
Effects of heavy metals on bacterial community surrounding Bijiashan mining area located in northwest China
-
Yuan Liu

 ,
Xueying Wang
,
Xueying Wang

Abstract
Heavy metal (HM) pollution is a severe and common environmental problem in mining area soil. It is imperative to understand the micro ecological characteristics of mining area soil for HM contaminated soil remediation. This study described the effects of HM pollution level and soil physical and chemical parameters on microbial diversity. In this study, high-throughput sequencing technology was used to study the effects of HM pollution on the diversity and composition of the soil microbial community. The soil groups were barren, exhibiting alkaline pH, low total nitrogen (TN), and total potassium (TK) according to soil fertility standard. Compared with the control group, there was severe multiple HM pollution in the other five groups, including lead (Pb), cadmium (Cd), zinc (Zn), and copper (Cu). The dominant phyla accounting for more than 1% of the overall community in all soil groups were Proteobacteria (34.432 ± 7.478%), Actinobacteria (22.947 ± 4.297%), Acidobacteria (10.47 ± 2.439%), Chloroflexi (7.89 ± 2.980%), Planctomycetota (5.993 ± 1.558%), Bacteroidota (4.275 ± 1.980%), Cyanobacteria (3.478 ± 2.196%), Myxococcus (2.888 ± 0.822%), Gemmatimonadota (2.448 ± 0.447%), Firmicutes (1.193 ± 0.634%), Patescibacteria (0.435 ± 0.813%), and Nitrospirota (0.612 ± 0.468%). Proteobacteria and Actinobacteria were predominant at the phylum level, which showed a certain tolerance to HMs. In addition, redundancy analysis (RDA) results showed that Pb, Cu, Zn, and Cd were strongly correlated with each other (P < 0.01). Other nutrient elements (except for TK) were significantly positively correlated with each other. Cu and nutrient element TK had an important impact on bacterial community structure. Therefore, bacteria with the function of HM tolerance and bioremediation in extreme environments should be researched, which provides a foundation for future ecological remediation of contaminated soil by using microbial remediation technology.
Graphical abstract
1 Introduction
Soil pollution by heavy metals (HMs), caused by rapid social and economic development activities, is one of the most serious environmental problems faced globally [1,2]. Although the development and utilization of mineral resources can guarantee the stable development of the economy, the limitation of management level and development technology causes a series of ecological environment problems, including local vegetation destruction, grassland degradation, soil erosion, and water pollution [3,4]. Acidic wastewater and solid waste are rich in HMs. They migrate and accumulate in the surrounding environment and within humans, causing severe harm to the environment and human health [5,6]. Bijiashan mining area is a typical sedimentary reformed lead–zinc (Pb–Zn) deposit in Chengxian County, Gansu Province [7].
Microorganisms play an essential role in soil. They can not only adsorb HMs [8] but change the rhizosphere nutrition conditions through their own metabolites. For example, iron carriers and plant growth promoters can enhance and transfer heavy metals in plants. Furthermore, microbes can repair the HM contaminated soil [9]. There are increasingly more studies on the interaction between HMs and microorganisms, but due to the soil environment’s complexity, few studies have characterized the in-situ soil bacterial community under HM stress [10].
The biological toxicity of HMs affects the community structure and function of microorganisms in the soil. Deng et al. investigated the microbial diversity in farmland, persistently polluted by HMs, and found that the abundance of fungi and bacteria decreased significantly [11]. Previous studies have found that HM pollution can reduce the microbial biomass in the soil [12]. However, with the increase in HM pollution, the relative abundance of microbial communities also increased due to a decrease in total biomass and competitive resources [13].
We studied the main physical and chemical properties and bacterial community structure in the soil from the Bijiashan mining area. The results showed that the soil in the mining area is seriously polluted by Pb, Zn, cadmium (Cd), and copper (Cu) compared with the control group. In this study, we chose the Bijiashan mining area in the Longnan City of Gansu Province, northwestern China, as the focus of our investigation and adopted the soil pollution load index method and 16S rRNA high-throughput sequencing technology to achieve the following research purposes: (1) To evaluate the HM pollution in the surrounding soil of Bijiashan mining area. (2) To reveal the diversity and distribution pattern of the bacterial community in the mining area. (3) To analyze the main environmental factors affecting bacterial community diversity in the mining area. (4) To screen potential bacteria for HM bioremediation in the mining area.
2 Materials and methods
2.1 Collection of soil samples
Soil samples were collected in the Bijiashan mining area (105°44′0″ E and 33°50′37″ N) located in Chengxian County, Longnan City, Gansu Province, northwestern China on September 19, 2020 (Figure 1 and Table 1). The soil samples were as follows: the ore drainage port (SO), the two rhizosphere soil and non-rhizosphere soil (S1, S2) in the mining area, and the soil samples 30 km away from the mining area as the control (SF) (Figure 1). The local temperature was 17°C, and the relative humidity was 72% that day. We first removed the surface debris and then collected 0–20 cm soil. Three equal amounts of soil samples were randomly taken at each sampling point. The samples were kept on ice and transported to the laboratory as soon as possible. Each sample was divided into two parts. One part was naturally dried to determine physical and chemical properties and HM contents. The other part was immediately stored at −80°C for biodiversity determination.
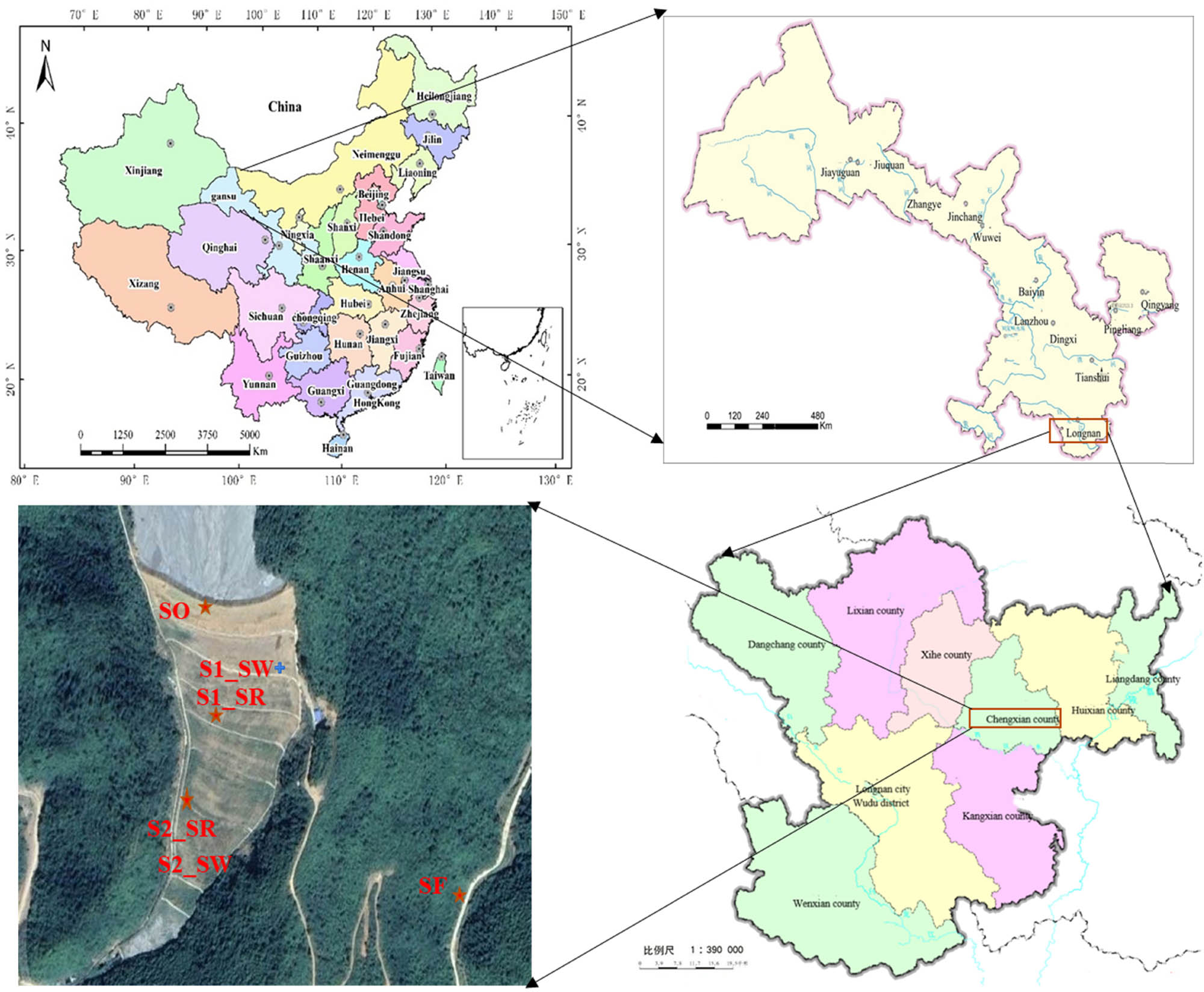
Distribution of sample area and sampling points.
Sampling point information
| Groups | Number | Average elevation/m | Longitude (E) | Latitude (N) | Oxygen content (%) |
|---|---|---|---|---|---|
| Ore discharge port | SO | 1230.99 | 105°44′0″ | 33°50′37″ | 18.96 |
| Rhizosphere soil No.1 | S1_SR | 1186.98 | 105°44′3″ | 33°50′37″ | 18.99 |
| Non-rhizosphere soil No.1 | S1_SW | ||||
| Rhizosphere soil No.2 | S2_SR | 1074.84 | 105°41′29″ | 33°55′28″ | 19.21 |
| Non-rhizosphere soil No.2 | S2_SW | ||||
| Control soil | SF | 992.08 | 105°43′49″ | 33°47′46″ | 19.44 |
2.2 Determination of soil physical and chemical properties
All soil samples were dried at room temperature and passed through a 2 mm sieve. The physicochemical properties of the soil samples were determined by conventional methods. Soil pH was measured at 2.5:1 (soil:water) using a pH meter (PHS-3E, Shanghai, China) [14]. Chemically stabilized organic matter (COM) was determined by oxygen reduction titration with butyrate [15]. Total nitrogen (TN) was investigated according to Kjeldahl method [16,17]. Total phosphorus (TP) was determined by sodium hydroxide alkali fusion molybdenum antimony method [18]. Total potassium (TK) was measured by flame photometer [19]. HM elements (Cu, Zn, Pb, and Cd) were detected by flame atomic absorption spectrophotometry (TAS-990F, Beijing, China) [20]. Soil nitrate-nitrogen (NO3 −N) and ammonium nitrogen (NH4 +_N) were extracted by 2 mol/L KCl solution (1:5 w/v) for 30 min, and the concentrations of the 2 compounds were determined by flow injection automatic analyzer [21]. The pollution load index (PLI) method was used to evaluate the comprehensive pollution status of HMs in the study area [4].
where
Background value of soil in Gansu Province
| Heavy metal elements | Cu | Zn | Cd | Pb |
|---|---|---|---|---|
| Soil content (mg/kg) | 24.1 | 68.5 | 0.116 | 18.8 |
2.3 DNA extraction and high-throughput sequencing
Total DNA was extracted from the soil using the FastDNA® Spin Kit for Soil (Beijing Lianlixin BioTech Co., Ltd, Beijing, China). The concentration and purity of DNA were determined by micro ultraviolet spectrophotometry (ZXHD/TL4, Beijing, China).
The v3–v4 region of the 16S rRNA gene in soil samples was amplified and sequenced by Takara kit (Takara Bio Inc., Japan). The specific primers, 515F (5′-ACTCCTACGGGAGGCAGCAG-3′) and 907R (5′-GGACTACHVGGGTWTCTAAT-3′) [23,24,25] were used in a total reaction volume of 25 µL, which consisted of 2.5 µL of 10× PCR buffer, 0.4 µM dNTPs, 2.0 µM 5 U Ex Taq enzyme, 0. 25 µL and 10 ng DNA with ddH2O [26]. The PCR amplification conditions were adjusted to pre-denaturation at 94°C for 3 min, denaturation at 94°C for 30 s, annealing at 56°C for 60 s, extension at 72°C for 30 s, then 30 cycles of 94°C for 30 s, and extension at 72°C for 10 min. The mixture of PCR products was migrated on a 2% agarose gel using electrophoresis and purified using Gene JET Genomic DNA Purification gel Recovery Kit (Thermo Fisher K0881, USA). After DNA extraction, soil samples were sent to Shanghai Meiji biological Co., Ltd, where 16S rRNA gene sequencing was undertaken on the Illumina MiSeq PE300 platform. The raw sequencing data obtained in this study has been submitted to the NCBI Sequence Read Archive database (Accession Number: PRJNA723126).
2.4 Bioinformatics and statistical analysis
Diversity indices, including Shannon, Simpson, Ace, and Chao, were calculated to evaluate the richness and diversity of the soil bacterial community. According to the number and abundance of Operational Taxonomic Units (OTUs), bacterial community richness (Chao and Ace) was adopted to reflect the richness of the species in the communities, and Shannon and Simpson indices were calculated by DPS v2.1.2 to evaluate the soil bacterial community diversity. Two-way analysis of variance (ANOVA) was used to compare the differences in the soil chemical properties, bacterial alpha (α) diversities, and relative abundances of bacteria taxonomy among the different soil groups. This was followed by Duncan’s multiple comparison test (P < 0.05), using SPSS 22.0 and the software Qiime v1.9.1 and R program. Principal component analysis (PCA) and hierarchical clustering based on Bray–Curtis distances (non-metric multidimensional scaling [NMDS]) were performed to investigate the similarities and differences in bacterial communities between samples. In addition, to identify the vital environmental variables influencing the bacterial community structure, redundancy analysis (RDA) was implemented. RDA was performed via the vegan package in R v3.4.2. Spearman’s correlation analysis was conducted to reveal the relationship between the soil chemical properties and the relative abundances of bacterial taxonomy using R v3.4.2 and SPSS 22.0 (SPSS Inc., Chicago, IL, USA).
3 Results
3.1 Analysis of soil physical and chemical properties and pollution assessment
Soil pH can regulate carbon mineralization through microbial activities and communities. The pH and HM content are shown in Table 3. The soil pH was similar among the six groups. All of the soil samples were alkaline.
Concentration and pollution index of heavy metals in soil
| Groups | Number | pH | Cu | Zn | Cd | Pb | PLI |
|---|---|---|---|---|---|---|---|
| Ore discharge port | SO | 8.40 ± 0.04a | 61.8 ± 7.104a | 748.66 ± 14.85d | 2.49 ± 0.04d | 453.03 ± 4.565cd | 10.897 |
| Rhizosphere soil No.1 | S1_SR | 8.96 ± 0.63a | 42.9 ± 7.398c | 2726.2 ± 163.6b | 9.15 ± 0.242b | 1199.593 ± 33.50b | 8.314 |
| Non-rhizosphere soil No.1 | S1_SW | 8.97 ± 0.36a | 58.4 ± 3.653b | 5045.0 ± 353a | 16.64 ± 0.83a | 1912.7 ± 114.5a | 41.290 |
| Rhizosphere soil No.2 | S2_SR | 8.85 ± 0.44a | 19.5 ± 3.201f | 257.7 ± 4.3b | 0.97 ± 0.05e | 89.7 ± 1.5d | 4.488 |
| Non-rhizosphere soil No.2 | S2_SW | 8.95 ± 0.45a | 30.5 ± 7.398d | 1814.67 ± 791c | 5.59 ± 2.848c | 609.5 ± 7.8c | 15.915 |
| Control soil | SF | 8.443 ± 0.065a | 23.7 ± 3.421e | 19.2 ± 12.9d | 0.653 ± 0.241e | 67.9 ± 1.3e | 1.53 |
Note: Cu, Pb, Zn, Cd (mg/kg). Different lower case letters in the same column indicated significant differences among soil samples from different sampling points (P < 0.05) followed by Duncan multiple comparison test.
The order of soil pollution load index was as follows: S1_SW > S2_SW > SO > S1_SR > S2_SR > SF. Among the six soil samples, the control sample (SF) was not polluted, and the other five samples were seriously polluted by HMs. The total Cu, Zn, and Pb concentrations under S1_SW was significantly higher than that in other soil groups (P < 0.05). The pollution degree of S1_SW was highest and the most concerning. The pollution degree of No.1 and No.2 non-rhizosphere soil samples was higher than the other four samples (Table 3) (P < 0.05).
The data are known as mean values ± standard deviations (n = 3). Different lowercase letters in the same column indicate that there are significant differences among soil samples from different sampling points (P < 0.05) by two-way analysis of variance (ANOVA) followed by Duncan’s multiple comparison test.
According to the Chinese soil fertility classification standard, we can accept that the cation exchange capacity (CEC) of S1_SR (9.92) was higher than low level (6.2). The CEC of the other five soil samples was very low. In addition to S1_SR (upper level) and S2_SW (middle level), the other four soil groups belonged to the low level (6–10). The TN content in S1_SR (1.772) was high level, and SO (0.343) and S1_SW (0.446) belonged to the very low level (<0.65). The TN content of S2_SR soil (0.859) was in the middle level (0.8–1.0). The TN content between S2_SW and SF was low (0.65–0.8). TP content of S1_SR and S2_SR was also at a low level (0.5–0.7). The TP content of the remaining four soils was very low (<0.5). The TK content of the six soil groups was at a very low level (<1). It can be seen that the soil fertility indices among SO, S1_SW, and S2_SW were at a low and very low level, indicating that the soil environment surrounding the mining area is extremely barren. The content of all physical and chemical properties in S1_SR was higher than S1_SW (P < 0.05), and the major difference of nutrient content between rhizosphere soil and non-rhizosphere soil indicates that the existence of plants improved the soil fertility in the mining area to a certain extent (P < 0.05).
The data are known as mean values ± standard deviations (n = 3). Different small letters in the same column indicated significant differences among soil samples from different sampling points (P < 0.05) followed by Duncan’s multiple comparison test.
3.2 Bacterial community structures
For this study, a total of 1,787 OTUs were detected in six soil groups and were classified into 30 phyla, 81 classes, 170 orders, 264 families, 477 genera, and 883 species.
The sequencing work was relatively comprehensive in covering bacterial diversity because the rarefaction curves tended to saturate (Figure 2a). Shannon curves showed that the data of diversity analysis was large enough to reflect the microbial species within the samples (Figure 2b).
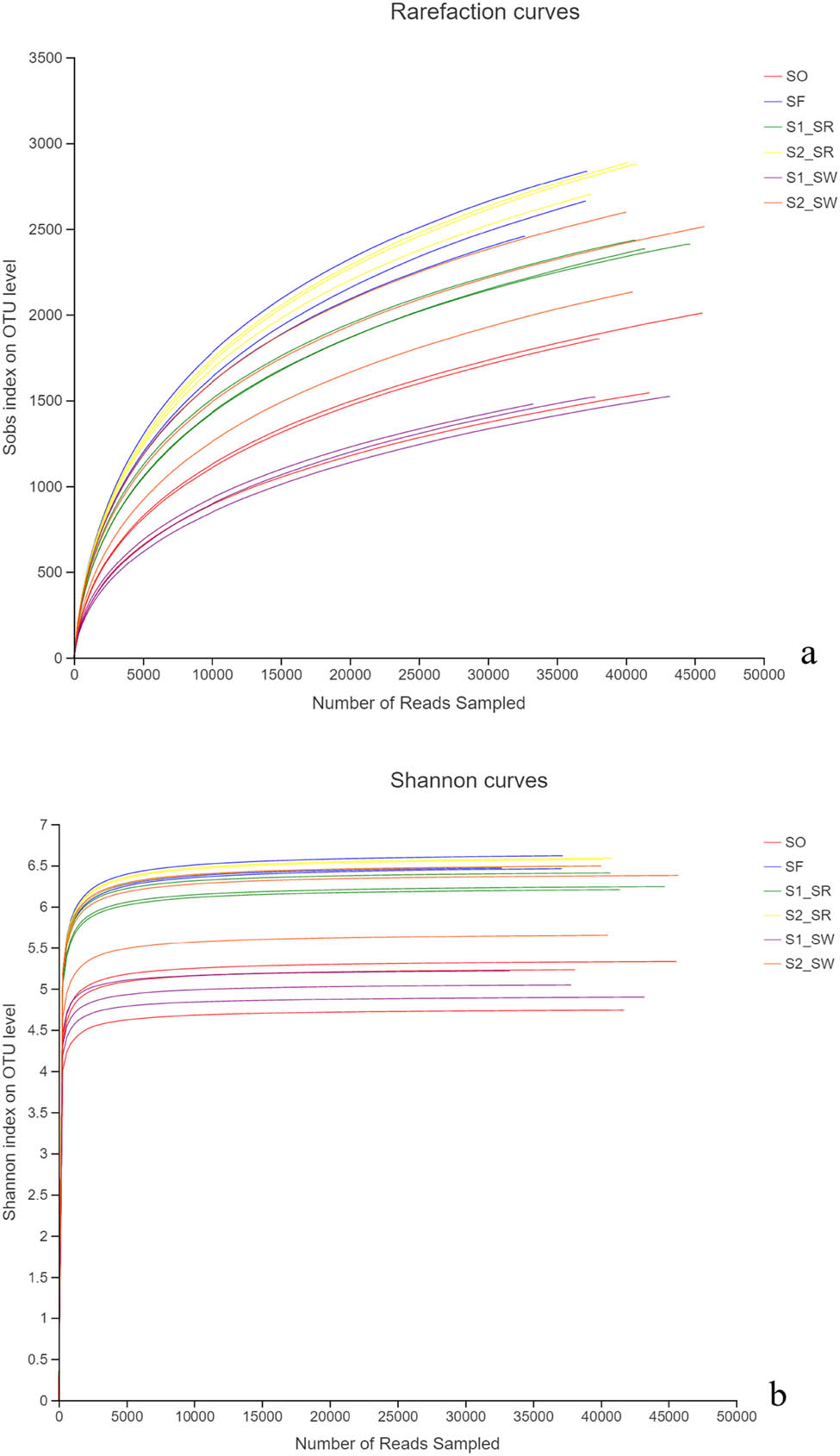
(a) Rarefaction curves and (b) Shannon–Wiener curve.
The length of rank abundance curves on the horizontal axis reflected the number of species among the six soil groups (Figure 3). The uniformity of the curves could indirectly reflect the low uniformity of species composition, indicating that the species distribution in the six sampling points is uneven.
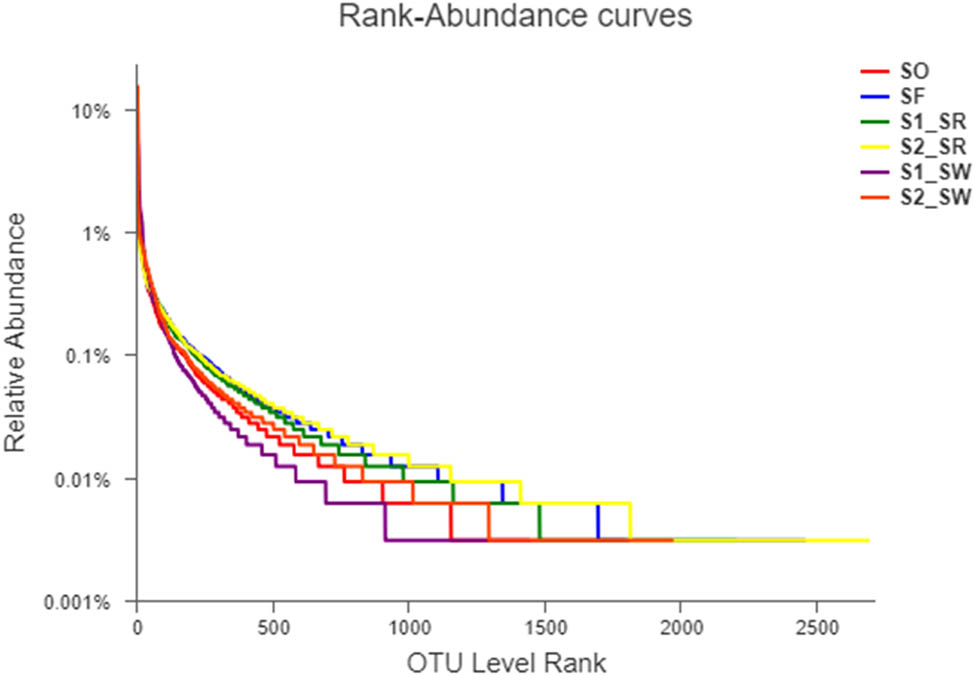
Rank abundance curves.
3.3 α Diversity
The α diversity indices of soil bacterial community were different in all soil samples (Table 4). Species richness index (OTU) and Chao index could reflect the species richness of the community, while the Simpson’s and Shannon indices could reflect the community species diversity. If the Shannon value is larger, the community diversity is higher. If the Simpson value is larger, this would indicate lower microbial community diversity.
Diversity indices of soil bacterial community in the six soil samples
| Index | SO | S1_SR | S1_SW | S2_SR | S2_SW | SF |
|---|---|---|---|---|---|---|
| Chao | 2670.473c | 3104.609b | 2372.538c | 3685.296a | 3145.042b | 3584.321a |
| Ace | 2588.136c | 3098.390b | 2165.975d | 3726.683a | 3168.627b | 3540.378ab |
| Shannon | 5.099b | 6.285a | 5.052b | 6.551a | 6.174a | 6.355a |
| Simpson | 0.038a | 0.005b | 0.027a | 0.004b | 0.011b | 0.006b |
Note: Different lower case letters in the same line indicated significant differences among soil samples from different sampling points (P < 0.05) followed by Duncan multiple comparison test.
Among the six soil samples, the indices of Chao, Ace, and Shannon from rhizosphere soil No.2 (S2_SR) were higher than the other five samples (P < 0.05), while those of rhizosphere soil No.1 (S1_SR) were lower than those of the other five samples (P < 0.05). It can be seen from the table that the community diversity of rhizosphere soil No.2 was the highest, and that of non-rhizosphere soil No.1 was the lowest (P < 0.05). Shannon diversity index was as follows: S2_SR > SF > S1_SR > S2_SW > SO > S1_SW. Shannon diversity index showed that there was a significant difference in α diversity index among control sample SF, SO, and S1_SW (P < 0.05), which indicated that HM pollution had an impact on α diversity of the bacterial community. Species diversity of bacteria community in the rhizosphere soil was higher than that of the non-rhizosphere soil (P < 0.05), but there was no significant difference in the four diversity indices between the S2_SR and the S2_SW (P < 0.05). Similarly, there was no significant difference in the Chao and Ace index between the SO and the S1_SR (P < 0.05). Therefore, the order of soil bacterial community richness at the six sampling sites was as follows: S2_SR > SF > S2_SW > S1_SR > SO > S1_SW. It can be seen that the richness and diversity of rhizosphere soil was greater than that of non-rhizosphere soil at the same sampling point (P < 0.05).
The Wayne map showed the differences of unique and common OTUs among the six soil samples (Figure 4a). The Venn diagram showed the number of OTU among six soil samples was as follows: S2_SR (4,000) > SF (3,728) > S2_SW (3,443) > S1_SR (3,397) > SO (2,735) > S1_SW (2,258).
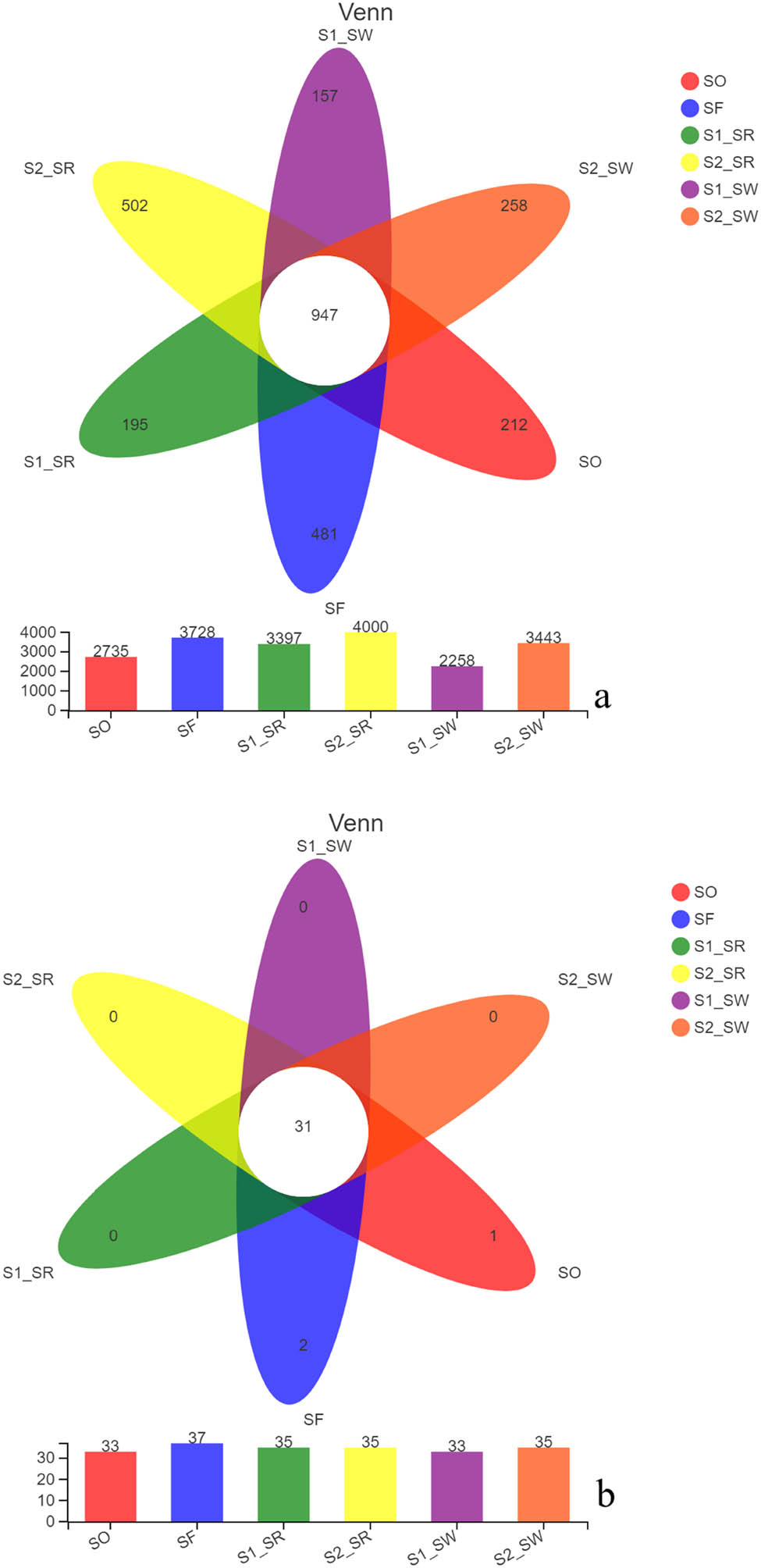
(a) OTU Wayne diagram of six soil samples and (b) Venn diagram of species in six soil samples.
From largest to smallest, the number of differences between the unique OTUs in the six soil groups was: S2_SR (502) > SF (481) > S2_SW (258) > SO (212) > S1_SR (195) > S1_SW (157). The number of OTUs jointly owned by the six soil groups was 947.
At the phylum level, from largest to smallest the number of species in six soil groups was as follows: SF (37) > S1_SR (35) = S2_SR (35) = S2_SW (35) > SO (33) = S1_SW (33). The number of species common among all samples was 31 (Figure 4b). SO and SF had one and two endemic species, respectively. The abundance of bacteria in rhizosphere soil was higher than that in non-rhizosphere soil, indicating that plant rhizosphere surrounding has a significant effect on the abundance of bacteria within the community (P < 0.05).
3.4 Beta (β) diversity
At the phylum level, based on the relative abundance of bacteria in soil samples, the PCA of soil bacterial communities in six sampling points was carried out (Figure 5a). The first two axes (PC1 and PC2) explained 21.88 and 20.01% of the total variance of bacteria in the six soil groups, respectively. The PCA result, which reflected the similarities and differences of bacterial community composition among samples, exhibited short distances between SO and SF (P < 0.001). The small dispersion of the SF indicated that the bacterial communities of SF are similar but there were obvious separation phenomena in the six sampling points. This clearly demonstrates that the soil bacterial communities are distinct from the six soil groups. Rhizosphere groups were independent of non-rhizosphere, indicating significant differences in community diversity between rhizosphere soil and non-rhizosphere soil.
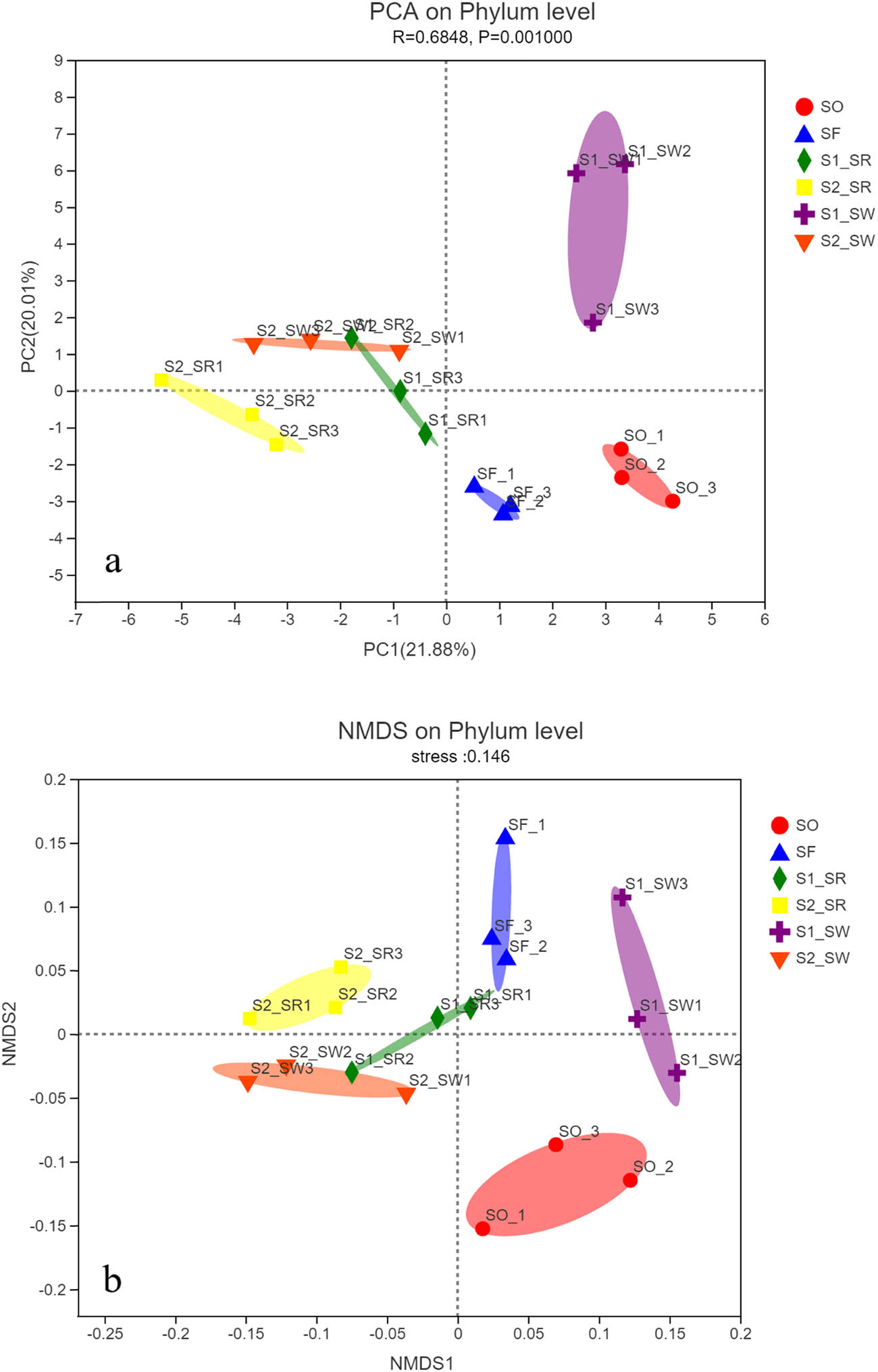
(a) PCA of the bacterial communities in sample and (b) NMDS of the bacterial communities in sample. Note: PCA of sampling points are based on Bray–Curtis microbial community distance. The horizontal and vertical axes represent two eigenvalues, which can best reflect the variance. Each point represents a sample, and the same color is the same sample point.
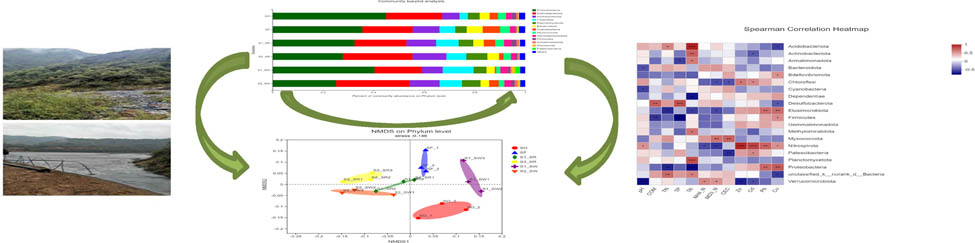
Phylum level clustering of samples using NMDS (stress = 0.146) indicated clear variation in the microbial profiles among the soil groups (Figure 5b).
3.5 The nature of bacterial community
The sequencing reads recovered from all soil groups classified at the phylum level were affiliated with 12 bacterial phyla (Figure 6). The dominant phyla accounting for more than 1% of the overall community in all soil groups were Proteobacteria (34.432 ± 7.478%), Actinobacteria (22.947 ± 4.297%), Acidobacteria (10.47 ± 2.439%), Chloroflexi (7.89 ± 2.980%), Planctomycetota (5.993 ± 1.558%), Bacteroidota (4.275 ± 1.980%), Cyanobacteria (3.478 ± 2.196%), Myxococcus (2.888 ± 0.822%), Gemmatimonadota (2.448 ± 0.447%), Firmicutes (1.193 ± 0.634%), Patescibacteria (0.435 ± 0.813%), and Nitrospirota (0.612 ± 0.468%). Among bacteria, Proteobacteria, Actinomycetes, and Acidobacteria were the three dominant phyla across all soil groups. Bacterial phyla with less abundance in all soil samples included Patescibacteria and Nitrospirota. Nevertheless, the relative abundance of dominant phyla differed among different soil samples. For example, all soil groups exhibited highest relative abundance of Proteobacteria, but the relative abundance of soil groups was as follows: SO (44.98%) > S1_SW (40.4%) > SF (35.6%) > S1_SR (32.5%) > S2_SR (27.89%) > S2_SW (25.22%).
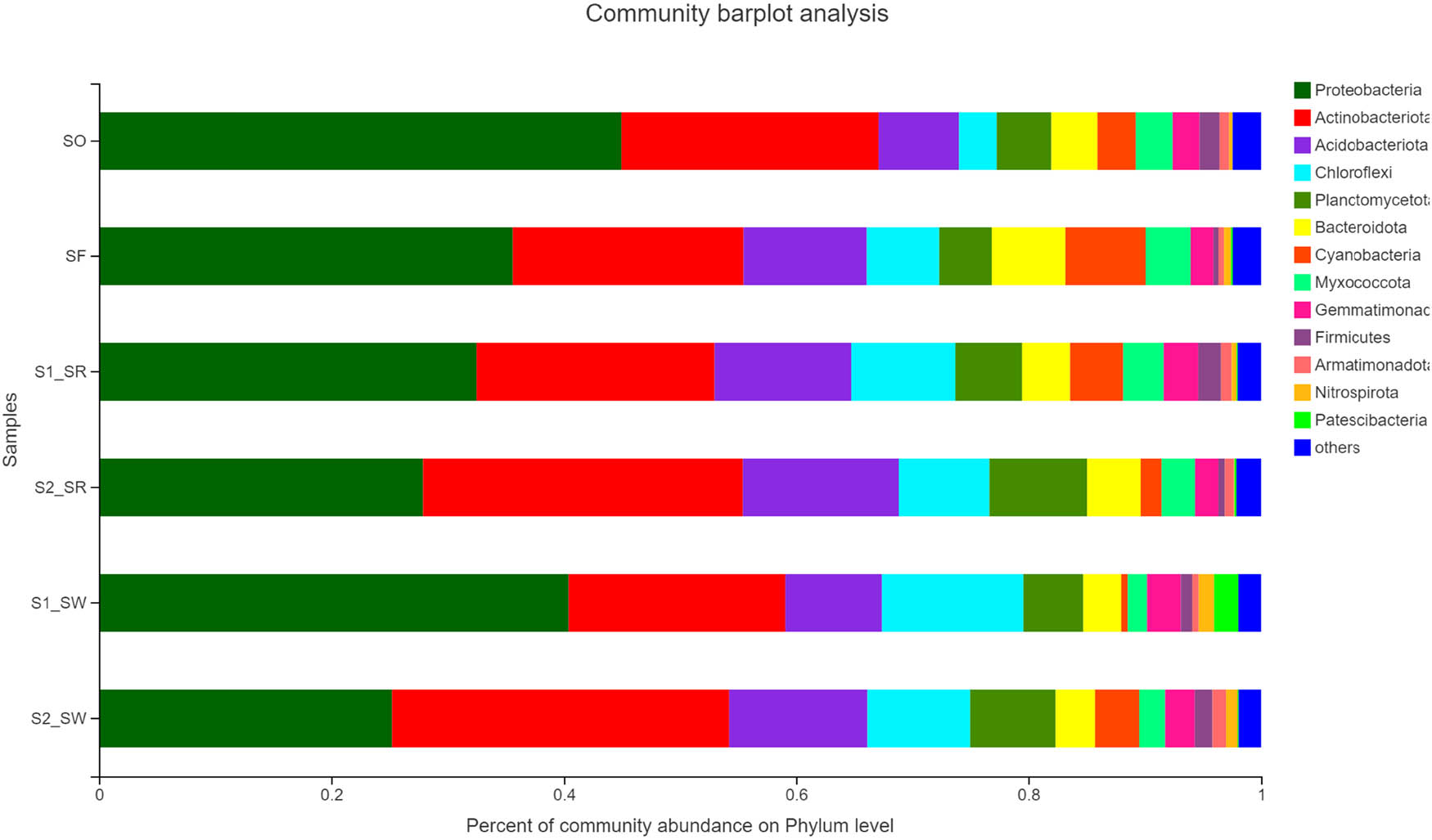
Relative abundance of soil bacterial community in six sampling sites (relative abundance > 0.1%) (Phylum level).
According to the analysis of the difference among soil groups (Figure 7), there are highly significant differences in Actinobacteria, Myxococcus, and Nitrospirota (P < 0.01). There were significant differences in the community structure among Chloroflexi, Acidobacteria, Firmicutes, and Proteobacteria (P < 0.05). In contrast, there are no significant differences in the community structure of Planctomycetota, Bacteroidota, Cyanobacteria, Gemmatimonadota, and Armatimonadota among the six soils. Second, the significant difference showed that Proteobacteria and Actinomycetes accounted for a large proportion of the bacterial community (P < 0.05). All results showed that the bacterial communities within soil samples at the same sampling point were similar, but the proportion of populations differed, which was supported by the results in Figure 6.
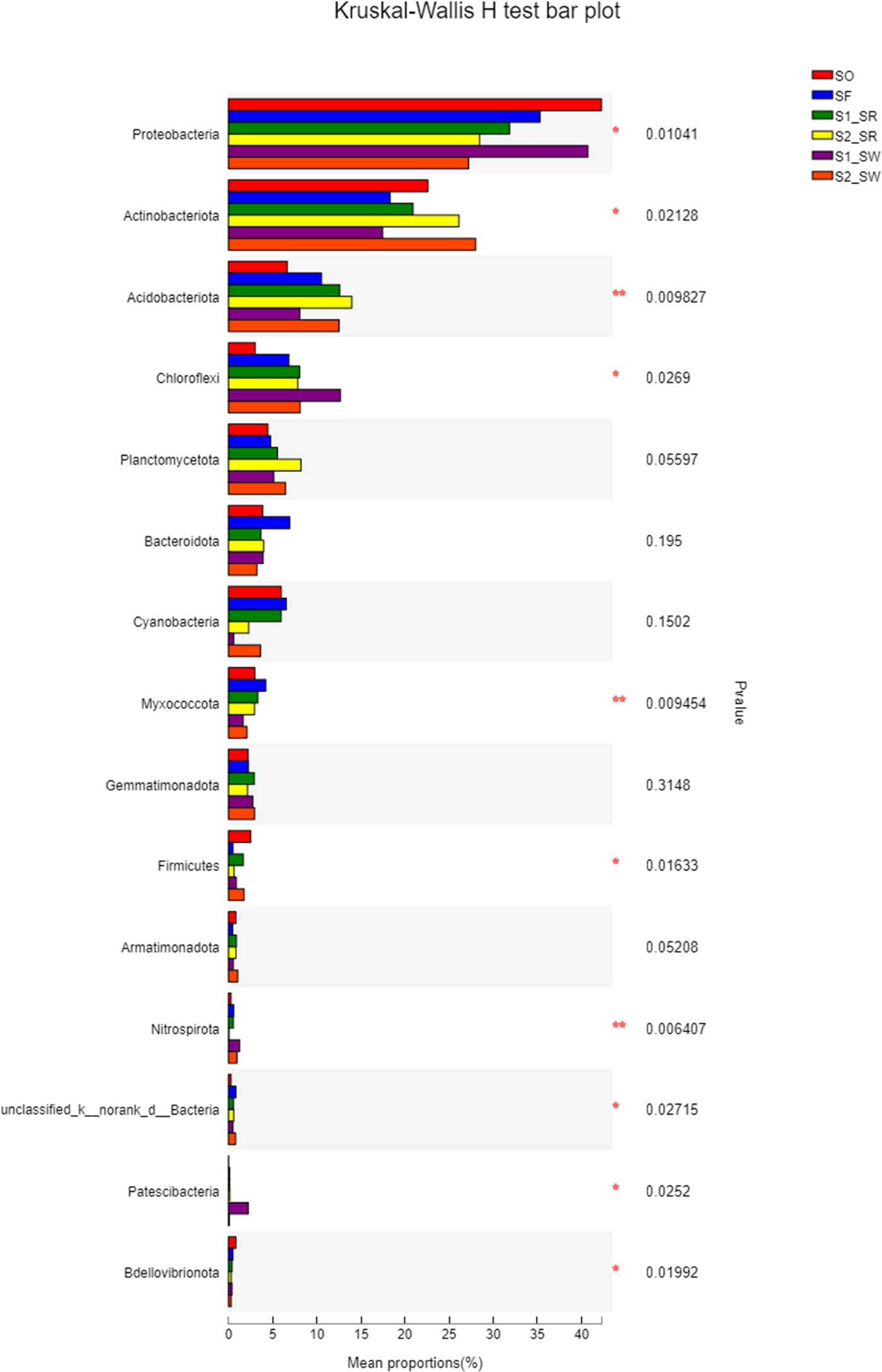
Analysis chart of significance test of difference between groups.
According to the Kruskal–Wallis H test, the number of asterisks in a row indicate the statistical significance between different soil samples (P < 0.05). One asterisk represents a significant difference (P < 0.05), and two asterisks represent an extremely significant difference (P < 0.01).
The Spearman correlation heat map showed that the soil bacterial communities in the six groups were different at the phylum level (Figure 8), which also supported the NMDS analysis (Figure 5b). The first 20 phyla were selected for multiple comparisons. At the phylum level, the abundance of Proteobacteria and Firmicutes in the S2_SR soil sample was the highest, and that of Chloroflexi was the lowest. The abundance of Gemmatimonadota and Firmicutes in SF soil samples was the lowest. The results are also supported by Figure 6. The significance test result between the groups found that the biomass of Gemmatimonadota and Firmicutes in non-polluted soil was lower than that in the heavily polluted soil. Therefore, it can be speculated that these two bacteria may not be subject to the stress of HMs, and the abundance of bacteria decreased in the soil containing high content of HMs.
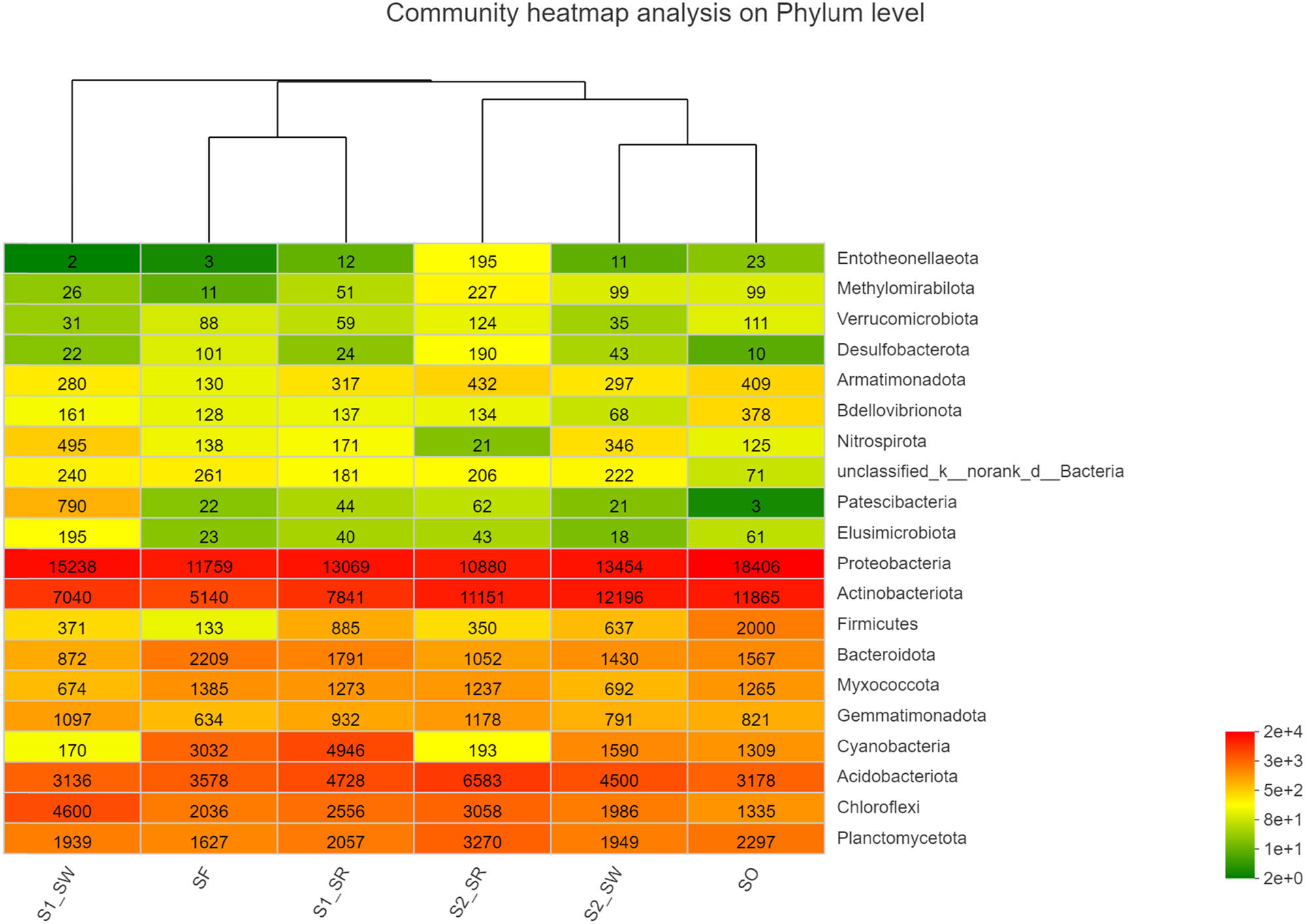
Heatmap of the relative abundance of bacteria community at phylum level in soil samples.
3.6 Relationship between bacterial community structure and environmental characteristics
To investigate the environmental characteristics affecting the bacterial communities, all the ten measured environmental variables were subjected to RDA (Figure 9). The RDA1 and RDA2 explained 37.89 and 15.41%, respectively, of the total variance of soil bacterial community composition at the phylum level (Figure 9a). Among these variables, HMs significantly influenced the bacterial communities, including Cu, Pb, Cd, and Zn (P < 0.05). There is a significant positive correlation among Cu, Pb, Cd, and Zn. There is a significant positive correlation between the four HMs (P < 0.05). The RDA1 and RDA2 explained 43.77 and 18.571%, respectively, of the total variances of soil bacterial community composition at the phylum level (Figure 9b). Among these variables, three types of nutrients significantly influenced the bacterial communities: TN, TP, and TK (P < 0.05). The most important environmental factors are Cu and TK. There is a significant positive correlation between the other nutrient element (except TK) factors (P < 0.05).
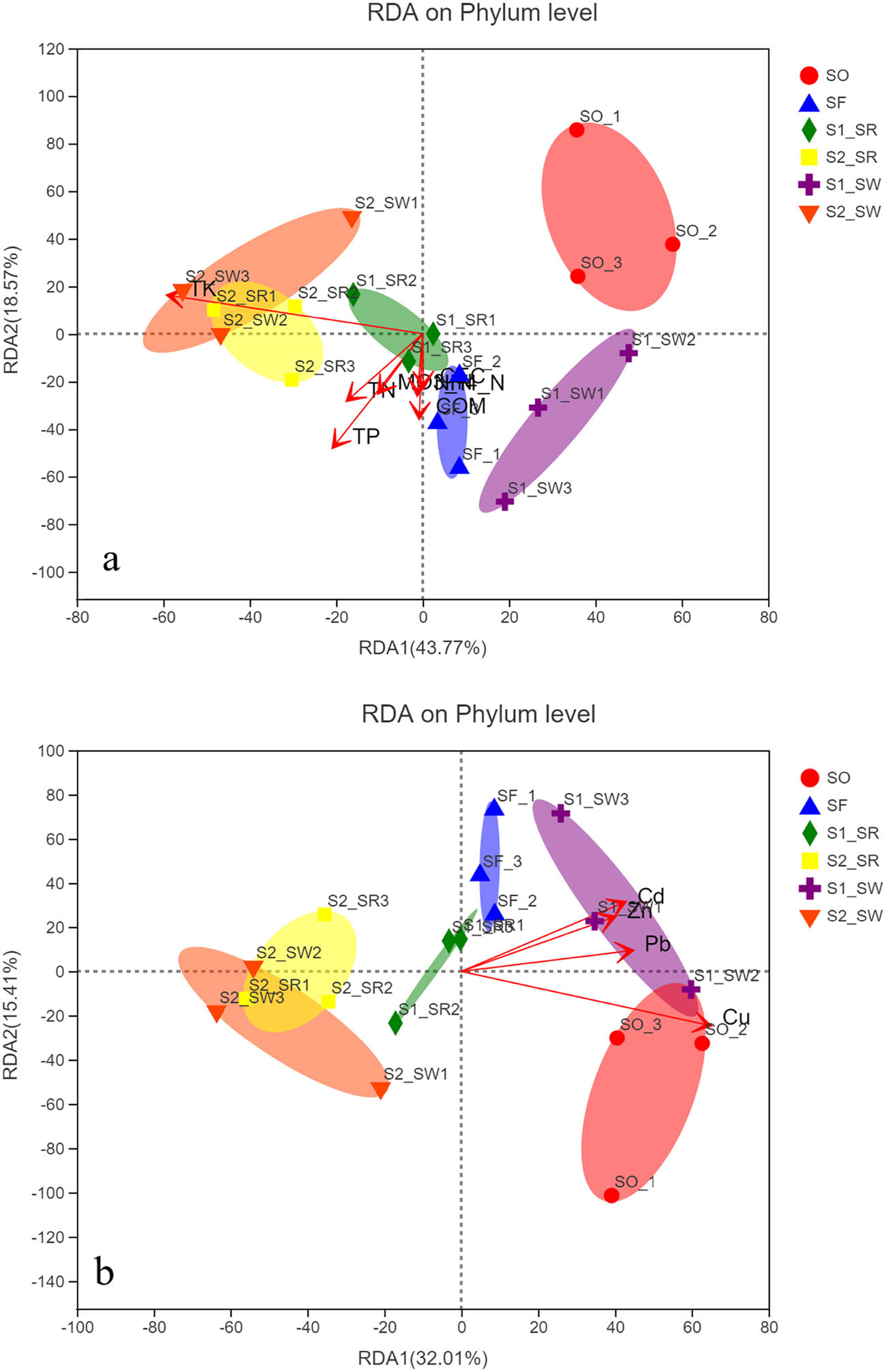
(a) RDA results of the soil physicochemical characteristics and the relative abundance of bacterial phyla and (b) RDA results of the soil HM factors and the relative abundance of bacterial phyla.
To analyze the β diversity, all soil sampling points were subjected to the Unweighted pair group method with arithmetic means (UPGMA) (Figure 10). According to a cluster of samples, the dominant OTUs showed a high similarity of bacterial communities in the soil groups between S1_SW and S2_SW. The same observation was noted for S1_SR and S2_SR, which suggested that the bacterial community structure of rhizosphere soil and non-rhizosphere soil were highly similar (P < 0.05). However, there were significant differences in bacterial community structure among SF and SO, and we can speculate that the large distance between the two soil groups is the reason for this phenomenon. The length of the branch among SF, S1, and S2 was short, indicating that the soil geographic position might have a significant influence on the bacterial community structure.
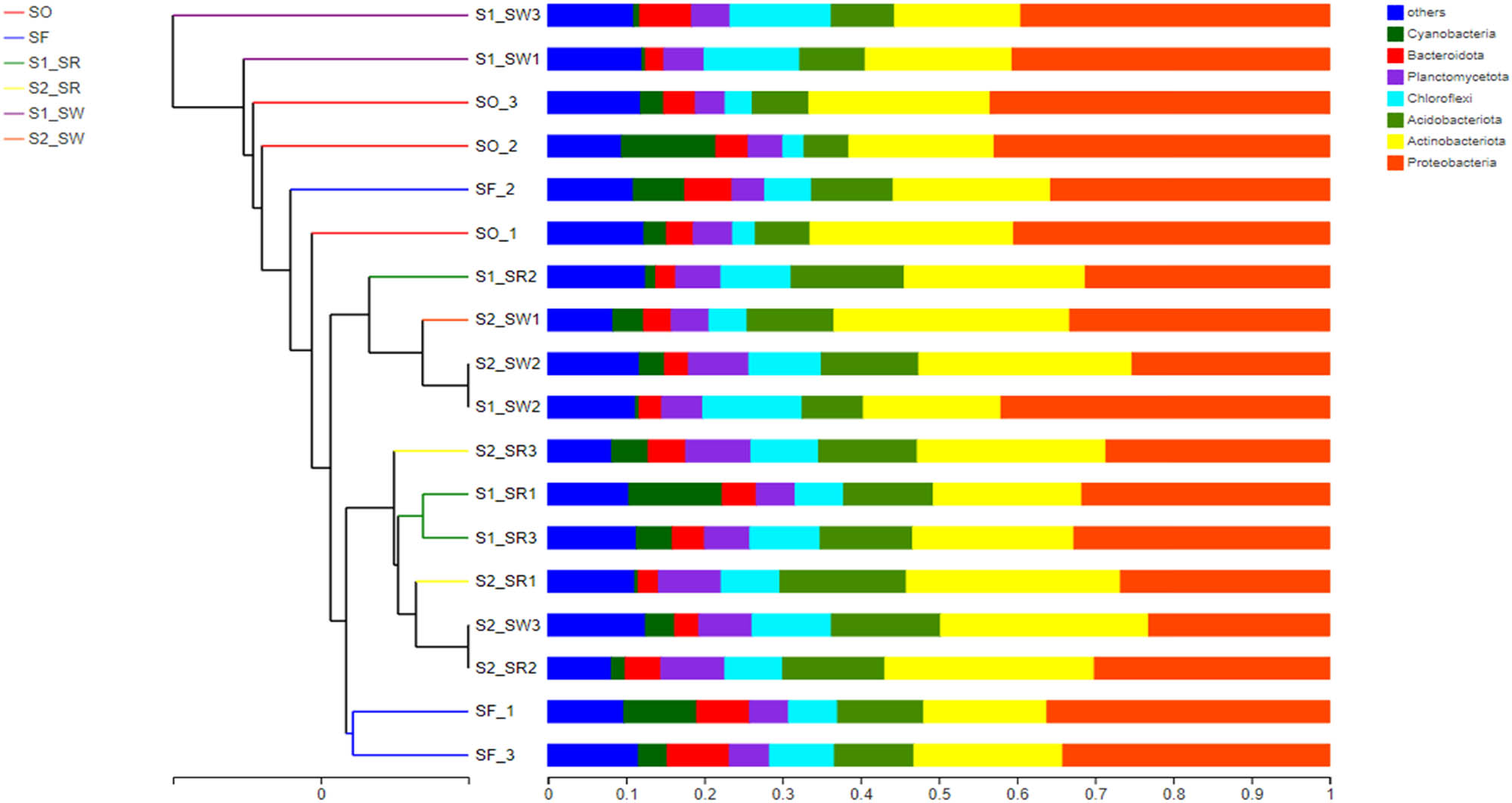
UPGMA clusters of different bacterial communities.
The correlation between dominant phyla bacteria and the physiochemical characteristics of soils is presented in Figure 11. The results showed that all eight variables, including COM, TN, TP, CEC, Cd, Zn, Cu, and Pb, had different effects on the structure of bacterial communities (P < 0.05). Soil bacterial richness was significantly correlated with the physical and chemical properties of the soil at the phylum level, including TN, TP, and TK. Strong positive correlation was found between HM and bacteria, which included Chloroflexi, Dependentiae, Elusimicrobiota, Gemmatimonadota, Nitrospirota, and Proteobacteria (P < 0.001, P < 0.01, or P < 0.05). A strong positive correlation was found between Nitrospirota and Zn and Cd (P < 0.001), indicating that Nitrospirota has a strong tolerance to the HMs. A strong negative correlation was found between HMs and bacteria, including Acidobacteriota, Actinobacteriota, Methylomirabilota, Cyanobacteria, Planctomycetota, and Verrucomicrobiota (P < 0.05 or P < 0.01). Myxococcota positively correlated with physicochemical factors, including COM, TN, TP, TK, NH4 +_N, NO3 −_N, and CEC (P < 0.05 or P < 0.01). In contrast, Proteobacteria and Elusimicrobiota, respectively, had negative correlation with physicochemical factors (P < 0.05 or P < 0.01).
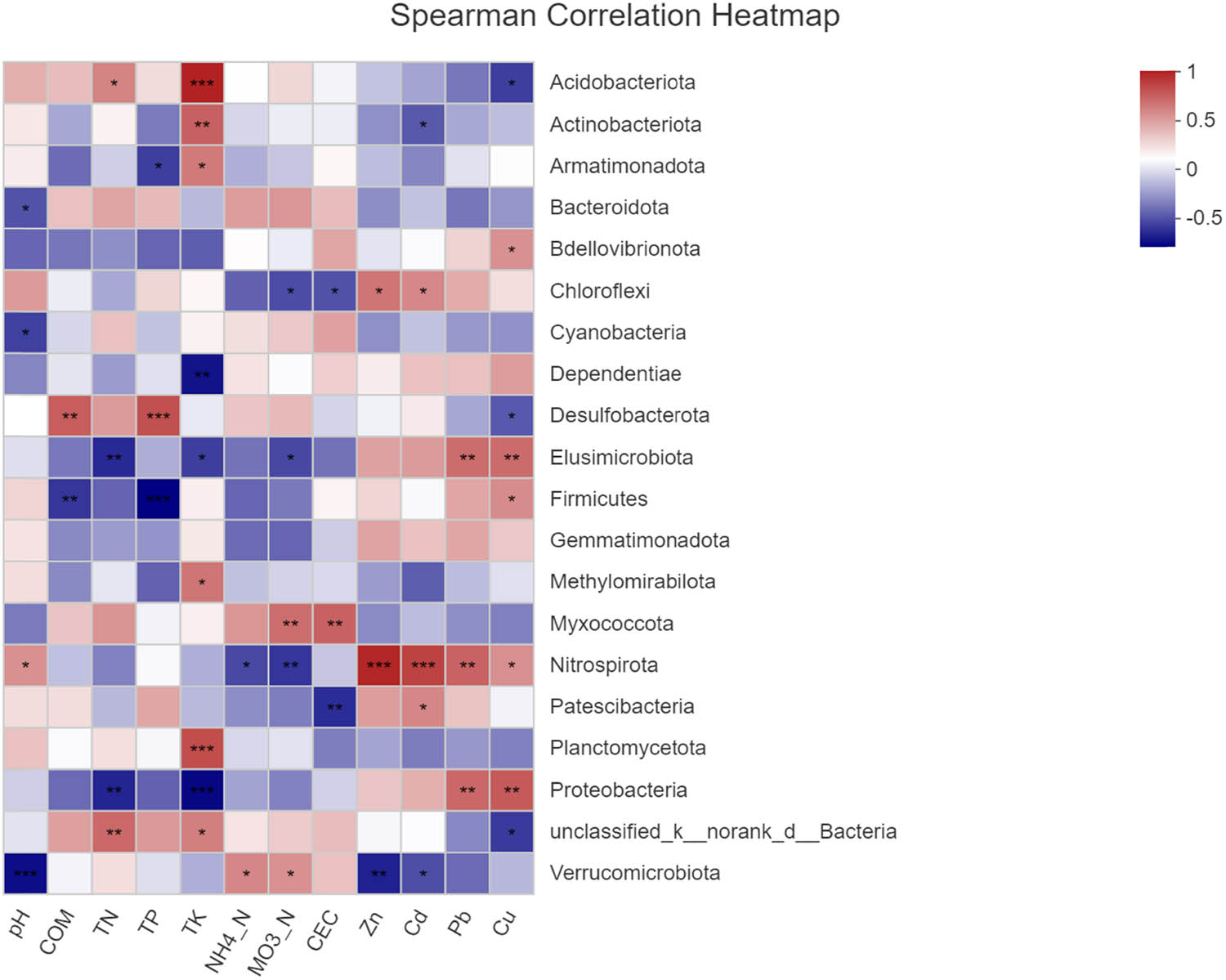
Heatmap analysis of the relationships between the fungal community composition at the phylum level and soil chemical properties. *P < 0.05, **P < 0.01, and ***P < 0.001.
4 Discussion
High-throughput sequencing was used to describe the diversity and community composition of bacteria in mining soil and evaluate the consequence of long-term pollution in mining soil on microbes. The results showed that HMs in soil had a significant positive effect on bacterial diversity, and there was no significant difference in the overall bacterial structure between the non-polluted and severely polluted soils. It is known that the pH value of tailings can be from highly acidic to highly alkaline (2–9), which depends on the influence of carbonate and other environmental factors in the process of soil mineral processing. Most of the research on tailing soil pH shows acidic conditions because of weathering, oxidation, and rain leaching, transforming most HMs from insoluble solid to soluble state and producing acidic substances. However, the soil pH in this study area is alkaline, which may be due to the climate type of desert steppe in the mainland and the soil salinization in Northwest China. This result is also supported by previous research reports [27,28]. Compared with the Chinese soil fertility standard, the content of soil nutrient elements, including CEC, TP, TK, TN, and COM, is generally low. It indicates that mining area soil is barren, and the lack of nutrients may affect the reproduction of soil microorganisms (including bacteria). S1_SW has the highest pollution level and the most serious pollution degree compared to other soil groups. These results indicate that HM does indeed cause serious soil concentration.
Soil environments usually have a stable microbial community. Soil pollution destroys the ecological balance, forcing the original microorganisms to adapt to the new environment, resulting in changes in microbial community structure and diversity [29,30,31]. Generally speaking, the dominant bacteria in the mining area include Proteobacteria, Actinobacillus, Acidobacteria, Bacteroides, and Nitrospirota [32,33]. Compared with S1_SW, the bacterial diversity (including Chao, Ace, and Shannon indices) was enhanced in S1_SR (Table 4). These can be explained by the abundant nutrients between S1_SR and S1_SW (Table 5). HM pollution not only reduced microbial biomass but also significantly changed the genetic diversity of the microbial community [34]. Chodak et al. proved a significant correlation between Chao index and Cd, Zn, and Pb [32]. Xu and Tang found that long-term exposure to HM polluted water environments has different effects on microbial diversity, and microorganisms gradually build a tolerance to metal [35].
Physical and chemical properties of soil in the mining area
| Index | SO | S1_SR | S1_SW | S2_SR | S2_SW | SF |
|---|---|---|---|---|---|---|
| COM | 7.80 ± 0.34c | 28.18 ± 0.31a | 8.30 ± 0.42c | 8.42 ± 0.59c | 12.0 ± 0.72b | 8.44 ± 0.68c |
| TN | 0.343 ± 0.014e | 1.772 ± 0.021a | 0.446 ± 0.031d | 0.859 ± 0.06f | 0.696 ± 0.049b | 0.731 ± 0.522c |
| TP | 0.281 ± 0.008e | 0.697 ± 0.002a | 0.426 ± 0.025d | 0.511 ± 0.041c | 0.462 ± 0.018b | 0.491 ± 0.132d |
| TK | 1.140 ± 0.02cd | 1.274 ± 0.016cd | 1.468 ± 0.059bc | 0.954 ± 0.067e | 1.844 ± 0.341ab | 1.397 ± 0.344a |
| NH4 +_N | 10.74 ± 0.06c | 21.3 ± 0.8a | 8.57 ± 0.425d | 13.46 ± 0.4d | 9.06 ± 0.77b | 11.93 ± 4.68d |
| NO3 −_N | 5.84 ± 0.182c | 13.99 ± 0.24a | 5.54 ± 0.22c | 4.25 ± 0.3 d | 9.23 ± 0.65b | 5.47 ± 0.44c |
| CEC | 2.95 ± 0.11b | 9.92 ± 0.18a | 2.69 ± 0.26bc | 0.8 ± 0.056 d | 2.52 ± 0.13c | 2.77 ± 0.167c |
Note: TN: total N (g/kg). COM: soil organic carbon (g/kg). TP: total P (g/kg). TK: total K (%). (NO3
−_N): nitrate N (mg/kg). (NH4
+_N): ammonium N (mg/kg). CEC: cation exchange capacity (
There are significant differences between communities of bacteria in HM polluted and non-polluted soils in this study. Proteobacteria was positively correlated with HMs Pb and Cu (P < 0.05), and Nitrospirota was positively correlated with four HMs (P < 0.05). This suggests that Proteobacteria and Nitrospirota have strong tolerance to HMs and can alleviate the hazardous toxicity from HMs via specialized mechanisms. Elusimicrobiota, Proteobacteria, and Verrucomicrobiota were negatively correlated with HM (P < 0.05), suggesting they have weak tolerance to HMs. Therefore, to mitigate pollution by HM in mining areas, we can speculate that the bacteria with strong HM tolerance become the dominant bacteria. Then, we can remediate HM pollution by using dominant bacteria.
The physical and chemical parameters of soil have a significant effect on the structure of bacterial communities. In this study, Proteobacteria was the dominant phylum in the six sampling sites, accounting for more than 30%. The diversity of Proteobacteria was positively correlated with HMs Pb and Cu (P < 0.05). As HM tolerant bacteria, Proteobacteria can survive in a variety of soil types (Karst soil, wetland, and mining soil) [33,36]. According to the comparison of bacterial diversity between rhizosphere soil and non-rhizosphere soil, it was found that the bacterial community structure was very similar, but the distribution and proportion of bacteria were different. It is speculated that the existence of plants may affect bacterial community distribution. However, the biomass of Acidobacteria in this study is not very high, which may be caused by the alkaline soil environment [28].
The soil’s physical and chemical properties significantly influence bacteria’s community structure and diversity, especially TN, TK, and TP. As important nutrient sources in soil, TP, TK, and TN play a vital role in bacteria’s metabolism, growth, and reproduction [37]. Microorganisms have the ability to adapt to fluctuating environmental changes and have an impact on the flow of energy and nutrients in the environment [38]. The soil in this study is from the mining area, and the soil environment was very complex [39]. The results showed that TN, TP, and TK significantly affected the bacterial community structure [40]. In our investigation, Myxococcota positively correlated with physicochemical factors, including COM, TN, TP, TK, NH4 +_N, NO3 −_N, and CEC (P < 0.05 or P < 0.01). In contrast, Proteobacteria and Elusimicrobiota, respectively, had negative correlation with physiochemical factor (P < 0.05 or P < 0.01). As important nutrient sources in soil, TN, TP, and TK play a key role in bacterial metabolism and reproduction. When the soil was polluted by HMs, more nutrients were mobilized by HM tolerant bacteria for transferring and metabolizing HMs, so TN, TP, and TK became the key environmental factors affecting the bacterial community structure [41,42].
In our present study, we analyzed the relationship of bacterial community structure and the mining area under severe pollution and non-pollution only. The bacterial community of the mining area with lower pollution was not specifically researched. In future work, further investigation should be undertaken to determine the relationship of bacterial community structure and diversity in different pollution levels. This will provide more scientific and reliable evidence for HM bioremediation of in-situ soil pollution.
-
Funding information: The research was funded by the National Natural Science Foundation of China (31860176 and 32160749) the Key Research and Development Program of Shanxi Province (2020ZDLSF06-06), the Key Research and Development Program of Gansu Province (20YF3FA037 and 20YF3FA043), and XAWLKYTD012.
-
Conflict of interest: Authors state no conflict of interest.
-
Data availability statement: The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
[1] Liu Y, Meng SR. Soil pollution: hidden reality. Ecol Econ. 2018;34(7):6–9.Suche in Google Scholar
[2] Gao TP, Wan ZD, Liu XX, Fu JW, Chang GH, Sun HL, et al. Effect of heavy metals on the salsola collina rhizosphere and bulk bacterial community in jinchuan mining area. Geomicrobiol J. 2021;38(1):1–11.10.1080/01490451.2021.1914784Suche in Google Scholar
[3] Xu ZX, Wu J, Li HZ, Chen YB, Xu J, Xiong LJ, et al. Characterizing heavy metals in combined sewer overflows and its influence on microbial diversity. Sci Total Envir. 2018;625:1272–82.10.1016/j.scitotenv.2017.12.338Suche in Google Scholar PubMed
[4] Li S, Wu JL, Huo YL, Zhao X, Xue LG. Profiling multiple heavy metal contamination and bacterial communities surrounding an iron tailing pond in Northwest China. Sci Total Envir. 2021;752:142827.10.1016/j.scitotenv.2020.141827Suche in Google Scholar PubMed
[5] Zhong QH, Wang YX, Ren SX, Zhou YT, Wu Y, Ying ML, et al. Speciation distribution and environment pollution assessment of heavy metals in sediments from wastewater pond of a pyrite mine. J Earth Environ. 2020;11(5):524–35.Suche in Google Scholar
[6] Xu W, Bai J, Shi ZM. Review on the environmental impact of mining in China and investigation methods. Acta Miner Sinica. 2013;33(S2):722–3.Suche in Google Scholar
[7] Du ZZ, Gao QH. Deep prospecting status and prospecting direction of Bijiashan lead-zinc deposit in Chengxian County. Gansu Metall. 2014;36(5):89–91.Suche in Google Scholar
[8] Ma Y, Luo YG, Teng Y, Li ZG. Plant growth promoting rhizobacteria and their role in phytoremediation of heavy metal contaminated soils. Acta Pedologica Sinica. 2013;50(5):1021–31.Suche in Google Scholar
[9] Snehlata BR. Role of rhizosphreic bacteria in phytoremediation of heavy metal contaminated soil. Inter J Cur Res. 2013;5(12):3897–907.Suche in Google Scholar
[10] Roane TM, Kellogg ST. Characterization of bacterial communities in heavy metal contaminated soils. Can J Microbiol. 1996;42(6):593–603.10.1139/m96-080Suche in Google Scholar PubMed
[11] Deng LJ, Zeng GG, Fan CZ, Lu LH, Chen XF, Chen M, et al. Response of rhizosphere microbial community structure and diversity to heavy metal co-pollution in arable soil. Appl Microbiol Biotechnol. 2015;99(19):8259–69.10.1007/s00253-015-6662-6Suche in Google Scholar PubMed
[12] Wang XH, Xu JM, Yao HY, Xie ZM. Effects of Cu, Zn, Cd and Pb compound contamination on soil microbial community. Cata Sci Cir. 2003;23(1):22–7.Suche in Google Scholar
[13] Griffiths BS, Philippot L. Insights into the resistance and resilience of the soil microbial community. FEMS Microbiol Rev. 2013;37(2):1–18.10.1111/j.1574-6976.2012.00343.xSuche in Google Scholar PubMed
[14] Hu H, Chen XJ, Hou FJ, Wu YP, Cheng YX. Bacterial and fungal community structures in loess plateau grasslands with different grazing intensities. Front Microbiol. 2017;8:606–16.10.3389/fmicb.2017.00606Suche in Google Scholar PubMed PubMed Central
[15] Annie JA, Abraham BM, Sridharan A. A critical re-examination of the factors influencing determination of organic matter in soils. Geotech Geol Eng. 2021;39:4287–93.10.1007/s10706-021-01758-3Suche in Google Scholar
[16] Bremner JM. Determination of nitrogen in soil by the Kjeldahl method. J Agr Sci. 1960;55(1):11–33.10.1017/S0021859600021572Suche in Google Scholar
[17] Edwards AH. The semi-micro Kjeldahl method for the determination of nitrogen in coal. J Appl Chem. 2007;4(6):330–4.10.1002/jctb.5010040610Suche in Google Scholar
[18] Nong BC. Improvement on determination of total phosphorus in soil by sodium hydroxide alkali fusion molybdenum antimony anti colorimetric. Guangxi For Sci. 1989;02:33–6.Suche in Google Scholar
[19] Tu ZH, Chen LH, Yu XX, Zheng YS. Rhizosphere soil enzymatic and microbial activities in bamboo forests in southeastern China. Soil Sci Plant Nutr. 2014;60(2):134–44.10.1080/00380768.2014.882219Suche in Google Scholar
[20] Wen YC, Li HY, Lin ZA, Zhao BQ, Sun ZB. Long-term fertilization alters soil properties and fungal community composition in fluvo-aquic soil of the North China Plain. Sci Re. 2020;10(1):7198–209.10.1038/s41598-020-64227-6Suche in Google Scholar
[21] Gu LM, Liu TN, Zhao J, Dong ST, Liu P, Zhang JW, et al. Nitrate leaching of winter wheat grown in lysimeters as affected by fertilizers and irrigation on the North China Plain. J Integr Agr. 2015;14(2):374–88.10.1016/S2095-3119(14)60747-4Suche in Google Scholar
[22] Jia SC, Xu YW. The characteristics and distribution of soil environment mechanical composition. Environ Monit. 1997;3:26–8.Suche in Google Scholar
[23] Kong ZY, Wu ZJ, Glick BR, He SY, Huang C, Wu L. Co-occurrence patterns of microbial communities affected by inoculants of plant growth-promoting bacteria during phytoremediation of heavy metal-contaminated soils. Ecotoxicol Environ Safety. 2019;183:109504.10.1016/j.ecoenv.2019.109504Suche in Google Scholar PubMed
[24] Wang P, Xu GC, Xu P. Spatiotemporal variation of dissolved oxygen and response characteristics of bacterial community in engineering recirculating aquaculture system of largemouth bass (Lateolabrax japonicus). Acta Hydrobiol Sinica. 2019;43(6):1290–9.Suche in Google Scholar
[25] Zhong Y, He L, Fu G, Xu YZ, Cheng YX, Zhao YY et al. Microbial community with the ability to biodegradation perchlorate in a bio-electrochemical reactor. The 4th International Conference on Energy and Environment Research Progress. China: Shenzhen; 2019.10.1051/e3sconf/201911801021Suche in Google Scholar
[26] Bourceret A, Cébron A, Tisserant E, Poupin P, Bauda P, Beguiristian T, et al. The bacterial and fungal diversity of an aged PAH- and heavy metal-contaminated soil is affected by plant cover and edaphic parameters. Micro Ecol. 2015;71(3):711–24.10.1007/s00248-015-0682-8Suche in Google Scholar PubMed
[27] Munshower FF. Practical handbook of disturbed land revegetation. Lewis Boca Rat Fl. 1994. p. 15–33.10.1201/9781351075923Suche in Google Scholar
[28] Li CH, Wu XF, Han JC, Guo YL. Research on pollution load evaluation model of heavy metals in kardt soil. Environ Pollut Control. 2021;43(1):91–3+102.Suche in Google Scholar
[29] Jurburg SD, Natal-Da-Luz T, Raimundo J, Morais PV, Sousa JP. Bacterial communities in soil become sensitive to drought under intensive grazing. Sci Total Environ. 2017;618:1638–46.10.1016/j.scitotenv.2017.10.012Suche in Google Scholar PubMed
[30] Philippot L, Raaijmakers JM, Lemanceau P, Putten WH. Going back to the roots: the microbial ecology of the rhizosphere. Nat Rev Microbiol. 2013;11(11):789–99.10.1038/nrmicro3109Suche in Google Scholar PubMed
[31] Jiang BH, Zhang B, Li L, Zhao Y, Shi Y, Jiang Q, et al. Analysis of microbial community structure and diversity in surrounding rock soil of different waste dump sites in Fushun western opencast mine. Chemosphere. 2021;269:128777.10.1016/j.chemosphere.2020.128777Suche in Google Scholar PubMed
[32] Chodak M, Gołębiewski M, Płoskonka MJ, Kuduk K, Niklińska M. Diversity of microorganisms from forest soils differently polluted with heavy metals. Appl Soil Ecol. 2013;64(Complete):7–14.10.1016/j.apsoil.2012.11.004Suche in Google Scholar
[33] Gao TP, Fu JW, Zuo MB, Liu YB, Xu DH, Chang GH, et al. Study on the Rhizosphere Soil Microbial Community Structure Associated with Five Land use Types in Jinchuan Mining Area.e3s Web of Conferences. 2021;237:01010. China: Internet; 2021. 10.051/e3sconf/202123701010.Suche in Google Scholar
[34] Zhao XW, Luo YM, Teng Y, Li ZG, Song J, Wu LH. Genetic diversity of microbial communities in farmland soils contaminated with mixed heavy metals. Acta Sci Cir. 2005;2:186–91.Suche in Google Scholar
[35] Xu S, Tang F. Study on soil environmental pollution and its remediation in mining area. Tech Innov and Appl. 2018;(21):81–2.Suche in Google Scholar
[36] Nie S. Studies on the relationship between heavy metals and soil physicochemical properties and wetland soil microbial biomass and bacterial community structure. (PhD thesis). China: Hunan university; 2016.Suche in Google Scholar
[37] Ikegami K, Hirose Y, Sakashita H, Maruyama R, Sugiyama T. Role of polyphenol in sugarcane molasses as a nutrient for hexavalent chromium bioremediation using bacteria. Chemosphere. 2020;250:126267.10.1016/j.chemosphere.2020.126267Suche in Google Scholar PubMed
[38] Schimel J, Balser TC, Wallenstein M. Microbial stress-response physiology and its implications for ecosystem function. Ecol. 2007;88(6):1386–94.10.1890/06-0219Suche in Google Scholar PubMed
[39] Lin YB, Ye YM, Wu CF, Hu YM, Shi HK. Response analysis of soil bacterial community to different heavy metal pollution levels in paddy fields: a case study of a county. Acta Sci Cir. 2020;40(1):224–33.Suche in Google Scholar
[40] Yu FM, Yao YW, Xie DY, Wang XR, Lin JM. Study on the soil microbial community structure associated with six land use in Siding mining area. China Environ Sci. 2020;v.40(5):409–16.Suche in Google Scholar
[41] Huang Q, Wang JM, Wang C, Wang Q. The 19-years inorganic fertilization increased bacterial diversity and altered bacterial community composition and potential functions in a paddy soil. Appl Soil Ecol. 2019;2019(144):60–7.10.1016/j.apsoil.2019.07.009Suche in Google Scholar
[42] Rajapaksha R. Heavy metal tolerance of culturable bacteria and fungi in a long-term cultivated tropical ultisol. Eur J Soil Biol. 2011;47(1):9–15.10.1016/j.ejsobi.2010.10.006Suche in Google Scholar
© 2022 Yuan Liu et al., published by De Gruyter
This work is licensed under the Creative Commons Attribution 4.0 International License.
Artikel in diesem Heft
- Biomedical Sciences
- Effects of direct oral anticoagulants dabigatran and rivaroxaban on the blood coagulation function in rabbits
- The mother of all battles: Viruses vs humans. Can humans avoid extinction in 50–100 years?
- Knockdown of G1P3 inhibits cell proliferation and enhances the cytotoxicity of dexamethasone in acute lymphoblastic leukemia
- LINC00665 regulates hepatocellular carcinoma by modulating mRNA via the m6A enzyme
- Association study of CLDN14 variations in patients with kidney stones
- Concanavalin A-induced autoimmune hepatitis model in mice: Mechanisms and future outlook
- Regulation of miR-30b in cancer development, apoptosis, and drug resistance
- Informatic analysis of the pulmonary microecology in non-cystic fibrosis bronchiectasis at three different stages
- Swimming attenuates tumor growth in CT-26 tumor-bearing mice and suppresses angiogenesis by mediating the HIF-1α/VEGFA pathway
- Characterization of intestinal microbiota and serum metabolites in patients with mild hepatic encephalopathy
- Functional conservation and divergence in plant-specific GRF gene family revealed by sequences and expression analysis
- Application of the FLP/LoxP-FRT recombination system to switch the eGFP expression in a model prokaryote
- Biomedical evaluation of antioxidant properties of lamb meat enriched with iodine and selenium
- Intravenous infusion of the exosomes derived from human umbilical cord mesenchymal stem cells enhance neurological recovery after traumatic brain injury via suppressing the NF-κB pathway
- Effect of dietary pattern on pregnant women with gestational diabetes mellitus and its clinical significance
- Potential regulatory mechanism of TNF-α/TNFR1/ANXA1 in glioma cells and its role in glioma cell proliferation
- Effect of the genetic mutant G71R in uridine diphosphate-glucuronosyltransferase 1A1 on the conjugation of bilirubin
- Quercetin inhibits cytotoxicity of PC12 cells induced by amyloid-beta 25–35 via stimulating estrogen receptor α, activating ERK1/2, and inhibiting apoptosis
- Nutrition intervention in the management of novel coronavirus pneumonia patients
- circ-CFH promotes the development of HCC by regulating cell proliferation, apoptosis, migration, invasion, and glycolysis through the miR-377-3p/RNF38 axis
- Bmi-1 directly upregulates glucose transporter 1 in human gastric adenocarcinoma
- Lacunar infarction aggravates the cognitive deficit in the elderly with white matter lesion
- Hydroxysafflor yellow A improved retinopathy via Nrf2/HO-1 pathway in rats
- Comparison of axon extension: PTFE versus PLA formed by a 3D printer
- Elevated IL-35 level and iTr35 subset increase the bacterial burden and lung lesions in Mycobacterium tuberculosis-infected mice
- A case report of CAT gene and HNF1β gene variations in a patient with early-onset diabetes
- Study on the mechanism of inhibiting patulin production by fengycin
- SOX4 promotes high-glucose-induced inflammation and angiogenesis of retinal endothelial cells by activating NF-κB signaling pathway
- Relationship between blood clots and COVID-19 vaccines: A literature review
- Analysis of genetic characteristics of 436 children with dysplasia and detailed analysis of rare karyotype
- Bioinformatics network analyses of growth differentiation factor 11
- NR4A1 inhibits the epithelial–mesenchymal transition of hepatic stellate cells: Involvement of TGF-β–Smad2/3/4–ZEB signaling
- Expression of Zeb1 in the differentiation of mouse embryonic stem cell
- Study on the genetic damage caused by cadmium sulfide quantum dots in human lymphocytes
- Association between single-nucleotide polymorphisms of NKX2.5 and congenital heart disease in Chinese population: A meta-analysis
- Assessment of the anesthetic effect of modified pentothal sodium solution on Sprague-Dawley rats
- Genetic susceptibility to high myopia in Han Chinese population
- Potential biomarkers and molecular mechanisms in preeclampsia progression
- Silencing circular RNA-friend leukemia virus integration 1 restrained malignancy of CC cells and oxaliplatin resistance by disturbing dyskeratosis congenita 1
- Endostar plus pembrolizumab combined with a platinum-based dual chemotherapy regime for advanced pulmonary large-cell neuroendocrine carcinoma as a first-line treatment: A case report
- The significance of PAK4 in signaling and clinicopathology: A review
- Sorafenib inhibits ovarian cancer cell proliferation and mobility and induces radiosensitivity by targeting the tumor cell epithelial–mesenchymal transition
- Characterization of rabbit polyclonal antibody against camel recombinant nanobodies
- Active legumain promotes invasion and migration of neuroblastoma by regulating epithelial-mesenchymal transition
- Effect of cell receptors in the pathogenesis of osteoarthritis: Current insights
- MT-12 inhibits the proliferation of bladder cells in vitro and in vivo by enhancing autophagy through mitochondrial dysfunction
- Study of hsa_circRNA_000121 and hsa_circRNA_004183 in papillary thyroid microcarcinoma
- BuyangHuanwu Decoction attenuates cerebral vasospasm caused by subarachnoid hemorrhage in rats via PI3K/AKT/eNOS axis
- Effects of the interaction of Notch and TLR4 pathways on inflammation and heart function in septic heart
- Monosodium iodoacetate-induced subchondral bone microstructure and inflammatory changes in an animal model of osteoarthritis
- A rare presentation of type II Abernethy malformation and nephrotic syndrome: Case report and review
- Rapid death due to pulmonary epithelioid haemangioendothelioma in several weeks: A case report
- Hepatoprotective role of peroxisome proliferator-activated receptor-α in non-cancerous hepatic tissues following transcatheter arterial embolization
- Correlation between peripheral blood lymphocyte subpopulations and primary systemic lupus erythematosus
- A novel SLC8A1-ALK fusion in lung adenocarcinoma confers sensitivity to alectinib: A case report
- β-Hydroxybutyrate upregulates FGF21 expression through inhibition of histone deacetylases in hepatocytes
- Identification of metabolic genes for the prediction of prognosis and tumor microenvironment infiltration in early-stage non-small cell lung cancer
- BTBD10 inhibits glioma tumorigenesis by downregulating cyclin D1 and p-Akt
- Mucormycosis co-infection in COVID-19 patients: An update
- Metagenomic next-generation sequencing in diagnosing Pneumocystis jirovecii pneumonia: A case report
- Long non-coding RNA HOXB-AS1 is a prognostic marker and promotes hepatocellular carcinoma cells’ proliferation and invasion
- Preparation and evaluation of LA-PEG-SPION, a targeted MRI contrast agent for liver cancer
- Proteomic analysis of the liver regulating lipid metabolism in Chaohu ducks using two-dimensional electrophoresis
- Nasopharyngeal tuberculosis: A case report
- Characterization and evaluation of anti-Salmonella enteritidis activity of indigenous probiotic lactobacilli in mice
- Aberrant pulmonary immune response of obese mice to periodontal infection
- Bacteriospermia – A formidable player in male subfertility
- In silico and in vivo analysis of TIPE1 expression in diffuse large B cell lymphoma
- Effects of KCa channels on biological behavior of trophoblasts
- Interleukin-17A influences the vulnerability rather than the size of established atherosclerotic plaques in apolipoprotein E-deficient mice
- Multiple organ failure and death caused by Staphylococcus aureus hip infection: A case report
- Prognostic signature related to the immune environment of oral squamous cell carcinoma
- Primary and metastatic squamous cell carcinoma of the thyroid gland: Two case reports
- Neuroprotective effects of crocin and crocin-loaded niosomes against the paraquat-induced oxidative brain damage in rats
- Role of MMP-2 and CD147 in kidney fibrosis
- Geometric basis of action potential of skeletal muscle cells and neurons
- Babesia microti-induced fulminant sepsis in an immunocompromised host: A case report and the case-specific literature review
- Role of cerebellar cortex in associative learning and memory in guinea pigs
- Application of metagenomic next-generation sequencing technique for diagnosing a specific case of necrotizing meningoencephalitis caused by human herpesvirus 2
- Case report: Quadruple primary malignant neoplasms including esophageal, ureteral, and lung in an elderly male
- Long non-coding RNA NEAT1 promotes angiogenesis in hepatoma carcinoma via the miR-125a-5p/VEGF pathway
- Osteogenic differentiation of periodontal membrane stem cells in inflammatory environments
- Knockdown of SHMT2 enhances the sensitivity of gastric cancer cells to radiotherapy through the Wnt/β-catenin pathway
- Continuous renal replacement therapy combined with double filtration plasmapheresis in the treatment of severe lupus complicated by serious bacterial infections in children: A case report
- Simultaneous triple primary malignancies, including bladder cancer, lymphoma, and lung cancer, in an elderly male: A case report
- Preclinical immunogenicity assessment of a cell-based inactivated whole-virion H5N1 influenza vaccine
- One case of iodine-125 therapy – A new minimally invasive treatment of intrahepatic cholangiocarcinoma
- S1P promotes corneal trigeminal neuron differentiation and corneal nerve repair via upregulating nerve growth factor expression in a mouse model
- Early cancer detection by a targeted methylation assay of circulating tumor DNA in plasma
- Calcifying nanoparticles initiate the calcification process of mesenchymal stem cells in vitro through the activation of the TGF-β1/Smad signaling pathway and promote the decay of echinococcosis
- Evaluation of prognostic markers in patients infected with SARS-CoV-2
- N6-Methyladenosine-related alternative splicing events play a role in bladder cancer
- Characterization of the structural, oxidative, and immunological features of testis tissue from Zucker diabetic fatty rats
- Effects of glucose and osmotic pressure on the proliferation and cell cycle of human chorionic trophoblast cells
- Investigation of genotype diversity of 7,804 norovirus sequences in humans and animals of China
- Characteristics and karyotype analysis of a patient with turner syndrome complicated with multiple-site tumors: A case report
- Aggravated renal fibrosis is positively associated with the activation of HMGB1-TLR2/4 signaling in STZ-induced diabetic mice
- Distribution characteristics of SARS-CoV-2 IgM/IgG in false-positive results detected by chemiluminescent immunoassay
- SRPX2 attenuated oxygen–glucose deprivation and reperfusion-induced injury in cardiomyocytes via alleviating endoplasmic reticulum stress-induced apoptosis through targeting PI3K/Akt/mTOR axis
- Aquaporin-8 overexpression is involved in vascular structure and function changes in placentas of gestational diabetes mellitus patients
- Relationship between CRP gene polymorphisms and ischemic stroke risk: A systematic review and meta-analysis
- Effects of growth hormone on lipid metabolism and sexual development in pubertal obese male rats
- Cloning and identification of the CTLA-4IgV gene and functional application of vaccine in Xinjiang sheep
- Antitumor activity of RUNX3: Upregulation of E-cadherin and downregulation of the epithelial–mesenchymal transition in clear-cell renal cell carcinoma
- PHF8 promotes osteogenic differentiation of BMSCs in old rat with osteoporosis by regulating Wnt/β-catenin pathway
- A review of the current state of the computer-aided diagnosis (CAD) systems for breast cancer diagnosis
- Bilateral dacryoadenitis in adult-onset Still’s disease: A case report
- A novel association between Bmi-1 protein expression and the SUVmax obtained by 18F-FDG PET/CT in patients with gastric adenocarcinoma
- The role of erythrocytes and erythroid progenitor cells in tumors
- Relationship between platelet activation markers and spontaneous abortion: A meta-analysis
- Abnormal methylation caused by folic acid deficiency in neural tube defects
- Silencing TLR4 using an ultrasound-targeted microbubble destruction-based shRNA system reduces ischemia-induced seizures in hyperglycemic rats
- Plant Sciences
- Seasonal succession of bacterial communities in cultured Caulerpa lentillifera detected by high-throughput sequencing
- Cloning and prokaryotic expression of WRKY48 from Caragana intermedia
- Novel Brassica hybrids with different resistance to Leptosphaeria maculans reveal unbalanced rDNA signal patterns
- Application of exogenous auxin and gibberellin regulates the bolting of lettuce (Lactuca sativa L.)
- Phytoremediation of pollutants from wastewater: A concise review
- Genome-wide identification and characterization of NBS-encoding genes in the sweet potato wild ancestor Ipomoea trifida (H.B.K.)
- Alleviative effects of magnetic Fe3O4 nanoparticles on the physiological toxicity of 3-nitrophenol to rice (Oryza sativa L.) seedlings
- Selection and functional identification of Dof genes expressed in response to nitrogen in Populus simonii × Populus nigra
- Study on pecan seed germination influenced by seed endocarp
- Identification of active compounds in Ophiopogonis Radix from different geographical origins by UPLC-Q/TOF-MS combined with GC-MS approaches
- The entire chloroplast genome sequence of Asparagus cochinchinensis and genetic comparison to Asparagus species
- Genome-wide identification of MAPK family genes and their response to abiotic stresses in tea plant (Camellia sinensis)
- Selection and validation of reference genes for RT-qPCR analysis of different organs at various development stages in Caragana intermedia
- Cloning and expression analysis of SERK1 gene in Diospyros lotus
- Integrated metabolomic and transcriptomic profiling revealed coping mechanisms of the edible and medicinal homologous plant Plantago asiatica L. cadmium resistance
- A missense variant in NCF1 is associated with susceptibility to unexplained recurrent spontaneous abortion
- Assessment of drought tolerance indices in faba bean genotypes under different irrigation regimes
- The entire chloroplast genome sequence of Asparagus setaceus (Kunth) Jessop: Genome structure, gene composition, and phylogenetic analysis in Asparagaceae
- Food Science
- Dietary food additive monosodium glutamate with or without high-lipid diet induces spleen anomaly: A mechanistic approach on rat model
- Binge eating disorder during COVID-19
- Potential of honey against the onset of autoimmune diabetes and its associated nephropathy, pancreatitis, and retinopathy in type 1 diabetic animal model
- FTO gene expression in diet-induced obesity is downregulated by Solanum fruit supplementation
- Physical activity enhances fecal lactobacilli in rats chronically drinking sweetened cola beverage
- Supercritical CO2 extraction, chemical composition, and antioxidant effects of Coreopsis tinctoria Nutt. oleoresin
- Functional constituents of plant-based foods boost immunity against acute and chronic disorders
- Effect of selenium and methods of protein extraction on the proteomic profile of Saccharomyces yeast
- Microbial diversity of milk ghee in southern Gansu and its effect on the formation of ghee flavor compounds
- Ecology and Environmental Sciences
- Effects of heavy metals on bacterial community surrounding Bijiashan mining area located in northwest China
- Microorganism community composition analysis coupling with 15N tracer experiments reveals the nitrification rate and N2O emissions in low pH soils in Southern China
- Genetic diversity and population structure of Cinnamomum balansae Lecomte inferred by microsatellites
- Preliminary screening of microplastic contamination in different marine fish species of Taif market, Saudi Arabia
- Plant volatile organic compounds attractive to Lygus pratensis
- Effects of organic materials on soil bacterial community structure in long-term continuous cropping of tomato in greenhouse
- Effects of soil treated fungicide fluopimomide on tomato (Solanum lycopersicum L.) disease control and plant growth
- Prevalence of Yersinia pestis among rodents captured in a semi-arid tropical ecosystem of south-western Zimbabwe
- Effects of irrigation and nitrogen fertilization on mitigating salt-induced Na+ toxicity and sustaining sea rice growth
- Bioengineering and Biotechnology
- Poly-l-lysine-caused cell adhesion induces pyroptosis in THP-1 monocytes
- Development of alkaline phosphatase-scFv and its use for one-step enzyme-linked immunosorbent assay for His-tagged protein detection
- Development and validation of a predictive model for immune-related genes in patients with tongue squamous cell carcinoma
- Agriculture
- Effects of chemical-based fertilizer replacement with biochar-based fertilizer on albic soil nutrient content and maize yield
- Genome-wide identification and expression analysis of CPP-like gene family in Triticum aestivum L. under different hormone and stress conditions
- Agronomic and economic performance of mung bean (Vigna radiata L.) varieties in response to rates of blended NPS fertilizer in Kindo Koysha district, Southern Ethiopia
- Influence of furrow irrigation regime on the yield and water consumption indicators of winter wheat based on a multi-level fuzzy comprehensive evaluation
- Discovery of exercise-related genes and pathway analysis based on comparative genomes of Mongolian originated Abaga and Wushen horse
- Lessons from integrated seasonal forecast-crop modelling in Africa: A systematic review
- Evolution trend of soil fertility in tobacco-planting area of Chenzhou, Hunan Province, China
- Animal Sciences
- Morphological and molecular characterization of Tatera indica Hardwicke 1807 (Rodentia: Muridae) from Pothwar, Pakistan
- Research on meat quality of Qianhua Mutton Merino sheep and Small-tail Han sheep
- SI: A Scientific Memoir
- Suggestions on leading an academic research laboratory group
- My scientific genealogy and the Toronto ACDC Laboratory, 1988–2022
- Erratum
- Erratum to “Changes of immune cells in patients with hepatocellular carcinoma treated by radiofrequency ablation and hepatectomy, a pilot study”
- Erratum to “A two-microRNA signature predicts the progression of male thyroid cancer”
- Retraction
- Retraction of “Lidocaine has antitumor effect on hepatocellular carcinoma via the circ_DYNC1H1/miR-520a-3p/USP14 axis”
Artikel in diesem Heft
- Biomedical Sciences
- Effects of direct oral anticoagulants dabigatran and rivaroxaban on the blood coagulation function in rabbits
- The mother of all battles: Viruses vs humans. Can humans avoid extinction in 50–100 years?
- Knockdown of G1P3 inhibits cell proliferation and enhances the cytotoxicity of dexamethasone in acute lymphoblastic leukemia
- LINC00665 regulates hepatocellular carcinoma by modulating mRNA via the m6A enzyme
- Association study of CLDN14 variations in patients with kidney stones
- Concanavalin A-induced autoimmune hepatitis model in mice: Mechanisms and future outlook
- Regulation of miR-30b in cancer development, apoptosis, and drug resistance
- Informatic analysis of the pulmonary microecology in non-cystic fibrosis bronchiectasis at three different stages
- Swimming attenuates tumor growth in CT-26 tumor-bearing mice and suppresses angiogenesis by mediating the HIF-1α/VEGFA pathway
- Characterization of intestinal microbiota and serum metabolites in patients with mild hepatic encephalopathy
- Functional conservation and divergence in plant-specific GRF gene family revealed by sequences and expression analysis
- Application of the FLP/LoxP-FRT recombination system to switch the eGFP expression in a model prokaryote
- Biomedical evaluation of antioxidant properties of lamb meat enriched with iodine and selenium
- Intravenous infusion of the exosomes derived from human umbilical cord mesenchymal stem cells enhance neurological recovery after traumatic brain injury via suppressing the NF-κB pathway
- Effect of dietary pattern on pregnant women with gestational diabetes mellitus and its clinical significance
- Potential regulatory mechanism of TNF-α/TNFR1/ANXA1 in glioma cells and its role in glioma cell proliferation
- Effect of the genetic mutant G71R in uridine diphosphate-glucuronosyltransferase 1A1 on the conjugation of bilirubin
- Quercetin inhibits cytotoxicity of PC12 cells induced by amyloid-beta 25–35 via stimulating estrogen receptor α, activating ERK1/2, and inhibiting apoptosis
- Nutrition intervention in the management of novel coronavirus pneumonia patients
- circ-CFH promotes the development of HCC by regulating cell proliferation, apoptosis, migration, invasion, and glycolysis through the miR-377-3p/RNF38 axis
- Bmi-1 directly upregulates glucose transporter 1 in human gastric adenocarcinoma
- Lacunar infarction aggravates the cognitive deficit in the elderly with white matter lesion
- Hydroxysafflor yellow A improved retinopathy via Nrf2/HO-1 pathway in rats
- Comparison of axon extension: PTFE versus PLA formed by a 3D printer
- Elevated IL-35 level and iTr35 subset increase the bacterial burden and lung lesions in Mycobacterium tuberculosis-infected mice
- A case report of CAT gene and HNF1β gene variations in a patient with early-onset diabetes
- Study on the mechanism of inhibiting patulin production by fengycin
- SOX4 promotes high-glucose-induced inflammation and angiogenesis of retinal endothelial cells by activating NF-κB signaling pathway
- Relationship between blood clots and COVID-19 vaccines: A literature review
- Analysis of genetic characteristics of 436 children with dysplasia and detailed analysis of rare karyotype
- Bioinformatics network analyses of growth differentiation factor 11
- NR4A1 inhibits the epithelial–mesenchymal transition of hepatic stellate cells: Involvement of TGF-β–Smad2/3/4–ZEB signaling
- Expression of Zeb1 in the differentiation of mouse embryonic stem cell
- Study on the genetic damage caused by cadmium sulfide quantum dots in human lymphocytes
- Association between single-nucleotide polymorphisms of NKX2.5 and congenital heart disease in Chinese population: A meta-analysis
- Assessment of the anesthetic effect of modified pentothal sodium solution on Sprague-Dawley rats
- Genetic susceptibility to high myopia in Han Chinese population
- Potential biomarkers and molecular mechanisms in preeclampsia progression
- Silencing circular RNA-friend leukemia virus integration 1 restrained malignancy of CC cells and oxaliplatin resistance by disturbing dyskeratosis congenita 1
- Endostar plus pembrolizumab combined with a platinum-based dual chemotherapy regime for advanced pulmonary large-cell neuroendocrine carcinoma as a first-line treatment: A case report
- The significance of PAK4 in signaling and clinicopathology: A review
- Sorafenib inhibits ovarian cancer cell proliferation and mobility and induces radiosensitivity by targeting the tumor cell epithelial–mesenchymal transition
- Characterization of rabbit polyclonal antibody against camel recombinant nanobodies
- Active legumain promotes invasion and migration of neuroblastoma by regulating epithelial-mesenchymal transition
- Effect of cell receptors in the pathogenesis of osteoarthritis: Current insights
- MT-12 inhibits the proliferation of bladder cells in vitro and in vivo by enhancing autophagy through mitochondrial dysfunction
- Study of hsa_circRNA_000121 and hsa_circRNA_004183 in papillary thyroid microcarcinoma
- BuyangHuanwu Decoction attenuates cerebral vasospasm caused by subarachnoid hemorrhage in rats via PI3K/AKT/eNOS axis
- Effects of the interaction of Notch and TLR4 pathways on inflammation and heart function in septic heart
- Monosodium iodoacetate-induced subchondral bone microstructure and inflammatory changes in an animal model of osteoarthritis
- A rare presentation of type II Abernethy malformation and nephrotic syndrome: Case report and review
- Rapid death due to pulmonary epithelioid haemangioendothelioma in several weeks: A case report
- Hepatoprotective role of peroxisome proliferator-activated receptor-α in non-cancerous hepatic tissues following transcatheter arterial embolization
- Correlation between peripheral blood lymphocyte subpopulations and primary systemic lupus erythematosus
- A novel SLC8A1-ALK fusion in lung adenocarcinoma confers sensitivity to alectinib: A case report
- β-Hydroxybutyrate upregulates FGF21 expression through inhibition of histone deacetylases in hepatocytes
- Identification of metabolic genes for the prediction of prognosis and tumor microenvironment infiltration in early-stage non-small cell lung cancer
- BTBD10 inhibits glioma tumorigenesis by downregulating cyclin D1 and p-Akt
- Mucormycosis co-infection in COVID-19 patients: An update
- Metagenomic next-generation sequencing in diagnosing Pneumocystis jirovecii pneumonia: A case report
- Long non-coding RNA HOXB-AS1 is a prognostic marker and promotes hepatocellular carcinoma cells’ proliferation and invasion
- Preparation and evaluation of LA-PEG-SPION, a targeted MRI contrast agent for liver cancer
- Proteomic analysis of the liver regulating lipid metabolism in Chaohu ducks using two-dimensional electrophoresis
- Nasopharyngeal tuberculosis: A case report
- Characterization and evaluation of anti-Salmonella enteritidis activity of indigenous probiotic lactobacilli in mice
- Aberrant pulmonary immune response of obese mice to periodontal infection
- Bacteriospermia – A formidable player in male subfertility
- In silico and in vivo analysis of TIPE1 expression in diffuse large B cell lymphoma
- Effects of KCa channels on biological behavior of trophoblasts
- Interleukin-17A influences the vulnerability rather than the size of established atherosclerotic plaques in apolipoprotein E-deficient mice
- Multiple organ failure and death caused by Staphylococcus aureus hip infection: A case report
- Prognostic signature related to the immune environment of oral squamous cell carcinoma
- Primary and metastatic squamous cell carcinoma of the thyroid gland: Two case reports
- Neuroprotective effects of crocin and crocin-loaded niosomes against the paraquat-induced oxidative brain damage in rats
- Role of MMP-2 and CD147 in kidney fibrosis
- Geometric basis of action potential of skeletal muscle cells and neurons
- Babesia microti-induced fulminant sepsis in an immunocompromised host: A case report and the case-specific literature review
- Role of cerebellar cortex in associative learning and memory in guinea pigs
- Application of metagenomic next-generation sequencing technique for diagnosing a specific case of necrotizing meningoencephalitis caused by human herpesvirus 2
- Case report: Quadruple primary malignant neoplasms including esophageal, ureteral, and lung in an elderly male
- Long non-coding RNA NEAT1 promotes angiogenesis in hepatoma carcinoma via the miR-125a-5p/VEGF pathway
- Osteogenic differentiation of periodontal membrane stem cells in inflammatory environments
- Knockdown of SHMT2 enhances the sensitivity of gastric cancer cells to radiotherapy through the Wnt/β-catenin pathway
- Continuous renal replacement therapy combined with double filtration plasmapheresis in the treatment of severe lupus complicated by serious bacterial infections in children: A case report
- Simultaneous triple primary malignancies, including bladder cancer, lymphoma, and lung cancer, in an elderly male: A case report
- Preclinical immunogenicity assessment of a cell-based inactivated whole-virion H5N1 influenza vaccine
- One case of iodine-125 therapy – A new minimally invasive treatment of intrahepatic cholangiocarcinoma
- S1P promotes corneal trigeminal neuron differentiation and corneal nerve repair via upregulating nerve growth factor expression in a mouse model
- Early cancer detection by a targeted methylation assay of circulating tumor DNA in plasma
- Calcifying nanoparticles initiate the calcification process of mesenchymal stem cells in vitro through the activation of the TGF-β1/Smad signaling pathway and promote the decay of echinococcosis
- Evaluation of prognostic markers in patients infected with SARS-CoV-2
- N6-Methyladenosine-related alternative splicing events play a role in bladder cancer
- Characterization of the structural, oxidative, and immunological features of testis tissue from Zucker diabetic fatty rats
- Effects of glucose and osmotic pressure on the proliferation and cell cycle of human chorionic trophoblast cells
- Investigation of genotype diversity of 7,804 norovirus sequences in humans and animals of China
- Characteristics and karyotype analysis of a patient with turner syndrome complicated with multiple-site tumors: A case report
- Aggravated renal fibrosis is positively associated with the activation of HMGB1-TLR2/4 signaling in STZ-induced diabetic mice
- Distribution characteristics of SARS-CoV-2 IgM/IgG in false-positive results detected by chemiluminescent immunoassay
- SRPX2 attenuated oxygen–glucose deprivation and reperfusion-induced injury in cardiomyocytes via alleviating endoplasmic reticulum stress-induced apoptosis through targeting PI3K/Akt/mTOR axis
- Aquaporin-8 overexpression is involved in vascular structure and function changes in placentas of gestational diabetes mellitus patients
- Relationship between CRP gene polymorphisms and ischemic stroke risk: A systematic review and meta-analysis
- Effects of growth hormone on lipid metabolism and sexual development in pubertal obese male rats
- Cloning and identification of the CTLA-4IgV gene and functional application of vaccine in Xinjiang sheep
- Antitumor activity of RUNX3: Upregulation of E-cadherin and downregulation of the epithelial–mesenchymal transition in clear-cell renal cell carcinoma
- PHF8 promotes osteogenic differentiation of BMSCs in old rat with osteoporosis by regulating Wnt/β-catenin pathway
- A review of the current state of the computer-aided diagnosis (CAD) systems for breast cancer diagnosis
- Bilateral dacryoadenitis in adult-onset Still’s disease: A case report
- A novel association between Bmi-1 protein expression and the SUVmax obtained by 18F-FDG PET/CT in patients with gastric adenocarcinoma
- The role of erythrocytes and erythroid progenitor cells in tumors
- Relationship between platelet activation markers and spontaneous abortion: A meta-analysis
- Abnormal methylation caused by folic acid deficiency in neural tube defects
- Silencing TLR4 using an ultrasound-targeted microbubble destruction-based shRNA system reduces ischemia-induced seizures in hyperglycemic rats
- Plant Sciences
- Seasonal succession of bacterial communities in cultured Caulerpa lentillifera detected by high-throughput sequencing
- Cloning and prokaryotic expression of WRKY48 from Caragana intermedia
- Novel Brassica hybrids with different resistance to Leptosphaeria maculans reveal unbalanced rDNA signal patterns
- Application of exogenous auxin and gibberellin regulates the bolting of lettuce (Lactuca sativa L.)
- Phytoremediation of pollutants from wastewater: A concise review
- Genome-wide identification and characterization of NBS-encoding genes in the sweet potato wild ancestor Ipomoea trifida (H.B.K.)
- Alleviative effects of magnetic Fe3O4 nanoparticles on the physiological toxicity of 3-nitrophenol to rice (Oryza sativa L.) seedlings
- Selection and functional identification of Dof genes expressed in response to nitrogen in Populus simonii × Populus nigra
- Study on pecan seed germination influenced by seed endocarp
- Identification of active compounds in Ophiopogonis Radix from different geographical origins by UPLC-Q/TOF-MS combined with GC-MS approaches
- The entire chloroplast genome sequence of Asparagus cochinchinensis and genetic comparison to Asparagus species
- Genome-wide identification of MAPK family genes and their response to abiotic stresses in tea plant (Camellia sinensis)
- Selection and validation of reference genes for RT-qPCR analysis of different organs at various development stages in Caragana intermedia
- Cloning and expression analysis of SERK1 gene in Diospyros lotus
- Integrated metabolomic and transcriptomic profiling revealed coping mechanisms of the edible and medicinal homologous plant Plantago asiatica L. cadmium resistance
- A missense variant in NCF1 is associated with susceptibility to unexplained recurrent spontaneous abortion
- Assessment of drought tolerance indices in faba bean genotypes under different irrigation regimes
- The entire chloroplast genome sequence of Asparagus setaceus (Kunth) Jessop: Genome structure, gene composition, and phylogenetic analysis in Asparagaceae
- Food Science
- Dietary food additive monosodium glutamate with or without high-lipid diet induces spleen anomaly: A mechanistic approach on rat model
- Binge eating disorder during COVID-19
- Potential of honey against the onset of autoimmune diabetes and its associated nephropathy, pancreatitis, and retinopathy in type 1 diabetic animal model
- FTO gene expression in diet-induced obesity is downregulated by Solanum fruit supplementation
- Physical activity enhances fecal lactobacilli in rats chronically drinking sweetened cola beverage
- Supercritical CO2 extraction, chemical composition, and antioxidant effects of Coreopsis tinctoria Nutt. oleoresin
- Functional constituents of plant-based foods boost immunity against acute and chronic disorders
- Effect of selenium and methods of protein extraction on the proteomic profile of Saccharomyces yeast
- Microbial diversity of milk ghee in southern Gansu and its effect on the formation of ghee flavor compounds
- Ecology and Environmental Sciences
- Effects of heavy metals on bacterial community surrounding Bijiashan mining area located in northwest China
- Microorganism community composition analysis coupling with 15N tracer experiments reveals the nitrification rate and N2O emissions in low pH soils in Southern China
- Genetic diversity and population structure of Cinnamomum balansae Lecomte inferred by microsatellites
- Preliminary screening of microplastic contamination in different marine fish species of Taif market, Saudi Arabia
- Plant volatile organic compounds attractive to Lygus pratensis
- Effects of organic materials on soil bacterial community structure in long-term continuous cropping of tomato in greenhouse
- Effects of soil treated fungicide fluopimomide on tomato (Solanum lycopersicum L.) disease control and plant growth
- Prevalence of Yersinia pestis among rodents captured in a semi-arid tropical ecosystem of south-western Zimbabwe
- Effects of irrigation and nitrogen fertilization on mitigating salt-induced Na+ toxicity and sustaining sea rice growth
- Bioengineering and Biotechnology
- Poly-l-lysine-caused cell adhesion induces pyroptosis in THP-1 monocytes
- Development of alkaline phosphatase-scFv and its use for one-step enzyme-linked immunosorbent assay for His-tagged protein detection
- Development and validation of a predictive model for immune-related genes in patients with tongue squamous cell carcinoma
- Agriculture
- Effects of chemical-based fertilizer replacement with biochar-based fertilizer on albic soil nutrient content and maize yield
- Genome-wide identification and expression analysis of CPP-like gene family in Triticum aestivum L. under different hormone and stress conditions
- Agronomic and economic performance of mung bean (Vigna radiata L.) varieties in response to rates of blended NPS fertilizer in Kindo Koysha district, Southern Ethiopia
- Influence of furrow irrigation regime on the yield and water consumption indicators of winter wheat based on a multi-level fuzzy comprehensive evaluation
- Discovery of exercise-related genes and pathway analysis based on comparative genomes of Mongolian originated Abaga and Wushen horse
- Lessons from integrated seasonal forecast-crop modelling in Africa: A systematic review
- Evolution trend of soil fertility in tobacco-planting area of Chenzhou, Hunan Province, China
- Animal Sciences
- Morphological and molecular characterization of Tatera indica Hardwicke 1807 (Rodentia: Muridae) from Pothwar, Pakistan
- Research on meat quality of Qianhua Mutton Merino sheep and Small-tail Han sheep
- SI: A Scientific Memoir
- Suggestions on leading an academic research laboratory group
- My scientific genealogy and the Toronto ACDC Laboratory, 1988–2022
- Erratum
- Erratum to “Changes of immune cells in patients with hepatocellular carcinoma treated by radiofrequency ablation and hepatectomy, a pilot study”
- Erratum to “A two-microRNA signature predicts the progression of male thyroid cancer”
- Retraction
- Retraction of “Lidocaine has antitumor effect on hepatocellular carcinoma via the circ_DYNC1H1/miR-520a-3p/USP14 axis”