 protects against renal fibrosis by inhibiting Wnt/β-catenin pathway
- Pan-cancer and single-cell analysis of actin cytoskeleton genes related to disulfidptosis
- Overexpression of miR-532-5p restrains oxidative stress response of chondrocytes in nontraumatic osteonecrosis of the femoral head by inhibiting ABL1
- Autologous liver transplantation for unresectable hepatobiliary malignancies in enhanced recovery after surgery model
- Clinical analysis of incomplete rupture of the uterus secondary to previous cesarean section
- Abnormal sleep duration is associated with sarcopenia in older Chinese people: A large retrospective cross-sectional study
- No genetic causality between obesity and benign paroxysmal vertigo: A two-sample Mendelian randomization study
- Identification and validation of autophagy-related genes in SSc
- Long non-coding RNA SRA1 suppresses radiotherapy resistance in esophageal squamous cell carcinoma by modulating glycolytic reprogramming
- Evaluation of quality of life in patients with schizophrenia: An inpatient social welfare institution-based cross-sectional study
- The possible role of oxidative stress marker glutathione in the assessment of cognitive impairment in multiple sclerosis
- Compilation of a self-management assessment scale for postoperative patients with aortic dissection
- Left atrial appendage closure in conjunction with radiofrequency ablation: Effects on left atrial functioning in patients with paroxysmal atrial fibrillation
- Effect of anterior femoral cortical notch grade on postoperative function and complications during TKA surgery: A multicenter, retrospective study
- Clinical characteristics and assessment of risk factors in patients with influenza A-induced severe pneumonia after the prevalence of SARS-CoV-2
- Analgesia nociception index is an indicator of laparoscopic trocar insertion-induced transient nociceptive stimuli
- High STAT4 expression correlates with poor prognosis in acute myeloid leukemia and facilitates disease progression by upregulating VEGFA expression
- Factors influencing cardiovascular system-related post-COVID-19 sequelae: A single-center cohort study
- HOXD10 regulates intestinal permeability and inhibits inflammation of dextran sulfate sodium-induced ulcerative colitis through the inactivation of the Rho/ROCK/MMPs axis
- Mesenchymal stem cell-derived exosomal miR-26a induces ferroptosis, suppresses hepatic stellate cell activation, and ameliorates liver fibrosis by modulating SLC7A11
- Endovascular thrombectomy versus intravenous thrombolysis for primary distal, medium vessel occlusion in acute ischemic stroke
- ANO6 (TMEM16F) inhibits gastrointestinal stromal tumor growth and induces ferroptosis
- Prognostic value of EIF5A2 in solid tumors: A meta-analysis and bioinformatics analysis
- The role of enhanced expression of Cx43 in patients with ulcerative colitis
- Choosing a COVID-19 vaccination site might be driven by anxiety and body vigilance
- Role of ICAM-1 in triple-negative breast cancer
- Cost-effectiveness of ambroxol in the treatment of Gaucher disease type 2
- HLA-DRB5 promotes immune thrombocytopenia via activating CD8+ T cells
- Efficacy and factors of myofascial release therapy combined with electrical and magnetic stimulation in the treatment of chronic pelvic pain syndrome
- Efficacy of tacrolimus monotherapy in primary membranous nephropathy
- Mechanisms of Tripterygium wilfordii Hook F on treating rheumatoid arthritis explored by network pharmacology analysis and molecular docking
- FBXO45 levels regulated ferroptosis renal tubular epithelial cells in a model of diabetic nephropathy by PLK1
- Optimizing anesthesia strategies to NSCLC patients in VATS procedures: Insights from drug requirements and patient recovery patterns
- Alpha-lipoic acid upregulates the PPARγ/NRF2/GPX4 signal pathway to inhibit ferroptosis in the pathogenesis of unexplained recurrent pregnancy loss
- Correlation between fat-soluble vitamin levels and inflammatory factors in paediatric community-acquired pneumonia: A prospective study
- CD1d affects the proliferation, migration, and apoptosis of human papillary thyroid carcinoma TPC-1 cells via regulating MAPK/NF-κB signaling pathway
- miR-let-7a inhibits sympathetic nerve remodeling after myocardial infarction by downregulating the expression of nerve growth factor
- Immune response analysis of solid organ transplantation recipients inoculated with inactivated COVID-19 vaccine: A retrospective analysis
- The H2Valdien derivatives regulate the epithelial–mesenchymal transition of hepatoma carcinoma cells through the Hedgehog signaling pathway
- Clinical efficacy of dexamethasone combined with isoniazid in the treatment of tuberculous meningitis and its effect on peripheral blood T cell subsets
- Comparison of short-segment and long-segment fixation in treatment of degenerative scoliosis and analysis of factors associated with adjacent spondylolisthesis
- Lycopene inhibits pyroptosis of endothelial progenitor cells induced by ox-LDL through the AMPK/mTOR/NLRP3 pathway
- Methylation regulation for FUNDC1 stability in childhood leukemia was up-regulated and facilitates metastasis and reduces ferroptosis of leukemia through mitochondrial damage by FBXL2
- Correlation of single-fiber electromyography studies and functional status in patients with amyotrophic lateral sclerosis
- Risk factors of postoperative airway obstruction complications in children with oral floor mass
- Expression levels and clinical significance of serum miR-19a/CCL20 in patients with acute cerebral infarction
- Physical activity and mental health trends in Korean adolescents: Analyzing the impact of the COVID-19 pandemic from 2018 to 2022
- Evaluating anemia in HIV-infected patients using chest CT
- Ponticulus posticus and skeletal malocclusion: A pilot study in a Southern Italian pre-orthodontic court
- Causal association of circulating immune cells and lymphoma: A Mendelian randomization study
- Assessment of the renal function and fibrosis indexes of conventional western medicine with Chinese medicine for dredging collaterals on treating renal fibrosis: A systematic review and meta-analysis
- Comprehensive landscape of integrator complex subunits and their association with prognosis and tumor microenvironment in gastric cancer
- New target-HMGCR inhibitors for the treatment of primary sclerosing cholangitis: A drug Mendelian randomization study
- Population pharmacokinetics of meropenem in critically ill patients
- Comparison of the ability of newly inflammatory markers to predict complicated appendicitis
- Comparative morphology of the cruciate ligaments: A radiological study
- Immune landscape of hepatocellular carcinoma: The central role of TP53-inducible glycolysis and apoptosis regulator
- Serum SIRT3 levels in epilepsy patients and its association with clinical outcomes and severity: A prospective observational study
- SHP-1 mediates cigarette smoke extract-induced epithelial–mesenchymal transformation and inflammation in 16HBE cells
- Acute hyper-hypoxia accelerates the development of depression in mice via the IL-6/PGC1α/MFN2 signaling pathway
- The GJB3 correlates with the prognosis, immune cell infiltration, and therapeutic responses in lung adenocarcinoma
- Physical fitness and blood parameters outcomes of breast cancer survivor in a low-intensity circuit resistance exercise program
- Exploring anesthetic-induced gene expression changes and immune cell dynamics in atrial tissue post-coronary artery bypass graft surgery
- Empagliflozin improves aortic injury in obese mice by regulating fatty acid metabolism
- Analysis of the risk factors of the radiation-induced encephalopathy in nasopharyngeal carcinoma: A retrospective cohort study
- Reproductive outcomes in women with BRCA 1/2 germline mutations: A retrospective observational study and literature review
- Evaluation of upper airway ultrasonographic measurements in predicting difficult intubation: A cross-section of the Turkish population
- Prognostic and diagnostic value of circulating IGFBP2 in pancreatic cancer
- Postural stability after operative reconstruction of the AFTL in chronic ankle instability comparing three different surgical techniques
- Research trends related to emergence agitation in the post-anaesthesia care unit from 2001 to 2023: A bibliometric analysis
- Frequency and clinicopathological correlation of gastrointestinal polyps: A six-year single center experience
- ACSL4 mediates inflammatory bowel disease and contributes to LPS-induced intestinal epithelial cell dysfunction by activating ferroptosis and inflammation
- Affibody-based molecular probe 99mTc-(HE)3ZHER2:V2 for non-invasive HER2 detection in ovarian and breast cancer xenografts
- Effectiveness of nutritional support for clinical outcomes in gastric cancer patients: A meta-analysis of randomized controlled trials
- The relationship between IFN-γ, IL-10, IL-6 cytokines, and severity of the condition with serum zinc and Fe in children infected with Mycoplasma pneumoniae
- Paraquat disrupts the blood–brain barrier by increasing IL-6 expression and oxidative stress through the activation of PI3K/AKT signaling pathway
- Sleep quality associate with the increased prevalence of cognitive impairment in coronary artery disease patients: A retrospective case–control study
- Dioscin protects against chronic prostatitis through the TLR4/NF-κB pathway
- Association of polymorphisms in FBN1, MYH11, and TGF-β signaling-related genes with susceptibility of sporadic thoracic aortic aneurysm and dissection in the Zhejiang Han population
- Application value of multi-parameter magnetic resonance image-transrectal ultrasound cognitive fusion in prostate biopsy
- Laboratory variables‐based artificial neural network models for predicting fatty liver disease: A retrospective study
- Decreased BIRC5-206 promotes epithelial–mesenchymal transition in nasopharyngeal carcinoma through sponging miR-145-5p
- Sepsis induces the cardiomyocyte apoptosis and cardiac dysfunction through activation of YAP1/Serpine1/caspase-3 pathway
- Assessment of iron metabolism and iron deficiency in incident patients on incident continuous ambulatory peritoneal dialysis
- Tibial periosteum flap combined with autologous bone grafting in the treatment of Gustilo-IIIB/IIIC open tibial fractures
- The application of intravenous general anesthesia under nasopharyngeal airway assisted ventilation undergoing ureteroscopic holmium laser lithotripsy: A prospective, single-center, controlled trial
- Long intergenic noncoding RNA for IGF2BP2 stability suppresses gastric cancer cell apoptosis by inhibiting the maturation of microRNA-34a
- Role of FOXM1 and AURKB in regulating keratinocyte function in psoriasis
- Parental control attitudes over their pre-school children’s diet
- The role of auto-HSCT in extranodal natural killer/T cell lymphoma
- Significance of negative cervical cytology and positive HPV in the diagnosis of cervical lesions by colposcopy
- Echinacoside inhibits PASMCs calcium overload to prevent hypoxic pulmonary artery remodeling by regulating TRPC1/4/6 and calmodulin
- ADAR1 plays a protective role in proximal tubular cells under high glucose conditions by attenuating the PI3K/AKT/mTOR signaling pathway
- The risk of cancer among insulin glargine users in Lithuania: A retrospective population-based study
- The unusual location of primary hydatid cyst: A case series study
- Intraoperative changes in electrophysiological monitoring can be used to predict clinical outcomes in patients with spinal cavernous malformation
- Obesity and risk of placenta accreta spectrum: A meta-analysis
- Shikonin alleviates asthma phenotypes in mice via an airway epithelial STAT3-dependent mechanism
- NSUN6 and HTR7 disturbed the stability of carotid atherosclerotic plaques by regulating the immune responses of macrophages
- The effect of COVID-19 lockdown on admission rates in Maternity Hospital
- Temporal muscle thickness is not a prognostic predictor in patients with high-grade glioma, an experience at two centers in China
- Luteolin alleviates cerebral ischemia/reperfusion injury by regulating cell pyroptosis
- Therapeutic role of respiratory exercise in patients with tuberculous pleurisy
- Effects of CFTR-ENaC on spinal cord edema after spinal cord injury
- Irisin-regulated lncRNAs and their potential regulatory functions in chondrogenic differentiation of human mesenchymal stem cells
- DMD mutations in pediatric patients with phenotypes of Duchenne/Becker muscular dystrophy
- Combination of C-reactive protein and fibrinogen-to-albumin ratio as a novel predictor of all-cause mortality in heart failure patients
- Significant role and the underly mechanism of cullin-1 in chronic obstructive pulmonary disease
- Ferroptosis-related prognostic model of mantle cell lymphoma
- Observation of choking reaction and other related indexes in elderly painless fiberoptic bronchoscopy with transnasal high-flow humidification oxygen therapy
- A bibliometric analysis of Prader-Willi syndrome from 2002 to 2022
- The causal effects of childhood sunburn occasions on melanoma: A univariable and multivariable Mendelian randomization study
- Oxidative stress regulates glycogen synthase kinase-3 in lymphocytes of diabetes mellitus patients complicated with cerebral infarction
- Role of COX6C and NDUFB3 in septic shock and stroke
- Trends in disease burden of type 2 diabetes, stroke, and hypertensive heart disease attributable to high BMI in China: 1990–2019
- Purinergic P2X7 receptor mediates hyperoxia-induced injury in pulmonary microvascular endothelial cells via NLRP3-mediated pyroptotic pathway
- Investigating the role of oviductal mucosa–endometrial co-culture in modulating factors relevant to embryo implantation
- Analgesic effect of external oblique intercostal block in laparoscopic cholecystectomy: A retrospective study
- Elevated serum miR-142-5p correlates with ischemic lesions and both NSE and S100β in ischemic stroke patients
- Correlation between the mechanism of arteriopathy in IgA nephropathy and blood stasis syndrome: A cohort study
- Risk factors for progressive kyphosis after percutaneous kyphoplasty in osteoporotic vertebral compression fracture
- Predictive role of neuron-specific enolase and S100-β in early neurological deterioration and unfavorable prognosis in patients with ischemic stroke
- The potential risk factors of postoperative cognitive dysfunction for endovascular therapy in acute ischemic stroke with general anesthesia
- Fluoxetine inhibited RANKL-induced osteoclastic differentiation in vitro
- Detection of serum FOXM1 and IGF2 in patients with ARDS and their correlation with disease and prognosis
- Rhein promotes skin wound healing by activating the PI3K/AKT signaling pathway
- Differences in mortality risk by levels of physical activity among persons with disabilities in South Korea
- Review Articles
- Cutaneous signs of selected cardiovascular disorders: A narrative review
- XRCC1 and hOGG1 polymorphisms and endometrial carcinoma: A meta-analysis
- A narrative review on adverse drug reactions of COVID-19 treatments on the kidney
- Emerging role and function of SPDL1 in human health and diseases
- Adverse reactions of piperacillin: A literature review of case reports
- Molecular mechanism and intervention measures of microvascular complications in diabetes
- Regulation of mesenchymal stem cell differentiation by autophagy
- Molecular landscape of borderline ovarian tumours: A systematic review
- Advances in synthetic lethality modalities for glioblastoma multiforme
- Investigating hormesis, aging, and neurodegeneration: From bench to clinics
- Frankincense: A neuronutrient to approach Parkinson’s disease treatment
- Sox9: A potential regulator of cancer stem cells in osteosarcoma
- Early detection of cardiovascular risk markers through non-invasive ultrasound methodologies in periodontitis patients
- Advanced neuroimaging and criminal interrogation in lie detection
- Maternal factors for neural tube defects in offspring: An umbrella review
- The chemoprotective hormetic effects of rosmarinic acid
- CBD’s potential impact on Parkinson’s disease: An updated overview
- Progress in cytokine research for ARDS: A comprehensive review
- Utilizing reactive oxygen species-scavenging nanoparticles for targeting oxidative stress in the treatment of ischemic stroke: A review
- NRXN1-related disorders, attempt to better define clinical assessment
- Lidocaine infusion for the treatment of complex regional pain syndrome: Case series and literature review
- Trends and future directions of autophagy in osteosarcoma: A bibliometric analysis
- Iron in ventricular remodeling and aneurysms post-myocardial infarction
- Case Reports
- Sirolimus potentiated angioedema: A case report and review of the literature
- Identification of mixed anaerobic infections after inguinal hernia repair based on metagenomic next-generation sequencing: A case report
- Successful treatment with bortezomib in combination with dexamethasone in a middle-aged male with idiopathic multicentric Castleman’s disease: A case report
- Complete heart block associated with hepatitis A infection in a female child with fatal outcome
- Elevation of D-dimer in eosinophilic gastrointestinal diseases in the absence of venous thrombosis: A case series and literature review
- Four years of natural progressive course: A rare case report of juvenile Xp11.2 translocations renal cell carcinoma with TFE3 gene fusion
- Advancing prenatal diagnosis: Echocardiographic detection of Scimitar syndrome in China – A case series
- Outcomes and complications of hemodialysis in patients with renal cancer following bilateral nephrectomy
- Anti-HMGCR myopathy mimicking facioscapulohumeral muscular dystrophy
- Recurrent opportunistic infections in a HIV-negative patient with combined C6 and NFKB1 mutations: A case report, pedigree analysis, and literature review
- Letter to the Editor
- Letter to the Editor: Total parenteral nutrition-induced Wernicke’s encephalopathy after oncologic gastrointestinal surgery
- Erratum
- Erratum to “Bladder-embedded ectopic intrauterine device with calculus”
- Retraction
- Retraction of “XRCC1 and hOGG1 polymorphisms and endometrial carcinoma: A meta-analysis”
- Corrigendum
- Corrigendum to “Investigating hormesis, aging, and neurodegeneration: From bench to clinics”
- Corrigendum to “Frankincense: A neuronutrient to approach Parkinson’s disease treatment”
- Special Issue The evolving saga of RNAs from bench to bedside - Part II
- Machine-learning-based prediction of a diagnostic model using autophagy-related genes based on RNA sequencing for patients with papillary thyroid carcinoma
- Unlocking the future of hepatocellular carcinoma treatment: A comprehensive analysis of disulfidptosis-related lncRNAs for prognosis and drug screening
- Elevated mRNA level indicates FSIP1 promotes EMT and gastric cancer progression by regulating fibroblasts in tumor microenvironment
- Special Issue Advancements in oncology: bridging clinical and experimental research - Part I
- Ultrasound-guided transperineal vs transrectal prostate biopsy: A meta-analysis of diagnostic accuracy and complication rates
- Assessment of diagnostic value of unilateral systematic biopsy combined with targeted biopsy in detecting clinically significant prostate cancer
- SENP7 inhibits glioblastoma metastasis and invasion by dissociating SUMO2/3 binding to specific target proteins
- MARK1 suppress malignant progression of hepatocellular carcinoma and improves sorafenib resistance through negatively regulating POTEE
- Analysis of postoperative complications in bladder cancer patients
- Carboplatin combined with arsenic trioxide versus carboplatin combined with docetaxel treatment for LACC: A randomized, open-label, phase II clinical study
- Special Issue Exploring the biological mechanism of human diseases based on MultiOmics Technology - Part I
- Comprehensive pan-cancer investigation of carnosine dipeptidase 1 and its prospective prognostic significance in hepatocellular carcinoma
- Identification of signatures associated with microsatellite instability and immune characteristics to predict the prognostic risk of colon cancer
- Single-cell analysis identified key macrophage subpopulations associated with atherosclerosis