The effect of Cladophora glomerata exudates on the amino acid composition of Cladophora fracta and Rhizoclonium sp.
-
Marta Pikosz


Abstract
Filamentous green algae (FGA) frequently forms dense mats which can be either mono- or polyspecies. While various defense mechanisms of competition in algae are known, little is known about the interactions between different species of FGA. An experiment in controlled laboratory conditions was conducted to gather data on the changes in amino acids (AA) concentrations in FGA species in the presence of exudates from different other species. The aim of the present study was to identify the AA whose concentrations showed significant changes and to assess if the changes could be adaptation to stress conditions. The major constituents of the AA pool in Cladophora glomerata, C. fracta and Rhizoclonium sp. were Glutamic acid (Glu), Aspartic acid (Asp) and Leucine (Leu). In response to chemical stress, that is the increasing presence of exudates, a significant increase in the concentrations Proline (Pro) and Tryptophan (Trp) was noted. The increase in Proline levels was observed in C. fracta and Rhizoclonium in response to chemical stress induced by C. glomerata exudates. As the concentration of exudates increased in the medium, there was a progressive shift in the pattern of AA group in FGA.
1 Introduction
Cosmopolitan filamentous green algae (FGA) such as branched Cladophora and unbranched Rhizoclonium often experience massive growth in eutrophic water ecosystems, forming dense, seasonal mats [1, 2, 3, 4]. The mats are varied in many ways and one of their characteristic features is the number of taxa forming them [5, 6, 7]. In some cases they are formed from only one taxon whose rapid growth monopolizes the space on the water; other species co-exist with the dominant one or disappear enhancing the dominant species [8, 9]. According to Khanum [8] Cladophora glomerata (L.) Kützing, the most common Chlorophyta in the world, forms mainly monoalgal mats but sometimes becomes dominant in an algal community. The C. glomerata algae may co-exist with other Cladophora species or with other FGA taxa such as Rhizoclonium, Oedogonium [3, 10]. Filaments of C. glomerata formed monoalgal mats in the Samica Stęszewska river, Lake Oporzynskie; they co-existed with Rhizoclonium sp. Kützing in Mogilnica river and with Cladophora fracta (L.) in Malta reservoir [11,12]. The wide ecological range and fast growth of C. glomerata is a strategy which allows it to establish dominance over other macroalgae in water ecosystems [13]. This fact, confirmed in previous studies, has shown that the species belonging to the genus Cladophora (C. glomerata, C. rivularis, C. fracta) well tolerates a wide range of habitat requirements [12,14]. Additionally, mat formation, morphological variability, and chemical defense strategies used by FGA are the components of its stress avoidance mechanisms.
Several types of interactions between algae are seen in nature, e.g. allelopathic, non-allelopathic interactions and responses to different factors like resource competition or environmental stresses [15]. In water environments, Cladophora displays a high level of interactions with other organisms (microalgae, macroalgae, macrophytes) related to competing for light exposure, nutrients, and attachment substrate [16]. Much of the available evidence demonstrates competition between macroalgae and phytoplankton including periphytic algae and macroalgae-macrophytes but not between macroalgae and macroalgae. C. glomerata can prevent overgrowth by periphytic diatoms because dense coverings of diatoms would interfere with nutrient uptake by Cladophora and therefore inhibit growth [16,17]. According to Tarmanowska [18] C. glomerata inhibits the growth of Elodea canadensis Michx. Moreover, Cladophora is an example of a freshwater chlorophytes species which is capable of mediating allelopathic interactions, causing an increase in allelochemical concentration in response to competitors [1,19]. The green alga C. glomerata synthesizes a wide variety of toxic fatty acids (capric and palmitoleic acids) which have been demonstrated to be allelopathic [1]. Extract from C. glomerata slightly reduces the photosynthetic rate of diatoms [17].
Although the morphological variability related to the effect of different biotic and abiotic factors is well-known, the changes in the contents of proteins synthesized in algae biomass are little known. According to literature, only changes in Proline accumulation in response to environmental stress have been studied. The defense of resources by FGA involves competition, which may influence biochemical and morphological changes in cells and filaments - which have not been yet sufficiently studied [20]. This study focuses on C. glomerata, the most common and dominant FGA in the world and other species interaction with C. fracta and Rhizolonium sp. We hypothesize that the presence of C. glomerata exudates have a significant impact on the amino acid content in C. fracta and Rhizoclonium sp. The results of this study could provide first data for understanding the mechanisms of C. glomerata response to competition and its strategy of stress avoidance.
2 Materials and methods
2.1 Raw materials and their collection
The study was carried out in June 2015 when freshwater populations of FGA were in the optimal phase of development. Samples of Cladophora glomerata were taken from the shallow Lake Oporzynskie (N52°55'; E17°9'), Cladophora fracta from the surface of the Malta Reservoir in Poznan (N52°24'; E16°57') and Rhizoclonium sp. from a pond in Poznan (N52°28'; E16°55') in Wielkopolska Province in Poland. All samples were randomly selected from the mat at a depth of about 10 cm from the water surface. The filamentous green algae samples were collected into a plastic container and transported in a refrigerated container (at 4°C) to a laboratory. Next, the filaments were rinsed repeatedly with distilled water.
2.2 Microscopic identification
Morphometric measurements of the length and width of cells, pyrenoids (stained with Lugol's) and nuclei (stained by 1% acetocarmine) were made using a light microscope (LM). Images of opportunistic C. glomerata were made using a transmission electron microscopy (TEM), scanning electron microscopy (SEM) and light microscopy (LM). TEM images were recorded of filaments in control samples and filaments in the presence of 100% exudates. For the TEM technique, the filaments of C. glomerata were fixed in a mixture of glutaraldehyde (3%) in 0.1 M phosphate buffer (pH= 7.2) for two hours, washed several times in a buffer and postfixed in 1% osmium tetroxide (OsO4) in the same buffer. The cells were dehydrated by an ascending ethanol series, according to the method proposed by Farias et al. [21]. Finally, the samples were embedded in Spurr's resin (Low viscosity, ProScitech) [22] to produce a solid block, which was cut with and ultramicrotome (Leica EM UC7) at 60 nm, the ultrathin sections were contrasting in uranyl acetate, according to methods described by Reynolds [23]. Observations and photographs were carried out on a TESLA BS 500 transmission electron microscope.
2.3 Determination of total phenolic content
The total phenolic content of the filamentous green algae extract was determined by the Folin-Ciocalteu spectrophotometric method [24]. First, 30 μL of crude extract (10 mg/mL) were diluted with distilled water to achieve 1.8 mL, mixed thoroughly with 1.5 mL of Folin-Ciocalteu, followed by the addition of 4.5 mL of 20% (w/v) sodium carbonate and 5.7 mL of distilled water. The mixture was allowed to stand for further 2 hours in the dark, and absorbance was measured at 760 nm. The total phenolic content was calculated from the calibration curve, and the results were expressed as mg of gallic acid equivalent per g dry weight.
2.4 Determination of pigments
Photosynthetic pigments: chlorophyll a, chlorophyll b and carotenoids concentrations in the filaments were analyzed according to the methods proposed by Yoshii et al. [25] and Dere et al. [26] and were determined spectrophotometrically using the equations proposed by Lichtenthaler and Welburn [27].
2.5 Experimental set-up
The first experiment was designed to observe the mutual impact of filamentous green algae (C. glomerata, C. fracta, Rhizoclonium sp.) exudates on each other and on the composition of their amino acids. For the laboratory experiments, ca. 500 g fresh weight (FW) of either C. glomerata, C.fracta and Rhizoclonium sp. were placed into 3 aquaria which were fully filled up with 10 L of filtered and sterilized freshwater. FGA were acclimated and maintained in aquaria for 24 h. Then, cultures of FGA species were incubated in Wang's medium at a temperature of 21°C and in a diurnal regime (16/8 h light/dark cycle, 250 μmol photons m-2 s-1) in phytothron (CONVIRON model CMP 6050). The cultures were left to rest for 7 days so that the green algae species secreted the exudes to the medium. Upon commencing the experiment, algal pieces of ca. 5.0 g FW (only filaments in good condition, without evidence of necrosis) were placed into 200 mL beakers. In the experiment the control sample contained Wang's medium (100 % medium) and three samples with the medium containing algae exudates at different concentrations (25, 50, 100% exudates in medium). Each level was studied in triplicate using glass containers with 200 mL of medium and 5 g of fresh algae biomass. Four series of experiments were performed; the first with C. fracta cultured in the medium with C. glomerata exudates; the second with C. glomerata cultured in the medium with C. fracta exudates; the third with Rhizoclonium sp. with C. glomerata exudates and the fourth with C. glomerata filaments in the presence of Rhizoclonium sp. exudates. Each time, after completion of the series of experiments, the FW of FGA filaments was determined, as well as their DW after drying in a laboratory oven for 2 hours at 105°C. Then, the filaments were put into plastic 100 mL containers.
2.6 Amino acids analysis in biomass
The algae samples were analyzed using the modified Kjeldahl procedure (AoAC 2007) for dry matter (method no. 934.01) and ash (method no. 942.05). Hydrolysis of total protein was performed according to the protocol described by Malmer and Schroeder [28]. The amino acid standards alanine (Ala), aspartic acid (Asp), arginine (Arg), cysteine (Cys), glutamic acid (Glu), glycine (Gly), histidine (His), isoleucine (Ile), leucine (Leu), lysine (Lys), methionine (Met), phenylalanine (Phe), proline (Pro), serine (Ser), tyrosine (Tyr), threonine (Thr) and valine (Val) were from Sigma-Aldrich (Steinheim, Germany), and the amino acid standard tryptophan (Trp) were purchased from Merck (Darmstadt, Germany). The ground algae samples were subjected to acid hydrolysis with 6 M HCl at the temperature of 383.15 K for 23 hours. Amino acids were determined using a AAA 400 amino acid analyzer (INGOS, Czech Republic) with ion exchange chromatography. Post column ninhydrin-based detection and sodium citrate buffer were used. The ninhydrin amino acid derivatives were detected using packing of column OSTION Lg ANB (column height: 35 x 0.37 cm) at 570 nm for primary amino acids and at 440 nm for secondary amino acids. Amino acid concentrations in samples were calculated from calibration plots obtained by analysis of working solutions of different concentration. The contents were expressed as g·100g-1 of total proteins.
2.7 Statistical analysis
The homogeneity of the variance was analyzed using Levene's test. A One-way ANOVA followed by Tukey's RIR post hoc test was used to determine significant differences between the control and treatment groups. Statistical tests were performed with the program STATISTICA (ver. 12). Differences were considered to be significant at p<0.05.
Ethical approval: The conducted research is not related to either human or animal use.
3 Results
3.1 Freshwater filamentous green algae: morphology and chemical composition
The studied FGA formed dense, monoalgal, free-floating mats in eutrophic freshwater ecosystems. The morphology of algae taxa was studied carefully, and the taxa were identified as C. glomerata, C. fracta and Rhizoclonium sp. classified as Order: Cladophorales, Class: Ulvophyceae and Phylum: Chlorophyta. The detailed taxonomic description is included in Table 1. The C. glomerata filaments were strongly branched and inhabited by large numbers of periphytic communities which were responsible for their roughness (Figure 1). Each cell in the branching filaments of C. glomerata is multinucleate and contains many discoid chloroplasts in peripheral arrangement. The pyrenoid is bilenticular and is divided into two hemispheres by a single thylakoid, and each hemisphere is covered by a bowl-shaped starch grain. Each chloroplast is densely packed with tightly appressed thylakoids which are often paired. Grains of starch are distributed through the stroma of the chloroplast. The nucleus has a very large nucleolus. Each cell contains a single Golgi body, and two, sometimes three large, elongated mitochondria with very long cristae. In the cell cross-section, the greater part of the cell can be seen occupied by a central vacuole surrounded by a thin layer of cytoplasm. The cell walls are multilayered and composed of microfibrils (12-22 nm in diameter) arranged in parallel to each other. In contrast, C. fracta cells were more delicate, its filaments were thinner than those of the other FGA studied and were like cotton to the touch. Filaments of Rhizoclonium sp. were characterized by the same width close to 50 μm. Among the studied algae, C. glomerata had got the highest content of pigments, amino acids and total phenolic content (Table 1). The chl a, chl b and carotenoids concentration in C. glomerata biomass was almost twice higher than in C. fracta or in Rhizoclonium sp. In the studied FGA the same 18 amino acids have been identified (Figure 2). The relative abundance of AA in C. fracta decreases in the order: Glu> Asp> Leu> Arg> Val> Ala> Gly> Lys> Thr> Ser> Pro> Phe> Ile> Tyr> His> Cys> Met> Trp. The analogous order for C. glomerata was: Glu> Asp> Leu> Arg> Val> Ala> Gly> Lys> Thr> Pro> Ser> Phe> Ile> Tyr> His> Cys> Met> Trp. In Rhizoclonium sp. the distribution order from higher to lower was: Glu> Asp> Leu> Arg> Val> Ala> Gly> Lys> Ser> Thr> Pro> Phe> Ile> Tyr> His> Cys> Met> Trp. The abundance of AA in the studied FGA decreased in similar order and varied only in the sequence of Thr, Ser and Pro (underlined). Moreover, a considerable difference in phenols content was noted for the taxa studied and its highest concentration was in C. glomerata.

Details of the ultrastructure of Cladophora glomerata thalli: a – general view of branched filaments (LM); b -surface of filament inhabited by periphytic diatoms (SEM); c –TEM micrograph of mitochondrion (M) in the vicinity of chloroplast (Chl) and pyrenoid (P) surrounded by chloroplast; the arrangement of thylakoids (Th) in chloroplast; internal singular vacuole (V) well organized; d – cell wall (CW), external layer and internal division of two cells; lipids on the chloroplast (Chl); (c, d – scale bar = 0.5 μm).
Morphology and chemical characteristic of freshwater green algae (values: minimum – maximum or mean±SD, n=30 for morphology data, n=3 for chemical data).
| Taxon Features | Cladophora glomerata | Cladophora fracta | Rhizoclonium sp. | |
|---|---|---|---|---|
| MORPHOLOGY | Filaments characteristic | dark green, rough in touch, often trichotomies and subtending branching | yellow green, delicate, branching frequent | dark green, long, rough in touch, mainly without branched or only with short branched |
| Cell shape | cylindrical, elongated | cylindrical | cylindrical | |
| Cell width (μm) | 30–70 | 20–31 | 20-55 | |
| Cell length (μm) | 125 –400 | 50–155 | 70-174 | |
| Shape of chloroplasts | reticulate | parietal | reticulate/parietal | |
| Diameter of pyrenoids (μm) | 4.5-7.2 | 2.8-5.6 | 2.9-5.5 | |
| Diameter of nuclei (μm) | 3.8-8.5 | 1.9-4.1 | 2.2-4.6 | |
| CHEMICAL | Pigments (μgmL-1): | |||
| COMPOUNDS | Chl a | 8.45±0.18 | 3.13±0.12 | 4.43±0.10 |
| Chl b | 6.36±0.11 | 2.95±0.07 | 2.8±0.08 | |
| Carotenoids | 2.32±0.03 | 0.72±0.05 | 1.15±0.03 | |
| Amino Acid | 18AA | 18 AA | 18 AA | |
| (g·100g-1) of total protein) | 91.17±0.24 | 90.33±0.28 | 90.30±0.37 | |
| Total phenolic content mg gallic acid equivalent (GAE)/g DW | 17.32±0.23 | 11.55±0.21 | 12.28±0.14 |
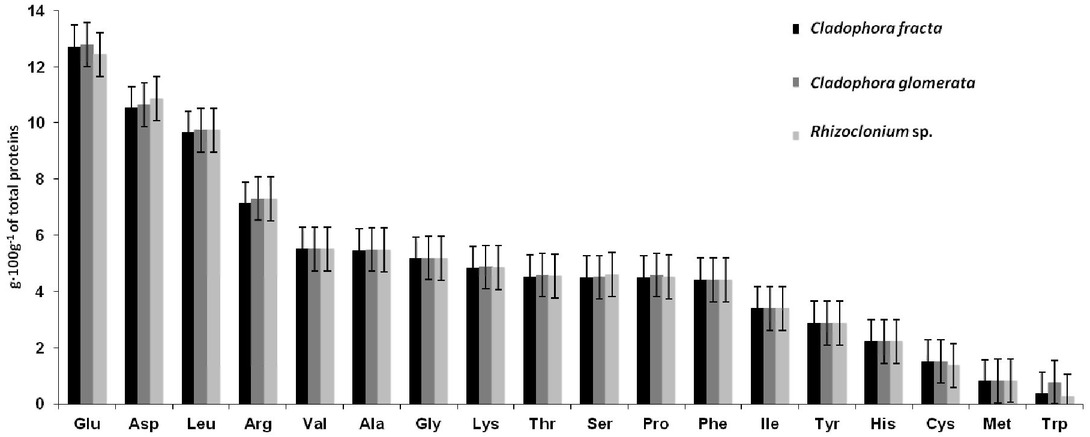
Amino acid profile in study algae (g·100g-1 of total protein).
3.2 Amino acid composition
3.2.1 Impact of exudates from C. glomerata on C. fracta amino acids production
In C. fracta filaments the presence of 18 amino acids was identified. Glutamic acid (Glu), Aspartic acid (Asp) and Leucine (Leu) were the most frequent (>10% of total AA content) while the least frequent (<1% of total AA content) was Methionine (Met) and Tryptophan (Trp) (Table 2). Production of AA in C. fracta cells was significantly higher for the filaments growing in the medium with C. glomerata exudates than in the control sample. For 12 AA the changes were statistically significant (Table 2). The concentration of 6 AA in the C. fracta cells was increased in response to increasing the exudates content. The greatest changes were observed for Proline: from 4.497 g·100g-1 of total protein in the control sample to 6.523 g·100g-1 of total protein in the medium with 100% of algal exudates and Tryptophan (Trp) from 0.359 to 0.466 g·100g-1, respectively. However the content of Asp, Glu significantly increased only in the medium with the highest exudates concentration (100%), in contrast to the contents of Ala, Lys and Phe which decreased (Table 2, S1).
The content of amino acids (AA) in Cladophora fracta biomass cultured in medium with exudates from C. glomerata in 3 series (g-100g-1 of total proteins; df=3; bold -statistically significant, P<0.05; gray selection -fluctuation in the content of amino acid >10%).
| Impact of exudates extracted from C. glomerata on C. fracta AA production | ||||||
|---|---|---|---|---|---|---|
| AA | Control | 25% exudate | 50% exudates | 100% exudate | F3,8 | P |
| Glutamic acid (Glu) | 12.690±0.008 | 12.706±0.007 | 12.704±0.002 | 12.731±0.003 | 8.939 | 0.000120 |
| Aspartic acid (Asp) | 10.527±0.002 | 10.528±0.004 | 10.532±0.002 | 10.567±0.004 | 1.561 | 0.000001 |
| Leucine(Leu) | 9.646±0.002 | 9.646±0.004 | 9.650±0.002 | 9.651±0.002 | 2.224 | 0.162901 |
| Arginine(Arg) | 7.412±0.001 | 7.409±0.002 | 7.413±0.002 | 7.414±0.001 | 5.333 | 0.026000 |
| Alanine (Ala) | 5.457±0.001 | 5.456±0.004 | 5.459±0.002 | 5.164±0.510 | 0.990 | 0.444947 |
| Valine (Val) | 5.512±0.001 | 5.510±0.002 | 5.512±0.002 | 5.517±0.005 | 4.444 | 0.040679 |
| Glycine (Gly) | 5.173±0.001 | 5.171±0.003 | 5.174±0.001 | 5.179±0.003 | 7.379 | 0.010840 |
| Lysine (Lys) | 4.824±0.001 | 4.825±0.001 | 4.827±0.002 | 4.799±0.003 | 1.186 | 0.000000 |
| Proline (Pro) | 4.496±0.002 | 4.553±0.005 | 5.464±0.004 | 6.523±0.03 | 5.290 | 0.000000 |
| Serine (Ser) | 4.502±0.001 | 4.504±0.002 | 4.506±0.002 | 4.506±0.003 | 2.119 | 0.176059 |
| Threonine (Thr) | 4.527±0.001 | 4.525±0.007 | 4.530±0.001 | 4.536±0.117 | 3.916 | 0.054420 |
| Phenylalanine(Phe) | 4.410±0.001 | 4.410±0.002 | 4.412±0.002 | 4.345±0.001 | 0.939 | 0.465777 |
| Isoleucine(lle) | 3.401±0.002 | 3.392±0.005 | 3.402±0.001 | 3.407±0.003 | 17.194 | 0.000756 |
| Tyrosine (Tyr) | 2.873±0.002 | 2.871±0.002 | 2.870±0.005 | 2.876±0.002 | 2.733 | 0.113590 |
| Histidine (His) | 2.207±0.001 | 2.209±0.002 | 2.209±0.002 | 2.212±0.003 | 4.471 | 0.040118 |
| Cysteine (Cys) | 1.505±0.002 | 1.501±0.003 | 1.503±0.001 | 1.509±0.005 | 6.925 | 0.012961 |
| Methionine (Met) | 0.798±0.002 | 0.797±0.003 | 0.811±0.001 | 0.815±0.002 | 26.589 | 0.000164 |
| Tryptophan (Trp) | 0.357±0.002 | 0.364±0.002 | 0.382±0.001 | 0.466±0.001 | 30.667 | 0.000000 |
3.2.2 Impact of exudates from C. fracta on C. glomerata amino acids production
The data in Table 3 show changes in the 18 amino acids levels in Cladophora glomerata thalli from the culture in the medium with C. fracta exudates. The most abundant amino acids in C. glomerata filaments were Glu, Asp, Leu and Arg. The results indicate a gradual increase in the contribution of all AA in C. glomerata, but significant changes occurred only for the samples from the medium containing 50% or 100% of C. fracta exudates. The effect was the same for all of amino acids in C. glomerata (Table S2). The content of Proline was increased from 4.5 to 4.9 g-100g-1 of total proteins and it was the highest change in the AA compositions.
The content of amino acids (AA) in Cladophora glomerata biomass (g·100g-1 of total proteins; df=3; bold -statistically significant, p<0.05; gray selection - fluctuation in the content of amino acid > 10%).
| Impact of exudates extracted from C. fracta on C. glomerata AA production | ||||||
|---|---|---|---|---|---|---|
| Control | 25% exudate | 50% exudate | 100% exudate | P | ||
| Glutamic acid (Glu) | 12.781±0.001 | 12.787±0.002 | 12.789±0.001 | 13.149±0.003 | 24.74 | 0.000000 |
| Aspartic acid (Asp) | 10.648±0.002 | 10.652±0.003 | 10.657±0.001 | 11.006±0.003 | 16.27 | 0.000000 |
| Leucine (Leu) | 9.735±0.001 | 9.740±0.001 | 9.747±0.001 | 9.967±0.002 | 21.19 | 0.000000 |
| Arginine(Arg) | 7.302±0.002 | 7.307±0.001 | 7.311±0.001 | 7.633±0.002 | 30.29 | 0.000000 |
| Alanine (Ala) | 5.492±0.001 | 5.503±0.002 | 5.509±0.001 | 5.586±0.002 | 28.03 | 0.000000 |
| Valine (Val) | 5.507±0.002 | 5.509±0.001 | 5.513±0.001 | 5.532±0.003 | 1.79 | 0.000000 |
| Glycine (Gly) | 5.185±0.002 | 5.193±0.002 | 5.200±0.001 | 5.224±0.001 | 2.48 | 0.000000 |
| Lysine (Lys) | 4.862±0.002 | 4.867±0.002 | 4.903±0.001 | 4.939±0.002 | 1.38 | 0.000000 |
| Proline (Pro) | 4.581±0.002 | 4.594±0.003 | 4.609±0.001 | 4.996±0.010 | 3.14 | 0.000000 |
| Serine (Ser) | 4.512±0.002 | 4.513±0.001 | 4.518±0.001 | 4.554±0.004 | 2.74 | 0.000000 |
| Threonine (Thr) | 4.586±0.002 | 4.590±0.001 | 4.596±0.001 | 4.627±0.002 | 5.44 | 0.000000 |
| Phenylalanine (Phe) | 4.406±0.002 | 4.411±0.002 | 4.417±0.001 | 4.445±0.003 | 1.93 | 0.000000 |
| Isoleucine(lle) | 3.392±0.002 | 3.404±0.003 | 3.743±0.001 | 3.767±0.578 | 0.77 | 0.544255 |
| Tyrosine (Tyr) | 2.875±0.002 | 2.878±0.004 | 2.886±0.001 | 2.917±0.001 | 2.69 | 0.000000 |
| Histidine (His) | 2.213±0.002 | 2.216±0.002 | 2.219±0.001 | 2.240±0.002 | 1.29 | 0.000000 |
| Cysteine (Cys) | 1.508±0.002 | 1.511±0.001 | 1.519±0.001 | 1.546±0.003 | 1.34 | 0.000000 |
| Methionine(Met) | 0.810±0.002 | 0.814±0.002 | 0.819±0.001 | 0.832±0.003 | 6.91 | 0.000000 |
| Tryptophan (Trp) | 0.765±0.002 | 0.836±0.001 | 0.861±0.001 | 0.943±0.002 | 6.89 | 0.000000 |
3.2.3 Impact of exudates from C. glomerata on Rhizoclonium sp. amino acids production
Amino acids in Rhizoclonium sp. showed a different response to addition of exudates from C. glomerata. The effect of experiment was irregular: the level of AA increased or decreased in response to addition of the exudates (Table 4). The contents of Proline and Lysine increased in the sample with the maximal concentration of macroalgal exudates when compared to those in the control sample.
The content of amino acids (AA) in Rhizoclonium sp. biomass (g·100g-1 of total proteins; df=3; bold -statistically significant, p<0.05; gray selection - fluctuation in the content of amino acid > 10%).
| Impact of exudates extracted C. glomerata on Rhizoclonium sp. AA production | ||||||
|---|---|---|---|---|---|---|
| Control | 25% exudate | 50% exudate | 100% exudate | F | P | |
| 3 , 8 | ||||||
| Glutamic acid (Glu) | 12.431±0.002 | 12.702±0.006 | 12.566±0.003 | 12.716±0.004 | 4.12 | 0.000000 |
| Aspartic acid (Asp) | 10.863±0.001 | 10.541±0.002 | 10.702±0.003 | 10.551±0.003 | 1.29 | 0.000000 |
| Leucine(Leu) | 9.731±0.002 | 9.697±0.002 | 9.714±0.003 | 9.721±0.002 | 1.64 | 0.000000 |
| Arginine(Arg) | 7.298±0.001 | 7.253±0.040 | 7.276±0.004 | 7.313±0.002 | 5.0 | 0.027661 |
| Alanine (Ala) | 5.469±0.001 | 5.470±0.002 | 5.469±0.004 | 5.481±0.002 | 1.3 | 0.002032 |
| Valine (Val) | 5.509±0.002 | 5.514±0.006 | 5.512±0.004 | 5.531±0.002 | 2.5 | 0.000192 |
| Glycine (Gly) | 5.178±0.001 | 5.182±0.002 | 5.180±0.001 | 5.207±0.002 | 1.57 | 0.000000 |
| Lysine (Lys) | 4.842±0.001 | 4.844±0.002 | 4.843±0.002 | 8.520±0.001 | 4.34 | 0.000000 |
| Proline (Pro) | 4.514±0.001 | 4.567±0.002 | 4.541±0.007 | 6.946±0.013 | 7.39 | 0.000000 |
| Serine (Ser) | 4.612±0.001 | 4.616±0.005 | 4.614±0.002 | 4.627±0.002 | 1.4 | 0.001374 |
| Threonine (Thr) | 4.536±0.001 | 4.538±0.002 | 4.537±0.001 | 4.547±0.002 | 3.9 | 0.000039 |
| Phenylalanine(Phe) | 4.402±0.001 | 3.387±0.002 | 3.895±0.002 | 3.406±0.004 | 12.88 | 0.000000 |
| Isoleucine(lle) | 3.387±0.002 | 2.872±0.002 | 3.130±0.003 | 2.898±0.008 | 9.9 | 0.000000 |
| Tyrosine (Tyr) | 2.869±0.001 | 2.212±0.003 | 2.541±0.002 | 2.225±0.002 | 7.89 | 0.000000 |
| Histidine (His) | 2.211±0.001 | 2.218±0.001 | 2.215±0.001 | 2.223±0.002 | 6.7 | 0.000005 |
| Cysteine (Cys) | 1.356±0.001 | 1.359±0.001 | 1.357±0.001 | 1.370±0.002 | 7.2 | 0.000004 |
| Methionine (Met) | 0.824±0.001 | 0.818±0.008 | 0.821±0.002 | 0.837±0.003 | 1.0 | 0.004162 |
| Tryptophan (Trp) | 0.268±0.001 | 0.275±0.002 | 0.271±0.003 | 0.314±0.002 | 3.31 | 0.000000 |
However, the contents of Asp, Ile and Tyr were reduced in the sample with 100% exudates relative to those in the control sample. The changes in the concentration of the other AA were less pronounced, although all changes in AA were statistically significant (Table S3).
3.2.4 Impact of exudates from Rhizoclonium sp. on C. glomerata amino acids production
The results shown in the Table 5 indicate a gradual increase in the contribution of all AAs in C. glomerata in response to the addition of Rhizoclonium sp. exudates. The results were statistically significant for all of the amino acids in C. glomerata. Changes in Proline content varied from 4.58 to 5.12 g·100g-1 of total proteins and its increase was the highest from among all AA studied (Table S4).
The content of amino acids (AA) in Cladophora glomerata biomass. (g·100g-1 of total proteins; df=3; bold -statistically significant, p<0.05; gray selection - fluctuation in the content of amino acid > 10%).
| Impact of exudates extracted Rhizoclonium sp . on C. glomerata AA production | ||||||
|---|---|---|---|---|---|---|
| Control | 25% exudate | 50% exudate | 100% exudate | F 3 , 8 | P | |
| Glutamic acid (Glu) | 12.774±0.002 | 12.778±0.002 | 12.783±0.002 | 12.896±0.002 | 2.64 | 0.000000 |
| Aspartic acid (Asp) | 10.639±0.001 | 10.643±0.002 | 10.646±0.001 | 10.970±0.003 | 2.84 | 0.000000 |
| Leucine(Leu) | 9.728±0.002 | 9.730±0.001 | 9.733±0.002 | 9.835±0.003 | 3.78 | 0.000000 |
| Arginine (Arg) | 7.302±0.001 | 7.306±0.002 | 7.309±0.002 | 7.431±0.002 | 4.60 | 0.000000 |
| Alanine (Ala) | 5.487±0.002 | 5.491±0.002 | 5.507±0.001 | 5.523±0.002 | 1.20 | 0.000001 |
| Valine (Val) | 5.501±0.001 | 5.504±0.002 | 5.518±0.001 | 5.521±0.002 | 3.50 | 0.000000 |
| Glycine (Gly) | 5.183±0.002 | 5.193±0.002 | 5.205±0.002 | 5.223±0.002 | 9.08 | 0.000000 |
| Lysine (Lys) | 4.861±0.001 | 4.871±0.002 | 4.895±0.001 | 4.925±0.002 | 41.47 | 0.000000 |
| Proline (Pro) | 4.579±0.001 | 4.599±0.001 | 4.637±0.002 | 5.123±0.004 | 4.47 | 0.000000 |
| Serine (Ser) | 4.508±0.001 | 4.511±0.001 | 4.519±0.003 | 4.548±0.001 | 7.05 | 0.000003 |
| Threonine (Thr) | 4.587±0.001 | 4.592±0.004 | 4.603±0.004 | 4.612±0.002 | 2.78 | 0.000000 |
| Phenylalanine(Phe) | 4.405±0.001 | 4.408±0.002 | 4.413±0.001 | 4.439±0.003 | 6.51 | 0.000000 |
| Isoleucine(lle) | 3.390±0.001 | 3.393±0.002 | 3.402±0.001 | 3.427±0.001 | 2.11 | 0.000000 |
| Tyrosine (Tyr) | 2.872±0.001 | 2.875±0.002 | 2.881±0.002 | 2.913±0.004 | 1.63 | 0.000001 |
| Histidine (His) | 2.207±0.001 | 2.211±0.002 | 2.221±0.003 | 2.233±0.002 | 1.46 | 0.000000 |
| Cysteine (Cys) | 1.508±0.001 | 1.512±0.001 | 1.524±0.001 | 1.529±0.002 | 1.17 | 0.000000 |
| Methionine (Met) | 0.805±0.001 | 0.810±0.002 | 0.810±0.001 | 0.827±0.002 | 2.16 | 0.000000 |
| Tryptophan (Trp) | 0.767±0.001 | 0.825±0.002 | 0.825±0.003 | 0.915±0.003 | 2.64 | 0.000000 |
3.3 Changes in Cladophora glomerata cell structure
Cell components such as chloroplasts, mitochondria, starch grains, lipid bodies, nuclei, and small vacuole were well organized and visible in the control samples. Changes caused by the presence of exudates were also observed in morphological features, i.e. the width of cell wall, shape and diameter of pyrenoids and in chloroplast arrangement. We observed (using TEM) these changes in C. glomerata in the control samples (Figure 3a,c,e) relative to the analogous features in the samples cultured in the medium with the highest concentration of exudates (Figure 3b,d,f). High magnification image of the cell wall clearly shows the parallel organization of the cellulose fibrils and the cell wall thinner (3.5 μm) in the control medium (Figure 3a) than (4.3 μm) in the medium with 100% exudates (Figure 4). The difference in diameter of pyrenoids were up to 3μm and the external layer was fuzzy (Figure 3c,d). Mitochondria were well organized in the control sample cells, and the parallel thylakoids membranes of the chloroplast were well preserved. Chloroplasts were in peripheral arrangement and were densely packed with tightly appressed thylakoids but under stress condition they were more rarely packed (Figure 3e,f).
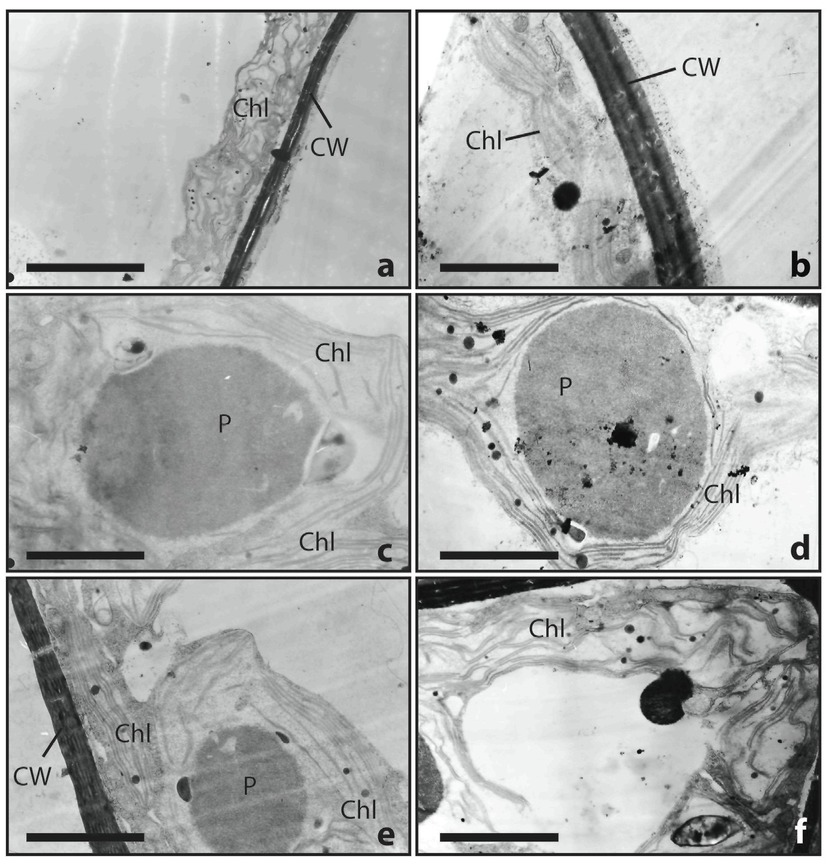
Ultrastructure analysis by transmission electron microscopy (TEM) of Cladophora glomerata with (right side) and without stress (left side): a – thin cell wall (CW) and normal chloroplast (Chl), b – multilayered cell walls (CW) and chloroplast (Chl); c, d – pyrenoid (P) surrounded by the chloroplast (Chl); e – multilayered cell wall (CW), chloroplast (Chl) and pyrenoid (P), f– cross section of filament showing chloroplast (Chl) (scale bar: 0.2 μm).
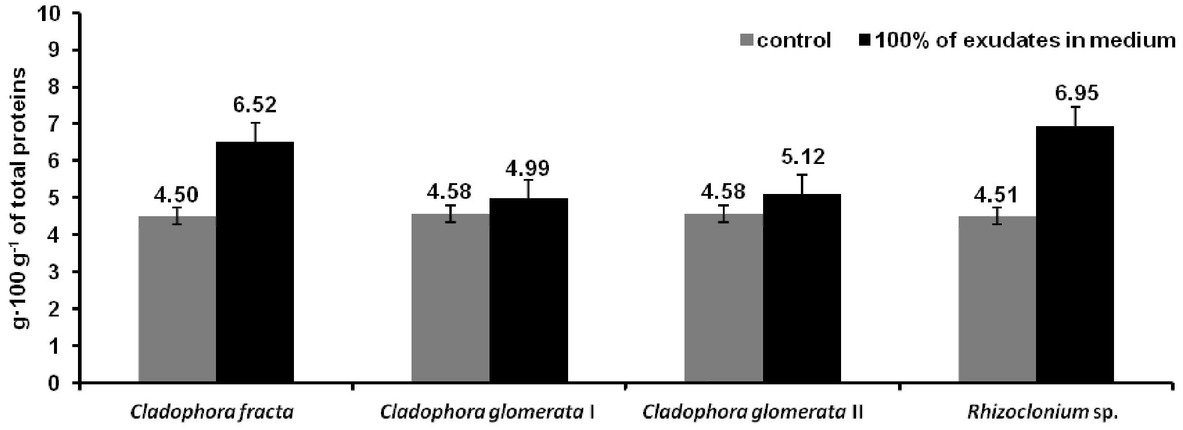
Diagram of Proline concentration in algae cell in control and in medium with 100% of exudates (C. fracta with 100% of C. glomerata exudates, C. glomerata I with 100% of exudates of C. fracta, C. glomerata II with 100% of exudates of Rhizoclonium sp. and Rhizoclonium sp. with 100% of C. glomerata exudates).
4 Disscusion
The results of our study have evidenced that filamentous green algae respond to the presence of other FGA species in the natural environment. The competition between different species of macroalgae may lead to the exchange of particular species in the community structure. We have shown for the first time the evidence of morphological changes at the cellular level in the filaments of C. glomerata in response to a biotic factor which is the appearance of exudates as a result of thalli accumulation. The effect of abiotic factors on morphological changes has been
partly described by Whitton [16], Pikosz and Messyasz [12]. Filamentous green algae often form monospecies mats in water ecosystems. Ecological characteristics of certain species may lead to their definite domination, as e.g. C. glomerata which is the most common FGA in the world [16]. Other species of filamentous green algae, as Cladophora rivularis (L) Kuntze and Rhizoclonium sp., do not occur so frequently or are less abundant [12]. The important outcome of our study is the evidence that the interactions between the macroalgae species can lead to changes in their biochemical structure. The green algae species selected for our study responded to the presence of exudates of other species recognized as competitors for the environment resources through changes in the quantitative composition of amino acids. In C. fracta and Rhizoclonium sp. the changes in AA profile were visible at the lower exudates concentration than in C. glomerata. It may explain, why C. fracta and Rhizoclonium sp. more rarely formed monoalgal mats and are rarely dominant in algae structures. Furthermore, different responses in AA contents were observed for C. rivularis and Rhizoclonium sp. under C. glomerata pressure. In C. fracta the changes in the content of three AA were more pronounced in the sample having the highest concentration of exudates (100%) than in the control sample. Ala decreased from 5.4 to 5.1 g·100g-1 of total proteins, Trp increased from 0.36 to 0.46 g·100g-1 of total proteins (30% of fluctuation in the content of amino acid) and the most pronounced changes were observed for Pro whose content increased from 4.49 to 6.5 g·100g-1 of total proteins (45%). However, in Rhizoclonium sp. the following changes in the contents of seven amino acids were observed, the concentrations of Pro (53% of fluctuation in the content of amino acid), Lys (75%), Trp (17%) and Glu increased, while those of Asp, Phe (22%) and Tyr (22%) decreased. The highest changes were noted for Lysine from 4.8 to 8.5 g·100g-1 of
total proteins (30%) and Proline increased from 4.8 to 6.9 g·100g-1 of total proteins (53%). An interesting observation is that the changes in the AA composition of C. glomerata in response to the presence of C. rivularis and Rhizoclonium sp. exudates are different. The changes in concentrations in AA in C. glomerata in response to the presence of C. fracta exudates were detected for seven amino acids (Leu, Glu, Asp, Arg, Pro 11%, Ile, Trp 19%), while in response to the Rhizoclonium sp. exudate only in one (Pro - 9% - 30% of fluctuation in the content of amino acid). This phenomenon confirms the C. glomerata plasticity and the fact that this species shows a wide ecological range [16].
The protein content of algae can vary according to the species, seasonal period, environmental condition and geographic area [29]. Amsler [30] determined the amino acid composition of 8 species of macroalgae and the most abundant AA were Asp, Glu, Lys, His, Val and Ala. C. glomerata contained 17 amino acids and Glutamic acid and Aspartic acid were detected in the highest contents in the filaments collected from the lake [11,31]. According to our results, the same AA were dominant, but we also observed the presence of Tryptophan. The major abundant amino acids (from 17 AA) identified in the freshwater Rhizoclonium hieroglyphicum (C. Agardh) Kützing from Northern Thailand were Tryptophan, Tyrosine and Phenylalanine [32]. Their content was lower than that of AA in freshwater Rhizoclonium sp. from our study, in which also Arg was noted. Interestingly, protein concentration in marine C. glomerata range about 20% of dry basic [33], while total protein content was slightly smaller and ranged from 12.75% to 14.45% in freshwater Cladophora [31].
Kaur and Asthir [34] suggest that accumulation of Proline is a common physiological response in plants exposed to various environmental stress, mainly high soil salinity, drought, oxidative stress, heavy metals presence. In the control samples for all individual taxa, the Proline content was at the same level (about 4.5 g·100g-1 of total proteins). The Proline content increased as a result of increasing content of exudation, a biotic Stressor, both in C. glomerata, C. fracta and Rhizoclonium sp. However in C. fracta and Rhizoclonium sp. there was a higher increase in Proline production than in C. glomerata (Figure 4). Slight differences were observed in the changes of Proline content in C. glomerata cultured in the medium with exudates of different macroalgae. The content of Proline in C. glomerata cultured with exudates of Rhizoclonium sp. (C. glomerata II in Figure 4) was higher than in C. glomerata cultured with exudates of C. fracta (C. glomerata I in Figure 4).
There are a lot of factors which influence the morphometric features of C. glomerata, including seasonality, stage of development, physicochemical parameters of water. The thickness of the C. glomerata cell wall may show seasonal variations, for example the samples from June had thinner cell walls than the samples from July [35]. However, results of this study indicate that also biotic factors may affect the morphological structure of the cell.
5 Conclusions
The results of the experiment with the three taxa of green algae C. glomerata, C. fracta and Rhizoclonium sp., show that the changes in amino acids production depend on FGA species and exudates concentration. Generally, the amount of some amino acids increased with increasing exudates concentrations. C. fracta and Rhizoclonium sp. react (by changes in AA profile) at the minimum C. glomerata exudates in medium, it was visible mainly in Proline concentration. We also observed, that C. glomerata produced more amino acids in response to exudates from the other FGA, but its response was noted only in the highest exudate concentration. Important are results which show that one species can have different strategies. To find out why C. glomerata monopolizes water surface, although this species occupies the same niche as the other filamentous green algae, a future study is needed to establish the composition of its secretions.
Acknowledgements
This project is financed in the framework of grant 2014/13/B/NZ8/04690 entitled: "Physico-chemical and biological causes of ecological dominance of filamentous green algae in freshwater ecosystems" attributed by the National Science Center.
Conflict of interest: Authors declare no conflict of interest
Supplemental Material: The online version of this article offers supplementary material (https://doi.org/10.1515/chem-2019-0032)
References
[1] Dodds W. K., Gudder D.A., The ecology of Cladophora, J. Phycol., 1992, 28, 415-27.10.1111/j.0022-3646.1992.00415.xSearch in Google Scholar
[2] Wetzel R.G., Limnology. Lake and River ecosystems, 3rd ed. Academic Press, 2001.Search in Google Scholar
[3] Pieczyńska E.., Eutrophication of shallow lakes – importance of macrophytes, Wiadomości Ekologiczne, 2008, 54(1), 3-28.Search in Google Scholar
[4] Mareš J., Leskinen E., Sitkowska M., Skácelová O., Blomster )., True identity of the European freshwater Ulva (Chlorophyta, Ulvophyceae) revealed by a combined molecular and morphological approach, J. Phycol., 2011, 47(5), 1177-1192.10.1111/j.1529-8817.2011.01048.xSearch in Google Scholar
[5] Saunders L.L., Metaphyton mat conditions and their effects on filamentous algal communities and their diatomd epiphytes. Thesis, Faculty of Drexel University, 2009.Search in Google Scholar
[6] Pikosz M., Oedogonium capillare (Chlorophyta, Oedogoniales) from the new site in Poland. Creative Sceince-Monography, Creative Time, 2012, 3, 7-9.Search in Google Scholar
[7] Messyasz B., Pikosz M., Treska E., Biology of freshwater macroalgae and their distribution, In: Chojnacka K., Wieczorek P.P., Schroeder G., Michalak I. (Eds), Algae Biomass: Characteristics and Applications. Towards algae-based products. Springer Publishers, Dondrecht, The Netherlands, 2018, 3, 17-31.10.1007/978-3-319-74703-3_3Search in Google Scholar
[8] Khanum A., An ecological study of freshwater algal mats, Bot. Bull. Academia Sinica, 1982, 23, 89-104.Search in Google Scholar
[9] Pikosz M., Messyasz B., Composition and seasonal changes in filamentous algae in floating mats, Oceanol Hydrobiol Stud, 2015, 44(2), 273-281.10.1515/ohs-2015-0026Search in Google Scholar
[10] Lembi C.A., Relative Tolerance of Mat-forming Algae to Copper, J. Aquat. Plant Manage, 2000, 38, 68-70.Search in Google Scholar
[11] Messyasz B., Leska B., Fabrowska J., Pikosz M., Roj E., Cieslak A., Schroeder G., Biomass of freshwater Cladophora as a raw material for agriculture and the cosmetic industry, Open Chemistry, 2015, 13, 1108-1118.10.1515/chem-2015-0124Search in Google Scholar
[12] Pikosz M., Messyasz B., Characteristic of Cladophora and coexisting filamentous algae against a background of environmental factors in freshwater ecosystems in Poland, Oceanol Hydrobiol Stud, 2016, 45(2), 202-215.10.1515/ohs-2016-0019Search in Google Scholar
[13] Ensminger I., Xyländer M., Hagen C., Braune W., Strategies providing success in a variable habitat: III. Dynamic control of photosynthesis in Cladophora glomerata, Plant Cell Environ., 2001, 24(8), 769-779.10.1046/j.1365-3040.2001.00725.xSearch in Google Scholar
[14] Pikosz M., Messyasz B., Gabka M., Functional structure of algal mat (Cladophora glomerata) in a freshwater in western Poland, Ecol Indic, 2017, 74, 1-9.10.1016/j.ecolind.2016.09.041Search in Google Scholar
[15] Irfanullah H. Md., Moss B., A filamentous green algae-dominated temperate shallow lake: Variations on the theme of clear-water stable states?, Arch. Hydrobiol., 2005, 163(1), 25-47.10.1127/0003-9136/2005/0163-0025Search in Google Scholar
[16] Whitton B.A., Biology of freshwater Cladophora, Water Res., 1970, 4, 457-476.10.1016/0043-1354(70)90061-8Search in Google Scholar
[17] Dodds W.K., Community interactions between the filamentous alga Cladophora glomerata (L.) Kuetzing, its epiphytes, and epiphyte grazers, Oecologia, 1991, 85, 572-80.10.1007/BF00323770Search in Google Scholar
[18] Tarmanowska A., Laboratory studies on the influence of living and decomposing filamentous algae on the growth of Elodea canadensis Michx., Acta Botanica Gallica, 1995, 142, 685-692.10.1080/12538078.1995.10515293Search in Google Scholar
[19] Fabrowska J., Messyasz B., Pankiewicz R., Wilinska P., Leska B., Seasonal differences in the content of phenols and pigments in thalli of freshwater Cladophora glomerata and its habitat, Water Res., 2018, 135, 66-74.10.1016/j.watres.2018.02.020Search in Google Scholar
[20] Messyasz B., Łęska B., Pikosz M., Fabrowska J., Cieślak A., Schroeder G., Effects of organic compounds on the macroalgae culture of freshwater Aegagropila, Open Chemistry, 2015, 13, 1040-1044.10.1515/chem-2015-0119Search in Google Scholar
[21] Farias D.R., Simioni C., Poltronieri E., Bouzon Z.L., Macleod C.K., Fine-tuning transmission electron microscopy methods to evaluate the cellular architecture of Ulvacean seaweeds (Chlorophyta), Micron, 2017, 96, 48-56.10.1016/j.micron.2017.02.003Search in Google Scholar
[22] Spurr A., A low viscosity epoxy resin embedding medium for electron microscopy, J Ultra Res., 1969, 26(1-2), 31-43.10.1016/S0022-5320(69)90033-1Search in Google Scholar
[23] Reynolds E.S., The use of lead citrate at high pH as an electron-opaque stain in electron microscopy, J Cell Biol., 1963, 17, 208-12.10.1083/jcb.17.1.208Search in Google Scholar
[24] Singleton V.L., Orthofer R., Lamuela-Raventos R.M., Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent, Methods Enzymol, 1999, 299, 152-178.10.1016/S0076-6879(99)99017-1Search in Google Scholar
[25] Yoshii Y., Hanyuda T., Wakana I., Miyaji K., Arai S., Ueda K., Inouye I., Carotenoid compositions of Cladophora balls (Aegagropila linnaei) and some members of the Cladophorales (Ulvophyceae, Chlorophyta): Their taxonomic and evolutionary implication, J. Phycol., 2004, 40, 1170-1177.10.1111/j.1529-8817.2004.03210.xSearch in Google Scholar
[26] Dere S., Günes T., Sivaci R., Spectrophotometric determination of chlorophyll-a, b and total carotenoid contents of some algae species using different solvents, Turk. J. Bot., 1998, 22, 13-17.Search in Google Scholar
[27] Lichtenthaler H.K., Wellburn A.R., Determination of total carotenoids and chlorophyll a and b of leaf extractin different solvents, Biochem. Soc. Trans., 1983, 11, 591-592.10.1042/bst0110591Search in Google Scholar
[28] Malmer M.F., Schroeder L.A., Amino acid analysis by high-performance liquid chromatography with methanesulfonic acid hydrolysis and 9-fluorenylmethyl-chloroformate derivatization, J. of Chromat. A, 1990, 514, 227-239.10.1016/S0021-9673(01)89394-8Search in Google Scholar
[29] Haroon A.M., The biochemical composition of Enteromorpha spp. from the Gulf of Gdansk coast on the southern Baltic Sea, Oceanologia, 2000, 42(1), 19-28.Search in Google Scholar
[30] Amsler, C.D., Algal Chemical Ecology, Springer Verlag, Berlin, Heidelberg, Germany, 2008, 32210.1007/978-3-540-74181-7Search in Google Scholar
[31] Messyasz B., Leska B., Pikosz M., Fabrowska J., Schroeder G., Characteristic of bioactive compounds from biomass of freshwater Cladophora glomerata, Eur J Phycol, 2015, 50,1, 140-141Search in Google Scholar
[32] Mungmai L., Jiranusornkul S., Peerapornpisal Y., Sirithunyalug B., Leelapornpisid P., Extraction characterization and biological activities of extracts from freshwater macroalga (Rhizoclonium hieroglyphicum (C. Agardh) Kützing) cultivated in northern Thailand, Chiang Mai J. Sci, 2014, 41(1), 14-26.Search in Google Scholar
[33] Manivannan K., Thirumaran G., Karthikai Devi G., Anantharaman P., Balasubramanian T., Proximate composition of different groups of seaweeds from Vedalai coastal waters (Gulf of Mannar): southeast coast of India, Middle-East, J. Sci. Res., 2009, 4 (2), 72–77.Search in Google Scholar
[34] Kaur G., Asthir B., Proline: a key player in plant abiotic stress tolerance, Biol Plant, 2015, 59, 609.10.1007/s10535-015-0549-3Search in Google Scholar
[35] Pankiewicz R., Leska B., Messyasz B., Fabrowska J., Sotoduch M., Pikosz M., First isolation of polysaccharidic ulvans from the cell walls of freshwater algae, Algal Research, 2016, 19, 348-354.10.1016/j.algal.2016.02.025Search in Google Scholar
© 2019 Marta Pikosz, Joanna Czerwik-Marcinkowska, Beata Messyasz, published by De Gruyter
This work is licensed under the Creative Commons Attribution 4.0 Public License.
Articles in the same Issue
- Regular Articles
- Research on correlation of compositions with oestrogenic activity of Cistanche based on LC/Q-TOF-MS/MS technology
- Efficacy of Pyrus elaeagnifolia subsp. elaeagnifolia in acetic acid–induced colitis model
- Anti-inflammatory and antinociceptive features of Bryonia alba L.: As a possible alternative in treating rheumatism
- High efficiency liposome fusion induced by reducing undesired membrane peptides interaction
- Prediction of the Blood-Brain Barrier Permeability Using RP-18 Thin Layer Chromatography
- Phytic Acid Extracted from Rice Bran as a Growth Promoter for Euglena gracilis
- Development of a validated spectrofluorimetric method for assay of sotalol hydrochloride in tablets and human plasma: application for stability-indicating studies
- Topological Indices of Hyaluronic Acid-Paclitaxel Conjugates’ Molecular Structure in Cancer Treatment
- Thermodynamic properties of the bubble growth process in a pool boiling of water-ethanol mixture two-component system
- Critical Roles of the PI3K-Akt-mTOR Signaling Pathway in Apoptosis and Autophagy of Astrocytes Induced by Methamphetamine
- Characteristics of Stable Hydrogen and Oxygen Isotopes of Soil Moisture under Different Land Use in Dry Hot Valley of Yuanmou
- Specific, highly sensitive and simple spectrofluorimetric method for quantification of daclatasvir in HCV human plasma patients and in tablets dosage form
- Chromium-modified cobalt molybdenum nitrides as catalysts for ammonia synthesis
- Langerhans cell-like dendritic cells treated with ginsenoside Rh2 regulate the differentiation of Th1 and Th2 cells in vivo
- Identification of Powdery Mildew Blumeria graminis f. sp. tritici Resistance Genes in Selected Wheat Varieties and Development of Multiplex PCR
- Computational Analysis of new Degree-based descriptors of oxide networks
- The Use Of Chemical Composition And Additives To Classify Petrol And Diesel Using Gas Chromatography–Mass Spectrometry And Chemometric Analysis: A Uk Study
- Minimal Energy Tree with 4 Branched Vertices
- Jatropha seed oil derived poly(esteramide-urethane)/ fumed silica nanocomposite coatings for corrosion protection
- Calculating topological indices of certain OTIS interconnection networks
- Energy storage analysis of R125 in UIO-66 and MOF-5 nanoparticles: A molecular simulation study
- Velvet Antler compounds targeting major cell signaling pathways in osteosarcoma - a new insight into mediating the process of invasion and metastasis in OS
- Effects of Azadirachta Indica Leaf Extract, Capping Agents, on the Synthesis of Pure And Cu Doped ZnO-Nanoparticles: A Green Approach and Microbial Activity
- Aqueous Micro-hydration of Na+(H2O)n=1-7 Clusters: DFT Study
- A proposed image-based detection of methamidophos pesticide using peroxyoxalate chemiluminescence system
- Phytochemical screening and estrogenic activity of total glycosides of Cistanche deserticola
- Biological evaluation of a series of benzothiazole derivatives as mosquitocidal agents
- Chemical pretreatments of Trapa bispinosa's peel (TBP) biosorbent to enhance adsorption capacity for Pb(ll)
- Dynamic Changes in MMP1 and TIMP1 in the Antifibrotic Process of Dahuang Zhechong Pill in Rats with Liver Fibrosis
- The Optimization and Production of Ginkgolide B Lipid Microemulsion
- Photodynamic Therapy Enhanced the Antitumor Effects of Berberine on HeLa Cells
- Chiral and Achiral Enantiomeric Separation of (±)-Alprenolol
- Correlation of Water Fluoride with Body Fluids, Dental Fluorosis and FT4, FT3 –TSH Disruption among Children in an Endemic Fluorosis area in Pakistan
- A one-step incubation ELISA kit for rapid determination of dibutyl phthalate in water, beverage and liquor
- Free Radical Scavenging Activity of Essential Oil of Eugenia caryophylata from Amboina Island and Derivatives of Eugenol
- Effects of Blue and Red Light On Growth And Nitrate Metabolism In Pakchoi
- miRNA-199a-5p functions as a tumor suppressor in prolactinomas
- Solar photodegradation of carbamazepine from aqueous solutions using a compound parabolic concentrator equipped with a sun tracking system
- Influence of sub-inhibitory concentration of selected plant essential oils on the physical and biochemical properties of Pseudomonas orientalis
- Preparation and spectroscopic studies of Fe(II), Ru(II), Pd(II) and Zn(II) complexes of Schiff base containing terephthalaldehyde and their transfer hydrogenation and Suzuki-Miyaura coupling reaction
- Complex formation in a liquid-liquid extraction-chromogenic system for vanadium(IV)
- Synthesis, characterization (IR, 1H, 13C & 31P NMR), fungicidal, herbicidal and molecular docking evaluation of steroid phosphorus compounds
- Analysis and Biological Evaluation of Arisaema Amuremse Maxim Essential Oil
- A preliminary assessment of potential ecological risk and soil contamination by heavy metals around a cement factory, western Saudi Arabia
- Anti- inflammatory effect of Prunus tomentosa Thunb total flavones in LPS-induced RAW264.7 cells
- Collaborative Influence of Elevated CO2 Concentration and High Temperature on Potato Biomass Accumulation and Characteristics
- Methods of extraction, physicochemical properties of alginates and their applications in biomedical field – a review
- Characteristics of liposomes derived from egg yolk
- Preparation of ternary ZnO/Ag/cellulose and its enhanced photocatalytic degradation property on phenol and benzene in VOCs
- Influence of Human Serum Albumin Glycation on the Binding Affinities for Natural Flavonoids
- Synthesis and antioxidant activity of 2-methylthio-pyrido[3,2-e][1,2,4] triazolo[1,5-a]pyrimidines
- Comparative study on the antioxidant activities of ten common flower teas from China
- Molecular Properties of Symmetrical Networks Using Topological Polynomials
- Synthesis of Co3O4 Nano Aggregates by Co-precipitation Method and its Catalytic and Fuel Additive Applications
- Phytochemical analysis, Antioxidant and Antiprotoscolices potential of ethanol extracts of selected plants species against Echinococcus granulosus: In-vitro study
- Silver nanoparticles enhanced fluorescence for sensitive determination of fluoroquinolones in water solutions
- Simultaneous Quantification of the New Psychoactive Substances 3-FMC, 3-FPM, 4-CEC, and 4-BMC in Human Blood using GC-MS
- Biodiesel Production by Lipids From Indonesian strain of Microalgae Chlorella vulgaris
- Miscibility studies of polystyrene/polyvinyl chloride blend in presence of organoclay
- Antibacterial Activities of Transition Metal complexes of Mesocyclic Amidine 1,4-diazacycloheptane (DACH)
- Novel 1,8-Naphthyridine Derivatives: Design, Synthesis and in vitro screening of their cytotoxic activity against MCF7 cell line
- Investigation of Stress Corrosion Cracking Behaviour of Mg-Al-Zn Alloys in Different pH Environments by SSRT Method
- Various Combinations of Flame Retardants for Poly (vinyl chloride)
- Phenolic compounds and biological activities of rye (Secale cereale L.) grains
- Oxidative degradation of gentamicin present in water by an electro-Fenton process and biodegradability improvement
- Optimizing Suitable Conditions for the Removal of Ammonium Nitrogen by a Microbe Isolated from Chicken Manure
- Anti-inflammatory, antipyretic, analgesic, and antioxidant activities of Haloxylon salicornicum aqueous fraction
- The anti-corrosion behaviour of Satureja montana L. extract on iron in NaCl solution
- Interleukin-4, hemopexin, and lipoprotein-associated phospholipase A2 are significantly increased in patients with unstable carotid plaque
- A comparative study of the crystal structures of 2-(4-(2-(4-(3-chlorophenyl)pipera -zinyl)ethyl) benzyl)isoindoline-1,3-dione by synchrotron radiation X-ray powder diffraction and single-crystal X-ray diffraction
- Conceptual DFT as a Novel Chemoinformatics Tool for Studying the Chemical Reactivity Properties of the Amatoxin Family of Fungal Peptides
- Occurrence of Aflatoxin M1 in Milk-based Mithae samples from Pakistan
- Kinetics of Iron Removal From Ti-Extraction Blast Furnace Slag by Chlorination Calcination
- Increasing the activity of DNAzyme based on the telomeric sequence: 2’-OMe-RNA and LNA modifications
- Exploring the optoelectronic properties of a chromene-appended pyrimidone derivative for photovoltaic applications
- Effect of He Qi San on DNA Methylation in Type 2 Diabetes Mellitus Patients with Phlegm-blood Stasis Syndrome
- Cyclodextrin potentiometric sensors based on selective recognition sites for procainamide: Comparative and theoretical study
- Greener synthesis of dimethyl carbonate from carbon dioxide and methanol using a tunable ionic liquid catalyst
- Nonisothermal Cold Crystallization Kinetics of Poly(lactic acid)/Bacterial Poly(hydroxyoctanoate) (PHO)/Talc
- Enhanced adsorption of sulfonamide antibiotics in water by modified biochar derived from bagasse
- Study on the Mechanism of Shugan Xiaozhi Fang on Cells with Non-alcoholic Fatty Liver Disease
- Comparative Effects of Salt and Alkali Stress on Antioxidant System in Cotton (Gossypium Hirsutum L.) Leaves
- Optimization of chromatographic systems for analysis of selected psychotropic drugs and their metabolites in serum and saliva by HPLC in order to monitor therapeutic drugs
- Electrocatalytic Properties of Ni-Doped BaFe12O19 for Oxygen Evolution in Alkaline Solution
- Study on the removal of high contents of ammonium from piggery wastewater by clinoptilolite and the corresponding mechanisms
- Phytochemistry and toxicological assessment of Bryonia dioica roots used in north-African alternative medicine
- The essential oil composition of selected Hemerocallis cultivars and their biological activity
- Mechanical Properties of Carbon Fiber Reinforced Nanocrystalline Nickel Composite Electroforming Deposit
- Anti-c-myc efficacy block EGFL7 induced prolactinoma tumorigenesis
- Topical Issue on Applications of Mathematics in Chemistry
- Zagreb Connection Number Index of Nanotubes and Regular Hexagonal Lattice
- The Sanskruti index of trees and unicyclic graphs
-
Valency-based molecular descriptors of Bakelite network
- Computing Topological Indices for Para-Line Graphs of Anthracene
- Zagreb Polynomials and redefined Zagreb indices of Dendrimers and Polyomino Chains
- Topological Descriptor of 2-Dimensional Silicon Carbons and Their Applications
- Topological invariants for the line graphs of some classes of graphs
- Words for maximal Subgroups of Fi24‘
- Generators of Maximal Subgroups of Harada-Norton and some Linear Groups
- Special Issue on POKOCHA 2018
- Influence of Production Parameters on the Content of Polyphenolic Compounds in Extruded Porridge Enriched with Chokeberry Fruit (Aronia melanocarpa (Michx.) Elliott)
- Effects of Supercritical Carbon Dioxide Extraction (SC-CO2) on the content of tiliroside in the extracts from Tilia L. flowers
- Impact of xanthan gum addition on phenolic acids composition and selected properties of new gluten-free maize-field bean pasta
- Impact of storage temperature and time on Moldavian dragonhead oil – spectroscopic and chemometric analysis
- The effect of selected substances on the stability of standard solutions in voltammetric analysis of ascorbic acid in fruit juices
- Determination of the content of Pb, Cd, Cu, Zn in dairy products from various regions of Poland
- Special Issue on IC3PE 2018 Conference
- The Photocatalytic Activity of Zns-TiO2 on a Carbon Fiber Prepared by Chemical Bath Deposition
- N-octyl chitosan derivatives as amphiphilic carrier agents for herbicide formulations
- Kinetics and Mechanistic Study of Hydrolysis of Adenosine Monophosphate Disodium Salt (AMPNa2) in Acidic and Alkaline Media
- Antimalarial Activity of Andrographis Paniculata Ness‘s N-hexane Extract and Its Major Compounds
- Special Issue on ABB2018 Conference
- Special Issue on ICCESEN 2017
- Theoretical Diagnostics of Second and Third-order Hyperpolarizabilities of Several Acid Derivatives
- Determination of Gamma Rays Efficiency Against Rhizoctonia solani in Potatoes
- Studies On Compatibilization Of Recycled Polyethylene/Thermoplastic Starch Blends By Using Different Compatibilizer
- Liquid−Liquid Extraction of Linalool from Methyl Eugenol with 1-Ethyl-3-methylimidazolium Hydrogen Sulfate [EMIM][HSO4] Ionic Liquid
- Synthesis of Graphene Oxide Through Ultrasonic Assisted Electrochemical Exfoliation
- Special Issue on ISCMP 2018
- Synthesis and antiproliferative evaluation of some 1,4-naphthoquinone derivatives against human cervical cancer cells
- The influence of the grafted aryl groups on the solvation properties of the graphyne and graphdiyne - a MD study
- Electrochemical modification of platinum and glassy carbon surfaces with pyridine layers and their use as complexing agents for copper (II) ions
- Effect of Electrospinning Process on Total Antioxidant Activity of Electrospun Nanofibers Containing Grape Seed Extract
- Effect Of Thermal Treatment Of Trepel At Temperature Range 800-1200˚C
- Topical Issue on Agriculture
- The effect of Cladophora glomerata exudates on the amino acid composition of Cladophora fracta and Rhizoclonium sp.
- Influence of the Static Magnetic Field and Algal Extract on the Germination of Soybean Seeds
- The use of UV-induced fluorescence for the assessment of homogeneity of granular mixtures
- The use of microorganisms as bio-fertilizers in the cultivation of white lupine
- Lyophilized apples on flax oil and ethyl esters of flax oil - stability and antioxidant evaluation
- Production of phosphorus biofertilizer based on the renewable materials in large laboratory scale
- Human health risk assessment of potential toxic elements in paddy soil and rice (Oryza sativa) from Ugbawka fields, Enugu, Nigeria
- Recovery of phosphates(V) from wastewaters of different chemical composition
- Special Issue on the 4th Green Chemistry 2018
- Dead zone for hydrogenation of propylene reaction carried out on commercial catalyst pellets
- Improved thermally stable oligoetherols from 6-aminouracil, ethylene carbonate and boric acid
- The role of a chemical loop in removal of hazardous contaminants from coke oven wastewater during its treatment
- Combating paraben pollution in surface waters with a variety of photocatalyzed systems: Looking for the most efficient technology
- Special Issue on Chemistry Today for Tomorrow 2019
- Applying Discriminant and Cluster Analyses to Separate Allergenic from Non-allergenic Proteins
- Chemometric Expertise Of Clinical Monitoring Data Of Prolactinoma Patients
- Chemomertic Risk Assessment of Soil Pollution
- New composite sorbent for speciation analysis of soluble chromium in textiles
- Photocatalytic activity of NiFe2O4 and Zn0.5Ni0.5Fe2O4 modified by Eu(III) and Tb(III) for decomposition of Malachite Green
- Photophysical and antibacterial activity of light-activated quaternary eosin Y
- Spectral properties and biological activity of La(III) and Nd(III) Monensinates
- Special Issue on Monitoring, Risk Assessment and Sustainable Management for the Exposure to Environmental Toxins
- Soil organic carbon mineralization in relation to microbial dynamics in subtropical red soils dominated by differently sized aggregates
- A potential reusable fluorescent aptasensor based on magnetic nanoparticles for ochratoxin A analysis
- Special Issue on 13th JCC 2018
- Fluorescence study of 5-nitroisatin Schiff base immobilized on SBA-15 for sensing Fe3+
- Thermal and Morphology Properties of Cellulose Nanofiber from TEMPO-oxidized Lower part of Empty Fruit Bunches (LEFB)
- Encapsulation of Vitamin C in Sesame Liposomes: Computational and Experimental Studies
- A comparative study of the utilization of synthetic foaming agent and aluminum powder as pore-forming agents in lightweight geopolymer synthesis
- Synthesis of high surface area mesoporous silica SBA-15 by adjusting hydrothermal treatment time and the amount of polyvinyl alcohol
- Review of large-pore mesostructured cellular foam (MCF) silica and its applications
- Ion Exchange of Benzoate in Ni-Al-Benzoate Layered Double Hydroxide by Amoxicillin
- Synthesis And Characterization Of CoMo/Mordenite Catalyst For Hydrotreatment Of Lignin Compound Models
- Production of Biodiesel from Nyamplung (Calophyllum inophyllum L.) using Microwave with CaO Catalyst from Eggshell Waste: Optimization of Transesterification Process Parameters
- The Study of the Optical Properties of C60 Fullerene in Different Organic Solvents
- Composite Material Consisting of HKUST-1 and Indonesian Activated Natural Zeolite and its Application in CO2 Capture
- Topical Issue on Environmental Chemistry
- Ionic liquids modified cobalt/ZSM-5 as a highly efficient catalyst for enhancing the selectivity towards KA oil in the aerobic oxidation of cyclohexane
- Application of Thermal Resistant Gemini Surfactants in Highly Thixotropic Water-in-oil Drilling Fluid System
- Screening Study on Rheological Behavior and Phase Transition Point of Polymer-containing Fluids produced under the Oil Freezing Point Temperature
- The Chemical Softening Effect and Mechanism of Low Rank Coal Soaked in Alkaline Solution
- The Influence Of NO/O2 On The NOx Storage Properties Over A Pt-Ba-Ce/γ-Al2O3 Catalyst
- Special Issue on the International conference CosCI 2018
- Design of SiO2/TiO2 that Synergistically Increases The Hydrophobicity of Methyltrimethoxysilane Coated Glass
- Antidiabetes and Antioxidant agents from Clausena excavata root as medicinal plant of Myanmar
- Development of a Gold Immunochromatographic Assay Method Using Candida Biofilm Antigen as a Bioreceptor for Candidiasis in Rats
- Special Issue on Applied Biochemistry and Biotechnology 2019
- Adsorption of copper ions on Magnolia officinalis residues after solid-phase fermentation with Phanerochaete chrysosporium
- Erratum
- Erratum to: Sand Dune Characterization For Preparing Metallurgical Grade Silicon
Articles in the same Issue
- Regular Articles
- Research on correlation of compositions with oestrogenic activity of Cistanche based on LC/Q-TOF-MS/MS technology
- Efficacy of Pyrus elaeagnifolia subsp. elaeagnifolia in acetic acid–induced colitis model
- Anti-inflammatory and antinociceptive features of Bryonia alba L.: As a possible alternative in treating rheumatism
- High efficiency liposome fusion induced by reducing undesired membrane peptides interaction
- Prediction of the Blood-Brain Barrier Permeability Using RP-18 Thin Layer Chromatography
- Phytic Acid Extracted from Rice Bran as a Growth Promoter for Euglena gracilis
- Development of a validated spectrofluorimetric method for assay of sotalol hydrochloride in tablets and human plasma: application for stability-indicating studies
- Topological Indices of Hyaluronic Acid-Paclitaxel Conjugates’ Molecular Structure in Cancer Treatment
- Thermodynamic properties of the bubble growth process in a pool boiling of water-ethanol mixture two-component system
- Critical Roles of the PI3K-Akt-mTOR Signaling Pathway in Apoptosis and Autophagy of Astrocytes Induced by Methamphetamine
- Characteristics of Stable Hydrogen and Oxygen Isotopes of Soil Moisture under Different Land Use in Dry Hot Valley of Yuanmou
- Specific, highly sensitive and simple spectrofluorimetric method for quantification of daclatasvir in HCV human plasma patients and in tablets dosage form
- Chromium-modified cobalt molybdenum nitrides as catalysts for ammonia synthesis
- Langerhans cell-like dendritic cells treated with ginsenoside Rh2 regulate the differentiation of Th1 and Th2 cells in vivo
- Identification of Powdery Mildew Blumeria graminis f. sp. tritici Resistance Genes in Selected Wheat Varieties and Development of Multiplex PCR
- Computational Analysis of new Degree-based descriptors of oxide networks
- The Use Of Chemical Composition And Additives To Classify Petrol And Diesel Using Gas Chromatography–Mass Spectrometry And Chemometric Analysis: A Uk Study
- Minimal Energy Tree with 4 Branched Vertices
- Jatropha seed oil derived poly(esteramide-urethane)/ fumed silica nanocomposite coatings for corrosion protection
- Calculating topological indices of certain OTIS interconnection networks
- Energy storage analysis of R125 in UIO-66 and MOF-5 nanoparticles: A molecular simulation study
- Velvet Antler compounds targeting major cell signaling pathways in osteosarcoma - a new insight into mediating the process of invasion and metastasis in OS
- Effects of Azadirachta Indica Leaf Extract, Capping Agents, on the Synthesis of Pure And Cu Doped ZnO-Nanoparticles: A Green Approach and Microbial Activity
- Aqueous Micro-hydration of Na+(H2O)n=1-7 Clusters: DFT Study
- A proposed image-based detection of methamidophos pesticide using peroxyoxalate chemiluminescence system
- Phytochemical screening and estrogenic activity of total glycosides of Cistanche deserticola
- Biological evaluation of a series of benzothiazole derivatives as mosquitocidal agents
- Chemical pretreatments of Trapa bispinosa's peel (TBP) biosorbent to enhance adsorption capacity for Pb(ll)
- Dynamic Changes in MMP1 and TIMP1 in the Antifibrotic Process of Dahuang Zhechong Pill in Rats with Liver Fibrosis
- The Optimization and Production of Ginkgolide B Lipid Microemulsion
- Photodynamic Therapy Enhanced the Antitumor Effects of Berberine on HeLa Cells
- Chiral and Achiral Enantiomeric Separation of (±)-Alprenolol
- Correlation of Water Fluoride with Body Fluids, Dental Fluorosis and FT4, FT3 –TSH Disruption among Children in an Endemic Fluorosis area in Pakistan
- A one-step incubation ELISA kit for rapid determination of dibutyl phthalate in water, beverage and liquor
- Free Radical Scavenging Activity of Essential Oil of Eugenia caryophylata from Amboina Island and Derivatives of Eugenol
- Effects of Blue and Red Light On Growth And Nitrate Metabolism In Pakchoi
- miRNA-199a-5p functions as a tumor suppressor in prolactinomas
- Solar photodegradation of carbamazepine from aqueous solutions using a compound parabolic concentrator equipped with a sun tracking system
- Influence of sub-inhibitory concentration of selected plant essential oils on the physical and biochemical properties of Pseudomonas orientalis
- Preparation and spectroscopic studies of Fe(II), Ru(II), Pd(II) and Zn(II) complexes of Schiff base containing terephthalaldehyde and their transfer hydrogenation and Suzuki-Miyaura coupling reaction
- Complex formation in a liquid-liquid extraction-chromogenic system for vanadium(IV)
- Synthesis, characterization (IR, 1H, 13C & 31P NMR), fungicidal, herbicidal and molecular docking evaluation of steroid phosphorus compounds
- Analysis and Biological Evaluation of Arisaema Amuremse Maxim Essential Oil
- A preliminary assessment of potential ecological risk and soil contamination by heavy metals around a cement factory, western Saudi Arabia
- Anti- inflammatory effect of Prunus tomentosa Thunb total flavones in LPS-induced RAW264.7 cells
- Collaborative Influence of Elevated CO2 Concentration and High Temperature on Potato Biomass Accumulation and Characteristics
- Methods of extraction, physicochemical properties of alginates and their applications in biomedical field – a review
- Characteristics of liposomes derived from egg yolk
- Preparation of ternary ZnO/Ag/cellulose and its enhanced photocatalytic degradation property on phenol and benzene in VOCs
- Influence of Human Serum Albumin Glycation on the Binding Affinities for Natural Flavonoids
- Synthesis and antioxidant activity of 2-methylthio-pyrido[3,2-e][1,2,4] triazolo[1,5-a]pyrimidines
- Comparative study on the antioxidant activities of ten common flower teas from China
- Molecular Properties of Symmetrical Networks Using Topological Polynomials
- Synthesis of Co3O4 Nano Aggregates by Co-precipitation Method and its Catalytic and Fuel Additive Applications
- Phytochemical analysis, Antioxidant and Antiprotoscolices potential of ethanol extracts of selected plants species against Echinococcus granulosus: In-vitro study
- Silver nanoparticles enhanced fluorescence for sensitive determination of fluoroquinolones in water solutions
- Simultaneous Quantification of the New Psychoactive Substances 3-FMC, 3-FPM, 4-CEC, and 4-BMC in Human Blood using GC-MS
- Biodiesel Production by Lipids From Indonesian strain of Microalgae Chlorella vulgaris
- Miscibility studies of polystyrene/polyvinyl chloride blend in presence of organoclay
- Antibacterial Activities of Transition Metal complexes of Mesocyclic Amidine 1,4-diazacycloheptane (DACH)
- Novel 1,8-Naphthyridine Derivatives: Design, Synthesis and in vitro screening of their cytotoxic activity against MCF7 cell line
- Investigation of Stress Corrosion Cracking Behaviour of Mg-Al-Zn Alloys in Different pH Environments by SSRT Method
- Various Combinations of Flame Retardants for Poly (vinyl chloride)
- Phenolic compounds and biological activities of rye (Secale cereale L.) grains
- Oxidative degradation of gentamicin present in water by an electro-Fenton process and biodegradability improvement
- Optimizing Suitable Conditions for the Removal of Ammonium Nitrogen by a Microbe Isolated from Chicken Manure
- Anti-inflammatory, antipyretic, analgesic, and antioxidant activities of Haloxylon salicornicum aqueous fraction
- The anti-corrosion behaviour of Satureja montana L. extract on iron in NaCl solution
- Interleukin-4, hemopexin, and lipoprotein-associated phospholipase A2 are significantly increased in patients with unstable carotid plaque
- A comparative study of the crystal structures of 2-(4-(2-(4-(3-chlorophenyl)pipera -zinyl)ethyl) benzyl)isoindoline-1,3-dione by synchrotron radiation X-ray powder diffraction and single-crystal X-ray diffraction
- Conceptual DFT as a Novel Chemoinformatics Tool for Studying the Chemical Reactivity Properties of the Amatoxin Family of Fungal Peptides
- Occurrence of Aflatoxin M1 in Milk-based Mithae samples from Pakistan
- Kinetics of Iron Removal From Ti-Extraction Blast Furnace Slag by Chlorination Calcination
- Increasing the activity of DNAzyme based on the telomeric sequence: 2’-OMe-RNA and LNA modifications
- Exploring the optoelectronic properties of a chromene-appended pyrimidone derivative for photovoltaic applications
- Effect of He Qi San on DNA Methylation in Type 2 Diabetes Mellitus Patients with Phlegm-blood Stasis Syndrome
- Cyclodextrin potentiometric sensors based on selective recognition sites for procainamide: Comparative and theoretical study
- Greener synthesis of dimethyl carbonate from carbon dioxide and methanol using a tunable ionic liquid catalyst
- Nonisothermal Cold Crystallization Kinetics of Poly(lactic acid)/Bacterial Poly(hydroxyoctanoate) (PHO)/Talc
- Enhanced adsorption of sulfonamide antibiotics in water by modified biochar derived from bagasse
- Study on the Mechanism of Shugan Xiaozhi Fang on Cells with Non-alcoholic Fatty Liver Disease
- Comparative Effects of Salt and Alkali Stress on Antioxidant System in Cotton (Gossypium Hirsutum L.) Leaves
- Optimization of chromatographic systems for analysis of selected psychotropic drugs and their metabolites in serum and saliva by HPLC in order to monitor therapeutic drugs
- Electrocatalytic Properties of Ni-Doped BaFe12O19 for Oxygen Evolution in Alkaline Solution
- Study on the removal of high contents of ammonium from piggery wastewater by clinoptilolite and the corresponding mechanisms
- Phytochemistry and toxicological assessment of Bryonia dioica roots used in north-African alternative medicine
- The essential oil composition of selected Hemerocallis cultivars and their biological activity
- Mechanical Properties of Carbon Fiber Reinforced Nanocrystalline Nickel Composite Electroforming Deposit
- Anti-c-myc efficacy block EGFL7 induced prolactinoma tumorigenesis
- Topical Issue on Applications of Mathematics in Chemistry
- Zagreb Connection Number Index of Nanotubes and Regular Hexagonal Lattice
- The Sanskruti index of trees and unicyclic graphs
-
Valency-based molecular descriptors of Bakelite network
- Computing Topological Indices for Para-Line Graphs of Anthracene
- Zagreb Polynomials and redefined Zagreb indices of Dendrimers and Polyomino Chains
- Topological Descriptor of 2-Dimensional Silicon Carbons and Their Applications
- Topological invariants for the line graphs of some classes of graphs
- Words for maximal Subgroups of Fi24‘
- Generators of Maximal Subgroups of Harada-Norton and some Linear Groups
- Special Issue on POKOCHA 2018
- Influence of Production Parameters on the Content of Polyphenolic Compounds in Extruded Porridge Enriched with Chokeberry Fruit (Aronia melanocarpa (Michx.) Elliott)
- Effects of Supercritical Carbon Dioxide Extraction (SC-CO2) on the content of tiliroside in the extracts from Tilia L. flowers
- Impact of xanthan gum addition on phenolic acids composition and selected properties of new gluten-free maize-field bean pasta
- Impact of storage temperature and time on Moldavian dragonhead oil – spectroscopic and chemometric analysis
- The effect of selected substances on the stability of standard solutions in voltammetric analysis of ascorbic acid in fruit juices
- Determination of the content of Pb, Cd, Cu, Zn in dairy products from various regions of Poland
- Special Issue on IC3PE 2018 Conference
- The Photocatalytic Activity of Zns-TiO2 on a Carbon Fiber Prepared by Chemical Bath Deposition
- N-octyl chitosan derivatives as amphiphilic carrier agents for herbicide formulations
- Kinetics and Mechanistic Study of Hydrolysis of Adenosine Monophosphate Disodium Salt (AMPNa2) in Acidic and Alkaline Media
- Antimalarial Activity of Andrographis Paniculata Ness‘s N-hexane Extract and Its Major Compounds
- Special Issue on ABB2018 Conference
- Special Issue on ICCESEN 2017
- Theoretical Diagnostics of Second and Third-order Hyperpolarizabilities of Several Acid Derivatives
- Determination of Gamma Rays Efficiency Against Rhizoctonia solani in Potatoes
- Studies On Compatibilization Of Recycled Polyethylene/Thermoplastic Starch Blends By Using Different Compatibilizer
- Liquid−Liquid Extraction of Linalool from Methyl Eugenol with 1-Ethyl-3-methylimidazolium Hydrogen Sulfate [EMIM][HSO4] Ionic Liquid
- Synthesis of Graphene Oxide Through Ultrasonic Assisted Electrochemical Exfoliation
- Special Issue on ISCMP 2018
- Synthesis and antiproliferative evaluation of some 1,4-naphthoquinone derivatives against human cervical cancer cells
- The influence of the grafted aryl groups on the solvation properties of the graphyne and graphdiyne - a MD study
- Electrochemical modification of platinum and glassy carbon surfaces with pyridine layers and their use as complexing agents for copper (II) ions
- Effect of Electrospinning Process on Total Antioxidant Activity of Electrospun Nanofibers Containing Grape Seed Extract
- Effect Of Thermal Treatment Of Trepel At Temperature Range 800-1200˚C
- Topical Issue on Agriculture
- The effect of Cladophora glomerata exudates on the amino acid composition of Cladophora fracta and Rhizoclonium sp.
- Influence of the Static Magnetic Field and Algal Extract on the Germination of Soybean Seeds
- The use of UV-induced fluorescence for the assessment of homogeneity of granular mixtures
- The use of microorganisms as bio-fertilizers in the cultivation of white lupine
- Lyophilized apples on flax oil and ethyl esters of flax oil - stability and antioxidant evaluation
- Production of phosphorus biofertilizer based on the renewable materials in large laboratory scale
- Human health risk assessment of potential toxic elements in paddy soil and rice (Oryza sativa) from Ugbawka fields, Enugu, Nigeria
- Recovery of phosphates(V) from wastewaters of different chemical composition
- Special Issue on the 4th Green Chemistry 2018
- Dead zone for hydrogenation of propylene reaction carried out on commercial catalyst pellets
- Improved thermally stable oligoetherols from 6-aminouracil, ethylene carbonate and boric acid
- The role of a chemical loop in removal of hazardous contaminants from coke oven wastewater during its treatment
- Combating paraben pollution in surface waters with a variety of photocatalyzed systems: Looking for the most efficient technology
- Special Issue on Chemistry Today for Tomorrow 2019
- Applying Discriminant and Cluster Analyses to Separate Allergenic from Non-allergenic Proteins
- Chemometric Expertise Of Clinical Monitoring Data Of Prolactinoma Patients
- Chemomertic Risk Assessment of Soil Pollution
- New composite sorbent for speciation analysis of soluble chromium in textiles
- Photocatalytic activity of NiFe2O4 and Zn0.5Ni0.5Fe2O4 modified by Eu(III) and Tb(III) for decomposition of Malachite Green
- Photophysical and antibacterial activity of light-activated quaternary eosin Y
- Spectral properties and biological activity of La(III) and Nd(III) Monensinates
- Special Issue on Monitoring, Risk Assessment and Sustainable Management for the Exposure to Environmental Toxins
- Soil organic carbon mineralization in relation to microbial dynamics in subtropical red soils dominated by differently sized aggregates
- A potential reusable fluorescent aptasensor based on magnetic nanoparticles for ochratoxin A analysis
- Special Issue on 13th JCC 2018
- Fluorescence study of 5-nitroisatin Schiff base immobilized on SBA-15 for sensing Fe3+
- Thermal and Morphology Properties of Cellulose Nanofiber from TEMPO-oxidized Lower part of Empty Fruit Bunches (LEFB)
- Encapsulation of Vitamin C in Sesame Liposomes: Computational and Experimental Studies
- A comparative study of the utilization of synthetic foaming agent and aluminum powder as pore-forming agents in lightweight geopolymer synthesis
- Synthesis of high surface area mesoporous silica SBA-15 by adjusting hydrothermal treatment time and the amount of polyvinyl alcohol
- Review of large-pore mesostructured cellular foam (MCF) silica and its applications
- Ion Exchange of Benzoate in Ni-Al-Benzoate Layered Double Hydroxide by Amoxicillin
- Synthesis And Characterization Of CoMo/Mordenite Catalyst For Hydrotreatment Of Lignin Compound Models
- Production of Biodiesel from Nyamplung (Calophyllum inophyllum L.) using Microwave with CaO Catalyst from Eggshell Waste: Optimization of Transesterification Process Parameters
- The Study of the Optical Properties of C60 Fullerene in Different Organic Solvents
- Composite Material Consisting of HKUST-1 and Indonesian Activated Natural Zeolite and its Application in CO2 Capture
- Topical Issue on Environmental Chemistry
- Ionic liquids modified cobalt/ZSM-5 as a highly efficient catalyst for enhancing the selectivity towards KA oil in the aerobic oxidation of cyclohexane
- Application of Thermal Resistant Gemini Surfactants in Highly Thixotropic Water-in-oil Drilling Fluid System
- Screening Study on Rheological Behavior and Phase Transition Point of Polymer-containing Fluids produced under the Oil Freezing Point Temperature
- The Chemical Softening Effect and Mechanism of Low Rank Coal Soaked in Alkaline Solution
- The Influence Of NO/O2 On The NOx Storage Properties Over A Pt-Ba-Ce/γ-Al2O3 Catalyst
- Special Issue on the International conference CosCI 2018
- Design of SiO2/TiO2 that Synergistically Increases The Hydrophobicity of Methyltrimethoxysilane Coated Glass
- Antidiabetes and Antioxidant agents from Clausena excavata root as medicinal plant of Myanmar
- Development of a Gold Immunochromatographic Assay Method Using Candida Biofilm Antigen as a Bioreceptor for Candidiasis in Rats
- Special Issue on Applied Biochemistry and Biotechnology 2019
- Adsorption of copper ions on Magnolia officinalis residues after solid-phase fermentation with Phanerochaete chrysosporium
- Erratum
- Erratum to: Sand Dune Characterization For Preparing Metallurgical Grade Silicon