Transcriptome analysis of effects of Tecrl deficiency on cardiometabolic and calcium regulation in cardiac tissue
-
Shujia Lin
 ,
Cuilan Hou
and
Lijian Xie
,
Cuilan Hou
and
Lijian Xie

Abstract
Catecholaminergic polymorphic ventricular tachycardia (CPVT) is a hereditary heart disease characterized by bidirectional or polymorphic ventricular tachycardia and an increased risk of sudden cardiac death. Although trans-2,3-enoyl-CoA reductase like (TECRL) is a newly reported pathogenic gene leading to CPVT that can influence intracellular calcium regulation, the unidentified mechanism underlying the pathogenesis of TECRL deficiency-mediated CPVT remains mainly elusive. In the present study, Tecrl knockout (KO) mice were established and the differentially expressed genes (DEGs) were investigated by RNA-sequencing from the heart tissues. In addition, 857 DEGs were identified in Tecrl KO mice. Subsequently, a weighted gene co-expression network analysis was conducted to discern the pivotal pathways implicated in the Tecrl-mediated regulatory network. Moreover, pathway mapping analyses demonstrated that essential metabolism-related pathways were significantly enriched, notably the fatty acid metabolic process and calcium regulation. Collectively, the data suggested a synergistic relationship between Tecrl deficiency and cardiometabolic and calcium regulation during the development of CPVT. Therefore, further studies on the potential function of TECRL in cardiac tissues would be beneficial to elucidate the pathogenesis of CPVT.
1 Introduction
Sudden death disease resulting from arrhythmia is one of the leading causes of mortality in patients without cardiac structural alterations. Catecholaminergic polymorphic ventricular tachycardia (CPVT) is an inherited ion channel disease that often occurs in prepuberal children (7–12 years old), with an incidence of 1/10,000 [1]. Its prominent clinical feature is bidirectional or polymorphic ventricular tachycardia caused by physical exercise or emotional stress and its first clinical manifestation is syncope or sudden death. CPVT is usually divided into several subtypes, including CPVT1, CPVT2, and CPVT3 and other subtypes [2]. Among them, CPVT1, which accounts for 50–60%, is one of the most common autosomal dominant forms caused by ryanodine receptor 2 (RYR2) mutation. CPVT2, which accounts for 5%, is an autosomal recessive form caused due to calcequestrin 2 (CASQ2) mutation [3–5], and CPVT3, which accounts for 1%, is also an autosomal recessive form caused by trans-2,3-enoyl-CoA reductase like (TECRL) mutation [6]. Our group reported a unique heterozygosity mutation in TECRL gene sequence of a boy diagnosed with CPVT3 and successfully generated a brand new Tecrl knockout (KO) mouse model based on this basis [7,8]. However, the mechanism of CPVT3 caused by TECRL requires further investigation.
TECRL localizes to the endoplasmic reticulum (ER) [9]. Previous studies have reported that TECRL deficiency was accompanied by reduction in the expression levels of RYR2 and CASQ2, leading to aberrant regulation of [Ca2+] i in human pluripotent stem cell-derived cardiomyocytes (hiPSCs-CMs) [6]. TECRL also has an important sequence identity with TECR, which participates in the elongation of very long-chain fatty acids [10]. Previous studies have shown that oxidation disorders of very long-chain fatty acid increase the incidence of delayed depolarizations (DADs) and diastolic [Ca2+] i , leading to the occurrence of arrhythmia [11]. As shown in our previous study, TECRL deficiency results in impaired mitochondrial function in both hearts and hiPSC-CMs [7]. In addition to the regulation of calcium processing, the potential of TECRL to interact with other pathways involved in the regulation of CPVT remains to be determined.
Recently, an increasing number of mouse models harboring CPVT-linked mutations have been established in Ryr2 (R4496C, N2386I, A165D) and Casq2 (D307H, DeltaE9/DeltaE9) [5,12–14]. In agreement with these findings, mouse genetic studies have suggested that these mutations are characterized by stress-induced arrhythmia with typical CPVT phenotypes notably abnormal calcium homeostasis and ultrastructural changes of mitochondria [5,12–14]. TECRL has been considered to be closely associated with [Ca2+] i , with the exception of animal models reported in the previous study conducted by our group [7]. In the past decade, the advancement of bioinformatic analyses utilizing high-throughput sequencing has significantly enhanced our comprehension of the underlying molecular mechanisms of CPVT through the identification of differentially expressed genes (DEGs). It is important to note that the diversity of algorithms utilized in bioinformatic analysis can heavily influence the results. The weighted gene co-expression network analysis (WGCNA) is a method that identifies gene modules with similar expression patterns and disease-related hub genes without the need for DEG analysis. Therefore, to further investigate the potential function of TECRL on cardiac myocytes, we performed RNA-seq on the hearts of Tecrl −/− mice followed by WGCNA. Herein, we reported that Tecrl deficiency created a reliance on fatty acid metabolism and Ca2+ handling. Therefore, TECRL may be useful as a therapeutic target in human CPVT.
2 Materials and methods
2.1 Animal model
Eight-week-old C57BL/6 mice and Tecrl KO mice were obtained from Shanghai Laboratory Animal Center. All mice had unrestricted access to water and food and were housed under 12 h dark–light conditions.
-
Ethics statement: All animal experiments were performed under licenses granted from the Ethics Committee of Experimental Research of Shanghai Children’s Hospital, School of Medicine, Shanghai Jiaotong University. This study was carried out in compliance with the Animal Research: Reporting of In vivo Experiments (ARRIVE) guidelines.
2.2 Western blotting
Heart tissues obtained from 8-week C57BL/6 mice were lysed in RIPA cell lysate buffer supplemented with protease (Roche Diagnostics) and phosphatase inhibitors (Roche Diagnostics); tissue lysates were subsequently centrifuged at 12,000 rpm for 10 min at 4°C. A bicinchoninic acid kit (Applygen Technologies, Inc.) was used to assess the protein concentration. Polyvinylidene fluoride membranes were incubated with the following primary antibodies overnight at 4°C: anti-TECRL (1:1,000; Aviva System Biology, Inc.) and anti-Tubulin (1:1,000; Cell Signaling Technology, Inc.). The secondary antibody (1:2,000; Cell Signaling Technology, Inc.) was diluted and incubated for 1 h at room temperature. An immunology scanner (GS-800, Bio-Rad Laboratories, Inc.) was used to measure the densities of the protein bands following exposure of the membranes to a chemiluminescent substrate (ECL, PerkinElmer, Inc.).
2.3 Sample collection and total RNA isolation
At 8 weeks of age, male and female mice were anesthetized with pentobarbital (100 mg/kg intraperitoneal dose) and subsequently sacrificed by cervical dislocation. Following verification of respiratory and cardiac arrest, animal death was confirmed. For subsequent processing, the heart tissues of each mouse were collected and stored at −80°C. Male and female mice were divided into a wild-type (WT) group and a Tecrl −/− group. Each group contained three mice. TRIzol® (Invitrogen; Thermo Fisher Scientific, Inc.) was added to the heart tissues of each group. Following sonication for three times (15 s each time), isopropyl alcohol was added to the mixture, which was allowed to stand for 10 min at room temperature. Subsequently, the mixture was centrifuged at 12,000 rpm for 10 min at 4°C. The supernatant was collected and washed with 75% alcohol. The final sample was stored at −80°C. RNA purity was assessed using the NanoPhotometer® spectrophotometer (IMPLEN). The RNA concentration was measured using Qubit® RNA Assay Kit in Qubit® 2.0 Fluorometer (Thermo Fisher Scientific, Inc.).
2.4 Library preparation for transcriptome sequencing
RNA for each sample was used at 1.5 µg total amounts as input material for sample preparations. Random hexamer primers were used to synthesize the first strand cDNA and the second strand cDNA synthesis was synthesized using DNA PolymeraseⅠand RNase H. Finally, the library quality was assessed on the Agilent Bioanalyzer 2100 system. The RNA-seq data produced in the course of this investigation have been archived in the Gene Expression Omnibus (GEO) database and the corresponding accession number is GSE191112. The residual data can be found in the article, appendix, or source data file. The source data file is included with this manuscript.
2.5 Gene expression level quantification and differential expression analysis
The cDNA libraries from each mouse were sequenced. The transcript expression levels of each gene were calculated using fragments per kilobase of transcript per million mapped reads (FPKM). The DESeq2 R package (1.26.0) was used to perform differential expression analysis. P < 0.05 and |log2(FoldChange)| ≥0.58 were used to indicate a significant differential expression.
2.6 DEGs identification and functional enrichment analysis
The cDNA libraries from each mouse were sequenced. The transcript expression level of each gene was calculated using FPKM. The DESeq2 R package (1.26.0) was used to perform differential expression analysis. A P value <0.05 and |log2(FoldChange)| ≥0.58 were set as the thresholds for significant differential expression. For functional annotation and classification, DEGs were enriched to the gene ontology (GO) terms based on the DAVID database. GO terms with P values less than 0.05 were considered significantly enriched by DEGs. The enrichment of DEGs in the KEGG pathway was tested using KOBAS v3.0 software. KEGG terms with P values less than 0.05 were considered significantly enriched by DEGs.
2.7 WGCNA
The functional modules in RNA-seq data were clustered using the WGCNA package in R software, with a defined cut-off height of 0.25 for merging similar modules. The co-expression network was inputted with group phenotypes (Tecrl KO vs control, mice gender), and genes with high hub modularity were identified as hub genes in the modular–trait correlation analysis. A heatmap was utilized to visualize the correlation between gene modules and clinical traits, followed by the identification of modules associated with co-expression patterns (midnight blue module and royal blue module) and phenotypes. The appendix material contains the analysis results of both modules.
2.8 Real- time PCR
Total RNA was extracted from mice tissue using TRIzol® reagent (Invitrogen; Thermo Fisher Scientific, Inc.). Complementary DNA was generated from 1,000 ng total RNA using oligo DT and SuperScript III reverse transcriptase (Invitrogen; Thermo Fisher Scientific, Inc.). Real-time PCR was performed using SYBR Green Master Mix (Roche Diagnostics) on a LightCycler 480 Instrument. The primers used are listed in Table A1.
2.9 Data collection from the GEO database
Transcription data of induced pluripotent stem cell-derived cardiomyocytes (iPSC-CMs) derived from patients diagnosed with Brugada Syndrome (BrS) were downloaded from the GEO database (GSE221945). According to the annotation information in the platform, the probes were converted to gene symbols. Four BrS samples (GSM6910336, GSM6910337, GSM6910338, and GSM6910339) and four control samples (GSM6910332, GSM6910333, GSM6910334, and GSM6910335) were identified in the GSE221945 dataset and were used in the present study. The data from GSE221945 were used to validate the function of Tecrl deficiency-related hub genes in fatty acid metabolism and their clinical implication in ventricular arrhythmia. The data procurement and application conformed to the GEO database principles and guidelines. The R packages “Pheatmap” was used to draw the heatmap.
2.10 Data analysis
Western blot parameters are presented as mean ± SEM. Data display was generated in GraphPad Prism 9.0 (GraphPad Software, Inc.). The DESeq2 R package (1.26.0) was used to analyze the differential expression analysis. For four-sample comparison in reverse transcription-quantitative PCR, one-way ANOVA was performed. P < 0.05 was considered to indicate the DEGs that demonstrated statistical significance.
3 Results
3.1 Tecrl expression in various tissues
In the previous study conducted by our group, Tecrl −/− mice were generated and the data indicated that Tecrl deficiency resulted in an abnormal electrocardiographic pattern and disrupted Ca2+ regulation [7]. To determine the tissue expression pattern of TECRL in WT mice, western blots were performed on multiple tissues of male mice. The protein expression analysis by western blots demonstrated that TECRL was ubiquitously expressed in heart, lung, kidney, brain, liver, and skeletal muscle (Figure 1a).
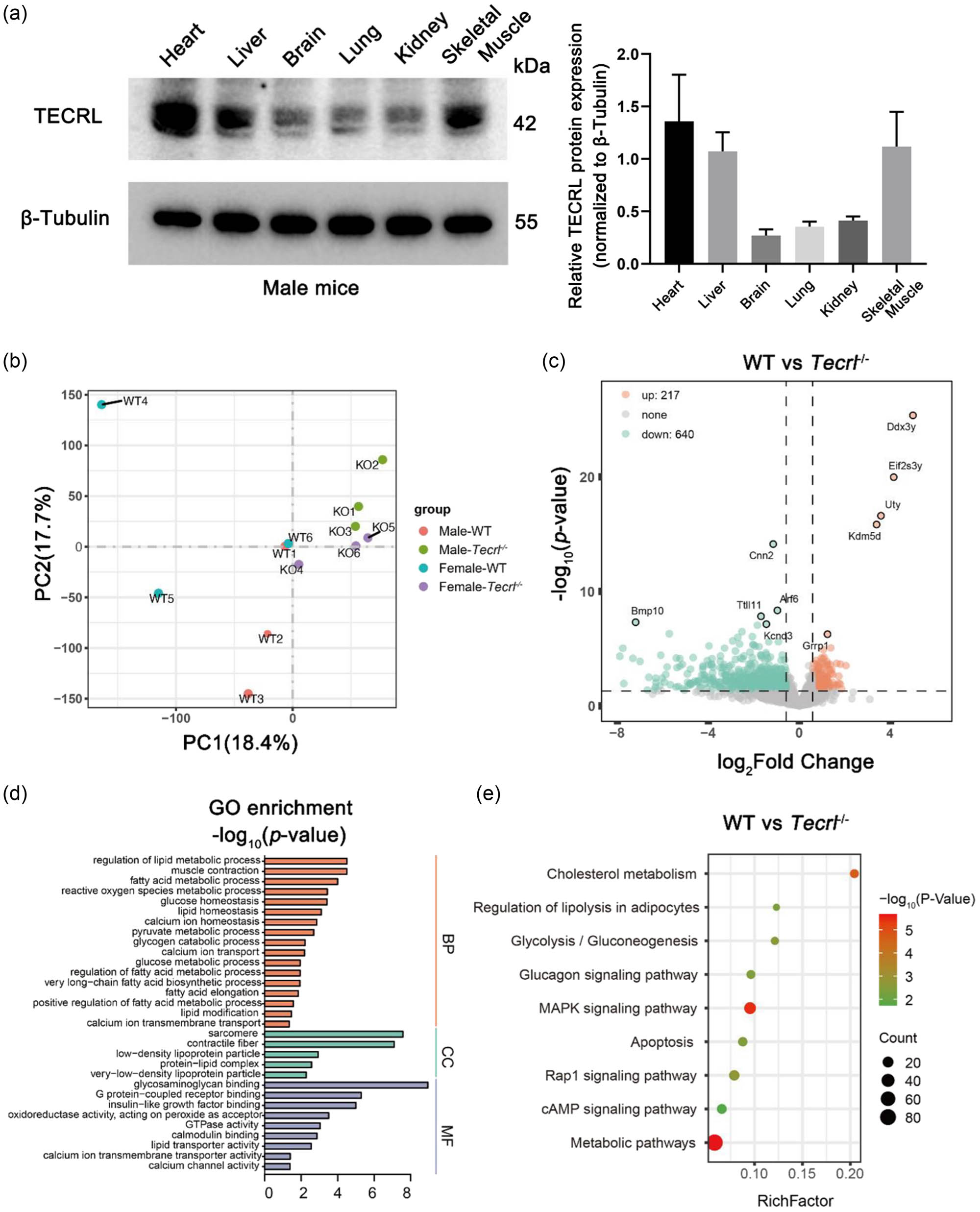
TECRL protein identification and functional enrichment analysis of DEGs. (a) Representative western blots of TECRL in different tissues of WT mice. (b) PCA plot of male and female mice samples. There were four groups, three mice in each group. An average of 5.28 ± 1.37 million raw reads per sample were obtained and PCA shows distinct patterns among groups. (c) Volcano plot of DEGs of male Tecrl −/− mice. Each point represents a single gene. Grey points represent the genes that have no significant changes, while the orange dots indicate the genes that are upregulated and green dots indicate the genes that are downregulated. (|log2(FoldChange)| ≥0.58, P value <0.05). (d) GO enriched terms associated with DEGs in male Tecrl −/− mice. (e) KEGG enriched pathways related to DEGs in male Tecrl −/− mice. BP, biological process; CC, cellular component; MF, molecular function.
3.2 Tecrl deficiency effects on gene expression variation in male and female mice
To further investigate the impact of Tecrl deficiency on the mice, total RNA was extracted from the hearts of Tecrl −/− (male, n = 3; female, n = 3) and WT mice (male, n = 3; female, n = 3). The RNA samples were analyzed by RNA-seq. The transcript expression level of each gene was determined using FPKM. To visualize the pattern of distribution, a principal component analysis was conducted; its value was 40.9% with the first principal component (PC1) = 23.2%, and the second principal component (PC2) = 17.7% (Figure 1b). Deseq2 package in R was used to analyze the DEGs between the different groups. Finally, 640 (74.70%) downregulated and 217 (25.30%) upregulated genes were identified in the Tecrl −/− male mice (Figure 1c).
3.3 GO analysis of Tecrl deficiency in mice
The identified DEGs were categorized into biological processes (BP), cellular components (CC), and molecular functions. The BP of GO analysis revealed that the majority of DEGs were mainly enriched in the pathways related to calcium regulation and metabolism, regulation of lipid metabolic process, muscle contraction, fatty acid metabolism process, glucose homeostasis, lipid homeostasis, calcium ion homeostasis, pyruvate metabolic process, glycogen catabolic process, and calcium ion transport. The CCs mainly consisted of the sarcomere, contractile fiber, low-density lipoprotein particles, protein–lipid complex, and very-low-density lipoprotein particles (Figure 1d).
3.4 Functional enrichment analysis of Tecrl deficiency in mice
KOBAS revealed the main pathways in which differentially expressed RNAs were enriched. In the male groups, KEGG analysis revealed the DEGs that were significantly enriched in 99 pathways. Among the pathways identified in the male Tecrl −/− group, metabolism-related pathways exhibited the highest number of DEGs, which included the metabolic pathway (86 genes), cholesterol metabolism (10 genes), regulation of lipolysis in adipocytes (7 genes), glycolysis/gluconeogenesis (8 genes), the glucagon signaling pathway (10 genes), the MAPK signaling pathway (28 genes), and the cyclic AMP signaling pathway (14 genes), most of which were related to fatty acid metabolism (Figure 1e). Gene set enriched analysis was also used to identify whether fatty acid beta-oxidation and calcium regulation of cardiac cells were related to Tecrl deficiency (Figure 2a and b). Subsequently, the expression levels of genes involved in fatty acid metabolism were shown by heatmap analysis (Figure 2c and d).

Hub gene selection and enrichment analysis: (a) GESA analysis of fatty acid beta-oxidation pathway, (b) GESA analysis of calcium regulation of cardiac cells pathway, (c) heatmap of fatty acid metabolism related genes in Tecrl −/− samples, and (d) heatmap of calcium handling related genes in Tecrl −/− samples.
3.5 Pivotal gene modules identification through WGCNA
To methodically elucidate crucial gene modules and potential mechanisms within the cardiac tissues of Tecrl KO or control mice of both genders, a WGCNA was performed. A hierarchical clustering approach was employed to generate numerous randomly color-coded modules for the cluster dendrogram (Figure 3a). The modular–trait correlation heatmap is presented in Figure 3b. The present study reveals a consistent negative correlation between the Tecrl KO trait and the midnight blue module as well as the royal blue module in mice heart tissue. This suggests that the deficiency of Tecrl results in a downregulated expression of genes associated with these functional modules. Notably, the midnight blue module comprises 74 genes, including CAV3, Rb1cc1, and Ryr2, while the royal blue module consists of 44 genes, including Psmc3, Psmd13, and Gsta4, all of which are hub genes. The shared pathways of these two modules were identified via Metascape (http://metascape.org/). The top GO terms of the hub genes included the following: response to muscle stretch, fatty acid and lipoprotein transport in hepatocytes, carbohydrate derivative catabolic process, and other metabolism-related pathways; the findings suggested the significant alteration of the metabolic process following Tecrl deficiency (Figure 3c and d). An analysis of DisNET (Figure 3e) indicated the participation of cardiovascular dysfunction, specifically left ventricular noncompaction cardiomyopathy, cardiomegaly, and electromyogram abnormality. Additionally, the MCODE analysis demonstrated an enrichment in the regulation of RAS by GAPs, RAF/MAP kinase cascade, negative epigenetic regulation of rRNA expression, and other pathways (Figure 3f). Collectively, the data indicated that the hub genes exhibiting a negative correlation with KO of Tecrl expression in heart tissues may significantly impact the metabolic process, particularly the fatty acid metabolic process, in the context of cardiovascular dysfunction.
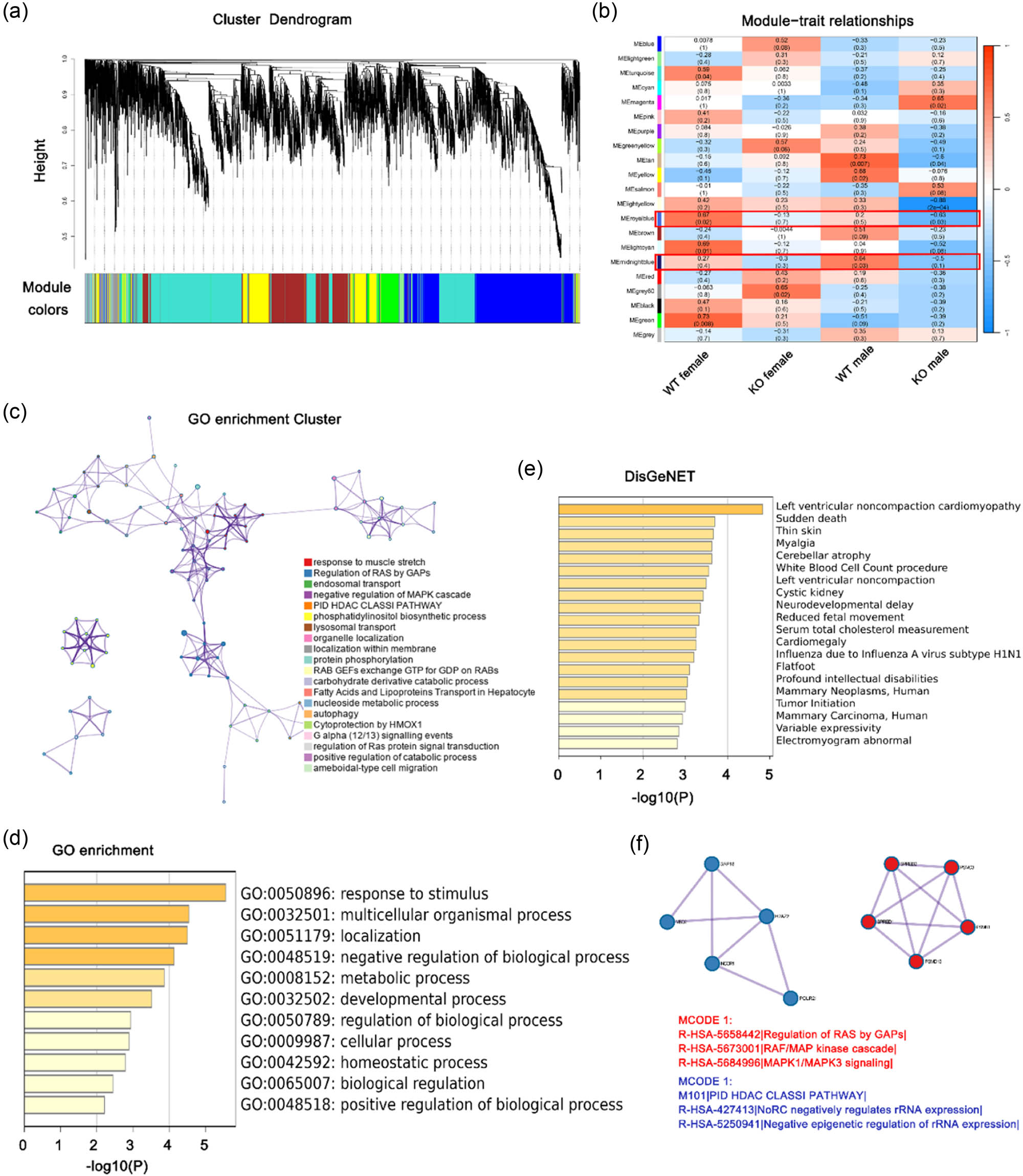
WGCNA revealing gene co-expression networks. (a) Utilizing WGCNA analysis, the dendrogram depicted the clusters of genes that were differentially expressed, based on various metrics. Each branch of the dendrogram represented an individual gene, while the colors beneath the branches represented a co-expression module. (b) The heatmap illustrated the correlation between gene modules and group phenotypes. The correlation coefficient within each cube indicated the degree of correlation between gene modules and traits, with a decreasing gradient from red to blue. (c) The functional enrichment analysis was conducted on Tecrl KO-related hub genes from the midnight blue and royal blue modules. (d) The DisGeNET terms of hub genes were enriched. (e) The hub genes underwent GO enrichment. The horizontal axis depicts the P-value of GO terms on Metascape using default parameters. (f) The top Molecular Complex Detection algorithm (MCODE) terms of hub genes related to Tecrl KO were identified. A network was formed by the protein–protein interactions (PPI) among Tecrl KO-related hub genes from the red module. The MCODE was utilized to detect the connected network components.
3.6 Tecrl deficiency effects on fatty acid metabolism in ventricular arrhythmia
To further explore the role of TECRL deficiency in the fatty acid metabolism and clinical implication in ventricular arrhythmia, the transcription data of iPSC-CMs were analyzed; iPSC-CMs were derived from patients diagnosed with BrS downloaded from GEO (GSE221945). TECRL was expressed at low levels in BrS samples (Figure 4a). Since fatty acid metabolism may play a crucial role in the pathogenesis of Tecrl −/− mice, the correlation of TECRL expression was analyzed with the key components (APOA1, CPT2, UCP2, UCP3) involved in the fatty acid metabolic pathways in BrS. In healthy iPSC-CMs, only UCP3 correlated positively with TECRL (Figure 4b). However, in iPSC-CMs derived from patients with BrS, all four genes were positively correlated with TECRL, which indicated the effects of TECRL on the fatty acid metabolism in patients with ventricular arrhythmia (Figure 4b–e). The transcription data also suggested that apolipoprotein A1 (APOA1) and fatty acid binding protein 5 (FABP5) exhibited a tendency to decrease in BrS, which was consistent with the results obtained from RNA-seq analysis (Figure 4f).

Tecrl deletion leads to significant representative changes. (a) TECRL expression in healthy and BrS patients. (b–e) The graphs of healthy group and Brs group showing a positive correlation of TECRL expression and (b) UCP3, (c) UCP2, (d) CPT2, and (e) APOA1. (f) The heatmap of 48 fatty acid metabolism related genes in BrS. (g) Changes in the mRNA level of Apoa1, Fabp5, and Camk2b.
In the present study, qPCR experiments were performed in WT and Tecrl −/− mice. The mRNA levels of Fabp5 and Apoa1 were significantly decreased in Tecrl −/− mice (Figure 4g). To assess the relationship of arrhythmia to calcium handling, qPCR was performed to determine the mRNA expression levels of Camk2b in male Tecrl −/− mice. The results revealed upregulation of Camk2b expression in Tecrl −/− mice (Figure 4g). In summary, these results indicated that TECRL deficiency was positively correlated to the fatty acid metabolism pathway and played a role in Ca2+ transport.
4 Discussion
Given the elusive pathogenic characteristics of CPVT, it is imperative to comprehend the potential mechanisms and identify innovative therapeutic targets for this ailment. It is established that mutation of the critical pathogenic gene TECRL could lead to CPVT, which is associated with Ca2+ processing and the development of arrhythmia. Our present in vivo transgenic mouse model confirmed that the progression of CPTV derived from Tecrl deficiency is caused by alteration of fatty acid metabolism and Ca2+ processing. Further investigation into the mechanism of TECRL-regulated fatty acid metabolism and the Ca2+ metabolic processing will deepen the understanding of TECRL-induced CPVT.
In our work, 857 DEGs were identified in male Tecrl −/− mice using RNA-seq analysis. An upregulation in the expression levels of Camk2b was noted. GO analysis revealed that DEGs in male and female Tecrl −/− mice were significantly recruited in calcium handling-related pathways such as calcium ion homeostasis and calcium ion transport. Moreover, WGCNA is widely regarded as a superior approach for identifying internal functional modules among key genes. Despite this, there have been few studies utilizing WGCNA to elucidate the underlying mechanisms of CPVT. RNA-seq analysis was used to identify two crucial gene modules that exhibited a consistent negative correlation with the Tecrl KO trait in mouse heart tissues. GO enrichment analysis of these modules revealed significant alterations in metabolic processes following Tecrl KO, indicating that a deficiency in Tecrl led to the downregulation of metabolism-related pathways, ultimately resulting in cardiovascular dysfunction. The top two enriched pathways related to the ion channel in Tecrl −/− mice were the MAPK signaling and the Ras-proximate-1 signaling pathway, suggesting that CPVT may be associated with dysfunction of the ion channel, notably the processing of Ca2+. Calcium/calmodulin-dependent protein kinase II (CaMKII) is one of the isoforms involved in the regulation of intracellular calcium levels [15,16], which plays a pivotal role in the release of Ca2+ from the ER and abnormal sarcoplasmic reticulum (SR) Ca2+ leakiness [17,18]. Inhibition of CaMKII normalized the CPVT phenotype including Ca2+ handling and electrocardiogram (ECG), which indicated that CaMKII played a key role in the incidence of CPVT [17]. In addition, CPVT is a channelopathy caused by the unregulated pathological calcium release [19] in parallel with a high spontaneous Ca2+ release and Ca2+ waves [20], which is consistent with the results presented in the current study. More importantly, Devalla’s group previously revealed that mutation in TECRL in hiPSC-CMs led to abnormalities in intracellular calcium ([Ca2+] i ) transients and increased susceptibility to trigger activity [6]. Our previous study also found that TECRL KO caused the alterations in calcium handling, which indicated that abnormal calcium handling may underly the mechanism by which TECRL deficiency induces CPVT (C. Hou and T. Xiao, unpublished data).
Another previous study reported that RYR2 may decrease glucose oxidation and increase glycolysis by inhibiting pyruvate dehydrogenase, as well as lowering Ca2+ signal amplitude and frequency [21]. Similarly, Santulli et al. found that the leaky RYR2 channels caused the impairment of insulin secretion, resulting in glucose intolerance [22]. Moreover, Li et al. expounded that BMAL1 regulates mitochondrial fission and mitophagy through mitochondrial protein BNIP3 and is critical in the development of dilated cardiomyopathy [23]. In our study, comparative analysis of differential gene expression showed that Tecrl deficiency was enriched in glucose metabolism, such as glucose metabolic process, glucose homeostasis, and glycogen catabolic process. Interestingly, Devalla et al. reported that TECRL deficiency was accompanied by a decrease in RYR2 in hiPSC-CMs [6], indicating that TECRL may have a potential effect on RYR2. Our previous study found that TECRL deficiency decreased the stability of RYR2, which suggested that TECRL deficiency may have an influence on RYR2. This suggested that TECRL played a key role in RYR2 expression, which may explain why DEGs were enriched in the glucose metabolism pathway.
Furthermore, we also observed that the most common pathway related to the DEGs was the fatty acid metabolism pathway in Tecrl −/− mice. And the TECRL was positively correlated with genes involved in the metabolism of fatty acids in BrS patients, which is a disorder also characterized by polymorphic ventricular tachycardia. Among the four genes, UCP2, UCP3, and CPT2 played a crucial role in fatty acid oxidation, it has proven that the downregulation of UCP2 and UCP3 impaired myocardial fatty acid oxidation and elevates the production of reactive oxygen species (ROS), subsequently increasing the incidence of arrhythmia [24,25]. We also identified the downregulation of Apoa1 and Fabp5 in male and female Tecrl −/− mice. Previous study revealed that Apoa1 deficiency results in the abnormal calcium ion transport, while Fabp5 deficiency increases oxidative stress [26,27]. Notably, TECRL has important individual sequences of TECR, which is a protein involved in the synthesis of fatty acids. Long chain fatty acids are responsible for the majority of energy for the normal human heart [28]. Impaired metabolism of fatty acids can contribute to the accumulation of the toxic lipids, which can in turn impair intracellular calcium handling and cardiac contractility [29]. Therefore, impaired metabolism of fatty acids is related to the occurrence of many cardiovascular diseases such as hypertrophic cardiomyopathy, diabetic cardiomyopathy, and arrhythmia [30–32]. Besides, previous study reported that very-long-fatty-acid deficiency may alter the ultrastructure of mitochondria and induce polymorphic ventricular tachycardia [33]. Priori’s group reported that mutation in Ryr2 led to the increased DADs and mitochondrial abnormalities [34]. While increased mitochondrial ROS can stimulate the RYR leak and CaMKII, which is a critical mechanism of the incidence of CPVT [35]. Similarly, in our previous study, we found that Tecrl deficiency impaired the ultrastructure of mitochondria and increased mitochondrial ROS [7]. These indicate that the mechanism by which TECRL causes CPVT may be related to impaired fatty acid metabolism and ultrastructure of mitochondria.
Like all scientific research, our study had some limitations. For example, in sample selection, the lack of human heart tissue of CPVT for clinical validation is a major limitation in our study. Due to the preciousness of the cardiac tissue, we are currently unable to address this issue. In addition, a limited number of studies have been conducted utilizing WGCNA to elucidate the underlying mechanisms of CPVT. Although we have identified two crucial gene modules that exhibited a consistent negative correlation with the Tecrl KO trait in mouse heart tissues by RNA-seq analysis, it requires more validation in the real world. Hence, more work and further validation in multicenter, large-sample cohorts are urgently needed.
5 Conclusion
Our findings suggest a possible association between Tecrl deficiency-mediated CPVT and fatty acid metabolism as well as Ca2+ handling. This study provides a novel perspective on the function of TECRL and its role in CPVT. Further investigation into the protein function of TECRL may offer potential therapeutic targets for the prevention and treatment of CPVT in the future.
Acknowledgements
The authors thank the imaging platform and electron microscope platform staff of Shanghai Institute Precision Medicine, Ninth People’s Hospital, Shanghai Jiao Tong University School of Medicine.
-
Funding information: This work was supported by the National Natural Science Foundation of China (NSFC) (No. 81900437, 82170518), the Shanghai Jiao Tong University medical technology crossing project (YG2021ZD26), the Shanghai Science and Technology Committee (22Y11909700), Shanghai key clinical specialty project (shslczdzk05705), Shanghai Children’s Hospital (2019YN006, 2020YGZQ10), and Jinshan Distinct key medical specialty project (JSZK2023A04). No benefit in any form has been or will be received from a commercial organization directly or indirectly.
-
Author contributions: Cuilan Hou, Tingting Xiao, and Lijian Xie designed and operated the project. Shujia Lin wrote the manuscript with input from Cuilan Hou, Lijian Xie, Shun Chen, and Qiuping Lin. All authors read and approved the final manuscript.
-
Conflict of interest: The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
-
Data availability statement: The datasets for this study can be found in online repositories. The datasets generated during the current study are available in the Gene Expression Omnibus repository, https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE191112 (enter token mjibgkqibliblyh into the box). GSE221945 can be found in the Gene Expression Omnibus repository, https://www.ncbi.nlm.nih.gov/geo/query/acc.cgi?acc=GSE221945.
Appendix
Sequence of the primers used for validation
| Gene name | Forward primer | Reverse primer |
|---|---|---|
| APOA1 | 5′-GTCCCAGTTTGAATCCTCCTCCTTG-3′ | 5′-AGGTTATCCCAGAAGTCCCGAGTC-3′ |
| FABP5 | 5′-GCTAGGAGTAGGACTGGCTCTTAGG-3′ | 5′-TCTTCACTGTGCTCTCGGTTTTGAC-3′ |
| CaMK2b | 5′-CCCTGCCCATCTCCGACTCTC-3′ | 5′-AATGAGCTGCTCTGTGGTCTTGATG-3′ |
| GAPDH | 5′-ACCCAGAAGACTGTGGATGG-3′ | 5′-CACATTGGGGGTAGGAACAC-3′ |
References
[1] Sumitomo N, Harada K, Nagashima M, Yasuda T, Nakamura Y, Aragaki Y, et al. Catecholaminergic polymorphic ventricular tachycardia: electrocardiographic characteristics and optimal therapeutic strategies to prevent sudden death. Heart. 2003;89:66–70.10.1136/heart.89.1.66Search in Google Scholar PubMed PubMed Central
[2] Priori SG, Mazzanti A, Santiago DJ, Kukavica D, Trancuccio A, Kovacic JC. Precision medicine in catecholaminergic polymorphic ventricular tachycardia: JACC focus seminar 5/5. J Am Coll Cardiol. 2021;77(20):2592–612.10.1016/j.jacc.2020.12.073Search in Google Scholar PubMed
[3] Päivi J, Laitinen B, Kevin M, Brown B, Kirsi Piippo M, Heikki Swan M, et al. Mutations of the cardiac ryanodine receptor (RyR2) gene in familial polymorphic ventricular tachycardia. Circulation. 2001;103:485–90.10.1161/01.CIR.103.4.485Search in Google Scholar PubMed
[4] Silvia G, Priori M, Carlo Napolitano M, Natascia Tiso P, Mirella Memmi P, Gabriele Vignati M, et al. Mutations in the cardiac ryanodine receptor gene (hRyR2) underlie catecholaminergic polymorphic ventricular tachycardia. Circulation. 2001;103:196–200.10.1161/01.CIR.103.2.196Search in Google Scholar PubMed
[5] Lahat H, Pras E, Olender T, Avidan N, Ben-Asher E, Man O, et al. A missense mutation in a highly conserved region of CASQ2 is associated with autosomal recessive catecholamine-induced polymorphic ventricular tachycardia in Bedouin families from Israel. Am J Hum Genet. 2001;69(6):1378–84.10.1086/324565Search in Google Scholar PubMed PubMed Central
[6] Devalla HD, Gelinas R, Aburawi EH, Beqqali A, Goyette P, Freund C, et al. TECRL, a new life-threatening inherited arrhythmia gene associated with overlapping clinical features of both LQTS and CPVT. EMBO Mol Med. 2016;8(12):1390–408.10.15252/emmm.201505719Search in Google Scholar PubMed PubMed Central
[7] Hou C, Jiang X, Zhang H, Zheng J, Qiu Q, Zhang Y, et al. TECRL deficiency results in aberrant mitochondrial function in cardiomyocytes. Commun Biol. 2022;5(1):470.10.1038/s42003-022-03414-9Search in Google Scholar PubMed PubMed Central
[8] Xie L, Hou C, Jiang X, Zhao J, Li Y, Xiao T. A compound heterozygosity of Tecrl gene confirmed in a catecholaminergic polymorphic ventricular tachycardia family. Eur J Med Genet. 2019;62(7):103631.10.1016/j.ejmg.2019.01.018Search in Google Scholar PubMed
[9] Moscu-Gregor A, Marschall C, Muntjes C, Schonecker A, Schuessler-Hahn F, Hohendanner F, et al. Novel variants in TECRL cause recessive inherited CPVT type 3 with severe and variable clinical symptoms. J Cardiovasc Electrophysiol. 2020;31(6):1527–35.10.1111/jce.14446Search in Google Scholar PubMed
[10] Moon YA, Horton JD. Identification of two mammalian reductases involved in the two-carbon fatty acyl elongation cascade. J Biol Chem. 2003;278(9):7335–43.10.1074/jbc.M211684200Search in Google Scholar PubMed
[11] Knottnerus SJG, Mengarelli I, Wust RCI, Baartscheer A, Bleeker JC, Coronel R, et al. Electrophysiological abnormalities in VLCAD deficient hiPSC-cardiomyocytes can be improved by lowering accumulation of fatty acid oxidation intermediates. Int J Mol Sci. 2020;21(7):2589.10.3390/ijms21072589Search in Google Scholar PubMed PubMed Central
[12] Kashimura T, Briston SJ, Trafford AW, Napolitano C, Priori SG, Eisner DA, et al. In the RyR2(R4496C) mouse model of CPVT, beta-adrenergic stimulation induces Ca waves by increasing SR Ca content and not by decreasing the threshold for Ca waves. Circ Res. 2010;107(12):1483–9.10.1161/CIRCRESAHA.110.227744Search in Google Scholar PubMed
[13] Shan J, Xie W, Betzenhauser M, Reiken S, Chen BX, Wronska A, et al. Calcium leak through ryanodine receptors leads to atrial fibrillation in 3 mouse models of catecholaminergic polymorphic ventricular tachycardia. Circ Res. 2012;111(6):708–17.10.1161/CIRCRESAHA.112.273342Search in Google Scholar PubMed PubMed Central
[14] Katz G, Shainberg A, Hochhauser E, Kurtzwald-Josefson E, Issac A, El-Ani D, et al. The role of mutant protein level in autosomal recessive catecholamine dependent polymorphic ventricular tachycardia (CPVT2). Biochem Pharmacol. 2013;86(11):1576–83.10.1016/j.bcp.2013.09.012Search in Google Scholar PubMed PubMed Central
[15] Akita T, Aoto K, Kato M, Shiina M, Mutoh H, Nakashima M, et al. De novo variants in CAMK2A and CAMK2B cause neurodevelopmental disorders. Ann Clin Transl Neurol. 2018;5(3):280–96.10.1002/acn3.528Search in Google Scholar PubMed PubMed Central
[16] Beckendorf J, van den Hoogenhof MMG, Backs J. Physiological and unappreciated roles of CaMKII in the heart. Basic Res Cardiol. 2018;113(4):29.10.1007/s00395-018-0688-8Search in Google Scholar PubMed PubMed Central
[17] Bezzerides VJ, Caballero A, Wang S, Ai Y, Hylind RJ, Lu F, et al. Gene therapy for catecholaminergic polymorphic ventricular tachycardia by inhibition of Ca(2+)/calmodulin-dependent kinase II. Circulation. 2019;140(5):405–19.10.1161/CIRCULATIONAHA.118.038514Search in Google Scholar PubMed PubMed Central
[18] Ai X, Curran JW, Shannon TR, Bers DM, Pogwizd SM. Ca2+/calmodulin-dependent protein kinase modulates cardiac ryanodine receptor phosphorylation and sarcoplasmic reticulum Ca2+ leak in heart failure. Circ Res. 2005;97(12):1314–22.10.1161/01.RES.0000194329.41863.89Search in Google Scholar PubMed
[19] Wleklinski MJ, Kannankeril PJ, Knollmann BC. Molecular and tissue mechanisms of catecholaminergic polymorphic ventricular tachycardia. J Physiol. 2020;598(14):2817–34.10.1113/JP276757Search in Google Scholar PubMed PubMed Central
[20] Fernández-Velasco M, Rueda A, Rizzi N, Benitah JP, Colombi B, Napolitano C, et al. Increased Ca2+ sensitivity of the ryanodine receptor mutant RyR2 R4496C underlies catecholaminergic polymorphic ventricular tachycardia. Circulation Res. 200;104(2):201–9.10.1161/CIRCRESAHA.108.177493Search in Google Scholar PubMed PubMed Central
[21] Bround MJ, Wambolt R, Cen H, Asghari P, Albu RF, Han J, et al. Cardiac ryanodine receptor (Ryr2)-mediated calcium signals specifically promote glucose oxidation via pyruvate dehydrogenase. J Biol Chem. 2016;291(45):23490–505.10.1074/jbc.M116.756973Search in Google Scholar PubMed PubMed Central
[22] Santulli G, Pagano G, Sardu C, Xie W, Reiken S, D’Ascia SL, et al. Calcium release channel RyR2 regulates insulin release and glucose homeostasis. J Clin Invest. 2015;125(5):1968–78.10.1172/JCI79273Search in Google Scholar PubMed PubMed Central
[23] Li E, Li X, Huang J, Xu C, Liang Q, Ren K, et al. BMAL1 regulates mitochondrial fission and mitophagy through mitochondrial protein BNIP3 and is critical in the development of dilated cardiomyopathy. Protein Cell. 2020;11(9):661–79.10.1007/s13238-020-00713-xSearch in Google Scholar PubMed PubMed Central
[24] Larbig R, Reda S, Paar V, Trost A, Leitner J, Weichselbaumer S, et al. Through modulation of cardiac Ca(2+) handling, UCP2 affects cardiac electrophysiology and influences the susceptibility for Ca(2+)-mediated arrhythmias. Exp Physiol. 2017;102(6):650–2.10.1113/EP086209Search in Google Scholar PubMed
[25] Edwards KS, Ashraf S, Lomax TM, Wiseman JM, Hall ME, Gava FN, et al. Uncoupling protein 3 deficiency impairs myocardial fatty acid oxidation and contractile recovery following ischemia/reperfusion. Basic Res Cardiol. 2018;113(6):47.10.1007/s00395-018-0707-9Search in Google Scholar PubMed PubMed Central
[26] Baartscheer A, Schumacher CA, Wekker V, Verkerk AO, Veldkamp MW, van Oort RJ, et al. Dyscholesterolemia protects against ischemia-induced ventricular arrhythmias. Circ Arrhythm Electrophysiol. 2015;8(6):1481–90.10.1161/CIRCEP.115.002751Search in Google Scholar PubMed
[27] Gao S, Li G, Shao Y, Wei Z, Huang S, Qi F, et al. FABP5 deficiency impairs mitochondrial function and aggravates pathological cardiac remodeling and dysfunction. Cardiovasc Toxicol. 2021;21(8):619–29.10.1007/s12012-021-09653-2Search in Google Scholar PubMed
[28] Lopaschuk GD. Metabolic modulators in heart disease: past, present, and future. Can J Cardiol. 2017;33(7):838–49.10.1016/j.cjca.2016.12.013Search in Google Scholar PubMed
[29] Carpentier AC. Abnormal myocardial dietary fatty acid metabolism and diabetic cardiomyopathy. Can J Cardiol. 2018;34(5):605–14.10.1016/j.cjca.2017.12.029Search in Google Scholar PubMed
[30] Zheng F, Xiao Y, Liu H, Fan Y, Dao M. Patient-specific organoid and organ-on-a-chip: 3D cell-culture meets 3D printing and numerical simulation. Adv Biol (Weinh). 2021;e2000024.10.1002/adbi.202000024Search in Google Scholar PubMed PubMed Central
[31] Aubert G, Martin OJ, Horton JL, Lai L, Vega RB, Leone TC, et al. The failing heart relies on ketone bodies as a fuel. Circulation. 2016;133(8):698–705.10.1161/CIRCULATIONAHA.115.017355Search in Google Scholar PubMed PubMed Central
[32] Nakamura M, Liu T, Husain S, Zhai P, Warren JS, Hsu CP, et al. Glycogen synthase kinase-3alpha promotes fatty acid uptake and lipotoxic cardiomyopathy. Cell Metab. 2019;29(5):1119–34 e12.10.1016/j.cmet.2019.01.005Search in Google Scholar PubMed PubMed Central
[33] Exil VJ, Roberts RL, Sims H, McLaughlin JE, Malkin RA, Gardner CD, et al. Very-long-chain acyl-coenzyme a dehydrogenase deficiency in mice. Circ Res. 2003;93(5):448–55.10.1161/01.RES.0000088786.19197.E4Search in Google Scholar PubMed
[34] Bongianino R, Denegri M, Mazzanti A, Lodola F, Vollero A, Boncompagni S, et al. Allele-specific silencing of mutant mRNA rescues ultrastructural and arrhythmic phenotype in mice carriers of the R4496C mutation in the ryanodine receptor gene (RYR2). Circ Res. 2017;121(5):525–36.10.1161/CIRCRESAHA.117.310882Search in Google Scholar PubMed
[35] Hegyi B, Polonen RP, Hellgren KT, Ko CY, Ginsburg KS, Bossuyt J, et al. Cardiomyocyte Na(+) and Ca(2+) mishandling drives vicious cycle involving CaMKII, ROS, and ryanodine receptors. Basic Res Cardiol. 2021;116(1):58.10.1007/s00395-021-00900-9Search in Google Scholar PubMed PubMed Central
© 2024 the author(s), published by De Gruyter
This work is licensed under the Creative Commons Attribution 4.0 International License.
Articles in the same Issue
- Research Articles
- EDNRB inhibits the growth and migration of prostate cancer cells by activating the cGMP-PKG pathway
- STK11 (LKB1) mutation suppresses ferroptosis in lung adenocarcinoma by facilitating monounsaturated fatty acid synthesis
- Association of SOX6 gene polymorphisms with Kashin-Beck disease risk in the Chinese Han population
- The pyroptosis-related signature predicts prognosis and influences the tumor immune microenvironment in dedifferentiated liposarcoma
- METTL3 attenuates ferroptosis sensitivity in lung cancer via modulating TFRC
- Identification and validation of molecular subtypes and prognostic signature for stage I and stage II gastric cancer based on neutrophil extracellular traps
- Novel lumbar plexus block versus femoral nerve block for analgesia and motor recovery after total knee arthroplasty
- Correlation between ABCB1 and OLIG2 polymorphisms and the severity and prognosis of patients with cerebral infarction
- Study on the radiotherapy effect and serum neutral granulocyte lymphocyte ratio and inflammatory factor expression of nasopharyngeal carcinoma
- Transcriptome analysis of effects of Tecrl deficiency on cardiometabolic and calcium regulation in cardiac tissue
- Aflatoxin B1 induces infertility, fetal deformities, and potential therapies
- Serum levels of HMW adiponectin and its receptors are associated with cytokine levels and clinical characteristics in chronic obstructive pulmonary disease
- METTL3-mediated methylation of CYP2C19 mRNA may aggravate clopidogrel resistance in ischemic stroke patients
- Understand how machine learning impact lung cancer research from 2010 to 2021: A bibliometric analysis
- Pressure ulcers in German hospitals: Analysis of reimbursement and length of stay
- Metformin plus L-carnitine enhances brown/beige adipose tissue activity via Nrf2/HO-1 signaling to reduce lipid accumulation and inflammation in murine obesity
- Downregulation of carbonic anhydrase IX expression in mouse xenograft nasopharyngeal carcinoma model via doxorubicin nanobubble combined with ultrasound
- Feasibility of 3-dimensional printed models in simulated training and teaching of transcatheter aortic valve replacement
- miR-335-3p improves type II diabetes mellitus by IGF-1 regulating macrophage polarization
- The analyses of human MCPH1 DNA repair machinery and genetic variations
- Activation of Piezo1 increases the sensitivity of breast cancer to hyperthermia therapy
- Comprehensive analysis based on the disulfidptosis-related genes identifies hub genes and immune infiltration for pancreatic adenocarcinoma
- Changes of serum CA125 and PGE2 before and after high-intensity focused ultrasound combined with GnRH-a in treatment of patients with adenomyosis
- The clinical value of the hepatic venous pressure gradient in patients undergoing hepatic resection for hepatocellular carcinoma with or without liver cirrhosis
- Development and validation of a novel model to predict pulmonary embolism in cardiology suspected patients: A 10-year retrospective analysis
- Downregulation of lncRNA XLOC_032768 in diabetic patients predicts the occurrence of diabetic nephropathy
- Circ_0051428 targeting miR-885-3p/MMP2 axis enhances the malignancy of cervical cancer
- Effectiveness of ginkgo diterpene lactone meglumine on cognitive function in patients with acute ischemic stroke
- The construction of a novel prognostic prediction model for glioma based on GWAS-identified prognostic-related risk loci
- Evaluating the impact of childhood BMI on the risk of coronavirus disease 2019: A Mendelian randomization study
- Lactate dehydrogenase to albumin ratio is associated with in-hospital mortality in patients with acute heart failure: Data from the MIMIC-III database
- CD36-mediated podocyte lipotoxicity promotes foot process effacement
- Efficacy of etonogestrel subcutaneous implants versus the levonorgestrel-releasing intrauterine system in the conservative treatment of adenomyosis
- FLRT2 mediates chondrogenesis of nasal septal cartilage and mandibular condyle cartilage
- Challenges in treating primary immune thrombocytopenia patients undergoing COVID-19 vaccination: A retrospective study
- Let-7 family regulates HaCaT cell proliferation and apoptosis via the ΔNp63/PI3K/AKT pathway
- Phospholipid transfer protein ameliorates sepsis-induced cardiac dysfunction through NLRP3 inflammasome inhibition
- Postoperative cognitive dysfunction in elderly patients with colorectal cancer: A randomized controlled study comparing goal-directed and conventional fluid therapy
- Long-pulsed ultrasound-mediated microbubble thrombolysis in a rat model of microvascular obstruction
- High SEC61A1 expression predicts poor outcome of acute myeloid leukemia
- Comparison of polymerase chain reaction and next-generation sequencing with conventional urine culture for the diagnosis of urinary tract infections: A meta-analysis
- Secreted frizzled-related protein 5 protects against renal fibrosis by inhibiting Wnt/β-catenin pathway
- Pan-cancer and single-cell analysis of actin cytoskeleton genes related to disulfidptosis
- Overexpression of miR-532-5p restrains oxidative stress response of chondrocytes in nontraumatic osteonecrosis of the femoral head by inhibiting ABL1
- Autologous liver transplantation for unresectable hepatobiliary malignancies in enhanced recovery after surgery model
- Clinical analysis of incomplete rupture of the uterus secondary to previous cesarean section
- Abnormal sleep duration is associated with sarcopenia in older Chinese people: A large retrospective cross-sectional study
- No genetic causality between obesity and benign paroxysmal vertigo: A two-sample Mendelian randomization study
- Identification and validation of autophagy-related genes in SSc
- Long non-coding RNA SRA1 suppresses radiotherapy resistance in esophageal squamous cell carcinoma by modulating glycolytic reprogramming
- Evaluation of quality of life in patients with schizophrenia: An inpatient social welfare institution-based cross-sectional study
- The possible role of oxidative stress marker glutathione in the assessment of cognitive impairment in multiple sclerosis
- Compilation of a self-management assessment scale for postoperative patients with aortic dissection
- Left atrial appendage closure in conjunction with radiofrequency ablation: Effects on left atrial functioning in patients with paroxysmal atrial fibrillation
- Effect of anterior femoral cortical notch grade on postoperative function and complications during TKA surgery: A multicenter, retrospective study
- Clinical characteristics and assessment of risk factors in patients with influenza A-induced severe pneumonia after the prevalence of SARS-CoV-2
- Analgesia nociception index is an indicator of laparoscopic trocar insertion-induced transient nociceptive stimuli
- High STAT4 expression correlates with poor prognosis in acute myeloid leukemia and facilitates disease progression by upregulating VEGFA expression
- Factors influencing cardiovascular system-related post-COVID-19 sequelae: A single-center cohort study
- HOXD10 regulates intestinal permeability and inhibits inflammation of dextran sulfate sodium-induced ulcerative colitis through the inactivation of the Rho/ROCK/MMPs axis
- Mesenchymal stem cell-derived exosomal miR-26a induces ferroptosis, suppresses hepatic stellate cell activation, and ameliorates liver fibrosis by modulating SLC7A11
- Endovascular thrombectomy versus intravenous thrombolysis for primary distal, medium vessel occlusion in acute ischemic stroke
- ANO6 (TMEM16F) inhibits gastrointestinal stromal tumor growth and induces ferroptosis
- Prognostic value of EIF5A2 in solid tumors: A meta-analysis and bioinformatics analysis
- The role of enhanced expression of Cx43 in patients with ulcerative colitis
- Choosing a COVID-19 vaccination site might be driven by anxiety and body vigilance
- Role of ICAM-1 in triple-negative breast cancer
- Cost-effectiveness of ambroxol in the treatment of Gaucher disease type 2
- HLA-DRB5 promotes immune thrombocytopenia via activating CD8+ T cells
- Efficacy and factors of myofascial release therapy combined with electrical and magnetic stimulation in the treatment of chronic pelvic pain syndrome
- Efficacy of tacrolimus monotherapy in primary membranous nephropathy
- Mechanisms of Tripterygium wilfordii Hook F on treating rheumatoid arthritis explored by network pharmacology analysis and molecular docking
- FBXO45 levels regulated ferroptosis renal tubular epithelial cells in a model of diabetic nephropathy by PLK1
- Optimizing anesthesia strategies to NSCLC patients in VATS procedures: Insights from drug requirements and patient recovery patterns
- Alpha-lipoic acid upregulates the PPARγ/NRF2/GPX4 signal pathway to inhibit ferroptosis in the pathogenesis of unexplained recurrent pregnancy loss
- Correlation between fat-soluble vitamin levels and inflammatory factors in paediatric community-acquired pneumonia: A prospective study
- CD1d affects the proliferation, migration, and apoptosis of human papillary thyroid carcinoma TPC-1 cells via regulating MAPK/NF-κB signaling pathway
- miR-let-7a inhibits sympathetic nerve remodeling after myocardial infarction by downregulating the expression of nerve growth factor
- Immune response analysis of solid organ transplantation recipients inoculated with inactivated COVID-19 vaccine: A retrospective analysis
- The H2Valdien derivatives regulate the epithelial–mesenchymal transition of hepatoma carcinoma cells through the Hedgehog signaling pathway
- Clinical efficacy of dexamethasone combined with isoniazid in the treatment of tuberculous meningitis and its effect on peripheral blood T cell subsets
- Comparison of short-segment and long-segment fixation in treatment of degenerative scoliosis and analysis of factors associated with adjacent spondylolisthesis
- Lycopene inhibits pyroptosis of endothelial progenitor cells induced by ox-LDL through the AMPK/mTOR/NLRP3 pathway
- Methylation regulation for FUNDC1 stability in childhood leukemia was up-regulated and facilitates metastasis and reduces ferroptosis of leukemia through mitochondrial damage by FBXL2
- Correlation of single-fiber electromyography studies and functional status in patients with amyotrophic lateral sclerosis
- Risk factors of postoperative airway obstruction complications in children with oral floor mass
- Expression levels and clinical significance of serum miR-19a/CCL20 in patients with acute cerebral infarction
- Physical activity and mental health trends in Korean adolescents: Analyzing the impact of the COVID-19 pandemic from 2018 to 2022
- Evaluating anemia in HIV-infected patients using chest CT
- Ponticulus posticus and skeletal malocclusion: A pilot study in a Southern Italian pre-orthodontic court
- Causal association of circulating immune cells and lymphoma: A Mendelian randomization study
- Assessment of the renal function and fibrosis indexes of conventional western medicine with Chinese medicine for dredging collaterals on treating renal fibrosis: A systematic review and meta-analysis
- Comprehensive landscape of integrator complex subunits and their association with prognosis and tumor microenvironment in gastric cancer
- New target-HMGCR inhibitors for the treatment of primary sclerosing cholangitis: A drug Mendelian randomization study
- Population pharmacokinetics of meropenem in critically ill patients
- Comparison of the ability of newly inflammatory markers to predict complicated appendicitis
- Comparative morphology of the cruciate ligaments: A radiological study
- Immune landscape of hepatocellular carcinoma: The central role of TP53-inducible glycolysis and apoptosis regulator
- Serum SIRT3 levels in epilepsy patients and its association with clinical outcomes and severity: A prospective observational study
- SHP-1 mediates cigarette smoke extract-induced epithelial–mesenchymal transformation and inflammation in 16HBE cells
- Acute hyper-hypoxia accelerates the development of depression in mice via the IL-6/PGC1α/MFN2 signaling pathway
- The GJB3 correlates with the prognosis, immune cell infiltration, and therapeutic responses in lung adenocarcinoma
- Physical fitness and blood parameters outcomes of breast cancer survivor in a low-intensity circuit resistance exercise program
- Exploring anesthetic-induced gene expression changes and immune cell dynamics in atrial tissue post-coronary artery bypass graft surgery
- Empagliflozin improves aortic injury in obese mice by regulating fatty acid metabolism
- Analysis of the risk factors of the radiation-induced encephalopathy in nasopharyngeal carcinoma: A retrospective cohort study
- Reproductive outcomes in women with BRCA 1/2 germline mutations: A retrospective observational study and literature review
- Evaluation of upper airway ultrasonographic measurements in predicting difficult intubation: A cross-section of the Turkish population
- Prognostic and diagnostic value of circulating IGFBP2 in pancreatic cancer
- Postural stability after operative reconstruction of the AFTL in chronic ankle instability comparing three different surgical techniques
- Research trends related to emergence agitation in the post-anaesthesia care unit from 2001 to 2023: A bibliometric analysis
- Frequency and clinicopathological correlation of gastrointestinal polyps: A six-year single center experience
- ACSL4 mediates inflammatory bowel disease and contributes to LPS-induced intestinal epithelial cell dysfunction by activating ferroptosis and inflammation
- Affibody-based molecular probe 99mTc-(HE)3ZHER2:V2 for non-invasive HER2 detection in ovarian and breast cancer xenografts
- Effectiveness of nutritional support for clinical outcomes in gastric cancer patients: A meta-analysis of randomized controlled trials
- The relationship between IFN-γ, IL-10, IL-6 cytokines, and severity of the condition with serum zinc and Fe in children infected with Mycoplasma pneumoniae
- Paraquat disrupts the blood–brain barrier by increasing IL-6 expression and oxidative stress through the activation of PI3K/AKT signaling pathway
- Sleep quality associate with the increased prevalence of cognitive impairment in coronary artery disease patients: A retrospective case–control study
- Dioscin protects against chronic prostatitis through the TLR4/NF-κB pathway
- Association of polymorphisms in FBN1, MYH11, and TGF-β signaling-related genes with susceptibility of sporadic thoracic aortic aneurysm and dissection in the Zhejiang Han population
- Application value of multi-parameter magnetic resonance image-transrectal ultrasound cognitive fusion in prostate biopsy
- Laboratory variables‐based artificial neural network models for predicting fatty liver disease: A retrospective study
- Decreased BIRC5-206 promotes epithelial–mesenchymal transition in nasopharyngeal carcinoma through sponging miR-145-5p
- Sepsis induces the cardiomyocyte apoptosis and cardiac dysfunction through activation of YAP1/Serpine1/caspase-3 pathway
- Assessment of iron metabolism and iron deficiency in incident patients on incident continuous ambulatory peritoneal dialysis
- Tibial periosteum flap combined with autologous bone grafting in the treatment of Gustilo-IIIB/IIIC open tibial fractures
- The application of intravenous general anesthesia under nasopharyngeal airway assisted ventilation undergoing ureteroscopic holmium laser lithotripsy: A prospective, single-center, controlled trial
- Long intergenic noncoding RNA for IGF2BP2 stability suppresses gastric cancer cell apoptosis by inhibiting the maturation of microRNA-34a
- Role of FOXM1 and AURKB in regulating keratinocyte function in psoriasis
- Parental control attitudes over their pre-school children’s diet
- The role of auto-HSCT in extranodal natural killer/T cell lymphoma
- Significance of negative cervical cytology and positive HPV in the diagnosis of cervical lesions by colposcopy
- Echinacoside inhibits PASMCs calcium overload to prevent hypoxic pulmonary artery remodeling by regulating TRPC1/4/6 and calmodulin
- ADAR1 plays a protective role in proximal tubular cells under high glucose conditions by attenuating the PI3K/AKT/mTOR signaling pathway
- The risk of cancer among insulin glargine users in Lithuania: A retrospective population-based study
- The unusual location of primary hydatid cyst: A case series study
- Intraoperative changes in electrophysiological monitoring can be used to predict clinical outcomes in patients with spinal cavernous malformation
- Obesity and risk of placenta accreta spectrum: A meta-analysis
- Shikonin alleviates asthma phenotypes in mice via an airway epithelial STAT3-dependent mechanism
- NSUN6 and HTR7 disturbed the stability of carotid atherosclerotic plaques by regulating the immune responses of macrophages
- The effect of COVID-19 lockdown on admission rates in Maternity Hospital
- Temporal muscle thickness is not a prognostic predictor in patients with high-grade glioma, an experience at two centers in China
- Luteolin alleviates cerebral ischemia/reperfusion injury by regulating cell pyroptosis
- Therapeutic role of respiratory exercise in patients with tuberculous pleurisy
- Effects of CFTR-ENaC on spinal cord edema after spinal cord injury
- Irisin-regulated lncRNAs and their potential regulatory functions in chondrogenic differentiation of human mesenchymal stem cells
- DMD mutations in pediatric patients with phenotypes of Duchenne/Becker muscular dystrophy
- Combination of C-reactive protein and fibrinogen-to-albumin ratio as a novel predictor of all-cause mortality in heart failure patients
- Significant role and the underly mechanism of cullin-1 in chronic obstructive pulmonary disease
- Ferroptosis-related prognostic model of mantle cell lymphoma
- Observation of choking reaction and other related indexes in elderly painless fiberoptic bronchoscopy with transnasal high-flow humidification oxygen therapy
- A bibliometric analysis of Prader-Willi syndrome from 2002 to 2022
- The causal effects of childhood sunburn occasions on melanoma: A univariable and multivariable Mendelian randomization study
- Oxidative stress regulates glycogen synthase kinase-3 in lymphocytes of diabetes mellitus patients complicated with cerebral infarction
- Role of COX6C and NDUFB3 in septic shock and stroke
- Trends in disease burden of type 2 diabetes, stroke, and hypertensive heart disease attributable to high BMI in China: 1990–2019
- Purinergic P2X7 receptor mediates hyperoxia-induced injury in pulmonary microvascular endothelial cells via NLRP3-mediated pyroptotic pathway
- Investigating the role of oviductal mucosa–endometrial co-culture in modulating factors relevant to embryo implantation
- Analgesic effect of external oblique intercostal block in laparoscopic cholecystectomy: A retrospective study
- Elevated serum miR-142-5p correlates with ischemic lesions and both NSE and S100β in ischemic stroke patients
- Correlation between the mechanism of arteriopathy in IgA nephropathy and blood stasis syndrome: A cohort study
- Risk factors for progressive kyphosis after percutaneous kyphoplasty in osteoporotic vertebral compression fracture
- Predictive role of neuron-specific enolase and S100-β in early neurological deterioration and unfavorable prognosis in patients with ischemic stroke
- The potential risk factors of postoperative cognitive dysfunction for endovascular therapy in acute ischemic stroke with general anesthesia
- Fluoxetine inhibited RANKL-induced osteoclastic differentiation in vitro
- Detection of serum FOXM1 and IGF2 in patients with ARDS and their correlation with disease and prognosis
- Rhein promotes skin wound healing by activating the PI3K/AKT signaling pathway
- Differences in mortality risk by levels of physical activity among persons with disabilities in South Korea
- Review Articles
- Cutaneous signs of selected cardiovascular disorders: A narrative review
- XRCC1 and hOGG1 polymorphisms and endometrial carcinoma: A meta-analysis
- A narrative review on adverse drug reactions of COVID-19 treatments on the kidney
- Emerging role and function of SPDL1 in human health and diseases
- Adverse reactions of piperacillin: A literature review of case reports
- Molecular mechanism and intervention measures of microvascular complications in diabetes
- Regulation of mesenchymal stem cell differentiation by autophagy
- Molecular landscape of borderline ovarian tumours: A systematic review
- Advances in synthetic lethality modalities for glioblastoma multiforme
- Investigating hormesis, aging, and neurodegeneration: From bench to clinics
- Frankincense: A neuronutrient to approach Parkinson’s disease treatment
- Sox9: A potential regulator of cancer stem cells in osteosarcoma
- Early detection of cardiovascular risk markers through non-invasive ultrasound methodologies in periodontitis patients
- Advanced neuroimaging and criminal interrogation in lie detection
- Maternal factors for neural tube defects in offspring: An umbrella review
- The chemoprotective hormetic effects of rosmarinic acid
- CBD’s potential impact on Parkinson’s disease: An updated overview
- Progress in cytokine research for ARDS: A comprehensive review
- Utilizing reactive oxygen species-scavenging nanoparticles for targeting oxidative stress in the treatment of ischemic stroke: A review
- NRXN1-related disorders, attempt to better define clinical assessment
- Lidocaine infusion for the treatment of complex regional pain syndrome: Case series and literature review
- Trends and future directions of autophagy in osteosarcoma: A bibliometric analysis
- Iron in ventricular remodeling and aneurysms post-myocardial infarction
- Case Reports
- Sirolimus potentiated angioedema: A case report and review of the literature
- Identification of mixed anaerobic infections after inguinal hernia repair based on metagenomic next-generation sequencing: A case report
- Successful treatment with bortezomib in combination with dexamethasone in a middle-aged male with idiopathic multicentric Castleman’s disease: A case report
- Complete heart block associated with hepatitis A infection in a female child with fatal outcome
- Elevation of D-dimer in eosinophilic gastrointestinal diseases in the absence of venous thrombosis: A case series and literature review
- Four years of natural progressive course: A rare case report of juvenile Xp11.2 translocations renal cell carcinoma with TFE3 gene fusion
- Advancing prenatal diagnosis: Echocardiographic detection of Scimitar syndrome in China – A case series
- Outcomes and complications of hemodialysis in patients with renal cancer following bilateral nephrectomy
- Anti-HMGCR myopathy mimicking facioscapulohumeral muscular dystrophy
- Recurrent opportunistic infections in a HIV-negative patient with combined C6 and NFKB1 mutations: A case report, pedigree analysis, and literature review
- Letter to the Editor
- Letter to the Editor: Total parenteral nutrition-induced Wernicke’s encephalopathy after oncologic gastrointestinal surgery
- Erratum
- Erratum to “Bladder-embedded ectopic intrauterine device with calculus”
- Retraction
- Retraction of “XRCC1 and hOGG1 polymorphisms and endometrial carcinoma: A meta-analysis”
- Corrigendum
- Corrigendum to “Investigating hormesis, aging, and neurodegeneration: From bench to clinics”
- Corrigendum to “Frankincense: A neuronutrient to approach Parkinson’s disease treatment”
- Special Issue The evolving saga of RNAs from bench to bedside - Part II
- Machine-learning-based prediction of a diagnostic model using autophagy-related genes based on RNA sequencing for patients with papillary thyroid carcinoma
- Unlocking the future of hepatocellular carcinoma treatment: A comprehensive analysis of disulfidptosis-related lncRNAs for prognosis and drug screening
- Elevated mRNA level indicates FSIP1 promotes EMT and gastric cancer progression by regulating fibroblasts in tumor microenvironment
- Special Issue Advancements in oncology: bridging clinical and experimental research - Part I
- Ultrasound-guided transperineal vs transrectal prostate biopsy: A meta-analysis of diagnostic accuracy and complication rates
- Assessment of diagnostic value of unilateral systematic biopsy combined with targeted biopsy in detecting clinically significant prostate cancer
- SENP7 inhibits glioblastoma metastasis and invasion by dissociating SUMO2/3 binding to specific target proteins
- MARK1 suppress malignant progression of hepatocellular carcinoma and improves sorafenib resistance through negatively regulating POTEE
- Analysis of postoperative complications in bladder cancer patients
- Carboplatin combined with arsenic trioxide versus carboplatin combined with docetaxel treatment for LACC: A randomized, open-label, phase II clinical study
- Special Issue Exploring the biological mechanism of human diseases based on MultiOmics Technology - Part I
- Comprehensive pan-cancer investigation of carnosine dipeptidase 1 and its prospective prognostic significance in hepatocellular carcinoma
- Identification of signatures associated with microsatellite instability and immune characteristics to predict the prognostic risk of colon cancer
- Single-cell analysis identified key macrophage subpopulations associated with atherosclerosis
Articles in the same Issue
- Research Articles
- EDNRB inhibits the growth and migration of prostate cancer cells by activating the cGMP-PKG pathway
- STK11 (LKB1) mutation suppresses ferroptosis in lung adenocarcinoma by facilitating monounsaturated fatty acid synthesis
- Association of SOX6 gene polymorphisms with Kashin-Beck disease risk in the Chinese Han population
- The pyroptosis-related signature predicts prognosis and influences the tumor immune microenvironment in dedifferentiated liposarcoma
- METTL3 attenuates ferroptosis sensitivity in lung cancer via modulating TFRC
- Identification and validation of molecular subtypes and prognostic signature for stage I and stage II gastric cancer based on neutrophil extracellular traps
- Novel lumbar plexus block versus femoral nerve block for analgesia and motor recovery after total knee arthroplasty
- Correlation between ABCB1 and OLIG2 polymorphisms and the severity and prognosis of patients with cerebral infarction
- Study on the radiotherapy effect and serum neutral granulocyte lymphocyte ratio and inflammatory factor expression of nasopharyngeal carcinoma
- Transcriptome analysis of effects of Tecrl deficiency on cardiometabolic and calcium regulation in cardiac tissue
- Aflatoxin B1 induces infertility, fetal deformities, and potential therapies
- Serum levels of HMW adiponectin and its receptors are associated with cytokine levels and clinical characteristics in chronic obstructive pulmonary disease
- METTL3-mediated methylation of CYP2C19 mRNA may aggravate clopidogrel resistance in ischemic stroke patients
- Understand how machine learning impact lung cancer research from 2010 to 2021: A bibliometric analysis
- Pressure ulcers in German hospitals: Analysis of reimbursement and length of stay
- Metformin plus L-carnitine enhances brown/beige adipose tissue activity via Nrf2/HO-1 signaling to reduce lipid accumulation and inflammation in murine obesity
- Downregulation of carbonic anhydrase IX expression in mouse xenograft nasopharyngeal carcinoma model via doxorubicin nanobubble combined with ultrasound
- Feasibility of 3-dimensional printed models in simulated training and teaching of transcatheter aortic valve replacement
- miR-335-3p improves type II diabetes mellitus by IGF-1 regulating macrophage polarization
- The analyses of human MCPH1 DNA repair machinery and genetic variations
- Activation of Piezo1 increases the sensitivity of breast cancer to hyperthermia therapy
- Comprehensive analysis based on the disulfidptosis-related genes identifies hub genes and immune infiltration for pancreatic adenocarcinoma
- Changes of serum CA125 and PGE2 before and after high-intensity focused ultrasound combined with GnRH-a in treatment of patients with adenomyosis
- The clinical value of the hepatic venous pressure gradient in patients undergoing hepatic resection for hepatocellular carcinoma with or without liver cirrhosis
- Development and validation of a novel model to predict pulmonary embolism in cardiology suspected patients: A 10-year retrospective analysis
- Downregulation of lncRNA XLOC_032768 in diabetic patients predicts the occurrence of diabetic nephropathy
- Circ_0051428 targeting miR-885-3p/MMP2 axis enhances the malignancy of cervical cancer
- Effectiveness of ginkgo diterpene lactone meglumine on cognitive function in patients with acute ischemic stroke
- The construction of a novel prognostic prediction model for glioma based on GWAS-identified prognostic-related risk loci
- Evaluating the impact of childhood BMI on the risk of coronavirus disease 2019: A Mendelian randomization study
- Lactate dehydrogenase to albumin ratio is associated with in-hospital mortality in patients with acute heart failure: Data from the MIMIC-III database
- CD36-mediated podocyte lipotoxicity promotes foot process effacement
- Efficacy of etonogestrel subcutaneous implants versus the levonorgestrel-releasing intrauterine system in the conservative treatment of adenomyosis
- FLRT2 mediates chondrogenesis of nasal septal cartilage and mandibular condyle cartilage
- Challenges in treating primary immune thrombocytopenia patients undergoing COVID-19 vaccination: A retrospective study
- Let-7 family regulates HaCaT cell proliferation and apoptosis via the ΔNp63/PI3K/AKT pathway
- Phospholipid transfer protein ameliorates sepsis-induced cardiac dysfunction through NLRP3 inflammasome inhibition
- Postoperative cognitive dysfunction in elderly patients with colorectal cancer: A randomized controlled study comparing goal-directed and conventional fluid therapy
- Long-pulsed ultrasound-mediated microbubble thrombolysis in a rat model of microvascular obstruction
- High SEC61A1 expression predicts poor outcome of acute myeloid leukemia
- Comparison of polymerase chain reaction and next-generation sequencing with conventional urine culture for the diagnosis of urinary tract infections: A meta-analysis
- Secreted frizzled-related protein 5 protects against renal fibrosis by inhibiting Wnt/β-catenin pathway
- Pan-cancer and single-cell analysis of actin cytoskeleton genes related to disulfidptosis
- Overexpression of miR-532-5p restrains oxidative stress response of chondrocytes in nontraumatic osteonecrosis of the femoral head by inhibiting ABL1
- Autologous liver transplantation for unresectable hepatobiliary malignancies in enhanced recovery after surgery model
- Clinical analysis of incomplete rupture of the uterus secondary to previous cesarean section
- Abnormal sleep duration is associated with sarcopenia in older Chinese people: A large retrospective cross-sectional study
- No genetic causality between obesity and benign paroxysmal vertigo: A two-sample Mendelian randomization study
- Identification and validation of autophagy-related genes in SSc
- Long non-coding RNA SRA1 suppresses radiotherapy resistance in esophageal squamous cell carcinoma by modulating glycolytic reprogramming
- Evaluation of quality of life in patients with schizophrenia: An inpatient social welfare institution-based cross-sectional study
- The possible role of oxidative stress marker glutathione in the assessment of cognitive impairment in multiple sclerosis
- Compilation of a self-management assessment scale for postoperative patients with aortic dissection
- Left atrial appendage closure in conjunction with radiofrequency ablation: Effects on left atrial functioning in patients with paroxysmal atrial fibrillation
- Effect of anterior femoral cortical notch grade on postoperative function and complications during TKA surgery: A multicenter, retrospective study
- Clinical characteristics and assessment of risk factors in patients with influenza A-induced severe pneumonia after the prevalence of SARS-CoV-2
- Analgesia nociception index is an indicator of laparoscopic trocar insertion-induced transient nociceptive stimuli
- High STAT4 expression correlates with poor prognosis in acute myeloid leukemia and facilitates disease progression by upregulating VEGFA expression
- Factors influencing cardiovascular system-related post-COVID-19 sequelae: A single-center cohort study
- HOXD10 regulates intestinal permeability and inhibits inflammation of dextran sulfate sodium-induced ulcerative colitis through the inactivation of the Rho/ROCK/MMPs axis
- Mesenchymal stem cell-derived exosomal miR-26a induces ferroptosis, suppresses hepatic stellate cell activation, and ameliorates liver fibrosis by modulating SLC7A11
- Endovascular thrombectomy versus intravenous thrombolysis for primary distal, medium vessel occlusion in acute ischemic stroke
- ANO6 (TMEM16F) inhibits gastrointestinal stromal tumor growth and induces ferroptosis
- Prognostic value of EIF5A2 in solid tumors: A meta-analysis and bioinformatics analysis
- The role of enhanced expression of Cx43 in patients with ulcerative colitis
- Choosing a COVID-19 vaccination site might be driven by anxiety and body vigilance
- Role of ICAM-1 in triple-negative breast cancer
- Cost-effectiveness of ambroxol in the treatment of Gaucher disease type 2
- HLA-DRB5 promotes immune thrombocytopenia via activating CD8+ T cells
- Efficacy and factors of myofascial release therapy combined with electrical and magnetic stimulation in the treatment of chronic pelvic pain syndrome
- Efficacy of tacrolimus monotherapy in primary membranous nephropathy
- Mechanisms of Tripterygium wilfordii Hook F on treating rheumatoid arthritis explored by network pharmacology analysis and molecular docking
- FBXO45 levels regulated ferroptosis renal tubular epithelial cells in a model of diabetic nephropathy by PLK1
- Optimizing anesthesia strategies to NSCLC patients in VATS procedures: Insights from drug requirements and patient recovery patterns
- Alpha-lipoic acid upregulates the PPARγ/NRF2/GPX4 signal pathway to inhibit ferroptosis in the pathogenesis of unexplained recurrent pregnancy loss
- Correlation between fat-soluble vitamin levels and inflammatory factors in paediatric community-acquired pneumonia: A prospective study
- CD1d affects the proliferation, migration, and apoptosis of human papillary thyroid carcinoma TPC-1 cells via regulating MAPK/NF-κB signaling pathway
- miR-let-7a inhibits sympathetic nerve remodeling after myocardial infarction by downregulating the expression of nerve growth factor
- Immune response analysis of solid organ transplantation recipients inoculated with inactivated COVID-19 vaccine: A retrospective analysis
- The H2Valdien derivatives regulate the epithelial–mesenchymal transition of hepatoma carcinoma cells through the Hedgehog signaling pathway
- Clinical efficacy of dexamethasone combined with isoniazid in the treatment of tuberculous meningitis and its effect on peripheral blood T cell subsets
- Comparison of short-segment and long-segment fixation in treatment of degenerative scoliosis and analysis of factors associated with adjacent spondylolisthesis
- Lycopene inhibits pyroptosis of endothelial progenitor cells induced by ox-LDL through the AMPK/mTOR/NLRP3 pathway
- Methylation regulation for FUNDC1 stability in childhood leukemia was up-regulated and facilitates metastasis and reduces ferroptosis of leukemia through mitochondrial damage by FBXL2
- Correlation of single-fiber electromyography studies and functional status in patients with amyotrophic lateral sclerosis
- Risk factors of postoperative airway obstruction complications in children with oral floor mass
- Expression levels and clinical significance of serum miR-19a/CCL20 in patients with acute cerebral infarction
- Physical activity and mental health trends in Korean adolescents: Analyzing the impact of the COVID-19 pandemic from 2018 to 2022
- Evaluating anemia in HIV-infected patients using chest CT
- Ponticulus posticus and skeletal malocclusion: A pilot study in a Southern Italian pre-orthodontic court
- Causal association of circulating immune cells and lymphoma: A Mendelian randomization study
- Assessment of the renal function and fibrosis indexes of conventional western medicine with Chinese medicine for dredging collaterals on treating renal fibrosis: A systematic review and meta-analysis
- Comprehensive landscape of integrator complex subunits and their association with prognosis and tumor microenvironment in gastric cancer
- New target-HMGCR inhibitors for the treatment of primary sclerosing cholangitis: A drug Mendelian randomization study
- Population pharmacokinetics of meropenem in critically ill patients
- Comparison of the ability of newly inflammatory markers to predict complicated appendicitis
- Comparative morphology of the cruciate ligaments: A radiological study
- Immune landscape of hepatocellular carcinoma: The central role of TP53-inducible glycolysis and apoptosis regulator
- Serum SIRT3 levels in epilepsy patients and its association with clinical outcomes and severity: A prospective observational study
- SHP-1 mediates cigarette smoke extract-induced epithelial–mesenchymal transformation and inflammation in 16HBE cells
- Acute hyper-hypoxia accelerates the development of depression in mice via the IL-6/PGC1α/MFN2 signaling pathway
- The GJB3 correlates with the prognosis, immune cell infiltration, and therapeutic responses in lung adenocarcinoma
- Physical fitness and blood parameters outcomes of breast cancer survivor in a low-intensity circuit resistance exercise program
- Exploring anesthetic-induced gene expression changes and immune cell dynamics in atrial tissue post-coronary artery bypass graft surgery
- Empagliflozin improves aortic injury in obese mice by regulating fatty acid metabolism
- Analysis of the risk factors of the radiation-induced encephalopathy in nasopharyngeal carcinoma: A retrospective cohort study
- Reproductive outcomes in women with BRCA 1/2 germline mutations: A retrospective observational study and literature review
- Evaluation of upper airway ultrasonographic measurements in predicting difficult intubation: A cross-section of the Turkish population
- Prognostic and diagnostic value of circulating IGFBP2 in pancreatic cancer
- Postural stability after operative reconstruction of the AFTL in chronic ankle instability comparing three different surgical techniques
- Research trends related to emergence agitation in the post-anaesthesia care unit from 2001 to 2023: A bibliometric analysis
- Frequency and clinicopathological correlation of gastrointestinal polyps: A six-year single center experience
- ACSL4 mediates inflammatory bowel disease and contributes to LPS-induced intestinal epithelial cell dysfunction by activating ferroptosis and inflammation
- Affibody-based molecular probe 99mTc-(HE)3ZHER2:V2 for non-invasive HER2 detection in ovarian and breast cancer xenografts
- Effectiveness of nutritional support for clinical outcomes in gastric cancer patients: A meta-analysis of randomized controlled trials
- The relationship between IFN-γ, IL-10, IL-6 cytokines, and severity of the condition with serum zinc and Fe in children infected with Mycoplasma pneumoniae
- Paraquat disrupts the blood–brain barrier by increasing IL-6 expression and oxidative stress through the activation of PI3K/AKT signaling pathway
- Sleep quality associate with the increased prevalence of cognitive impairment in coronary artery disease patients: A retrospective case–control study
- Dioscin protects against chronic prostatitis through the TLR4/NF-κB pathway
- Association of polymorphisms in FBN1, MYH11, and TGF-β signaling-related genes with susceptibility of sporadic thoracic aortic aneurysm and dissection in the Zhejiang Han population
- Application value of multi-parameter magnetic resonance image-transrectal ultrasound cognitive fusion in prostate biopsy
- Laboratory variables‐based artificial neural network models for predicting fatty liver disease: A retrospective study
- Decreased BIRC5-206 promotes epithelial–mesenchymal transition in nasopharyngeal carcinoma through sponging miR-145-5p
- Sepsis induces the cardiomyocyte apoptosis and cardiac dysfunction through activation of YAP1/Serpine1/caspase-3 pathway
- Assessment of iron metabolism and iron deficiency in incident patients on incident continuous ambulatory peritoneal dialysis
- Tibial periosteum flap combined with autologous bone grafting in the treatment of Gustilo-IIIB/IIIC open tibial fractures
- The application of intravenous general anesthesia under nasopharyngeal airway assisted ventilation undergoing ureteroscopic holmium laser lithotripsy: A prospective, single-center, controlled trial
- Long intergenic noncoding RNA for IGF2BP2 stability suppresses gastric cancer cell apoptosis by inhibiting the maturation of microRNA-34a
- Role of FOXM1 and AURKB in regulating keratinocyte function in psoriasis
- Parental control attitudes over their pre-school children’s diet
- The role of auto-HSCT in extranodal natural killer/T cell lymphoma
- Significance of negative cervical cytology and positive HPV in the diagnosis of cervical lesions by colposcopy
- Echinacoside inhibits PASMCs calcium overload to prevent hypoxic pulmonary artery remodeling by regulating TRPC1/4/6 and calmodulin
- ADAR1 plays a protective role in proximal tubular cells under high glucose conditions by attenuating the PI3K/AKT/mTOR signaling pathway
- The risk of cancer among insulin glargine users in Lithuania: A retrospective population-based study
- The unusual location of primary hydatid cyst: A case series study
- Intraoperative changes in electrophysiological monitoring can be used to predict clinical outcomes in patients with spinal cavernous malformation
- Obesity and risk of placenta accreta spectrum: A meta-analysis
- Shikonin alleviates asthma phenotypes in mice via an airway epithelial STAT3-dependent mechanism
- NSUN6 and HTR7 disturbed the stability of carotid atherosclerotic plaques by regulating the immune responses of macrophages
- The effect of COVID-19 lockdown on admission rates in Maternity Hospital
- Temporal muscle thickness is not a prognostic predictor in patients with high-grade glioma, an experience at two centers in China
- Luteolin alleviates cerebral ischemia/reperfusion injury by regulating cell pyroptosis
- Therapeutic role of respiratory exercise in patients with tuberculous pleurisy
- Effects of CFTR-ENaC on spinal cord edema after spinal cord injury
- Irisin-regulated lncRNAs and their potential regulatory functions in chondrogenic differentiation of human mesenchymal stem cells
- DMD mutations in pediatric patients with phenotypes of Duchenne/Becker muscular dystrophy
- Combination of C-reactive protein and fibrinogen-to-albumin ratio as a novel predictor of all-cause mortality in heart failure patients
- Significant role and the underly mechanism of cullin-1 in chronic obstructive pulmonary disease
- Ferroptosis-related prognostic model of mantle cell lymphoma
- Observation of choking reaction and other related indexes in elderly painless fiberoptic bronchoscopy with transnasal high-flow humidification oxygen therapy
- A bibliometric analysis of Prader-Willi syndrome from 2002 to 2022
- The causal effects of childhood sunburn occasions on melanoma: A univariable and multivariable Mendelian randomization study
- Oxidative stress regulates glycogen synthase kinase-3 in lymphocytes of diabetes mellitus patients complicated with cerebral infarction
- Role of COX6C and NDUFB3 in septic shock and stroke
- Trends in disease burden of type 2 diabetes, stroke, and hypertensive heart disease attributable to high BMI in China: 1990–2019
- Purinergic P2X7 receptor mediates hyperoxia-induced injury in pulmonary microvascular endothelial cells via NLRP3-mediated pyroptotic pathway
- Investigating the role of oviductal mucosa–endometrial co-culture in modulating factors relevant to embryo implantation
- Analgesic effect of external oblique intercostal block in laparoscopic cholecystectomy: A retrospective study
- Elevated serum miR-142-5p correlates with ischemic lesions and both NSE and S100β in ischemic stroke patients
- Correlation between the mechanism of arteriopathy in IgA nephropathy and blood stasis syndrome: A cohort study
- Risk factors for progressive kyphosis after percutaneous kyphoplasty in osteoporotic vertebral compression fracture
- Predictive role of neuron-specific enolase and S100-β in early neurological deterioration and unfavorable prognosis in patients with ischemic stroke
- The potential risk factors of postoperative cognitive dysfunction for endovascular therapy in acute ischemic stroke with general anesthesia
- Fluoxetine inhibited RANKL-induced osteoclastic differentiation in vitro
- Detection of serum FOXM1 and IGF2 in patients with ARDS and their correlation with disease and prognosis
- Rhein promotes skin wound healing by activating the PI3K/AKT signaling pathway
- Differences in mortality risk by levels of physical activity among persons with disabilities in South Korea
- Review Articles
- Cutaneous signs of selected cardiovascular disorders: A narrative review
- XRCC1 and hOGG1 polymorphisms and endometrial carcinoma: A meta-analysis
- A narrative review on adverse drug reactions of COVID-19 treatments on the kidney
- Emerging role and function of SPDL1 in human health and diseases
- Adverse reactions of piperacillin: A literature review of case reports
- Molecular mechanism and intervention measures of microvascular complications in diabetes
- Regulation of mesenchymal stem cell differentiation by autophagy
- Molecular landscape of borderline ovarian tumours: A systematic review
- Advances in synthetic lethality modalities for glioblastoma multiforme
- Investigating hormesis, aging, and neurodegeneration: From bench to clinics
- Frankincense: A neuronutrient to approach Parkinson’s disease treatment
- Sox9: A potential regulator of cancer stem cells in osteosarcoma
- Early detection of cardiovascular risk markers through non-invasive ultrasound methodologies in periodontitis patients
- Advanced neuroimaging and criminal interrogation in lie detection
- Maternal factors for neural tube defects in offspring: An umbrella review
- The chemoprotective hormetic effects of rosmarinic acid
- CBD’s potential impact on Parkinson’s disease: An updated overview
- Progress in cytokine research for ARDS: A comprehensive review
- Utilizing reactive oxygen species-scavenging nanoparticles for targeting oxidative stress in the treatment of ischemic stroke: A review
- NRXN1-related disorders, attempt to better define clinical assessment
- Lidocaine infusion for the treatment of complex regional pain syndrome: Case series and literature review
- Trends and future directions of autophagy in osteosarcoma: A bibliometric analysis
- Iron in ventricular remodeling and aneurysms post-myocardial infarction
- Case Reports
- Sirolimus potentiated angioedema: A case report and review of the literature
- Identification of mixed anaerobic infections after inguinal hernia repair based on metagenomic next-generation sequencing: A case report
- Successful treatment with bortezomib in combination with dexamethasone in a middle-aged male with idiopathic multicentric Castleman’s disease: A case report
- Complete heart block associated with hepatitis A infection in a female child with fatal outcome
- Elevation of D-dimer in eosinophilic gastrointestinal diseases in the absence of venous thrombosis: A case series and literature review
- Four years of natural progressive course: A rare case report of juvenile Xp11.2 translocations renal cell carcinoma with TFE3 gene fusion
- Advancing prenatal diagnosis: Echocardiographic detection of Scimitar syndrome in China – A case series
- Outcomes and complications of hemodialysis in patients with renal cancer following bilateral nephrectomy
- Anti-HMGCR myopathy mimicking facioscapulohumeral muscular dystrophy
- Recurrent opportunistic infections in a HIV-negative patient with combined C6 and NFKB1 mutations: A case report, pedigree analysis, and literature review
- Letter to the Editor
- Letter to the Editor: Total parenteral nutrition-induced Wernicke’s encephalopathy after oncologic gastrointestinal surgery
- Erratum
- Erratum to “Bladder-embedded ectopic intrauterine device with calculus”
- Retraction
- Retraction of “XRCC1 and hOGG1 polymorphisms and endometrial carcinoma: A meta-analysis”
- Corrigendum
- Corrigendum to “Investigating hormesis, aging, and neurodegeneration: From bench to clinics”
- Corrigendum to “Frankincense: A neuronutrient to approach Parkinson’s disease treatment”
- Special Issue The evolving saga of RNAs from bench to bedside - Part II
- Machine-learning-based prediction of a diagnostic model using autophagy-related genes based on RNA sequencing for patients with papillary thyroid carcinoma
- Unlocking the future of hepatocellular carcinoma treatment: A comprehensive analysis of disulfidptosis-related lncRNAs for prognosis and drug screening
- Elevated mRNA level indicates FSIP1 promotes EMT and gastric cancer progression by regulating fibroblasts in tumor microenvironment
- Special Issue Advancements in oncology: bridging clinical and experimental research - Part I
- Ultrasound-guided transperineal vs transrectal prostate biopsy: A meta-analysis of diagnostic accuracy and complication rates
- Assessment of diagnostic value of unilateral systematic biopsy combined with targeted biopsy in detecting clinically significant prostate cancer
- SENP7 inhibits glioblastoma metastasis and invasion by dissociating SUMO2/3 binding to specific target proteins
- MARK1 suppress malignant progression of hepatocellular carcinoma and improves sorafenib resistance through negatively regulating POTEE
- Analysis of postoperative complications in bladder cancer patients
- Carboplatin combined with arsenic trioxide versus carboplatin combined with docetaxel treatment for LACC: A randomized, open-label, phase II clinical study
- Special Issue Exploring the biological mechanism of human diseases based on MultiOmics Technology - Part I
- Comprehensive pan-cancer investigation of carnosine dipeptidase 1 and its prospective prognostic significance in hepatocellular carcinoma
- Identification of signatures associated with microsatellite instability and immune characteristics to predict the prognostic risk of colon cancer
- Single-cell analysis identified key macrophage subpopulations associated with atherosclerosis