lncRNA WT1-AS attenuates hypoxia/ischemia-induced neuronal injury during cerebral ischemic stroke via miR-186-5p/XIAP axis
-
Jianquan You
 und
Bin Gu
und
Bin Gu

Abstract
This study aimed to investigate the role and mechanism of long non-coding RNA (lncRNA) WT1 antisense RNA (WT1-AS) in cerebral ischemic stroke. The Starbase database and dual-luciferase reporter gene assay were used to analyze the interaction between lncRNA WT1 antisense RNA (lncRNA WT1-AS) and microRNA-186-5p (miR-186-5p). Reverse transcription-quantitative PCR analysis was performed to determine lncRNA WT1-AS and miR-186-5p levels. An oxygen glucose deprivation (OGD)-induced SH-SY5Y cell injury model was established. Cell viability and apoptosis were determined using 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide and flow cytometric assays, respectively. Caspase 3 activity was evaluated using a caspase 3 activity detection kit. The results showed that miR-186-5p is a direct target of the lncRNA WT1-AS. In addition, lncRNA WT1-AS levels were downregulated and miR-186-5p levels were upregulated in the blood samples of patients with ischemic stroke and OGD-induced SH-SY5Y cells. WT1-AS overexpression promoted OGD-induced cell viability and reduced the cell apoptosis and caspase 3 activity. However, these effects were reversed by miR-186-5p overexpression. Furthermore, the results demonstrated that the X-linked inhibitor of apoptosis (XIAP) was directly targeted by miR-186-5p. Similarly, transfection with the miR-186-5p inhibitor reduced OGD-induced neuronal damage by upregulating XIAP expression. In conclusion, lncRNA WT1-AS attenuates hypoxia/ischemia-induced neuronal injury in cerebral ischemic stroke through the miR-186-5p/XIAP axis.
1 Introduction
Stroke is the leading cause of cerebral dysfunction and mortality worldwide [1]. Approximately 795,000 new cases of stroke are recorded in the United States annually, while more than 140,000 individuals die from stroke [2]. Eighty percent of stroke cases are attributed to ischemic stroke [3]. The main features of ischemic stroke include interruption of blood flow and lack of oxygen and glucose supply in the brain cells, eventually leading to impaired neuronal cell function. Current treatment approaches for stroke mainly focus on stimulating nerve replacement and timely resolving the infarction to limit continuous hypoxic stress-mediated damage, which eventually results in neuronal injury [4]. To date, the mechanisms and treatment strategies for ischemia/hypoxia-induced neuronal damage remain elusive. Recently, numerous studies have shown that long non-coding RNAs (lncRNAs) and miRNAs are involved in mechanisms underlying ischemia/hypoxia-induced neuronal damage.
lncRNAs are approximately 200 nucleotides in length. Among lncRNAs, genomic DNA can be transcribed into RNA, but it cannot be translated into proteins [5]. lncRNAs play crucial roles in several biological processes, including the regulation of the cell cycle and differentiation and epigenetic regulation [6,7]. The roles of various lncRNAs in different diseases have been extensively reported. Several studies have shown that lncRNA expression is upregulated or downregulated in several diseases, including cancer, neurological diseases, and diabetes [8,9]. WT1 antisense RNA (WT1-AS) encodes a zinc finger transcription domain [10]. Emerging evidence has shown that WT1-AS is involved in the onset of numerous types of cancers, including breast cancer, non-small-cell lung cancer, cervical cancer, and glioma [11,12,13,14]. This study aimed to investigate the role of WT1-AS in cerebral ischemic stroke.
miRNA is a general term for a class of small non-coding RNAs, ∼20–22 nucleotides in length, that are not translated into proteins and inhibit the expression of their target genes [15]. microRNA-186-5p (miR-186-5p), a cancer-related miRNA, is involved in the occurrence and development of several types of cancers [16,17]. Zhu et al. [16] showed that miR-186-5p acts as a tumor suppressor gene that is downregulated in neuroblastoma. In addition, Jones et al. [17] demonstrated that miR-186-5p silencing inhibits the proliferation, growth, and invasion of metastatic cancer cells. Tao et al. [18] indicated that miR-186-5p may be involved in the development of atherosclerosis. However, the role of miR-186-5p in cerebral ischemic stroke has not yet been investigated. Therefore, the current study aimed to investigate whether lncRNA WT1 antisense RNA (lncRNA WT1-AS), miR-186-5p, and X-linked inhibitor of apoptosis (XIAP) could interact with each other and to uncover the underlying mechanism of action of these molecules in cerebral ischemic stroke.
2 Methods
2.1 Clinical samples
Blood samples were obtained from 30 patients with ischemic stroke within 3 h of stroke onset. In addition, 30 blood samples collected from healthy volunteers served as the control group. The characteristics of patients are shown in Table 1. The levels of lncRNA WT1-AS and miR-186-5p in the plasma of patients with ischemic stroke and healthy volunteers were determined using reverse transcription-quantitative PCR (RT-qPCR). Inclusion criteria were as follows: patients who demonstrated new-onset cerebral infarction on magnetic resonance imaging within 3 h from the time of admission were included in the study. The exclusion criteria were evidence of prior cerebral infarcts, diabetes mellitus, coronary artery disease, hypertension, kidney diseases, circulatory disorders, or autoimmune diseases. The study protocol was approved by the Ethics Committee of Taizhou People’s Hospital. All the patients in the current study approved the use of their specimens.
Characteristics of patients
| Characteristic | Healthy control | Stroke | P value |
|---|---|---|---|
| Age (year) | 63–72 | 64–75 | >0.05 |
| Gender (male), n (%) | 15 (50.0) | 15 (50.0) | >0.05 |
| BMI (kg/m2) | 23.3 ± 0.34 | 25.1 ± 0.48 | >0.05 |
| Smoking, n (%) | 6 (20.0) | 5 (16.7) | >0.05 |
| Drinking, n (%) | 3 (10.0) | 4 (13.3) | >0.05 |
| Hypertension, n (%) | 4 (13.4) | 17 (56.7) | <0.05 |
| Diabetes mellitus, n (%) | 3 (10.0) | 4 (13.3) | >0.05 |
| Total cholesterol (mm) | 4.25 ± 0.13 | 4.35 ± 0.26 | >0.05 |
| Triglycerides (mm) | 1.53 ± 0.13 | 1.41 ± 0.12 | >0.05 |
| LDL (mm) | 2.63 ± 0.15 | 2.70 ± 0.17 | >0.05 |
| HDL (mm) | 1.14 ± 0.05 | 1.31 ± 0.07 | >0.05 |
| NIHSS score | |||
| 1–4 | 9 (30.0%) | ||
| 5–15 | 14 (46.7%) | ||
| 15–20 | 6 (20%) | ||
| 21–42 | 1 (3.3%) | ||
BMI, body mass index; HDL, high-density lipoprotein; LDL, low-density lipoprotein; NIHSS, National Institute of Health Stroke Scale.
2.2 Dual luciferase reporter assay
The wild-type (WT) or mutant (MUT) 3′-untranslated region (3′-UTR) of WT1-AS were subcloned into the pmiRGLO vector (cat. no. E1330; Promega Corporation) to assess the association between WT1-AS and miR-186-5p. Subsequently, 293T cells were co-transfected with miR-186-5p, control mimics, or the above plasmids. Following transfection for 48 h, luciferase activity was measured using a dual-luciferase assay kit (cat. no. E1910; Promega Corporation). To evaluate the association between miR-186-5p and XIAP, XIAP-WT and XIAP-MUT 3′-UTR luciferase reporter plasmids were constructed. Then, 293T cells were co-transfected with Renilla luciferase, luciferase reporter plasmids, and miR-186-5p or control mimics for 48 h. Luciferase activity was determined using a dual luciferase assay kit (Promega Corporation), according to the manufacturer’s instructions.
2.3 Cell culture and transfection
The neuroblastoma cell line, SH-SY5Y, was obtained from the American Tissue Culture Collection (cat. no. CRL-2266). Cells were cultured in Dulbecco’s modified Eagle’s medium (DMEM) supplemented with 10% fetal bovine serum (both from Gibco; Thermo Fisher Scientific, Inc.) in a cell culture incubator with 5% CO2 at 37°C. SH-SY5Y cells were then transfected with WT1-AS plasmid, control plasmid, mimic control (5′-UUCUCCGAACGUGUCACGUTT-3′; Shanghai GenePharma Co., Ltd., China), miR-186-5p mimics (5′-CAAAGAAUUCUCCUUUUGGGCU-3′; Shanghai GenePharma Co., Ltd., China), WT1-AS plasmid + mimic control, WT1-AS plasmid + miR-186-5p mimics, inhibitor control (5′-GCCUCCGGCUUCGCACCUCU-3′; Shanghai GenePharma Co., Ltd., China), miR-186-5p inhibitor (5′-AGCCCAAAAGGAGAAUUCUUUG-3′; Shanghai GenePharma Co., Ltd., China), control small interfering (si)-RNA, XIAP small-interfering RNA (siRNA), miR-186-5p inhibitor + control siRNA, or miR-186-5p inhibitor + XIAP siRNA for 48 h using Lipofectamine™ 2000 transfection reagent (cat. no. 11668019; Invitrogen; Thermo Fisher Scientific, Inc.), according to the manufacturer’s instructions. The cells were incubated for 48 h prior to subsequent experiments.
2.4 RT-qPCR assay
Total RNA was extracted from SH-SY5Y cells using TRIzol reagent (cat. no. 9108; Takara Bio, Inc.), following standard operating procedures. Total RNA was reverse transcribed into cDNA using a reverse transcriptase kit (cat no. R211-01; Vazyme Biotech Co., Ltd.). Subsequently, qPCR was performed using the SYBR Green PCR kit (cat. no. Q311-02; Vazyme Biotech Co., Ltd.), according to the manufacturer’s instructions. Glyceraldehyde-3-phosphate dehydrogenase (for mRNA) and uracil 6 (for miRNA) were used as endogenous controls. The relative gene expression was quantified using the 2−ΔΔCq method. Primer sequences are listed in Table 2.
Primer sequences for PCR
| Gene name | Sequences: 5′–3′ |
|---|---|
| lncRNA WT1-AS | Forward, 5′-GCCTCTCTGTCCTCTTCTTTG-3′ |
| Reverse, 5′-GCTGTGAGTCCTGGTGCTTA-3′ | |
| miR-186-5p | Forward, 5′-TCAAAGAATTCTCCTTTTGGGCT-3′ |
| Reverse, 5′-CGCTTCACGAATTTGCGTGTCAT-3′ | |
| GAPDH | Forward, 5′-ATCACTGCCACCCAGAAGAC-3′ |
| Reverse, 5′-TTTCTAGACGGCAGGTCAGG-3′ | |
| U6 | Forward, 5′-CTCGCTTCGGCAGCACA-3′ |
| Reverse, 5′-AACGCTTCACGAATTTGCGT-3′ | |
| XIAP | Forward, 5′-ACCGTGCGGTGCTTTAGTT-3′ |
| Reverse, 5′-TGCGTGGCACTATTTTCAAGATA-3′ |
2.5 Establishment of the oxygen glucose deprivation/reoxygenation (OGD/R) cell model
To establish an in vitro OGD/R model, the cells were cultured in glucose-free DMEM at 37°C in an atmosphere of 1% O2, 94% N2, and 5% CO2 for 4 h. Subsequently, the cells were cultured under normoxic conditions of 95% air and 5% CO2 for an additional 24 h.
2.6 Western blotting analysis
The cells were lysed with the radioimmunoprecipitation assay buffer (cat. no. R0010; Beijing Solarbio Science & Technology Co., Ltd.), and the protein concentration was measured using a bicinchoninic acid kit (cat no. 23225; Pierce; Thermo Fisher Scientific, Inc.). Equal amounts of proteins were separated via 12% sodium dodecyl sulfate-polyacrylamide gel electrophoresis and transferred onto polyvinylidene fluoride membranes. Following blocking with 5% nonfat milk to prevent nonspecific binding, the membranes were incubated with primary antibodies against XIAP (cat. no. ab21278; dilution, 1:1,000) and GAPDH (cat. no. ab9485; dilution, 1:1,000; both from Abcam) at 4°C overnight. The next day, membranes were incubated with the corresponding secondary antibodies for 2 h. Protein bands were visualized using the ECL method (cat. no. 34579; Applygen Technologies, Inc.).
2.7 Flow cytometric assay
Cell apoptosis was evaluated using the Annexin-V-fluorescein isothiocyanate (FITC)/propidium iodide (PI) Apoptosis Detection Kit (cat. no. 556570; BD Bioscience). Briefly, after treatment, the cells were collected and centrifuged at a high speed and low temperature. Following centrifugation, the supernatant was discarded and the cells were resuspended in 100 μL FITC-binding buffer. Subsequently, the cell suspension was supplemented with 5 μL ready-to-use annexin V-FITC and 5 μL PI. The cells were incubated for 30 min at room temperature in the dark. Cell apoptosis rate was assessed using a BD FACSCalibur flow cytometer (BD Technologies).
2.8 3-(4,5-Dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) assay
Following treatment, cells were seeded into a 96-well plate and incubated for 24 h. Subsequently, each well was supplemented with 20 μL MTT reagent (5 mg/mL; cat. no. CT02; Sigma-Aldrich), and the cells were cultured for an additional 4 h. The supernatant was discarded and each well was supplemented with 200 µL dimethyl sulfoxide. The absorbance of each well was measured at 570 nm wavelength.
2.9 Caspase 3 activity detection
Caspase 3 activity was assessed using the corresponding detection kit (cat. no. C1116; Beyotime Institute of Biotechnology). Briefly, transfected cells were collected into tubes and centrifuged at 600×g for 5 min at 4°C. The cells were resuspended in an appropriate volume of lysis buffer and lysed for 15 min in an ice bath. Following centrifugation for 10 min, the supernatant was transferred to a pre-cooled centrifuge tube and incubated on ice. The enzymatic activity of caspase 3 was measured immediately at a wavelength of 405 nm.
2.10 Statistical analysis
All experiments were repeated at least three times. All data were analyzed using GraphPad Prism software (version 6.0; GraphPad Software, Inc.). Statistically significant differences between two groups were evaluated using unpaired Student’s t-test, while those among multiple groups were evaluated using one-way analysis of variance followed by Tukey’s post hoc test. Data are expressed as the mean ± standard deviation from three independent experiments. P < 0.05 was considered to be statistically significant.
3 Results
3.1 lncRNA WT1-AS directly interacts with miR-186-5p
First, the association between WT1-AS and miR-186-5p was predicted using the StarBase bioinformatics tool. Bioinformatics analysis revealed that miR-186-5p and lncRNA WT1-AS shared mutual binding sites (Figure 1a), suggesting that miR-186-5p could bind to the WT1-AS 3′-UTR. Subsequently, 293T cells were cotransfected with WT1-AS-WT, WT1-AS-MUT, miR-186-5p, or mimic control for 48 h. A dual-luciferase reporter assay was then performed to evaluate the luciferase activity. The results revealed that miR-186-5p mimics inhibited the activity of WT1-AS-WT but not that of WT1-AS-MUT (Figure 1b). These findings indicate that miR-186-5p is directly targeted by the lncRNA WT1-AS. Subsequently, blood samples from 30 patients with ischemic stroke were collected within 3 h of stroke onset. RT-qPCR assay results demonstrated that, compared with the healthy control group, lncRNA WT1-AS was substantially downregulated (Figure 1c) and miR-186-5p was upregulated (Figure 1d) in the blood samples of patients with ischemic stroke.

Expression levels of lncRNA WT1-AS and miR-186-5p in patients with ischemic stroke. (a) Interaction between miR-186-5p and WT1-AS 3′-UTR was predicted using the Starbase prediction software. (b) Dual luciferase reporter gene assay was used to verify the interaction between WT1-AS and miR-186-5p in 293T cells co-transfected with miR-186-5p mimics and WT or MUT WT1-AS 3′-UTR reporter plasmids. (c and d) Expression levels of WT1-AS and miR-186-5p in blood samples of patients with ischemic stroke and healthy individuals. ** P < 0.01 vs control mimics group; ## P < 0.01 vs healthy control group. GraphPad Prism 6.0 software (GraphPad Software, Inc.) was used for creation of the figure.
3.2 lncRNA WT1-AS levels are downregulated and miR-186-5p levels are upregulated in OGD-induced SH-SY5Y cells
The results showed that, compared with the control group, the expression of lncRNA WT1-AS was significantly reduced in OGD-induced SH-SY5Y cells (Figure 2a), while miR-186-5p expression was significantly upregulated (Figure 2b).
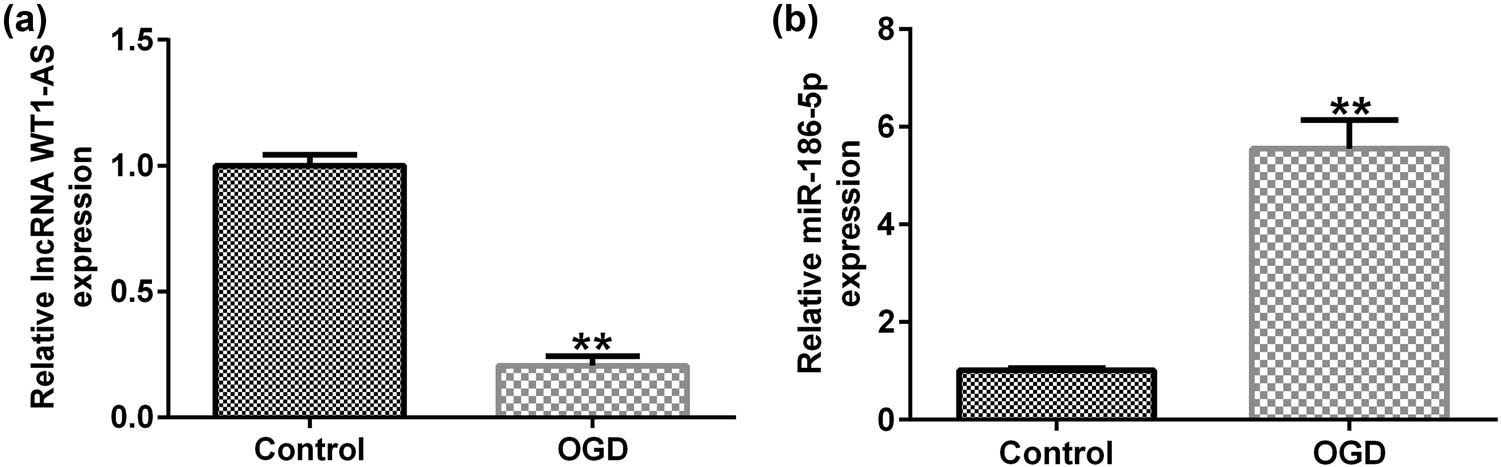
Expression levels of lncRNA WT1-AS/miR-186-5p in OGD-induced SH-SY5Y cells. (a) RT-qPCR analysis results showing lncRNA WT1-AS expression levels. (b) RT-qPCR analysis results showing miR-186-5p expression levels. ** P < 0.01 vs control group. GraphPad Prism 6.0 software (GraphPad Software, Inc.) was used for creation of the figure.
3.3 WT1-AS negatively regulates miR-186-5p levels in SH-SY5Y cells
Compared with the control plasmid group, cell transfection with the WT1-AS plasmid successfully overexpressed WT1-AS in SH-SY5Y cells (Figure 3a). In addition, compared to the mimic control group, transfection with miR-186-5p mimics notably increased miR-186-5p expression in SH-SY5Y cells (Figure 3b). There were no significant changes in the expression of WT1-AS in WT1-AS plasmid + mimic control and WT1-AS plasmid + miR-186-5p mimics groups (Figure 3c). Finally, WT1-AS overexpression inhibited miR-186-5p expression compared to the control plasmid group, while this effect was notably reversed by miR-186-5p overexpression (Figure 3d). The data indicated that WT1-AS negatively regulates miR-186-5p levels in SH-SY5Y cells.

lncRNA WT1-AS negatively regulates miR-186-5p expression levels in SH-SY5Y cells. (a) RT-qPCR analysis results showing WT1-AS expression levels in SH-SY5Y cells transfected with the control or WT1-AS-plasmids. (b) RT-qPCR analysis results showing miR-186-5p expression levels in SH-SY5Y cells transfected with the control or miR-186-5p mimics. (c) RT-qPCR analysis showing the expression levels of WT1-AS in SH-SY5Y cells transfected with the control plasmid, miR-186-5p mimics, WT1-AS plasmid + control mimics, or WT1-AS plasmid + miR-186-5p mimics. (d) RT-qPCR analysis showing the expression levels of miR-186-5p in SH-SY5Y cells transfected with the control plasmid, miR-186-5p mimics, WT1-AS plasmid + control mimics, or WT1-AS plasmid + miR-186-5p mimics. ** P < 0.01 vs control plasmid group; ## P < 0.01 vs control mimics group; && P < 0.01 vs WT1-AS plasmid + control mimics group. GraphPad Prism 6.0 software (GraphPad Software, Inc.) was used for creation of the figure.
3.4 lncRNA WT1-AS attenuates OGD-induced neuronal injury by downregulating miR-186-5p levels
Compared to the control group, miR-186-5p expression was significantly enhanced in the OGD group. In addition, the expression of miR-186-5p was significantly reduced in the OGD + WT1-AS plasmid group compared to that in the OGD + control plasmid group. However, this effect was reversed by miR-186-5p overexpression (Figure 4a). Furthermore, RT-qPCR analysis showed that compared to the control group, the expression of lncRNA WT1-AS decreased in the OGD group. In addition, compared to the OGD + control plasmid group, lncRNA WT1-AS was upregulated in the OGD + WT1-AS plasmid group (Figure 4b). MTT assay and flow cytometric analysis revealed that compared to the control group, cell viability decreased (Figure 4c) and cell apoptosis increased (Figure 4d and e), respectively, in the OGD group. Compared to the control group, the activity of caspase 3 was enhanced in the OGD group (Figure 4f). Compared to the OGD + control plasmid group, cell viability was significantly increased (Figure 4c), apoptosis (Figure 4d and e) was reduced, and caspase 3 activity was attenuated (Figure 4f) in the OGD + WT1-AS plasmid group. Furthermore, all the aforementioned effects caused by WT1-AS plasmid were reversed following miR-186-5p overexpression. These findings suggested that lncRNA WT1-AS attenuates OGD-induced neuronal injury by downregulating miR-186-5p levels.
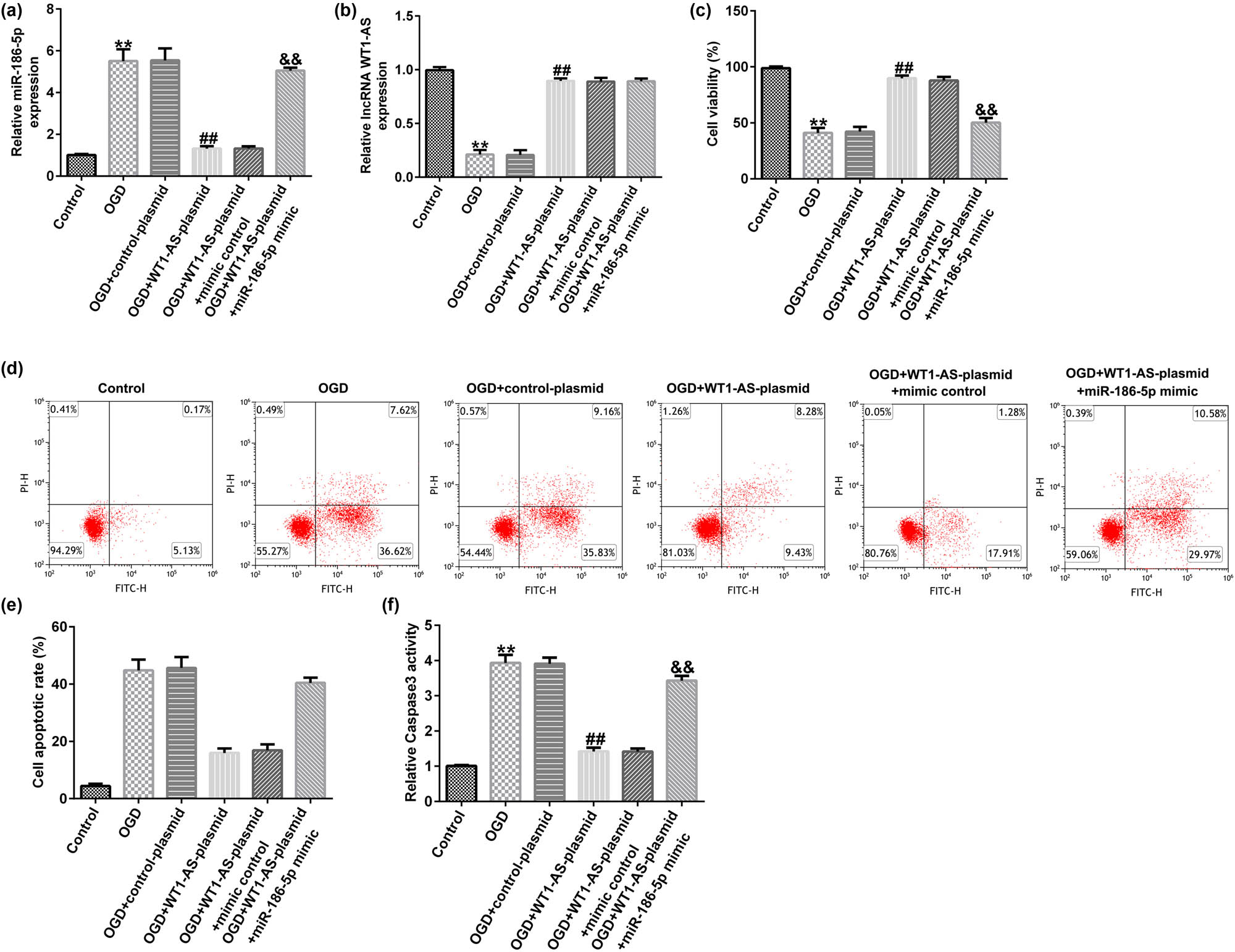
Effect of lncRNA WT1-AS on OGD-induced nerve cell injury. (a) RT-qPCR analysis results showing microRNA-186-5p expression levels. (b) RT-qPCR analysis results showing lncRNA WT1-AS expression levels. (c) A MTT assay was used to assess the cell viability. (d) A flow cytometric assay was used to evaluate cell apoptosis. (e) The apoptosis ratio is presented. (f) Caspase 3 activity was detected using the corresponding kit. ** P < 0.01 vs control group; ## P < 0.01 vs OGD + control mimics group; && P < 0.01 vs OGD + WT1-AS plasmid + control mimics group. GraphPad Prism 6.0 software (GraphPad Software, Inc.) was used for creation of the figure.
3.5 miR-186-5p directly interacts with XIAP
A literature review [19] and bioinformatic analysis using the TargetScan database identified a mutual binding site between miR-186-5p and XIAP (Figure 5a). Furthermore, a dual-luciferase reporter assay verified the association between miR-186-5p and XIAP. As shown in Figure 5b, miR-186-5p mimics inhibited the activity of XIAP-WT but not that of XIAP-MUT. Then, we determined the level of XIAP in the blood samples of patients with ischemic stroke. The results indicated that compared with the healthy control group, XIAP was significantly downregulated (Figure 5c) in the blood samples of patients with ischemic stroke. Taken together, these findings suggest that XIAP is directly targeted by miR-186-5p, and it is downregulated in the blood samples of patients with ischemic stroke.
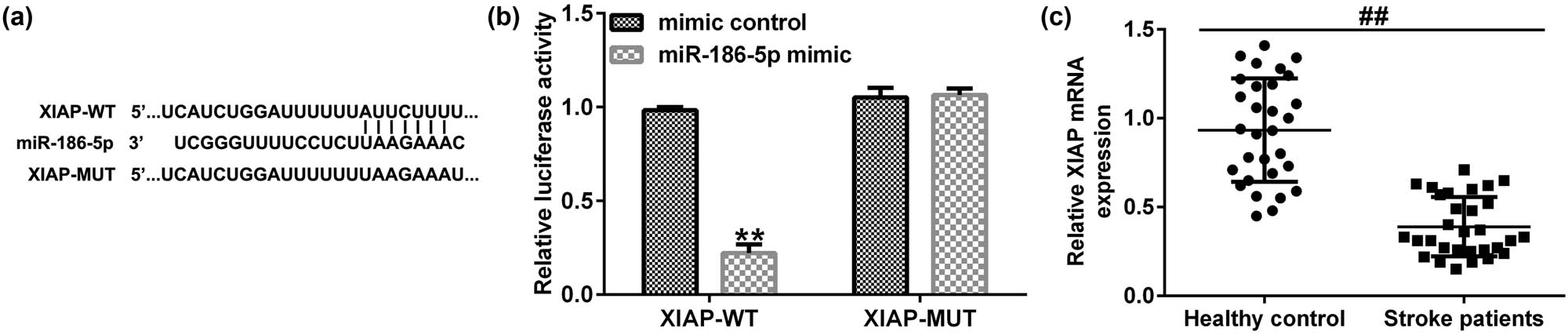
XIAP is directly targeted by miR-186-5p. (a) The interaction between miR-186-5p and XIAP 3′-UTR was predicted by TargetScan. (b) A dual luciferase reporter gene assay was performed to verify the interaction between miR-186-5p and XIAP in 293T cells co-transfected with miR-186-5p mimics and WT or MUT XIAP 3′-UTR reporter plasmids. (c) Expression levels of XIAP in blood samples of patients with ischemic stroke and healthy individuals.** P < 0.01 vs control mimics group; ## P < 0.01 vs Healthy control group. GraphPad Prism 6.0 software (GraphPad Software, Inc.) was used for creation of the figure.
3.6 miR-186-5p negatively regulated XIAP expression in SH-SY5Y cells
SH-SY5Y cells were transfected with inhibitor control, miR-186-5p inhibitor, control siRNA, XIAP siRNA, miR-186-5p inhibitor + control siRNA, or miR-186-5p inhibitor + XIAP siRNA for 24 h. RT-qPCR was performed to assess the transfection efficiency. Compared to the inhibitor control group, transfection with the miR-186-5p inhibitor significantly reduced miR-186-5p expression in SH-SY5Y cells (Figure 6a). Consistently, compared to the control siRNA group, transfection with XIAP siRNA notably attenuated XIAP expression (Figure 6b). However, miR-186-5p silencing significantly increased the mRNA and protein expression levels of XIAP compared to those in the control inhibitor group, and this effect was reversed by XIAP knockdown (Figure 6c–e). These findings suggest that XIAP is negatively regulated by miR-186-5p in SH-SY5Y cells.
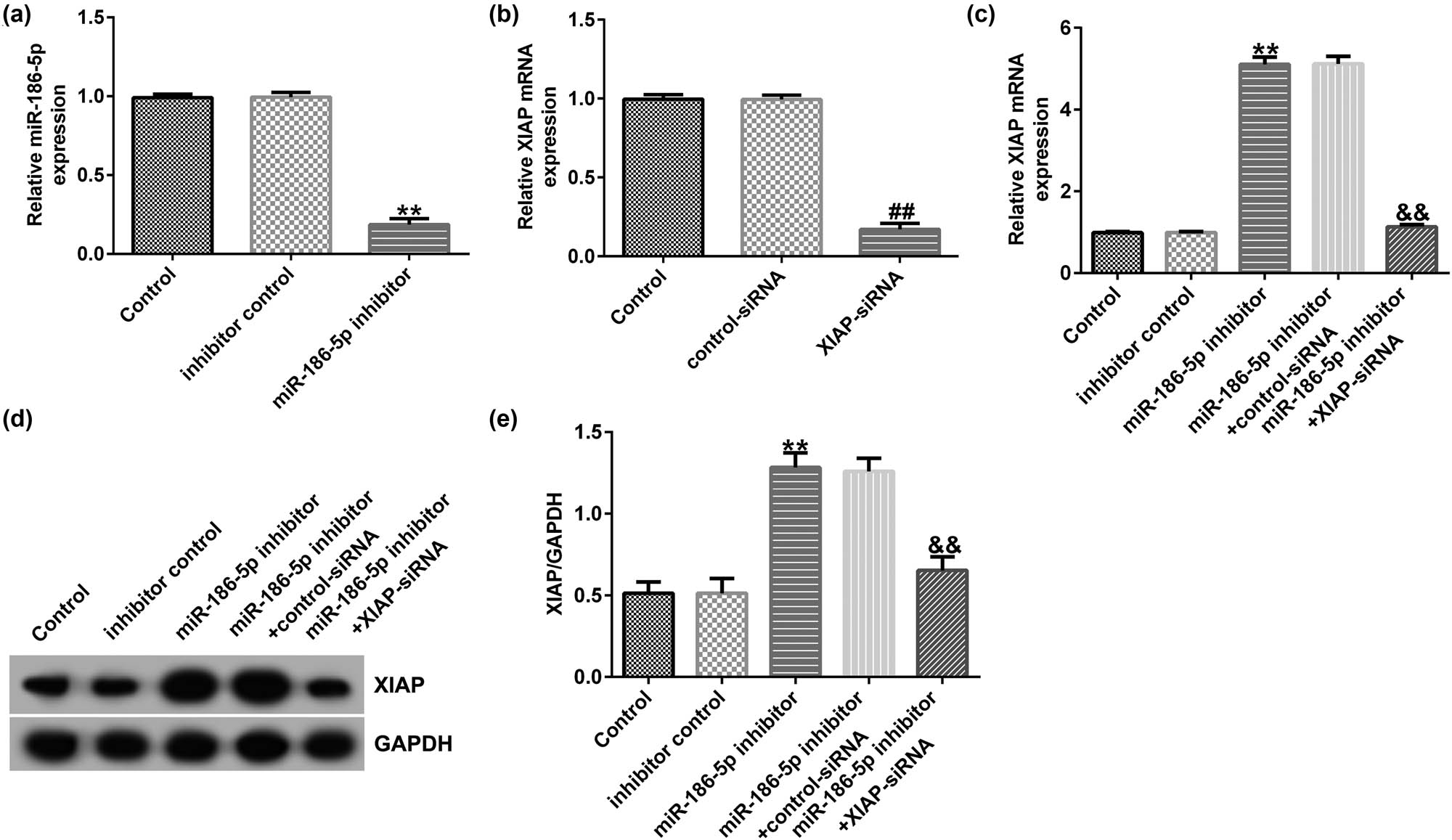
miR-186-5p negatively regulates XIAP expression levels in SH-SY5Y cells. (a) RT-qPCR assay results showing miR-186-5p expression levels in SH-SY5Y cells transfected with the inhibitor control or miR-186-5p inhibitors. (b) RT-qPCR assay results showing XIAP expression levels in SH-SY5Y cells transfected with the control or XIAP siRNAs. (c) RT-qPCR assay results showing XIAP expression levels in SH-SY5Y cells transfected with the miR-186-5p inhibitor + control siRNA or miR-186-5p inhibitor + XIAP siRNA. (d) Western blotting analysis results showing the protein expression levels of XIAP. (e) XIAP/GAPDH. ** P < 0.01 vs control inhibitor group; ## P < 0.01 vs control siRNA group; && P < 0.01 vs miR-186-5p inhibitor + control siRNA group. GraphPad Prism 6.0 software (GraphPad Software, Inc.) was used for creation of the figure.
3.7 Effect of miR-186-5p silencing on OGD-induced nerve cell injury
Following cell transfection with inhibitor control, miR-186-5p inhibitor, miR-186-5p inhibitor + control-siRNA, or miR-186-5p inhibitor + XIAP-siRNA for 24 h, an OGD-induced cell injury model was established in SH-SY5Y cells. The results demonstrated that miR-186-5p levels were upregulated (Figure 7a) and XIAP levels were downregulated (Figure 7b) in the OGD group compared to those in the control group. In addition, compared with the OGD + inhibitor control group, the expression of miR-186-5p and XIAP decreased and increased, respectively, in the OGD + miR-186-5p inhibitor group. Finally, compared with the OGD + inhibitor control group, cell viability was improved (Figure 7c) and cell apoptosis (Figure 7d and e) and caspase 3 activity (Figure 7f) were reduced in the OGD + miR-186-5p inhibitor group, and all these effects were reversed by the knockdown of XIAP. In general, miR-186-5p silencing relieves OGD-induced nerve cell injury via up-regulating XIAP expression.

miR-186-5p inhibitor attenuates OGD-induced neuronal injury by upregulating XIAP expression. (a) RT-qPCR analysis results showing miR-186-5p expression levels. (b) RT-qPCR analysis results showing XIAP expression levels. (c) MTT assay was used to assess the cell viability. (d) Flow cytometric assay was carried out to evaluate cell apoptosis. (e) The cell apoptosis ratio is presented. (f) Caspase 3 activity was detected using the corresponding kit. ** P < 0.01 vs control group; ## P < 0.01 vs OGD + inhibitor control group; && P < 0.01 vs OGD + miR-186-5p inhibitor + control siRNA group. GraphPad Prism 6.0 software (GraphPad Software, Inc.) was used for creation of the figure.
4 Discussion
Acute cerebral ischemic stroke is characterized by an increased mortality rate in humans and is commonly accompanied by several complications, such as behavioral, social, attention, cognitive, and functional motor dysfunctions [20,21]. The pathological process of ischemic stroke is complicated and the factors contributing to the disease are diverse. Neuroinflammation and oxidative stress have been reported to play significant roles in ischemic brain injury [22,23]. Although several treatment strategies have been previously applied to treat ischemic stroke, the therapeutic potential of currently available treatment approaches remains unsatisfactory [24,25,26]. Therefore, novel treatment methods for ischemic stroke are urgently required.
The current study aimed to investigate the role of the lncRNA WT1-AS in cerebral ischemic stroke. The roles of lncRNAs in several diseases, including cerebral ischemic stroke, have attracted increasing attention. Shan et al. [27] showed that the lncRNA taurine-upregulated gene 1 (TUG1) is involved in the development of cerebral ischemic stroke, and TUG1 silencing can diminish OGD/R-induced injury. Another study by Wen et al. [28] indicated that lncRNA Gm4419 overexpression upregulates TNF-α, IL-1β, and IL-6 levels, thereby enhancing OGD/R damage. Hu et al. [29] showed that knockdown of lncRNA small nucleolar RNA host gene 15 could improve ischemia/hypoxia-induced neuronal injury and microglial inflammation by targeting the miR-302a-3p/signal transducer and activator of transcription 1 axis. Furthermore, Xiao et al. [30] demonstrated that lncRNA H19 was significantly upregulated in patients with ischemic stroke, whereas its silencing could attenuate neuronal apoptosis in OGD-induced neuronal cells by targeting miR-19a. In this study, bioinformatics analysis and dual-luciferase reporter assays were performed to verify the interaction between lncRNA WT1-AS and miR-186-5p.
lncRNA WT1-AS is involved in cancer development [31,32,33]. To the best of our knowledge, the effects of lncRNA WT1-AS on ischemic stroke have not yet been reported. Consistent with previous studies, the results of the present study demonstrated that WT1-AS levels were downregulated and miR-186-5p levels were upregulated in the blood samples of patients with ischemic stroke. Similarly, the expression levels of WT1-AS and miR-186-5p were enhanced and reduced, respectively, in OGD-treated SH-SY5Y cells in vitro.
Subsequently, the mechanisms underlying the effects of WT1-AS and miR-186-5p on OGD-induced SH-SY5Y cell injury were investigated. Du et al. [34] showed that LINC01705 was directly targeted by miR-186-5p and was involved in breast cancer development. In addition, Zhu et al. [35] demonstrated that HLA complex P5 could promote neuroblastoma cell proliferation by downregulating miR-186-5p expression. In osteosarcoma, miR-186-5p attenuates cell proliferation, invasion, and metastasis by targeting Forkhead Box K1 [36]. In this study, WT1-AS overexpression attenuated OGD-induced neuronal injury by downregulating the expression of miR-186-5p. Furthermore, the results of the current study revealed that XIAP was a direct target of miR-186-5p.
XIAP, a member of the inhibitor of apoptosis protein family, is a cytoplasmic inhibitor of caspases 3, 7, and 9 [37]. As a target for cancer treatment, XIAP is abnormally expressed in cancer, thus playing a significant role in regulating patient mortality [38]. Huang et al. [39] revealed that miR-377-3p can suppress colorectal cancer by regulating XIAP expression. Deng et al. [40] demonstrated that XIAP is involved in the development of ischemic stroke, and its expression is reduced in middle cerebral artery occlusion (MCAO) model rats. Therefore, XIAP silencing can reverse the effects of miR-130a downregulation on neurological function and angiogenesis in MCAO model rats. In this study, miR-186-5p silencing alleviated the OGD-induced SH-SY5Y cell injury. Besides, activation of caspase3 was found to be mechanism of XIAP silencing-induced apoptosis. Since caspase3 is activated by proteolytic cleavage, total caspase3 and cleaved caspase3 levels could be determined. This study did not analyze the total caspase3 and cleaved caspase3 level, which was a limitation of this study. Overall, the results of the present study indicate that WT1-AS plays a crucial role in cerebral ischemic stroke by regulating the miR-186-5p/XIAP axis. lncRNA WT1-AS attenuates hypoxia/ischemia-induced neuronal injury in cerebral ischemic stroke via the miR-186-5p/XIAP axis.
However, to further elucidate the role of lncRNA WT1-AS in ischemic stroke, future in-depth studies are needed. For example, more cell lines may be required to confirm that the effect of IncRNA WT1-AS on neuronal damage is general activity and not SH-SY5Y specific. The effects of XIAP over-expression on OGD induced neuronal injury should be further clarified. Besides, the role of lncRNA WT1-AS in ischemic stroke should be explored using animal models. In addition, the correlation between lncRNA WT1-AS expression and clinicopathological parameters of ischemic stroke patients should be investigated. These issues will be addressed in the future.
5 Conclusion
In this study, we found that lncRNA WT1-AS reduced OGD-induced SH-SY5Y cell injury via the miR-186-5p/XIAP axis, indicating the protective role of lncRNA WT1-AS in hypoxia/ischemia-induced neuronal injury in cerebral ischemic stroke.
Abbreviations
- FITC
-
fluorescein isothiocyanate
- lncRNA
-
long non-coding RNA
- MTT
-
3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide
- OGD
-
oxygen glucose deprivation
- PI
-
propidium iodide
- RT-qPCR
-
reverse transcription-quantitative PCR
- WT1-AS
-
WT1 antisense RNA
- XIAP
-
X-linked inhibitor of apoptosis
Acknowledgments
Not applicable.
-
Funding information: This study was supported by the Taizhou Science and Technology Support Plan (Social Development, grant no. SSF20210117), the Scientific Research Foundation of Taizhou People’s Hospital (Grant No. ZL202015) and the Project of Taizhou City Personnel Bureau (Grant No. RCPY201937).
-
Conflict of interest: Authors state no conflict of interest.
-
Data availability statement: The datasets used and/or analyzed during the present study are available from the corresponding author on reasonable request.
References
[1] Meschia JF, Bushnell C, Boden-Albala B, Braun LT, Bravata DM, Chaturvedi S, et al. Guidelines for the primary prevention of stroke. Stroke. 2014;45:3754–832.10.1161/STR.0000000000000046Suche in Google Scholar PubMed PubMed Central
[2] Virani SS, Alonso A, Benjamin EJ, Bittencourt MS, Callaway CW, Carson AP, et al. Heart disease and stroke statistics-2020 update: a report from the american heart association. Circulation. 2020;141:e139–596.10.1161/CIR.0000000000000757Suche in Google Scholar PubMed
[3] Thrift AG, Dewey HM, Macdonell RA, McNeil JJ, Donnan GA. Incidence of the major stroke subtypes: initial findings from the North East Melbourne stroke incidence study (NEMESIS). Stroke. 2001;32:1732–8.10.1161/01.STR.32.8.1732Suche in Google Scholar PubMed
[4] Kuriakose D, Xiao Z. Pathophysiology and treatment of stroke: Present status and future perspectives. Int J Mol Sci. 2020;21:7609.10.3390/ijms21207609Suche in Google Scholar PubMed PubMed Central
[5] Rinn JL, Chang HY. Long noncoding RNAs: molecular modalities to organismal functions. Annu Rev Biochem. 2020;89:283–308.10.1146/annurev-biochem-062917-012708Suche in Google Scholar PubMed
[6] Lekka E, Hall J. Noncoding RNAs in disease. FEBS Lett. 2018;592:2884–900.10.1002/1873-3468.13182Suche in Google Scholar PubMed PubMed Central
[7] Kopp F, Mendell JT. Functional classification and experimental dissection of long noncoding RNAs. Cell. 2018;172:393–407.10.1016/j.cell.2018.01.011Suche in Google Scholar PubMed PubMed Central
[8] Carter G, Miladinovic B, Patel AA, Deland L, Mastorides S, Patel NA. Circulating long noncoding RNA GAS5 levels are correlated to prevalence of type 2 diabetes mellitus. BBA Clin. 2015;4:102–7.10.1016/j.bbacli.2015.09.001Suche in Google Scholar PubMed PubMed Central
[9] Zheng Y, Song D, Xiao K, Yang C, Ding Y, Deng W, et al. LncRNA GAS5 contributes to lymphatic metastasis in colorectal cancer. Oncotarget. 2016;7:83727–34.10.18632/oncotarget.13384Suche in Google Scholar PubMed PubMed Central
[10] Le F, Luo P, Ouyang Q, Zhong X. LncRNA WT1-AS downregulates survivin by upregulating miR-203 in papillary thyroid carcinoma. Cancer Manag Res. 2020 Jan 20;12:443–9.10.2147/CMAR.S232294Suche in Google Scholar PubMed PubMed Central
[11] Wang J, Xi C, Yang X, Lu X, Yu K, Zhang Y, et al. LncRNA WT1-AS inhibits triple-negative breast cancer cell migration and invasion by downregulating transforming growth factor beta1. Cancer Biother Radiopharm. 2019;34:671–5.10.1089/cbr.2019.2925Suche in Google Scholar PubMed
[12] Wu C, Yang J, Li R, Lin X, Wu J, Wu J. LncRNA WT1-AS/miR-494-3p regulates cell proliferation, apoptosis, migration and invasion via PTEN/PI3K/AKT signaling pathway in non-small cell lung cancer. Onco Targets Ther. 2021;14:891–904.10.2147/OTT.S278233Suche in Google Scholar PubMed PubMed Central
[13] Qiu G, Tong W, Jiang C, Xie Q, Zou J, Luo C, et al. Long noncoding RNA WT1-AS inhibit cell malignancy via miR-494-3p in glioma. Technol Cancer Res Treat. 2020;19:1533033820919759.10.1177/1533033820919759Suche in Google Scholar PubMed PubMed Central
[14] Luo H, Zhang J, He Z, Wu S. Long noncoding RNA WT1-AS inhibits the progression of cervical cancer by sponging miR-205. Cancer Biother Radiopharm. 2021;36:491–500.10.1089/cbr.2019.3279Suche in Google Scholar PubMed
[15] Saliminejad K, Khorram Khorshid HR, Soleymani Fard S, Ghaffari SH. An overview of microRNAs: Biology, functions, therapeutics, and analysis methods. J Cell Physiol. 2019;234:5451–65.10.1002/jcp.27486Suche in Google Scholar PubMed
[16] Zhu K, Su Y, Xu B, Wang Z, Sun H, Wang L, et al. MicroRNA-186-5p represses neuroblastoma cell growth via downregulation of Eg5. Am J Transl Res. 2019;11:2245–56.Suche in Google Scholar
[17] Jones DZ, Schmidt ML, Suman S, Hobbing KR, Barve SS, Gobejishvili L, et al. Micro-RNA-186-5p inhibition attenuates proliferation, anchorage independent growth and invasion in metastatic prostate cancer cells. BMC Cancer. 2018;18:421.10.1186/s12885-018-4258-0Suche in Google Scholar PubMed PubMed Central
[18] Tao Z, Cao Z, Wang X, Pan D, Jia Q. Long noncoding RNA SNHG14 regulates ox-LDL-induced atherosclerosis cell proliferation and apoptosis by targeting miR-186-5p/WIPF2 axis. Hum Exp Toxicol. 2021;40(1):47–59.10.1177/0960327120940363Suche in Google Scholar PubMed
[19] Zhou Y, Li T, Chen Z, Huang J, Qin Z, Li L. Overexpression of lncRNA TUG1 alleviates NLRP3 inflammasome-mediated cardiomyocyte pyroptosis through targeting the miR-186-5p/XIAP axis in coronary microembolization-induced myocardial damage. Front Immunol. 2021;12:637598.10.3389/fimmu.2021.637598Suche in Google Scholar PubMed PubMed Central
[20] Ren C, Li N, Gao C, Zhang W, Yang Y, Li S, et al. Ligustilide provides neuroprotection by promoting angiogenesis after cerebral ischemia. Neurol Res. 2020;42:683–92.10.1080/01616412.2020.1782122Suche in Google Scholar PubMed
[21] Li B, Concepcion K, Meng X, Zhang L. Brain-immune interactions in perinatal hypoxic-ischemic brain injury. Prog Neurobiol. 2017;159:50–68.10.1016/j.pneurobio.2017.10.006Suche in Google Scholar PubMed PubMed Central
[22] Tobin MK, Bonds JA, Minshall RD, Pelligrino DA, Testai FD, Lazarov O. Neurogenesis and inflammation after ischemic stroke: what is known and where we go from here. J Cereb Blood Flow Metab. 2014;34:1573–84.10.1038/jcbfm.2014.130Suche in Google Scholar PubMed PubMed Central
[23] Yu W, Gao D, Jin W, Liu S, Qi S. Propofol prevents oxidative stress by decreasing the ischemic accumulation of succinate in focalcerebral ischemia-reperfusion injury. Neurochem Res. 2018;43:420–9.10.1007/s11064-017-2437-zSuche in Google Scholar PubMed
[24] Rabinstein AA. Update on treatment of acute ischemic stroke. Contin (Minneap Minn). 2020;26:268–86.10.1212/CON.0000000000000840Suche in Google Scholar PubMed
[25] Warach SJ, Dula AN, Milling TJ Jr. Tenecteplase thrombolysis for acute ischemic stroke. Stroke. 2020;51:3440–51.10.1161/STROKEAHA.120.029749Suche in Google Scholar PubMed PubMed Central
[26] Paul S, Candelario-Jalil E. Emerging neuroprotective strategies for the treatment of ischemic stroke: An overview of clinical and preclinical studies. Exp Neurol. 2021;335:113518.10.1016/j.expneurol.2020.113518Suche in Google Scholar PubMed PubMed Central
[27] Shan W, Chen W, Zhao X, Pei A, Chen M, Yu Y, et al. Long noncoding RNA TUG1 contributes to cerebral ischaemia/reperfusion injury by sponging mir-145 to up-regulate AQP4 expression. J Cell Mol Med. 2020;24:250–9.10.1111/jcmm.14712Suche in Google Scholar PubMed PubMed Central
[28] Wen Y, Yu Y, Fu X. LncRNA Gm4419 contributes to OGD/R injury of cerebral microglial cells via IκB phosphorylation and NF-κB activation. Biochem Biophys Res Commun. 2017;487:923–9.10.1016/j.bbrc.2017.05.005Suche in Google Scholar PubMed
[29] Hu C, Li C, Ma Q, Wang R, He Y, Wang H, et al. Inhibition of long noncoding RNA SNHG15 ameliorates hypoxia/ischemia-induced neuronal damage by regulating miR-302a-3p/STAT1/NF-κB axis. Yonsei Med J. 2021;62:325–37.10.3349/ymj.2021.62.4.325Suche in Google Scholar PubMed PubMed Central
[30] Xiao Z, Qiu Y, Lin Y, Medina R, Zhuang S, Rosenblum JS, et al. Blocking lncRNA H19-miR-19a-Id2 axis attenuates hypoxia/ischemia induced neuronal injury. Aging (Albany NY). 2019;11(11):3600.10.18632/aging.101999Suche in Google Scholar PubMed PubMed Central
[31] Cui L, Nai M, Zhang K, Li L, Li R. LncRNA WT1-AS inhibits the aggressiveness of cervical cancer cell via regulating p53 expression via sponging miR-330-5p. Cancer Manag Res. 2019;11:651–67.10.2147/CMAR.S176525Suche in Google Scholar PubMed PubMed Central
[32] Li F, Luo P, Ouyang Q, Zhong XM. LncRNA WT1-AS downregulates survivin by upregulating miR-203 in papillary thyroid carcinoma. Cancer Manag Res. 2020;12:443–9.10.2147/CMAR.S232294Suche in Google Scholar PubMed PubMed Central
[33] Zhang Y, Na R, Wang X. LncRNA WT1-AS up-regulates p53 to inhibit the proliferation of cervical squamous carcinoma cells. BMC Cancer. 2019;19:1052.10.1186/s12885-019-6264-2Suche in Google Scholar PubMed PubMed Central
[34] Du C, Zhang J, Wang Y, Zhang Y, Zhang J, Zhang L, Li J. The long non-coding RNA LINC01705 regulates the development of breast cancer by sponging miR-186-5p to mediate TPR expression as a competitive endogenous RNA. Front Genet. 2020;11:799.10.3389/fgene.2020.00779Suche in Google Scholar PubMed PubMed Central
[35] Zhu K, Wang L, Zhang X, Sun H, Chen T, Sun C, et al. LncRNA HCP5 promotes neuroblastoma proliferation by regulating miR-186-5p/MAP3K2 signal axis. J Pediatr Surg. 2021;56:778–87.10.1016/j.jpedsurg.2020.10.011Suche in Google Scholar PubMed
[36] Zhang Z, Zhang W, Mao J, Xu Z, Fan M. miR-186-5p functions as a tumor suppressor in human osteosarcoma by targeting FOXK1. Cell Physiol Biochem. 2019;52:553–64.10.33594/000000039Suche in Google Scholar PubMed
[37] Jost PJ, Vucic D. Regulation of cell death and immunity by XIAP. Cold Spring Harb Perspect Biol. 2020;12:a036426.10.1101/cshperspect.a036426Suche in Google Scholar PubMed PubMed Central
[38] Tu H, Costa M. XIAP’s profile in human cancer. Biomolecules. 2020;10:1493.10.3390/biom10111493Suche in Google Scholar PubMed PubMed Central
[39] Huang L, Liu Z, Hu J, Luo Z, Zhang C, Wang L, et al. MiR-377-3p suppresses colorectal cancer through negative regulation on Wnt/beta-catenin signaling by targeting XIAP and ZEB2. Pharmacol Res. 2020;156:104774.10.1016/j.phrs.2020.104774Suche in Google Scholar PubMed
[40] Deng W, Fan C, Zhao Y, Mao Y, Li J, Zhang Y, et al. MicroRNA-130a regulates neurological deficit and angiogenesis in rats with ischaemic stroke by targeting XIAP. J Cell Mol Med. 2020;24:10987–1000.10.1111/jcmm.15732Suche in Google Scholar PubMed PubMed Central
© 2022 Jianquan You et al., published by De Gruyter
This work is licensed under the Creative Commons Attribution 4.0 International License.
Artikel in diesem Heft
- Research Articles
- AMBRA1 attenuates the proliferation of uveal melanoma cells
- A ceRNA network mediated by LINC00475 in papillary thyroid carcinoma
- Differences in complications between hepatitis B-related cirrhosis and alcohol-related cirrhosis
- Effect of gestational diabetes mellitus on lipid profile: A systematic review and meta-analysis
- Long noncoding RNA NR2F1-AS1 stimulates the tumorigenic behavior of non-small cell lung cancer cells by sponging miR-363-3p to increase SOX4
- Promising novel biomarkers and candidate small-molecule drugs for lung adenocarcinoma: Evidence from bioinformatics analysis of high-throughput data
- Plasmapheresis: Is it a potential alternative treatment for chronic urticaria?
- The biomarkers of key miRNAs and gene targets associated with extranodal NK/T-cell lymphoma
- Gene signature to predict prognostic survival of hepatocellular carcinoma
- Effects of miRNA-199a-5p on cell proliferation and apoptosis of uterine leiomyoma by targeting MED12
- Does diabetes affect paraneoplastic thrombocytosis in colorectal cancer?
- Is there any effect on imprinted genes H19, PEG3, and SNRPN during AOA?
- Leptin and PCSK9 concentrations are associated with vascular endothelial cytokines in patients with stable coronary heart disease
- Pericentric inversion of chromosome 6 and male fertility problems
- Staple line reinforcement with nebulized cyanoacrylate glue in laparoscopic sleeve gastrectomy: A propensity score-matched study
- Retrospective analysis of crescent score in clinical prognosis of IgA nephropathy
- Expression of DNM3 is associated with good outcome in colorectal cancer
- Activation of SphK2 contributes to adipocyte-induced EOC cell proliferation
- CRRT influences PICCO measurements in febrile critically ill patients
- SLCO4A1-AS1 mediates pancreatic cancer development via miR-4673/KIF21B axis
- lncRNA ACTA2-AS1 inhibits malignant phenotypes of gastric cancer cells
- circ_AKT3 knockdown suppresses cisplatin resistance in gastric cancer
- Prognostic value of nicotinamide N-methyltransferase in human cancers: Evidence from a meta-analysis and database validation
- GPC2 deficiency inhibits cell growth and metastasis in colon adenocarcinoma
- A pan-cancer analysis of the oncogenic role of Holliday junction recognition protein in human tumors
- Radiation increases COL1A1, COL3A1, and COL1A2 expression in breast cancer
- Association between preventable risk factors and metabolic syndrome
- miR-29c-5p knockdown reduces inflammation and blood–brain barrier disruption by upregulating LRP6
- Cardiac contractility modulation ameliorates myocardial metabolic remodeling in a rabbit model of chronic heart failure through activation of AMPK and PPAR-α pathway
- Quercitrin protects human bronchial epithelial cells from oxidative damage
- Smurf2 suppresses the metastasis of hepatocellular carcinoma via ubiquitin degradation of Smad2
- circRNA_0001679/miR-338-3p/DUSP16 axis aggravates acute lung injury
- Sonoclot’s usefulness in prediction of cardiopulmonary arrest prognosis: A proof of concept study
- Four drug metabolism-related subgroups of pancreatic adenocarcinoma in prognosis, immune infiltration, and gene mutation
- Decreased expression of miR-195 mediated by hypermethylation promotes osteosarcoma
- LMO3 promotes proliferation and metastasis of papillary thyroid carcinoma cells by regulating LIMK1-mediated cofilin and the β-catenin pathway
- Cx43 upregulation in HUVECs under stretch via TGF-β1 and cytoskeletal network
- Evaluation of menstrual irregularities after COVID-19 vaccination: Results of the MECOVAC survey
- Histopathologic findings on removed stomach after sleeve gastrectomy. Do they influence the outcome?
- Analysis of the expression and prognostic value of MT1-MMP, β1-integrin and YAP1 in glioma
- Optimal diagnosis of the skin cancer using a hybrid deep neural network and grasshopper optimization algorithm
- miR-223-3p alleviates TGF-β-induced epithelial-mesenchymal transition and extracellular matrix deposition by targeting SP3 in endometrial epithelial cells
- Clinical value of SIRT1 as a prognostic biomarker in esophageal squamous cell carcinoma, a systematic meta-analysis
- circ_0020123 promotes cell proliferation and migration in lung adenocarcinoma via PDZD8
- miR-22-5p regulates the self-renewal of spermatogonial stem cells by targeting EZH2
- hsa-miR-340-5p inhibits epithelial–mesenchymal transition in endometriosis by targeting MAP3K2 and inactivating MAPK/ERK signaling
- circ_0085296 inhibits the biological functions of trophoblast cells to promote the progression of preeclampsia via the miR-942-5p/THBS2 network
- TCD hemodynamics findings in the subacute phase of anterior circulation stroke patients treated with mechanical thrombectomy
- Development of a risk-stratification scoring system for predicting risk of breast cancer based on non-alcoholic fatty liver disease, non-alcoholic fatty pancreas disease, and uric acid
- Tollip promotes hepatocellular carcinoma progression via PI3K/AKT pathway
- circ_0062491 alleviates periodontitis via the miR-142-5p/IGF1 axis
- Human amniotic fluid as a source of stem cells
- lncRNA NONRATT013819.2 promotes transforming growth factor-β1-induced myofibroblastic transition of hepatic stellate cells by miR24-3p/lox
- NORAD modulates miR-30c-5p-LDHA to protect lung endothelial cells damage
- Idiopathic pulmonary fibrosis telemedicine management during COVID-19 outbreak
- Risk factors for adverse drug reactions associated with clopidogrel therapy
- Serum zinc associated with immunity and inflammatory markers in Covid-19
- The relationship between night shift work and breast cancer incidence: A systematic review and meta-analysis of observational studies
- LncRNA expression in idiopathic achalasia: New insight and preliminary exploration into pathogenesis
- Notoginsenoside R1 alleviates spinal cord injury through the miR-301a/KLF7 axis to activate Wnt/β-catenin pathway
- Moscatilin suppresses the inflammation from macrophages and T cells
- Zoledronate promotes ECM degradation and apoptosis via Wnt/β-catenin
- Epithelial-mesenchymal transition-related genes in coronary artery disease
- The effect evaluation of traditional vaginal surgery and transvaginal mesh surgery for severe pelvic organ prolapse: 5 years follow-up
- Repeated partial splenic artery embolization for hypersplenism improves platelet count
- Low expression of miR-27b in serum exosomes of non-small cell lung cancer facilitates its progression by affecting EGFR
- Exosomal hsa_circ_0000519 modulates the NSCLC cell growth and metastasis via miR-1258/RHOV axis
- miR-455-5p enhances 5-fluorouracil sensitivity in colorectal cancer cells by targeting PIK3R1 and DEPDC1
- The effect of tranexamic acid on the reduction of intraoperative and postoperative blood loss and thromboembolic risk in patients with hip fracture
- Isocitrate dehydrogenase 1 mutation in cholangiocarcinoma impairs tumor progression by sensitizing cells to ferroptosis
- Artemisinin protects against cerebral ischemia and reperfusion injury via inhibiting the NF-κB pathway
- A 16-gene signature associated with homologous recombination deficiency for prognosis prediction in patients with triple-negative breast cancer
- Lidocaine ameliorates chronic constriction injury-induced neuropathic pain through regulating M1/M2 microglia polarization
- MicroRNA 322-5p reduced neuronal inflammation via the TLR4/TRAF6/NF-κB axis in a rat epilepsy model
- miR-1273h-5p suppresses CXCL12 expression and inhibits gastric cancer cell invasion and metastasis
- Clinical characteristics of pneumonia patients of long course of illness infected with SARS-CoV-2
- circRNF20 aggravates the malignancy of retinoblastoma depending on the regulation of miR-132-3p/PAX6 axis
- Linezolid for resistant Gram-positive bacterial infections in children under 12 years: A meta-analysis
- Rack1 regulates pro-inflammatory cytokines by NF-κB in diabetic nephropathy
- Comprehensive analysis of molecular mechanism and a novel prognostic signature based on small nuclear RNA biomarkers in gastric cancer patients
- Smog and risk of maternal and fetal birth outcomes: A retrospective study in Baoding, China
- Let-7i-3p inhibits the cell cycle, proliferation, invasion, and migration of colorectal cancer cells via downregulating CCND1
- β2-Adrenergic receptor expression in subchondral bone of patients with varus knee osteoarthritis
- Possible impact of COVID-19 pandemic and lockdown on suicide behavior among patients in Southeast Serbia
- In vitro antimicrobial activity of ozonated oil in liposome eyedrop against multidrug-resistant bacteria
- Potential biomarkers for inflammatory response in acute lung injury
- A low serum uric acid concentration predicts a poor prognosis in adult patients with candidemia
- Antitumor activity of recombinant oncolytic vaccinia virus with human IL2
- ALKBH5 inhibits TNF-α-induced apoptosis of HUVECs through Bcl-2 pathway
- Risk prediction of cardiovascular disease using machine learning classifiers
- Value of ultrasonography parameters in diagnosing polycystic ovary syndrome
- Bioinformatics analysis reveals three key genes and four survival genes associated with youth-onset NSCLC
- Identification of autophagy-related biomarkers in patients with pulmonary arterial hypertension based on bioinformatics analysis
- Protective effects of glaucocalyxin A on the airway of asthmatic mice
- Overexpression of miR-100-5p inhibits papillary thyroid cancer progression via targeting FZD8
- Bioinformatics-based analysis of SUMOylation-related genes in hepatocellular carcinoma reveals a role of upregulated SAE1 in promoting cell proliferation
- Effectiveness and clinical benefits of new anti-diabetic drugs: A real life experience
- Identification of osteoporosis based on gene biomarkers using support vector machine
- Tanshinone IIA reverses oxaliplatin resistance in colorectal cancer through microRNA-30b-5p/AVEN axis
- miR-212-5p inhibits nasopharyngeal carcinoma progression by targeting METTL3
- Association of ST-T changes with all-cause mortality among patients with peripheral T-cell lymphomas
- LINC00665/miRNAs axis-mediated collagen type XI alpha 1 correlates with immune infiltration and malignant phenotypes in lung adenocarcinoma
- The perinatal factors that influence the excretion of fecal calprotectin in premature-born children
- Effect of femoral head necrosis cystic area on femoral head collapse and stress distribution in femoral head: A clinical and finite element study
- Does the use of 3D-printed cones give a chance to postpone the use of megaprostheses in patients with large bone defects in the knee joint?
- lncRNA HAGLR modulates myocardial ischemia–reperfusion injury in mice through regulating miR-133a-3p/MAPK1 axis
- Protective effect of ghrelin on intestinal I/R injury in rats
- In vivo knee kinematics of an innovative prosthesis design
- Relationship between the height of fibular head and the incidence and severity of knee osteoarthritis
- lncRNA WT1-AS attenuates hypoxia/ischemia-induced neuronal injury during cerebral ischemic stroke via miR-186-5p/XIAP axis
- Correlation of cardiac troponin T and APACHE III score with all-cause in-hospital mortality in critically ill patients with acute pulmonary embolism
- LncRNA LINC01857 reduces metastasis and angiogenesis in breast cancer cells via regulating miR-2052/CENPQ axis
- Endothelial cell-specific molecule 1 (ESM1) promoted by transcription factor SPI1 acts as an oncogene to modulate the malignant phenotype of endometrial cancer
- SELENBP1 inhibits progression of colorectal cancer by suppressing epithelial–mesenchymal transition
- Visfatin is negatively associated with coronary artery lesions in subjects with impaired fasting glucose
- Treatment and outcomes of mechanical complications of acute myocardial infarction during the Covid-19 era: A comparison with the pre-Covid-19 period. A systematic review and meta-analysis
- Neonatal stroke surveillance study protocol in the United Kingdom and Republic of Ireland
- Oncogenic role of TWF2 in human tumors: A pan-cancer analysis
- Mean corpuscular hemoglobin predicts the length of hospital stay independent of severity classification in patients with acute pancreatitis
- Association of gallstone and polymorphisms of UGT1A1*27 and UGT1A1*28 in patients with hepatitis B virus-related liver failure
- TGF-β1 upregulates Sar1a expression and induces procollagen-I secretion in hypertrophic scarring fibroblasts
- Antisense lncRNA PCNA-AS1 promotes esophageal squamous cell carcinoma progression through the miR-2467-3p/PCNA axis
- NK-cell dysfunction of acute myeloid leukemia in relation to the renin–angiotensin system and neurotransmitter genes
- The effect of dilution with glucose and prolonged injection time on dexamethasone-induced perineal irritation – A randomized controlled trial
- miR-146-5p restrains calcification of vascular smooth muscle cells by suppressing TRAF6
- Role of lncRNA MIAT/miR-361-3p/CCAR2 in prostate cancer cells
- lncRNA NORAD promotes lung cancer progression by competitively binding to miR-28-3p with E2F2
- Noninvasive diagnosis of AIH/PBC overlap syndrome based on prediction models
- lncRNA FAM230B is highly expressed in colorectal cancer and suppresses the maturation of miR-1182 to increase cell proliferation
- circ-LIMK1 regulates cisplatin resistance in lung adenocarcinoma by targeting miR-512-5p/HMGA1 axis
- LncRNA SNHG3 promoted cell proliferation, migration, and metastasis of esophageal squamous cell carcinoma via regulating miR-151a-3p/PFN2 axis
- Risk perception and affective state on work exhaustion in obstetrics during the COVID-19 pandemic
- lncRNA-AC130710/miR-129-5p/mGluR1 axis promote migration and invasion by activating PKCα-MAPK signal pathway in melanoma
- SNRPB promotes cell cycle progression in thyroid carcinoma via inhibiting p53
- Xylooligosaccharides and aerobic training regulate metabolism and behavior in rats with streptozotocin-induced type 1 diabetes
- Serpin family A member 1 is an oncogene in glioma and its translation is enhanced by NAD(P)H quinone dehydrogenase 1 through RNA-binding activity
- Silencing of CPSF7 inhibits the proliferation, migration, and invasion of lung adenocarcinoma cells by blocking the AKT/mTOR signaling pathway
- Ultrasound-guided lumbar plexus block versus transversus abdominis plane block for analgesia in children with hip dislocation: A double-blind, randomized trial
- Relationship of plasma MBP and 8-oxo-dG with brain damage in preterm
- Identification of a novel necroptosis-associated miRNA signature for predicting the prognosis in head and neck squamous cell carcinoma
- Delayed femoral vein ligation reduces operative time and blood loss during hip disarticulation in patients with extremity tumors
- The expression of ASAP3 and NOTCH3 and the clinicopathological characteristics of adult glioma patients
- Longitudinal analysis of factors related to Helicobacter pylori infection in Chinese adults
- HOXA10 enhances cell proliferation and suppresses apoptosis in esophageal cancer via activating p38/ERK signaling pathway
- Meta-analysis of early-life antibiotic use and allergic rhinitis
- Marital status and its correlation with age, race, and gender in prognosis of tonsil squamous cell carcinomas
- HPV16 E6E7 up-regulates KIF2A expression by activating JNK/c-Jun signal, is beneficial to migration and invasion of cervical cancer cells
- Amino acid profiles in the tissue and serum of patients with liver cancer
- Pain in critically ill COVID-19 patients: An Italian retrospective study
- Immunohistochemical distribution of Bcl-2 and p53 apoptotic markers in acetamiprid-induced nephrotoxicity
- Estradiol pretreatment in GnRH antagonist protocol for IVF/ICSI treatment
- Long non-coding RNAs LINC00689 inhibits the apoptosis of human nucleus pulposus cells via miR-3127-5p/ATG7 axis-mediated autophagy
- The relationship between oxygen therapy, drug therapy, and COVID-19 mortality
- Monitoring hypertensive disorders in pregnancy to prevent preeclampsia in pregnant women of advanced maternal age: Trial mimicking with retrospective data
- SETD1A promotes the proliferation and glycolysis of nasopharyngeal carcinoma cells by activating the PI3K/Akt pathway
- The role of Shunaoxin pills in the treatment of chronic cerebral hypoperfusion and its main pharmacodynamic components
- TET3 governs malignant behaviors and unfavorable prognosis of esophageal squamous cell carcinoma by activating the PI3K/AKT/GSK3β/β-catenin pathway
- Associations between morphokinetic parameters of temporary-arrest embryos and the clinical prognosis in FET cycles
- Long noncoding RNA WT1-AS regulates trophoblast proliferation, migration, and invasion via the microRNA-186-5p/CADM2 axis
- The incidence of bronchiectasis in chronic obstructive pulmonary disease
- Integrated bioinformatics analysis shows integrin alpha 3 is a prognostic biomarker for pancreatic cancer
- Inhibition of miR-21 improves pulmonary vascular responses in bronchopulmonary dysplasia by targeting the DDAH1/ADMA/NO pathway
- Comparison of hospitalized patients with severe pneumonia caused by COVID-19 and influenza A (H7N9 and H1N1): A retrospective study from a designated hospital
- lncRNA ZFAS1 promotes intervertebral disc degeneration by upregulating AAK1
- Pathological characteristics of liver injury induced by N,N-dimethylformamide: From humans to animal models
- lncRNA ELFN1-AS1 enhances the progression of colon cancer by targeting miR-4270 to upregulate AURKB
- DARS-AS1 modulates cell proliferation and migration of gastric cancer cells by regulating miR-330-3p/NAT10 axis
- Dezocine inhibits cell proliferation, migration, and invasion by targeting CRABP2 in ovarian cancer
- MGST1 alleviates the oxidative stress of trophoblast cells induced by hypoxia/reoxygenation and promotes cell proliferation, migration, and invasion by activating the PI3K/AKT/mTOR pathway
- Bifidobacterium lactis Probio-M8 ameliorated the symptoms of type 2 diabetes mellitus mice by changing ileum FXR-CYP7A1
- circRNA DENND1B inhibits tumorigenicity of clear cell renal cell carcinoma via miR-122-5p/TIMP2 axis
- EphA3 targeted by miR-3666 contributes to melanoma malignancy via activating ERK1/2 and p38 MAPK pathways
- Pacemakers and methylprednisolone pulse therapy in immune-related myocarditis concomitant with complete heart block
- miRNA-130a-3p targets sphingosine-1-phosphate receptor 1 to activate the microglial and astrocytes and to promote neural injury under the high glucose condition
- Review Articles
- Current management of cancer pain in Italy: Expert opinion paper
- Hearing loss and brain disorders: A review of multiple pathologies
- The rationale for using low-molecular weight heparin in the therapy of symptomatic COVID-19 patients
- Amyotrophic lateral sclerosis and delayed onset muscle soreness in light of the impaired blink and stretch reflexes – watch out for Piezo2
- Interleukin-35 in autoimmune dermatoses: Current concepts
- Recent discoveries in microbiota dysbiosis, cholangiocytic factors, and models for studying the pathogenesis of primary sclerosing cholangitis
- Advantages of ketamine in pediatric anesthesia
- Congenital adrenal hyperplasia. Role of dentist in early diagnosis
- Migraine management: Non-pharmacological points for patients and health care professionals
- Atherogenic index of plasma and coronary artery disease: A systematic review
- Physiological and modulatory role of thioredoxins in the cellular function
- Case Reports
- Intrauterine Bakri balloon tamponade plus cervical cerclage for the prevention and treatment of postpartum haemorrhage in late pregnancy complicated with acute aortic dissection: Case series
- A case of successful pembrolizumab monotherapy in a patient with advanced lung adenocarcinoma: Use of multiple biomarkers in combination for clinical practice
- Unusual neurological manifestations of bilateral medial medullary infarction: A case report
- Atypical symptoms of malignant hyperthermia: A rare causative mutation in the RYR1 gene
- A case report of dermatomyositis with the missed diagnosis of non-small cell lung cancer and concurrence of pulmonary tuberculosis
- A rare case of endometrial polyp complicated with uterine inversion: A case report and clinical management
- Spontaneous rupturing of splenic artery aneurysm: Another reason for fatal syncope and shock (Case report and literature review)
- Fungal infection mimicking COVID-19 infection – A case report
- Concurrent aspergillosis and cystic pulmonary metastases in a patient with tongue squamous cell carcinoma
- Paraganglioma-induced inverted takotsubo-like cardiomyopathy leading to cardiogenic shock successfully treated with extracorporeal membrane oxygenation
- Lineage switch from lymphoma to myeloid neoplasms: First case series from a single institution
- Trismus during tracheal extubation as a complication of general anaesthesia – A case report
- Simultaneous treatment of a pubovesical fistula and lymph node metastasis secondary to multimodal treatment for prostate cancer: Case report and review of the literature
- Two case reports of skin vasculitis following the COVID-19 immunization
- Ureteroiliac fistula after oncological surgery: Case report and review of the literature
- Synchronous triple primary malignant tumours in the bladder, prostate, and lung harbouring TP53 and MEK1 mutations accompanied with severe cardiovascular diseases: A case report
- Huge mucinous cystic neoplasms with adhesion to the left colon: A case report and literature review
- Commentary
- Commentary on “Clinicopathological features of programmed cell death-ligand 1 expression in patients with oral squamous cell carcinoma”
- Rapid Communication
- COVID-19 fear, post-traumatic stress, growth, and the role of resilience
- Erratum
- Erratum to “Tollip promotes hepatocellular carcinoma progression via PI3K/AKT pathway”
- Erratum to “Effect of femoral head necrosis cystic area on femoral head collapse and stress distribution in femoral head: A clinical and finite element study”
- Erratum to “lncRNA NORAD promotes lung cancer progression by competitively binding to miR-28-3p with E2F2”
- Retraction
- Expression and role of ABIN1 in sepsis: In vitro and in vivo studies
- Retraction to “miR-519d downregulates LEP expression to inhibit preeclampsia development”
- Special Issue Computational Intelligence Methodologies Meets Recurrent Cancers - Part II
- Usefulness of close surveillance for rectal cancer patients after neoadjuvant chemoradiotherapy
Artikel in diesem Heft
- Research Articles
- AMBRA1 attenuates the proliferation of uveal melanoma cells
- A ceRNA network mediated by LINC00475 in papillary thyroid carcinoma
- Differences in complications between hepatitis B-related cirrhosis and alcohol-related cirrhosis
- Effect of gestational diabetes mellitus on lipid profile: A systematic review and meta-analysis
- Long noncoding RNA NR2F1-AS1 stimulates the tumorigenic behavior of non-small cell lung cancer cells by sponging miR-363-3p to increase SOX4
- Promising novel biomarkers and candidate small-molecule drugs for lung adenocarcinoma: Evidence from bioinformatics analysis of high-throughput data
- Plasmapheresis: Is it a potential alternative treatment for chronic urticaria?
- The biomarkers of key miRNAs and gene targets associated with extranodal NK/T-cell lymphoma
- Gene signature to predict prognostic survival of hepatocellular carcinoma
- Effects of miRNA-199a-5p on cell proliferation and apoptosis of uterine leiomyoma by targeting MED12
- Does diabetes affect paraneoplastic thrombocytosis in colorectal cancer?
- Is there any effect on imprinted genes H19, PEG3, and SNRPN during AOA?
- Leptin and PCSK9 concentrations are associated with vascular endothelial cytokines in patients with stable coronary heart disease
- Pericentric inversion of chromosome 6 and male fertility problems
- Staple line reinforcement with nebulized cyanoacrylate glue in laparoscopic sleeve gastrectomy: A propensity score-matched study
- Retrospective analysis of crescent score in clinical prognosis of IgA nephropathy
- Expression of DNM3 is associated with good outcome in colorectal cancer
- Activation of SphK2 contributes to adipocyte-induced EOC cell proliferation
- CRRT influences PICCO measurements in febrile critically ill patients
- SLCO4A1-AS1 mediates pancreatic cancer development via miR-4673/KIF21B axis
- lncRNA ACTA2-AS1 inhibits malignant phenotypes of gastric cancer cells
- circ_AKT3 knockdown suppresses cisplatin resistance in gastric cancer
- Prognostic value of nicotinamide N-methyltransferase in human cancers: Evidence from a meta-analysis and database validation
- GPC2 deficiency inhibits cell growth and metastasis in colon adenocarcinoma
- A pan-cancer analysis of the oncogenic role of Holliday junction recognition protein in human tumors
- Radiation increases COL1A1, COL3A1, and COL1A2 expression in breast cancer
- Association between preventable risk factors and metabolic syndrome
- miR-29c-5p knockdown reduces inflammation and blood–brain barrier disruption by upregulating LRP6
- Cardiac contractility modulation ameliorates myocardial metabolic remodeling in a rabbit model of chronic heart failure through activation of AMPK and PPAR-α pathway
- Quercitrin protects human bronchial epithelial cells from oxidative damage
- Smurf2 suppresses the metastasis of hepatocellular carcinoma via ubiquitin degradation of Smad2
- circRNA_0001679/miR-338-3p/DUSP16 axis aggravates acute lung injury
- Sonoclot’s usefulness in prediction of cardiopulmonary arrest prognosis: A proof of concept study
- Four drug metabolism-related subgroups of pancreatic adenocarcinoma in prognosis, immune infiltration, and gene mutation
- Decreased expression of miR-195 mediated by hypermethylation promotes osteosarcoma
- LMO3 promotes proliferation and metastasis of papillary thyroid carcinoma cells by regulating LIMK1-mediated cofilin and the β-catenin pathway
- Cx43 upregulation in HUVECs under stretch via TGF-β1 and cytoskeletal network
- Evaluation of menstrual irregularities after COVID-19 vaccination: Results of the MECOVAC survey
- Histopathologic findings on removed stomach after sleeve gastrectomy. Do they influence the outcome?
- Analysis of the expression and prognostic value of MT1-MMP, β1-integrin and YAP1 in glioma
- Optimal diagnosis of the skin cancer using a hybrid deep neural network and grasshopper optimization algorithm
- miR-223-3p alleviates TGF-β-induced epithelial-mesenchymal transition and extracellular matrix deposition by targeting SP3 in endometrial epithelial cells
- Clinical value of SIRT1 as a prognostic biomarker in esophageal squamous cell carcinoma, a systematic meta-analysis
- circ_0020123 promotes cell proliferation and migration in lung adenocarcinoma via PDZD8
- miR-22-5p regulates the self-renewal of spermatogonial stem cells by targeting EZH2
- hsa-miR-340-5p inhibits epithelial–mesenchymal transition in endometriosis by targeting MAP3K2 and inactivating MAPK/ERK signaling
- circ_0085296 inhibits the biological functions of trophoblast cells to promote the progression of preeclampsia via the miR-942-5p/THBS2 network
- TCD hemodynamics findings in the subacute phase of anterior circulation stroke patients treated with mechanical thrombectomy
- Development of a risk-stratification scoring system for predicting risk of breast cancer based on non-alcoholic fatty liver disease, non-alcoholic fatty pancreas disease, and uric acid
- Tollip promotes hepatocellular carcinoma progression via PI3K/AKT pathway
- circ_0062491 alleviates periodontitis via the miR-142-5p/IGF1 axis
- Human amniotic fluid as a source of stem cells
- lncRNA NONRATT013819.2 promotes transforming growth factor-β1-induced myofibroblastic transition of hepatic stellate cells by miR24-3p/lox
- NORAD modulates miR-30c-5p-LDHA to protect lung endothelial cells damage
- Idiopathic pulmonary fibrosis telemedicine management during COVID-19 outbreak
- Risk factors for adverse drug reactions associated with clopidogrel therapy
- Serum zinc associated with immunity and inflammatory markers in Covid-19
- The relationship between night shift work and breast cancer incidence: A systematic review and meta-analysis of observational studies
- LncRNA expression in idiopathic achalasia: New insight and preliminary exploration into pathogenesis
- Notoginsenoside R1 alleviates spinal cord injury through the miR-301a/KLF7 axis to activate Wnt/β-catenin pathway
- Moscatilin suppresses the inflammation from macrophages and T cells
- Zoledronate promotes ECM degradation and apoptosis via Wnt/β-catenin
- Epithelial-mesenchymal transition-related genes in coronary artery disease
- The effect evaluation of traditional vaginal surgery and transvaginal mesh surgery for severe pelvic organ prolapse: 5 years follow-up
- Repeated partial splenic artery embolization for hypersplenism improves platelet count
- Low expression of miR-27b in serum exosomes of non-small cell lung cancer facilitates its progression by affecting EGFR
- Exosomal hsa_circ_0000519 modulates the NSCLC cell growth and metastasis via miR-1258/RHOV axis
- miR-455-5p enhances 5-fluorouracil sensitivity in colorectal cancer cells by targeting PIK3R1 and DEPDC1
- The effect of tranexamic acid on the reduction of intraoperative and postoperative blood loss and thromboembolic risk in patients with hip fracture
- Isocitrate dehydrogenase 1 mutation in cholangiocarcinoma impairs tumor progression by sensitizing cells to ferroptosis
- Artemisinin protects against cerebral ischemia and reperfusion injury via inhibiting the NF-κB pathway
- A 16-gene signature associated with homologous recombination deficiency for prognosis prediction in patients with triple-negative breast cancer
- Lidocaine ameliorates chronic constriction injury-induced neuropathic pain through regulating M1/M2 microglia polarization
- MicroRNA 322-5p reduced neuronal inflammation via the TLR4/TRAF6/NF-κB axis in a rat epilepsy model
- miR-1273h-5p suppresses CXCL12 expression and inhibits gastric cancer cell invasion and metastasis
- Clinical characteristics of pneumonia patients of long course of illness infected with SARS-CoV-2
- circRNF20 aggravates the malignancy of retinoblastoma depending on the regulation of miR-132-3p/PAX6 axis
- Linezolid for resistant Gram-positive bacterial infections in children under 12 years: A meta-analysis
- Rack1 regulates pro-inflammatory cytokines by NF-κB in diabetic nephropathy
- Comprehensive analysis of molecular mechanism and a novel prognostic signature based on small nuclear RNA biomarkers in gastric cancer patients
- Smog and risk of maternal and fetal birth outcomes: A retrospective study in Baoding, China
- Let-7i-3p inhibits the cell cycle, proliferation, invasion, and migration of colorectal cancer cells via downregulating CCND1
- β2-Adrenergic receptor expression in subchondral bone of patients with varus knee osteoarthritis
- Possible impact of COVID-19 pandemic and lockdown on suicide behavior among patients in Southeast Serbia
- In vitro antimicrobial activity of ozonated oil in liposome eyedrop against multidrug-resistant bacteria
- Potential biomarkers for inflammatory response in acute lung injury
- A low serum uric acid concentration predicts a poor prognosis in adult patients with candidemia
- Antitumor activity of recombinant oncolytic vaccinia virus with human IL2
- ALKBH5 inhibits TNF-α-induced apoptosis of HUVECs through Bcl-2 pathway
- Risk prediction of cardiovascular disease using machine learning classifiers
- Value of ultrasonography parameters in diagnosing polycystic ovary syndrome
- Bioinformatics analysis reveals three key genes and four survival genes associated with youth-onset NSCLC
- Identification of autophagy-related biomarkers in patients with pulmonary arterial hypertension based on bioinformatics analysis
- Protective effects of glaucocalyxin A on the airway of asthmatic mice
- Overexpression of miR-100-5p inhibits papillary thyroid cancer progression via targeting FZD8
- Bioinformatics-based analysis of SUMOylation-related genes in hepatocellular carcinoma reveals a role of upregulated SAE1 in promoting cell proliferation
- Effectiveness and clinical benefits of new anti-diabetic drugs: A real life experience
- Identification of osteoporosis based on gene biomarkers using support vector machine
- Tanshinone IIA reverses oxaliplatin resistance in colorectal cancer through microRNA-30b-5p/AVEN axis
- miR-212-5p inhibits nasopharyngeal carcinoma progression by targeting METTL3
- Association of ST-T changes with all-cause mortality among patients with peripheral T-cell lymphomas
- LINC00665/miRNAs axis-mediated collagen type XI alpha 1 correlates with immune infiltration and malignant phenotypes in lung adenocarcinoma
- The perinatal factors that influence the excretion of fecal calprotectin in premature-born children
- Effect of femoral head necrosis cystic area on femoral head collapse and stress distribution in femoral head: A clinical and finite element study
- Does the use of 3D-printed cones give a chance to postpone the use of megaprostheses in patients with large bone defects in the knee joint?
- lncRNA HAGLR modulates myocardial ischemia–reperfusion injury in mice through regulating miR-133a-3p/MAPK1 axis
- Protective effect of ghrelin on intestinal I/R injury in rats
- In vivo knee kinematics of an innovative prosthesis design
- Relationship between the height of fibular head and the incidence and severity of knee osteoarthritis
- lncRNA WT1-AS attenuates hypoxia/ischemia-induced neuronal injury during cerebral ischemic stroke via miR-186-5p/XIAP axis
- Correlation of cardiac troponin T and APACHE III score with all-cause in-hospital mortality in critically ill patients with acute pulmonary embolism
- LncRNA LINC01857 reduces metastasis and angiogenesis in breast cancer cells via regulating miR-2052/CENPQ axis
- Endothelial cell-specific molecule 1 (ESM1) promoted by transcription factor SPI1 acts as an oncogene to modulate the malignant phenotype of endometrial cancer
- SELENBP1 inhibits progression of colorectal cancer by suppressing epithelial–mesenchymal transition
- Visfatin is negatively associated with coronary artery lesions in subjects with impaired fasting glucose
- Treatment and outcomes of mechanical complications of acute myocardial infarction during the Covid-19 era: A comparison with the pre-Covid-19 period. A systematic review and meta-analysis
- Neonatal stroke surveillance study protocol in the United Kingdom and Republic of Ireland
- Oncogenic role of TWF2 in human tumors: A pan-cancer analysis
- Mean corpuscular hemoglobin predicts the length of hospital stay independent of severity classification in patients with acute pancreatitis
- Association of gallstone and polymorphisms of UGT1A1*27 and UGT1A1*28 in patients with hepatitis B virus-related liver failure
- TGF-β1 upregulates Sar1a expression and induces procollagen-I secretion in hypertrophic scarring fibroblasts
- Antisense lncRNA PCNA-AS1 promotes esophageal squamous cell carcinoma progression through the miR-2467-3p/PCNA axis
- NK-cell dysfunction of acute myeloid leukemia in relation to the renin–angiotensin system and neurotransmitter genes
- The effect of dilution with glucose and prolonged injection time on dexamethasone-induced perineal irritation – A randomized controlled trial
- miR-146-5p restrains calcification of vascular smooth muscle cells by suppressing TRAF6
- Role of lncRNA MIAT/miR-361-3p/CCAR2 in prostate cancer cells
- lncRNA NORAD promotes lung cancer progression by competitively binding to miR-28-3p with E2F2
- Noninvasive diagnosis of AIH/PBC overlap syndrome based on prediction models
- lncRNA FAM230B is highly expressed in colorectal cancer and suppresses the maturation of miR-1182 to increase cell proliferation
- circ-LIMK1 regulates cisplatin resistance in lung adenocarcinoma by targeting miR-512-5p/HMGA1 axis
- LncRNA SNHG3 promoted cell proliferation, migration, and metastasis of esophageal squamous cell carcinoma via regulating miR-151a-3p/PFN2 axis
- Risk perception and affective state on work exhaustion in obstetrics during the COVID-19 pandemic
- lncRNA-AC130710/miR-129-5p/mGluR1 axis promote migration and invasion by activating PKCα-MAPK signal pathway in melanoma
- SNRPB promotes cell cycle progression in thyroid carcinoma via inhibiting p53
- Xylooligosaccharides and aerobic training regulate metabolism and behavior in rats with streptozotocin-induced type 1 diabetes
- Serpin family A member 1 is an oncogene in glioma and its translation is enhanced by NAD(P)H quinone dehydrogenase 1 through RNA-binding activity
- Silencing of CPSF7 inhibits the proliferation, migration, and invasion of lung adenocarcinoma cells by blocking the AKT/mTOR signaling pathway
- Ultrasound-guided lumbar plexus block versus transversus abdominis plane block for analgesia in children with hip dislocation: A double-blind, randomized trial
- Relationship of plasma MBP and 8-oxo-dG with brain damage in preterm
- Identification of a novel necroptosis-associated miRNA signature for predicting the prognosis in head and neck squamous cell carcinoma
- Delayed femoral vein ligation reduces operative time and blood loss during hip disarticulation in patients with extremity tumors
- The expression of ASAP3 and NOTCH3 and the clinicopathological characteristics of adult glioma patients
- Longitudinal analysis of factors related to Helicobacter pylori infection in Chinese adults
- HOXA10 enhances cell proliferation and suppresses apoptosis in esophageal cancer via activating p38/ERK signaling pathway
- Meta-analysis of early-life antibiotic use and allergic rhinitis
- Marital status and its correlation with age, race, and gender in prognosis of tonsil squamous cell carcinomas
- HPV16 E6E7 up-regulates KIF2A expression by activating JNK/c-Jun signal, is beneficial to migration and invasion of cervical cancer cells
- Amino acid profiles in the tissue and serum of patients with liver cancer
- Pain in critically ill COVID-19 patients: An Italian retrospective study
- Immunohistochemical distribution of Bcl-2 and p53 apoptotic markers in acetamiprid-induced nephrotoxicity
- Estradiol pretreatment in GnRH antagonist protocol for IVF/ICSI treatment
- Long non-coding RNAs LINC00689 inhibits the apoptosis of human nucleus pulposus cells via miR-3127-5p/ATG7 axis-mediated autophagy
- The relationship between oxygen therapy, drug therapy, and COVID-19 mortality
- Monitoring hypertensive disorders in pregnancy to prevent preeclampsia in pregnant women of advanced maternal age: Trial mimicking with retrospective data
- SETD1A promotes the proliferation and glycolysis of nasopharyngeal carcinoma cells by activating the PI3K/Akt pathway
- The role of Shunaoxin pills in the treatment of chronic cerebral hypoperfusion and its main pharmacodynamic components
- TET3 governs malignant behaviors and unfavorable prognosis of esophageal squamous cell carcinoma by activating the PI3K/AKT/GSK3β/β-catenin pathway
- Associations between morphokinetic parameters of temporary-arrest embryos and the clinical prognosis in FET cycles
- Long noncoding RNA WT1-AS regulates trophoblast proliferation, migration, and invasion via the microRNA-186-5p/CADM2 axis
- The incidence of bronchiectasis in chronic obstructive pulmonary disease
- Integrated bioinformatics analysis shows integrin alpha 3 is a prognostic biomarker for pancreatic cancer
- Inhibition of miR-21 improves pulmonary vascular responses in bronchopulmonary dysplasia by targeting the DDAH1/ADMA/NO pathway
- Comparison of hospitalized patients with severe pneumonia caused by COVID-19 and influenza A (H7N9 and H1N1): A retrospective study from a designated hospital
- lncRNA ZFAS1 promotes intervertebral disc degeneration by upregulating AAK1
- Pathological characteristics of liver injury induced by N,N-dimethylformamide: From humans to animal models
- lncRNA ELFN1-AS1 enhances the progression of colon cancer by targeting miR-4270 to upregulate AURKB
- DARS-AS1 modulates cell proliferation and migration of gastric cancer cells by regulating miR-330-3p/NAT10 axis
- Dezocine inhibits cell proliferation, migration, and invasion by targeting CRABP2 in ovarian cancer
- MGST1 alleviates the oxidative stress of trophoblast cells induced by hypoxia/reoxygenation and promotes cell proliferation, migration, and invasion by activating the PI3K/AKT/mTOR pathway
- Bifidobacterium lactis Probio-M8 ameliorated the symptoms of type 2 diabetes mellitus mice by changing ileum FXR-CYP7A1
- circRNA DENND1B inhibits tumorigenicity of clear cell renal cell carcinoma via miR-122-5p/TIMP2 axis
- EphA3 targeted by miR-3666 contributes to melanoma malignancy via activating ERK1/2 and p38 MAPK pathways
- Pacemakers and methylprednisolone pulse therapy in immune-related myocarditis concomitant with complete heart block
- miRNA-130a-3p targets sphingosine-1-phosphate receptor 1 to activate the microglial and astrocytes and to promote neural injury under the high glucose condition
- Review Articles
- Current management of cancer pain in Italy: Expert opinion paper
- Hearing loss and brain disorders: A review of multiple pathologies
- The rationale for using low-molecular weight heparin in the therapy of symptomatic COVID-19 patients
- Amyotrophic lateral sclerosis and delayed onset muscle soreness in light of the impaired blink and stretch reflexes – watch out for Piezo2
- Interleukin-35 in autoimmune dermatoses: Current concepts
- Recent discoveries in microbiota dysbiosis, cholangiocytic factors, and models for studying the pathogenesis of primary sclerosing cholangitis
- Advantages of ketamine in pediatric anesthesia
- Congenital adrenal hyperplasia. Role of dentist in early diagnosis
- Migraine management: Non-pharmacological points for patients and health care professionals
- Atherogenic index of plasma and coronary artery disease: A systematic review
- Physiological and modulatory role of thioredoxins in the cellular function
- Case Reports
- Intrauterine Bakri balloon tamponade plus cervical cerclage for the prevention and treatment of postpartum haemorrhage in late pregnancy complicated with acute aortic dissection: Case series
- A case of successful pembrolizumab monotherapy in a patient with advanced lung adenocarcinoma: Use of multiple biomarkers in combination for clinical practice
- Unusual neurological manifestations of bilateral medial medullary infarction: A case report
- Atypical symptoms of malignant hyperthermia: A rare causative mutation in the RYR1 gene
- A case report of dermatomyositis with the missed diagnosis of non-small cell lung cancer and concurrence of pulmonary tuberculosis
- A rare case of endometrial polyp complicated with uterine inversion: A case report and clinical management
- Spontaneous rupturing of splenic artery aneurysm: Another reason for fatal syncope and shock (Case report and literature review)
- Fungal infection mimicking COVID-19 infection – A case report
- Concurrent aspergillosis and cystic pulmonary metastases in a patient with tongue squamous cell carcinoma
- Paraganglioma-induced inverted takotsubo-like cardiomyopathy leading to cardiogenic shock successfully treated with extracorporeal membrane oxygenation
- Lineage switch from lymphoma to myeloid neoplasms: First case series from a single institution
- Trismus during tracheal extubation as a complication of general anaesthesia – A case report
- Simultaneous treatment of a pubovesical fistula and lymph node metastasis secondary to multimodal treatment for prostate cancer: Case report and review of the literature
- Two case reports of skin vasculitis following the COVID-19 immunization
- Ureteroiliac fistula after oncological surgery: Case report and review of the literature
- Synchronous triple primary malignant tumours in the bladder, prostate, and lung harbouring TP53 and MEK1 mutations accompanied with severe cardiovascular diseases: A case report
- Huge mucinous cystic neoplasms with adhesion to the left colon: A case report and literature review
- Commentary
- Commentary on “Clinicopathological features of programmed cell death-ligand 1 expression in patients with oral squamous cell carcinoma”
- Rapid Communication
- COVID-19 fear, post-traumatic stress, growth, and the role of resilience
- Erratum
- Erratum to “Tollip promotes hepatocellular carcinoma progression via PI3K/AKT pathway”
- Erratum to “Effect of femoral head necrosis cystic area on femoral head collapse and stress distribution in femoral head: A clinical and finite element study”
- Erratum to “lncRNA NORAD promotes lung cancer progression by competitively binding to miR-28-3p with E2F2”
- Retraction
- Expression and role of ABIN1 in sepsis: In vitro and in vivo studies
- Retraction to “miR-519d downregulates LEP expression to inhibit preeclampsia development”
- Special Issue Computational Intelligence Methodologies Meets Recurrent Cancers - Part II
- Usefulness of close surveillance for rectal cancer patients after neoadjuvant chemoradiotherapy