TGF-β1-overexpressing mesenchymal stem cells reciprocally regulate Th17/Treg cells by regulating the expression of IFN-γ
-
Ruixue Li


Abstract
Transforming growth factor (TGF)-β1 and mesenchymal stromal cells (MSCs) are two effective immunosuppressive agents for organ transplantation technology. This study aims to explore the molecular mechanism of TGF-β1-overexpressed MSCs on T cell immunosuppression. To achieve that, BM-MSCs were isolated from canine bone marrow, and their osteogenic differentiation and surface markers were detected. The TGF-β1 gene was transferred into lentivirus and modified MSCs (TGF-β1/MSCs) by lentivirus transfection. Furthermore, TGF-β1/MSCs were co-cultured with T cells to investigate their effect on differentiation and immune regulation. Results showed that TGF-β1/MSCs significantly downregulated the proportion of CD4+ CD8+ T cells in lymphocytes and significantly upregulated the proportion of CD4+ CD25+ T cells. Moreover, TGF-β1/MSCs significantly upregulated the expression of IL-10 in CD4+ T cells and downregulated the expression of IL-17A, IL-21, and IL-22. Meanwhile, interferon-γ (IFN-γ) neutralizing antibody blocked the effects of TGF-β1/MSCs on the differentiation inhibition of Th17. Overall, our results confirm the strong immunosuppressive effect of TGF-β1/MSCs in vitro and demonstrate that IFN-γ mediates the immunosuppressive effect of TGF-β1/MSC.
1 Introduction
With the continuous development and progress of modern medicine, the transplantation of allogeneic tissues and organs has become necessary for various diseases. However, the acute or chronic rejection that often occurs after allotransplantation seriously restricts the further development and application of transplantation technology. At present, immunosuppressants are mainly used to inhibit rejection after transplantation to prolong the survival time of grafts or transplant recipients; however, recipients need to take immunosuppressant drugs for life. Moreover, it may produce various side effects and increase the risk of opportunistic infection, causing a significant burden to patients, which seriously limits the scope of its clinical application. Therefore, it is a crucial problem to be solved urgently in transplantation technology to find a method that can induce allograft immune tolerance, prolong graft survival time, have low side effects, and high safety factors.
Bone marrow mesenchymal stem cells (MSCs) belong to nonterminally differentiated cells, which have the characteristics of mesenchymal cells and the potential of stem cell self-renewal and multidirectional differentiation. Animal experiments and preliminary clinical transplantation studies have found that MSCs play an essential role in supporting stem cell implantation and regulating transplantation immunity. After the infusion of MSCs to patients receiving allogeneic skin transplantation, it was found that MSCs could significantly prolong the survival time of allogeneic skin transplantation [1]. A study on the use of mesenchymal stem cells in cartilage regeneration in osteoarthritis has shown that MSCs have special low immunogenicity and immunomodulatory ability [2]. Another study of 47 patients with intractable graft-versus-host disease showed that bone marrow MSCs isolated from third parties effectively treated intractable graft-versus-host disease [3]. The unique immunological characteristics of MSCs enable them to survive for a long time in an allogeneic or even xenogeneic body. After transplantation, MSCs still maintain the potential of multidirectional differentiation but its antigenicity is not increased, which plays a vital role in studying allogeneic or mismatched transplantation. Preliminary studies have shown that autologous or allogeneic bone marrow-derived MSC transplantation does not cause significant adverse reactions. The low immunogenicity of MSCs enables them to evade immune recognition in immune response and gain the ability to survive in the receptor. Besides, MSCs can inhibit T cell proliferation through direct cell-to-cell contact and the indirect effect of cytokines secreted by MSC; moreover, the inhibitory effect of MSCs is dose-dependent, that is, the greater the number of MSCs, the stronger the inhibitory effect [4].
Transforming growth factor β (TGF-β) is a widely used immunosuppressive agent regulating proliferation, differentiation, and biological function of many kinds of immunoreactive cells [5]. Its role in immunity has attracted wide attention, such as inducing B cells to secrete IgA [6]. Particularly, it can stimulate immature CD4+ T cells to differentiate into Foxp3 Treg and inhibit the activation and proliferation of effector T cells [7]. TGF-β leads to cell inactivation and plays a negative regulatory role, thus maintaining immune tolerance [8,9]. Johnston et al. proved that TGF-β upregulated regulatory T cells (Treg), induce immune tolerance, and prolong grafts’ survival time [10]. The activity of TGF-β1 and MSCs can regulate rejection after transplantation.
Based on the above, this study explored the molecular mechanism of TGF-β1 modification of MSCs on T cell immunosuppression. Compared with the blank control group, TGF-β1/MSCs significantly downregulated the proportion of CD4+ CD8+ T cells in lymphocytes and significantly upregulated the proportion of CD4+ CD25+ T cells in lymphocytes. In addition, we also found that TGF-β1/MSCs significantly upregulated the expression of IL-10 in CD4+ T cells and downregulated the expression of IL-17A, IL-21, and IL-22, which explained the immunosuppressive effect of TGF-β1/MSC. Meanwhile, IFN-γ neutralizing antibody blocked the effects of TGF-β1/MSCs on the differentiation inhibition of Th17. Overall, our results confirmed the strong immunosuppressive effect of TGF-β1/MSCs in vitro and found that IFN-γ mediated the immunosuppressive effect of TGF-β1/MSC.
2 Material and methods
2.1 Preparation of canine bone marrow mesenchymal stem cells and gene transduction
Under an aseptic operation, the experimental dogs were anesthetized and punctured at the left or right skeletal crest with a bone marrow puncture needle. First, 3 mL of the bone marrow fluid was extracted into a preprepared 15 mL centrifuge tube containing anticoagulants, and then 3 mL of PBS buffer containing double antibodies was diluted to make 6 mL bone marrow dilution. Clumps were removed by filtering through a 70 μm cell strainer (Thermo-Fisher Scientific). Cells were then centrifuged at 400×g for 5 min and then resuspended in the complete culture medium of stem cells. The cell suspension was inoculated in a 60 mm Petri dish and cultured at 37°C in a 5% CO2 incubator for 24 h. Lentiviral vectors for canine TGF-β1 gene overexpression were constructed by and purchased from Genscript (Nanjing, China). The cells were transduced with the recombinant virus for 12 h and then cultured in the viral vector-free medium for another 48 h. Subsequently, the MSCs transduced with TGF-β1 were exposed to 5 mg/mL puromycin for 3 days to obtain stable transduction TGF-β1/MSCs. To neutralize the IFN-γ, neutralizing antibodies (NA) for IFN-γ (10 μg/mL) were added to the co-culture; NA to IFN-γ were obtained from R&D Systems (Minneapolis, MN).
-
Ethical approval: The research related to animal use has been complied with all the relevant national regulations and institutional policies for the care and use of animals and was approved by the Animal Ethics Committee of the Second Affiliated Hospital of Kunming Medical University.
2.2 Identification of osteogenic differentiation of MSCs
When the cell density reaches 80%, 1/3rd of bone marrow MSCs were digested with trypsin. The cell concentration was adjusted to 3 × 104 cells/mL and seeded in 6-well plates. About 2 mL of growth medium (stem cell complete medium) was added to each well and cultured at 37°C in 5% CO2 incubator until the cells were covered with culture plates. After 24 h of induction and differentiation, the growth medium was carefully discarded, and 2 mL of the osteogenic induction medium was added to the experimental group. After alizarin red staining, the experimental group cells were induced to form calcium nodules as far as possible. The induced differentiation solute ion was discarded entirely, washed with PBS, and fixed with 4% formaldehyde solution (2 mL for 30 min); after fixation, it was washed twice with PBS and washed 2–3 times with 1 mL of alizarin red staining solution for 3 min. The staining results were observed under an inverted microscope and photographed.
2.3 Flow cytometry cell sorting and analysis
For cell sorting, splenocytes from 7- to 8-week-old BALB/C mice were used to purify CD4+, CD8+ or CD25+ T cells according to the manufacturer’s instructions (BD FACSAria Ⅲ, BD Biosciences, Germany). The purity of T cells was confirmed by flow cytometry. For cell surface staining, the cells were resuspended in PBS (1 × 105 cells/mL). Fluorescence-labeled monoclonal antibodies anti-CD4, anti-CD25, anti-CD34, anti-CD105, anti-CD44, anti-CD29, and anti-CD90 were, respectively, added and incubated in the refrigerator at 4°C for 30 min. All antibodies for flow cytometry were purchased from BD Biosciences. Flow cytometry analysis was performed with a FACS Aria II Cell Sorter (BD Biosciences) and data analysis was carried out using FlowJo software (TreeStar, Ashland, USA).
2.4 Western blotting
Proteins were extracted and subjected to Western blot analysis. Equal amounts of proteins (20 μg) were conducted by SDS-PAGE (12%) and transferred to the PVDF membrane (Epizyme, Shanghai, China) by semidry blotting. After blocking with 5% (w/v) non-fat milk powder, membranes were probed with the primary antibody against β-actin (1:5,000 dilution), IL-17 (1:1,000 dilution), Foxp3 (1:1,000 dilution), and RORγt (1:1,000 dilution). All antibodies were purchased from CST (Shanghai, China). The secondary antibodies were HRP conjugated goat anti-rabbit IgG (1:5,000 dilutions, Proteintech, Wuhan, China). Signals were detected by the chemiluminescence procedure (Pierce, Rockford, Illinois, USA) with BioMax films (Kodak).
2.5 Real-time quantitative PCR
RNA extraction and quantitative real-time PCR were performed for gene expression analysis. In short, according to the manufacturer’s protocols, the total RNA was extracted from the cell sample with the TRIzol reagent (Invitrogen, Carlsbad, CA, USA). The RNA concentration was measured using a spectrophotometer, and all samples were balanced by reverse transcription with a cDNA synthesis kit (Fermentas, St. Leon-Rot, Germany). Using cDNA as templates, SYBR Green I dye-labeled fluorescence quantitative detection was performed at 95°C for 10 min, followed by 40 cycles of 95°C, 20 s and 60°C, 30 s. Assays were performed in duplicate or triplicate as 20 µL reactions in 96 well plates using the 7,500 Fast Real-Time PCR System (Applied Biosystems, Foster City, CA, USA). Using the 2−ΔΔCt method, GAPDH was used as an internal reference to quantify the expression level of target genes in each experimental group. The primers used were synthesized as follows: GAPDH, 5′-AGA CAG CCG CAT CTT CTT GT-3′ (forward), GAPDH, 5′-TGA TGG CAA CAA TGT CCA CT-3′ (reverse); IL-17, 5′-CTC CAG AAG GCC CTC AGA CTAC-3′ (forward), IL-17, 5′-AGC TTT CCC TCC GCA TTG ACA CAG-3′ (reverse); T-bet, 5′-TCA CTA AGC AAG GAC GGA GAA TG-3′ (forward), T-bet, 5′-ATA AGC GGT TCC CTG GCA TAC-3′ (reverse); Foxp3, 5′-CAA GTT CCA CAA CAT GCG AC-3′ (forward), Foxp3, 5′-ATT GAG TGT CCG CTG CTT CT-3′ (reverse); IL-17F, 5′-ACC AAG GCT GCT CTG TTT CT-3′ (forward), IL-17F, 5′-GGT AAG GAG TGG CAT TTC TA-3′ (reverse); RORγt, 5′-TGG AAG TGG TGC TGG TTA GGA TG-3′ (forward), RORγt, 5′-GGA GTG GGA GAA GTC AAA GAT GGA-3′ (reverse); RORα, 5′-CTA CAT TGA CGG GCA CACC-3′ (forward), RORα, 5′-ACA CAG TTG GGG AAG TCT CG-3′ (reverse).
2.6 Quantification of cytokines
After the cells were treated according to the experimental conditions, the culture medium was collected. Then, the culture medium was centrifuged at 4°C for 10 min. The contents of IFN-γ, IL-4, IL-10, IL-21, IL-22, and IL-17A in the supernatant were detected using the ELISA kit (Dakewe, Beijing, China).
2.7 Inhibitory effects of TGF-β1/MSCs on T cells
CD4+ T and CD8+ T cells were isolated from the bone marrow MSCs. All 2.5 × 105 CD4+ T cells per well were co-cultured with MSCs (3 × 105) or TGF-β1/MSCs. The proliferation of CD4+ T cells was measured after 3, 5, or 7 days using the MTT assay [11]. The percentage of suppressive capacity was calculated by defining the absorption of the control as 100%. All CD8+ T cells were co-cultivated with IL-2 (40.0 U/mL) and co-cultured with MSCs or TGF-β1/MSCs. After 3, 5, or 7 days, an LDH assay (Beyotime Biotechnology, Nantong, China) was performed to analyze the cytotoxic lymphocyte (CTL) activity. Then, MDCK cells were added to the effector cells. All cells were cultured under 5% CO2 at 37°C for 24 h, and the cell-free supernatant was then retrieved after centrifugation at 500×g for 5 min to analyze LDH. Then, the CTL killing activity was calculated as previously described [12].
2.8 Statistical analysis
Origin 9.0 for Windows was employed in the data analysis. GraphPad Prism 5 was also adopted for preliminary data analysis. All data were expressed as mean ± SD of individual values. The statistical analysis was determined using Student’s t-test. P < 0.05 was considered statistically significant.
3 Results
3.1 Isolation and identification of BM-MSCs
The cell suspensions isolated from the bone marrow of the femur and tibia of mice were cultured. The next day, the nonadherent cells and impurities were removed by changing the culture medium. With the growth and proliferation of the cells, the adherent cells gradually became fusiform and showed colony-like growth (Figure 1a). To confirm the stemness of MSCs, the osteogenic and adipogenic differentiation potential were determined. After osteogenic induction and differentiation culture, granular substances could be observed in the cells after about 5–7 days, and their levels increased gradually. After about 3 weeks, a large number of granular substances could be observed in the cells, evenly suspended in the culture medium, and alizarin red staining showed positive results, which proved the osteogenic differentiation ability of the extracted cells. Similarly, oil red staining results also showed many lipid droplets, indicating that the isolated MSCs were functional (Figure 1b). The flow cytometry results showed that CD44, CD90, and CD29 were positively expressed, while CD105 and CD34 were negatively expressed (Figure 1c). These results were consistent with the characteristics of MSCs.
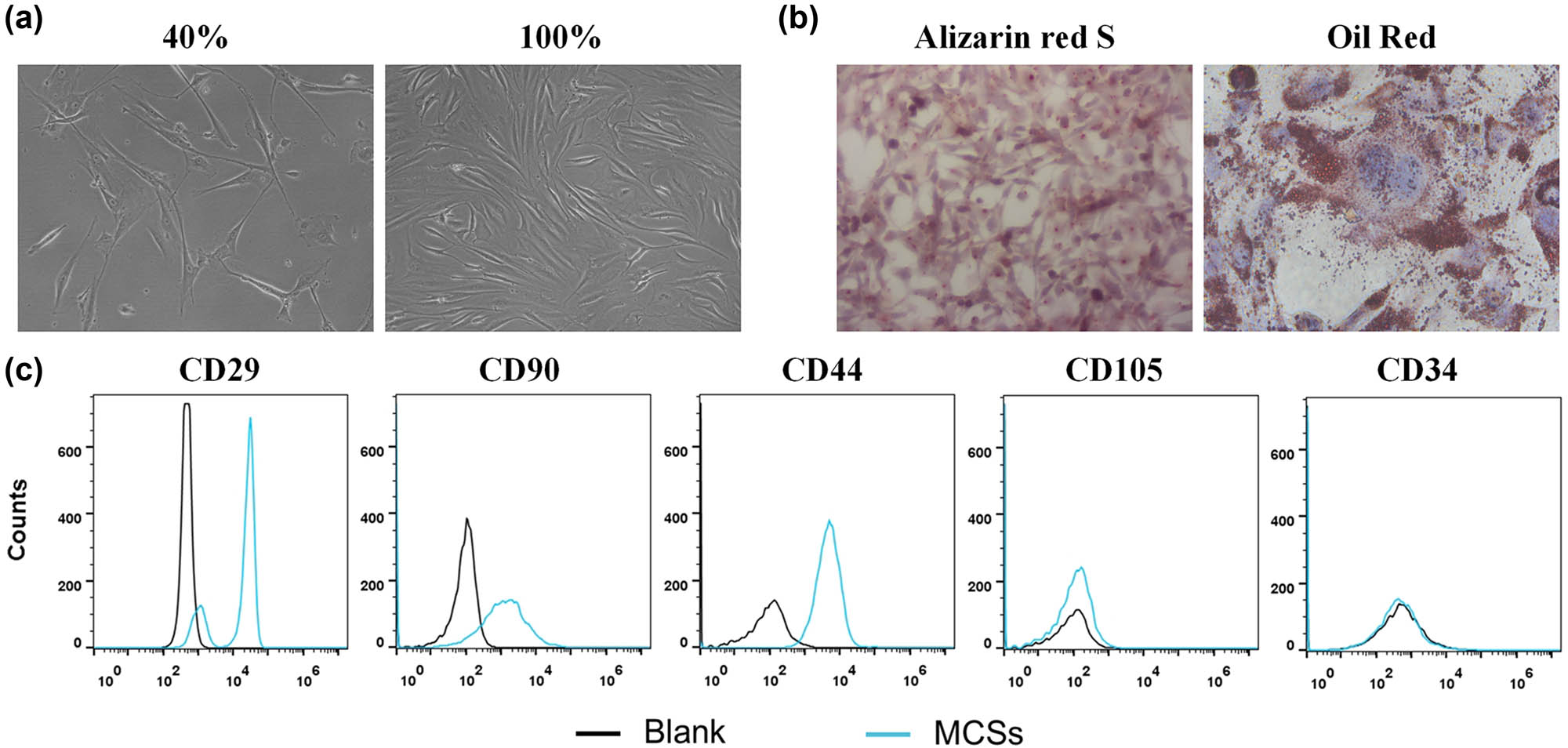
Characterization of BM-MSCs. (a) The basic morphology of BM-MSCs. (b) Identification of osteogenic differentiation ability of BM-MSCs. (c) Phenotype identification of BM-MSCs was conducted by flow cytometry.
3.2 Detection of transfection efficiency
After 96 h of TGF-β1-lentivirus transfection, massive green fluorescence (GFP expression) was observed under a fluorescence microscope in the TGF-β1/MSCs group (Figure 2a), which was also confirmed by flow cytometry (Figure 2b). We further detected the expression of TGF-β1. The results showed that the mRNA level and protein expression of TGF-β1 were significantly increased in TGF- β1/MSCs compared to MSCs.
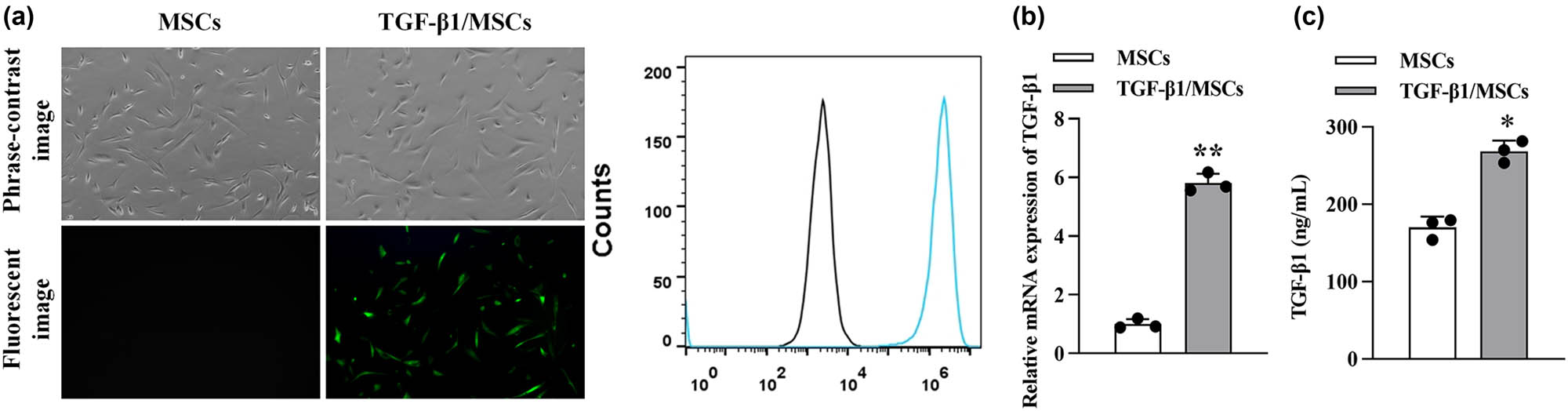
Characterization of TGF-β1/MSCs. (a) MSCs were transduced with lentivirus-GFP and lentivirus-TGF-β1. (b) The transduction efficiency was analyzed using flow cytometry. (c) The mRNA level of TGF-β1 in MSCs and TGF-β1/MSCs. (d) The concentration of TGF-β1 in the supernatants of MSCs and TGF-β1/MSCs. Data are expressed as the mean ± SD. Three independent experiments were analyzed using unpaired Student’s t-tests. *P < 0.05; **P < 0.01.
3.3 TGF-β1/MSCs inhibited T lymphocyte proliferation and the regulation of T lymphocyte immune status
T lymphocytes were isolated and purified, and the inhibition of T lymphocyte proliferation and T lymphocyte immune status regulation was observed in MSCs and TGF-β1/MSCs. The proportion of CD4+ CD25+ T cells in mixed lymphocytes was measured by flow cytometry. As shown in Figure 3a, the proportion of CD4+ CD25+ T cells increased under the stimulation of MSCs and TGF-β1/MSCs, and the TGF-β1/MSCs group increased more than the MSCs group. To evaluate the effect of TGF-β1/MSCs on allogeneic stimulation, the mixed lymphocyte reaction of CD4+ T cells and the CTL activity of CD8+ T cells were evaluated. Figure 4b shows that CD4+ T cells from the TGF-β1/MSCs group had markedly decreased proliferative functions than those from the MSCs group and control group. Further, the control group showed a significant increase in the allospecific CTL activity (Figure 4c).
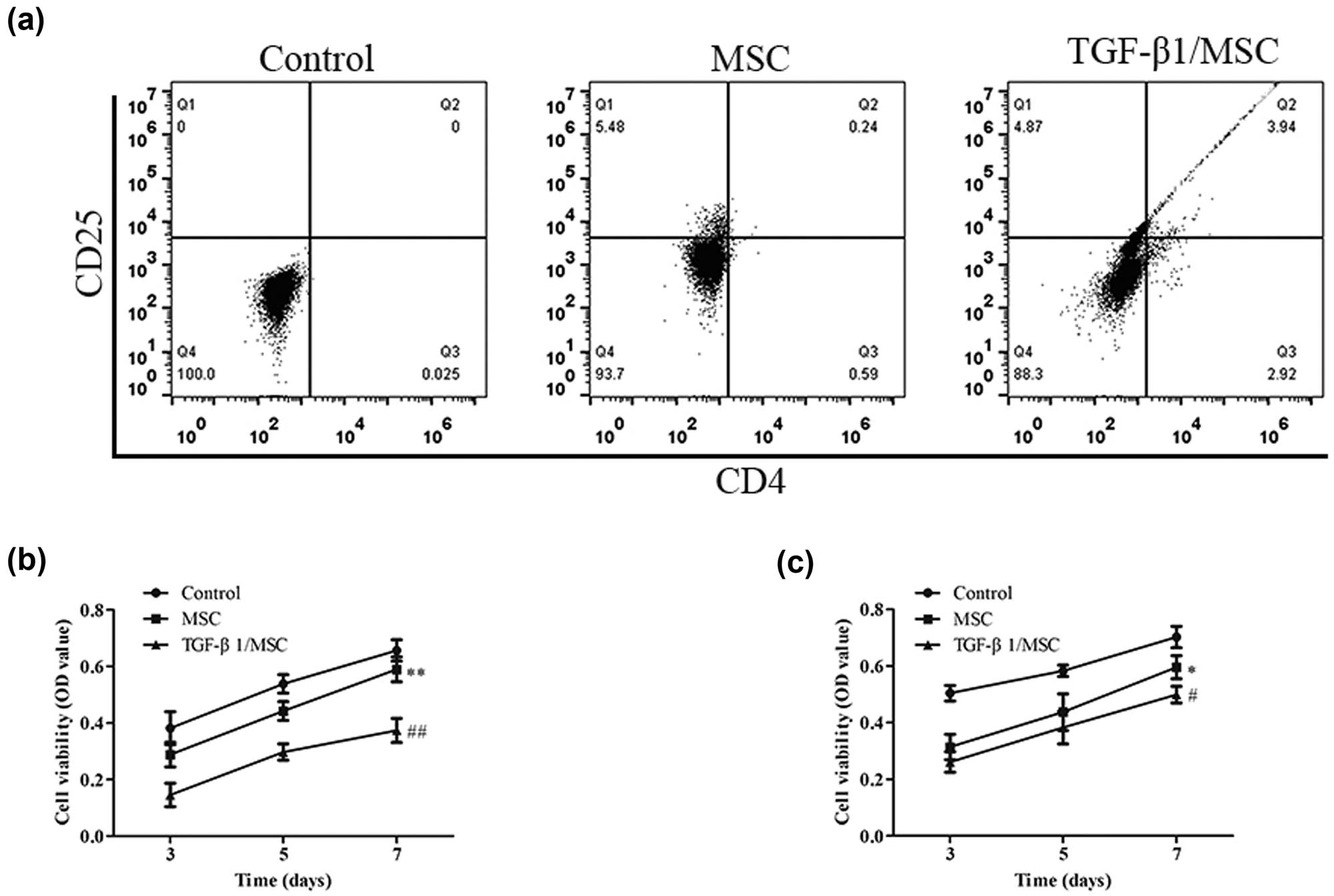
The effect of TGF-β1/MSCs on T cells. (a) T cells were co-cultured with MSCs or TGF-β1/MSCs, and the proportion of CD4+ CD25+ T cells in lymphocytes was detected by flow cytometry. (b) CD4+ T cells were co-cultured with MSCs or TGF-β1/MSCs for 3, 5, or 7 days, and proliferative responses were measured. (c) CD8+ T cells were co-cultured with MSCs or TGF-β1/MSCs in the presence of IL-2 after 3, 5, or 7 days. Live cells were collected, and the cytotoxic activity against MDCK cells was assessed. Data are expressed as the mean ± SD. Three independent experiments were analyzed using unpaired Student’s t-tests. *P < 0.05; **P < 0.01; MSCs group vs control group. # P < 0.05; ## P < 0.01; TGF-β1/MSCs group vs MSCs group.
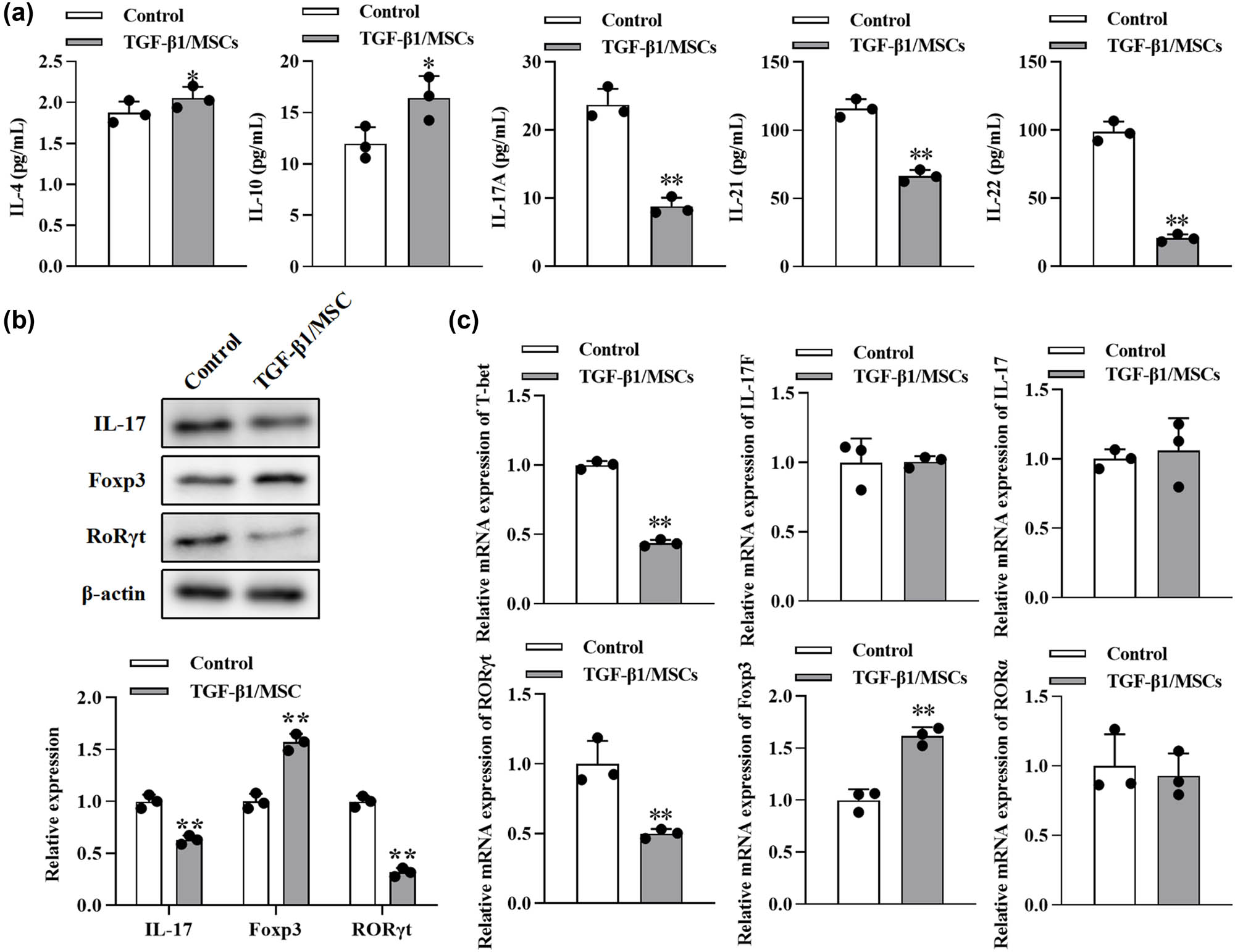
The effect of TGF-β1/MSCs on Th17 differentiation of T cells. (a) After CD4+ CD25− T cells were cultured with TGF-β1/MSCs for 3 days in the presence of IL-2, cytokines (IL-4, IL-10, IL-17A, IL-21, and IL-22) were measured by ELISA from the culture supernatant. (b) The expression of IL17, Foxp3, and RORγt in T cells was measured by Western blotting. (c) The mRNA levels of T-bet, IL-17, IL-17F, RORγt, Foxp3, and RORα were detected using RT-PCR. Data are expressed as the mean ± SD. Three independent experiments were analyzed using unpaired Student’s t-tests. *P < 0.05; **P < 0.01; TGF-β1/MSCs group vs control group.
3.4 TGF-β1/MSCs promoted the generation of Treg cells and suppressed the differentiation to Th17
We further tested the effects of TGF-β1/MSCs on the differentiation of T cells. In order to exclude the effect of TGF-β1 transfection on the proliferation of MSCs, we conducted the CCK-8 assay. The proliferation of MSCs after overexpression of TGF- β1 was not significantly different from that of the control group at 1–5 days but was inhibited at 5–7 days (Figure A1). Next, CD4+ CD25− T cells were co-cultured with TGF-β1/MSCs, and the results showed that the level of IL-10 in the culture supernatant was higher than that in the control group (Figure 4a). However, the levels of IL-17A, IL-21, and IL-22 were lower than those in the control group, and the level of IL-4 was similar to that in the control group. TGF-β1/MSCs could increase the expression of Foxp3 cells but suppressed the expression of IL17 and RORγt in CD4+ CD25− T cells (Figure 4b). Furthermore, CD4+ CD25− T cells in these co-cultures expressed a higher mRNA level of Foxp3 but lower mRNA level of T-bet and RORγt (Figure 4c). These results revealed that TGF-β1/MSCs co-cultured with CD4+ CD25− T cells promoted the generation of Treg cells and suppressed the differentiation to Th17 cells.
3.5 TGF-β1/MSCs inhibited Th17 cell differentiation via IFN-γ
Next, we further explored the intrinsic mechanisms underlying the inhibition of Th17 differentiation. A high level of IFN-γ was observed in the co-culture (Figure 5a), so we conjectured that IFN-γ might be related to the differentiation of Th17. Hence, to neutralize IFN-γ, NA for IFN-γ (10 μg/mL) was added to the co-culture. In the supernatant in the IFN-γ + group, higher levels of IL-17A and IL-22but lower levels of IL-10 were found (Figure 5b). Besides, more IL-17 and RORγt but fewer Foxp3 were detected using Western blot (Figure 5c). These results indicated that IFN-γ mediated the differentiation inhibition of Th17 cells.

TGF-β1/MSCs regulate Th17 cell differentiation through IFN-γ. (a) CD4+ T cells were co-cultured with TGF-β1/MSCs, and the concentration of IFN-γ in the supernatants was measured. *P < 0.05; **P < 0.01; TGF-β1/MSCs group vs control group. (b) Cytokines were measured by ELISA using the culture supernatant in which CD4+ T cells were co-cultured with TGF-β1/MSCs in the presence of IFN-γ NA. *P < 0.05; **P < 0.01; IFN-γ + group vs IFN-γ- group. (c) The expression of IL17, Foxp3, and RORγt in T cells was measured by Western blotting in the presence of IFN-γ NA. *P < 0.05; **P < 0.01; TGF-β1/MSCs group vs control group. Data are expressed as the mean ± SD. Three independent experiments were analyzed using unpaired Student’s t-tests.
4 Discussion
Organ transplantation is the most effective method for the treatment of patients with end-stage organ failure. However, the therapeutic effect is affected by the possible rejection after the operation and the possible complications of lifelong use of immunosuppressants. Because of their immunosuppressive effect, MSCs have excellent potential in inducing graft immune tolerance to inhibit rejection and reduce the dosage of immunosuppressants. In the study of the ischemia-reperfusion renal transplantation model in rats, it was found that MSCs could reduce the chemotaxis of antigen-presenting cells (APCs), thus effectively reducing the inflammatory reaction in the process of transplantation [13]. Thus, the role of MSCs in renal transplantation has been confirmed. Studies by Miceli et al. [14] have shown that MSCs can reduce the inflammatory response caused by ischemia during renal transplantation, which is beneficial to graft survival. Mohr et al. [15] also showed that infusion of MSCs could significantly improve the decrease of superoxide dismutase (SOD) and glutathione peroxidase (GSH-Px) activity caused by ischemia-reperfusion, which is beneficial to the improvement of acute renal failure caused by ischemia-reperfusion.
Treg was first found to inhibit immune response and induce immune tolerance in the 1970s [16]. Natural regulatory T cell (Treg) accounts for about 5–10% of CD4+ T cells, expressing both CD4 and CD25 [17], and specifically expressing Foxp3 [17]. Treg can inhibit the activation and proliferation of CD4+ and CD8+ T lymphocytes, which is related to the direct contact between cells and paracrine cytokines IL-10 and TGF-β [18]. Interleukin-2 (IL-2) and other immunoregulatory molecules have been shown to control the generation of Tregs [19]. It can also indirectly inhibit the immune response by inhibiting the activity of APC while maintaining the microenvironment of immune tolerance. The most significant advantage of Treg in transplantation immunity is that it can actively transplant tolerance, that is, the Treg of the recipient who had reached immune tolerance to organ transplantation can be transferred to other recipients, and the recipient can also achieve the effect of immune tolerance. Sayitoglu et al. [20] confirmed this in an experiment on a mouse skin graft model and found that rejection would occur again after the removal of Treg. Studies of Park [19] and Tahvildari [21] also confirmed that transplantation of Treg could induce immune tolerance to MHC-mismatched heart or skin grafts. Thomann et al. [22] used donor-derived Treg in the islet transplantation model, which also significantly prolonged the survival time of the graft.
TGF-β is a crucial regulator of the immune response, regulating the occurrence and termination of immune response by regulating lymphocyte differentiation and apoptosis. A large number of studies have revealed the mechanism of this regulation. TGF-β can inhibit T lymphocyte proliferation mediated by IL-2 through the Smad3 pathway and directly inhibit T lymphocyte proliferation by downregulating cyclin D2, cyclin E, and c-myc [23,24]; upregulate the expression of FoxP3 and induce the differentiation of Treg. Given the immunosuppressive effect of TGF-β, its application in transplantation immunity has been widely studied. The study of mouse liver transplantation also confirmed a close relationship between TGF-β and liver regeneration [25]. In renal transplantation, TGF-β can significantly inhibit the inflammatory response in the early stage of acute inhibition of rejection, thus achieving the inhibition of chronic rejection [26]. Also, increasing the expression of islet B cell TGF-β in the type 1 diabetic mouse model is helpful to improve the condition of diabetes, which may be related to the increase of T lymphocyte apoptosis [27].
As a well-known immune enhancement factor, IFN-γ is the first discovered cytokine, a necessary condition for stimulating Th1 differentiation [28,29]. At the same time, it can also upregulate the expression of cellular MHC-I molecules, thus improving the ability of immune surveillance in vivo [30]. It can also mediate the activation of macrophages, enhance the lethality of macrophages and NK cells, promote the proliferation of B lymphocytes and the production of antibodies, and upregulate chemokines to promote the chemotaxis of inflammatory cells. According to the report, a large amount of IFN-γ can be detected in the supernatant of the co-culture system of CD4+ T cells and MSCs. Liu et al. believe that this phenomenon is caused by the high expression of TGF-β1 in MSCs [31]. In this study, the IFN-γ-neutralizing antibody significantly blocked the effects of MSCs on T cells, indicating that IFN-γ mediated the differentiation inhibition of Th17.
5 Conclusion
The present study demonstrated that TGF-β1/MSCs inhibited T lymphocyte proliferation and T lymphocyte immune status regulation. Moreover, TGF-β1/MSCs promoted the generation of Treg cells and suppressed the differentiation of Th17 cells. Besides, IFN-γ mediated the differentiation inhibition of Th17.
-
Funding information: This work was supported by the National Natural Science Foundation of China (81460132) and Yunnan Provincial Science and Technology Department – Kunming Medical University Joint Project (2018FE001).
-
Author contributions: Ruixue Li and Hong Zhu drafted the manuscript, conceived and designed the study, and revised the manuscript for important intellectual content. Renyong Wang, Shijie Zhong, and Farhan Asghar performed the acquisition of data and conducted the statistical analysis. Tiehan Li and Lei Zhu performed the analysis and interpretation of data. Ruixue Li, Renyong Wang, and Hong Zhu obtained funding. All authors read and approved the final manuscript.
-
Conflict of interest: The authors state no conflict of interest.
-
Data availability statement: The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
Appendix

The effects of overexpressing TGFβ1 on the proliferation of MSCs. The growth curves of the cells after transduction for 7 days were evaluated by the CCK-8 assay. *P < 0.05, **P < 0.01; TGF-β1/MSCs group vs MSCs group.
References
[1] Kim YJ, Shin S, Han DJ, Kim YH, Lee JY, Yoon YH, et al. Long-term effects of pancreas transplantation on diabetic retinopathy and incidence and predictive risk factors for early worsening. Transplantation. 2018;102(1):e30–8.10.1097/TP.0000000000001958Search in Google Scholar PubMed
[2] Kristjánsson B, Honsawek S. Mesenchymal stem cells for cartilage regeneration in osteoarthritis. World J Orthopedics. 2017;8(9):674–80.10.5312/wjo.v8.i9.674Search in Google Scholar PubMed PubMed Central
[3] Zhao K, Lou R, Huang F, Peng Y, Jiang Z, Huang K, et al. Immunomodulation effects of mesenchymal stromal cells on acute graft-versus-host disease after hematopoietic stem cell transplantation. Biol Blood Marrow Transplant. 2015;21(1):97–104.10.1016/j.bbmt.2014.09.030Search in Google Scholar PubMed
[4] Fan XL, Zeng QX, Li X, Li CL, Xu ZB, Deng XQ, et al. Induced pluripotent stem cell-derived mesenchymal stem cells activate quiescent T cells and elevate regulatory T cell response via NF-κB in allergic rhinitis patients. Stem Cell Res Ther. 2018;9(1):1–15.10.1186/s13287-018-0896-zSearch in Google Scholar PubMed PubMed Central
[5] Istomine R, Alvarez F, Almadani Y, Philip A, Piccirillo CA. The deubiquitinating enzyme ubiquitin-specific peptidase 11 potentiates TGF-β signaling in CD4+ T cells to facilitate Foxp3 + regulatory T and TH17 cell differentiation. J Immunology. 2019;203(9):2388–400.10.4049/jimmunol.1801689Search in Google Scholar PubMed
[6] Weissler KA, Frischmeyer-Guerrerio PA. Genetic evidence for the role of transforming growth factor-β in atopic phenotypes. Curr Opt Immunol. 2019;60:54–62.10.1016/j.coi.2019.05.002Search in Google Scholar PubMed PubMed Central
[7] Zhang XX, Qiao YC, Li W, Zou X, Chen YL, Shen J, et al. Human amylin induces CD4+ Foxp3 + regulatory T cells in the protection from autoimmune diabetes. Immunologic Res. 2018;66(1):179–86.10.1007/s12026-017-8956-5Search in Google Scholar PubMed
[8] Huai G, Markmann JF, Deng S, Rickert CG. TGF‐β‐secreting regulatory B cells: unsung players in immune regulation. Clin Transl Immunol. 2021;10(4):e1270.10.1002/cti2.1270Search in Google Scholar PubMed PubMed Central
[9] Guzman-Genuino RM, Diener KR. Regulatory B cells in pregnancy: lessons from autoimmunity, graft tolerance, and cancer. Front Immunol. 2017;8:172.10.3389/fimmu.2017.00172Search in Google Scholar PubMed PubMed Central
[10] Johnston CJ, Smyth DJ, Dresser DW, Maizels RM. TGF-β in tolerance, development and regulation of immunity. Cell Immunol. 2016;299:14–22.10.1016/j.cellimm.2015.10.006Search in Google Scholar PubMed PubMed Central
[11] Kuca-Warnawin E, Janicka I, Szczęsny P, Olesińska M, Bonek K, Głuszko P, et al. Modulation of T-Cell activation markers expression by the adipose tissue–derived mesenchymal stem cells of patients with rheumatic diseases. Cell Transplant. 2020;29:0963689720945682.10.1177/0963689720945682Search in Google Scholar PubMed PubMed Central
[12] Niu J, Wang Y, Liu B, Yao Y. Mesenchymal stem cells prolong the survival of orthotopic liver transplants by regulating the expression of TGF-β1. Turkish J Gastroenterol. 2018;29(5):601–9.10.5152/tjg.2018.17395Search in Google Scholar PubMed PubMed Central
[13] Bai L, Shao H, Wang H, Zhang Z, Su C, Dong L, et al. Effects of mesenchymal stem cell-derived exosomes on experimental autoimmune uveitis. Sci Rep. 2017;7(1):1–11.10.1038/s41598-017-04559-ySearch in Google Scholar PubMed PubMed Central
[14] Miceli V, Bertani A, Chinnici CM, Bulati M, Pampalone M, Amico G, et al. Conditioned medium from human amnion-derived mesenchymal stromal/stem cells attenuating the effects of cold ischemia-reperfusion injury in an in vitro model using human alveolar epithelial cells. Int J Mol Sci. 2021;22(2):510.10.3390/ijms22020510Search in Google Scholar PubMed PubMed Central
[15] Mohr A, Zwacka R. The future of mesenchymal stem cell-based therapeutic approaches for cancer–from cells to ghosts. Cancer Lett. 2018;414:239–49.10.1016/j.canlet.2017.11.025Search in Google Scholar PubMed
[16] Lin S, Wu H, Wang C, Xiao Z, Xu F. Regulatory T cells and acute lung injury: cytokines, uncontrolled inflammation, and therapeutic implications. Front Immunol. 2018;9:1545.10.3389/fimmu.2018.01545Search in Google Scholar PubMed PubMed Central
[17] Ferrandino F, Grazioli P, Bellavia D, Campese AF, Screpanti I, Felli MP. Notch and NF-κB: coach and players of regulatory T-cell response in cancer. Front Immunol. 2018;9:2165.10.3389/fimmu.2018.02165Search in Google Scholar PubMed PubMed Central
[18] Apert C, Romagnoli P, van Meerwijk JPM. IL-2 and IL-15 dependent thymic development of Foxp3-expressing regulatory T lymphocytes. Protein Cell. 2018;9(4):322–32.10.1007/s13238-017-0425-3Search in Google Scholar PubMed PubMed Central
[19] Park MJ, Baek JA, Kim SY, Jung KA, Choi JW, Park SH, et al. Myeloid-derived suppressor cells therapy enhance immunoregulatory properties in acute graft versus host disease with combination of regulatory T cells. J Transl Med. 2020;18(1):1–14.10.1186/s12967-020-02657-6Search in Google Scholar PubMed PubMed Central
[20] Sayitoglu EC, Freeborn RA, Roncarolo MG. The Yin and Yang of type 1 regulatory T cells: from discovery to clinical application. Front Immunol. 2021;12:2199.10.3389/fimmu.2021.693105Search in Google Scholar PubMed PubMed Central
[21] Tahvildari M, Inomata T, Amouzegar A, Dana R. Regulatory T cell modulation of cytokine and cellular networks in corneal graft rejection. Curr Ophthalmol Rep. 2018;6(4):266–74.10.1007/s40135-018-0191-2Search in Google Scholar
[22] Thomann AS, Schneider T, Cyran L, Eckert IN, Kerstan A, Lutz MB. Conversion of anergic T cells into Foxp3-IL-10 + regulatory T cells by a second antigen stimulus in vivo. Front Immunol. 2021;12:2598.10.3389/fimmu.2021.704578Search in Google Scholar PubMed PubMed Central
[23] Geisinger S. Transforming growth factor beta suppression of CD8 + T Cell proliferation. Loyola University Chicago: ProQuest Dissertations Publishing; 2020.Search in Google Scholar
[24] Takeuchi H, Konnai S, Maekawa N, Takagi S, Ohta H, Sasaki N, et al. Canine transforming growth factor-β receptor 2-Ig: a potential candidate biologic for melanoma treatment that reverses transforming growth factor-β1 immunosuppression. Front Veterinary Sci. 2021;8:656715.10.3389/fvets.2021.656715Search in Google Scholar PubMed PubMed Central
[25] Yin L, Guo X, Zhang C, Cai Z, Xu C. In silico analysis of expression data during the early priming stage of liver regeneration after partial hepatectomy in rat. Oncotarget. 2018;9(14):11794–804.10.18632/oncotarget.24370Search in Google Scholar PubMed PubMed Central
[26] Park MS, Kim SK, Park HJ, Seok H, Kang SW, Lee SH, et al. Association studies of bone morphogenetic protein 2 gene polymorphisms with acute rejection in kidney transplantation recipients. Transplant Proc. 2017;49(5):1012–7.10.1016/j.transproceed.2017.03.019Search in Google Scholar PubMed
[27] Miani M, Le Naour J, Waeckel-Enée E, Verma SC, Straube M, Emond P, et al. Gut microbiota-stimulated innate lymphoid cells support β-defensin 14 expression in pancreatic endocrine cells, preventing autoimmune diabetes. Cell Metab. 2018;28(4):557–72.e6.10.1016/j.cmet.2018.06.012Search in Google Scholar PubMed
[28] Chatterjee S, Chatterjee A, Jana S, Dey S, Roy H, Das MK, et al. Transforming growth factor beta orchestrates PD-L1 enrichment in tumor-derived exosomes and mediates CD8 T-cell dysfunction regulating early phosphorylation of TCR signalome in breast cancer. Carcinogenesis. 2021;42(1):38–47.10.1093/carcin/bgaa092Search in Google Scholar PubMed
[29] Okada K, Sadahiro S, Chan LF, Ogimi T, Miyakita H, Saito G, et al. The number of natural killer cells in the largest diameter lymph nodes is associated with the number of retrieved lymph nodes and lymph node size, and is an independent prognostic factor in patients with stage II colon cancer. Oncology. 2018;95(5):288–96.10.1159/000491019Search in Google Scholar PubMed PubMed Central
[30] Qin XY, Zhang YL, Chi YF, Yan B, Zeng XJ, Li HH, et al. Angiotensin II regulates Th1 T cell differentiation through angiotensin II type 1 receptor-PKA-mediated activation of proteasome. Cell Physiol Biochem. 2018;45(4):1366–76.10.1159/000487562Search in Google Scholar PubMed
[31] Liu X, Ren S, Qu X, Ge C, Cheng K, Zhao RC. Mesenchymal stem cells inhibit Th17 cells differentiation via IFN-γ-mediated SOCS3 activation. Immunologic Res. 2015;61(3):219–29.10.1007/s12026-014-8612-2Search in Google Scholar PubMed
© 2021 Ruixue Li et al., published by De Gruyter
This work is licensed under the Creative Commons Attribution 4.0 International License.
Articles in the same Issue
- Biomedical Sciences
- Research progress on the mechanism of orexin in pain regulation in different brain regions
- Adriamycin-resistant cells are significantly less fit than adriamycin-sensitive cells in cervical cancer
- Exogenous spermidine affects polyamine metabolism in the mouse hypothalamus
- Iris metastasis of diffuse large B-cell lymphoma misdiagnosed as primary angle-closure glaucoma: A case report and review of the literature
- LncRNA PVT1 promotes cervical cancer progression by sponging miR-503 to upregulate ARL2 expression
- Two new inflammatory markers related to the CURB-65 score for disease severity in patients with community-acquired pneumonia: The hypersensitive C-reactive protein to albumin ratio and fibrinogen to albumin ratio
- Circ_0091579 enhances the malignancy of hepatocellular carcinoma via miR-1287/PDK2 axis
- Silencing XIST mitigated lipopolysaccharide (LPS)-induced inflammatory injury in human lung fibroblast WI-38 cells through modulating miR-30b-5p/CCL16 axis and TLR4/NF-κB signaling pathway
- Protocatechuic acid attenuates cerebral aneurysm formation and progression by inhibiting TNF-alpha/Nrf-2/NF-kB-mediated inflammatory mechanisms in experimental rats
- ABCB1 polymorphism in clopidogrel-treated Montenegrin patients
- Metabolic profiling of fatty acids in Tripterygium wilfordii multiglucoside- and triptolide-induced liver-injured rats
- miR-338-3p inhibits cell growth, invasion, and EMT process in neuroblastoma through targeting MMP-2
- Verification of neuroprotective effects of alpha-lipoic acid on chronic neuropathic pain in a chronic constriction injury rat model
- Circ_WWC3 overexpression decelerates the progression of osteosarcoma by regulating miR-421/PDE7B axis
- Knockdown of TUG1 rescues cardiomyocyte hypertrophy through targeting the miR-497/MEF2C axis
- MiR-146b-3p protects against AR42J cell injury in cerulein-induced acute pancreatitis model through targeting Anxa2
- miR-299-3p suppresses cell progression and induces apoptosis by downregulating PAX3 in gastric cancer
- Diabetes and COVID-19
- Discovery of novel potential KIT inhibitors for the treatment of gastrointestinal stromal tumor
- TEAD4 is a novel independent predictor of prognosis in LGG patients with IDH mutation
- circTLK1 facilitates the proliferation and metastasis of renal cell carcinoma by regulating miR-495-3p/CBL axis
- microRNA-9-5p protects liver sinusoidal endothelial cell against oxygen glucose deprivation/reperfusion injury
- Long noncoding RNA TUG1 regulates degradation of chondrocyte extracellular matrix via miR-320c/MMP-13 axis in osteoarthritis
- Duodenal adenocarcinoma with skin metastasis as initial manifestation: A case report
- Effects of Loofah cylindrica extract on learning and memory ability, brain tissue morphology, and immune function of aging mice
- Recombinant Bacteroides fragilis enterotoxin-1 (rBFT-1) promotes proliferation of colorectal cancer via CCL3-related molecular pathways
- Blocking circ_UBR4 suppressed proliferation, migration, and cell cycle progression of human vascular smooth muscle cells in atherosclerosis
- Gene therapy in PIDs, hemoglobin, ocular, neurodegenerative, and hemophilia B disorders
- Downregulation of circ_0037655 impedes glioma formation and metastasis via the regulation of miR-1229-3p/ITGB8 axis
- Vitamin D deficiency and cardiovascular risk in type 2 diabetes population
- Circ_0013359 facilitates the tumorigenicity of melanoma by regulating miR-136-5p/RAB9A axis
- Mechanisms of circular RNA circ_0066147 on pancreatic cancer progression
- lncRNA myocardial infarction-associated transcript (MIAT) knockdown alleviates LPS-induced chondrocytes inflammatory injury via regulating miR-488-3p/sex determining region Y-related HMG-box 11 (SOX11) axis
- Identification of circRNA circ-CSPP1 as a potent driver of colorectal cancer by directly targeting the miR-431/LASP1 axis
- Hyperhomocysteinemia exacerbates ischemia-reperfusion injury-induced acute kidney injury by mediating oxidative stress, DNA damage, JNK pathway, and apoptosis
- Potential prognostic markers and significant lncRNA–mRNA co-expression pairs in laryngeal squamous cell carcinoma
- Gamma irradiation-mediated inactivation of enveloped viruses with conservation of genome integrity: Potential application for SARS-CoV-2 inactivated vaccine development
- ADHFE1 is a correlative factor of patient survival in cancer
- The association of transcription factor Prox1 with the proliferation, migration, and invasion of lung cancer
- Is there a relationship between the prevalence of autoimmune thyroid disease and diabetic kidney disease?
- Immunoregulatory function of Dictyophora echinovolvata spore polysaccharides in immunocompromised mice induced by cyclophosphamide
- T cell epitopes of SARS-CoV-2 spike protein and conserved surface protein of Plasmodium malariae share sequence homology
- Anti-obesity effect and mechanism of mesenchymal stem cells influence on obese mice
- Long noncoding RNA HULC contributes to paclitaxel resistance in ovarian cancer via miR-137/ITGB8 axis
- Glucocorticoids protect HEI-OC1 cells from tunicamycin-induced cell damage via inhibiting endoplasmic reticulum stress
- Prognostic value of the neutrophil-to-lymphocyte ratio in acute organophosphorus pesticide poisoning
- Gastroprotective effects of diosgenin against HCl/ethanol-induced gastric mucosal injury through suppression of NF-κβ and myeloperoxidase activities
- Silencing of LINC00707 suppresses cell proliferation, migration, and invasion of osteosarcoma cells by modulating miR-338-3p/AHSA1 axis
- Successful extracorporeal membrane oxygenation resuscitation of patient with cardiogenic shock induced by phaeochromocytoma crisis mimicking hyperthyroidism: A case report
- Effects of miR-185-5p on replication of hepatitis C virus
- Lidocaine has antitumor effect on hepatocellular carcinoma via the circ_DYNC1H1/miR-520a-3p/USP14 axis
- Primary localized cutaneous nodular amyloidosis presenting as lymphatic malformation: A case report
- Multimodal magnetic resonance imaging analysis in the characteristics of Wilson’s disease: A case report and literature review
- Therapeutic potential of anticoagulant therapy in association with cytokine storm inhibition in severe cases of COVID-19: A case report
- Neoadjuvant immunotherapy combined with chemotherapy for locally advanced squamous cell lung carcinoma: A case report and literature review
- Rufinamide (RUF) suppresses inflammation and maintains the integrity of the blood–brain barrier during kainic acid-induced brain damage
- Inhibition of ADAM10 ameliorates doxorubicin-induced cardiac remodeling by suppressing N-cadherin cleavage
- Invasive ductal carcinoma and small lymphocytic lymphoma/chronic lymphocytic leukemia manifesting as a collision breast tumor: A case report and literature review
- Clonal diversity of the B cell receptor repertoire in patients with coronary in-stent restenosis and type 2 diabetes
- CTLA-4 promotes lymphoma progression through tumor stem cell enrichment and immunosuppression
- WDR74 promotes proliferation and metastasis in colorectal cancer cells through regulating the Wnt/β-catenin signaling pathway
- Down-regulation of IGHG1 enhances Protoporphyrin IX accumulation and inhibits hemin biosynthesis in colorectal cancer by suppressing the MEK-FECH axis
- Curcumin suppresses the progression of gastric cancer by regulating circ_0056618/miR-194-5p axis
- Scutellarin-induced A549 cell apoptosis depends on activation of the transforming growth factor-β1/smad2/ROS/caspase-3 pathway
- lncRNA NEAT1 regulates CYP1A2 and influences steroid-induced necrosis
- A two-microRNA signature predicts the progression of male thyroid cancer
- Isolation of microglia from retinas of chronic ocular hypertensive rats
- Changes of immune cells in patients with hepatocellular carcinoma treated by radiofrequency ablation and hepatectomy, a pilot study
- Calcineurin Aβ gene knockdown inhibits transient outward potassium current ion channel remodeling in hypertrophic ventricular myocyte
- Aberrant expression of PI3K/AKT signaling is involved in apoptosis resistance of hepatocellular carcinoma
- Clinical significance of activated Wnt/β-catenin signaling in apoptosis inhibition of oral cancer
- circ_CHFR regulates ox-LDL-mediated cell proliferation, apoptosis, and EndoMT by miR-15a-5p/EGFR axis in human brain microvessel endothelial cells
- Resveratrol pretreatment mitigates LPS-induced acute lung injury by regulating conventional dendritic cells’ maturation and function
- Ubiquitin-conjugating enzyme E2T promotes tumor stem cell characteristics and migration of cervical cancer cells by regulating the GRP78/FAK pathway
- Carriage of HLA-DRB1*11 and 1*12 alleles and risk factors in patients with breast cancer in Burkina Faso
- Protective effect of Lactobacillus-containing probiotics on intestinal mucosa of rats experiencing traumatic hemorrhagic shock
- Glucocorticoids induce osteonecrosis of the femoral head through the Hippo signaling pathway
- Endothelial cell-derived SSAO can increase MLC20 phosphorylation in VSMCs
- Downregulation of STOX1 is a novel prognostic biomarker for glioma patients
- miR-378a-3p regulates glioma cell chemosensitivity to cisplatin through IGF1R
- The molecular mechanisms underlying arecoline-induced cardiac fibrosis in rats
- TGF-β1-overexpressing mesenchymal stem cells reciprocally regulate Th17/Treg cells by regulating the expression of IFN-γ
- The influence of MTHFR genetic polymorphisms on methotrexate therapy in pediatric acute lymphoblastic leukemia
- Red blood cell distribution width-standard deviation but not red blood cell distribution width-coefficient of variation as a potential index for the diagnosis of iron-deficiency anemia in mid-pregnancy women
- Small cell neuroendocrine carcinoma expressing alpha fetoprotein in the endometrium
- Superoxide dismutase and the sigma1 receptor as key elements of the antioxidant system in human gastrointestinal tract cancers
- Molecular characterization and phylogenetic studies of Echinococcus granulosus and Taenia multiceps coenurus cysts in slaughtered sheep in Saudi Arabia
- ITGB5 mutation discovered in a Chinese family with blepharophimosis-ptosis-epicanthus inversus syndrome
- ACTB and GAPDH appear at multiple SDS-PAGE positions, thus not suitable as reference genes for determining protein loading in techniques like Western blotting
- Facilitation of mouse skin-derived precursor growth and yield by optimizing plating density
- 3,4-Dihydroxyphenylethanol ameliorates lipopolysaccharide-induced septic cardiac injury in a murine model
- Downregulation of PITX2 inhibits the proliferation and migration of liver cancer cells and induces cell apoptosis
- Expression of CDK9 in endometrial cancer tissues and its effect on the proliferation of HEC-1B
- Novel predictor of the occurrence of DKA in T1DM patients without infection: A combination of neutrophil/lymphocyte ratio and white blood cells
- Investigation of molecular regulation mechanism under the pathophysiology of subarachnoid hemorrhage
- miR-25-3p protects renal tubular epithelial cells from apoptosis induced by renal IRI by targeting DKK3
- Bioengineering and Biotechnology
- Green fabrication of Co and Co3O4 nanoparticles and their biomedical applications: A review
- Agriculture
- Effects of inorganic and organic selenium sources on the growth performance of broilers in China: A meta-analysis
- Crop-livestock integration practices, knowledge, and attitudes among smallholder farmers: Hedging against climate change-induced shocks in semi-arid Zimbabwe
- Food Science and Nutrition
- Effect of food processing on the antioxidant activity of flavones from Polygonatum odoratum (Mill.) Druce
- Vitamin D and iodine status was associated with the risk and complication of type 2 diabetes mellitus in China
- Diversity of microbiota in Slovak summer ewes’ cheese “Bryndza”
- Comparison between voltammetric detection methods for abalone-flavoring liquid
- Composition of low-molecular-weight glutenin subunits in common wheat (Triticum aestivum L.) and their effects on the rheological properties of dough
- Application of culture, PCR, and PacBio sequencing for determination of microbial composition of milk from subclinical mastitis dairy cows of smallholder farms
- Investigating microplastics and potentially toxic elements contamination in canned Tuna, Salmon, and Sardine fishes from Taif markets, KSA
- From bench to bar side: Evaluating the red wine storage lesion
- Establishment of an iodine model for prevention of iodine-excess-induced thyroid dysfunction in pregnant women
- Plant Sciences
- Characterization of GMPP from Dendrobium huoshanense yielding GDP-D-mannose
- Comparative analysis of the SPL gene family in five Rosaceae species: Fragaria vesca, Malus domestica, Prunus persica, Rubus occidentalis, and Pyrus pyrifolia
- Identification of leaf rust resistance genes Lr34 and Lr46 in common wheat (Triticum aestivum L. ssp. aestivum) lines of different origin using multiplex PCR
- Investigation of bioactivities of Taxus chinensis, Taxus cuspidata, and Taxus × media by gas chromatography-mass spectrometry
- Morphological structures and histochemistry of roots and shoots in Myricaria laxiflora (Tamaricaceae)
- Transcriptome analysis of resistance mechanism to potato wart disease
- In silico analysis of glycosyltransferase 2 family genes in duckweed (Spirodela polyrhiza) and its role in salt stress tolerance
- Comparative study on growth traits and ions regulation of zoysiagrasses under varied salinity treatments
- Role of MS1 homolog Ntms1 gene of tobacco infertility
- Biological characteristics and fungicide sensitivity of Pyricularia variabilis
- In silico/computational analysis of mevalonate pyrophosphate decarboxylase gene families in Campanulids
- Identification of novel drought-responsive miRNA regulatory network of drought stress response in common vetch (Vicia sativa)
- How photoautotrophy, photomixotrophy, and ventilation affect the stomata and fluorescence emission of pistachios rootstock?
- Apoplastic histochemical features of plant root walls that may facilitate ion uptake and retention
- Ecology and Environmental Sciences
- The impact of sewage sludge on the fungal communities in the rhizosphere and roots of barley and on barley yield
- Domestication of wild animals may provide a springboard for rapid variation of coronavirus
- Response of benthic invertebrate assemblages to seasonal and habitat condition in the Wewe River, Ashanti region (Ghana)
- Molecular record for the first authentication of Isaria cicadae from Vietnam
- Twig biomass allocation of Betula platyphylla in different habitats in Wudalianchi Volcano, northeast China
- Animal Sciences
- Supplementation of probiotics in water beneficial growth performance, carcass traits, immune function, and antioxidant capacity in broiler chickens
- Predators of the giant pine scale, Marchalina hellenica (Gennadius 1883; Hemiptera: Marchalinidae), out of its natural range in Turkey
- Honey in wound healing: An updated review
- NONMMUT140591.1 may serve as a ceRNA to regulate Gata5 in UT-B knockout-induced cardiac conduction block
- Radiotherapy for the treatment of pulmonary hydatidosis in sheep
- Retraction
- Retraction of “Long non-coding RNA TUG1 knockdown hinders the tumorigenesis of multiple myeloma by regulating microRNA-34a-5p/NOTCH1 signaling pathway”
- Special Issue on Reuse of Agro-Industrial By-Products
- An effect of positional isomerism of benzoic acid derivatives on antibacterial activity against Escherichia coli
- Special Issue on Computing and Artificial Techniques for Life Science Applications - Part II
- Relationship of Gensini score with retinal vessel diameter and arteriovenous ratio in senile CHD
- Effects of different enantiomers of amlodipine on lipid profiles and vasomotor factors in atherosclerotic rabbits
- Establishment of the New Zealand white rabbit animal model of fatty keratopathy associated with corneal neovascularization
- lncRNA MALAT1/miR-143 axis is a potential biomarker for in-stent restenosis and is involved in the multiplication of vascular smooth muscle cells
Articles in the same Issue
- Biomedical Sciences
- Research progress on the mechanism of orexin in pain regulation in different brain regions
- Adriamycin-resistant cells are significantly less fit than adriamycin-sensitive cells in cervical cancer
- Exogenous spermidine affects polyamine metabolism in the mouse hypothalamus
- Iris metastasis of diffuse large B-cell lymphoma misdiagnosed as primary angle-closure glaucoma: A case report and review of the literature
- LncRNA PVT1 promotes cervical cancer progression by sponging miR-503 to upregulate ARL2 expression
- Two new inflammatory markers related to the CURB-65 score for disease severity in patients with community-acquired pneumonia: The hypersensitive C-reactive protein to albumin ratio and fibrinogen to albumin ratio
- Circ_0091579 enhances the malignancy of hepatocellular carcinoma via miR-1287/PDK2 axis
- Silencing XIST mitigated lipopolysaccharide (LPS)-induced inflammatory injury in human lung fibroblast WI-38 cells through modulating miR-30b-5p/CCL16 axis and TLR4/NF-κB signaling pathway
- Protocatechuic acid attenuates cerebral aneurysm formation and progression by inhibiting TNF-alpha/Nrf-2/NF-kB-mediated inflammatory mechanisms in experimental rats
- ABCB1 polymorphism in clopidogrel-treated Montenegrin patients
- Metabolic profiling of fatty acids in Tripterygium wilfordii multiglucoside- and triptolide-induced liver-injured rats
- miR-338-3p inhibits cell growth, invasion, and EMT process in neuroblastoma through targeting MMP-2
- Verification of neuroprotective effects of alpha-lipoic acid on chronic neuropathic pain in a chronic constriction injury rat model
- Circ_WWC3 overexpression decelerates the progression of osteosarcoma by regulating miR-421/PDE7B axis
- Knockdown of TUG1 rescues cardiomyocyte hypertrophy through targeting the miR-497/MEF2C axis
- MiR-146b-3p protects against AR42J cell injury in cerulein-induced acute pancreatitis model through targeting Anxa2
- miR-299-3p suppresses cell progression and induces apoptosis by downregulating PAX3 in gastric cancer
- Diabetes and COVID-19
- Discovery of novel potential KIT inhibitors for the treatment of gastrointestinal stromal tumor
- TEAD4 is a novel independent predictor of prognosis in LGG patients with IDH mutation
- circTLK1 facilitates the proliferation and metastasis of renal cell carcinoma by regulating miR-495-3p/CBL axis
- microRNA-9-5p protects liver sinusoidal endothelial cell against oxygen glucose deprivation/reperfusion injury
- Long noncoding RNA TUG1 regulates degradation of chondrocyte extracellular matrix via miR-320c/MMP-13 axis in osteoarthritis
- Duodenal adenocarcinoma with skin metastasis as initial manifestation: A case report
- Effects of Loofah cylindrica extract on learning and memory ability, brain tissue morphology, and immune function of aging mice
- Recombinant Bacteroides fragilis enterotoxin-1 (rBFT-1) promotes proliferation of colorectal cancer via CCL3-related molecular pathways
- Blocking circ_UBR4 suppressed proliferation, migration, and cell cycle progression of human vascular smooth muscle cells in atherosclerosis
- Gene therapy in PIDs, hemoglobin, ocular, neurodegenerative, and hemophilia B disorders
- Downregulation of circ_0037655 impedes glioma formation and metastasis via the regulation of miR-1229-3p/ITGB8 axis
- Vitamin D deficiency and cardiovascular risk in type 2 diabetes population
- Circ_0013359 facilitates the tumorigenicity of melanoma by regulating miR-136-5p/RAB9A axis
- Mechanisms of circular RNA circ_0066147 on pancreatic cancer progression
- lncRNA myocardial infarction-associated transcript (MIAT) knockdown alleviates LPS-induced chondrocytes inflammatory injury via regulating miR-488-3p/sex determining region Y-related HMG-box 11 (SOX11) axis
- Identification of circRNA circ-CSPP1 as a potent driver of colorectal cancer by directly targeting the miR-431/LASP1 axis
- Hyperhomocysteinemia exacerbates ischemia-reperfusion injury-induced acute kidney injury by mediating oxidative stress, DNA damage, JNK pathway, and apoptosis
- Potential prognostic markers and significant lncRNA–mRNA co-expression pairs in laryngeal squamous cell carcinoma
- Gamma irradiation-mediated inactivation of enveloped viruses with conservation of genome integrity: Potential application for SARS-CoV-2 inactivated vaccine development
- ADHFE1 is a correlative factor of patient survival in cancer
- The association of transcription factor Prox1 with the proliferation, migration, and invasion of lung cancer
- Is there a relationship between the prevalence of autoimmune thyroid disease and diabetic kidney disease?
- Immunoregulatory function of Dictyophora echinovolvata spore polysaccharides in immunocompromised mice induced by cyclophosphamide
- T cell epitopes of SARS-CoV-2 spike protein and conserved surface protein of Plasmodium malariae share sequence homology
- Anti-obesity effect and mechanism of mesenchymal stem cells influence on obese mice
- Long noncoding RNA HULC contributes to paclitaxel resistance in ovarian cancer via miR-137/ITGB8 axis
- Glucocorticoids protect HEI-OC1 cells from tunicamycin-induced cell damage via inhibiting endoplasmic reticulum stress
- Prognostic value of the neutrophil-to-lymphocyte ratio in acute organophosphorus pesticide poisoning
- Gastroprotective effects of diosgenin against HCl/ethanol-induced gastric mucosal injury through suppression of NF-κβ and myeloperoxidase activities
- Silencing of LINC00707 suppresses cell proliferation, migration, and invasion of osteosarcoma cells by modulating miR-338-3p/AHSA1 axis
- Successful extracorporeal membrane oxygenation resuscitation of patient with cardiogenic shock induced by phaeochromocytoma crisis mimicking hyperthyroidism: A case report
- Effects of miR-185-5p on replication of hepatitis C virus
- Lidocaine has antitumor effect on hepatocellular carcinoma via the circ_DYNC1H1/miR-520a-3p/USP14 axis
- Primary localized cutaneous nodular amyloidosis presenting as lymphatic malformation: A case report
- Multimodal magnetic resonance imaging analysis in the characteristics of Wilson’s disease: A case report and literature review
- Therapeutic potential of anticoagulant therapy in association with cytokine storm inhibition in severe cases of COVID-19: A case report
- Neoadjuvant immunotherapy combined with chemotherapy for locally advanced squamous cell lung carcinoma: A case report and literature review
- Rufinamide (RUF) suppresses inflammation and maintains the integrity of the blood–brain barrier during kainic acid-induced brain damage
- Inhibition of ADAM10 ameliorates doxorubicin-induced cardiac remodeling by suppressing N-cadherin cleavage
- Invasive ductal carcinoma and small lymphocytic lymphoma/chronic lymphocytic leukemia manifesting as a collision breast tumor: A case report and literature review
- Clonal diversity of the B cell receptor repertoire in patients with coronary in-stent restenosis and type 2 diabetes
- CTLA-4 promotes lymphoma progression through tumor stem cell enrichment and immunosuppression
- WDR74 promotes proliferation and metastasis in colorectal cancer cells through regulating the Wnt/β-catenin signaling pathway
- Down-regulation of IGHG1 enhances Protoporphyrin IX accumulation and inhibits hemin biosynthesis in colorectal cancer by suppressing the MEK-FECH axis
- Curcumin suppresses the progression of gastric cancer by regulating circ_0056618/miR-194-5p axis
- Scutellarin-induced A549 cell apoptosis depends on activation of the transforming growth factor-β1/smad2/ROS/caspase-3 pathway
- lncRNA NEAT1 regulates CYP1A2 and influences steroid-induced necrosis
- A two-microRNA signature predicts the progression of male thyroid cancer
- Isolation of microglia from retinas of chronic ocular hypertensive rats
- Changes of immune cells in patients with hepatocellular carcinoma treated by radiofrequency ablation and hepatectomy, a pilot study
- Calcineurin Aβ gene knockdown inhibits transient outward potassium current ion channel remodeling in hypertrophic ventricular myocyte
- Aberrant expression of PI3K/AKT signaling is involved in apoptosis resistance of hepatocellular carcinoma
- Clinical significance of activated Wnt/β-catenin signaling in apoptosis inhibition of oral cancer
- circ_CHFR regulates ox-LDL-mediated cell proliferation, apoptosis, and EndoMT by miR-15a-5p/EGFR axis in human brain microvessel endothelial cells
- Resveratrol pretreatment mitigates LPS-induced acute lung injury by regulating conventional dendritic cells’ maturation and function
- Ubiquitin-conjugating enzyme E2T promotes tumor stem cell characteristics and migration of cervical cancer cells by regulating the GRP78/FAK pathway
- Carriage of HLA-DRB1*11 and 1*12 alleles and risk factors in patients with breast cancer in Burkina Faso
- Protective effect of Lactobacillus-containing probiotics on intestinal mucosa of rats experiencing traumatic hemorrhagic shock
- Glucocorticoids induce osteonecrosis of the femoral head through the Hippo signaling pathway
- Endothelial cell-derived SSAO can increase MLC20 phosphorylation in VSMCs
- Downregulation of STOX1 is a novel prognostic biomarker for glioma patients
- miR-378a-3p regulates glioma cell chemosensitivity to cisplatin through IGF1R
- The molecular mechanisms underlying arecoline-induced cardiac fibrosis in rats
- TGF-β1-overexpressing mesenchymal stem cells reciprocally regulate Th17/Treg cells by regulating the expression of IFN-γ
- The influence of MTHFR genetic polymorphisms on methotrexate therapy in pediatric acute lymphoblastic leukemia
- Red blood cell distribution width-standard deviation but not red blood cell distribution width-coefficient of variation as a potential index for the diagnosis of iron-deficiency anemia in mid-pregnancy women
- Small cell neuroendocrine carcinoma expressing alpha fetoprotein in the endometrium
- Superoxide dismutase and the sigma1 receptor as key elements of the antioxidant system in human gastrointestinal tract cancers
- Molecular characterization and phylogenetic studies of Echinococcus granulosus and Taenia multiceps coenurus cysts in slaughtered sheep in Saudi Arabia
- ITGB5 mutation discovered in a Chinese family with blepharophimosis-ptosis-epicanthus inversus syndrome
- ACTB and GAPDH appear at multiple SDS-PAGE positions, thus not suitable as reference genes for determining protein loading in techniques like Western blotting
- Facilitation of mouse skin-derived precursor growth and yield by optimizing plating density
- 3,4-Dihydroxyphenylethanol ameliorates lipopolysaccharide-induced septic cardiac injury in a murine model
- Downregulation of PITX2 inhibits the proliferation and migration of liver cancer cells and induces cell apoptosis
- Expression of CDK9 in endometrial cancer tissues and its effect on the proliferation of HEC-1B
- Novel predictor of the occurrence of DKA in T1DM patients without infection: A combination of neutrophil/lymphocyte ratio and white blood cells
- Investigation of molecular regulation mechanism under the pathophysiology of subarachnoid hemorrhage
- miR-25-3p protects renal tubular epithelial cells from apoptosis induced by renal IRI by targeting DKK3
- Bioengineering and Biotechnology
- Green fabrication of Co and Co3O4 nanoparticles and their biomedical applications: A review
- Agriculture
- Effects of inorganic and organic selenium sources on the growth performance of broilers in China: A meta-analysis
- Crop-livestock integration practices, knowledge, and attitudes among smallholder farmers: Hedging against climate change-induced shocks in semi-arid Zimbabwe
- Food Science and Nutrition
- Effect of food processing on the antioxidant activity of flavones from Polygonatum odoratum (Mill.) Druce
- Vitamin D and iodine status was associated with the risk and complication of type 2 diabetes mellitus in China
- Diversity of microbiota in Slovak summer ewes’ cheese “Bryndza”
- Comparison between voltammetric detection methods for abalone-flavoring liquid
- Composition of low-molecular-weight glutenin subunits in common wheat (Triticum aestivum L.) and their effects on the rheological properties of dough
- Application of culture, PCR, and PacBio sequencing for determination of microbial composition of milk from subclinical mastitis dairy cows of smallholder farms
- Investigating microplastics and potentially toxic elements contamination in canned Tuna, Salmon, and Sardine fishes from Taif markets, KSA
- From bench to bar side: Evaluating the red wine storage lesion
- Establishment of an iodine model for prevention of iodine-excess-induced thyroid dysfunction in pregnant women
- Plant Sciences
- Characterization of GMPP from Dendrobium huoshanense yielding GDP-D-mannose
- Comparative analysis of the SPL gene family in five Rosaceae species: Fragaria vesca, Malus domestica, Prunus persica, Rubus occidentalis, and Pyrus pyrifolia
- Identification of leaf rust resistance genes Lr34 and Lr46 in common wheat (Triticum aestivum L. ssp. aestivum) lines of different origin using multiplex PCR
- Investigation of bioactivities of Taxus chinensis, Taxus cuspidata, and Taxus × media by gas chromatography-mass spectrometry
- Morphological structures and histochemistry of roots and shoots in Myricaria laxiflora (Tamaricaceae)
- Transcriptome analysis of resistance mechanism to potato wart disease
- In silico analysis of glycosyltransferase 2 family genes in duckweed (Spirodela polyrhiza) and its role in salt stress tolerance
- Comparative study on growth traits and ions regulation of zoysiagrasses under varied salinity treatments
- Role of MS1 homolog Ntms1 gene of tobacco infertility
- Biological characteristics and fungicide sensitivity of Pyricularia variabilis
- In silico/computational analysis of mevalonate pyrophosphate decarboxylase gene families in Campanulids
- Identification of novel drought-responsive miRNA regulatory network of drought stress response in common vetch (Vicia sativa)
- How photoautotrophy, photomixotrophy, and ventilation affect the stomata and fluorescence emission of pistachios rootstock?
- Apoplastic histochemical features of plant root walls that may facilitate ion uptake and retention
- Ecology and Environmental Sciences
- The impact of sewage sludge on the fungal communities in the rhizosphere and roots of barley and on barley yield
- Domestication of wild animals may provide a springboard for rapid variation of coronavirus
- Response of benthic invertebrate assemblages to seasonal and habitat condition in the Wewe River, Ashanti region (Ghana)
- Molecular record for the first authentication of Isaria cicadae from Vietnam
- Twig biomass allocation of Betula platyphylla in different habitats in Wudalianchi Volcano, northeast China
- Animal Sciences
- Supplementation of probiotics in water beneficial growth performance, carcass traits, immune function, and antioxidant capacity in broiler chickens
- Predators of the giant pine scale, Marchalina hellenica (Gennadius 1883; Hemiptera: Marchalinidae), out of its natural range in Turkey
- Honey in wound healing: An updated review
- NONMMUT140591.1 may serve as a ceRNA to regulate Gata5 in UT-B knockout-induced cardiac conduction block
- Radiotherapy for the treatment of pulmonary hydatidosis in sheep
- Retraction
- Retraction of “Long non-coding RNA TUG1 knockdown hinders the tumorigenesis of multiple myeloma by regulating microRNA-34a-5p/NOTCH1 signaling pathway”
- Special Issue on Reuse of Agro-Industrial By-Products
- An effect of positional isomerism of benzoic acid derivatives on antibacterial activity against Escherichia coli
- Special Issue on Computing and Artificial Techniques for Life Science Applications - Part II
- Relationship of Gensini score with retinal vessel diameter and arteriovenous ratio in senile CHD
- Effects of different enantiomers of amlodipine on lipid profiles and vasomotor factors in atherosclerotic rabbits
- Establishment of the New Zealand white rabbit animal model of fatty keratopathy associated with corneal neovascularization
- lncRNA MALAT1/miR-143 axis is a potential biomarker for in-stent restenosis and is involved in the multiplication of vascular smooth muscle cells