Molecular and phenotypic markers for pyramiding multiple traits in rice
-
 ,
,

Abstract
Gene pyramiding has been developed for effective resistant genes, such as bacterial blight resistance genes. However, in this report, pyramiding for multiple traits is presented. We evaluated 28 pyramided rice lines derived from 7 hybridizations of valuable parents for high yield potential, early maturity, aroma, high grain quality, and resistance to brown planthopper using both simple-sequence repeat markers and phenotypic markers. Pyramided rice genotypes were evaluated with RM282 molecular marker linked to GW3 gene associated with grain weight, RM259 (number of panicles), RM3600 (length of panicles), RM3701 (short panicles), RM19414 (Hd3), RM7601 (Hd2), RM190, and SSIIa (waxy), IFAP and ESP (aroma), RM586 (Bph3 and Bph4), and RM8213 (Qpbh4 and Bph17(t)). Eight genotypes (i.e., #9, #15, #16, #22, #25, #26, #27, and #28) were confirmed to have desired genes associated with multiple important traits based on molecular markers and morpho-agronomic traits. A strong correlation was found between RM19414 and heading date, between specific markers for aroma and sensory test. Meanwhile, a weak correlation was found between RM259 with panicle number, RM586, and RM8213 with trichome density. As presented in this study, gene pyramiding promises a new hope to combine multiple traits in rice breeding. Promising rice lines will be incorporated in the multiple traits breeding program.
1 Introduction
Rice (Oryza sativa L.) is the primary source of carbohydrates for more than half of the world’s population. The world population is now progressively growing, while on the other side, agricultural lands have been declining. Both tendencies require increasing rice production through breeding high-yielding rice genotypes. Selection for high-yield (>10 tons/ha), early maturity (<100 days after planting), and high-quality traits are the main objectives in rice breeding programs. However, early maturity trait has been reported to be negatively correlated with grain yield [1]. In rice varieties with very early maturing (<48 days flowering), most of their productivity are still low, and rice varieties with high productivity, in general, have intermediate maturity [2,3,4]. Therefore, developing early maturing rice with high yield potential remains to be the main challenge for rice breeders.
To overcome this obstacle introgression of early maturity traits into superior (high-yielding) cultivar to create a new cultivar with a short life cycle and high grain yield potential is necessary. Meanwhile, improving yield potential is an effort in increasing rice production [5]. The cultivars frequently used as a genetic resource for early maturity is cv. Kitaake, with extremely early maturity, originated from Hokkaido, Japan. Flowering initiates 7–8 weeks after sowing, seeds are harvested in 11–12 weeks, grains are moister and more glutinous (sticky), and fewer tillers are formed (i.e., seven tillers per plant) [6]. We have developed superior rice with early maturity by hybridization between cv. Ciapus and cv. Kitaake. Ciapus (indica) is a high-yielding cultivar (>8.2 ton/ha) with an average weight of 31 g per 1000 grains. Molecular and phenotypic markers have selected early maturity genotypes derived from this hybridization. However, these selected genotypes were susceptible to brown planthopper (BPH, Nilaparvata lugens Stal.), which was a weakness for the rice genotype since both parents do not carry any BPH-resistance genes. The BPH is a major pest in rice-producing areas and may cause yearly yield loss [7].
The major genes that control BPH-resistance in several cultivated and wild species have been identified and mapped with DNA markers [5,8,9,10,11]. The Indian rice variety PTB 33 has been known to carry BPH-resistance genes (i.e., Bph2, Bph3, and Bph4) in chromosome 6, and some BPH-resistance genes that are assumed as quantitative trait loci (QTL) [12,13,14]. In addition, a wax layer on its leaf surface (cuticle) can disturb when BPH sucks the leaf [5]. PTB 33 has been used as a BPH-resistance donor in conventional breeding. However, due to its poor grain quality and other traits such as high amylose content, low gel consistency, chalky endosperm, no fragrance, and late maturity, PTB 33 is avoided by both farmers and consumers. Amylose content is controlled by the Wxa allele of the waxy locus (approximately 380 kb), which is located very close to the Bph3 gene in PTB 33 [15]. The amylose content and gel consistency are physicochemical properties of endosperm that affect cooking and eating quality, which is associated with the Wx locus at the short arm of chromosome 6 [16,17,18]. Hu et al. [19] also reported that the line carrying Bph3 has amylose content of 24.7–27.8%, which indicates that Bph3 is linked to the Wx allele controlling amylose synthesis. Therefore, obtaining rice genotype with Bph3 resistance gene and intermediate amylose content (20–24%) is difficult. Rice with intermediate amylose is preferred since it becomes soft and fluffy after cooking [20,21]. However, this effort needs longer time when conventional breeding approach is used. Breeding rice resistant to BPHs with intermediate amylose content, high yielding, and early maturity is an effort of pyramiding multiple traits into one rice genotype.
At present, research on gene pyramiding in rice has been successfully conducted by many researchers. In this crop, pyramiding schemes include genes for the resistance to blight (i.e., xa5, xa13, Xa21 [20]), and blast (i.e., Pi(2)t, Piz-5, and Pi(t)a [22]). In wheat cultivar Yang158, successful pyramiding schemes have been developed for powdery mildew resistance gene (i.e., Pm2 + Pm4a, Pm2 + Pm21, Pm4a + Pm21 [23]). However, all these pyramiding efforts have been conducted for various biotypes or strains of a single pathogen with is different biotype or strain.
Our study here demonstrates a pyramiding effort of different superior morpho-agronomic traits, such as high yield (grain weight, number of panicles, and panicle length), early maturity, rice quality (intermediate amylose content and aroma), and resistance to BPH in a common breeding line. In previous studies, we have successfully developed some genotypes that combined superior traits such as, early maturity (i.e., genotype CAKA (cv. Ciapus x KA) with Hd3 and Hd4; genotypes SP87-4 (Sintanur x PTB 33), PP48-3 (Pandanwangi x PTB 33) with fgr, Wx, SSIIa, Bph3, Bph4, Qbph4, and Bph17(t), IP158-5 (IR-64 x PTB33) with Bph3, Bph4, Qbph4, and Bph17(t)) [24,25].
Using conventional breeding approaches, gene pyramiding is challenging due to combining two or more genes into one genotype, and identifying genes inserted in the pyramided genotype is more difficult [23,26]. However, using molecular markers closely linked with the desired genes are more powerful in the identification of plants with two and three inserted genes [20]. Molecular markers can be used to identify favorable genes and multiple alleles in the gene pools of diverse genotypes [27]. However, in this study we employed simple sequence repeat (SSR) markers followed by phenotypic assessment. Numerous rice SSR markers were available in the previous study to identify, map, and select traits. Here we used SSR markers representing five traits in the pyramiding line. It is believed that both approaches (molecular markers and phenotypic assessment) are efficient and accurate in predicting the genetic composition of rice genotypes. Another important aspect is the correlation between the molecular markers and desired traits, which indicate whether the molecular markers are suitable for identifying the phenotypic traits in a pyramiding population [28]. Therefore, this study aims to identify rice genotypes with high yield potential and desirable agronomic traits (i.e., early maturity, intermediate amylose content, aromatic, and resistance to BPH) using molecular markers and phenotypic characteristics. The information collected will be helpful for rice breeders since the obtained rice lines will correspond to farmers’ and consumers’ preferences.
2 Materials and methods
2.1 Plant materials
Twenty-eight pyramided rice genotypes derived from seven combinations of hybridization including CAKA41 x PP48-3; PP48-3 x CAKA41; CAKA41 x SP87-4; SP87-4 x CAKA41; SP87-4 x PP48-3; PP48-3 x SP87-4; and PP48-3 x IP158-5 were used in this study. Plants were grown in the screen-house with a standard cultivation method.
2.2 Target genes and molecular markers used
Twelve genes encoding the desired morpho-agronomical traits were chosen for molecular marker-assisted selection (Table 1). A total of ten SSR markers and one specific marker were chosen to identify desirable traits.
SSR and Gene-specific PCR primers used in this experiment
| Marker | Linked to | Chr. | Primer sequences | PCR product size (bp) | |
|---|---|---|---|---|---|
| Forward | Reverse | ||||
| RM282 | GW3 | 3 | gctccacctgcttaagcatc | tgaagaccatgttctgcagg | 162–226 |
| RM259 | qPN1 | 1 | tggagtttgagaggaggg | cttgttgcatggtgccatgt | 162 |
| RM3600 | LP1 | 9 | tgcccacacatgatgagc | aacgggcaagagatcttctg | 130 |
| RM3701 | SP1 | 11 | gagctagagggaggaggtgc | ttgactgatagccgattggg | 174 |
| RM7601 | Hd2 | 6 | gcctcgctgtcgctaatatac | cagcctctccttgtgttgtg | 133 |
| RM19414 | Hd3 | 6 | gtcagaacttcaacaccaagg | ctgtatagcttgatctaggagtagc | 504 |
| RM190 | Wx | 6 | ctttgtctatctcaagacac | ttgcagatgttcttcctgatg | 104–124 |
| SSIIa | SSIIa | 6 | caaggagagctggagggggc | acatgccgcgcacctggaaa | 300 and 500 |
| RM586 | Bph3, Bph 4 | 6S and 6 | acctcgcgttattaggtaccc | gagatacgccaacgagatacc | 271 |
| RM8213 | Qbph4, Bph17(t) | 4 and 4S | agcccagtgatacaaagatg | gcgaggagataccaagaaag | 177 |
| IFAP and ESP | Aromatic (fgr) | 8 | 5′ cataggagcagctgaaatatatacc 3′ | 257 | |
| 5′ ttgtttggagcttgctgatg 3′ | 257 | ||||
| INSP and EAP | Non-aromatic | 8 | 5′ ctggtaaaaagattatggcttca 3′ | 355 | |
| 5′ agtgctttacaaagtcccgc 3′ | 355 | ||||
2.3 DNA extraction and molecular marker detection
Genomic DNA was isolated from fresh leaves using the Cetyl trimethyl ammonium bromide (CTAB) method [29] with slight modification. The quality of DNA was estimated by 0.8% agarose gel electrophoresis. The PCR reaction was performed in 12.5 µl of reaction mixture containing 1 µl of 25ng DNA template, 1 µl of 0.4 μM each of forward and reverse primers, and 9.5 µl of KAPA2GTM Fast ReadyMix (0.25 U/μl Taq DNA polymerase, 0.2 mM dNTPs, 1.5 mM MgCl2). The PCR products were separated on an agarose (1.5%) and polyacrylamide (0.8%) gels, stained with ethidium bromide (EtBr), and visualized with UV-transilluminator (Syngene Type G-Box). Agarose gel was used to distinguish DNA fragment with sizes of more than 100 bp, and for a size of less than 100 bp, polyacrylamide gel was used. Band size estimation was performed using GeneTools software (Syngene, UK). Molecular data were then scored as presence (1) and absence (0) according to the bands in parent lines.
2.4 Characterization of morpho-agronomic traits
The morpho-agronomic traits measured were plant height, total number of tillers, number of panicles, panicle length, number of spikelets per panicle, weight of spikelet per panicle, weight of 100 grains, and total weight of spikelet per plant. Early maturity was identified by observing heading date in the field. Observation and scoring were based on descriptor for rice based on Bioversity International. The aromatic trait was identified by a sensory test using 1.7% KOH solution [30]. Evaluation for trichome density, related to a defense system against BPH [31], was performed under a binocular microscope (magnification of 400×). Individual selection of plant was done as detected by both methods (molecular and phenotypic). Spearman’s Rank correlation coefficient was applied to assess the strength of the relationship between molecular and phenotypic data.
3 Results
3.1 Characterization of pyramided genotypes for multiple traits using molecular markers
A total of 28 pyramided rice genotypes were evaluated using SSR markers considering the presence or absence of the selected genes of interest (i.e., GW3, qPN1, LP1, Hd3, Hd4, fgr, Wx, SSIIa, Bph3, Bph4, Qbph4, and Bph17(t)). Twelve genotypes (#9, #15, #16, #19, #20, #23, #22, #23, #24, #25, #26, and #27) presented all desired genes, indicating that the twelve selected genes were successfully inserted into these plant’s genomes (Table 2).
Evaluation of pyramided genotypes for the selected morpho-agronomical traits using molecular markers
 |
+ presence of the expected band; – absence of the expected band; H – presence of two amplification bands; green highlight – genotypes selected.
Five genotypes from CAKA41 x PP48-3 (#1, #2, #3, #4, #5) had five genes (GW3, LP1, Hd2, Hd3, and Wx), while other three (#6, #7, and #8) had seven genes (GW3, LP1, Hd2, Hd3, Wx, Bph3, and Bph4). Four genotypes from PP48-3 x CAKA41 (#10, #11, #12, #13) exhibited ten of the selected genes [GW3, LP1, Hd2, Wx, SSIIa, fgr, Bph3, Bph4, Qbph4, and Bph17(t)], while another genotype (#14) exhibited seven genes [GW3, LP1, Hd2, Hd3, Wx, Bph3, and Bph4]. One genotype, from SP87-4 x CAKA41 (#17), was found to have seven genes (GW3, FLO, LP1, Hd2, Wx, SSIIa, fgr), with constitution genetic of BPH was heterozygous based on RM586 molecular marker (Bph3 and Bph4). One genotype (#18) from SP87-4 x PP48-3 revealed the presence of six selected genes (LP1, Hd2, Wx, SSIIa, Qbph4, and Bph17(t)), while another (#21) revealed the presence of nine (GW3, LP1, Hd2, Wx, fgr, Bph3, Bph4, Qbph4, and Bph17(t) genes) (Figures 1–3).
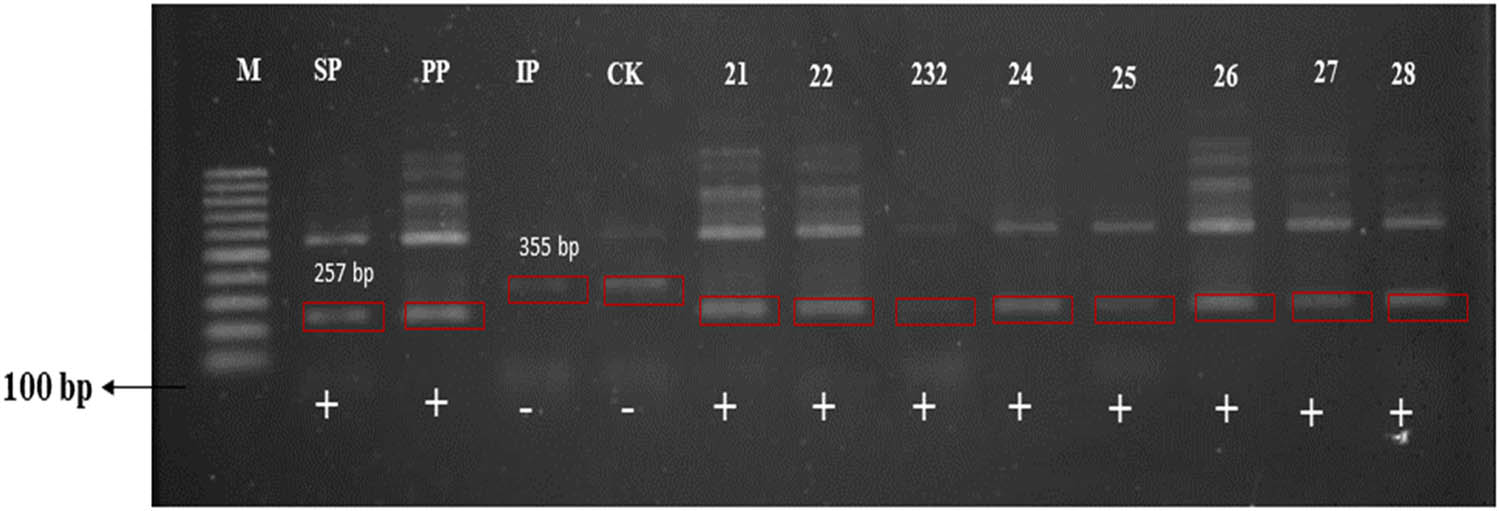
Amplification results using molecular marker for aroma (insert the name of the marker). M – DNA ladder; SP – Sintanur x PTB 33; PP – Pandanwangi x PTB 33; IP – IR64 x PTB33; CK – x KA; + aroma ; – non-aroma.
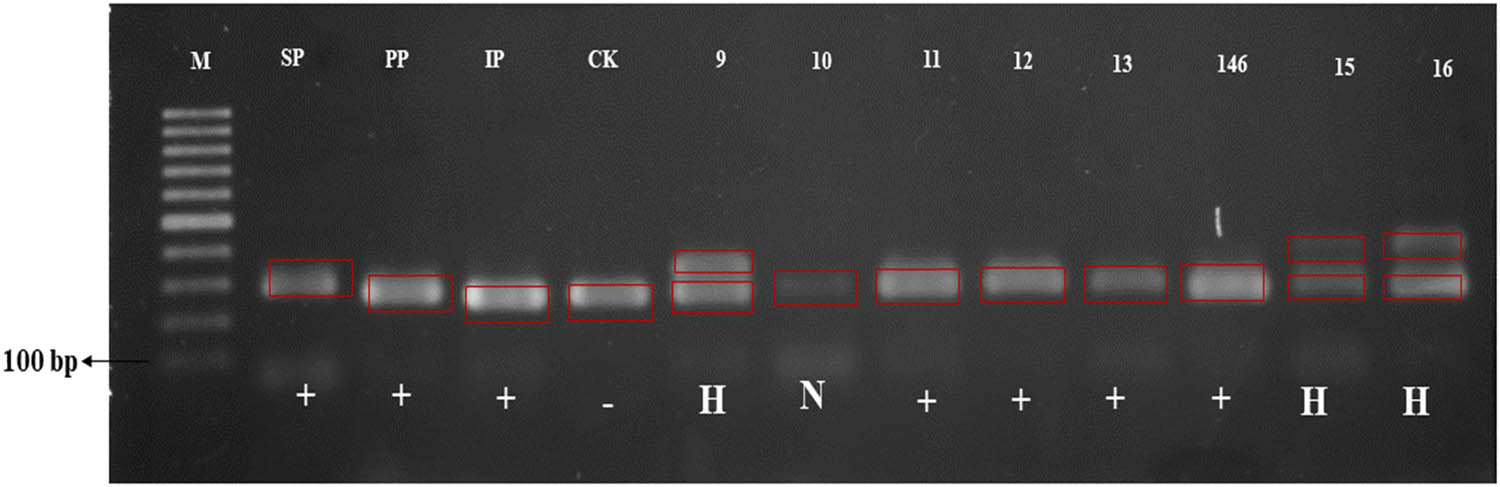
Amplification results using molecular marker for BPH-resistance (RM586). M – DNA ladder; SP – Sintanur x PTB 33; PP – Pandanwangi x PTB 33; IP – IR64 x PTB 33; CK – Ciapus x Kitaake; + resistance to BPH; – susceptible to BPH; H – heterozygous resistance to BPH.
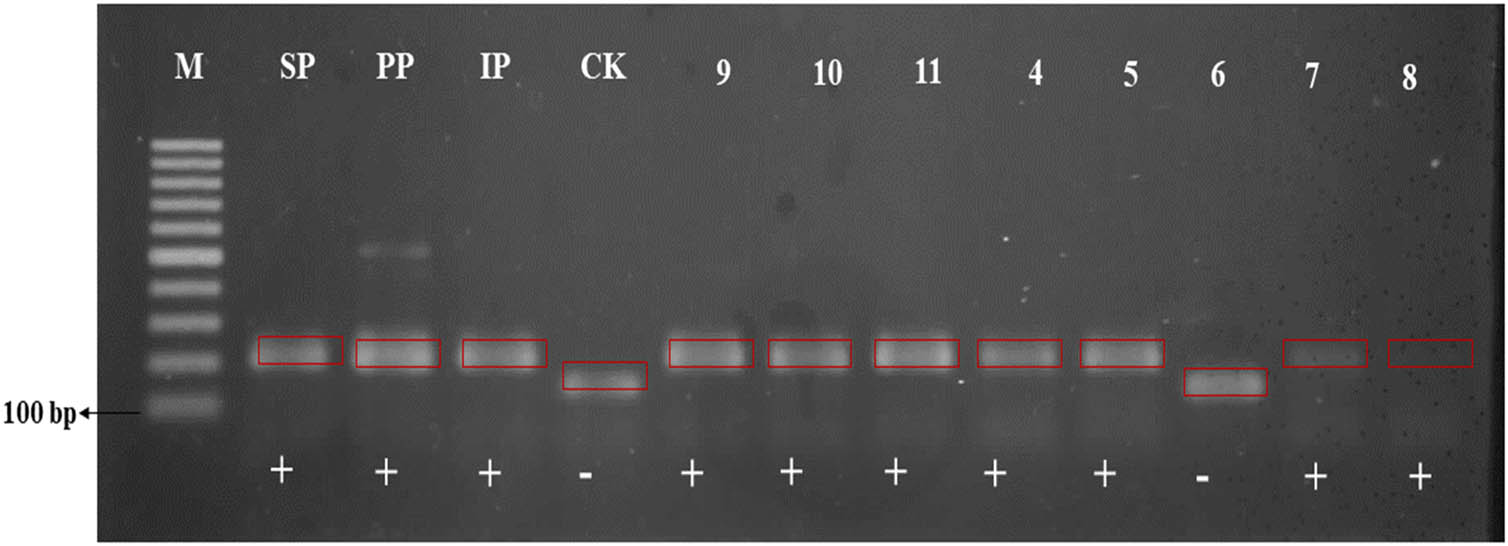
Amplification results using molecular marker for BPH-resistance (RM8213). M – DNA ladder; SP – Sintanur x PTB 33; PP – Pandanwangi x PTB 33; IP – IR64 x PTB 33; CK – Ciapus x Kitaake; + resistance to BPH; – susceptible to BPH.
3.2 Characterization of pyramided genotypes for multiple traits by phenotypic assessment
Six genotypes (#3, #6, #7, #9, #14, and #21) presented a high number of tillers, total number of panicles, long panicles, medium and high number of spikelets per panicle, and high grain weight (Table 3). Sixteen genotypes (#1, #2, #4, #5, #8, #10, #13, #15, #16, #17, #18, #22, #25, #26, #27, and #28) exhibited very high total number of tillers and panicles, medium panicle length, medium number of spikelets per panicle, high spikelet per panicle, and high grain weight. Grain weight was affected by grain size and the number of grains per panicle (Table 3).
Comparison of morpho-agronomic traits among pyramided genotypes
 |
GW/P – grain weight per panicle, W100 – weight of 100 grains, PN – panicle number, PL – panicle length, Hd – heading date, Ar – Aroma, Tr – number of trichomes, B – Heavy, R – light, Vh – very high, G – good, L – long, M – medium, Gj – mature, A – aromatic, S – less aromatic, N – non-aromatic, blue highlight – genotypes selected.
All evaluated genotypes were categorized as early maturing genotypes (<100 days after planting), according to a previous study [32]. The assessment of aroma revealed that 4 pyramided genotypes were not aromatic, 5 were medium aromatic, and 19 genotypes were aromatic.
Trichome density on parents and introgression lines show that trichome exists, but in lesser density than PTB 33, a BPH-resist cultivar (Figure 4).

Trichome density in parental line: (a) SP87-4; (b) PP48-3; (c) IP158-5; (d) CAKA41; and (e) PTB 33 (400 magnification).
3.3 The association between molecular markers and phenotypic assessment
The association between molecular markers and phenotypic assessment is presented in Table 4. RM19414 marker and specific marker of aroma were positively correlated with heading date (0.645) and sensory test (0.620). RM259 showed a positive correlation with the number of panicles (r = 0.377*), although the linkage of both marker relationships was low. The phenotypic assessment shows that most pyramided genotypes developed a high number of panicles (Table 3). Some genotypes did not present the qPN1 gene based on RM259. RM8213 showed no correlation with trichome density (r = 0.370), whereas RM586 showed non-significant negative correlation with trichome density (r = −0.360). Pyramided genotypes presented fewer trichomes than cv. PTB 33, but have longer trichome (Figure 4).
Correlation between molecular markers and phenotypic characteristics by Spearman’s rho (rs)
| Molecular marker | Phenotypic marker | Spearman’s rho (rs) | |
|---|---|---|---|
| P-value | r | ||
| RM259 | Number of panicles | 0.034 | 0.377* |
| RM19414 | Heading date | 0.000 | 0.645** |
| ESP/IFAP and INSP/EAP | Sensory test | 0.001 | 0.620** |
| RM586 | Trichome density | 0.840 | −0.360ns |
| RM8213 | Trichome density | 0.838 | 0.370 |
ESP – external sense primer, IFAP – internal fragrant antisense primer, INSP – internal non-fragrant sense primer, EAP – external antisense primer, ns – non significant, * significant at 0.05, and ** significant at 0.01.
4 Discussion
Identifying pyramided genes using molecular markers has become a straightforward approach for improving crop yield. Yield components refer to the structures of the rice plant that are directly converted into yield, such as grain weight, panicle number (PN), and panicle length. In this study, the grain weight was identified using RM282 (chromosome 3), supposedly linked to GW3, the gene controlling grain weight. RM282 confirmed the presence of the GW3 gene in female and male parents and all pyramided genotypes, indicating that the progenies can inherit the grain weight trait from parental lines. SP87-4, IP158-5, and CAKA41 parent lines present high grain weight. It has been known that progeny from gene pyramiding effort have GW3 gene related to high grain weight character since this gene is an inherited trait (40–60% transferable).
Pyramided rice genotypes were also identified using RM259 associated with PN [33]. PN has a direct positive effect on grain yield [34]. SP87-4 and IP158-5 parental lines with a high number of panicles were also linked to RM259. Rice exhibiting a high number of panicles with a high seed set will present a high yield. Agustina et al. [35] also concluded that there is a significant positive correlation between the grain yield and the total number of tillers and panicles. In addition, panicle length also influences the number of grains per panicle [36]. These genotypes correspond to rice varieties associated with superior traits and high productivity. However, in our study, the existence of qPN1 gene in some pyramided genotypes were not confirmed using RM 296 marker. The number of panicles is coded by many genes [22,37]. In a previous study, the performance of PN was identified as a QTL influenced by environmental factors (i.e., soil nutrients, planting distance, light intensity and duration, temperature, humidity, water supply, and other environmental conditions) [38]. No QTL of PN in rice has yet been cloned. Using mapping technology, QTL-linked PN per rice plant has been reported [39]. Moreover, using chromosome segment substituted lines derived from Nipponbare and Guangluai 4, Zhu et al. [40] successfully found the qPN1 gene, which influences PN per plant and several other agronomical traits.
Panicle length was evaluated using RM3600 (to identify long panicle) and RM3701 (short panicle) markers. Using RM3600, both parental lines and all pyramided progenies were confirmed to contain the LP1 gene associated with longer panicle phenotypes. No genotypes from pyramiding exhibited the SP1 gene associated with a shorter panicle. Length of panicle is quantitatively inherited and controlled by major and minor QTLs [41], presenting additive gene interaction. Akinwale et al. [42] also reported that panicle length is highly heritable and the selection of this character can be achieved directly based on their phenotypic performance. Characters with high heritability are dominant in the appearance of plant genetics.
Early maturity was identified using two markers, RM7601 (linked to Hd2) and RM19414 (linked to Hd3a). Hd3a induces rice flowering faster than usual in short-day conditions [43]. SP87-4, PP48-3, and IP158-5 were used as parental lines, while CAKA41 was used as a donor parent to Hd2 and Hd3. RM7601 revealed the presence of Hd2 in both parental lines, and all genotypes. RM19414 showed that Hd3a was present in 14 genotypes, while absent in 14 other genotypes. Hd3 gene is one of the essential genes controlling heading date. Eight genotypes from CAKA41 x PP48-3 presented Hd3a, while five genotypes from PP48-3 x CAKA41 did not contain this gene. Hd3a was detected in one genotype from CAKA41 x SP87-4.
Amylose content in rice was evaluated using two molecular markers: RM190 and SSIIa. RM190 detects intermediate amylose content due to the presence of Wx int allele in chromosome 6 in cv. Fajr and Domsiah [44]. SSIIa detects an alternative allele associated with amylose content [45]. In this work, RM190 revealed that 19 pyramided genotypes presented the Wx gene (low/intermediate amylose content) similar to SP87-4 and PP48-3 parental lines. Nine genotypes presented high amylose content similar to the CAKA41 parental line. SSIIa revealed that 19 pyramided genotypes exhibited low/intermediate amylose content similar to SP87-4 and PP48-3 parental lines, and nine other genotypes exhibited a high amylose content, similar to IP158-5 and CAKA41 parental lines. Therefore, two alleles control amylose content, presenting differential effects on this characteristic [46].
Rice aroma in parental lines and pyramided genotypes was qualitatively evaluated for the presence or absence of aroma. The specific markers ESP and IFAP are associated with the fgr locus used to distinguish aromatic and non-aromatic rice. The fgr gene was present in both parental lines and pyramided genotypes. Two, out of seven combinations of hybridization, all progenies from crosses CAKA41 x PP48-3 and CAKA41 x SP87-4, did not detect the presence of the fgr gene. These lines were developed from non-aromatic (CAKA41) and aromatic (PP48-3 and SP87-4) rice genotypes; meanwhile, the pyramided rice genotype was obtained through crossing between aromatic lines (SP87-4 and PP48-3) and non-aromatic lines (CAKA41 and IP158-5), resulting in progenies with homozygous aromatic lines (Table 2), indicating that the aroma genes in each of these two aromatic lines are recessive homozygous and two aromatic lines are heterozygous. Previous research reports that a single recessive gene controls aromatic compounds in cultivar Della [47]. According to Yoshihashi et al. [48], aromatic expression is controlled by the fgr gene inherited from parental lines, assuming that this aromatic character will be presented in pyramided genotype if present in the parental lines (predominantly in the female parental line). Crosses between non-aroma (CAKA41 and IP158-5) with aromatic lines (SP87-4 and PP48-3) resulted in a progeny of non-aromatic homozygous (AA), indicating that female parental non-aromatic lines were heterozygous and donor aromatic parental lines were recessive homozygous, so the aromatic trait was not inherited. However, it has been reported that other genes, such as badh2 (chromosomes 8 and 3) and badh1 (chromosome 4), are alternative genes that control aroma [49]. Other genes proposed are aro3-1 (chromosome 3), aro4-1 (chromosome 4), and aro8-1 (chromosome 8) [50]. Therefore, aromatic genotypes may be undetected using IFAP/ESP and ENSP/EAP markers [51]. Moreover, different gene locations may be responsible for the observed aroma in rice or the presence of another major aromatic compound [52]. ESP and IFAP have been reported as specific primers for the aromatic gene (257 bp), while INSP and EAP as non-aromatic genes (355 bp), amplifying in heterozygous conditions [53]. The sensory test using KOH solution is also a suitable method for rapid identification of aroma in rice [54]. The aroma in rice results from volatile compounds, such as the 2-acetyl-1-pyrroline (2-AP) [55]. According to Bradbury et al. [56], the 2-AP compound is synthesized by the action of a betaine aldehyde dehydrogenase 2 (BAD2) enzyme. BAD expression is controlled by the fgr gene that is easily transferred to the offspring. An aroma-specific marker will also detect aromatic traits of pyramided genotypes as detected by a sensory test. In this study, we obtained some pyramided genotypes with intermediate aroma. The intensity of the fragrance in rice is influenced by the presence of 2-AP, its concentration being influenced by several environmental conditions (e.g., soil characteristics, soil tillage, climate condition during the grain filling, handling postharvest, and duration of storage) [55,57]. The intensity of the aroma in aromatic rice decreases and may disappear depending on the stage of the plant under disadvantageous conditions. For instance, high temperature significantly affects the expression of aroma-associated genes, decreasing 2-AP accumulation, especially at the grain filling and ripening stages [57].
BPH-resistance evaluation was conducted by observing morphological traits associated with trichome density, a physical resistance mechanism [58,59]. Pest-resistant rice plants present traits that allow pest avoidance or recovery from pest attacks. The presence of high density of trichome is a physical anti-xenosis resistance mechanism since plant morphology does not favor pest development, BPHs are incapable of laying their egg, breeding, or eating. BPH resistance was identified using two markers, RM586 (linked to Bph3 and Bph4) and RM8213 [linked to Qbph4 and Bph17(t)]. RM586 marker revealed that PP48-3, 291 SP87-4, and IP158-5 parental lines were BPH-resistant, while CAKA41 was not. Those three parent genotypes resulted from crosses where one of their parental lines was PTB 33, linked to Bph3 and Bph4 genes. Six pyramided genotypes (i.e., #1, #2, #3, #4 #5, and #18) did not present the BPH-resistance gene; eight genotypes (i.e., #9, #15, #16, #17, #20, #22, #23, and #24) were heterozygous, and 14 genotypes (i.e., #6, #7, #8, #10, #11, #12, #13, #14, #19, #21, #25, #26, #27, #28) presented this gene. Plant with Bph3 and Bph6 genes were resistant to BPH biotype 4 [60]. Similar results were obtained with RM8213. The parental lines PP48-3, SP87- 299 4, and IP158-5 presented the BPH-resistance gene, while in CAKA41, the gene was absent. Ten pyramided genotypes (i.e., #1, #2, #3, #4 #5, #6, #7, #8, #14, and #17) did present the BPH-resistance gene, two genotypes (i.e., #16 and #27) were heterozygous, and 17 genotypes (i.e., #9 #10, #11, #12, #13, #15, #18, #19, #20, #21, #22, #23, #24, #25, #26, #27, and #28) presented this gene. Results indicate that BPH resistance in these pyramided genotypes is controlled by a dominant gene (i.e., Qbph4 and Bph17(t)). Recently, a new important resistance gene, which is tentatively designated as Bph17, has been closely linked to markers RM8213, RM6487, and RM401 on chromosome 4 [61]. The existence of Bph3, Bph4, Qbph4, and Bph17(t) genes in pyramided genotypes is beneficial since accumulating several BPH-resistance genes in a single cultivar improves resistance to BPH biotypes [62].
Trichomes density is controlled by several genes, such as OsGL1 (chromosome 5) and OsGL6 (chromosome 6) [63,64]. Moreover, other factors can also influence the density distribution and trichome morphology. Meanwhile, different organs will vary in trichome density. Different leaf types also have different trichome densities. The difference in results may be attributed to the difference in the genetic background of genotypes used and the environmental conditions of the experiment. The 12 closely linked SSR markers identified in this study would be helpful in maker-assisted breeding programs aimed at developing new, improved superior cultivars. These results showed a highly significant correlation, which indicated that these molecular markers could be used to predict the existence of genes linked to traits in all pyramided genotypes.
5 Conclusion
New superior rice introgression lines with desired traits have been developed. In this study, molecular markers were used to validate the presence of favorable alleles in pyramided genotypes. Eight pyramided genotypes (i.e., #9, #15, #16, #22, #25, #26, #27, and #28) presented multiple desirable traits as assessed by molecular markers and the observation of phenotypic characteristics. These superior genotypes presented high yield, aroma, early maturity, high grain quality, and BPH-resistance. Strong correlations were observed between RM19414 and heading date and EAP, and INSP and sensory test. Weak correlations were observed between RM259 and PNs, RM586 and RM8213, and trichome density. Molecular markers were strongly correlated with specific traits, and the selected genotypes will be utilized in future rice breeding programs.
Acknowledgments
Authors wish to thank Directorate General of Higher Education, Ministry of Education, Culture, Research and Technology for awarding a basic research grant (Hibah Penelitian Dasar) to NC.
-
Funding information: The study was financed by a basic research grant (Hibah Penelitian Dasar), donated by Directorate General of Higher Education, Ministry of Education, Culture, Research and Technology.
-
Conflict of interest: The authors state no conflict of interest
-
Data availability statement: The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
[1] Pramudyawardani EF, Suprihatno J, Mejaya MJ. Yield of promising rice lines of very early and early maturing rice lines. J Penelit Pertan Tanam Pangan. 2015;34(1):1–11. (Indonesian). 10.21082/jpptp.v34n1.2015.p1-11.Search in Google Scholar
[2] Ying J, Peng S, He Q, Yang H, Yang C, Visperas RM, et al. Comparison of high-yield rice in tropical and subtropical environments: I. Determinants of grain and dry matter yields. Field Crop Res. 1998;57:71–84. 10.1016/s0378-4290(98)00077-x.Search in Google Scholar
[3] Peng S, Cassman KG, Virmani SS, Sheehy J, Khush GS. Yield potential trends of tropical rice since the release of IR8 and the challenge of increasing rice yield potential. Crop Sci. 1999 Nov;39(6):1552–9. 10.2135/cropsci1999.3961552x.Search in Google Scholar
[4] Takai T, Matsura S, Nishio T, Ohsumi A, Shiralwa T, Horie T. Rice yield potential is closly realted to crop growth rate during late reproductive period. Field Crop Res. 2006;96(2):328–9. 10.1016/j.fcr.2005.08.001.Search in Google Scholar
[5] Jairin J, Phengrat K, Teangdeerith S, Vanavichit A, Toojinda T. Mapping of a broad-spectrum brown planthopper resistance gene, Bph3, on rice chromosome 6. Mol Breed. 2007;19:35–44. 10.1007/s11032-006-9040-3.Search in Google Scholar
[6] Kim SL, Choi M, Jung KH, An G. Analysis of the early-flowering mechanisms and generation of T-DNA tagging lines in Kitaake, a model rice cultivar. J Exp Botany. 2013;64(14):4169–82. 10.1093/jxb/ert226.Search in Google Scholar PubMed PubMed Central
[7] Dyck VA, Thomas B. The brown planthopper problem. In: Brown planthopper: threat to rice production in Asia. Los Banos, Laguna, Philippines: IRRI; 1979. p. 4–17. http://books.irri.org/9711040220_content.pdf.Search in Google Scholar
[8] Huang Z, He G, Shu L, Li X, Zhang Q. Identification and mapping of two brown planthopper resistance genes in rice. Theor Appl Genet. 2002;102:929–34. 10.1007/s001220000455.Search in Google Scholar
[9] Jena KK, Pasalu IC, Rao YK, Varalaxmi Y, Krishnaiah K, Khush GS, et al. Molecular tagging of a gene for resistance to brown planthopper in rice (Oryza sativa L.). Euphytica. 2003;129:81–8. 10.1023/A:1021590025240.Search in Google Scholar
[10] Jena KK, Jeung JU, Lee JH, Choi HC, Brar DS. High-resolution mapping of a new brown planthopper (BPH) resistance gene, Bph18(t), and marker-assisted selection for BPH resistance in rice (Oryza sativa L.). Theor Appl Genet. 2006;112:288–97. 10.1007/s00122-005-0127-8.Search in Google Scholar PubMed
[11] Rongbai L, Xueyi Q, Sumei W, Pandey MP, Pathak PK, Fenguan H, et al. Inheritance of resistance to brown planthopper in an Oryza rufipogon (Griff.)-derived line in rice. Curr Sci. 2001;80(11):1421–23. http://www.jstor.org/stable/24104967.Search in Google Scholar
[12] Sun L, Su C, Wang C, Zhai H, Wan J. Mapping of a major resistance gene to the brown planthopper in the rice cultivar Rathu Heenati. Breed Sci. 2005;55:391–6. 10.1270/jsbbs.55.391.Search in Google Scholar
[13] Santhanalakshmi S, Saikumar S, Rao S, SaiHarini A, Khera P, Shashidar HE, et al. Mapping genetic locus linked to BPH resistance in rice. Int J Plant Breed Genet. 2010;4(1):13–22. 10.3923/ijpbg.2010.13.22.Search in Google Scholar
[14] Nugaliyadde L, Des Abeysiriwardena DS, Samanmalee LG, Pathirana R, Wilkins RM. Inheritance of resistance in rice to brown planthopper: its implications on rice varietal improvement in Sri Lanka; 2001. http://delphacid.s3.amazonaws.com/7674.pdf.Search in Google Scholar
[15] Jairin J, Kotchasatit U, Saleeto S, Jearakongman S, Srivilai K, Chamarerk V, et al. Application of marker-assisted breeding to improve biotic stress resistance for rainfed lowland rice in northeastern Thailand. SABRAO J Breed Genet. 2017;49(2):168–78. file:///E:/download/SABRAO-J-Breed-Genet-49-2-168-178-Jairin.pdf.Search in Google Scholar
[16] Itoh K, Ozaki H, Okada K, Hori H, Takeda Y, Mitsu T. Introduction of Wx transgene into rice wx mutants leads to both high- and low-amylose rice. Plant Cell Physiol. 2003 May;44(5):473–80. 10.1093/pcp/pcg068.Search in Google Scholar PubMed
[17] Lanceras J, Huang ZL, Naivikul O, Vanavichit A, Ruanjaichon V, Tragoonrung S. Mapping of genes for cooking and eating qualities in Thai jasmine rice (KDML105). DNA Res. 2000 Apr 28;7(2):93–101. 10.1093/dnares/7.2.93.Search in Google Scholar PubMed
[18] Zhou PH, Tan YF, He YQ, Xu CG, Zhang Q. Simultaneous improvement for four quality traits of Zhenshan 97, an elite parent of hybrid rice, by molecular marker-assisted selection. Theor Appl Genet. 2003;106:326–33. 10.1007/s00122-002-1023-0.Search in Google Scholar PubMed
[19] Hu W, Xiao H, Hu K, Jiang Y, Zhang Y. Application of marker-assisted backcross to introgress Bph3, Bph14 and Bph15 into an elite indica rice variety for improving its resistance to brown planthopper. Plant Breed. 2016;135:291–300. 10.1111/pbr.12364.Search in Google Scholar
[20] Singh S, Sidhu JS, Huang N, Vikal Y, Li Z, Brar DS, et al. Pyramiding three bacterial blight resistance genes (xa5, xa13 and Xa21) using marker-assisted selection into indica rice cultivar PR106. Theor Appl Genet. 2001;102:1011–5. 10.1007/s001220000495.Search in Google Scholar
[21] Joseph M, Gopalkrishnan S, Sharma RK, Singh VP, Singh AK, Singh NK, et al. Combining bacterial blight resistance and Basmati quality characteristics by phenotypic and molecular marker-assisted selection in rice. Mol Breed. 2004;13:377–87. 10.1023/B:MOLB.0000034093.63593.4c.Search in Google Scholar
[22] Hittalmani S, Parco A, Mew TV, Zeigler RS, Huang N. Fine mapping and DNA marker-assisted pyramiding of the three major genes for blast resistance in rice. Theor Appl Genet. 2000;100:1121–8. 10.1007/s001220051395.Search in Google Scholar
[23] Liu J, Liu D, Tao W, Li W, Wang S, Chen P, et al. Molecular marker-facilitated pyramiding of different genes for powdery mildew resistance in wheat. Plant Breed. 2000;119:21–4. 10.1046/j.1439-0523.2000.00431.x.Search in Google Scholar
[24] Zaelani A. Identification of early maturing gene in F2 generation of rice derived from cross between cv. Ciapus X Kitaake by using two SSR markers and its correlation with heading date. Bachelor thesis. Universitas Padjadjaran; 2013. unpublished. (Indonesian).Search in Google Scholar
[25] Asri AP. Screening of promising rice genotypes for resistance to the brown planthopper based on standard seedbox screening test (SSST) and simple sequence repeats (SSR) molecular marker. thesis; 2016. unpublished. (Indonesian).Search in Google Scholar
[26] Rajpurohit D, Kumar R, Kumar M, Paul P, Awasthi AA, Bash PO, et al. Pyramiding of two bacterial blight resistance and a semidwarfing gene in Type 3 Basmati using marker-assisted selection. Euphytica. 2001;178:111–26. 10.1007/s10681-010-0279-8.Search in Google Scholar
[27] Jena KK, Mackill DJ. Molecular markers and their use in marker-assisted selection in rice. Crop Sci. 2008;48:1266–76. 10.2135/cropsci2008.02.0082.Search in Google Scholar
[28] Ramakrishnan AP, Meyer SE, Waters J, Stevens MR, Coleman CE, Fairbanks DJ. Correlation between molecular markers and adaptively significant genetic variation in Bromus tectorum (Poaceae), an inbreeding annual grass. Am J Botany. 2004;91(6):797–803. 10.3732/ajb.91.6.797.Search in Google Scholar PubMed
[29] Dellaporta SL, Wood J, Hicks JB. A plant DNA minipreparation: Version II. Plant Mol Biol Rep. 1983;1:19–21. 10.1007/BF02712670.Search in Google Scholar
[30] Sood BC, Siddiq EA. A rapid technique for scent determination in rice. Indian J Genet Plant Breed. 1978;38(2):268–75. https://agris.fao.org/agris-search/search.do?recordID=IN19790415936.Search in Google Scholar
[31] Chandaramani P, Rajendran R, Sivasubramania P, Muthiah C. Impact of biophysical factors as influenced by organic sources of nutrients on major pests of rice. J Biopesticides. 2009;2(1):1–5, http://www.jbiopest.com/users/LW8/efiles/1-5.pdf.10.57182/jbiopestic.2.1.01-05Search in Google Scholar
[32] Bioversity International, IRRI and WARDA. Descriptors for Wild and Cultivated Rice (Oryza spp). Via dei Tre Denari, 472/a. 00057 Maccarese Rome, Italy; 2007. https://www.bioversityinternational.org/fileadmin/_migrated/uploads/tx_news/Rice_232.pdf.Search in Google Scholar
[33] Carsono N, Lukman PN, Damayanti F, Susanto U, Sari S. Identification of polymorphisms of molecular markers that supposed to be associated with high yielding on 30 rice genotypes. Chim et Nat Acta. 2014;2(1):91–5. file:///E:/download/9141-15486-1-PB.pdf. (Indonesian).10.24198/cna.v2.n1.9141Search in Google Scholar
[34] Ma X, Li F, Zhang Q, Wang X, Guo H, Xie J, et al. Genetic architecture to cause dynamic change in tiller and panicle numbers revealed by genome-wide association study and transcriptome profile in rice. Plant J. 2020;104:1603–16. 10.1111/tpj.15023.Search in Google Scholar PubMed
[35] Agustina M, Sutjahjo SH, Koesoemaningtyas T, Jagau Y. Genetic parameter estimation on upland rice agronomic characters in ultisols through diallel analysis. Hayati. 2005;12(3):98–102. 10.4308/hjb.12.3.98 (Indonesian).Search in Google Scholar
[36] Masruroh F, Samanhudi S, Yunus A. Improvement of rice (Oryza sativa L.) var. Ciherang and Cempo Ireng productivity using gamma irradiation. J Agric Sci Technol. 2016;6(5):289–94. 17265/2161-6264/2016.05.001.10.17265/2161-6264/2016.05.001Search in Google Scholar
[37] Li S, Qian Q, Fu Z, Zeng D, Meng X, Kyozuka J, et al. Short panicle 1 encodes a putative PTR family transporter and determines rice panicle size. Plant J. 2009;58:592–605. 10.1111/j.1365-313X.2009.03799.x.Search in Google Scholar PubMed
[38] Liu E, Liu Y, Wu G, Zeng S, Thi TGT, Liang L, et al. Identification of a candidate gene for panicle length in rice (Oryza sativa L.) via association and linkage analysis. Front Plant Sci. 2016;7:1–13. 10.3389/fpls.2016.00596.Search in Google Scholar PubMed PubMed Central
[39] Yan JQ, Zhu J, He CX, Benmoussa M, Wu P. Quantitative trait loci analysis for the developmental behaviour of tiller number in rice (Oryza sativa L.). Theor Appl Genet. 1998;97:267–74. 10.1007/s001220050895.Search in Google Scholar
[40] Zhu J, Zhou JY, Liu Y, Wang Z, Tan Z, Yi C, et al. Fine mapping of a major QTL controlling panicle number in rice. Mol Breed. 2011;27:171–80. 10.1007/s11032-010-9420-6.Search in Google Scholar
[41] Liu T, Li L, Zhang Y, Xu C, Li X, Xing Y. Comparison of quantitative trait loci for rice yield, panicle length and spikelet density across three connected populations. J Genet. 2011;90:377–82. 10.1007/s12041-011-0083-9.Search in Google Scholar PubMed
[42] Akinwale MG, Gregorio G, Nwilene F, Akinyele BO, Ogunbayo SA, Odiyi AC. Heritability and correlation coefficient analysis for yield and its components in rice (Oryza sativa L.). Afr J Plant Sci. 2011 March;5(3):207–12. 10.5897/AJPS.9000137.Search in Google Scholar
[43] Song Y, Gao Z, Luan W. Interaction between temperature and photoperiod in regulation of flowering time in rice. Sci China Life Sci. 2012;3:241–9. 10.1007/s11427-012-4300-4.Search in Google Scholar PubMed
[44] Katalani K, Nematzadeh G, Kiani G, Hashdemi SH. Marker assisted selection for quality improvement of Ghaem rice variety in multiple crosses at segregating population. Int J Agron Plant Prod. 2013;4(9):2420–8, https://www.researchgate.net/publication/261512329.Search in Google Scholar
[45] Jin L, Lu Y, Shao Y, Zhang G, Xiao P, Shen S, et al. Molecular marker assisted selection for improvement of the eating, cooking and sensory quality of rice (Oryza sativa L.). J Cereal Sci. 2010;51(1):159–64. 10.1016/j.jcs.2009.11.007.Search in Google Scholar
[46] Pooni HS, Kumar I, Khush GS. Genetic control of amylose content in diallel set of rice crosses. Heredity. 1993;71:603–13. 10.1038/hdy.1993.185.Search in Google Scholar
[47] Berner DK, Hoff BJ. Inheritance of scent in American long grain rice. Crop Sci. 1986;26(5):876–8. 10.2135/cropsci1986.0011183X002600050008x.Search in Google Scholar
[48] Yoshihashi T, Huong NTT, Kabaki N. Quality evaluation of Khao Dawk Mali 105 an aromatic rice cultivar of Northeast Thailand. JIRCAS Working Rep. 2002;30:151–60, https://eurekamag.com/research/003/903/003903906.php.Search in Google Scholar
[49] Singh R, Singh AK, Sharma TR, Singh A, Singh NK. Fine mapping of aroma QTLs in Basmati rice (Oryza sativa L) on chromosomes 3, 4 and 8. J Plant Biochem Biotechnol. 2007;16(2):75–82. 10.1007/BF03321978.Search in Google Scholar
[50] Amarawathi Y, Singh R, Singh AK, Singh VP, Mohapatra T, Sharma TR, et al. Mapping of quantitative trait loci for basmati quality traits in rice (Oryza sativa L.). Mol Breed. 2008;21:49–65. 10.1007/s11032-007-9108-8.Search in Google Scholar
[51] Bradbury LMT, Fitzgerald TL, Henry RJ, Jin Q, Waters DLE. The gene for fragrance in rice. Plant Biotechnol J. 2005;3(3):363–70. 10.1111/j.1467-7652.2005.00131.x.Search in Google Scholar PubMed
[52] Bounphanousay C, Jaisil P, Sanitchon J, Fitzgerald M, Hamilton NRS. Chemical and molecular characterization of fragrance in black glutinous rice from Loa PDR. Asian J Plant Sci. 2008;7(1):1–7. 10.3923/ajps.2008.1.7.Search in Google Scholar
[53] Bradbury LMT, Henry RJ, Jin Q, Reinke RF, Waters DLE. A perfect marker for fragrance genotyping in rice. Mol Breed. 2005;16:279–83. 10.1007/s11032-005-0776-y.Search in Google Scholar
[54] Sarhadi WA, Hien NL, Zanjani M, Yosofzai W, Yoshihashi T. Comparative analyses for aroma and agronomic traits of native rice cultivars from Central Asia. J Crop Sci Biotech. 2002;11(1):17–22. https://www.researchgate.net/publication/241070155_Comparative_Analyses_for_Aroma_and_Agronomic_Traits_of_Native_Rice_Cultivars_from_Central_Asia.Search in Google Scholar
[55] Champagne ET. Rice aroma and flavour: a literature review. Cereal Chem. 2008;85(4):445–54. 10.1094/CCHEM-85-4-0445.Search in Google Scholar
[56] Bradbury LMT, Gillies SA, Brushett DJ, Waters DLE, Henry RJ. Inactivation of an aminoaldehyde dehydrogenase is responsible for fragrance in rice. Plant Mol Biol. 2008;68:439–49. 10.1007/s11103-008-9381-x.Search in Google Scholar PubMed
[57] Itani T, Tamaki M, Hayata Y, Fushimi T, Hashizume K. Variation of 2-Acetyl-1-Pyrroline concentration in aromatic rice grains collected in the same region in Japan and factors affecting its concentration. Plant Prod Sci. 2004;7(2):178–83. 10.1626/pps.7.178.Search in Google Scholar
[58] Wang Y, Wang X, Yuan H, Chen R, Zhu L, He R, et al. Responses of two contrasting genotypes of rice to brown planthopper. Mol Plant Microbe Interact. 2008;21(1):122–32. 10.1094/MPMI-21-1-0122.Search in Google Scholar PubMed
[59] Prayoga GI. Physiological Trait Analysis of Parents and Application of Molecular Marker Selection (MAS) on F2 Progeny in Assembling of Brown Planthopper Resistance Rice Promising Lines. Thesis; 2013. Unpublished. Indonesian.Search in Google Scholar
[60] Baehaki SE, Munawar D. Biotype testing for brown planthopper, Nilaparvata lugens STAL on rice growing area. Rice Natl Semin. 2008;347–59. http://www.litbang.pertanian.go.id/special/padi/bbpadi_2008_prosb132.pdf Indonesian.Search in Google Scholar
[61] Sun L, Su C, Wang C, Zai H, Wan J. Mapping of a major resistance gene to brown planthopper in the rice cultivar Rathu Heenati. Breed Sci. 2005;55(4):391–6. 10.1270/jsbbs.55.391.Search in Google Scholar
[62] Maqbool SB, Riazuddin S, Loc NT, Gatehouse AMR, Gatehouse JA, Christou P. Expression of multiple insecticidal genes confers broad resistance against a range of different rice pests. Mol Breed. 2001;7(1):85–93. 10.1023/A:1009644712157.Search in Google Scholar
[63] Zeng YH, Zhu YS, Lian L, Xie HG, Zhang JF, Xie HA. Genetic analysis and fine mapping of the pubescence gene GL6 in rice (Oryza sativa L.). Chin Sci Bull. 2013;58:2992–9. 10.1007/s11434-013-5737-y.Search in Google Scholar
[64] Qin BX, Tang D, Huang J, Li M, Wu XR, Lu LL, et al. Rice OsGL1-1 is involved in leaf cuticular wax and cuticle membrane. Mol Plant. 2011;4(6):985–95. 10.1093/mp/ssr028.Search in Google Scholar PubMed
© 2023 the author(s), published by De Gruyter
This work is licensed under the Creative Commons Attribution 4.0 International License.
Articles in the same Issue
- Regular Articles
- The impact of COVID-19 pandemic on business risks and potato commercial model
- Effects of potato (Solanum tuberosum L.)–Mucuna pruriens intercropping pattern on the agronomic performances of potato and the soil physicochemical properties of the western highlands of Cameroon
- Machine learning-based prediction of total phenolic and flavonoid in horticultural products
- Revamping agricultural sector and its implications on output and employment generation: Evidence from Nigeria
- Does product certification matter? A review of mechanism to influence customer loyalty in the poultry feed industry
- Farmer regeneration and knowledge co-creation in the sustainability of coconut agribusiness in Gorontalo, Indonesia
- Lablab purpureus: Analysis of landraces cultivation and distribution, farming systems, and some climatic trends in production areas in Tanzania
- The effects of carrot (Daucus carota L.) waste juice on the performances of native chicken in North Sulawesi, Indonesia
- Properties of potassium dihydrogen phosphate and its effects on plants and soil
- Factors influencing the role and performance of independent agricultural extension workers in supporting agricultural extension
- The fate of probiotic species applied in intensive grow-out ponds in rearing water and intestinal tracts of white shrimp, Litopenaeus vannamei
- Yield stability and agronomic performances of provitamin A maize (Zea mays L.) genotypes in South-East of DR Congo
- Diallel analysis of length and shape of rice using Hayman and Griffing method
- Physicochemical and microbiological characteristics of various stem bark extracts of Hopea beccariana Burck potential as natural preservatives of coconut sap
- Correlation between descriptive and group type traits in the system of cow’s linear classification of Ukrainian Brown dairy breed
- Meta-analysis of the influence of the substitution of maize with cassava on performance indices of broiler chickens
- Bacteriocin-like inhibitory substance (BLIS) produced by Enterococcus faecium MA115 and its potential use as a seafood biopreservative
- Meta-analysis of the benefits of dietary Saccharomyces cerevisiae intervention on milk yield and component characteristics in lactating small ruminants
- Growth promotion potential of Bacillus spp. isolates on two tomato (Solanum lycopersicum L.) varieties in the West region of Cameroon
- Prioritizing IoT adoption strategies in millennial farming: An analytical network process approach
- Soil fertility and pomelo yield influenced by soil conservation practices
- Soil macrofauna under laying hens’ grazed fields in two different agroecosystems in Portugal
- Factors affecting household carbohydrate food consumption in Central Java: Before and during the COVID-19 pandemic
- Properties of paper coated with Prunus serotina (Ehrh.) extract formulation
- Fertiliser cost prediction in European Union farms: Machine-learning approaches through artificial neural networks
- Molecular and phenotypic markers for pyramiding multiple traits in rice
- Natural product nanofibers derived from Trichoderma hamatum K01 to control citrus anthracnose caused by Colletotrichum gloeosporioides
- Role of actors in promoting sustainable peatland management in Kubu Raya Regency, West Kalimantan, Indonesia
- Small-scale coffee farmers’ perception of climate-adapted attributes in participatory coffee breeding: A case study of Gayo Highland, Aceh, Indonesia
- Optimization of extraction using surface response methodology and quantification of cannabinoids in female inflorescences of marijuana (Cannabis sativa L.) at three altitudinal floors of Peru
- Production factors, technical, and economic efficiency of soybean (Glycine max L. Merr.) farming in Indonesia
- Economic performance of smallholder soya bean production in Kwara State, Nigeria
- Indonesian rice farmers’ perceptions of different sources of information and their effect on farmer capability
- Feed preference, body condition scoring, and growth performance of Dohne Merino ram fed varying levels of fossil shell flour
- Assessing the determinant factors of risk strategy adoption to mitigate various risks: An experience from smallholder rubber farmers in West Kalimantan Province, Indonesia
- Analysis of trade potential and factors influencing chili export in Indonesia
- Grade-C kenaf fiber (poor quality) as an alternative material for textile crafts
- Technical efficiency changes of rice farming in the favorable irrigated areas of Indonesia
- Palm oil cluster resilience to enhance indigenous welfare by innovative ability to address land conflicts: Evidence of disaster hierarchy
- Factors determining cassava farmers’ accessibility to loan sources: Evidence from Lampung, Indonesia
- Tailoring business models for small-medium food enterprises in Eastern Africa can drive the commercialization and utilization of vitamin A rich orange-fleshed sweet potato puree
- Revitalizing sub-optimal drylands: Exploring the role of biofertilizers
- Effects of salt stress on growth of Quercus ilex L. seedlings
- Design and fabrication of a fish feed mixing cum pelleting machine for small-medium scale aquaculture industry
- Indicators of swamp buffalo business sustainability using partial least squares structural equation modelling
- Effect of arbuscular mycorrhizal fungi on early growth, root colonization, and chlorophyll content of North Maluku nutmeg cultivars
- How intergenerational farmers negotiate their identity in the era of Agriculture 4.0: A multiple-case study in Indonesia
- Responses of broiler chickens to incremental levels of water deprivation: Growth performance, carcass characteristics, and relative organ weights
- The improvement of horticultural villages sustainability in Central Java Province, Indonesia
- Effect of short-term grazing exclusion on herbage species composition, dry matter productivity, and chemical composition of subtropical grasslands
- Analysis of beef market integration between consumer and producer regions in Indonesia
- Analysing the sustainability of swamp buffalo (Bubalus bubalis carabauesis) farming as a protein source and germplasm
- Toxicity of Calophyllum soulattri, Piper aduncum, Sesamum indicum and their potential mixture for control Spodoptera frugiperda
- Consumption profile of organic fruits and vegetables by a Portuguese consumer’s sample
- Phenotypic characterisation of indigenous chicken in the central zone of Tanzania
- Diversity and structure of bacterial communities in saline and non-saline rice fields in Cilacap Regency, Indonesia
- Isolation and screening of lactic acid bacteria producing anti-Edwardsiella from the gastrointestinal tract of wild catfish (Clarias gariepinus) for probiotic candidates
- Effects of land use and slope position on selected soil physicochemical properties in Tekorsh Sub-Watershed, East Gojjam Zone, Ethiopia
- Design of smart farming communication and web interface using MQTT and Node.js
- Assessment of bread wheat (Triticum aestivum L.) seed quality accessed through different seed sources in northwest Ethiopia
- Estimation of water consumption and productivity for wheat using remote sensing and SEBAL model: A case study from central clay plain Ecosystem in Sudan
- Agronomic performance, seed chemical composition, and bioactive components of selected Indonesian soybean genotypes (Glycine max [L.] Merr.)
- The role of halal requirements, health-environmental factors, and domestic interest in food miles of apple fruit
- Subsidized fertilizer management in the rice production centers of South Sulawesi, Indonesia: Bridging the gap between policy and practice
- Factors affecting consumers’ loyalty and purchase decisions on honey products: An emerging market perspective
- Inclusive rice seed business: Performance and sustainability
- Design guidelines for sustainable utilization of agricultural appropriate technology: Enhancing human factors and user experience
- Effect of integrate water shortage and soil conditioners on water productivity, growth, and yield of Red Globe grapevines grown in sandy soil
- Synergic effect of Arbuscular mycorrhizal fungi and potassium fertilizer improves biomass-related characteristics of cocoa seedlings to enhance their drought resilience and field survival
- Control measure of sweet potato weevil (Cylas formicarius Fab.) (Coleoptera: Curculionidae) in endemic land of entisol type using mulch and entomopathogenic fungus Beauveria bassiana
- In vitro and in silico study for plant growth promotion potential of indigenous Ochrobactrum ciceri and Bacillus australimaris
- Effects of repeated replanting on yield, dry matter, starch, and protein content in different potato (Solanum tuberosum L.) genotypes
- Review Articles
- Nutritional and chemical composition of black velvet tamarind (Dialium guineense Willd) and its influence on animal production: A review
- Black pepper (Piper nigrum Lam) as a natural feed additive and source of beneficial nutrients and phytochemicals in chicken nutrition
- The long-crowing chickens in Indonesia: A review
- A transformative poultry feed system: The impact of insects as an alternative and transformative poultry-based diet in sub-Saharan Africa
- Short Communication
- Profiling of carbonyl compounds in fresh cabbage with chemometric analysis for the development of freshness assessment method
- Special Issue of The 4th International Conference on Food Science and Engineering (ICFSE) 2022 - Part I
- Non-destructive evaluation of soluble solid content in fruits with various skin thicknesses using visible–shortwave near-infrared spectroscopy
- Special Issue on FCEM - International Web Conference on Food Choice & Eating Motivation - Part I
- Traditional agri-food products and sustainability – A fruitful relationship for the development of rural areas in Portugal
- Consumers’ attitudes toward refrigerated ready-to-eat meat and dairy foods
- Breakfast habits and knowledge: Study involving participants from Brazil and Portugal
- Food determinants and motivation factors impact on consumer behavior in Lebanon
- Comparison of three wine routes’ realities in Central Portugal
- Special Issue on Agriculture, Climate Change, Information Technology, Food and Animal (ACIFAS 2020)
- Environmentally friendly bioameliorant to increase soil fertility and rice (Oryza sativa) production
- Enhancing the ability of rice to adapt and grow under saline stress using selected halotolerant rhizobacterial nitrogen fixer
Articles in the same Issue
- Regular Articles
- The impact of COVID-19 pandemic on business risks and potato commercial model
- Effects of potato (Solanum tuberosum L.)–Mucuna pruriens intercropping pattern on the agronomic performances of potato and the soil physicochemical properties of the western highlands of Cameroon
- Machine learning-based prediction of total phenolic and flavonoid in horticultural products
- Revamping agricultural sector and its implications on output and employment generation: Evidence from Nigeria
- Does product certification matter? A review of mechanism to influence customer loyalty in the poultry feed industry
- Farmer regeneration and knowledge co-creation in the sustainability of coconut agribusiness in Gorontalo, Indonesia
- Lablab purpureus: Analysis of landraces cultivation and distribution, farming systems, and some climatic trends in production areas in Tanzania
- The effects of carrot (Daucus carota L.) waste juice on the performances of native chicken in North Sulawesi, Indonesia
- Properties of potassium dihydrogen phosphate and its effects on plants and soil
- Factors influencing the role and performance of independent agricultural extension workers in supporting agricultural extension
- The fate of probiotic species applied in intensive grow-out ponds in rearing water and intestinal tracts of white shrimp, Litopenaeus vannamei
- Yield stability and agronomic performances of provitamin A maize (Zea mays L.) genotypes in South-East of DR Congo
- Diallel analysis of length and shape of rice using Hayman and Griffing method
- Physicochemical and microbiological characteristics of various stem bark extracts of Hopea beccariana Burck potential as natural preservatives of coconut sap
- Correlation between descriptive and group type traits in the system of cow’s linear classification of Ukrainian Brown dairy breed
- Meta-analysis of the influence of the substitution of maize with cassava on performance indices of broiler chickens
- Bacteriocin-like inhibitory substance (BLIS) produced by Enterococcus faecium MA115 and its potential use as a seafood biopreservative
- Meta-analysis of the benefits of dietary Saccharomyces cerevisiae intervention on milk yield and component characteristics in lactating small ruminants
- Growth promotion potential of Bacillus spp. isolates on two tomato (Solanum lycopersicum L.) varieties in the West region of Cameroon
- Prioritizing IoT adoption strategies in millennial farming: An analytical network process approach
- Soil fertility and pomelo yield influenced by soil conservation practices
- Soil macrofauna under laying hens’ grazed fields in two different agroecosystems in Portugal
- Factors affecting household carbohydrate food consumption in Central Java: Before and during the COVID-19 pandemic
- Properties of paper coated with Prunus serotina (Ehrh.) extract formulation
- Fertiliser cost prediction in European Union farms: Machine-learning approaches through artificial neural networks
- Molecular and phenotypic markers for pyramiding multiple traits in rice
- Natural product nanofibers derived from Trichoderma hamatum K01 to control citrus anthracnose caused by Colletotrichum gloeosporioides
- Role of actors in promoting sustainable peatland management in Kubu Raya Regency, West Kalimantan, Indonesia
- Small-scale coffee farmers’ perception of climate-adapted attributes in participatory coffee breeding: A case study of Gayo Highland, Aceh, Indonesia
- Optimization of extraction using surface response methodology and quantification of cannabinoids in female inflorescences of marijuana (Cannabis sativa L.) at three altitudinal floors of Peru
- Production factors, technical, and economic efficiency of soybean (Glycine max L. Merr.) farming in Indonesia
- Economic performance of smallholder soya bean production in Kwara State, Nigeria
- Indonesian rice farmers’ perceptions of different sources of information and their effect on farmer capability
- Feed preference, body condition scoring, and growth performance of Dohne Merino ram fed varying levels of fossil shell flour
- Assessing the determinant factors of risk strategy adoption to mitigate various risks: An experience from smallholder rubber farmers in West Kalimantan Province, Indonesia
- Analysis of trade potential and factors influencing chili export in Indonesia
- Grade-C kenaf fiber (poor quality) as an alternative material for textile crafts
- Technical efficiency changes of rice farming in the favorable irrigated areas of Indonesia
- Palm oil cluster resilience to enhance indigenous welfare by innovative ability to address land conflicts: Evidence of disaster hierarchy
- Factors determining cassava farmers’ accessibility to loan sources: Evidence from Lampung, Indonesia
- Tailoring business models for small-medium food enterprises in Eastern Africa can drive the commercialization and utilization of vitamin A rich orange-fleshed sweet potato puree
- Revitalizing sub-optimal drylands: Exploring the role of biofertilizers
- Effects of salt stress on growth of Quercus ilex L. seedlings
- Design and fabrication of a fish feed mixing cum pelleting machine for small-medium scale aquaculture industry
- Indicators of swamp buffalo business sustainability using partial least squares structural equation modelling
- Effect of arbuscular mycorrhizal fungi on early growth, root colonization, and chlorophyll content of North Maluku nutmeg cultivars
- How intergenerational farmers negotiate their identity in the era of Agriculture 4.0: A multiple-case study in Indonesia
- Responses of broiler chickens to incremental levels of water deprivation: Growth performance, carcass characteristics, and relative organ weights
- The improvement of horticultural villages sustainability in Central Java Province, Indonesia
- Effect of short-term grazing exclusion on herbage species composition, dry matter productivity, and chemical composition of subtropical grasslands
- Analysis of beef market integration between consumer and producer regions in Indonesia
- Analysing the sustainability of swamp buffalo (Bubalus bubalis carabauesis) farming as a protein source and germplasm
- Toxicity of Calophyllum soulattri, Piper aduncum, Sesamum indicum and their potential mixture for control Spodoptera frugiperda
- Consumption profile of organic fruits and vegetables by a Portuguese consumer’s sample
- Phenotypic characterisation of indigenous chicken in the central zone of Tanzania
- Diversity and structure of bacterial communities in saline and non-saline rice fields in Cilacap Regency, Indonesia
- Isolation and screening of lactic acid bacteria producing anti-Edwardsiella from the gastrointestinal tract of wild catfish (Clarias gariepinus) for probiotic candidates
- Effects of land use and slope position on selected soil physicochemical properties in Tekorsh Sub-Watershed, East Gojjam Zone, Ethiopia
- Design of smart farming communication and web interface using MQTT and Node.js
- Assessment of bread wheat (Triticum aestivum L.) seed quality accessed through different seed sources in northwest Ethiopia
- Estimation of water consumption and productivity for wheat using remote sensing and SEBAL model: A case study from central clay plain Ecosystem in Sudan
- Agronomic performance, seed chemical composition, and bioactive components of selected Indonesian soybean genotypes (Glycine max [L.] Merr.)
- The role of halal requirements, health-environmental factors, and domestic interest in food miles of apple fruit
- Subsidized fertilizer management in the rice production centers of South Sulawesi, Indonesia: Bridging the gap between policy and practice
- Factors affecting consumers’ loyalty and purchase decisions on honey products: An emerging market perspective
- Inclusive rice seed business: Performance and sustainability
- Design guidelines for sustainable utilization of agricultural appropriate technology: Enhancing human factors and user experience
- Effect of integrate water shortage and soil conditioners on water productivity, growth, and yield of Red Globe grapevines grown in sandy soil
- Synergic effect of Arbuscular mycorrhizal fungi and potassium fertilizer improves biomass-related characteristics of cocoa seedlings to enhance their drought resilience and field survival
- Control measure of sweet potato weevil (Cylas formicarius Fab.) (Coleoptera: Curculionidae) in endemic land of entisol type using mulch and entomopathogenic fungus Beauveria bassiana
- In vitro and in silico study for plant growth promotion potential of indigenous Ochrobactrum ciceri and Bacillus australimaris
- Effects of repeated replanting on yield, dry matter, starch, and protein content in different potato (Solanum tuberosum L.) genotypes
- Review Articles
- Nutritional and chemical composition of black velvet tamarind (Dialium guineense Willd) and its influence on animal production: A review
- Black pepper (Piper nigrum Lam) as a natural feed additive and source of beneficial nutrients and phytochemicals in chicken nutrition
- The long-crowing chickens in Indonesia: A review
- A transformative poultry feed system: The impact of insects as an alternative and transformative poultry-based diet in sub-Saharan Africa
- Short Communication
- Profiling of carbonyl compounds in fresh cabbage with chemometric analysis for the development of freshness assessment method
- Special Issue of The 4th International Conference on Food Science and Engineering (ICFSE) 2022 - Part I
- Non-destructive evaluation of soluble solid content in fruits with various skin thicknesses using visible–shortwave near-infrared spectroscopy
- Special Issue on FCEM - International Web Conference on Food Choice & Eating Motivation - Part I
- Traditional agri-food products and sustainability – A fruitful relationship for the development of rural areas in Portugal
- Consumers’ attitudes toward refrigerated ready-to-eat meat and dairy foods
- Breakfast habits and knowledge: Study involving participants from Brazil and Portugal
- Food determinants and motivation factors impact on consumer behavior in Lebanon
- Comparison of three wine routes’ realities in Central Portugal
- Special Issue on Agriculture, Climate Change, Information Technology, Food and Animal (ACIFAS 2020)
- Environmentally friendly bioameliorant to increase soil fertility and rice (Oryza sativa) production
- Enhancing the ability of rice to adapt and grow under saline stress using selected halotolerant rhizobacterial nitrogen fixer