HAGLR promotes neuron differentiation through the miR-130a-3p-MeCP2 axis
-
Bo Wei


Abstract
Parkinson’s disease (PD) is a prevalent neurodegenerative disease. Currently, the molecular mechanisms underlying the progressions of PD are not fully understood. The human neuroblastoma cell line SH-SY5Y has been widely used as an in vitro model for PD. This study aims to investigate the molecular mechanisms of the non-coding RNA-mediated SH-SY5Y differentiation induced by retinoic acid (RA). By microArray analysis, lncRNA HAGLR was observed to be significantly upregulated during the RA-induced SH-SY5Y differentiation. Silencing HAGLR blocked the RA-induced SH-SY5Y differentiation. Moreover, bioinformatical analysis illustrated that miR-130a-3p contains binding sites for HAGLR. The RNA-pull down assay and luciferase assay demonstrated that HAGLR functioned as a ceRNA of miR-130a-3p in SH-SY5Y cells. Overexpression of miR-130a-3p effectively inhibited SH-SY5Y differentiation. We identified MeCP2, a vital molecule in neuronal diseases, to be a direct target of miR-130a-3p in SH-SY5Y cells by western blot and luciferase assays. The rescue experiments verified that recovery of miR-130a-3p in HAGLR-overexpressing SH-SY5Y cells could successfully overcome the RA-induced SH-SY5Y differentiation by targeting MeCP2. In summary, this study reveals a potential molecular mechanism for the lncRNA-HAGLR-promoted in vitro neuron differentiation by targeting the miR-130a-3p-MeCP2 axis, contributing to the understanding of the pathogenesis and progression of PD.
1 Introduction
Parkinson’s disease (PD), a neurodegenerative disorder, is one of the most prevalent neurodegenerative diseases worldwide [1]. Symptoms of PD generally develop over years, resulting in socioeconomic burden [1,2]. The molecular mechanisms underlying PD and the effective disease-modifying treatments are still under clinical investigation [3]. The human neuroblastoma SH-SY5Y cell line is an in vitro model for PD research since it synthesizes both dopamine (DA) and noradrenaline (NA) [4]. In addition, retinoic acid (RA) has been used to induce the in vitro neuronal phenotypes of SH-SY5Y cells by inducing terminal neural differentiation [5]. However, the precise molecular mechanisms for the RA-induced SH-SY5Y differentiation are not fully understood.
lncRNAs, which are non-coding RNAs with relatively larger sizes (>200 nucleotides), have been extensively studied as important regulators for diverse diseases [6,7]. Accumulating evidence revealed that lncRNAs function by specifically binding to DNA, RNA, or proteins to regulate gene expressions [8]. Moreover, studies uncovered that lncRNAs act as competitive endogenous RNAs (ceRNA) of miRNAs by sponging them to de-repress the expressions of miRNA target genes [9]. lncRNA HAGLR (HOXD antisense growth-associated long non-coding RNA) has been reported to be positively associated with various diseases [10,11,12,13]. Currently, the biological roles of HAGLR in neuronal diseases such as PD have not been elucidated.
Methyl CpG binding protein 2 (MeCP2), which specifically binds to methylated cytosines on DNA, is a member of the methyl-CpG-binding domain (MBD) protein family [14]. Studies reported that MeCP2 was the most abundant molecule in the adult brain and tightly correlated with diverse neuronal processes during neurodevelopment [15]. Moreover, mutations in the MeCP2 gene cause the Rett syndrome (RTT), which is a neurologic condition affecting young girls [16] and lead to the MeCP2 duplication syndrome (MDS) [17]. These studies suggest MeCP2 is an important diagnostic biomarker and therapeutic target for neuronal diseases. However, the precise roles and molecular mechanisms of MeCP2 in PD are still under investigation.
In this study, we investigated the biological functions of HAGLR and MeCP2 during the RA-induced neuron differentiation of the SH-SY5Y neuroblastoma cell line. From lncRNA-microarray analysis, for the first time, we discovered HAGLR was upregulated from the in vitro neuronal differentiation model. The lncRNA–miRNA ceRNA network and the targets of miRNA will be identified.
2 Materials and methods
2.1 Cell culture and treatment
Human SH-SY5Y and SK-N-MC cells were cultured in Dulbecco’s modified Eagle’s medium/ F12 medium (DMEM/F12) (Thermo Fisher Scientific, Shanghai, China) supplemented with 10% fetal bovine serum (FBS) (Thermo Fisher Scientific, Shanghai, China) with 100 units/mL penicillin and 100 μg/mL streptomycin (Thermo Fisher Scientific, Shanghai, China) at 37°C in a humidified atmosphere with 5% CO2. RA induction was performed as previously described [18]. Cells were treated with 5 µM RA for 3–7 days. Morphological changes were monitored under a brightfield microscope. RA was purchased from Sigma-Aldrich (Shanghai, China). Mouse anti-MeCP2 (#M6818) and β-actin antibodies (#A5441) were purchased from Sigma-Aldrich (Shanghai, China).
2.2 Transfections of shRNA, miRNA, and plasmid DNA
Transfections were performed using Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA) according to the manufacturer’s protocols. SH-SY5Y cells (1 × 106) were plated in 60 mm wells for 24 h. siRNA, miRNAs, shRNA, and their negative controls were synthesized by GenePharma Inc. (Shanghai, China) and transfected at 50 nM for 48 h. Plasmid DNA was transfected at 2 µg/well for 48 h. Cells were then treated for 48 h with RA. Experiments were conducted in triplicate and repeated at least three times.
2.3 lncRNA–miRNA and miRNA–mRNA interaction analysis
The HAGLR-miR-130a-3p and miR-130a-3p-MeCP2 3′UTR bindings were predicted from Targetscan.org and starBase of ENCORI http://starbase.sysu.edu.cn/ according to previous reports [19]. The online non-coding RNA service predicts binging of miRNA with lncRNAs or mRNA targets of miRNA by searching for the presence of conserved 8mer, 7mer, and 6mer sites that match the seed region of each miRNA. Prediction of starBase was from 108 CLIP-Seq (PAR-CLIP, HITS-CLIP, iCLIP, and CLASH) data sets generated by 37 independent studies.
2.4 Quantitative RT-PCR
Total RNA was extracted from SH-SY5Y cells using the RNeasy Mini Kit (Qiagen, Shanghai, China) according to the manufacturer’s instructions. The quality and concentration of RNA samples were determined using a Nanodrop 2000 spectrophotometer (Thermo Fisher Scientific, Shanghai, China). Total RNA was treated with DNase I, followed by reverse transcription into cDNA using the PrimeScript RT Master Mix (TaKaRa, Dalian, China) according to the manufacturer’s instructions. The qRT-PCRs were performed using the SYBR Green qPCR Master Mix (ThermoFisher Scientific, Waltham, MA, USA) in an ABI PRISM 7500 Real-Time System. A total of 40 amplification cycles were applied with the following settings: denaturation at 95°C for 15 s, annealing at 58°C for 30 s, and extension at 72°C for 42 s. The relative gene expression was determined by the 2−ΔΔCt method. β-Actin was an internal control for lncRNA and MeCP2 mRNA detection, and U6 was an internal control for miRNA detection. Experiments were performed in triplicate and repeated three times.
2.5 RNA pull-down assay
The RNA pull-down assay was performed as described previously. Briefly, biotin-labeled scramble, sense, or antisense lncRNA HAGLR DNA oligomers (RiboBio) were incubated with the SH-SY5Y cell lysate followed by incubation for 1 h; then streptavidin-coupled agarose beads (Thermo Fisher Scientific, Shanghai, China) were added to pull down the RNA–RNA complexes. The miR-130a-3p expressions were analyzed from the RNA–RNA complexes by quantitative RT-PCR (qRT-PCR). Experiments were repeated three times.
2.6 Luciferase assay
The wild-type (WT) and mutated (Mut) HAGLR or MeCP2 3′UTR were amplified and cloned into the pGL3-control luciferase reporter vector system (Promega, Madison, WI, USA). SH-SY5Y cells were co-transfected with control miRNA or miR-130a-3p with WT or Mut HAGLR or MeCP2 3′UTR using Lipofectamine 2000 (Invitrogen, Carlsbad, CA, USA) for 72 h. Luciferase activity was measured using a Dual-Luciferase reporter assay system (Promega, Madison, WI, USA) according to the protocol from the kit. The experiment was performed in triplicate and repeated three times.
2.7 Microarray analysis
Total RNA was extracted from SH-SY5Y cells using the RNeasy Mini Kit (Qiagen, Shanghai, China) according to the manufacturer’s instructions. RNA was quantified using a Nanodrop 2000 spectrophotometer (Thermo Fisher Scientific, Shanghai, China). Integrities of RNA samples were assessed using an Agilent Bioanalyzer 2100 (Agilent Technologies, Santa Clara, CA, USA). RNA was amplified and labeled. The Cy3-labeled complementary RNA was hybridized using the Gene Expression Hybridization Kit (Agilent Technologies, Santa Clara, CA, USA). Slides were scanned using an Agilent Microarray Scanner (Agilent Technologies, Santa Clara, CA, USA). The MicroArray results were analyzed using Agilent Feature Extraction software (Agilent Technologies, Inc.) and GeneSpring GX v11.5.1 software (Agilent Technologies, Inc.).
2.8 Western blot
Proteins from SH-SY5Y cells were extracted using RIPA buffer (ThermoFisher Scientific, Inc.) with a 1× protease inhibitor cocktail (Thermo Fisher Scientific Inc.). After 20 min incubation on ice, lysates were centrifuged at 10,000 g for 10 min at 4°C. An equal amount of protein from each sample was separated by SDS-PAGE and transferred to polyvinylidene difluoride membranes (PVDFs) (Millipore, Bedford, MA, USA) followed by blockage for 1 h at room temperature with 4% bovine serum albumin (BSA). Membranes were incubated with primary antibodies at 1:1,000 overnight at 4°C. After washing with phosphate buffered saline with tween 20 (PBST), membranes were incubated with secondary antibody (1:3,000) for 1 h at room temperature. Blots were visualized with enhanced chemiluminescence (Millipore, Bedford, MA, USA). β-Actin was used as a loading control. Experiments were repeated three times.
2.9 Statistical analysis
Data were analyzed using Prism version 7.0 (GraphPad Software, La Jolla, CA, USA). Data were expressed as mean ± standard deviation (SD). Experiments were repeated three times. Independent unpaired Student’s t-test was applied to compare differences between two groups. Values of p < 0.05 were considered statistically significant.
3 Results
3.1 HAGLR is upregulated during the RA-induced neuron differentiation
We evaluated the cellular mechanisms of neuron differentiation using RA-induction of SH-SY5Y cells as an in vitro model, which has been widely used in PD research [4]. The results showed that the RA treatment (5 μM) of SH-SY5Y cells for 4 days induced remarkable morphologic differentiation with neurite extension compared with control cells that showed no morphological differentiation (Figure 1a and Figure S1a). Moreover, the neuron differentiation marker, GAP43, was induced under RA treatments (Figure S1b), indicating successful neuronal induction by RA. Accumulating studies revealed that non-coding RNAs play vital roles during neuron differentiation [7,8]. To investigate the molecular mechanisms, we performed a lncRNA microArray analysis to profile lncRNA expressions using SH-SY5Y cells with or without RA induction. Among the differentially expressed lncRNAs in SH-SY5Y cells with RA induction, we observed that lncRNA HAGLR was significantly upregulated by the RA treatment (Figure 1b), suggesting that HALGR is positively associated with neuron differentiation. The results from microArray were further validated by qRT-PCR in SH-SY5Y cells with RA induction at 3, 5, and 7 days. In addition, the RA-induced HAGLR upregulation at 5 and 7 days was observed in another neuroblastoma cell line, SK-N-MC (Figure S2a). To assess the biological functions of HAGLR in neuron differentiation, HAGLR was knocked down in SH-SY5Y cells (Figure 1d). As we expected, SH-SY5Y and SK-N-MC cells with HAGLR silencing displayed little response to RA treatments compared with the control shRNA transfected cells (Figure 1e and Figure S2b). Taken together, these results consistently demonstrated that HAGLR promotes neuron differentiation from an RA-induced in vitro SH-SY5Y model.

Roles of HAGLR in RA-induced neuron differentiation. (a) Effects of RA treatment on cell morphology. SH-SY5Y cells were treated with 5 µM RA for 5 days. (b) SH-SY5Y cells were treated with RA using the above protocol. RNAs were isolated and lncRNA-microArray analysis was performed in duplicate. (c) SH-SY5Y cells were treated with 5 µM RA for 3, 5, and 7 days. HAGLR expressions were examined by qRT-PCR. (d) SH-SY5Y cells were transfected with control shRNA or HAGLR shRNA for 48 h, and expression of HAGLR was determined by qRT-PCR. (e) SH-SY5Y cells were transfected with control shRNA or HAGLR shRNA for 48 h, followed by treatments with 5 µM RA for 5 and 7 days. Cell morphologies were examined under a microscope, and the neurite length was measured. *p < 0.05; **p < 0.01; and ***p < 0.001.
3.2 HAGLR sponges miR-130a-3p as a ceRNA in SH-SY5Y cells
We next explored the molecular mechanisms underlying the HAGLR-promoted neuron differentiation. Previous studies uncovered that lncRNAs function by interfering with target miRNAs as molecular sponges [9]. The downregulation of miRNAs by lncRNA leads to de-repression of their target mRNAs [20]. To identify the potential miRNA targets of HAGLR, we analyzed the HAGLR–miRNA interaction through the non-coding RNA database, starBase2.0. We observed miR-130a-3p, which plays an important role during neuron development and neuronal diseases [21], contains HAGLR binding sites (Figure 2a). We then evaluated the functional roles of miR-130a-3p during RA-induced neuron differentiation. qPCR results remarkedly showed a reverse phenotype between HAGLR and miR-130a-3p, which was significantly downregulated in SH-SY5Y and SK-N-MC cells with RA induction at 3, 5, and 7 days (Figure 2b and Figure S3a). Moreover, although RA treatments suppressed miR-130a-3p expressions, exogenous overexpression of miR-130a-3p (Figure S3b and c) rendered cells to maintain relatively high miR-130a-3p expression levels (Figure S3d and e). Expectedly, SH-SY5Y and SK-N-MC cells with exogenous overexpression of miR-130a-3p showed no effective morphological changes in the neurite extension under RA treatments (Figure 2c and Figure S3f). To assess whether HAGLR downregulates miR-130a-3p, HAGLR was overexpressed or silenced in SH-SY5Y cells. Expectedly, overexpression of HAGLR effectively blocked the miR-130a-3p expressions, while knockdown of HAGLR significantly upregulated miR-130a-3p expressions (Figure 2d). To validate the binding of miR-130a-3p on HAGLR, we performed an RNA pull-down assay. The biotin-labeled scramble, sense, or antisense DNA probe of HAGLR was incubated with SH-SY5Y cell lysates. The qRT-PCR results demonstrated that only the antisense HAGLR DNA probe pulled down the enriched miR-130a-3p (Figure 2e). The endogenous miR-130a-3p could not be effectively pulled down by the scramble or sense HAGLR probe (Figure 2e), suggesting that HAGLR specifically interacts with miR-130a-3p in neuroblastoma cells. To validate whether HAGLR directly binds on the seeding region of miR-130a-3p, SH-SY5Y cells were co-transfected with a luciferase vector containing the wild-type HAGLR (WT-HAGLR) or the binding site mutant HAGLR (Mut-HAGLR) and control miRNA or miR-130a-3p. Luciferase activity of cells with WT-HAGLR and miR-130a-3p co-transfection was significantly suppressed compared with that from Mut-HAGLR and miRNA-130a-3p co-transfection (Figure 2f). In summary, these results demonstrated that HAGLR sponges miR-130a-3p by forming a ceRNA network.

HAGLR sponges miR-130a-3p to negatively regulate its expression. (a) Bioinformatics analysis of potential binding of HAGLR on miR-130a-3p. (b) SH-SY5Y cells were treated with 5 µM RA for 3, 5, and 7 days. miR-130a-3p expressions were examined by qRT-PCR. (c) SH-SY5Y cells were transfected with control miRNA or miR-130a-3p for 48 h, followed by treatments with 5 µM RA for 5 and 7 days. Cell morphologies were examined under a microscope, and the neurite length was measured. (d) SH-SY5Y cells were transfected with control, the HAGLR overexpression plasmid, or HAGLR shRNA for 48 h; miR-130a-3p expressions were examined by qRT-PCR. (e) SH-SY5Y cell lysates were incubated with biotin-labeled scramble, sense, or antisense HAGLR DNA probes. Biotin pull-down assay was performed, and miR-130a-3p expression was assessed by qRT-PCR. (f) Dual-luciferase reporter assay was performed in SH-SY5Y cells with co-transfection of WT-HAGLR or Mut-HAGLR with control miRNA, or miR-130a-3p. *p < 0.05; **p < 0.01; and ***p < 0.001.
3.3 miR-130a-3p directly targets MeCP2 and inhibits the RA-induced neuron differentiation
Since miRNAs function via targeting mRNA 3′UTR to downregulate target mRNA expressions [20], we performed miRNA–mRNA interaction analysis from the non-coding RNA database, starBase 2.0. Interestingly, the 3′UTR of MeCP2, which is an important molecule in neuron differentiation and neuronal diseases, contains conserved miR-130a-3p binding sites through multiple species (Figure 3a and b). To examine whether miR-130a-3p could suppress protein expression of MeCP2, miR-130a-3p or control miRNA was transfected into SH-SY5Y cells. The results from western blot showed that overexpression of miR-130a-3p effectively blocked MeCP2 protein expression in SH-SY5Y cells (Figure 3d). To validate the direct binding of miR-130a-3p on 3′UTR of MeCP2 mRNA, SH-SY5Y cells were co-transfected with luciferase vector containing WT-MeCP2 3′UTR or the miR-130a-3p binding site mutant MeCP2 3′UTR (Mut-MeCP2) and control miRNA or miR-130a-3p. Luciferase activity of the vector containing WT-MeCP2 was remarkedly blocked by miR-130a-3p (Figure 3d). Expectedly, the luciferase activity of the Mut-MeCP2 vector has no significant change by miR-130a-3p overexpression (Figure 3d). Together, these results demonstrated that miR-130a-3p could directly target 3′UTR of MeCP2.

MeCP2 is directly targeted by miR-130a-3p. (a) Bioinformatics analysis of potential binding of miR-130a-3p on 3′UTR of MeCP2. (b) The conserved binding sites of miR-130a-3p on 3′UTR of MeCP2 in multiple species. (c) SH-SY5Y cells were transfected with control or miR-130a-3p for 48 h; the protein expression of MeCP2 was detected by western blot. (d) Dual-luciferase reporter assay was performed in SH-SY5Y cells with co-transfection of WT-MeCP2 or Mut-MeCP2 with control miRNA, or miR-130a-3p. **p < 0.01.
3.4 MeCP2 is induced and promotes the RA-induced neuron differentiation
We continue to evaluate the roles of MeCP2 in RA-induced neuron differentiation. Under RA treatments, protein and mRNA expressions of MeCP2 were significantly upregulated (Figure 4a and b and Figure S4a). Furthermore, MeCP2 was knocked down by siRNA in SH-SY5Y cells. The results in Figure 4c and Figure S4b illustrate that SH-SY5Y cells with downregulated MeCP2 expression exhibited unchanged neurite extension in response to RA induction compared with control cells, indicating that MeCP2 promotes the process of neuron differentiation.

Roles of MeCP2 in the RA-induced neuron differentiation. (a) SH-SY5Y cells were treated with 5 µM RA for 3, 5, and 7 days. Protein and (b) mRNA expressions of MeCP2 were detected. (c) SH-SY5Y cells were transfected with control siRNA or MeCP2 siRNA for 48 h, followed by treatments with 5 µM RA for 5 and 7 days. Cell morphologies were examined under a microscope, and the neurite length was measured. *p < 0.05; **p < 0.01; and ***p < 0.001.
3.5 Restoration of MeCP2 in miR-130a-3p overexpressing SH-SY5Y cells rescues the RA-induced neuron differentiation
Rescue experiments were performed to validate whether miR-130a-3p blocked the RA-induced neuron differentiation through targeting MeCP2. SH-SY5Y cells were transfected with control miRNAs, miR-130a-3p alone or with MeCP2 overexpression plasmid. Co-transfection of miR-130a-3p with MeCP2 successfully recovered MeCP2 protein levels compared with miR-130a-3p overexpression cells (Figure 5a). Moreover, the expected results demonstrated that SH-SY5Y cells with the restoration of MeCP2 rescued the RA-induced neuron differentiation phenotype (Figure 5b). These rescue experiments further confirmed that miR-130a-3p directly targets MeCP2 to inhibit neuron differentiation.
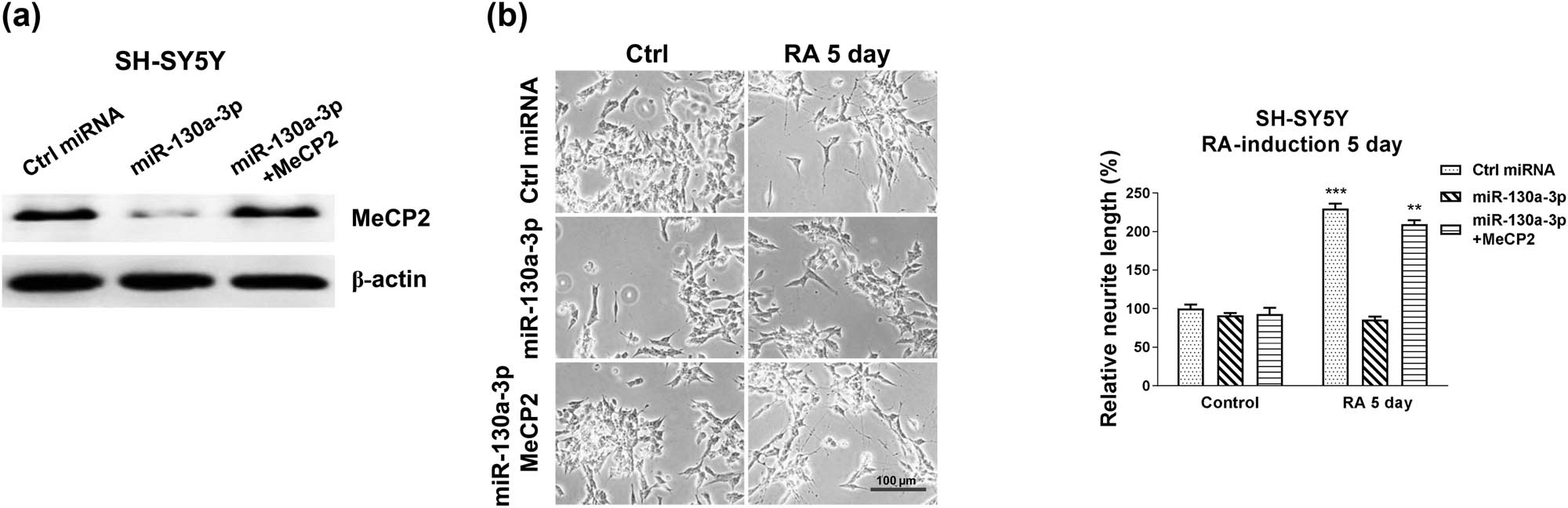
Restoration of MeCP2 rescues the miR-130a-3p suppressed neuron differentiation. (a) SH-SY5Y cells were transfected with control, miR-130a-3p alone, or with MeCP2 overexpression plasmid for 48 h; protein expression of MeCP2 was determined by western blot. (b) The above transfected cells were treated with 5 µM RA for 5 and 7 days. Cell morphologies were examined under a microscope, and the neurite length was measured. *p < 0.05 and **p < 0.01.
3.6 HAGLR promoted neuron differentiation by targeting the miR-130a-3p-MeCP2 axis
Finally, we assessed whether neuron differentiation was regulated by the HAGLR-miR-130a-3p-MeCP2 axis. SH-SY5Y cells were transfected with control, HAGLR overexpression plasmid alone, or with miR-130a-3p. qRT-PCR and western blot results demonstrated that co-transfection of HAGLR with miR-130a-3p successfully restored miR-130a-3p and MeCP2 protein levels compared with HAGLR overexpressing cells (Figure 6a and b). As expected, SH-SY5Y cells with co-transfection of HAGLR and miR-130a-3p re-suppressed the RA-induced neuron differentiation phenotype compared with that from HAGLR overexpressing cells (Figure 6c). In summary, these mechanism rescue experiments consolidated a HAGLR-miR-130a-3p-MeCP2 axis that contributes to the processes of neuron differentiation.
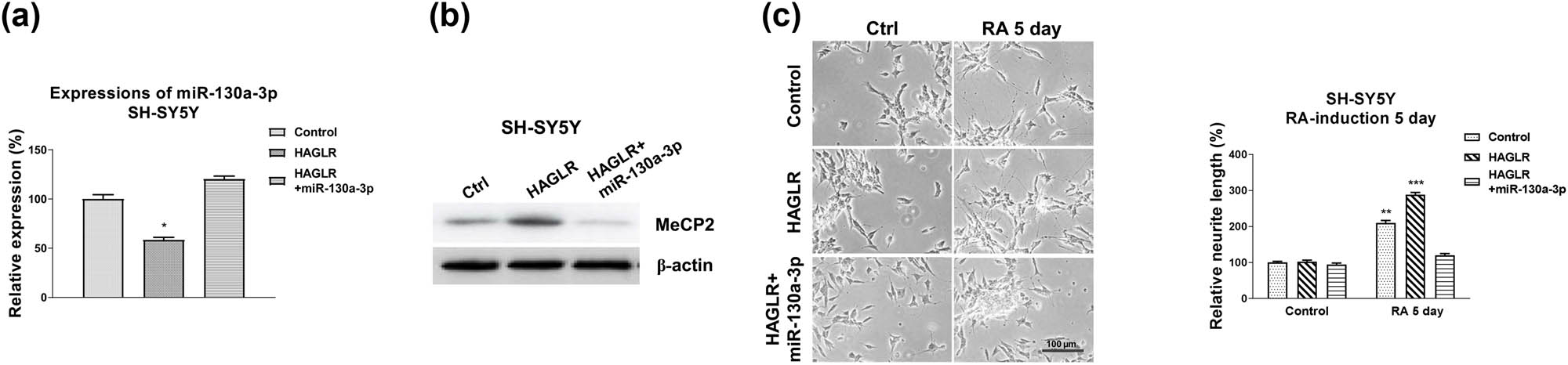
The HAGLR-miR-130a-3p-MeCP2 axis in neuron differentiation. (a) SH-SY5Y cells were transfected with control, HAGLR alone, or with miR-130a-3p for 48 h; expression of miR-130a-3p was detected by qRT-PCR and (b) protein expression of MeCP2 was determined by western blot. (c) The above transfected cells were treated with 5 µM RA for 5 and 7 days. Cell morphologies were examined under a microscope, and the neurite length was measured. *p < 0.05 and **p < 0.01.
4 Discussion
PD is one of the most prevalent neurodegenerative diseases worldwide [1,2]. Currently, the molecular mechanisms underlying PD and the effective disease-modifying approaches are not fully understood. The human neuroblastoma SH-SY5Y cell line is widely studied for investigating the molecular and cellular mechanisms underlying PD pathogenesis and progression since it synthesizes both DA and NA [4]. Moreover, although the SH-SY5Y cell line displays a number of cancerous characteristics, most genes regulations and signal pathways in PD pathogenesis are intact [4]. It is difficult to obtain and maintain human dopaminergic neurons as primary cells, which are mainly affected in PD [4]. Thus, the neuroblastoma SH-SY5Y cell line is a widely used in vitro model for PD research. Currently, the biological roles of lncRNA HAGLR in neuronal diseases remain unclear. In this study, we first reported the non-coding RNA-regulated neuron differentiation using SH-SY5Y cells as an in vitro model. Under RA induction, we detected that HAGLR was significantly upregulated in SH-SY5Y cells from both microArray analysis and qPCR results. In addition, silencing HAGLR effectively blocked the RA-induced SH-SY5Y differentiation based on the observations that the neurite extension of SH-SY5Y cells with lower HAGLR expression showed little response to RA treatments, suggesting an essential role of HAGLR in SH-SY5Y cell differentiation.
Accumulating studies demonstrated that miRNAs play regulatory roles in neuron development and neuronal diseases [20]. Moreover, the lncRNA–miRNA interaction has been shown to control vital functions during the molecular and cellular processes of human malignancies, including neuronal diseases [8,9]. A recent study reported that miR-130a-3p regulates VEGFR-2 expression in sensory and motor neurons during development [22]. Moreover, the miR-130a-3p/DAPK1 axis was known to regulate the pathophysiology of neonatal hypoxic-ischemia encephalopathy [23], suggesting miR-130-3p to be a potential therapeutic target for the hypoxic ischemia encephalopathy treatment. However, the miR-130a-3p-mediated neuron differentiation has not been investigated. Here, we show that miR-130a-3p was remarkedly downregulated in SH-SY5Y cells under RA induction. Overexpression of miR-130a-3p inhibited the morphological changes of SH-SY5Y cells under RA treatments. Bioinformatics analysis revealed that HAGLR contains miR-130a-3p binding sites. The predicted ceRNA network was further validated by the RNA pull-down assay and luciferase assay.
MeCP2 is the most abundant methyl-DNA binding domain family member in the adult brain and is tightly correlated with diverse neuronal processes during neurodevelopment [15]. It was known that mutations in the MeCP2 gene cause Rett syndrome (RTT), a neurologic condition affecting primarily young girls [16]. Interestingly, girls with RTT exhibiting motor deficits showed similar phenotypes to those in PD [24], suggesting MeCP2 is involved in the defects of the nigrostriatal pathway. A recent study used the 6-hydroxydopamine-induced human neuroblastoma cell (SH-SY5Y cell) injury as a cell model of PD [25]. They described that overexpression of MeCP2 was able to ameliorate the 6-hydroxydopamine-induced apoptosis of SH-SY5Y cells, suggesting that MeCP2 is a potential therapeutic target for the treatment of PD [25]. Currently, the precise roles and molecular mechanisms of MeCP2 in PD have not been elucidated. Here, we described that the expressions of MeCP2 were significantly upregulated in SH-SY5Y cells by RA induction. One advantage of this study is that we illustrated the direct binding of miR-130a-3p on 3′UTR of MeCP2 in SH-SY5Y cells. Furthermore, the rescue experiments validated that the miR-130a-3p-inhibited SH-SY5Y cell differentiation was by direct targeting of MeCP2. Thus, it was possible to target the above signaling pathways in neuronal disease to improve the clinical therapeutic outcomes. These strategies still have limitations owing to the fact that the neuroblastoma cell line is not purely neuron cells since it is oncogenically transformed with catecholaminergic, resulting in different physiological characteristics compared to those from the normal DAergic neuronal features. In addition, the above in vitro signaling pathway needs to be verified in animal models.
In summary, this study reports a non-coding RNA-based molecular mechanism for the PD using SH-SY5Y cells as an in vitro model. HAGLR positively regulates SH-SY5Y cell differentiation, which is induced by RA via targeting the miR-130a-3p-MeCP2 axis, contributing to an extensive understanding of the pathogenesis and progression of PD.
Acknowledgement
The authors appreciate the contributions from all medical doctors and research scientists from the Department of Neurology, Shaoxing People’s Hospital, for generously supporting this study.
-
Funding information: No funds.
-
Author contributions: B.W. and Y.Q.X. designed the experiments and wrote the manuscript. B.W., G.R.X., Ch.L.W., and Y.Q.X. carried out the experiments and analyzed the data.
-
Conflict of interest: The authors declare that there are no conflicts of interest.
-
Data availability statement: The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Appendix

Effects of retinoic acid treatment on neuron differentiation. (a) SH-SY5Y cells were treated with RA at 5 µM for 5 days, the relative neurite length was calculated. (b) Expressions of GAP43 in SH-SY5Y cells under RA treatments at 5 days and 7 days were determined by qRT-PCR. **, p < 0.01; ***, p < 0.001.

Effects of retinoic acid treatment on SK-N-MC cells. (a) Expressions of HAGLR in SK-N-MC cells under RA treatments for 3, 5 and 7 days were determined by qRT-PCR. (b) The relative neurite length was calculated in SK-N-MC cells without or with HAGLR silencing under RA treatments for 5 and 7 days. *, p < 0.05; **, p < 0.01; ***, p < 0.001.

miR-130a-3p suppresses RA-induced neuron differentiation. (a) Expressions of miR-130a-3p in SK-N-MC cells under RA treatments at 3, 5 and 7 days were determined by qRT-PCR. (b) Effects of miR-130a-3p overexpression in SH-SY5Y and (c) SK-N-MC cells. (d) Detection of miR-130a-3p expressions in SH-SY5Y and (e) SK-N-MC cells without or with miR-130a-3p overexpression under RA treatments for 5 and 7 days. (f) The relative neurite length was calculated in SK-N-MC cells without or with miR-130a-3p overexpression under RA treatments for 5 and 7 days. *, p < 0.05; **, p < 0.01; ***, p < 0.001.

Silencing MeCP2 suppresses RA-induced neuron differentiation. (a) SK-N-MC cells under RA treatments for 3, 5 and 7 days, expressions of MeCP2 were determined by qRT-PCR. (b) The relative neurite length was calculated in SK-N-MC cells without or with MeCP2 silencing under RA treatments for 5 and 7 days. *, p < 0.05; **, p < 0.01; ***, p < 0.001.
References
[1] Reich SG , Savitt JM . Parkinson’s disease. Med Clin North Am. 2019;103:337–50.10.1016/B978-0-323-05674-8.50194-8Suche in Google Scholar
[2] Ascherio A , Schwarzschild MA . The epidemiology of Parkinson’s disease: risk factors and prevention. Lancet Neurol. 2016;15:1257–72.10.1016/S1474-4422(16)30230-7Suche in Google Scholar
[3] Raza C , Anjum R , Shakeel NUA . Parkinson’s disease: mechanisms, translational models and management strategies. Life Sci. 2019;226:77–90.10.1016/j.lfs.2019.03.057Suche in Google Scholar PubMed
[4] Xicoy H , Wieringa B , Martens GJ . The SH-SY5Y cell line in Parkinson’s disease research: a systematic review. Mol Neurodegener. 2017;12:10.10.1186/s13024-017-0149-0Suche in Google Scholar PubMed PubMed Central
[5] Gonulalan EM , Bayazeid O , Yalcin FN , Demirezer LO . The roles of valerenic acid on BDNF expression in the SH-SY5Y cell. Saudi Pharm J. 2018;26:960–4.10.1016/j.jsps.2018.05.005Suche in Google Scholar PubMed PubMed Central
[6] Peng WX , Koirala P , Mo YY . lncRNA-mediated regulation of cell signaling in cancer. Oncogene. 2017;36:5661–7.10.1038/onc.2017.184Suche in Google Scholar PubMed PubMed Central
[7] Lyu Y , Bai L , Qin C . Long noncoding RNAs in neurodevelopment and Parkinson’s disease. Animal Model Exp Med. 2019;2:239–51.10.1002/ame2.12093Suche in Google Scholar PubMed PubMed Central
[8] Riva P , Ratti A , Venturin M . The long non-coding RNAs in neurodegenerative diseases: novel mechanisms of pathogenesis. Curr Alzheimer Res. 2016;13:1219–31.10.2174/1567205013666160622112234Suche in Google Scholar PubMed
[9] Paraskevopoulou MD , Hatzigeorgiou AG . Analyzing MiRNA-lncRNA Interactions. Methods Mol Biol. 2016;1402:271–86.10.1007/978-1-4939-3378-5_21Suche in Google Scholar PubMed
[10] Yang C , Shen S , Zheng X , Ye K , Sun Y , Lu Y , et al. Long noncoding RNA HAGLR acts as a microRNA-143-5p sponge to regulate epithelial-mesenchymal transition and metastatic potential in esophageal cancer by regulating LAMP3. FASEB J. 2019;33:10490–504.10.1096/fj.201802543RRSuche in Google Scholar PubMed
[11] Lu C , Ma J , Cai D . Increased HAGLR expression promotes non-small cell lung cancer proliferation and invasion via enhanced de novo lipogenesis. Tumour Biol. 2017;39:1010428317697574.10.1177/1010428317697574Suche in Google Scholar PubMed
[12] Sun W , Nie W , Wang Z , Zhang H , Li Y , Fang X . Lnc HAGLR promotes colon cancer progression through sponging miR-185-5p and activating CDK4 and CDK6 in vitro and in vivo. Onco Targets Ther. 2020;13:5913–25.10.2147/OTT.S246092Suche in Google Scholar PubMed PubMed Central
[13] Zakany J , Darbellay F , Mascrez B , Necsulea A , Duboule D . Control of growth and gut maturation by HoxD genes and the associated lncRNA Haglr. Proc Natl Acad Sci USA. 2017;114:E9290–9.10.1073/pnas.1712511114Suche in Google Scholar PubMed PubMed Central
[14] Horvath PM , Monteggia LM . MeCP2 as an activator of gene expression. Trends Neurosci. 2018;41:72–4.10.1016/j.tins.2017.11.005Suche in Google Scholar PubMed PubMed Central
[15] Sharma K , Singh J , Frost EE , Pillai PP . MeCP2 in central nervous system glial cells: current updates. Acta Neurobiol Exp (Wars). 2018;78:30–40.10.21307/ane-2018-007Suche in Google Scholar
[16] Shah RR , Bird AP . MeCP2 mutations: progress towards understanding and treating Rett syndrome. Genome Med. 2017;9:17.10.1186/s13073-017-0411-7Suche in Google Scholar PubMed PubMed Central
[17] Gulmez Karaca K , Brito DVC , Oliveira AMM . MeCP2: A critical regulator of chromatin in neurodevelopment and adult brain function. Int J Mol Sci. 2019;20:4577.10.3390/ijms20184577Suche in Google Scholar PubMed PubMed Central
[18] Teppola H , Sarkanen JR , Jalonen TO , Linne ML . Morphological differentiation towards neuronal phenotype of SH-SY5Y neuroblastoma cells by estradiol, retinoic acid and cholesterol. Neurochem Res. 2016;41:731–47.10.1007/s11064-015-1743-6Suche in Google Scholar PubMed PubMed Central
[19] Li JH , Liu S , Zhou H , Qu LH , Yang JH . StarBase v2.0: decoding miRNA-ceRNA, miRNA-ncRNA and protein-RNA interaction networks from large-scale CLIP-Seq data. Nucleic Acids Res. 2014;42:D92–7.10.1093/nar/gkt1248Suche in Google Scholar PubMed PubMed Central
[20] Mohr AM , Mott JL . Overview of microRNA biology. Semin Liver Dis. 2015;35:3–11.10.1055/s-0034-1397344Suche in Google Scholar PubMed PubMed Central
[21] Glaesel K , May C , Marcus K , Matschke V , Theiss C , Theis V . miR-129-5p and miR-130a-3p regulate VEGFR-2 expression in sensory and motor neurons during development. Int J Mol Sci. 2020;21:3839.10.3390/ijms21113839Suche in Google Scholar PubMed PubMed Central
[22] Glaesel K , May C , Marcus K , Matschke V , Theiss C , Theis V . miR-129-5p and miR-130a-3p regulate VEGFR-2 expression in sensory and motor neurons during development. Int J Mol Sci. 2020;21:3839.10.3390/ijms21113839Suche in Google Scholar
[23] Feng M , Zhu X , Zhuo C . H19/miR-130a-3p/DAPK1 axis regulates the pathophysiology of neonatal hypoxic-ischemia encephalopathy. Neurosci Res. 2021;163:52–62.10.1016/j.neures.2020.03.005Suche in Google Scholar PubMed
[24] Gantz SC , Ford CP , Neve KA , Williams JT . Loss of Mecp2 in substantia nigra dopamine neurons compromises the nigrostriatal pathway. J Neurosci. 2011;31:12629–37.10.1523/JNEUROSCI.0684-11.2011Suche in Google Scholar PubMed PubMed Central
[25] Xie T , Zhang J , Yuan X , Yang J , Ding W , Huang X , et al. Is X-linked methyl-CpG binding protein 2 a new target for the treatment of Parkinson’s disease. Neural Regen Res. 2013;8:1948–57.Suche in Google Scholar
© 2021 Bo Wei et al., published by De Gruyter
This work is licensed under the Creative Commons Attribution 4.0 International License.
Artikel in diesem Heft
- Research Articles
- Identification of ZG16B as a prognostic biomarker in breast cancer
- Behçet’s disease with latent Mycobacterium tuberculosis infection
- Erratum
- Erratum to “Suffering from Cerebral Small Vessel Disease with and without Metabolic Syndrome”
- Research Articles
- GPR37 promotes the malignancy of lung adenocarcinoma via TGF-β/Smad pathway
- Expression and role of ABIN1 in sepsis: In vitro and in vivo studies
- Additional baricitinib loading dose improves clinical outcome in COVID-19
- The co-treatment of rosuvastatin with dapagliflozin synergistically inhibited apoptosis via activating the PI3K/AKt/mTOR signaling pathway in myocardial ischemia/reperfusion injury rats
- SLC12A8 plays a key role in bladder cancer progression and EMT
- LncRNA ATXN8OS enhances tamoxifen resistance in breast cancer
- Case Report
- Serratia marcescens as a cause of unfavorable outcome in the twin pregnancy
- Spleno-adrenal fusion mimicking an adrenal metastasis of a renal cell carcinoma: A case report and embryological background
- Research Articles
- TRIM25 contributes to the malignancy of acute myeloid leukemia and is negatively regulated by microRNA-137
- CircRNA circ_0004370 promotes cell proliferation, migration, and invasion and inhibits cell apoptosis of esophageal cancer via miR-1301-3p/COL1A1 axis
- LncRNA XIST regulates atherosclerosis progression in ox-LDL-induced HUVECs
- Potential role of IFN-γ and IL-5 in sepsis prediction of preterm neonates
- Rapid Communication
- COVID-19 vaccine: Call for employees in international transportation industries and international travelers as the first priority in global distribution
- Case Report
- Rare squamous cell carcinoma of the kidney with concurrent xanthogranulomatous pyelonephritis: A case report and review of the literature
- An infertile female delivered a baby after removal of primary renal carcinoid tumor
- Research Articles
- Hypertension, BMI, and cardiovascular and cerebrovascular diseases
- Case Report
- Coexistence of bilateral macular edema and pale optic disc in the patient with Cohen syndrome
- Research Articles
- Correlation between kinematic sagittal parameters of the cervical lordosis or head posture and disc degeneration in patients with posterior neck pain
- Review Articles
- Hepatoid adenocarcinoma of the lung: An analysis of the Surveillance, Epidemiology, and End Results (SEER) database
- Research Articles
- Thermography in the diagnosis of carpal tunnel syndrome
- Pemetrexed-based first-line chemotherapy had particularly prominent objective response rate for advanced NSCLC: A network meta-analysis
- Comparison of single and double autologous stem cell transplantation in multiple myeloma patients
- The influence of smoking in minimally invasive spinal fusion surgery
- Impact of body mass index on left atrial dimension in HOCM patients
- Expression and clinical significance of CMTM1 in hepatocellular carcinoma
- miR-142-5p promotes cervical cancer progression by targeting LMX1A through Wnt/β-catenin pathway
- Comparison of multiple flatfoot indicators in 5–8-year-old children
- Early MRI imaging and follow-up study in cerebral amyloid angiopathy
- Intestinal fatty acid-binding protein as a biomarker for the diagnosis of strangulated intestinal obstruction: A meta-analysis
- miR-128-3p inhibits apoptosis and inflammation in LPS-induced sepsis by targeting TGFBR2
- Dynamic perfusion CT – A promising tool to diagnose pancreatic ductal adenocarcinoma
- Biomechanical evaluation of self-cinching stitch techniques in rotator cuff repair: The single-loop and double-loop knot stitches
- Review Articles
- The ambiguous role of mannose-binding lectin (MBL) in human immunity
- Case Report
- Membranous nephropathy with pulmonary cryptococcosis with improved 1-year follow-up results: A case report
- Fertility problems in males carrying an inversion of chromosome 10
- Acute myeloid leukemia with leukemic pleural effusion and high levels of pleural adenosine deaminase: A case report and review of literature
- Metastatic renal Ewing’s sarcoma in adult woman: Case report and review of the literature
- Burkitt-like lymphoma with 11q aberration in a patient with AIDS and a patient without AIDS: Two cases reports and literature review
- Skull hemophilia pseudotumor: A case report
- Judicious use of low-dosage corticosteroids for non-severe COVID-19: A case report
- Adult-onset citrullinaemia type II with liver cirrhosis: A rare cause of hyperammonaemia
- Clinicopathologic features of Good’s syndrome: Two cases and literature review
- Fatal immune-related hepatitis with intrahepatic cholestasis and pneumonia associated with camrelizumab: A case report and literature review
- Research Articles
- Effects of hydroxyethyl starch and gelatin on the risk of acute kidney injury following orthotopic liver transplantation: A multicenter retrospective comparative clinical study
- Significance of nucleic acid positive anal swab in COVID-19 patients
- circAPLP2 promotes colorectal cancer progression by upregulating HELLS by targeting miR-335-5p
- Ratios between circulating myeloid cells and lymphocytes are associated with mortality in severe COVID-19 patients
- Risk factors of left atrial appendage thrombus in patients with non-valvular atrial fibrillation
- Clinical features of hypertensive patients with COVID-19 compared with a normotensive group: Single-center experience in China
- Surgical myocardial revascularization outcomes in Kawasaki disease: systematic review and meta-analysis
- Decreased chromobox homologue 7 expression is associated with epithelial–mesenchymal transition and poor prognosis in cervical cancer
- FGF16 regulated by miR-520b enhances the cell proliferation of lung cancer
- Platelet-rich fibrin: Basics of biological actions and protocol modifications
- Accurate diagnosis of prostate cancer using logistic regression
- miR-377 inhibition enhances the survival of trophoblast cells via upregulation of FNDC5 in gestational diabetes mellitus
- Prognostic significance of TRIM28 expression in patients with breast carcinoma
- Integrative bioinformatics analysis of KPNA2 in six major human cancers
- Exosomal-mediated transfer of OIP5-AS1 enhanced cell chemoresistance to trastuzumab in breast cancer via up-regulating HMGB3 by sponging miR-381-3p
- A four-lncRNA signature for predicting prognosis of recurrence patients with gastric cancer
- Knockdown of circ_0003204 alleviates oxidative low-density lipoprotein-induced human umbilical vein endothelial cells injury: Circulating RNAs could explain atherosclerosis disease progression
- Propofol postpones colorectal cancer development through circ_0026344/miR-645/Akt/mTOR signal pathway
- Knockdown of lncRNA TapSAKI alleviates LPS-induced injury in HK-2 cells through the miR-205/IRF3 pathway
- COVID-19 severity in relation to sociodemographics and vitamin D use
- Clinical analysis of 11 cases of nocardiosis
- Cis-regulatory elements in conserved non-coding sequences of nuclear receptor genes indicate for crosstalk between endocrine systems
- Four long noncoding RNAs act as biomarkers in lung adenocarcinoma
- Real-world evidence of cytomegalovirus reactivation in non-Hodgkin lymphomas treated with bendamustine-containing regimens
- Relation between IL-8 level and obstructive sleep apnea syndrome
- circAGFG1 sponges miR-28-5p to promote non-small-cell lung cancer progression through modulating HIF-1α level
- Nomogram prediction model for renal anaemia in IgA nephropathy patients
- Effect of antibiotic use on the efficacy of nivolumab in the treatment of advanced/metastatic non-small cell lung cancer: A meta-analysis
- NDRG2 inhibition facilitates angiogenesis of hepatocellular carcinoma
- A nomogram for predicting metabolic steatohepatitis: The combination of NAMPT, RALGDS, GADD45B, FOSL2, RTP3, and RASD1
- Clinical and prognostic features of MMP-2 and VEGF in AEG patients
- The value of miR-510 in the prognosis and development of colon cancer
- Functional implications of PABPC1 in the development of ovarian cancer
- Prognostic value of preoperative inflammation-based predictors in patients with bladder carcinoma after radical cystectomy
- Sublingual immunotherapy increases Treg/Th17 ratio in allergic rhinitis
- Prediction of improvement after anterior cruciate ligament reconstruction
- Effluent Osteopontin levels reflect the peritoneal solute transport rate
- circ_0038467 promotes PM2.5-induced bronchial epithelial cell dysfunction
- Significance of miR-141 and miR-340 in cervical squamous cell carcinoma
- Association between hair cortisol concentration and metabolic syndrome
- Microvessel density as a prognostic indicator of prostate cancer: A systematic review and meta-analysis
- Characteristics of BCR–ABL gene variants in patients of chronic myeloid leukemia
- Knee alterations in rheumatoid arthritis: Comparison of US and MRI
- Long non-coding RNA TUG1 aggravates cerebral ischemia and reperfusion injury by sponging miR-493-3p/miR-410-3p
- lncRNA MALAT1 regulated ATAD2 to facilitate retinoblastoma progression via miR-655-3p
- Development and validation of a nomogram for predicting severity in patients with hemorrhagic fever with renal syndrome: A retrospective study
- Analysis of COVID-19 outbreak origin in China in 2019 using differentiation method for unusual epidemiological events
- Laparoscopic versus open major liver resection for hepatocellular carcinoma: A case-matched analysis of short- and long-term outcomes
- Travelers’ vaccines and their adverse events in Nara, Japan
- Association between Tfh and PGA in children with Henoch–Schönlein purpura
- Can exchange transfusion be replaced by double-LED phototherapy?
- circ_0005962 functions as an oncogene to aggravate NSCLC progression
- Circular RNA VANGL1 knockdown suppressed viability, promoted apoptosis, and increased doxorubicin sensitivity through targeting miR-145-5p to regulate SOX4 in bladder cancer cells
- Serum intact fibroblast growth factor 23 in healthy paediatric population
- Algorithm of rational approach to reconstruction in Fournier’s disease
- A meta-analysis of exosome in the treatment of spinal cord injury
- Src-1 and SP2 promote the proliferation and epithelial–mesenchymal transition of nasopharyngeal carcinoma
- Dexmedetomidine may decrease the bupivacaine toxicity to heart
- Hypoxia stimulates the migration and invasion of osteosarcoma via up-regulating the NUSAP1 expression
- Long noncoding RNA XIST knockdown relieves the injury of microglia cells after spinal cord injury by sponging miR-219-5p
- External fixation via the anterior inferior iliac spine for proximal femoral fractures in young patients
- miR-128-3p reduced acute lung injury induced by sepsis via targeting PEL12
- HAGLR promotes neuron differentiation through the miR-130a-3p-MeCP2 axis
- Phosphoglycerate mutase 2 is elevated in serum of patients with heart failure and correlates with the disease severity and patient’s prognosis
- Cell population data in identifying active tuberculosis and community-acquired pneumonia
- Prognostic value of microRNA-4521 in non-small cell lung cancer and its regulatory effect on tumor progression
- Mean platelet volume and red blood cell distribution width is associated with prognosis in premature neonates with sepsis
- 3D-printed porous scaffold promotes osteogenic differentiation of hADMSCs
- Association of gene polymorphisms with women urinary incontinence
- Influence of COVID-19 pandemic on stress levels of urologic patients
- miR-496 inhibits proliferation via LYN and AKT pathway in gastric cancer
- miR-519d downregulates LEP expression to inhibit preeclampsia development
- Comparison of single- and triple-port VATS for lung cancer: A meta-analysis
- Fluorescent light energy modulates healing in skin grafted mouse model
- Silencing CDK6-AS1 inhibits LPS-induced inflammatory damage in HK-2 cells
- Predictive effect of DCE-MRI and DWI in brain metastases from NSCLC
- Severe postoperative hyperbilirubinemia in congenital heart disease
- Baicalin improves podocyte injury in rats with diabetic nephropathy by inhibiting PI3K/Akt/mTOR signaling pathway
- Clinical factors predicting ureteral stent failure in patients with external ureteral compression
- Novel H2S donor proglumide-ADT-OH protects HUVECs from ox-LDL-induced injury through NF-κB and JAK/SATA pathway
- Triple-Endobutton and clavicular hook: A propensity score matching analysis
- Long noncoding RNA MIAT inhibits the progression of diabetic nephropathy and the activation of NF-κB pathway in high glucose-treated renal tubular epithelial cells by the miR-182-5p/GPRC5A axis
- Serum exosomal miR-122-5p, GAS, and PGR in the non-invasive diagnosis of CAG
- miR-513b-5p inhibits the proliferation and promotes apoptosis of retinoblastoma cells by targeting TRIB1
- Fer exacerbates renal fibrosis and can be targeted by miR-29c-3p
- The diagnostic and prognostic value of miR-92a in gastric cancer: A systematic review and meta-analysis
- Prognostic value of α2δ1 in hypopharyngeal carcinoma: A retrospective study
- No significant benefit of moderate-dose vitamin C on severe COVID-19 cases
- circ_0000467 promotes the proliferation, metastasis, and angiogenesis in colorectal cancer cells through regulating KLF12 expression by sponging miR-4766-5p
- Downregulation of RAB7 and Caveolin-1 increases MMP-2 activity in renal tubular epithelial cells under hypoxic conditions
- Educational program for orthopedic surgeons’ influences for osteoporosis
- Expression and function analysis of CRABP2 and FABP5, and their ratio in esophageal squamous cell carcinoma
- GJA1 promotes hepatocellular carcinoma progression by mediating TGF-β-induced activation and the epithelial–mesenchymal transition of hepatic stellate cells
- lncRNA-ZFAS1 promotes the progression of endometrial carcinoma by targeting miR-34b to regulate VEGFA expression
- Anticoagulation is the answer in treating noncritical COVID-19 patients
- Effect of late-onset hemorrhagic cystitis on PFS after haplo-PBSCT
- Comparison of Dako HercepTest and Ventana PATHWAY anti-HER2 (4B5) tests and their correlation with silver in situ hybridization in lung adenocarcinoma
- VSTM1 regulates monocyte/macrophage function via the NF-κB signaling pathway
- Comparison of vaginal birth outcomes in midwifery-led versus physician-led setting: A propensity score-matched analysis
- Treatment of osteoporosis with teriparatide: The Slovenian experience
- New targets of morphine postconditioning protection of the myocardium in ischemia/reperfusion injury: Involvement of HSP90/Akt and C5a/NF-κB
- Superenhancer–transcription factor regulatory network in malignant tumors
- β-Cell function is associated with osteosarcopenia in middle-aged and older nonobese patients with type 2 diabetes: A cross-sectional study
- Clinical features of atypical tuberculosis mimicking bacterial pneumonia
- Proteoglycan-depleted regions of annular injury promote nerve ingrowth in a rabbit disc degeneration model
- Effect of electromagnetic field on abortion: A systematic review and meta-analysis
- miR-150-5p affects AS plaque with ASMC proliferation and migration by STAT1
- MALAT1 promotes malignant pleural mesothelioma by sponging miR-141-3p
- Effects of remifentanil and propofol on distant organ lung injury in an ischemia–reperfusion model
- miR-654-5p promotes gastric cancer progression via the GPRIN1/NF-κB pathway
- Identification of LIG1 and LIG3 as prognostic biomarkers in breast cancer
- MitoQ inhibits hepatic stellate cell activation and liver fibrosis by enhancing PINK1/parkin-mediated mitophagy
- Dissecting role of founder mutation p.V727M in GNE in Indian HIBM cohort
- circATP2A2 promotes osteosarcoma progression by upregulating MYH9
- Prognostic role of oxytocin receptor in colon adenocarcinoma
- Review Articles
- The function of non-coding RNAs in idiopathic pulmonary fibrosis
- Efficacy and safety of therapeutic plasma exchange in stiff person syndrome
- Role of cesarean section in the development of neonatal gut microbiota: A systematic review
- Small cell lung cancer transformation during antitumor therapies: A systematic review
- Research progress of gut microbiota and frailty syndrome
- Recommendations for outpatient activity in COVID-19 pandemic
- Rapid Communication
- Disparity in clinical characteristics between 2019 novel coronavirus pneumonia and leptospirosis
- Use of microspheres in embolization for unruptured renal angiomyolipomas
- COVID-19 cases with delayed absorption of lung lesion
- A triple combination of treatments on moderate COVID-19
- Social networks and eating disorders during the Covid-19 pandemic
- Letter
- COVID-19, WHO guidelines, pedagogy, and respite
- Inflammatory factors in alveolar lavage fluid from severe COVID-19 pneumonia: PCT and IL-6 in epithelial lining fluid
- COVID-19: Lessons from Norway tragedy must be considered in vaccine rollout planning in least developed/developing countries
- What is the role of plasma cell in the lamina propria of terminal ileum in Good’s syndrome patient?
- Case Report
- Rivaroxaban triggered multifocal intratumoral hemorrhage of the cabozantinib-treated diffuse brain metastases: A case report and review of literature
- CTU findings of duplex kidney in kidney: A rare duplicated renal malformation
- Synchronous primary malignancy of colon cancer and mantle cell lymphoma: A case report
- Sonazoid-enhanced ultrasonography and pathologic characters of CD68 positive cell in primary hepatic perivascular epithelioid cell tumors: A case report and literature review
- Persistent SARS-CoV-2-positive over 4 months in a COVID-19 patient with CHB
- Pulmonary parenchymal involvement caused by Tropheryma whipplei
- Mediastinal mixed germ cell tumor: A case report and literature review
- Ovarian female adnexal tumor of probable Wolffian origin – Case report
- Rare paratesticular aggressive angiomyxoma mimicking an epididymal tumor in an 82-year-old man: Case report
- Perimenopausal giant hydatidiform mole complicated with preeclampsia and hyperthyroidism: A case report and literature review
- Primary orbital ganglioneuroblastoma: A case report
- Primary aortic intimal sarcoma masquerading as intramural hematoma
- Sustained false-positive results for hepatitis A virus immunoglobulin M: A case report and literature review
- Peritoneal loose body presenting as a hepatic mass: A case report and review of the literature
- Chondroblastoma of mandibular condyle: Case report and literature review
- Trauma-induced complete pacemaker lead fracture 8 months prior to hospitalization: A case report
- Primary intradural extramedullary extraosseous Ewing’s sarcoma/peripheral primitive neuroectodermal tumor (PIEES/PNET) of the thoracolumbar spine: A case report and literature review
- Computer-assisted preoperative planning of reduction of and osteosynthesis of scapular fracture: A case report
- High quality of 58-month life in lung cancer patient with brain metastases sequentially treated with gefitinib and osimertinib
- Rapid response of locally advanced oral squamous cell carcinoma to apatinib: A case report
- Retrieval of intrarenal coiled and ruptured guidewire by retrograde intrarenal surgery: A case report and literature review
- Usage of intermingled skin allografts and autografts in a senior patient with major burn injury
- Retraction
- Retraction on “Dihydromyricetin attenuates inflammation through TLR4/NF-kappa B pathway”
- Special Issue Computational Intelligence Methodologies Meets Recurrent Cancers - Part I
- An artificial immune system with bootstrap sampling for the diagnosis of recurrent endometrial cancers
- Breast cancer recurrence prediction with ensemble methods and cost-sensitive learning
Artikel in diesem Heft
- Research Articles
- Identification of ZG16B as a prognostic biomarker in breast cancer
- Behçet’s disease with latent Mycobacterium tuberculosis infection
- Erratum
- Erratum to “Suffering from Cerebral Small Vessel Disease with and without Metabolic Syndrome”
- Research Articles
- GPR37 promotes the malignancy of lung adenocarcinoma via TGF-β/Smad pathway
- Expression and role of ABIN1 in sepsis: In vitro and in vivo studies
- Additional baricitinib loading dose improves clinical outcome in COVID-19
- The co-treatment of rosuvastatin with dapagliflozin synergistically inhibited apoptosis via activating the PI3K/AKt/mTOR signaling pathway in myocardial ischemia/reperfusion injury rats
- SLC12A8 plays a key role in bladder cancer progression and EMT
- LncRNA ATXN8OS enhances tamoxifen resistance in breast cancer
- Case Report
- Serratia marcescens as a cause of unfavorable outcome in the twin pregnancy
- Spleno-adrenal fusion mimicking an adrenal metastasis of a renal cell carcinoma: A case report and embryological background
- Research Articles
- TRIM25 contributes to the malignancy of acute myeloid leukemia and is negatively regulated by microRNA-137
- CircRNA circ_0004370 promotes cell proliferation, migration, and invasion and inhibits cell apoptosis of esophageal cancer via miR-1301-3p/COL1A1 axis
- LncRNA XIST regulates atherosclerosis progression in ox-LDL-induced HUVECs
- Potential role of IFN-γ and IL-5 in sepsis prediction of preterm neonates
- Rapid Communication
- COVID-19 vaccine: Call for employees in international transportation industries and international travelers as the first priority in global distribution
- Case Report
- Rare squamous cell carcinoma of the kidney with concurrent xanthogranulomatous pyelonephritis: A case report and review of the literature
- An infertile female delivered a baby after removal of primary renal carcinoid tumor
- Research Articles
- Hypertension, BMI, and cardiovascular and cerebrovascular diseases
- Case Report
- Coexistence of bilateral macular edema and pale optic disc in the patient with Cohen syndrome
- Research Articles
- Correlation between kinematic sagittal parameters of the cervical lordosis or head posture and disc degeneration in patients with posterior neck pain
- Review Articles
- Hepatoid adenocarcinoma of the lung: An analysis of the Surveillance, Epidemiology, and End Results (SEER) database
- Research Articles
- Thermography in the diagnosis of carpal tunnel syndrome
- Pemetrexed-based first-line chemotherapy had particularly prominent objective response rate for advanced NSCLC: A network meta-analysis
- Comparison of single and double autologous stem cell transplantation in multiple myeloma patients
- The influence of smoking in minimally invasive spinal fusion surgery
- Impact of body mass index on left atrial dimension in HOCM patients
- Expression and clinical significance of CMTM1 in hepatocellular carcinoma
- miR-142-5p promotes cervical cancer progression by targeting LMX1A through Wnt/β-catenin pathway
- Comparison of multiple flatfoot indicators in 5–8-year-old children
- Early MRI imaging and follow-up study in cerebral amyloid angiopathy
- Intestinal fatty acid-binding protein as a biomarker for the diagnosis of strangulated intestinal obstruction: A meta-analysis
- miR-128-3p inhibits apoptosis and inflammation in LPS-induced sepsis by targeting TGFBR2
- Dynamic perfusion CT – A promising tool to diagnose pancreatic ductal adenocarcinoma
- Biomechanical evaluation of self-cinching stitch techniques in rotator cuff repair: The single-loop and double-loop knot stitches
- Review Articles
- The ambiguous role of mannose-binding lectin (MBL) in human immunity
- Case Report
- Membranous nephropathy with pulmonary cryptococcosis with improved 1-year follow-up results: A case report
- Fertility problems in males carrying an inversion of chromosome 10
- Acute myeloid leukemia with leukemic pleural effusion and high levels of pleural adenosine deaminase: A case report and review of literature
- Metastatic renal Ewing’s sarcoma in adult woman: Case report and review of the literature
- Burkitt-like lymphoma with 11q aberration in a patient with AIDS and a patient without AIDS: Two cases reports and literature review
- Skull hemophilia pseudotumor: A case report
- Judicious use of low-dosage corticosteroids for non-severe COVID-19: A case report
- Adult-onset citrullinaemia type II with liver cirrhosis: A rare cause of hyperammonaemia
- Clinicopathologic features of Good’s syndrome: Two cases and literature review
- Fatal immune-related hepatitis with intrahepatic cholestasis and pneumonia associated with camrelizumab: A case report and literature review
- Research Articles
- Effects of hydroxyethyl starch and gelatin on the risk of acute kidney injury following orthotopic liver transplantation: A multicenter retrospective comparative clinical study
- Significance of nucleic acid positive anal swab in COVID-19 patients
- circAPLP2 promotes colorectal cancer progression by upregulating HELLS by targeting miR-335-5p
- Ratios between circulating myeloid cells and lymphocytes are associated with mortality in severe COVID-19 patients
- Risk factors of left atrial appendage thrombus in patients with non-valvular atrial fibrillation
- Clinical features of hypertensive patients with COVID-19 compared with a normotensive group: Single-center experience in China
- Surgical myocardial revascularization outcomes in Kawasaki disease: systematic review and meta-analysis
- Decreased chromobox homologue 7 expression is associated with epithelial–mesenchymal transition and poor prognosis in cervical cancer
- FGF16 regulated by miR-520b enhances the cell proliferation of lung cancer
- Platelet-rich fibrin: Basics of biological actions and protocol modifications
- Accurate diagnosis of prostate cancer using logistic regression
- miR-377 inhibition enhances the survival of trophoblast cells via upregulation of FNDC5 in gestational diabetes mellitus
- Prognostic significance of TRIM28 expression in patients with breast carcinoma
- Integrative bioinformatics analysis of KPNA2 in six major human cancers
- Exosomal-mediated transfer of OIP5-AS1 enhanced cell chemoresistance to trastuzumab in breast cancer via up-regulating HMGB3 by sponging miR-381-3p
- A four-lncRNA signature for predicting prognosis of recurrence patients with gastric cancer
- Knockdown of circ_0003204 alleviates oxidative low-density lipoprotein-induced human umbilical vein endothelial cells injury: Circulating RNAs could explain atherosclerosis disease progression
- Propofol postpones colorectal cancer development through circ_0026344/miR-645/Akt/mTOR signal pathway
- Knockdown of lncRNA TapSAKI alleviates LPS-induced injury in HK-2 cells through the miR-205/IRF3 pathway
- COVID-19 severity in relation to sociodemographics and vitamin D use
- Clinical analysis of 11 cases of nocardiosis
- Cis-regulatory elements in conserved non-coding sequences of nuclear receptor genes indicate for crosstalk between endocrine systems
- Four long noncoding RNAs act as biomarkers in lung adenocarcinoma
- Real-world evidence of cytomegalovirus reactivation in non-Hodgkin lymphomas treated with bendamustine-containing regimens
- Relation between IL-8 level and obstructive sleep apnea syndrome
- circAGFG1 sponges miR-28-5p to promote non-small-cell lung cancer progression through modulating HIF-1α level
- Nomogram prediction model for renal anaemia in IgA nephropathy patients
- Effect of antibiotic use on the efficacy of nivolumab in the treatment of advanced/metastatic non-small cell lung cancer: A meta-analysis
- NDRG2 inhibition facilitates angiogenesis of hepatocellular carcinoma
- A nomogram for predicting metabolic steatohepatitis: The combination of NAMPT, RALGDS, GADD45B, FOSL2, RTP3, and RASD1
- Clinical and prognostic features of MMP-2 and VEGF in AEG patients
- The value of miR-510 in the prognosis and development of colon cancer
- Functional implications of PABPC1 in the development of ovarian cancer
- Prognostic value of preoperative inflammation-based predictors in patients with bladder carcinoma after radical cystectomy
- Sublingual immunotherapy increases Treg/Th17 ratio in allergic rhinitis
- Prediction of improvement after anterior cruciate ligament reconstruction
- Effluent Osteopontin levels reflect the peritoneal solute transport rate
- circ_0038467 promotes PM2.5-induced bronchial epithelial cell dysfunction
- Significance of miR-141 and miR-340 in cervical squamous cell carcinoma
- Association between hair cortisol concentration and metabolic syndrome
- Microvessel density as a prognostic indicator of prostate cancer: A systematic review and meta-analysis
- Characteristics of BCR–ABL gene variants in patients of chronic myeloid leukemia
- Knee alterations in rheumatoid arthritis: Comparison of US and MRI
- Long non-coding RNA TUG1 aggravates cerebral ischemia and reperfusion injury by sponging miR-493-3p/miR-410-3p
- lncRNA MALAT1 regulated ATAD2 to facilitate retinoblastoma progression via miR-655-3p
- Development and validation of a nomogram for predicting severity in patients with hemorrhagic fever with renal syndrome: A retrospective study
- Analysis of COVID-19 outbreak origin in China in 2019 using differentiation method for unusual epidemiological events
- Laparoscopic versus open major liver resection for hepatocellular carcinoma: A case-matched analysis of short- and long-term outcomes
- Travelers’ vaccines and their adverse events in Nara, Japan
- Association between Tfh and PGA in children with Henoch–Schönlein purpura
- Can exchange transfusion be replaced by double-LED phototherapy?
- circ_0005962 functions as an oncogene to aggravate NSCLC progression
- Circular RNA VANGL1 knockdown suppressed viability, promoted apoptosis, and increased doxorubicin sensitivity through targeting miR-145-5p to regulate SOX4 in bladder cancer cells
- Serum intact fibroblast growth factor 23 in healthy paediatric population
- Algorithm of rational approach to reconstruction in Fournier’s disease
- A meta-analysis of exosome in the treatment of spinal cord injury
- Src-1 and SP2 promote the proliferation and epithelial–mesenchymal transition of nasopharyngeal carcinoma
- Dexmedetomidine may decrease the bupivacaine toxicity to heart
- Hypoxia stimulates the migration and invasion of osteosarcoma via up-regulating the NUSAP1 expression
- Long noncoding RNA XIST knockdown relieves the injury of microglia cells after spinal cord injury by sponging miR-219-5p
- External fixation via the anterior inferior iliac spine for proximal femoral fractures in young patients
- miR-128-3p reduced acute lung injury induced by sepsis via targeting PEL12
- HAGLR promotes neuron differentiation through the miR-130a-3p-MeCP2 axis
- Phosphoglycerate mutase 2 is elevated in serum of patients with heart failure and correlates with the disease severity and patient’s prognosis
- Cell population data in identifying active tuberculosis and community-acquired pneumonia
- Prognostic value of microRNA-4521 in non-small cell lung cancer and its regulatory effect on tumor progression
- Mean platelet volume and red blood cell distribution width is associated with prognosis in premature neonates with sepsis
- 3D-printed porous scaffold promotes osteogenic differentiation of hADMSCs
- Association of gene polymorphisms with women urinary incontinence
- Influence of COVID-19 pandemic on stress levels of urologic patients
- miR-496 inhibits proliferation via LYN and AKT pathway in gastric cancer
- miR-519d downregulates LEP expression to inhibit preeclampsia development
- Comparison of single- and triple-port VATS for lung cancer: A meta-analysis
- Fluorescent light energy modulates healing in skin grafted mouse model
- Silencing CDK6-AS1 inhibits LPS-induced inflammatory damage in HK-2 cells
- Predictive effect of DCE-MRI and DWI in brain metastases from NSCLC
- Severe postoperative hyperbilirubinemia in congenital heart disease
- Baicalin improves podocyte injury in rats with diabetic nephropathy by inhibiting PI3K/Akt/mTOR signaling pathway
- Clinical factors predicting ureteral stent failure in patients with external ureteral compression
- Novel H2S donor proglumide-ADT-OH protects HUVECs from ox-LDL-induced injury through NF-κB and JAK/SATA pathway
- Triple-Endobutton and clavicular hook: A propensity score matching analysis
- Long noncoding RNA MIAT inhibits the progression of diabetic nephropathy and the activation of NF-κB pathway in high glucose-treated renal tubular epithelial cells by the miR-182-5p/GPRC5A axis
- Serum exosomal miR-122-5p, GAS, and PGR in the non-invasive diagnosis of CAG
- miR-513b-5p inhibits the proliferation and promotes apoptosis of retinoblastoma cells by targeting TRIB1
- Fer exacerbates renal fibrosis and can be targeted by miR-29c-3p
- The diagnostic and prognostic value of miR-92a in gastric cancer: A systematic review and meta-analysis
- Prognostic value of α2δ1 in hypopharyngeal carcinoma: A retrospective study
- No significant benefit of moderate-dose vitamin C on severe COVID-19 cases
- circ_0000467 promotes the proliferation, metastasis, and angiogenesis in colorectal cancer cells through regulating KLF12 expression by sponging miR-4766-5p
- Downregulation of RAB7 and Caveolin-1 increases MMP-2 activity in renal tubular epithelial cells under hypoxic conditions
- Educational program for orthopedic surgeons’ influences for osteoporosis
- Expression and function analysis of CRABP2 and FABP5, and their ratio in esophageal squamous cell carcinoma
- GJA1 promotes hepatocellular carcinoma progression by mediating TGF-β-induced activation and the epithelial–mesenchymal transition of hepatic stellate cells
- lncRNA-ZFAS1 promotes the progression of endometrial carcinoma by targeting miR-34b to regulate VEGFA expression
- Anticoagulation is the answer in treating noncritical COVID-19 patients
- Effect of late-onset hemorrhagic cystitis on PFS after haplo-PBSCT
- Comparison of Dako HercepTest and Ventana PATHWAY anti-HER2 (4B5) tests and their correlation with silver in situ hybridization in lung adenocarcinoma
- VSTM1 regulates monocyte/macrophage function via the NF-κB signaling pathway
- Comparison of vaginal birth outcomes in midwifery-led versus physician-led setting: A propensity score-matched analysis
- Treatment of osteoporosis with teriparatide: The Slovenian experience
- New targets of morphine postconditioning protection of the myocardium in ischemia/reperfusion injury: Involvement of HSP90/Akt and C5a/NF-κB
- Superenhancer–transcription factor regulatory network in malignant tumors
- β-Cell function is associated with osteosarcopenia in middle-aged and older nonobese patients with type 2 diabetes: A cross-sectional study
- Clinical features of atypical tuberculosis mimicking bacterial pneumonia
- Proteoglycan-depleted regions of annular injury promote nerve ingrowth in a rabbit disc degeneration model
- Effect of electromagnetic field on abortion: A systematic review and meta-analysis
- miR-150-5p affects AS plaque with ASMC proliferation and migration by STAT1
- MALAT1 promotes malignant pleural mesothelioma by sponging miR-141-3p
- Effects of remifentanil and propofol on distant organ lung injury in an ischemia–reperfusion model
- miR-654-5p promotes gastric cancer progression via the GPRIN1/NF-κB pathway
- Identification of LIG1 and LIG3 as prognostic biomarkers in breast cancer
- MitoQ inhibits hepatic stellate cell activation and liver fibrosis by enhancing PINK1/parkin-mediated mitophagy
- Dissecting role of founder mutation p.V727M in GNE in Indian HIBM cohort
- circATP2A2 promotes osteosarcoma progression by upregulating MYH9
- Prognostic role of oxytocin receptor in colon adenocarcinoma
- Review Articles
- The function of non-coding RNAs in idiopathic pulmonary fibrosis
- Efficacy and safety of therapeutic plasma exchange in stiff person syndrome
- Role of cesarean section in the development of neonatal gut microbiota: A systematic review
- Small cell lung cancer transformation during antitumor therapies: A systematic review
- Research progress of gut microbiota and frailty syndrome
- Recommendations for outpatient activity in COVID-19 pandemic
- Rapid Communication
- Disparity in clinical characteristics between 2019 novel coronavirus pneumonia and leptospirosis
- Use of microspheres in embolization for unruptured renal angiomyolipomas
- COVID-19 cases with delayed absorption of lung lesion
- A triple combination of treatments on moderate COVID-19
- Social networks and eating disorders during the Covid-19 pandemic
- Letter
- COVID-19, WHO guidelines, pedagogy, and respite
- Inflammatory factors in alveolar lavage fluid from severe COVID-19 pneumonia: PCT and IL-6 in epithelial lining fluid
- COVID-19: Lessons from Norway tragedy must be considered in vaccine rollout planning in least developed/developing countries
- What is the role of plasma cell in the lamina propria of terminal ileum in Good’s syndrome patient?
- Case Report
- Rivaroxaban triggered multifocal intratumoral hemorrhage of the cabozantinib-treated diffuse brain metastases: A case report and review of literature
- CTU findings of duplex kidney in kidney: A rare duplicated renal malformation
- Synchronous primary malignancy of colon cancer and mantle cell lymphoma: A case report
- Sonazoid-enhanced ultrasonography and pathologic characters of CD68 positive cell in primary hepatic perivascular epithelioid cell tumors: A case report and literature review
- Persistent SARS-CoV-2-positive over 4 months in a COVID-19 patient with CHB
- Pulmonary parenchymal involvement caused by Tropheryma whipplei
- Mediastinal mixed germ cell tumor: A case report and literature review
- Ovarian female adnexal tumor of probable Wolffian origin – Case report
- Rare paratesticular aggressive angiomyxoma mimicking an epididymal tumor in an 82-year-old man: Case report
- Perimenopausal giant hydatidiform mole complicated with preeclampsia and hyperthyroidism: A case report and literature review
- Primary orbital ganglioneuroblastoma: A case report
- Primary aortic intimal sarcoma masquerading as intramural hematoma
- Sustained false-positive results for hepatitis A virus immunoglobulin M: A case report and literature review
- Peritoneal loose body presenting as a hepatic mass: A case report and review of the literature
- Chondroblastoma of mandibular condyle: Case report and literature review
- Trauma-induced complete pacemaker lead fracture 8 months prior to hospitalization: A case report
- Primary intradural extramedullary extraosseous Ewing’s sarcoma/peripheral primitive neuroectodermal tumor (PIEES/PNET) of the thoracolumbar spine: A case report and literature review
- Computer-assisted preoperative planning of reduction of and osteosynthesis of scapular fracture: A case report
- High quality of 58-month life in lung cancer patient with brain metastases sequentially treated with gefitinib and osimertinib
- Rapid response of locally advanced oral squamous cell carcinoma to apatinib: A case report
- Retrieval of intrarenal coiled and ruptured guidewire by retrograde intrarenal surgery: A case report and literature review
- Usage of intermingled skin allografts and autografts in a senior patient with major burn injury
- Retraction
- Retraction on “Dihydromyricetin attenuates inflammation through TLR4/NF-kappa B pathway”
- Special Issue Computational Intelligence Methodologies Meets Recurrent Cancers - Part I
- An artificial immune system with bootstrap sampling for the diagnosis of recurrent endometrial cancers
- Breast cancer recurrence prediction with ensemble methods and cost-sensitive learning