Apoplastic histochemical features of plant root walls that may facilitate ion uptake and retention
-
 and
and

Abstract
We used brightfield and epifluorescence microscopy, as well as permeability tests, to investigate the apoplastic histochemical features of plant roots associated with ion hyperaccumulation, invasion, and tolerance of oligotrophic conditions. In hyperaccumulator species with a hypodermis (exodermis absent), ions penetrated the root apex, including the root cap. By contrast, in non-hyperaccumulator species possessing an exodermis, ions did not penetrate the root cap. In vivo, the lignified hypodermis blocked the entry of ions into the cortex, while root exodermis absorbed ions and restricted them to the cortex. The roots of the hyperaccumulators Pteris vittata and Cardamine hupingshanensis, as well as the aquatic invasives Alternanthera philoxeroides, Eichhornia crassipes, and Pistia stratiotes, contained lignin and pectins. These compounds may trap and store ions before hypodermis maturation, facilitating ion hyperaccumulation and retention in the apoplastic spaces of the roots. These apoplastic histochemical features were consistent with certain species-specific characters, including ion hyperaccumulation, invasive behaviors in aquatic environments, or tolerance of oligotrophic conditions. We suggest that apoplastic histochemical features of the root may act as invasion mechanisms, allowing these invasive aquatic plants to outcompete indigenous plants for ions.
1 Introduction
Several species in the genus Pteris (Pteridaceae), including Pteris vittata, hyperaccumulate ions such as arsenic (As) and chromium (Cr) [1,2,3,4]. The uptake, transport, translocation, and detoxification of heavy metals in the roots and fronds of these species have been well studied [2,3,4]. Pteris species have also evolved various anatomical features and hyperaccumulator functions to adapt to terrestrial, xeric, epiphytic, and rupicolous environments [3,4,5,6,7]. Cardamine hupingshanensis (Brassicaceae), which is found in Selenium (Se)-rich environments, is another well-known hyperaccumulator of ions, including Se and cadmium (Cd) [8,9,10]. Some invasive aquatic plants, including Alternanthera philoxeroides (Amaranthaceae), Eichhornia crassipes (Pontederiaceae), and Pistia stratiotes (Araceae), also hyperaccumulate ions: these species purify eutrophic water bodies and tend to outcompete indigenous plants in similar hostile environments [11,12,13,14,15,16,17,18,19,20]. Similarly, plants in the Proteaceae have evolved cluster roots with lignified or phenol-rich cortical walls to adapt to environments deficient in phosphorus and other nutrients [21,22,23]. Plants with cluster roots potentially facilitate alterations in plant community structure [24] and outcompete species without cluster roots [25]. Thus, cluster roots are highly desirable in crop breeding [26]. Paspalum distichum (Poaceae), a typical amphibious plant, is a non-hyperaccumulator with an endodermis and an exodermis in its roots [27].
In vascular plants, the lignified, suberized endodermis and exodermis act as apoplastic barriers, restricting water-solute exchange, reducing oxygen loss after submersion, and supporting adaptation to terrestrial environments [10,27,28,29,30,31,32,33,34,35,36,37]. The exodermis has Casparian bands in the primary walls and has suberin lamellae and/or lignin in the secondary walls [27,28,29,30,32,33,34,35,38]. Permeability tests showed that the lignified cortex and the hypodermis block ion exchange in Alternanthera philoxeroides and brassicas [18,39,40,41,42,43,44]. The cortical walls of the cluster roots in the Proteaceae contain soluble phenolic or lignin-like compounds that retain fluorescent agents (e.g., fluorol yellow 088); the presence of these compounds reflects an adaptation to nutrient deprivation [21,45,46,47,48,49,50,51,52]. In addition, the velamen, rhizodermis, and hairs of epiphytic orchids have pectins, which also facilitate ion uptake [53,54,55]. The surfaces of the mucilage hairs of Brasenia schreberi (Cabombaceae) have polysaccharides in various patterns that absorb berberine during different development stages in vivo [31,56].
In this study, we aimed to identify the apoplastic histochemical features of the root cortical walls that facilitate ion uptake and retention, leading to ion hyperaccumulation and reflecting an adaptation to nutrient-deprived environments. To identify these features, we investigated the roots of seven representative hyperaccumulator, invasive, and/or oligotrophic plants: the aerial species, Pteris vittata and Chlorophytum comosum; the wetland species, Cardamine hupingshanensis and Paspalum distichum; and the aquatic species, Alternanthera philoxeroides, Eichhornia crassipes, and Pistia stratiotes. We also tested the apoplastic permeability of Pteris vittata and Paspalum distichum. An improved understanding of these plant roots’ apoplastic histochemical features might help explain how these plants become invasive, tolerate oligotrophic conditions, and hyperaccumulate ions [4,5,8,10,11,12,19,20,22,23,25,26]. These data will support the development of plants that can be used for the phytoremediation of ion-contaminated soils and oligotrophic water. Our results will also provide suggestions for the breeding of crops that can outcompete weed species [3,8,11,12,14,19,23,25,26].
2 Materials and methods
2.1 Plant sourcing and collection
Mature specimens of Pteris vittata, Paspalum distichum, Chlorophytum comosum, Cardamine hupingshanensis, Alternanthera philoxeroides, Eichhornia crassipes, and Pistia stratiotes were identified in the Testing Ground of Yangtze University (Jingzhou City, Hubei Province, China) in October 2020. We collected samples of the adventitious aerial roots of Pteris vittata, which grow on walls in the cracks between bricks, and of Chlorophytum comosum, which propagate via shoots with adventitious aerial roots. We collected the roots of Cardamine hupingshanensis and Paspalum distichum from a wetland area. We collected the roots of Alternanthera philoxeroides, Eichhornia crassipes, and Pistia stratiotes from ponds. Ten roots were collected from each species of five plants and immediately fixed in formaldehyde-alcohol-acetic acid [57]. Eight fresh, intact specimens of Pteris vittata and Paspalum distichum were used for the apoplastic permeability tests [18,30,33,34,35].
2.2 Microstructure and histochemistry
Root tissues were sectioned freehand, using a two-sided razor blade, under a stereoscope (JNOEC JSZ6, China). Root sections were cut at 10 and 20 mm from the root tip, as well as at the point where the cortex began to slough off. Sections were divided into three sets, such that each set included sections of each plant and at same distance from the root tip. Each set of sections was then stained with one of three stains: 0.1% (w/v) berberine hemisulfate-aniline blue (BAB) to test for Casparian bands and lignin in the cell walls [38,58], phloroglucinol-HCl to test for lignin in the cell walls [59], and 0.02% (w/v) ruthenium red to test for pectin in the cell walls [55,60].
All sections were washed 2–3 times with sterile water, mounted with sterile water, and examined using brightfield microscopy under a Leica DME microscope (Germany). Specimens were photographed with a digital camera and a micrometer (Nikon E5400, Japan). Specimens stained with BAB were viewed under ultraviolet light on an Olympus IX71 epifluorescence microscope with excitation filter G 365 nm, absorption filter barriers U-WB (blue light), dichromatic mirror DM 500, compensation excitation filter BP 450–480, and compensation absorption filter BA 515. BAB-stained specimens were photographed using a digital camera and a micrometer (RZ200C-21, Ruizhi Cop., China) [27].
2.3 Apoplastic permeability
We tested the apoplastic permeability of whole fresh specimens of Pteris vittata and Paspalum distichum. We tested ion uptake using the apoplastic permeability tests of Seago et al., Meyer et al., and Meyer and Peterson [38,61,62], with modifications. In brief, we immersed the roots of the whole plants in the berberine solution without separating the roots from the plants; the plants remained intact. This modification allowed us to use the permeability tests to assess how the plants absorbed ions. Three intact plant roots were left unstained as the negative control. Three additional intact plants roots (tracer control) were immersed in 100 mL of 0.05% berberine hemisulfate for 1 h and washed with sterile water. The final three intact plant roots were immersed in 100 mL of 0.05% berberine hemisulfate for 1 h, washed with sterile water, immersed in 0.05 M potassium thiocyanate for 0.5 h, and washed again with sterile water. Roots were sectioned freehand and viewed under UV light as described by Seago et al. [38].
3 Results and discussion
At 10 mm from the tips of the adventitious aerial roots of Pteris vittata, the root wall contained pectins from the endodermis to the rhizodermis and hairs (Figure 1a); the inner cortex had lignin-rich sclerenchyma layers and retained berberine around the endodermis (Figure 1b); and the surfaces of the rhizodermis and hairs accumulated substantial amounts of berberine or berberine thiocyanate crystals (Figure 1b–d). Berberine penetrated to the cortex of the Pteris vittata roots close to the root tips (Figure 1c and d), as indicated by the intense yellow fluorescence from the rhizodermis to the cortex. Similarly, intense yellow fluorescence was observed close to the tips of the roots of Paspalum distichum (Figure 1e), but berberine did not penetrate the root cap of this species. The walls of the adventitious aerial roots of Chlorophytum comosum also contained pectins from the endodermis to the rhizodermis and hairs (Figure 1f). Similar to Pteris vittata, the surfaces of the rhizodermis and hairs accumulated large amounts of berberine before metaxylem development (Figure 1g). After metaxylem development, the hairs were nearly sloughed off, but the exodermis and the rhizodermis surface continued to retain berberine (Figure 1h).

Photomicrographs of the adventitious roots of (a–d) Pteris vittata, (e) Paspalum distichum, and (f–h) Chlorophytum comosum. Scale bars = 50 μm. (a) Sectioned at 10 mm from root tip. Endodermis (arrowhead), cortex, rhizodermis, and hairs. Stain: ruthenium red. (b) Sectioned at 10 mm from root tip. Protoxylem, endodermis (arrowhead), lignified cortex (*), cortex, rhizodermis, and hairs. Stain: BAB. (c) Sectioned at 10 mm from root tip. Protoxylem, lignified thickened cortex (*), hypodermis, rhizodermis, and hairs, showing heavy accumulation of berberine thiocyanate. Stain: berberine (apoplastic tracer) and potassium thiocyanate. (d) Root tip showing root cap and entrance of berberine thiocyanate (arrowhead) close to the root tip; rhizodermis and hairs showing berberine thiocyanate accumulation (*). Stain: berberine (apoplastic tracer) and potassium thiocyanate. (e) Root tip showing root cap and entrance of berberine thiocyanate (arrowhead) close to the root tip; rhizodermis with limited berberine thiocyanate accumulation (*). Stain: berberine (apoplastic tracer) and potassium thiocyanate. (f) Sectioned at 10 mm from root tip. Pith, endodermis (arrowhead), cortex, hypodermis, rhizodermis, and hairs. Stain: ruthenium red. (g) Sectioned at 10 mm from root tip. Protoxylem, endodermis (arrowhead), cortex, exodermis (arrow), rhizodermis, and hairs. Stain: BAB. (h) Sectioned at 20 mm from root tip. Protoxylem, metaxylem, endodermis (arrowhead), cortex, exodermis (arrow), rhizodermis, and hairs. Stain: BAB. Abbreviations: ae, aerenchyma; co, cortex; h, hairs; hy, hypodermis; ic, intercellular space; mx, metaxylem; pa, parenchyma; pi, pith; px, protoxylem; rc, root cap; rh, rhizodermis; sc, sclerenchyma layer; sx, secondary xylem.
Before the cortex sloughed off, the adventitious roots of Cardamine hupingshanensis had pectins and lignin with even and Φ thickenings from the endodermis to the rhizodermis walls (Figure 2a–c). Similarly, pectins and lignified even thickenings were found from the endodermis to the rhizodermis walls in the adventitious roots of the aquatic plants Alternanthera philoxeroides (Figure 2d–f), Eichhornia crassipes (Figure 3a–c), and Pistia stratiotes (Figure 3d–f). In the adventitious roots of Alternanthera philoxeroides (Figure 2d–f) and Pistia stratiotes (Figure 3d–f), the cortex had typical radial schizogenous aerenchyma, while in the adventitious roots of Cardamine hupingshanensis (Figure 2a–c) and Eichhornia crassipes (Figure 3a–c), the cortex had radial lysigenous aerenchyma. The hypodermis of Eichhornia crassipes had lignified sclerenchyma layers (Figure 3a and c).
![Figure 2
Photomicrographs of the adventitious roots of (a–c) Cardamine hupingshanensis and (d–f) Alternanthera philoxeroides. Scale bars = 50 μm. (a) Sectioned at 10 mm from root tip. Protoxylem, metaxylem, endodermis (arrowhead), cortex, aerenchyma, hypodermis, and rhizodermis. Stain: ruthenium red. (b) Sectioned at 10 mm from root tip. Protoxylem, metaxylem, endodermis (arrowhead), cortex, lignified cortex (*), cortical lignified Φ thickenings (arrows), intercellular space, hypodermis, and rhizodermis. Stain: BAB. Image from [10] used with the permission of Open Life Sciences. (c) Sectioned at 50 mm from root tip. Secondary xylem, parenchyma, endodermis (white arrowhead), cortex, inner cortical lignified Φ thickening (black arrowhead), outer cortical lignified Φ thickenings (black arrows), lignified cortex (*), intercellular space, and hypodermis. Stain: phloroglucinol-HCl. Image from [10] used with the permission of Open Life Sciences. (d) Sectioned at 10 mm from root tip. Protoxylem, endodermis (arrowhead), cortex, aerenchyma, hypodermis, and rhizodermis. Stain: ruthenium red. (e) Sectioned at 10 mm from root tip. Protoxylem, endodermis (arrowhead), cortex, aerenchyma, lignified cortex (*), hypodermis, and rhizodermis. Stain: BAB. Image from [18] used with the permission of Flora. (f) Sectioned at 10 mm from root tip. Protoxylem, endodermis (arrowhead), cortex, aerenchyma, lignified cortex (*), and hypodermis. Stain: phloroglucinol-HCl.](/document/doi/10.1515/biol-2021-0137/asset/graphic/j_biol-2021-0137_fig_002.jpg)
Photomicrographs of the adventitious roots of (a–c) Cardamine hupingshanensis and (d–f) Alternanthera philoxeroides. Scale bars = 50 μm. (a) Sectioned at 10 mm from root tip. Protoxylem, metaxylem, endodermis (arrowhead), cortex, aerenchyma, hypodermis, and rhizodermis. Stain: ruthenium red. (b) Sectioned at 10 mm from root tip. Protoxylem, metaxylem, endodermis (arrowhead), cortex, lignified cortex (*), cortical lignified Φ thickenings (arrows), intercellular space, hypodermis, and rhizodermis. Stain: BAB. Image from [10] used with the permission of Open Life Sciences. (c) Sectioned at 50 mm from root tip. Secondary xylem, parenchyma, endodermis (white arrowhead), cortex, inner cortical lignified Φ thickening (black arrowhead), outer cortical lignified Φ thickenings (black arrows), lignified cortex (*), intercellular space, and hypodermis. Stain: phloroglucinol-HCl. Image from [10] used with the permission of Open Life Sciences. (d) Sectioned at 10 mm from root tip. Protoxylem, endodermis (arrowhead), cortex, aerenchyma, hypodermis, and rhizodermis. Stain: ruthenium red. (e) Sectioned at 10 mm from root tip. Protoxylem, endodermis (arrowhead), cortex, aerenchyma, lignified cortex (*), hypodermis, and rhizodermis. Stain: BAB. Image from [18] used with the permission of Flora. (f) Sectioned at 10 mm from root tip. Protoxylem, endodermis (arrowhead), cortex, aerenchyma, lignified cortex (*), and hypodermis. Stain: phloroglucinol-HCl.
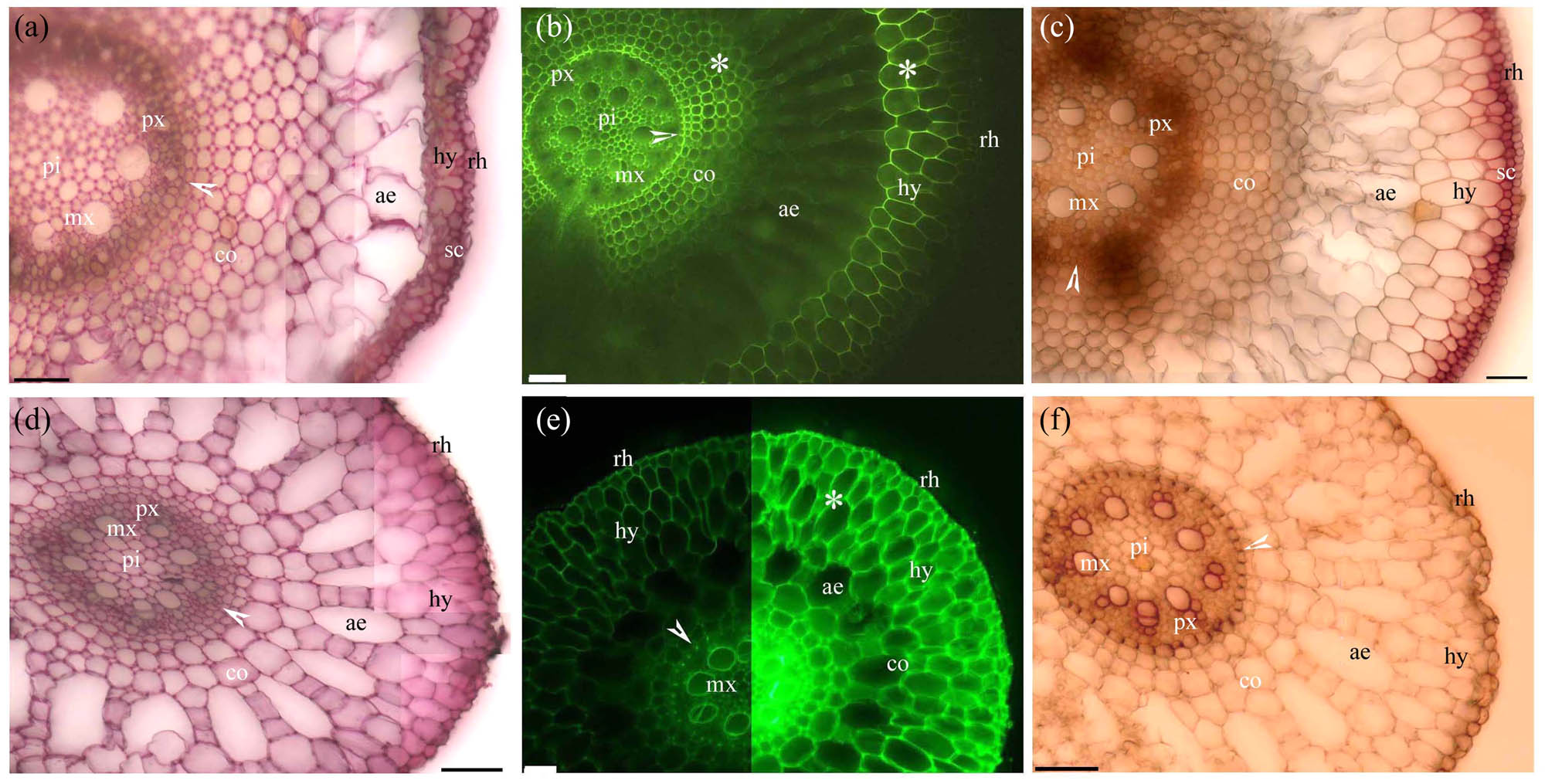
Photomicrographs of the adventitious roots of (a–c) Eichhornia crassipes and (d–f) Pistia stratiotes. Scale bars = 50 μm. (a) Sectioned at 10 mm from root tip. Pith, protoxylem, metaxylem, endodermis (arrowhead), cortex, aerenchyma, hypodermis, sclerenchyma layer, and rhizodermis. Stain: ruthenium red. (b) Sectioned at 10 mm from root tip. Pith, protoxylem, metaxylem, endodermis (arrowhead), cortex, aerenchyma, lignified cortex (*), hypodermis, and rhizodermis. Stain: BAB. (c) Sectioned at 10 mm from root tip. Pith, protoxylem, metaxylem, endodermis (white arrowhead), cortex, aerenchyma, lignified cortex (*), hypodermis, sclerenchyma layer, and rhizodermis. Stain: phloroglucinol-HCl. (d) Sectioned at 10 mm from root tip. Pith, protoxylem, metaxylem, endodermis (arrowhead), cortex, aerenchyma, hypodermis, and rhizodermis. Stain: ruthenium red. (e) Sectioned at 10 mm from root tip. Metaxylem, endodermis (arrowhead), cortex, aerenchyma, lignified cortex (*), hypodermis, and rhizodermis. Stain: BAB. (f) Sectioned at 10 mm from root tip. Pith, protoxylem, metaxylem, endodermis (arrowhead), cortex, aerenchyma, lignified cortex (*), hypodermis, and rhizodermis. Stain: phloroglucinol-HCl.
The endodermis and the exodermis are key impermeable apoplastic barriers that are common in vascular plant roots [28,29,36,37,63,64,65,66,67,68]. Apoplastic barriers can be histochemically characterized by the presence of Casparian bands, suberin lamellae, and lignin; these barriers protect tissues from oxygen shortages and inhibit water-solute exchanges [18,23,30,34,35,69,70,71,72,73,74]. All the species examined in this study had roots with an endodermis, but only the roots of Paspalum distichum and Chlorophytum comosum had an exodermis [27,75]. The roots of the other species included in this study (Pteris vittata, Cardamine hupingshanensis, Alternanthera philoxeroides, Eichhornia crassipes, and Pistia stratiotes) lacked an exodermis but possessed a hypodermis with lignin, as has been described in a variety of other plants, including Adiantum reniforme var. sinense, Brassica sp., Oenanthe javanica, Lycopodium obscurum, Pelargonium hortorum, Platycerium bifurcatum, and Selaginella sp. [10,37,39,40,41,42,43,44,66,76,77,78,79]. The cluster roots of various genera in the Proteaceae (e.g., Banksia, Grevillea, and Hakea) have a hypodermis containing soluble phenolic or lignin-like compounds, which have similar histochemical features with lignin of hypodermis in this studied species [21,49,50,51,52].
Lignified Φ and even thickenings that are distributed between the endodermis and the hypodermis of the cortical walls are typical of roots in the Brassicaceae, including in the genera Brassica, Noccaea, and Cardamine [10,39,40,41,42,43,44]. These thickenings act as barriers to ion transport. Unlike plants in the Brassicaceae, Pelargonium hortorum has larger Φ thickenings at the hypodermis [77]. The roots of Platycerium bifurcatum, Pleopeltis sp., and Doryopteris triphylla have lignified sclerenchyma layers around the endodermis [78,79,82,83,84], while the roots of Selaginella sp. have an exodermis [79]. In the heavy metal hyperaccumulator Noccaea caerulescens, the inner cortical walls of roots contain pectins and lignin [43]. Here lignified cortical thickenings were found in the roots of Pteris vittata [85], Cardamine hupingshanensis [10], Alternanthera philoxeroides [18], Eichhornia crassipes, and Pistia stratiotes.
In Pteris vittata and Chlorophytum comosum, the rhizodermis and hair walls contained pectins and accumulated a large amount of berberine. Similarly, the orchid root velamen also contains pectins and accumulates ions [53,54,55]. The root rhizodermis and hair walls of the hyperaccumulating ecotype of Sedum alfredii accumulated substantial Leadmium Green AM dye [80]; roots in this species also have thin inner cortical walls that contain large amounts of highly methylated pectin [81]. The root surfaces of Chlorophytum comosum retained berberine, similar to retention of polysaccharides by the smooth, immature mucilage hairs of Brasenia schreberi; the retention of berberine thiocyanate grains by the Pteris vittata root surfaces was similar to that of the mature mucilage hairs of Brasenia schreberi in vivo [31,56]. The rhizodermis surface retains little berberine in Adiantum reniforme var. sinense [37] and retains none in Metasequoia glyptostroboides [32], Cardamine hupingshanensis [10], and Alternanthera philoxeroides [18]. By contrast, our results showed that the surface of the root rhizodermis in Pteris vittata and Chlorophytum comosum retained substantial berberine. Pectins were present from the endodermis to the rhizodermis walls in Cardamine hupingshanensis, Alternanthera philoxeroides, Eichhornia crassipes, and Pistia stratiotes. However, pectins are only found in the peri-endodermal thickenings of Noccaea caerulescens [39,43].
In the apoplastic permeability test, the berberine tracer penetrated to the cortex of both Pteris vittata (exodermis absent) and Paspalum distichum (exodermis present) near the root tips [27,75], similar to what has been shown in Iris germanica (exodermis present) [61,86]. The berberine tracer also penetrated the root caps of Pteris vittata, similar to the results in Vicia faba (exodermis absent) [86]. However, the berberine tracer was unable to penetrate the root cap of Paspalum distichum, similar to what has been shown in Zea mays (exodermis present) and Iris germanica (exodermis present) [61,86]. Many berberine thiocyanate grains adhered to the mature hypodermis of Pteris vittata. By contrast, few berberine thiocyanate grains adhered to the mature exodermis of Paspalum distichum at the root surface [27,75]. The lignified hypodermis of Alternanthera philoxeroides blocks the entrance of ions into the cortex [18]. The root exodermis has only been shown to absorb berberine in vivo in Phalaris arundinacea, Zizania latifolia, and Artemisia spp. [30,34,35].
Based on the apoplastic histochemical features of the roots and their permeability, we hypothesize that the root hairs of Pteris vittata and Chlorophytum comosum have pectins that capture ions from the atmosphere, which helps these plants to survive in an oligotrophic aerial environment. Like Pteris vittata and Chlorophytum comosum, the epiphytic Orchidaceae use pectins to capture ions from the atmosphere [53,54,55]. Similarly, Brasenia schreberi uses polysaccharides to capture ions [7,31,39,53,54,55,56]. The carpet-like root system of Pteris vittata has many adventitious roots that absorb captured ions, leading to the hyperaccumulation of ions such as As and Cr [1,2,3,4,7,31,39,53,54,55,56,80,81,86,87]. We suggest that the lignified thickenings and pectins in the roots of Cardamine hupingshanensis, Alternanthera philoxeroides, Eichhornia crassipes, and Pistia stratiotes may trap ions before the hypodermis matures. These ions are then retained in the lignified walls, giving these species a competitive advantage over indigenous plants, particularly in oligotrophic environments [8,9,10,11,12,13,14,15,16,17,18,19,20,21,22,23,24,25,26,39,40,41,42,43,44]. Finally, the dense, fine roots of Cardamine hupingshanensis may allow the plant to hyperaccumulate Se in a manner that is similar to the hyperaccumulation of Cd in the dense root hairs of certain Arabidopsis thaliana genotypes [10,88] and in the phenol-rich cluster roots of species in the Proteaceae [21,22,23,24,25,49,50,51,52]. This ability to hyperaccumulate ions supports the adaptation of these plants to phosphorus deprivation and/or nutrient-poor environments [22,23,26,50,52].
4 Conclusion
Histochemical analyses indicate that pectins and lignin are present in several parts of the plant root walls, including the cortex, endodermis, exodermis or hypodermis, rhizodermis, and hairs [8,9,10,11,12,13,14,15,16,17,18,19,20,31,39,40,41,42,43,44,52,53,54,55,56,80,81,86]. These compounds, including the polysaccharides and phenolics, may facilitate ion uptake and retention in plants [8,9,10,11,12,13,14,15,16,17,18,19,20,21,22,23,24,25,26,27,28,29,30,31,34,35,39,40,41,42,43,44,52,53,54,55,56, 61,66,75,80,81,86]. In hyperaccumulator species without an exodermis (hypodermis), ions penetrate the root apex as well as the root cap [66,86]. By contrast, ions do not penetrate the root cap in non-hyperaccumulator species possessing an exodermis [27,61,66,75,86]. It has been shown in vivo that the lignified hypodermis of the root blocks the entry of ions into the cortex [18,86], while the root exodermis absorbs ions, trapping them within the exodermis walls [30,34,35,61,75,86]. The root hairs of Pteris vittata and Chlorophytum comosum are pectin-rich, reflecting an adaptation to the oligotrophic aerial environment [31,39,43,53,54,55,56,80,81]. The roots of the hyperaccumulators Pteris vittata and Cardamine hupingshanensis, as well as those of the invasive aquatic plants Alternanthera philoxeroides, Eichhornia crassipes, and Pistia stratiotes, have lignin and pectins in the cortex and rhizodermis; these structures may trap and store ions before hypodermis maturation. This hyperaccumulation of ions supports the survival of these plants in oligotrophic environments [8,9,10,11,12,13,14,15,16,17,18,19,20,21,22,23,24,25,26,39,40,41,42,43,44,52]. We suggest that the apoplastic histochemical features of invasive aquatic plant roots may allow such plants to acquire ions more efficiently than indigenous plants, and these features can thus be considered invasive mechanisms [11,12,13,14,15,16,17,18,19,20]. The histochemical features associated with hyperaccumulation are highly desirable for crop improvement, as well as when designing plants for the phytoremediation of ion-contaminated soils and for the population of eutrophic environments [11,12,13,14,15,16,17,18,19,20,26,50,51,52,89,90,91,92,93,94].
-
Funding information: This work was supported by the Hubei Province Key Laboratory of Conservation Biology for Shennongjia Golden Monkey Opening Fund (2019SNJ003), the Engineering Research Center of Ecology and Agriculture Use of Wetland, Ministry of Education Opening Fund, Yangtze University (KFT202004), and the Major Program on Technology Innovation of Hubei Province (2018ACA132).
-
Conflict of interest: The authors state no conflict of interest.
-
Data availability statement: The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
[1] Zhao FJ, Dunham SJ, Mcgrath SP. Arsenic hyperaccumulation by different fern species. N Phytologist. 2002;156(1):27–31.10.1046/j.1469-8137.2002.00493.xSearch in Google Scholar
[2] Oliveira LMD, Gress J, De J, Rathinasabapathi B, Marchi G, Chen YS, et al. Sulfate and chromate increased each other’s uptake and translocation in As-hyperaccumulator Pteris vittata. Chemosphere. 2016;147:36–43.10.1016/j.chemosphere.2015.12.088Search in Google Scholar
[3] Souri Z, Karimi N, Sandalio LM. Arsenic hyperaccumulation strategies: an overview. Front Cell Develop Biol. 2017;5:67.10.3389/fcell.2017.00067Search in Google Scholar
[4] Wagner S, Hoefer C, Puschenreiter M, Wenzel WW, Oburgera E, Hann S, et al. Arsenic redox transformations and cycling in the rhizosphere of Pteris vittata and Pteris quadriaurita. Environ Exp Botany. 2020;177:104122.10.1016/j.envexpbot.2020.104122Search in Google Scholar
[5] Sridhar BBM, Han FX, Diehl SV, Monts DL, Su Y. Effect of phytoaccumulation of arsenic and chromium on structural and ultrastructural changes of brake fern (Pteris vittata). Brazilian. J Plant Physiol. 2011;23:285–93.10.1590/S1677-04202011000400006Search in Google Scholar
[6] Martínez OG, Vilte I. The structure of petioles in Pteris (Pteridaceae). Am Fern J. 2012;102(1):1–11.10.1640/0002-8444-102.1.1Search in Google Scholar
[7] Gaafar AA, Ali SI, Faried AM, El-Hallouty SM. An insight into chemical content, biological effect and morphological features of Pteris vittata L. rarely growing in Egypt. Res J Chem Environ. 2018;22(10):47–55.Search in Google Scholar
[8] Bai HF, Li XM. Cadmium accumulation in hyperaccumulator Cardamine hupingshanensis. Jiangsu J Agric Sci. 2012;28(1):76–9.Search in Google Scholar
[9] Yuan LX, Zhu YY, Lin ZQ, Banuelos G, Li W, Yin XB. A novel selenocystine-accumulating plant in selenium-mine drainage area in Enshi, China. PLoS One. 2013;8(6):e65615.10.1371/journal.pone.0065615Search in Google Scholar
[10] Xiang JQ, Ming JJ, Yin HQ, Zhu YF, Li YJ, Long L, et al. Anatomy and histochemistry of the roots and shoots in the aquatic Selenium hyperaccumulator Cardamine hupingshanensis (Brassicaceae). Open Life Sci. 2019;14(1):318–26.10.1515/biol-2019-0035Search in Google Scholar
[11] Zhao YQ, Lu JB, Zhu L, Fu ZH. Effects of nutrient levels on growth characteristics and competitive ability of water hyacinth (Eichhornia crassipes), an aquatic invasive plant. Biodiversity Sci. 2006;14(2):159–64. 10.3321/j.issn:1005-0094.2006.02.010.Search in Google Scholar
[12] Liu HQ, Gao YQ, Song W, Han SQ, Huang JP. A experimental study on the removal of the nitrogen phosphorus of eutrophication-water and restraint of algae by Alternanthera Philoxeroides. Moden Agric Sci. 2008;15(12):89–92. CNKI:SUN:NCSY.0.2008-12-036.Search in Google Scholar
[13] Liao JX, Tao M, Jiang MX. Spatial arrangements affect suppression of invasive Alternanthera philoxeroides by native Hemarthria compressa. Acta Oecol. 2014;59:46–51. 10.1016/j.actao.2014.05.010.Search in Google Scholar
[14] Zhan JH, Li Q, Deng S, Cai Q, Zhao SM, Pan XJ. Purification Effect of eutrophic water body by purple-root water hyacinth. Earth Environ. 2014;42(3):389–96. CNKI:SUN:DZDQ.0.2014-03-018.Search in Google Scholar
[15] Singh AK, Majumder CB, Mishra S. Removal of arsenic from contaminated wastewater using Eichhornia crassipes. Integr Res Adv. 2015;2(1):1–4.Search in Google Scholar
[16] Lu L, Dong BC, Peter A, Yu FH. Effects of soil substrate heterogeneity and moisture on interspecific competition between Alternanthera philoxeroides and four native species. J Plant Ecol. 2016;10(3):528–37.Search in Google Scholar
[17] Wu H, Ismail M, Ding J. Global warming increases the interspecific competitiveness of the invasive plant alligator weed, Alternanthera philoxeroides. Sci Total Environ. 2016;575:1415–22.10.1016/j.scitotenv.2016.09.226Search in Google Scholar
[18] Yang CD, Yang XL, Zhang X, Zhou CY, Zhang F, Wang XE, et al. Anatomical structures of alligator weed (Alternanthera philoxeroides) suggest it is well adapted to the aquatic–terrestrial transition zone. Flora. 2019;253:27–34.10.1016/j.flora.2019.02.013Search in Google Scholar
[19] Schwantes D, Gonçalves AC, Schiller Ada P, Manfrin J, Campagnolo MA, Somavilla E. Pistia stratiotes in the phytoremediation and post-treatment of domestic sewage. Int J Phytoremediat. 2019;21(7):714–23.10.1080/15226514.2018.1556591Search in Google Scholar
[20] Abbasi T, Patnaik P, Tauseef SM, Abbasi SA. SHEFROL bioreactor enhances the ability of pistia (pistia stratiotes) in the phytoremediation of greywater. Int J Environ Anal Chem. 2020. 10.1080/03067319.2020.1738420.Search in Google Scholar
[21] Shane MW, Lambers H. Cluster roots: a curiosity in context. Plant Soil. 2005;274:101–25.10.1007/1-4020-4099-7_5Search in Google Scholar
[22] Hayes PE, Clode PL, Oliveira RS, Lambers H. Proteaceae from phosphorus-impoverished habitats preferentially allocate phosphorus to photosynthetic cells: an adaptation improving phosphorus-use efficiency. Plant Cell Environ. 2018;41(3):605–19.10.1111/pce.13124Search in Google Scholar
[23] Zúñiga-Feest A, Sepúlveda F, Delgado SM, Muñoz VG, Reyes-Diaz MPM. Gevuina avellana and Lomatia dentata, two Proteaceae species from evergreen temperate forests of South America exhibit contrasting physiological responses under nutrient deprivation. Plant Soil. 2020;464:29–44. 10.1007/s11104-020-04640-y.Search in Google Scholar
[24] Piper FI, Fajardo A, Baeza G, Cavieres LA. The association between a nurse cushion plant and a cluster-root bearing tree species alters the plant community structure. J Ecol. 2019;107(5):2182–96.10.1111/1365-2745.13188Search in Google Scholar
[25] Fajardo A, Piper FI. Cluster-root bearing Proteaceae species show a competitive advantage over non-cluster root species. Ann Botany. 2019;124(6):1121–31.10.1093/aob/mcz128Search in Google Scholar
[26] Lambers H, Shane MW, Cramer MD, Pearse SJ, Veneklaas EJ. Root structure and functioning for efficient acquisition of phosphorus: matching morphological and physiological traits. Ann Botany. 2006;98(4):693–713.10.1093/aob/mcl114Search in Google Scholar PubMed PubMed Central
[27] Yang CD, Zhang X, Zhou CY, Seago Jr JL. Root and stem anatomy and histochemistry of four grasses from the Jianghan Floodplain along the Yangtze River. China Flora. 2011;206:653–61.10.1016/j.flora.2010.11.011Search in Google Scholar
[28] Enstone DE, Peterson CA, Ma F. Root endodermis and exodermis: structure, function, and responses to the environment. J Plant Growth Regul. 2003;21:335–51.10.1007/s00344-003-0002-2Search in Google Scholar
[29] Geldner N. The endodermis. Annu Rev Plant Biol. 2013;64:531–58.10.1146/annurev-arplant-050312-120050Search in Google Scholar PubMed
[30] Yang CD, Zhang X, Li JK, Bao MZ, Ni DJ, Seago Jr JL. Anatomy and histochemistry of roots and shoots in wild rice (Zizania latifolia Griseb.). J Botany. 2014;2014:1–9.10.1155/2014/181727Search in Google Scholar
[31] Yang CD, Zhang X, Zhang F, Wang XE, Wang QF. Structure and ion physiology of Brasenia schreberi glandular trichomes in vivo. PeerJ. 2019;7:e7288.10.7717/peerj.7288Search in Google Scholar PubMed PubMed Central
[32] Yang CD, Zhang X, Wang T, Hu SS, Zhou CY, Zhang J, et al. Phenotypic plasticity in the structure of fine adventitious Metasequoia glyptostroboides roots allows adaptation to aquatic and terrestrial environments. Plants. 2019;8(11):501.10.3390/plants8110501Search in Google Scholar PubMed PubMed Central
[33] Yang CD, Zhang X, Seago Jr JL, Wang QF. Anatomical and histochemical features of Brasenia schreberi (Cabombaceae) shoots. Flora. 2020;263:151524.10.1016/j.flora.2019.151524Search in Google Scholar
[34] Zhang X, Hu LJ, Yang CD, Zhou CY, Yuan LY, Chen ZY, et al. Structural features of Phalaris arundinacea L. in the Jianghan Floodplain of the Yangtze River. China Flora. 2017;229:100–6.10.1016/j.flora.2017.02.016Search in Google Scholar
[35] Zhang X, Yang CD, Seago Jr JL. Anatomical and histochemical traits of roots and stems of Artemisia lavandulaefolia and A. selengensis (Asteraceae) in the Jianghan Floodplain, China. Flora. 2018;239:87–97.10.1016/j.flora.2017.11.009Search in Google Scholar
[36] Seago Jr JL. Revisiting the occurrence and evidence of endodermis in angiosperm shoots. Flora. 2020;273:151709.10.1016/j.flora.2020.151709Search in Google Scholar
[37] Wu D, Li L, Ma X, Huang G, Yang C. Morphological and anatomical adaptations to dry, shady environments in Adiantum reniforme var. sinense (Pteridaceae). PeerJ. 2020;8:e9937.10.7717/peerj.9937Search in Google Scholar PubMed PubMed Central
[38] Seago Jr JL, Peterson CA, Enstone DE, Scholey CA. Development of the endodermis and hypodermis of Typha glauca Godr. and T. angustifolia L. roots. Can J Botany. 1999;77(1):122–34.10.1139/b98-173Search in Google Scholar
[39] Zelko I, Lux A, Czibula K. Difference in the root structure of hyperaccumulator Thlaspi caerulescens and non-hyperaccumulator Thlaspi arvense. Int J Environ Pollut. 2008;33(2):123–32.10.1504/IJEP.2008.019387Search in Google Scholar
[40] Lopez-Perez L, Fernandez-Garcia N, Olmos E, Carvajal M. The phi thickening in roots of broccoli plants. An adaptation mechanism to salinity. Int J Plant Sci. 2007;168(8):1141–9.10.1086/520722Search in Google Scholar
[41] Fernandez-Garcia N, López-Perez L, Hernandez M, Olmos E. Role of phi cells and the endodermis under salt stress in Brassica oleracea. N Phytologist. 2009;181(2):347–60.10.1111/j.1469-8137.2008.02674.xSearch in Google Scholar PubMed
[42] Aleamotua M, McCurdy DW, Collings DA. Phi thickenings in roots: novel secondary wall structures responsive to biotic and abiotic stresses. J Exp Botany. 2019;70(18):4631–42.10.1093/jxb/erz240Search in Google Scholar PubMed
[43] Kováč J, Lux A, Soukup M, Weidinger M, Gruber D, Lichtscheidl-Schultz I, et al. A new insight on structural and some functional aspects of peri-endodermal thickenings, a specific layer in Noccaea caerulescens roots. Ann Botany. 2020;126(3):423–34.10.1093/aob/mcaa069Search in Google Scholar PubMed PubMed Central
[44] Aleamotuʻa M, Tai YT, McCurdy DW, Collings DA. Developmental biology and induction of phi thickenings by abiotic stress in roots of the Brassicaceae. Plants. 2018;7(2):47. 10.3390/plants7020047.Search in Google Scholar PubMed PubMed Central
[45] Kolattukudy PE. Biochemistry and function of cutin and suberin. Can J Botany. 2011;62(12):2918–33.10.1139/b84-391Search in Google Scholar
[46] Biggs AR, Miles NW. Association of suberin formation in uninoculated wounds with susceptibility to Leucostoma cincta and L. persoonii in various peach cultivars. Phytopathology. 1988;78(8):1070–4.10.1094/Phyto-78-1070Search in Google Scholar
[47] Lulai EC, Corsini DL. Differential deposition of suberin phenolic and aliphatic domains and their roles in resistance to infection during potato tuber (Solanum tuberosum L.) wound healing. Physioll Mol Plant Pathol. 1998;53:209–22.10.1006/pmpp.1998.0179Search in Google Scholar
[48] Thomas R, Fang X, Ranathunge K, Anderson TR, Peterson CA, Bernards MA. Soybean root suberin: anatomical distribution, chemical composition, and relationship to partial resistance to Phytophthora sojae. Plant Physiol. 2007;144(1):299–311.10.1104/pp.106.091090Search in Google Scholar PubMed PubMed Central
[49] Skene KR, Sutherland JM, Sprent RJI. Cluster root development in Grevillea robusta (Proteaceae). II. The development of the endodermis in a determinate root and in an indeterminate, lateral root. N Phytol. 1998;138(4):733–42.10.1046/j.1469-8137.1998.00153.xSearch in Google Scholar
[50] Lambers H, Albornoz FE, Kotula L, Laliberté E, Ranathunge K, Teste FP, et al. How below-ground interactions contribute to the coexistence of mycorrhizal and non-mycorrhizal species in severely phosphorus-impoverished hyperdiverse ecosystems. Plant Soil. 2018;424:11–33.10.1007/s11104-017-3427-2Search in Google Scholar
[51] Lambers H, Albornoz FE, Arruda AJ, Barker T, Finnegan PM, Gille C, et al. Nutrient acquisition strategies. In Lambers H. editor, A jewel in the crown of a global biodiversity hotspot. Perth: Kwongan Foundation and the Western Australian Naturalists’ Club Inc; 2019. p. 227–48.Search in Google Scholar
[52] Kotula L, Clode PL, Ranathunge K, Lambers H. Role of roots in adaptation of soil-indifferent Proteaceae to calcareous soils in south-western Australia. J Exp Botany. 2020;72:1490–505. 10.1093/jxb/eraa515.Search in Google Scholar PubMed
[53] Albersheim P, Darvill A, Roberts K, Sederoff R, Staehelin A. Plant cell walls: from chemistry to biology. New York: Garland Science; 2011.10.1201/9780203833476Search in Google Scholar
[54] Joca TAC, De Oliveira DC, Zotz G, Winkler U, Moreira ASFP. The velamen of epiphytic orchids: variation in structure and correlations with nutrient absorption. Flora. 2017;230:66–74.10.1016/j.flora.2017.03.009Search in Google Scholar
[55] Joca TAC, De Oliveira DC, Zotz G, Winkler U, Moreira ASFP. Chemical composition of cell walls in velamentous roots of epiphytic Orchidaceae. Protoplasma. 2020;257:103–18.10.1007/s00709-019-01421-ySearch in Google Scholar PubMed
[56] Feng S, Ning K, Luan D, Lu S, Sun P. Chemical composition and antioxidant capacities analysis of different parts of Brasenia schreberi. J Food Process Preservation. 2019;43(4):e14014.10.1111/jfpp.14014Search in Google Scholar
[57] Ruzin SE. Plant microtechnique and microscopy. New York: Oxford University Press; 1999.Search in Google Scholar
[58] Brundrett MC, Enstone DE, Peterson CA. A berberine–aniline blue fluorescent staining procedure for suberin, lignin and callose in plant tissue. Protoplasma. 1988;146:133–42.10.1007/BF01405922Search in Google Scholar
[59] Jensen WA. Botanical histochemistry–principles and practice. San Francisco, CA, USA: W.H. Freeman; 1962.Search in Google Scholar
[60] Peterson RL, Peterson CA, Melville LH. Teaching plant anatomy through creative laboratory exercise. Ontartio, Ottawa: N.P.C. Press; 2008.Search in Google Scholar
[61] Meyer CJ, Seago Jr JL, Peterson CA. Environmental effects on the maturation of the endodermis and multiseriate exodermis of Iris germanica roots. Ann Botany. 2009;103:687–702.10.1093/aob/mcn255Search in Google Scholar PubMed PubMed Central
[62] Meyer CJ, Peterson CA. Casparian bands occur in the periderm of Pelargonium hortorum stem and root. Ann Botany. 2011;107:591–8.10.1093/aob/mcq267Search in Google Scholar PubMed PubMed Central
[63] Fahn A. Plant anatomy. 4th edn. Oxford, UK: Pergamon Press; 1990.Search in Google Scholar
[64] Steudle E, Peterson CA. How does water get through roots? J Exp Botany. 1998;49(322):775–88.10.1093/jxb/49.322.775Search in Google Scholar
[65] Zimmermann HM, Steudle E. Apoplastic transport across young maize roots: effect of the exodermis. Planta. 1998;206:7–19. 10.1007/s004250050368.Search in Google Scholar
[66] Hose E, Clarkson DT, Steudle E, Schreiber L, Hartung W. The exodermis: a variable apoplastic barrier. J Exp Botany. 2001;52(365):2245–64.10.1093/jexbot/52.365.2245Search in Google Scholar PubMed
[67] Evert RF. Esau’s plant anatomy: meristems, cells, and tissues of the plant body: their structure, function, and development. 3rd edn. Hoboken, New Jersey, USA: Wiley-Interscience; 2006.10.1002/0470047380Search in Google Scholar
[68] Crang R, Lyons-Sobaski S, Wise R. Plant anatomy: a concept-based approach to the structure of seed plants. 1st edn. Switzerland: Springer, Gewerbestrasse; 2019.10.1007/978-3-319-77315-5_20Search in Google Scholar
[69] Colmer TD, Gibberd MR, Wiengweera A, Tinh TK. The barrier to radial oxygen loss from roots of rice (Oryza sativa L.) is induced by growth in stagnant solutions. J Exp Botany. 1998;49(325):1431–6.10.1093/jxb/49.325.1431Search in Google Scholar
[70] Seago Jr JL, Marsh LC, Stevens KJ, Soukup A, Votrubová O, Enstone DE. A re-examination of the root cortex in wetland flowering plants with respect to aerenchyma. Ann Botany. 2005;96(4):565–79.10.1093/aob/mci211Search in Google Scholar PubMed PubMed Central
[71] Armstrong J, Jones RE, Armstrong W. Rhizome phyllosphere oxygenation in Phragmites and other species in relation to redox potential convective gas flow, submergence and aeration pathways. N Phytologist. 2006;172(4):719–31.10.1111/j.1469-8137.2006.01878.xSearch in Google Scholar PubMed
[72] Soukup A, Armstrong W, Schreiber L, Rochus F, Votrubová O. Apoplastic barriers to radial oxygen loss and solute penetration: a chemical and functional comparison of the exodermis of two wetland species, Phragmites australis and Glyceria maxima. N Phytologist. 2007;173(2):264–78.10.1111/j.1469-8137.2006.01907.xSearch in Google Scholar PubMed
[73] Kotula L, Ranathunge K, Schreiber L, Steudle E. Functional and chemical comparison of apoplastic barriers to radial oxygen loss in roots of rice (Oryza sativa L.) grown in aerated or deoxygenated solution. J Exp Botany. 2009;60(7):2155–67.10.1093/jxb/erp089Search in Google Scholar PubMed
[74] Ranathunge K, Lin J, Steudle E, Schreiber L. Stagnant deoxygenated growth enhances root suberization and lignifications, but differentially affects water and NaCl permeabilities in rice (Oryza sativa L.) roots. Plant Cell Environ. 2011;34(8):1223–40.10.1111/j.1365-3040.2011.02318.xSearch in Google Scholar PubMed
[75] Yang CD, Zhang X. Permeability and supplement structures of stems of Paspalum distichum. Bull Botanical Res. 2013;33(5):564–8.Search in Google Scholar
[76] Zhang X, Hu LJ, Zhou CY, Yang CD. Studies on anatomy and apoplastic barrier histochemistry characters of Oenanthe javanica (Bl.) DC. Adapted to wetland environment. China Veg. 2016;2016(7):52–8.Search in Google Scholar
[77] Peterson CA, Emanuel ME, Weerdenburg CA. The permeability of phi thickenings in apple (Pyrus malus) and geranium (Pelargonium hortorum) roots to an apoplastic fluorescent dye tracer. Can J Botany. 1981;59(6):1107–10.10.1139/b81-149Search in Google Scholar
[78] Chapple CCS, Peterson RL. Root structure in the fern Platycerium bifurcatum (Cav.) C. Chr. (Polypodiaceae). Bot Gaz. 1987;148(2):180–7.10.1086/337646Search in Google Scholar
[79] Damus M, Peterson RL, Enstone DE, Peterson CA. Modifications of cortical cell walls in roots of seedless vascular plants. Bot Acta. 1997;110(2):190–5.10.1111/j.1438-8677.1997.tb00628.xSearch in Google Scholar
[80] Tao Q, Liu YK, Li M, Li JX, Luo JP, Lux A, et al. Cd-induced difference in root characteristics along root apex contributes to variation in Cd uptake and accumulation between two contrasting ecotypes of Sedum alfredii. Chemosphere. 2019;243:125290.10.1016/j.chemosphere.2019.125290Search in Google Scholar PubMed
[81] Guo X, Luo J, Du Y, Li J, Liu Y, Liang Y, et al. Coordination between root cell wall thickening and pectin modification is involved in cadmium accumulation in Sedum alfredii. Environ Pollut. 2021;268:115665.10.1016/j.envpol.2020.115665Search in Google Scholar PubMed
[82] Hernández MA, Teran L, Mata M, Martinez OG, Prado J. Helical cell wall thickenings in root cortical cells of Polypodiaceae species from Northwestern Argentina. Am Fern J. 2013;103(4):225–40.10.1640/0002-8444-103.4.225Search in Google Scholar
[83] Neira DA, Andrada AR, Páez VDLÁ, Rodriguez AMDV, Ríos NF, Martínez OG, et al. Anatomical, histochemical and cytogenetic features of Doryopteris triphylla (Pteridaceae). Am J Plant Sci. 2017;8(4):907–20.10.4236/ajps.2017.84061Search in Google Scholar
[84] Lagoria MDLÁ, Avila G, Neira DA, Rodríguez AM, Ríos NF, Prado J, et al. Morphoanatomical and histochemical characteristics of the epiphytic fern Pleopeltis macrocarpa (Polypodiaceae). Braz J Botany. 2018;41:739–50.10.1007/s40415-018-0474-8Search in Google Scholar
[85] Bondada B, Tu C, Ma L. Surface structure and anatomical aspects of Chinese brake fern (Pteris vittata; Pteridaceae). Brittonia. 2006;58(3):217–28.10.1663/0007-196X(2006)58[217:SSAAAO]2.0.CO;2Search in Google Scholar
[86] Enstone D, Peterson C. The apoplastic permeability of root apices. Can J Botany. 1992;70(7):1502–12.10.1139/b92-188Search in Google Scholar
[87] Pecková E, Tylová E, Soukup A. Tracing root permeability: comparison of tracer methods. Biol Plant. 2016;60(4):695–705.10.1007/s10535-016-0634-2Search in Google Scholar
[88] Kohanova J, Martinka M, Vaculik M, White PJ, Hauser MT, Lux A. Root hair abundance impacts cadmium accumulation in Arabidopsis thaliana shoots. Ann Botany. 2018;122(5):903–14.10.1093/aob/mcx220Search in Google Scholar
[89] Pask AJD, Sylvester-Bradley R, Jamieson PD, Foulkes MJ. Quantifying how winter wheat crops accumulate and use nitrogen reserves during growth. Field Crop Res. 2012;126:104–18.10.1016/j.fcr.2011.09.021Search in Google Scholar
[90] Lynch JP. Roots of the second green revolution. Aust J Botany. 2007;55(5):493–512.10.1071/BT06118Search in Google Scholar
[91] Lynch JP. Root phenotypes for improved nutrient capture: an underexploited opportunity for global agriculture. N Phytol. 2019;223(2):548–64.10.1111/nph.15738Search in Google Scholar
[92] Paez-Garcia A, Motes CM, Scheible WR, Chen RJ, Blancaflor EB, Monteros MJ. Root traits and phenotyping strategies for plant improvement. Plants. 2015;4(2):334–55.10.3390/plants4020334Search in Google Scholar
[93] White PJ. Root traits benefitting crop production in environments with limited water and nutrient availability. Ann Botany. 2019;124(6):883–90.10.1093/aob/mcz162Search in Google Scholar PubMed PubMed Central
[94] Griffiths M, York LM. Targeting root ion uptake kinetics to increase plant productivity and nutrient use efficiency. Plant Physiol. 2020;182:1854–68.10.1104/pp.19.01496Search in Google Scholar PubMed PubMed Central
© 2021 Di Wu et al., published by De Gruyter
This work is licensed under the Creative Commons Attribution 4.0 International License.
Articles in the same Issue
- Biomedical Sciences
- Research progress on the mechanism of orexin in pain regulation in different brain regions
- Adriamycin-resistant cells are significantly less fit than adriamycin-sensitive cells in cervical cancer
- Exogenous spermidine affects polyamine metabolism in the mouse hypothalamus
- Iris metastasis of diffuse large B-cell lymphoma misdiagnosed as primary angle-closure glaucoma: A case report and review of the literature
- LncRNA PVT1 promotes cervical cancer progression by sponging miR-503 to upregulate ARL2 expression
- Two new inflammatory markers related to the CURB-65 score for disease severity in patients with community-acquired pneumonia: The hypersensitive C-reactive protein to albumin ratio and fibrinogen to albumin ratio
- Circ_0091579 enhances the malignancy of hepatocellular carcinoma via miR-1287/PDK2 axis
- Silencing XIST mitigated lipopolysaccharide (LPS)-induced inflammatory injury in human lung fibroblast WI-38 cells through modulating miR-30b-5p/CCL16 axis and TLR4/NF-κB signaling pathway
- Protocatechuic acid attenuates cerebral aneurysm formation and progression by inhibiting TNF-alpha/Nrf-2/NF-kB-mediated inflammatory mechanisms in experimental rats
- ABCB1 polymorphism in clopidogrel-treated Montenegrin patients
- Metabolic profiling of fatty acids in Tripterygium wilfordii multiglucoside- and triptolide-induced liver-injured rats
- miR-338-3p inhibits cell growth, invasion, and EMT process in neuroblastoma through targeting MMP-2
- Verification of neuroprotective effects of alpha-lipoic acid on chronic neuropathic pain in a chronic constriction injury rat model
- Circ_WWC3 overexpression decelerates the progression of osteosarcoma by regulating miR-421/PDE7B axis
- Knockdown of TUG1 rescues cardiomyocyte hypertrophy through targeting the miR-497/MEF2C axis
- MiR-146b-3p protects against AR42J cell injury in cerulein-induced acute pancreatitis model through targeting Anxa2
- miR-299-3p suppresses cell progression and induces apoptosis by downregulating PAX3 in gastric cancer
- Diabetes and COVID-19
- Discovery of novel potential KIT inhibitors for the treatment of gastrointestinal stromal tumor
- TEAD4 is a novel independent predictor of prognosis in LGG patients with IDH mutation
- circTLK1 facilitates the proliferation and metastasis of renal cell carcinoma by regulating miR-495-3p/CBL axis
- microRNA-9-5p protects liver sinusoidal endothelial cell against oxygen glucose deprivation/reperfusion injury
- Long noncoding RNA TUG1 regulates degradation of chondrocyte extracellular matrix via miR-320c/MMP-13 axis in osteoarthritis
- Duodenal adenocarcinoma with skin metastasis as initial manifestation: A case report
- Effects of Loofah cylindrica extract on learning and memory ability, brain tissue morphology, and immune function of aging mice
- Recombinant Bacteroides fragilis enterotoxin-1 (rBFT-1) promotes proliferation of colorectal cancer via CCL3-related molecular pathways
- Blocking circ_UBR4 suppressed proliferation, migration, and cell cycle progression of human vascular smooth muscle cells in atherosclerosis
- Gene therapy in PIDs, hemoglobin, ocular, neurodegenerative, and hemophilia B disorders
- Downregulation of circ_0037655 impedes glioma formation and metastasis via the regulation of miR-1229-3p/ITGB8 axis
- Vitamin D deficiency and cardiovascular risk in type 2 diabetes population
- Circ_0013359 facilitates the tumorigenicity of melanoma by regulating miR-136-5p/RAB9A axis
- Mechanisms of circular RNA circ_0066147 on pancreatic cancer progression
- lncRNA myocardial infarction-associated transcript (MIAT) knockdown alleviates LPS-induced chondrocytes inflammatory injury via regulating miR-488-3p/sex determining region Y-related HMG-box 11 (SOX11) axis
- Identification of circRNA circ-CSPP1 as a potent driver of colorectal cancer by directly targeting the miR-431/LASP1 axis
- Hyperhomocysteinemia exacerbates ischemia-reperfusion injury-induced acute kidney injury by mediating oxidative stress, DNA damage, JNK pathway, and apoptosis
- Potential prognostic markers and significant lncRNA–mRNA co-expression pairs in laryngeal squamous cell carcinoma
- Gamma irradiation-mediated inactivation of enveloped viruses with conservation of genome integrity: Potential application for SARS-CoV-2 inactivated vaccine development
- ADHFE1 is a correlative factor of patient survival in cancer
- The association of transcription factor Prox1 with the proliferation, migration, and invasion of lung cancer
- Is there a relationship between the prevalence of autoimmune thyroid disease and diabetic kidney disease?
- Immunoregulatory function of Dictyophora echinovolvata spore polysaccharides in immunocompromised mice induced by cyclophosphamide
- T cell epitopes of SARS-CoV-2 spike protein and conserved surface protein of Plasmodium malariae share sequence homology
- Anti-obesity effect and mechanism of mesenchymal stem cells influence on obese mice
- Long noncoding RNA HULC contributes to paclitaxel resistance in ovarian cancer via miR-137/ITGB8 axis
- Glucocorticoids protect HEI-OC1 cells from tunicamycin-induced cell damage via inhibiting endoplasmic reticulum stress
- Prognostic value of the neutrophil-to-lymphocyte ratio in acute organophosphorus pesticide poisoning
- Gastroprotective effects of diosgenin against HCl/ethanol-induced gastric mucosal injury through suppression of NF-κβ and myeloperoxidase activities
- Silencing of LINC00707 suppresses cell proliferation, migration, and invasion of osteosarcoma cells by modulating miR-338-3p/AHSA1 axis
- Successful extracorporeal membrane oxygenation resuscitation of patient with cardiogenic shock induced by phaeochromocytoma crisis mimicking hyperthyroidism: A case report
- Effects of miR-185-5p on replication of hepatitis C virus
- Lidocaine has antitumor effect on hepatocellular carcinoma via the circ_DYNC1H1/miR-520a-3p/USP14 axis
- Primary localized cutaneous nodular amyloidosis presenting as lymphatic malformation: A case report
- Multimodal magnetic resonance imaging analysis in the characteristics of Wilson’s disease: A case report and literature review
- Therapeutic potential of anticoagulant therapy in association with cytokine storm inhibition in severe cases of COVID-19: A case report
- Neoadjuvant immunotherapy combined with chemotherapy for locally advanced squamous cell lung carcinoma: A case report and literature review
- Rufinamide (RUF) suppresses inflammation and maintains the integrity of the blood–brain barrier during kainic acid-induced brain damage
- Inhibition of ADAM10 ameliorates doxorubicin-induced cardiac remodeling by suppressing N-cadherin cleavage
- Invasive ductal carcinoma and small lymphocytic lymphoma/chronic lymphocytic leukemia manifesting as a collision breast tumor: A case report and literature review
- Clonal diversity of the B cell receptor repertoire in patients with coronary in-stent restenosis and type 2 diabetes
- CTLA-4 promotes lymphoma progression through tumor stem cell enrichment and immunosuppression
- WDR74 promotes proliferation and metastasis in colorectal cancer cells through regulating the Wnt/β-catenin signaling pathway
- Down-regulation of IGHG1 enhances Protoporphyrin IX accumulation and inhibits hemin biosynthesis in colorectal cancer by suppressing the MEK-FECH axis
- Curcumin suppresses the progression of gastric cancer by regulating circ_0056618/miR-194-5p axis
- Scutellarin-induced A549 cell apoptosis depends on activation of the transforming growth factor-β1/smad2/ROS/caspase-3 pathway
- lncRNA NEAT1 regulates CYP1A2 and influences steroid-induced necrosis
- A two-microRNA signature predicts the progression of male thyroid cancer
- Isolation of microglia from retinas of chronic ocular hypertensive rats
- Changes of immune cells in patients with hepatocellular carcinoma treated by radiofrequency ablation and hepatectomy, a pilot study
- Calcineurin Aβ gene knockdown inhibits transient outward potassium current ion channel remodeling in hypertrophic ventricular myocyte
- Aberrant expression of PI3K/AKT signaling is involved in apoptosis resistance of hepatocellular carcinoma
- Clinical significance of activated Wnt/β-catenin signaling in apoptosis inhibition of oral cancer
- circ_CHFR regulates ox-LDL-mediated cell proliferation, apoptosis, and EndoMT by miR-15a-5p/EGFR axis in human brain microvessel endothelial cells
- Resveratrol pretreatment mitigates LPS-induced acute lung injury by regulating conventional dendritic cells’ maturation and function
- Ubiquitin-conjugating enzyme E2T promotes tumor stem cell characteristics and migration of cervical cancer cells by regulating the GRP78/FAK pathway
- Carriage of HLA-DRB1*11 and 1*12 alleles and risk factors in patients with breast cancer in Burkina Faso
- Protective effect of Lactobacillus-containing probiotics on intestinal mucosa of rats experiencing traumatic hemorrhagic shock
- Glucocorticoids induce osteonecrosis of the femoral head through the Hippo signaling pathway
- Endothelial cell-derived SSAO can increase MLC20 phosphorylation in VSMCs
- Downregulation of STOX1 is a novel prognostic biomarker for glioma patients
- miR-378a-3p regulates glioma cell chemosensitivity to cisplatin through IGF1R
- The molecular mechanisms underlying arecoline-induced cardiac fibrosis in rats
- TGF-β1-overexpressing mesenchymal stem cells reciprocally regulate Th17/Treg cells by regulating the expression of IFN-γ
- The influence of MTHFR genetic polymorphisms on methotrexate therapy in pediatric acute lymphoblastic leukemia
- Red blood cell distribution width-standard deviation but not red blood cell distribution width-coefficient of variation as a potential index for the diagnosis of iron-deficiency anemia in mid-pregnancy women
- Small cell neuroendocrine carcinoma expressing alpha fetoprotein in the endometrium
- Superoxide dismutase and the sigma1 receptor as key elements of the antioxidant system in human gastrointestinal tract cancers
- Molecular characterization and phylogenetic studies of Echinococcus granulosus and Taenia multiceps coenurus cysts in slaughtered sheep in Saudi Arabia
- ITGB5 mutation discovered in a Chinese family with blepharophimosis-ptosis-epicanthus inversus syndrome
- ACTB and GAPDH appear at multiple SDS-PAGE positions, thus not suitable as reference genes for determining protein loading in techniques like Western blotting
- Facilitation of mouse skin-derived precursor growth and yield by optimizing plating density
- 3,4-Dihydroxyphenylethanol ameliorates lipopolysaccharide-induced septic cardiac injury in a murine model
- Downregulation of PITX2 inhibits the proliferation and migration of liver cancer cells and induces cell apoptosis
- Expression of CDK9 in endometrial cancer tissues and its effect on the proliferation of HEC-1B
- Novel predictor of the occurrence of DKA in T1DM patients without infection: A combination of neutrophil/lymphocyte ratio and white blood cells
- Investigation of molecular regulation mechanism under the pathophysiology of subarachnoid hemorrhage
- miR-25-3p protects renal tubular epithelial cells from apoptosis induced by renal IRI by targeting DKK3
- Bioengineering and Biotechnology
- Green fabrication of Co and Co3O4 nanoparticles and their biomedical applications: A review
- Agriculture
- Effects of inorganic and organic selenium sources on the growth performance of broilers in China: A meta-analysis
- Crop-livestock integration practices, knowledge, and attitudes among smallholder farmers: Hedging against climate change-induced shocks in semi-arid Zimbabwe
- Food Science and Nutrition
- Effect of food processing on the antioxidant activity of flavones from Polygonatum odoratum (Mill.) Druce
- Vitamin D and iodine status was associated with the risk and complication of type 2 diabetes mellitus in China
- Diversity of microbiota in Slovak summer ewes’ cheese “Bryndza”
- Comparison between voltammetric detection methods for abalone-flavoring liquid
- Composition of low-molecular-weight glutenin subunits in common wheat (Triticum aestivum L.) and their effects on the rheological properties of dough
- Application of culture, PCR, and PacBio sequencing for determination of microbial composition of milk from subclinical mastitis dairy cows of smallholder farms
- Investigating microplastics and potentially toxic elements contamination in canned Tuna, Salmon, and Sardine fishes from Taif markets, KSA
- From bench to bar side: Evaluating the red wine storage lesion
- Establishment of an iodine model for prevention of iodine-excess-induced thyroid dysfunction in pregnant women
- Plant Sciences
- Characterization of GMPP from Dendrobium huoshanense yielding GDP-D-mannose
- Comparative analysis of the SPL gene family in five Rosaceae species: Fragaria vesca, Malus domestica, Prunus persica, Rubus occidentalis, and Pyrus pyrifolia
- Identification of leaf rust resistance genes Lr34 and Lr46 in common wheat (Triticum aestivum L. ssp. aestivum) lines of different origin using multiplex PCR
- Investigation of bioactivities of Taxus chinensis, Taxus cuspidata, and Taxus × media by gas chromatography-mass spectrometry
- Morphological structures and histochemistry of roots and shoots in Myricaria laxiflora (Tamaricaceae)
- Transcriptome analysis of resistance mechanism to potato wart disease
- In silico analysis of glycosyltransferase 2 family genes in duckweed (Spirodela polyrhiza) and its role in salt stress tolerance
- Comparative study on growth traits and ions regulation of zoysiagrasses under varied salinity treatments
- Role of MS1 homolog Ntms1 gene of tobacco infertility
- Biological characteristics and fungicide sensitivity of Pyricularia variabilis
- In silico/computational analysis of mevalonate pyrophosphate decarboxylase gene families in Campanulids
- Identification of novel drought-responsive miRNA regulatory network of drought stress response in common vetch (Vicia sativa)
- How photoautotrophy, photomixotrophy, and ventilation affect the stomata and fluorescence emission of pistachios rootstock?
- Apoplastic histochemical features of plant root walls that may facilitate ion uptake and retention
- Ecology and Environmental Sciences
- The impact of sewage sludge on the fungal communities in the rhizosphere and roots of barley and on barley yield
- Domestication of wild animals may provide a springboard for rapid variation of coronavirus
- Response of benthic invertebrate assemblages to seasonal and habitat condition in the Wewe River, Ashanti region (Ghana)
- Molecular record for the first authentication of Isaria cicadae from Vietnam
- Twig biomass allocation of Betula platyphylla in different habitats in Wudalianchi Volcano, northeast China
- Animal Sciences
- Supplementation of probiotics in water beneficial growth performance, carcass traits, immune function, and antioxidant capacity in broiler chickens
- Predators of the giant pine scale, Marchalina hellenica (Gennadius 1883; Hemiptera: Marchalinidae), out of its natural range in Turkey
- Honey in wound healing: An updated review
- NONMMUT140591.1 may serve as a ceRNA to regulate Gata5 in UT-B knockout-induced cardiac conduction block
- Radiotherapy for the treatment of pulmonary hydatidosis in sheep
- Retraction
- Retraction of “Long non-coding RNA TUG1 knockdown hinders the tumorigenesis of multiple myeloma by regulating microRNA-34a-5p/NOTCH1 signaling pathway”
- Special Issue on Reuse of Agro-Industrial By-Products
- An effect of positional isomerism of benzoic acid derivatives on antibacterial activity against Escherichia coli
- Special Issue on Computing and Artificial Techniques for Life Science Applications - Part II
- Relationship of Gensini score with retinal vessel diameter and arteriovenous ratio in senile CHD
- Effects of different enantiomers of amlodipine on lipid profiles and vasomotor factors in atherosclerotic rabbits
- Establishment of the New Zealand white rabbit animal model of fatty keratopathy associated with corneal neovascularization
- lncRNA MALAT1/miR-143 axis is a potential biomarker for in-stent restenosis and is involved in the multiplication of vascular smooth muscle cells
Articles in the same Issue
- Biomedical Sciences
- Research progress on the mechanism of orexin in pain regulation in different brain regions
- Adriamycin-resistant cells are significantly less fit than adriamycin-sensitive cells in cervical cancer
- Exogenous spermidine affects polyamine metabolism in the mouse hypothalamus
- Iris metastasis of diffuse large B-cell lymphoma misdiagnosed as primary angle-closure glaucoma: A case report and review of the literature
- LncRNA PVT1 promotes cervical cancer progression by sponging miR-503 to upregulate ARL2 expression
- Two new inflammatory markers related to the CURB-65 score for disease severity in patients with community-acquired pneumonia: The hypersensitive C-reactive protein to albumin ratio and fibrinogen to albumin ratio
- Circ_0091579 enhances the malignancy of hepatocellular carcinoma via miR-1287/PDK2 axis
- Silencing XIST mitigated lipopolysaccharide (LPS)-induced inflammatory injury in human lung fibroblast WI-38 cells through modulating miR-30b-5p/CCL16 axis and TLR4/NF-κB signaling pathway
- Protocatechuic acid attenuates cerebral aneurysm formation and progression by inhibiting TNF-alpha/Nrf-2/NF-kB-mediated inflammatory mechanisms in experimental rats
- ABCB1 polymorphism in clopidogrel-treated Montenegrin patients
- Metabolic profiling of fatty acids in Tripterygium wilfordii multiglucoside- and triptolide-induced liver-injured rats
- miR-338-3p inhibits cell growth, invasion, and EMT process in neuroblastoma through targeting MMP-2
- Verification of neuroprotective effects of alpha-lipoic acid on chronic neuropathic pain in a chronic constriction injury rat model
- Circ_WWC3 overexpression decelerates the progression of osteosarcoma by regulating miR-421/PDE7B axis
- Knockdown of TUG1 rescues cardiomyocyte hypertrophy through targeting the miR-497/MEF2C axis
- MiR-146b-3p protects against AR42J cell injury in cerulein-induced acute pancreatitis model through targeting Anxa2
- miR-299-3p suppresses cell progression and induces apoptosis by downregulating PAX3 in gastric cancer
- Diabetes and COVID-19
- Discovery of novel potential KIT inhibitors for the treatment of gastrointestinal stromal tumor
- TEAD4 is a novel independent predictor of prognosis in LGG patients with IDH mutation
- circTLK1 facilitates the proliferation and metastasis of renal cell carcinoma by regulating miR-495-3p/CBL axis
- microRNA-9-5p protects liver sinusoidal endothelial cell against oxygen glucose deprivation/reperfusion injury
- Long noncoding RNA TUG1 regulates degradation of chondrocyte extracellular matrix via miR-320c/MMP-13 axis in osteoarthritis
- Duodenal adenocarcinoma with skin metastasis as initial manifestation: A case report
- Effects of Loofah cylindrica extract on learning and memory ability, brain tissue morphology, and immune function of aging mice
- Recombinant Bacteroides fragilis enterotoxin-1 (rBFT-1) promotes proliferation of colorectal cancer via CCL3-related molecular pathways
- Blocking circ_UBR4 suppressed proliferation, migration, and cell cycle progression of human vascular smooth muscle cells in atherosclerosis
- Gene therapy in PIDs, hemoglobin, ocular, neurodegenerative, and hemophilia B disorders
- Downregulation of circ_0037655 impedes glioma formation and metastasis via the regulation of miR-1229-3p/ITGB8 axis
- Vitamin D deficiency and cardiovascular risk in type 2 diabetes population
- Circ_0013359 facilitates the tumorigenicity of melanoma by regulating miR-136-5p/RAB9A axis
- Mechanisms of circular RNA circ_0066147 on pancreatic cancer progression
- lncRNA myocardial infarction-associated transcript (MIAT) knockdown alleviates LPS-induced chondrocytes inflammatory injury via regulating miR-488-3p/sex determining region Y-related HMG-box 11 (SOX11) axis
- Identification of circRNA circ-CSPP1 as a potent driver of colorectal cancer by directly targeting the miR-431/LASP1 axis
- Hyperhomocysteinemia exacerbates ischemia-reperfusion injury-induced acute kidney injury by mediating oxidative stress, DNA damage, JNK pathway, and apoptosis
- Potential prognostic markers and significant lncRNA–mRNA co-expression pairs in laryngeal squamous cell carcinoma
- Gamma irradiation-mediated inactivation of enveloped viruses with conservation of genome integrity: Potential application for SARS-CoV-2 inactivated vaccine development
- ADHFE1 is a correlative factor of patient survival in cancer
- The association of transcription factor Prox1 with the proliferation, migration, and invasion of lung cancer
- Is there a relationship between the prevalence of autoimmune thyroid disease and diabetic kidney disease?
- Immunoregulatory function of Dictyophora echinovolvata spore polysaccharides in immunocompromised mice induced by cyclophosphamide
- T cell epitopes of SARS-CoV-2 spike protein and conserved surface protein of Plasmodium malariae share sequence homology
- Anti-obesity effect and mechanism of mesenchymal stem cells influence on obese mice
- Long noncoding RNA HULC contributes to paclitaxel resistance in ovarian cancer via miR-137/ITGB8 axis
- Glucocorticoids protect HEI-OC1 cells from tunicamycin-induced cell damage via inhibiting endoplasmic reticulum stress
- Prognostic value of the neutrophil-to-lymphocyte ratio in acute organophosphorus pesticide poisoning
- Gastroprotective effects of diosgenin against HCl/ethanol-induced gastric mucosal injury through suppression of NF-κβ and myeloperoxidase activities
- Silencing of LINC00707 suppresses cell proliferation, migration, and invasion of osteosarcoma cells by modulating miR-338-3p/AHSA1 axis
- Successful extracorporeal membrane oxygenation resuscitation of patient with cardiogenic shock induced by phaeochromocytoma crisis mimicking hyperthyroidism: A case report
- Effects of miR-185-5p on replication of hepatitis C virus
- Lidocaine has antitumor effect on hepatocellular carcinoma via the circ_DYNC1H1/miR-520a-3p/USP14 axis
- Primary localized cutaneous nodular amyloidosis presenting as lymphatic malformation: A case report
- Multimodal magnetic resonance imaging analysis in the characteristics of Wilson’s disease: A case report and literature review
- Therapeutic potential of anticoagulant therapy in association with cytokine storm inhibition in severe cases of COVID-19: A case report
- Neoadjuvant immunotherapy combined with chemotherapy for locally advanced squamous cell lung carcinoma: A case report and literature review
- Rufinamide (RUF) suppresses inflammation and maintains the integrity of the blood–brain barrier during kainic acid-induced brain damage
- Inhibition of ADAM10 ameliorates doxorubicin-induced cardiac remodeling by suppressing N-cadherin cleavage
- Invasive ductal carcinoma and small lymphocytic lymphoma/chronic lymphocytic leukemia manifesting as a collision breast tumor: A case report and literature review
- Clonal diversity of the B cell receptor repertoire in patients with coronary in-stent restenosis and type 2 diabetes
- CTLA-4 promotes lymphoma progression through tumor stem cell enrichment and immunosuppression
- WDR74 promotes proliferation and metastasis in colorectal cancer cells through regulating the Wnt/β-catenin signaling pathway
- Down-regulation of IGHG1 enhances Protoporphyrin IX accumulation and inhibits hemin biosynthesis in colorectal cancer by suppressing the MEK-FECH axis
- Curcumin suppresses the progression of gastric cancer by regulating circ_0056618/miR-194-5p axis
- Scutellarin-induced A549 cell apoptosis depends on activation of the transforming growth factor-β1/smad2/ROS/caspase-3 pathway
- lncRNA NEAT1 regulates CYP1A2 and influences steroid-induced necrosis
- A two-microRNA signature predicts the progression of male thyroid cancer
- Isolation of microglia from retinas of chronic ocular hypertensive rats
- Changes of immune cells in patients with hepatocellular carcinoma treated by radiofrequency ablation and hepatectomy, a pilot study
- Calcineurin Aβ gene knockdown inhibits transient outward potassium current ion channel remodeling in hypertrophic ventricular myocyte
- Aberrant expression of PI3K/AKT signaling is involved in apoptosis resistance of hepatocellular carcinoma
- Clinical significance of activated Wnt/β-catenin signaling in apoptosis inhibition of oral cancer
- circ_CHFR regulates ox-LDL-mediated cell proliferation, apoptosis, and EndoMT by miR-15a-5p/EGFR axis in human brain microvessel endothelial cells
- Resveratrol pretreatment mitigates LPS-induced acute lung injury by regulating conventional dendritic cells’ maturation and function
- Ubiquitin-conjugating enzyme E2T promotes tumor stem cell characteristics and migration of cervical cancer cells by regulating the GRP78/FAK pathway
- Carriage of HLA-DRB1*11 and 1*12 alleles and risk factors in patients with breast cancer in Burkina Faso
- Protective effect of Lactobacillus-containing probiotics on intestinal mucosa of rats experiencing traumatic hemorrhagic shock
- Glucocorticoids induce osteonecrosis of the femoral head through the Hippo signaling pathway
- Endothelial cell-derived SSAO can increase MLC20 phosphorylation in VSMCs
- Downregulation of STOX1 is a novel prognostic biomarker for glioma patients
- miR-378a-3p regulates glioma cell chemosensitivity to cisplatin through IGF1R
- The molecular mechanisms underlying arecoline-induced cardiac fibrosis in rats
- TGF-β1-overexpressing mesenchymal stem cells reciprocally regulate Th17/Treg cells by regulating the expression of IFN-γ
- The influence of MTHFR genetic polymorphisms on methotrexate therapy in pediatric acute lymphoblastic leukemia
- Red blood cell distribution width-standard deviation but not red blood cell distribution width-coefficient of variation as a potential index for the diagnosis of iron-deficiency anemia in mid-pregnancy women
- Small cell neuroendocrine carcinoma expressing alpha fetoprotein in the endometrium
- Superoxide dismutase and the sigma1 receptor as key elements of the antioxidant system in human gastrointestinal tract cancers
- Molecular characterization and phylogenetic studies of Echinococcus granulosus and Taenia multiceps coenurus cysts in slaughtered sheep in Saudi Arabia
- ITGB5 mutation discovered in a Chinese family with blepharophimosis-ptosis-epicanthus inversus syndrome
- ACTB and GAPDH appear at multiple SDS-PAGE positions, thus not suitable as reference genes for determining protein loading in techniques like Western blotting
- Facilitation of mouse skin-derived precursor growth and yield by optimizing plating density
- 3,4-Dihydroxyphenylethanol ameliorates lipopolysaccharide-induced septic cardiac injury in a murine model
- Downregulation of PITX2 inhibits the proliferation and migration of liver cancer cells and induces cell apoptosis
- Expression of CDK9 in endometrial cancer tissues and its effect on the proliferation of HEC-1B
- Novel predictor of the occurrence of DKA in T1DM patients without infection: A combination of neutrophil/lymphocyte ratio and white blood cells
- Investigation of molecular regulation mechanism under the pathophysiology of subarachnoid hemorrhage
- miR-25-3p protects renal tubular epithelial cells from apoptosis induced by renal IRI by targeting DKK3
- Bioengineering and Biotechnology
- Green fabrication of Co and Co3O4 nanoparticles and their biomedical applications: A review
- Agriculture
- Effects of inorganic and organic selenium sources on the growth performance of broilers in China: A meta-analysis
- Crop-livestock integration practices, knowledge, and attitudes among smallholder farmers: Hedging against climate change-induced shocks in semi-arid Zimbabwe
- Food Science and Nutrition
- Effect of food processing on the antioxidant activity of flavones from Polygonatum odoratum (Mill.) Druce
- Vitamin D and iodine status was associated with the risk and complication of type 2 diabetes mellitus in China
- Diversity of microbiota in Slovak summer ewes’ cheese “Bryndza”
- Comparison between voltammetric detection methods for abalone-flavoring liquid
- Composition of low-molecular-weight glutenin subunits in common wheat (Triticum aestivum L.) and their effects on the rheological properties of dough
- Application of culture, PCR, and PacBio sequencing for determination of microbial composition of milk from subclinical mastitis dairy cows of smallholder farms
- Investigating microplastics and potentially toxic elements contamination in canned Tuna, Salmon, and Sardine fishes from Taif markets, KSA
- From bench to bar side: Evaluating the red wine storage lesion
- Establishment of an iodine model for prevention of iodine-excess-induced thyroid dysfunction in pregnant women
- Plant Sciences
- Characterization of GMPP from Dendrobium huoshanense yielding GDP-D-mannose
- Comparative analysis of the SPL gene family in five Rosaceae species: Fragaria vesca, Malus domestica, Prunus persica, Rubus occidentalis, and Pyrus pyrifolia
- Identification of leaf rust resistance genes Lr34 and Lr46 in common wheat (Triticum aestivum L. ssp. aestivum) lines of different origin using multiplex PCR
- Investigation of bioactivities of Taxus chinensis, Taxus cuspidata, and Taxus × media by gas chromatography-mass spectrometry
- Morphological structures and histochemistry of roots and shoots in Myricaria laxiflora (Tamaricaceae)
- Transcriptome analysis of resistance mechanism to potato wart disease
- In silico analysis of glycosyltransferase 2 family genes in duckweed (Spirodela polyrhiza) and its role in salt stress tolerance
- Comparative study on growth traits and ions regulation of zoysiagrasses under varied salinity treatments
- Role of MS1 homolog Ntms1 gene of tobacco infertility
- Biological characteristics and fungicide sensitivity of Pyricularia variabilis
- In silico/computational analysis of mevalonate pyrophosphate decarboxylase gene families in Campanulids
- Identification of novel drought-responsive miRNA regulatory network of drought stress response in common vetch (Vicia sativa)
- How photoautotrophy, photomixotrophy, and ventilation affect the stomata and fluorescence emission of pistachios rootstock?
- Apoplastic histochemical features of plant root walls that may facilitate ion uptake and retention
- Ecology and Environmental Sciences
- The impact of sewage sludge on the fungal communities in the rhizosphere and roots of barley and on barley yield
- Domestication of wild animals may provide a springboard for rapid variation of coronavirus
- Response of benthic invertebrate assemblages to seasonal and habitat condition in the Wewe River, Ashanti region (Ghana)
- Molecular record for the first authentication of Isaria cicadae from Vietnam
- Twig biomass allocation of Betula platyphylla in different habitats in Wudalianchi Volcano, northeast China
- Animal Sciences
- Supplementation of probiotics in water beneficial growth performance, carcass traits, immune function, and antioxidant capacity in broiler chickens
- Predators of the giant pine scale, Marchalina hellenica (Gennadius 1883; Hemiptera: Marchalinidae), out of its natural range in Turkey
- Honey in wound healing: An updated review
- NONMMUT140591.1 may serve as a ceRNA to regulate Gata5 in UT-B knockout-induced cardiac conduction block
- Radiotherapy for the treatment of pulmonary hydatidosis in sheep
- Retraction
- Retraction of “Long non-coding RNA TUG1 knockdown hinders the tumorigenesis of multiple myeloma by regulating microRNA-34a-5p/NOTCH1 signaling pathway”
- Special Issue on Reuse of Agro-Industrial By-Products
- An effect of positional isomerism of benzoic acid derivatives on antibacterial activity against Escherichia coli
- Special Issue on Computing and Artificial Techniques for Life Science Applications - Part II
- Relationship of Gensini score with retinal vessel diameter and arteriovenous ratio in senile CHD
- Effects of different enantiomers of amlodipine on lipid profiles and vasomotor factors in atherosclerotic rabbits
- Establishment of the New Zealand white rabbit animal model of fatty keratopathy associated with corneal neovascularization
- lncRNA MALAT1/miR-143 axis is a potential biomarker for in-stent restenosis and is involved in the multiplication of vascular smooth muscle cells