Bacteriocin-like inhibitory substance (BLIS) produced by Enterococcus faecium MA115 and its potential use as a seafood biopreservative
-
 ,
,

Abstract
Introduction: Listeria monocytogenes is an opportunistic foodborne pathogen that can cause death to infected humans. The bacterial pathogen was frequently associated with food-borne origins including seafood. Thus, the present study aimed at screening anti-listeria-producing lactic acid bacteria (AL-LAB) isolated from the intestinal tracts of wild gilt-head seabream (Sparus aurata).
Materials and Methods: Lactic acid bacteria (LAB) were isolated from the intestinal tracts of 15 wild gilt-head seabreams and screened for antimicrobial activity against Listeria monocytogenes. LAB isolated with the strongest anti-listerial activity was identified and further characterized for its anti-listerial compounds, followed by testing its protecting capacity on experimentally contaminated salmon fillets.
Results: Three out of 52 LAB showed antagonistic activities against the foodborne pathogen. Of these 3, an isolate with the strongest inhibitory activity was obtained from strain MA115 (>400 AU). Based on its 16S rDNA sequence, strain MA115 had 99% similarity to Enterococcus faecium (Acc. Nb: MG461637.1). Further in vitro assays showed that the anti-listerial compound was very sensitive to proteinase K which suggested that it is a bacteriocin-like inhibitory substance (BLIS). In addition, the BLIS was resistant to a wide range of pH (2–10), and low (4 to −20°C) as well as high temperatures (30−121°C). In addition, the BLIS showed bactericidal activity on L. monocytogenes in artificially contaminated Salmon fillets stored at 0°C.
Conclusion: The BLIS produced by Enterococcus faecium strain MA115 has the potential for application to improve food safety, although further studies are still required to specifically identify the BLIS, for biopreservation purposes.
1 Introduction
Listeria monocytogenes is an opportunistic foodborne pathogen that can cause death to infected humans. Several studies reported that this bacterial pathogen was frequently associated with food-borne origins including seafood [1]. In seafood, L. monocytogenes has been commonly reported from frozen and raw salmon fillets, crab meat, shellfish, and shrimp [2]. According to the Center for disease control and prevention (CDC), approximately 1,600 cases of listeriosis have been reported yearly until 2015 in the United States, of which 260 cases resulted in death (https://www.cdc.gov/). The bacterial pathogen mostly infects adults aged 65 or older, and pregnant women and their newborns. Acknowledging the hazardous effect of this pathogen, the Codex Alimentarius Commission has set a zero count for products with favorable conditions for the pathogen growth and 100 CFU g−1 for foods with unfavorable growth environments [3]. However, achieving a zero count of L. monocytogenes is difficult, and remains a challenge because of the pathogen’s ability to survive and persist even in several environmental stressors such as low (−0.1 to −0.4°C) and a high temperature (71°C), low pH, and smoked as well as brined food products [4].
Various strategies have been developed to control the incidence of L. monocytogenes in seafood including chemicals and bacteriocins [5]. In general, bacteriocins have gained the most interest due to: being considered as natural biopreservatives that are generally recognized as safe (GRAS) substances, especially bacteriocins produced by lactic acid bacteria (LAB). They are inactive and nontoxic to eukaryote cells, pH and heat tolerant, and have bactericidal activity against bacterial pathogens [6]. Currently, nisin and a class II bacteriocin (pediocin ALTA 2341®) are bacteriocins that are available commercially to control L. monocytogenes [7]. However, these bacteriocins have been questioned due to their low solubility and instability in high pH values [8,9]. Accordingly, other studies confirmed that nisin has been reported to be unable to inhibit the growth of L. monocytogenes due to its sensitivity to acidic and basic environments and salinity [10,11]. In addition, some studies have reported the emergence of Listeria strains which are resistant to nisin, and some class II bacteriocins [12]. Thus, new types of bacteriocins with more stability in different environments are required to control L. monocytogenes in food products.
This study aimed at screening anti-listerial compounds from LAB strains isolated from the intestinal tracts of wild gilt-head seabream (Sparus aurata). The anti-listerial compound was further characterized by investigating susceptibility to proteinase K, pH, and heat as well as determining their molecular size. In addition, the capacity of the anti-listerial substances to suppress the growth or kill L. monocytogenes was studied in experimentally contaminated salmon fillets.
2 Materials and method
2.1 Isolation of LAB
LAB were isolated from intestinal tracts of gilt-head seabreams (Sparus aurata) as previously described by Amin [13]. Pure bacterial isolates were phenotypically tested, and only those isolates which were gram-positive, catalase-negative, oxidase-negative, and able to ferment glucose were considered as members of LAB. The purified LAB were stored in MRS broth (CM0359, Oxoid) supplemented with 15% glycerol at −20°C until further use.
-
Ethical approval: The research related to animal use has been complied with all the relevant national regulations and institutional policies for the care and use of animals.
2.2 Preparation of neutralized cell-free supernatant (CFSn)
CFSn of each LAB was prepared as previously described by Amin et al. [14]. The sterilized supernatant obtained from each tested LAB was afterward kept in a refrigerator (4°C) until further use.
2.3 Preparation of L. monocytogenes inoculum
L. monocytogenes was obtained from the Microbiology Laboratory, Faculty of Health Sciences and Community Care, University of Tasmania, Newnham campus. The isolate was originally human isolates (Storage code, respectively) stored as coated beads in a cryogenic freezer at –80°C. Then, the following steps were performed according to a protocol described by Amin et al. [14]. Cell concentration of the targeted pathogen was set at ∼1.0 × 106 CFU ml−1 cell concentration for further screening assay.
2.4 Screening for anti-listerial activity
The screening for anti-listerial activity was performed according to a protocol previously described by Amin et al. [14]. The CFSn exhibiting the widest clearance zone was chosen for further assays.
In addition, the activity of anti-listeria was performed by measuring arbitrary units (AUs) according to a protocol of Saeed et al. [15] with slight modification. CFSn from the LAB isolate with the largest inhibition zone was diluted 1–5 fold with phosphate-buffered saline (PBS, pH 7.2). The anti-listerial activity from each dilution was determined using the agar well diffusion assay, and an AU was calculated according to the below formula [15].
where AU is an arbitrary unit (AU ml−1), V is the volume of CFSn used, and D is the highest dilution that still inhibited the cell growth of L. monocytogenes.
2.5 Susceptibility of the BLIS to proteinase K, heat, pH, and storage condition
The susceptibility of anti-listeria compounds to proteinase K was evaluated according to a modified protocol of Abrams et al. [16] with slight modification. Briefly, the CFSn solution from the selected LAB was mixed with proteinase K (Tritirachium, Sigma-Aldrich: 1 mg ml−1 final concentration) for 1 h. The remained anti-listerial activity was measured by pipetting 200 µl containing ∼ 1.0 × 106 CFU ml−1 into wells of a 96 well plate, followed by the addition of 10 µl of proteinase K-treated CFSn. Each treatment had four replicate wells including a control which was only the pathogen solution without CFSn addition. Afterward, the pathogen growth (L. monocytogenes) was measured by a microplate reader (Tecan Infinite M200 Pro) at OD600 nm every hour for 15 h.
The effect of temperature on the antimicrobial compound was evaluated according to a modified method by Vidhyasagar and Jeevaratnam [17]. In brief, the CFSn was heated for 30 min at different temperatures of 30, 50, 80, and 100°C, or for 15 min at 121°C by autoclaving. In addition, the effect of pH on CFSn stability was tested by adjusting the CFSn to pH values of 2, 4, 6, 8, and 10 using either 1 M hydrochloric acid (HCL) or 1 M NaOH. Thereafter, all CFSn were incubated at 37°C for 1 h. Afterward, the antimicrobial activity of CFSn at different values of pH and temperatures was evaluated against the foodborne pathogen using the microtiter plate assay as previously described.
In addition, the effect of storage conditions on the antimicrobial compounds was assessed by storing the CFSn at 4°C and −20°C for 2 weeks. Then, the residual antimicrobial activity was determined using the microtiter plate assay as previously described.
2.5.1 Identification of LAB
Each LAB was sub cultured in MRS agar and a single pure colony was picked with a micropipette tip as DNA template and added into 30 µl of master mix. The master mix consisted of 4.0 µl of 5× HF buffer, 2 µl of MgCl2, 2.0 µl of 10 µM deoxynucleotide triphosphate (dNTP), 0.3 µl of 4 U µl−1 Taq polymerase, and 17 µl of sterilized milliQ water, and 2.0 µl of 10 µM of each universal forward and reverse primers. The PCR amplification was performed using the universal primers: forward primer (27 F), and reverse primer (1492 R) [18]. DNA amplification was performed in a PCR thermocycler with a protocol previously described by Amin [13]. PCR products were purified using Promega DNA purification kit according to the manufacturer’s protocol, and 15 µl of the purified DNA fragments (∼50 ng l−1) were sent for sequencing. The sequenced isolate was aligned to published sequences using the Basic Local Alignment Search Tool (BLAST) to find the closest-known species. In addition, a phylogenetic tree was constructed between the sequenced isolate of anti-listerial-producing LAB strains and several closest known species derived from GeneBank databases using a Joining–neighboring method of Genious software version 5.3.6.
2.6 Control of L. monocytogenes in experimentally contaminated salmon fillets
Six kilograms of freshly harvested Atlantic salmon (Salmo salar L.) was purchased from the Van Diemen aquaculture farm and transported in ice within 2 h of the harvest to the laboratory for fillet processing. The fish was filleted aseptically and then cut into 50 ± 0.5 g pieces. Then, replicate salmon fillets were immersed into CFSn for 5 s, while sterile MRS broth was used as the control. Thereafter, each fillet was inserted into a sterile resealable plastic bag and sprayed ten times (∼1 ml) with L. monocytogenes inoculum (∼1.0 × 106 CFU ml−1) before storing at 0°C for 6 days. Enumeration of L. monocytogenes was carried out on day 0, and day 6 by weighing 20 g of salmon fillet and mixing it with 180 ml of bacteriological peptone water (LP0037, Oxoid), and homogenizing in a stomacher (Lab Blender400) for 30 s. Afterward, 5-fold dilutions were made and 0.1 ml of appropriate dilution was plated in duplicate on Listeria selective agar (CM0856, Oxoid) with Listeria Selective Supplement (SR0140, Oxoid). Dark colonies were counted as L. monocytogenes after 48 h of incubation at 37°C.
2.7 Data analysis
The diameters of clearance zones observed using a well-diffusion assay from three LAB isolates, and the growth of L. monocytogenes obtained using a microtiter plate assay (in terms of OD600nm values) after the CFSn treatments which were previously treated with heat, pH proteinase K, and storage temperatures were compared using one-way analysis of variance (ANOVA), with a Tukey post hoc test. Viable counts of L. monocytogenes in salmon fillets at day 0 and day 6 were statistically compared using a paired sample t-test, both with SPSS software version 22.
3 Results
3.1 LAB isolation and selection of strains with anti-listerial activity
A total of 52 bacterial isolates were identified as members of LAB due to being catalase-negative, oxidase-negative, gram-positive, rod or cocci in cell shapes, and were able to ferment glucose. Out of these isolates, three strains displayed antagonistic activity against L. monocytogenes, as indicated by >10 mm diameter inhibition zones (Figure 1).

Inhibition zones of CFSn extracted from three LAB strains on the surface of MH agar plate layered with L. monocytogenes using the agar well diffusion assay after incubation at 37°C for 24 h. The CFSn of three LAB isolates with anti-listerial activity: isolate MA115, isolate MA114, and isolate MA111, and sterilized MRS broth as control (Ctrl). R1 and R2 are replicates of each CFSn.
Among these, the strongest anti-listerial activity was observed from LAB strain MA115, with >400 AU ml−1. Based on the partial sequence of 16S rDNA, LAB strain MA115 showed 99% similarity to Enterococcus faecium strain GR7. This bacterial 16S rRNA gene sequence has been deposited in the National Center for Biotechnology Information with GenBank accession no: MG461637.1. Phylogenetic evaluation based on the partial sequence of the 16S rRNA gene also indicated that the isolate MA115 was very close to E. faecium GR7 confirming the previous result obtained from the BLAST function (Figure 2).

Phylogenetic tree of anti-listeria compound producing E. faecium MA115 isolated from the gastrointestinal tract of gilthead seabream. Bracket (NR_═) after the species name is the accession number in the gene bank.
3.2 Characterization of anti-listerial compound
The anti-listerial compounds in the CFSn of E. faecium MA115 were highly susceptible to proteinase K, (Table 1), but remained active at wide ranges of pH (2, 4, 6, 8, or 10), indicated by no growth of L. monocytogenes grown in the microtiter assay for 24 h, Table 1. The high sensitivity of anti-listerial compound in CFSn indicate that the E. facium MA115 produced bacteiocin-like inhibitory substance (BLIS). Furthermore, when heated at 30, 50, 80, and 100°C for 30 min, or 121°C for 15 min, the BLIS of E. faecium strain MA115 maintained its anti-listerial activity, confirmed by no growth of L. monocytogenes in the microtiter plate assay, Table 1. In addition, the anti-listerial compound of the BLIS remained stable after being stored at 4°C for 2 weeks (OD of L. monocytogenes was 0.16 ± 0.02 at 0 h and 0.14 ± 0.001 after 15 h at room temperature), Table 1. Another result showed that, the anti-listerial activity of BLIS decreased after being stored at −20°C for 2 weeks, indicated by a slight increase in the OD value of L. monocytogenes (from 0.17 ± 0.01 at 0 h to 0.25 ± 0.01 after 15 h incubation), Table 1. However, the OD values of L. monocytogenes treated with −20°C − stored BLIS were still significantly lower compared to the OD values of L. monocytogenes in the control from 0.17 ± 0.01 at 0 h to 0.45 ± 0.03 after 15 h incubation, F = 277.8, df = 2.12, and p < 0.001.
Effect of proteinase K, pH, and temperatures on the anti-listeria activity of BLIS produced by E. faecium MA115
| Treatments and incubation time | Anti-listeria activity |
|---|---|
| Proteinase K/1 h | — |
| pH | |
|
++ |
|
++ |
|
++ |
|
++ |
|
++ |
| High temperatures | |
|
++ |
|
++ |
|
++ |
|
++ |
|
++ |
| Low temperatures | |
|
++ |
|
+ |
− anti-listeria activity was completely inactivated. ++ anti-listeria activity remained stable indicated by no growth of L. monocytogenes after being treated with the BLIS. + BLIS activity decreased slightly, indicated by the increase in OD value at 600 nm which represents the growth of L. monocytogenes. However, the growth of the pathogen was still significantly lower than the growth in the control (p < 0.05).
3.3 Control of L. monocytogenes in experimentally contaminated salmon fillets
The BLIS of E. faecium MA115 showed bactericidal activity on L. monocytogenes as indicated by a significant reduction in the pathogen’s viable cells on salmon fillets after treatment (Figure 3). In the absence of the BLIS, L. monocytogenes was able to survive on the salmon fillets stored at 0°C for 6 days, indicated by no significant difference in the number of L. monocytogenes cells (4.27 log units at day 0 and 4.35 log units at day 6 (t = 0.42, df = 3, p = 0.71)). However, viable cells of L. monocytogenes in the salmon fillets after 5 s immersion in the neutralized BLIS of E. faecium MA115 significantly decreased after 6 days (t = 9.95, df = 3, p = 0.002). The viable cells of L. monocytogenes were recorded at 4.22 log units at day 0 to only 0.46 log units at day 6 (0°C). This shows that the BLIS produced by E. faecium was bactericidal to the foodborne pathogen.
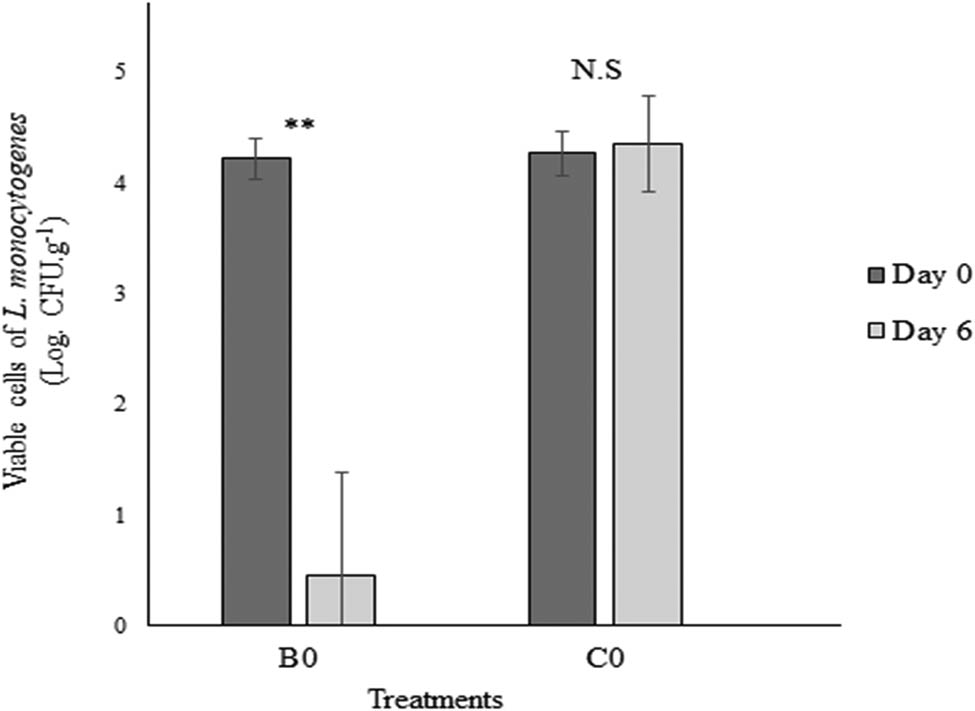
Viability of L. monocytogenes in Atlantic salmon fillets after immersion in BLIS liquid and stored at 0°C. B0 was salmon fillets immersed in BLIS, inoculated with L. monocytogenes, and incubated at 0°C, C0 was a control in which the salmon fillets were immersed in sterile MRS broth prior to inoculation. ** represents a significant difference between TVC at day 0 and after 6 days stored at 0°C (p < 0.05).
4 Discussion
L. monocytogenes is a food-borne pathogen which is very difficult to control due to its ability to persist and grow in a wide range of environmental conditions [19]. Some authors have suggested the use of bacteriocins produced by LAB to control the growth of the foodborne pathogen due to them being regarded as natural and safe food preservatives [20]. However, current bacteriocins such as nisin and pediocin still have some weaknesses [10,12,21]; therefore, the search for new types of bacteriocin is still needed. The present study screened 52 LAB isolated from the intestinal tracts of gilt-head seabream for anti-listerial activity. The results show that 3 out of the 52 LAB displayed anti-listerial activity. One of the three isolates, LAB strain MA115, was selected for further studies as it exhibited the strongest anti-listerial activity. Based on phenotypic assays and its 16S rDNA sequence, LAB strain MA115 was identified as Enterococcus faecium.
E. faecium is a member of LAB commonly found as part of the intestinal microbiota of animals including aquatic species [14,22,23]. In addition, many strains of E. faecium have been reported to have antagonistic activity against several bacterial pathogens: V. parahaemolyticus, V. vulnificus, Escherichia coli, and also L. monocytogenes [24,25]. Other studies have reported that antimicrobial activity can be in several mechanisms including competition for nutrients or the production of metabolites such as ethanol or carbon dioxide organic acids [26], hydrogen peroxide, and bacteriocins [24,27]. Among these antimicrobial substances, bacteriocins gain the most interest due to several factors including having a narrower spectrum of killing activity [28], GRAS substances, inactive and nontoxic to eukaryote cells, and being pH and heat tolerance [6]. The antagonistic activity of lactic and acetic acids has been ruled out because the CFSn was neutralized for the antimicrobial assay in our study. And the antagonistic activity of hydrogen peroxide (H2O2) is excluded because the CFSn was collected from anaerobic culture conditions. The remaining possibility is the presence of BLIS, indicated by sensitivity to proteinase K.
Numerous studies have reported the capacity of E. faecium to produce diverse bacteriocins such as Enterocin A and B [24], Enterocin 3Da and 3Dab, Enterocin P [25], and Bacteriocin T8 [29]. Bacteriocins produced by E. faecium are generally grouped into class II bacteriocins, non-lantibiotics due to these common characteristics: having a small molecular size (3–8 kDa), being heat stable, and having a narrow range of the inhibitory spectrum [29]. This study shows that the BLIS produced by this new strain of E. faecium appeared to be very stable under a wide range of environmental conditions including, a wide range of pH (2–10), heat (30–121°C), and storage condition (4°C). The stability of this BLIS in high pH appeared to be better than one of current commercial bacteriocin (nisin), which has been reported to have low solubility and less stability in pH value of above 5 and high temperatures [8,9]. Based on these parameters especially heat stability, the BLIS could be a class II bacteriocin. However, further studies to characterize the BLIS are still required.
According to Valenzuela et al. [30], the use of bacteriocins produced by enterococci for biocontrol of L. monocytogenes in seafood and processed products has become increasingly popular in the last few decades. A study by de Carvalho et al. [20] confirmed that a class II bacteriocin synthesized by E. faecium was more effective in killing L. monocytogenes than other control agents including NaNO3. In addition, BLIS produced by Lactobacillus pentosus 39 was reported to reduce the viable count of L. monocytogenes on fresh salmon fillets after being incubated at 4°C for 6 days, from ∼5.8 log units to ∼4.8 log units [31]. The anti-listerial activity of BLIS in our study appeared to be much stronger than in these previous studies. As the consumer demands for more “natural” preservatives of foods are increasing, this BLIS is worth further study as an alternative for food biopreservatives, especially in controlling L. monocytogenes in salmon fillets.
Other studies have described bacteriocins as being limited by short activity in foods [31]. The reason for inactivation of bacteriocin activity was explained as due to: (1) the degradation of bacteriocin by endogenous proteases in the food matrix [32], or (2) being absorbed by the meat and fat particles which lead to the inactivation of bacteriocin [33,34]. The application of pentocin 31-1 produced by L. pentosus 31-1 had decreased viable counts of L. monocytogenes to only ∼1 log unit for 6 days, and the viable counts of the pathogens increased afterward. In addition, Katla et al. [35] reported low activity of sakacin P when applied to control L. monocytogenes in chicken cold cuts. In contradiction, the present study demonstrated the bactericidal activity of BLIS synthesized by E. faecium MA115 remains stable over the 6-day incubation period, which was a promising result for food biopreservative candidates.
Acknowledging various foodborne pathogens such as L. monocytogenes will be still considered a major problem in many food products including seafood or dairy products in the future, natural techniques such as the use of bacteriocins will still gain considerable interest in food preservatives. Many researchers nowadays keep searching for specific bacteriocins from LAB groups from diverse sources including animals [24,36], vegetables [37], and dairy products [38]. Various types of bacteriocins have also been found in a wide range of LAB members. Some of those bacteriocins are very stable at a wide range of environmental conditions including temperature, pH, and enzymes. Future studies would be to find the best method on how these bacteriocins could keep their bactericidal activity stable in food products. A recent paper by Malheiros et al. [39], for instance, encapsulated bacteriocin-A produced by E. faecium using liposomal nanovesicles to suppress the growth of L. monocytogenes in goat milk. These types of studies are performed due to many bacteriocins showing antagonistic activities by in vitro assays show no activity when applied in food due to being degraded by endogenous proteases in the food matrix [32], or being absorbed by food particles [33,34].
The BLIS of E. faecium MA115 showed bactericidal activity on L. monocytogenes as indicated by a significant reduction in the pathogen’s viable cells on salmon fillets after treatment (Figure 3). In the absence of the BLIS, L. monocytogenes was able to survive on the salmon fillets stored at 0°C for 6 days, indicated by no significant difference in the number of viable cells (4.27 log units at day 0 and 4.35 log units at day 6 (t = 0.42, df = 3, p = 0.71)). However, viable cells of L. monocytogenes in the salmon fillets after 5 s immersion in the neutralized BLIS of E. faecium MA115 significantly decreased after 6 days (t = 9.95, df = 3, p = 0.002). The viable cells of L. monocytogenes were recorded at 4.22 log units at day 0 to only 0.46 log units on day 6 (0°C). This shows that the BLIS produced by E. faecium was bactericidal to the foodborne pathogen.
5 Conclusion
Members of LAB are potential sources to find natural biopreservatives. The BLIS produced by E. faecium strain MA115 obtained in the present study could be a good candidate for seafood biopreservatives, especially to control L. monocytogenes in seafood due to exhibiting a strong antimicrobial activity against the foodborne pathogen by in vitro test. However, more studies are still required to characterize this BLIS and to develop it as a food biopreservative as the consumer demands for more “natural” preservatives of foods are increasing.
Acknowledgments
The authors thank all members of the Aquatic Microbiology Laboratory, Institute for Marine and Antarctic Studies (IMAS) Launceston, and Laboratory of Microbiology, Human Life Science Faculty, the University of Tasmania for all technical advice during the experiment.
-
Funding information: This research was funded by the Institute for Marine and Antarctic Studies (IMAS), University of Tasmania, Australia, and supported by Universitas Airlangga, Indonesia.
-
Author contributions: M.A – Conceptualization, methodology, data collection, bacterial sequencing, data analysis, writing original draft, funding acquisition, and submission; O.A.O – Conceptualization, methodology, data collection, data analysis, editing; validation, and approved the final manuscript; F.R.D – Conceptualization, methodology, funding acquisition, data collection, review, editing, and approved the final manuscript; C.M.B. – Conceptualization, methodology, supervision, data analysis, review, validation, and approved the final manuscript.
-
Conflict of interest: The authors state no conflict of interest.
-
Data availability statement: The datasets generated during and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
[1] Basha KA, Kumar NR, Das V, Reshmi K, Rao BM, Lalitha K, et al. Prevalence, molecular characterization, genetic heterogeneity and antimicrobial resistance of Listeria monocytogenes associated with fish and fishery environment in Kerala, India. Lett Appl Microbiol. 2019;69(4):286–93.10.1111/lam.13205Search in Google Scholar PubMed
[2] Mohan V, Cruz CD, van Vliet AH, Pitman AR, Visnovsky SB, Rivas L, et al. Genomic diversity of Listeria monocytogenes isolates from seafood, horticulture and factory environments in New Zealand. Int J Food Microbiol. 2021;347:109166.10.1016/j.ijfoodmicro.2021.109166Search in Google Scholar PubMed
[3] Commission CA. Joint FAO/WHO Food Standards Programme CODEX Committee on Food Hygiene. 40th Session ftp://ftp fao org/codex/ccfh40/fh40_03e pdf. 2008.Search in Google Scholar
[4] Cheng C, Yang Y, Dong Z, Wang X, Fang C, Yang M, et al. Listeria monocytogenes varies among strains to maintain intracellular pH homeostasis under stresses by different acids as analyzed by a high-throughput microplate-based fluorometry. Front Microbiol. 2015;6:15.10.3389/fmicb.2015.00015Search in Google Scholar PubMed PubMed Central
[5] Pingitore EV, Todorov SD, Sesma F, Franco BDGD. Application of bacteriocinogenic Enterococcus mundtii CRL35 and Enterococcus faecium ST88Ch in the control of Listeria monocytogenes in fresh Minas cheese. Food Microbiol. 2012;32(1):38–47.10.1016/j.fm.2012.04.005Search in Google Scholar PubMed
[6] Hwanhlem N, Chobert JM, H-Kittikun A. Bacteriocin-producing lactic acid bacteria isolated from mangrove forests in southern Thailand as potential bio-control agents in food: Isolation, screening and optimization. Food Control. 2014;41:202–11.10.1016/j.foodcont.2014.01.021Search in Google Scholar
[7] Zhao X, Chen L, Wu JE, He Y, Yang H. Elucidating antimicrobial mechanism of nisin and grape seed extract against Listeria monocytogenes in broth and on shrimp through NMR-based metabolomics approach. Int J Food Microbiol. 2020;319:108494.10.1016/j.ijfoodmicro.2019.108494Search in Google Scholar PubMed
[8] Delves-Broughton J, Blackburn P, Evans RJ, Hugenholtz J. Applications of the bacteriocin, nisin. Antonie van Leeuwenhoek. 1996;69(2):193–202.10.1007/BF00399424Search in Google Scholar PubMed
[9] Rollema HS, Kuipers OP, Both O, Devos WM, Siezen RJ. Improvement of solubility and stability of the antimicrobial peptide nisin by protein engineering. Appl Env Microbiol. 1995;61(8):2873–8.10.1128/aem.61.8.2873-2878.1995Search in Google Scholar PubMed PubMed Central
[10] Yamaki S, Shirahama S, Kobayashi T, Kawai Y, Yamazaki K. Combined effect of nisin and commercial pectin-hydrolysate treatment on survival and growth of Listeria monocytogenes in soy-seasoned salmon roe products. Food Sci Technol Res. 2015;21(5):751–5.10.3136/fstr.21.751Search in Google Scholar
[11] Chung K-T, Dickson JS, Crouse JD. Effects of nisin on growth of bacteria attached to meat. Appl Env Microbiol. 1989;55(6):1329–33.10.1128/aem.55.6.1329-1333.1989Search in Google Scholar PubMed PubMed Central
[12] Naghmouchi K, Kheadr E, Lacroix C, Fliss I. Class I/Class IIa bacteriocin cross-resistance phenomenon in Listeria monocytogenes. Food Microbiol. 2007;24(7):718–27.10.1016/j.fm.2007.03.012Search in Google Scholar PubMed
[13] Amin M. Isolation and selection of probiotic candidates from gastrointestinal tracts of teleosts and molluscs. PhD dissertation, University of Tasmania, Australia; 2018.Search in Google Scholar
[14] Amin M, Liliyanti MA, Nufus NH, Ali M, editors. Screening of antivibrio-producing lactic acid bacteria originated from aquatic animals as probiotic candidates. IOP Conf Ser: Earth Environ Sci. Vol. 441. IOP Publishing; 2020. p. 012092.10.1088/1755-1315/441/1/012092Search in Google Scholar
[15] Saeed S, Ahmad S, Rasool SA. Antimicrobial spectrum, production and mode of action of staphylococcin 188 produced by Staphylococcus aureus 188. Pak J Pharm Sci. 2004;17:1–8.Search in Google Scholar
[16] Abrams D, Barbosa J, Albano H, Silva J, Gibbs PA, Teixeira P. Characterization of bacPPK34 a bacteriocin produced by Pediococcus pentosaceus strain K34 isolated from “Alheira.” Food Control. 2011;22(6):940–6.10.1016/j.foodcont.2010.11.029Search in Google Scholar
[17] Vidhyasagar V, Jeevaratnam K. Bacteriocin activity against various pathogens produced by Pediococcus pentosaceus VJ13 isolated from Idly batter. Biomed Chromatogr. 2013;27(11):1497–502.10.1002/bmc.2948Search in Google Scholar PubMed
[18] Lane D. 16S/23S rRNA sequencing. In: Stackebrandt E, Goodfellow M, editors. Nucleic acid techniques in bacterial systematics. New York: John Wiley & Sons; 1991. p. 115–75.Search in Google Scholar
[19] Cheng C, Arritt F, Stevenson C. Controlling Listeria monocytogenes in cold smoked salmon with the antimicrobial peptide salmine. J Food Sci. 2015;80(6):M1314–M8.10.1111/1750-3841.12886Search in Google Scholar PubMed
[20] de Carvalho AAT, de Paula RA, Mantovani HC, de Moraes CA. Inhibition of Listeria monocytogenes by a lactic acid bacterium isolated from Italian salami. Food Microbiol. 2006;23(3):213–9.10.1016/j.fm.2005.05.009Search in Google Scholar PubMed
[21] Huang Y, Luo YB, Zhai ZY, Zhang HX, Yang CX, Tian HT, et al. Characterization and application of an anti-Listeria bacteriocin produced by Pediococcus pentosaceus 05-10 isolated from Sichuan Pickle, a traditionally fermented vegetable product from China. Food Control. 2009;20(11):1030–5.10.1016/j.foodcont.2008.12.008Search in Google Scholar
[22] Barros J, Andrade M, Radhouani H, Lopez M, Igrejas G, Poeta P, et al. Detection of vanA-containing Enterococcus species in faecal microbiota of gilthead seabream (Sparus aurata). Microbes Env. 2012;27(4):509–11.10.1264/jsme2.ME11346Search in Google Scholar PubMed PubMed Central
[23] Amin M, Adams MB, Burke CM, Bolch CJ. Isolation and screening of lactic acid bacteria associated with the gastrointestinal tracts of abalone at various life stages for probiotic candidates. Aquacult Rep. 2020;17:100378.10.1016/j.aqrep.2020.100378Search in Google Scholar
[24] Aspri M, O’Connor PM, Field D, Cotter PD, Ross P, Hill C, et al. Application of bacteriocin-producing Enterococcus faecium isolated from donkey milk, in the bio-control of Listeria monocytogenes in fresh whey cheese. Int Dairy J. 2017;73:1–9.10.1016/j.idairyj.2017.04.008Search in Google Scholar
[25] Bayoub K, Mardad I, Ammar E, Serrano A, Soukri A. Isolation and purification of two bacteriocins 3D produced by Enterococcus faecium with inhibitory activity against Listeria monocytogenes. Curr Microbiol. 2011;62(2):479–85.10.1007/s00284-010-9732-0Search in Google Scholar PubMed
[26] Vazquez JA, Gonzalez MP, Murado MA. Effects of lactic acid bacteria cultures on pathogenic microbiota from fish. Aquaculture. 2005;245(1–4):149–61.10.1016/j.aquaculture.2004.12.008Search in Google Scholar
[27] Lin YH, Chen YS, Wu HC, Pan SF, Yu B, Chiang CM, et al. Screening and characterization of LAB-produced bacteriocin-like substances from the intestine of grey mullet (Mugil cephalus L.) as potential biocontrol agents in aquaculture. J Appl Microbiol. 2013;114(2):299–307.10.1111/jam.12041Search in Google Scholar PubMed
[28] Riley MA, Wertz JE. Bacteriocins: evolution, ecology, and application. Annu Rev Microbiol. 2002;56(1):117–37.10.1146/annurev.micro.56.012302.161024Search in Google Scholar PubMed
[29] De Kwaadsteniet M, Fraser T, Van Reenen CA, Dicks LM. Bacteriocin T8, a novel class IIa sec-dependent bacteriocin produced by Enterococcus faecium T8, isolated from vaginal secretions of children infected with human immunodeficiency virus. Appl Env Microbiol. 2006;72(7):4761–6.10.1128/AEM.00436-06Search in Google Scholar PubMed PubMed Central
[30] Valenzuela AS, Benomar N, Abriouel H, Cañamero MM, Gálvez A. Isolation and identification of Enterococcus faecium from seafoods: antimicrobial resistance and production of bacteriocin-like substances. Food Microbiol. 2010;27(7):955–61.10.1016/j.fm.2010.05.033Search in Google Scholar PubMed
[31] Anacarso I, Messi P, Condò C, Iseppi R, Bondi M, Sabia C, et al. A bacteriocin-like substance produced from Lactobacillus pentosus 39 is a natural antagonist for the control of Aeromonas hydrophila and Listeria monocytogenes in fresh salmon fillets. LWT - Food Sci Technol. 2014;55(2):604–11.10.1016/j.lwt.2013.10.012Search in Google Scholar
[32] Zhang J, Liu G, Li P, Qu Y. Pentocin 31-1, a novel meat-borne bacteriocin and its application as biopreservative in chill-stored tray-packaged pork meat. Food Control. 2010;21(2):198–202.10.1016/j.foodcont.2009.05.010Search in Google Scholar
[33] Katla T, Møretrø T, Aasen I, Holck A, Axelsson L, Naterstad K. Inhibition of Listeria monocytogenes in cold smoked salmon by addition of sakacin P and/or live Lactobacillus sakei cultures. Food Microbiol. 2001;18(4):431–9.10.1006/fmic.2001.0420Search in Google Scholar
[34] Aasen IM, Markussen S, Møretrø T, Katla T, Axelsson L, Naterstad K. Interactions of the bacteriocins sakacin P and nisin with food constituents. Int J Food Microbiol. 2003;87(1–2):35–43.10.1016/S0168-1605(03)00047-3Search in Google Scholar PubMed
[35] Katla T, Møretrø T, Sveen I, Aasen I, Axelsson L, Rørvik L, et al. Inhibition of Listeria monocytogenes in chicken cold cuts by addition of sakacin P and sakacin P‐producing Lactobacillus sakei. J Appl Microbiol. 2002;93(2):191–6.10.1046/j.1365-2672.2002.01675.xSearch in Google Scholar PubMed
[36] Kubašová I, Diep DB, Ovchinnikov KV, Lauková A, Strompfová V. Bacteriocin production and distribution of bacteriocin-encoding genes in enterococci from dogs. Int J Antimicrob Agents. 2020;55(2):105859.10.1016/j.ijantimicag.2019.11.016Search in Google Scholar PubMed
[37] Qiao X, Du R, Wang Y, Han Y, Zhou Z. Purification, characterization and mode of action of enterocin, a novel bacteriocin produced by Enterococcus faecium TJUQ1. Int J Biol Macromol. 2020;144:151–9.10.1016/j.ijbiomac.2019.12.090Search in Google Scholar PubMed
[38] Lianou A, Kakouri A, Pappa EC, Samelis J. Growth interactions and antilisterial effects of the bacteriocinogenic Lactococcus lactis subsp. cremoris M104 and Enterococcus faecium KE82 strains in thermized milk in the presence or absence of a commercial starter culture. Food Microbiol. 2017;64:145–54.10.1016/j.fm.2016.12.019Search in Google Scholar PubMed
[39] Malheiros PS, Cuccovia IM, Franco BD. Inhibition of Listeria monocytogenes in vitro and in goat milk by liposomal nanovesicles containing bacteriocins produced by Lactobacillus sakei subsp. sakei 2a. Food Control. 2016;63:158–64.10.1016/j.foodcont.2015.11.037Search in Google Scholar
© 2023 the author(s), published by De Gruyter
This work is licensed under the Creative Commons Attribution 4.0 International License.
Articles in the same Issue
- Regular Articles
- The impact of COVID-19 pandemic on business risks and potato commercial model
- Effects of potato (Solanum tuberosum L.)–Mucuna pruriens intercropping pattern on the agronomic performances of potato and the soil physicochemical properties of the western highlands of Cameroon
- Machine learning-based prediction of total phenolic and flavonoid in horticultural products
- Revamping agricultural sector and its implications on output and employment generation: Evidence from Nigeria
- Does product certification matter? A review of mechanism to influence customer loyalty in the poultry feed industry
- Farmer regeneration and knowledge co-creation in the sustainability of coconut agribusiness in Gorontalo, Indonesia
- Lablab purpureus: Analysis of landraces cultivation and distribution, farming systems, and some climatic trends in production areas in Tanzania
- The effects of carrot (Daucus carota L.) waste juice on the performances of native chicken in North Sulawesi, Indonesia
- Properties of potassium dihydrogen phosphate and its effects on plants and soil
- Factors influencing the role and performance of independent agricultural extension workers in supporting agricultural extension
- The fate of probiotic species applied in intensive grow-out ponds in rearing water and intestinal tracts of white shrimp, Litopenaeus vannamei
- Yield stability and agronomic performances of provitamin A maize (Zea mays L.) genotypes in South-East of DR Congo
- Diallel analysis of length and shape of rice using Hayman and Griffing method
- Physicochemical and microbiological characteristics of various stem bark extracts of Hopea beccariana Burck potential as natural preservatives of coconut sap
- Correlation between descriptive and group type traits in the system of cow’s linear classification of Ukrainian Brown dairy breed
- Meta-analysis of the influence of the substitution of maize with cassava on performance indices of broiler chickens
- Bacteriocin-like inhibitory substance (BLIS) produced by Enterococcus faecium MA115 and its potential use as a seafood biopreservative
- Meta-analysis of the benefits of dietary Saccharomyces cerevisiae intervention on milk yield and component characteristics in lactating small ruminants
- Growth promotion potential of Bacillus spp. isolates on two tomato (Solanum lycopersicum L.) varieties in the West region of Cameroon
- Prioritizing IoT adoption strategies in millennial farming: An analytical network process approach
- Soil fertility and pomelo yield influenced by soil conservation practices
- Soil macrofauna under laying hens’ grazed fields in two different agroecosystems in Portugal
- Factors affecting household carbohydrate food consumption in Central Java: Before and during the COVID-19 pandemic
- Properties of paper coated with Prunus serotina (Ehrh.) extract formulation
- Fertiliser cost prediction in European Union farms: Machine-learning approaches through artificial neural networks
- Molecular and phenotypic markers for pyramiding multiple traits in rice
- Natural product nanofibers derived from Trichoderma hamatum K01 to control citrus anthracnose caused by Colletotrichum gloeosporioides
- Role of actors in promoting sustainable peatland management in Kubu Raya Regency, West Kalimantan, Indonesia
- Small-scale coffee farmers’ perception of climate-adapted attributes in participatory coffee breeding: A case study of Gayo Highland, Aceh, Indonesia
- Optimization of extraction using surface response methodology and quantification of cannabinoids in female inflorescences of marijuana (Cannabis sativa L.) at three altitudinal floors of Peru
- Production factors, technical, and economic efficiency of soybean (Glycine max L. Merr.) farming in Indonesia
- Economic performance of smallholder soya bean production in Kwara State, Nigeria
- Indonesian rice farmers’ perceptions of different sources of information and their effect on farmer capability
- Feed preference, body condition scoring, and growth performance of Dohne Merino ram fed varying levels of fossil shell flour
- Assessing the determinant factors of risk strategy adoption to mitigate various risks: An experience from smallholder rubber farmers in West Kalimantan Province, Indonesia
- Analysis of trade potential and factors influencing chili export in Indonesia
- Grade-C kenaf fiber (poor quality) as an alternative material for textile crafts
- Technical efficiency changes of rice farming in the favorable irrigated areas of Indonesia
- Palm oil cluster resilience to enhance indigenous welfare by innovative ability to address land conflicts: Evidence of disaster hierarchy
- Factors determining cassava farmers’ accessibility to loan sources: Evidence from Lampung, Indonesia
- Tailoring business models for small-medium food enterprises in Eastern Africa can drive the commercialization and utilization of vitamin A rich orange-fleshed sweet potato puree
- Revitalizing sub-optimal drylands: Exploring the role of biofertilizers
- Effects of salt stress on growth of Quercus ilex L. seedlings
- Design and fabrication of a fish feed mixing cum pelleting machine for small-medium scale aquaculture industry
- Indicators of swamp buffalo business sustainability using partial least squares structural equation modelling
- Effect of arbuscular mycorrhizal fungi on early growth, root colonization, and chlorophyll content of North Maluku nutmeg cultivars
- How intergenerational farmers negotiate their identity in the era of Agriculture 4.0: A multiple-case study in Indonesia
- Responses of broiler chickens to incremental levels of water deprivation: Growth performance, carcass characteristics, and relative organ weights
- The improvement of horticultural villages sustainability in Central Java Province, Indonesia
- Effect of short-term grazing exclusion on herbage species composition, dry matter productivity, and chemical composition of subtropical grasslands
- Analysis of beef market integration between consumer and producer regions in Indonesia
- Analysing the sustainability of swamp buffalo (Bubalus bubalis carabauesis) farming as a protein source and germplasm
- Toxicity of Calophyllum soulattri, Piper aduncum, Sesamum indicum and their potential mixture for control Spodoptera frugiperda
- Consumption profile of organic fruits and vegetables by a Portuguese consumer’s sample
- Phenotypic characterisation of indigenous chicken in the central zone of Tanzania
- Diversity and structure of bacterial communities in saline and non-saline rice fields in Cilacap Regency, Indonesia
- Isolation and screening of lactic acid bacteria producing anti-Edwardsiella from the gastrointestinal tract of wild catfish (Clarias gariepinus) for probiotic candidates
- Effects of land use and slope position on selected soil physicochemical properties in Tekorsh Sub-Watershed, East Gojjam Zone, Ethiopia
- Design of smart farming communication and web interface using MQTT and Node.js
- Assessment of bread wheat (Triticum aestivum L.) seed quality accessed through different seed sources in northwest Ethiopia
- Estimation of water consumption and productivity for wheat using remote sensing and SEBAL model: A case study from central clay plain Ecosystem in Sudan
- Agronomic performance, seed chemical composition, and bioactive components of selected Indonesian soybean genotypes (Glycine max [L.] Merr.)
- The role of halal requirements, health-environmental factors, and domestic interest in food miles of apple fruit
- Subsidized fertilizer management in the rice production centers of South Sulawesi, Indonesia: Bridging the gap between policy and practice
- Factors affecting consumers’ loyalty and purchase decisions on honey products: An emerging market perspective
- Inclusive rice seed business: Performance and sustainability
- Design guidelines for sustainable utilization of agricultural appropriate technology: Enhancing human factors and user experience
- Effect of integrate water shortage and soil conditioners on water productivity, growth, and yield of Red Globe grapevines grown in sandy soil
- Synergic effect of Arbuscular mycorrhizal fungi and potassium fertilizer improves biomass-related characteristics of cocoa seedlings to enhance their drought resilience and field survival
- Control measure of sweet potato weevil (Cylas formicarius Fab.) (Coleoptera: Curculionidae) in endemic land of entisol type using mulch and entomopathogenic fungus Beauveria bassiana
- In vitro and in silico study for plant growth promotion potential of indigenous Ochrobactrum ciceri and Bacillus australimaris
- Effects of repeated replanting on yield, dry matter, starch, and protein content in different potato (Solanum tuberosum L.) genotypes
- Review Articles
- Nutritional and chemical composition of black velvet tamarind (Dialium guineense Willd) and its influence on animal production: A review
- Black pepper (Piper nigrum Lam) as a natural feed additive and source of beneficial nutrients and phytochemicals in chicken nutrition
- The long-crowing chickens in Indonesia: A review
- A transformative poultry feed system: The impact of insects as an alternative and transformative poultry-based diet in sub-Saharan Africa
- Short Communication
- Profiling of carbonyl compounds in fresh cabbage with chemometric analysis for the development of freshness assessment method
- Special Issue of The 4th International Conference on Food Science and Engineering (ICFSE) 2022 - Part I
- Non-destructive evaluation of soluble solid content in fruits with various skin thicknesses using visible–shortwave near-infrared spectroscopy
- Special Issue on FCEM - International Web Conference on Food Choice & Eating Motivation - Part I
- Traditional agri-food products and sustainability – A fruitful relationship for the development of rural areas in Portugal
- Consumers’ attitudes toward refrigerated ready-to-eat meat and dairy foods
- Breakfast habits and knowledge: Study involving participants from Brazil and Portugal
- Food determinants and motivation factors impact on consumer behavior in Lebanon
- Comparison of three wine routes’ realities in Central Portugal
- Special Issue on Agriculture, Climate Change, Information Technology, Food and Animal (ACIFAS 2020)
- Environmentally friendly bioameliorant to increase soil fertility and rice (Oryza sativa) production
- Enhancing the ability of rice to adapt and grow under saline stress using selected halotolerant rhizobacterial nitrogen fixer
Articles in the same Issue
- Regular Articles
- The impact of COVID-19 pandemic on business risks and potato commercial model
- Effects of potato (Solanum tuberosum L.)–Mucuna pruriens intercropping pattern on the agronomic performances of potato and the soil physicochemical properties of the western highlands of Cameroon
- Machine learning-based prediction of total phenolic and flavonoid in horticultural products
- Revamping agricultural sector and its implications on output and employment generation: Evidence from Nigeria
- Does product certification matter? A review of mechanism to influence customer loyalty in the poultry feed industry
- Farmer regeneration and knowledge co-creation in the sustainability of coconut agribusiness in Gorontalo, Indonesia
- Lablab purpureus: Analysis of landraces cultivation and distribution, farming systems, and some climatic trends in production areas in Tanzania
- The effects of carrot (Daucus carota L.) waste juice on the performances of native chicken in North Sulawesi, Indonesia
- Properties of potassium dihydrogen phosphate and its effects on plants and soil
- Factors influencing the role and performance of independent agricultural extension workers in supporting agricultural extension
- The fate of probiotic species applied in intensive grow-out ponds in rearing water and intestinal tracts of white shrimp, Litopenaeus vannamei
- Yield stability and agronomic performances of provitamin A maize (Zea mays L.) genotypes in South-East of DR Congo
- Diallel analysis of length and shape of rice using Hayman and Griffing method
- Physicochemical and microbiological characteristics of various stem bark extracts of Hopea beccariana Burck potential as natural preservatives of coconut sap
- Correlation between descriptive and group type traits in the system of cow’s linear classification of Ukrainian Brown dairy breed
- Meta-analysis of the influence of the substitution of maize with cassava on performance indices of broiler chickens
- Bacteriocin-like inhibitory substance (BLIS) produced by Enterococcus faecium MA115 and its potential use as a seafood biopreservative
- Meta-analysis of the benefits of dietary Saccharomyces cerevisiae intervention on milk yield and component characteristics in lactating small ruminants
- Growth promotion potential of Bacillus spp. isolates on two tomato (Solanum lycopersicum L.) varieties in the West region of Cameroon
- Prioritizing IoT adoption strategies in millennial farming: An analytical network process approach
- Soil fertility and pomelo yield influenced by soil conservation practices
- Soil macrofauna under laying hens’ grazed fields in two different agroecosystems in Portugal
- Factors affecting household carbohydrate food consumption in Central Java: Before and during the COVID-19 pandemic
- Properties of paper coated with Prunus serotina (Ehrh.) extract formulation
- Fertiliser cost prediction in European Union farms: Machine-learning approaches through artificial neural networks
- Molecular and phenotypic markers for pyramiding multiple traits in rice
- Natural product nanofibers derived from Trichoderma hamatum K01 to control citrus anthracnose caused by Colletotrichum gloeosporioides
- Role of actors in promoting sustainable peatland management in Kubu Raya Regency, West Kalimantan, Indonesia
- Small-scale coffee farmers’ perception of climate-adapted attributes in participatory coffee breeding: A case study of Gayo Highland, Aceh, Indonesia
- Optimization of extraction using surface response methodology and quantification of cannabinoids in female inflorescences of marijuana (Cannabis sativa L.) at three altitudinal floors of Peru
- Production factors, technical, and economic efficiency of soybean (Glycine max L. Merr.) farming in Indonesia
- Economic performance of smallholder soya bean production in Kwara State, Nigeria
- Indonesian rice farmers’ perceptions of different sources of information and their effect on farmer capability
- Feed preference, body condition scoring, and growth performance of Dohne Merino ram fed varying levels of fossil shell flour
- Assessing the determinant factors of risk strategy adoption to mitigate various risks: An experience from smallholder rubber farmers in West Kalimantan Province, Indonesia
- Analysis of trade potential and factors influencing chili export in Indonesia
- Grade-C kenaf fiber (poor quality) as an alternative material for textile crafts
- Technical efficiency changes of rice farming in the favorable irrigated areas of Indonesia
- Palm oil cluster resilience to enhance indigenous welfare by innovative ability to address land conflicts: Evidence of disaster hierarchy
- Factors determining cassava farmers’ accessibility to loan sources: Evidence from Lampung, Indonesia
- Tailoring business models for small-medium food enterprises in Eastern Africa can drive the commercialization and utilization of vitamin A rich orange-fleshed sweet potato puree
- Revitalizing sub-optimal drylands: Exploring the role of biofertilizers
- Effects of salt stress on growth of Quercus ilex L. seedlings
- Design and fabrication of a fish feed mixing cum pelleting machine for small-medium scale aquaculture industry
- Indicators of swamp buffalo business sustainability using partial least squares structural equation modelling
- Effect of arbuscular mycorrhizal fungi on early growth, root colonization, and chlorophyll content of North Maluku nutmeg cultivars
- How intergenerational farmers negotiate their identity in the era of Agriculture 4.0: A multiple-case study in Indonesia
- Responses of broiler chickens to incremental levels of water deprivation: Growth performance, carcass characteristics, and relative organ weights
- The improvement of horticultural villages sustainability in Central Java Province, Indonesia
- Effect of short-term grazing exclusion on herbage species composition, dry matter productivity, and chemical composition of subtropical grasslands
- Analysis of beef market integration between consumer and producer regions in Indonesia
- Analysing the sustainability of swamp buffalo (Bubalus bubalis carabauesis) farming as a protein source and germplasm
- Toxicity of Calophyllum soulattri, Piper aduncum, Sesamum indicum and their potential mixture for control Spodoptera frugiperda
- Consumption profile of organic fruits and vegetables by a Portuguese consumer’s sample
- Phenotypic characterisation of indigenous chicken in the central zone of Tanzania
- Diversity and structure of bacterial communities in saline and non-saline rice fields in Cilacap Regency, Indonesia
- Isolation and screening of lactic acid bacteria producing anti-Edwardsiella from the gastrointestinal tract of wild catfish (Clarias gariepinus) for probiotic candidates
- Effects of land use and slope position on selected soil physicochemical properties in Tekorsh Sub-Watershed, East Gojjam Zone, Ethiopia
- Design of smart farming communication and web interface using MQTT and Node.js
- Assessment of bread wheat (Triticum aestivum L.) seed quality accessed through different seed sources in northwest Ethiopia
- Estimation of water consumption and productivity for wheat using remote sensing and SEBAL model: A case study from central clay plain Ecosystem in Sudan
- Agronomic performance, seed chemical composition, and bioactive components of selected Indonesian soybean genotypes (Glycine max [L.] Merr.)
- The role of halal requirements, health-environmental factors, and domestic interest in food miles of apple fruit
- Subsidized fertilizer management in the rice production centers of South Sulawesi, Indonesia: Bridging the gap between policy and practice
- Factors affecting consumers’ loyalty and purchase decisions on honey products: An emerging market perspective
- Inclusive rice seed business: Performance and sustainability
- Design guidelines for sustainable utilization of agricultural appropriate technology: Enhancing human factors and user experience
- Effect of integrate water shortage and soil conditioners on water productivity, growth, and yield of Red Globe grapevines grown in sandy soil
- Synergic effect of Arbuscular mycorrhizal fungi and potassium fertilizer improves biomass-related characteristics of cocoa seedlings to enhance their drought resilience and field survival
- Control measure of sweet potato weevil (Cylas formicarius Fab.) (Coleoptera: Curculionidae) in endemic land of entisol type using mulch and entomopathogenic fungus Beauveria bassiana
- In vitro and in silico study for plant growth promotion potential of indigenous Ochrobactrum ciceri and Bacillus australimaris
- Effects of repeated replanting on yield, dry matter, starch, and protein content in different potato (Solanum tuberosum L.) genotypes
- Review Articles
- Nutritional and chemical composition of black velvet tamarind (Dialium guineense Willd) and its influence on animal production: A review
- Black pepper (Piper nigrum Lam) as a natural feed additive and source of beneficial nutrients and phytochemicals in chicken nutrition
- The long-crowing chickens in Indonesia: A review
- A transformative poultry feed system: The impact of insects as an alternative and transformative poultry-based diet in sub-Saharan Africa
- Short Communication
- Profiling of carbonyl compounds in fresh cabbage with chemometric analysis for the development of freshness assessment method
- Special Issue of The 4th International Conference on Food Science and Engineering (ICFSE) 2022 - Part I
- Non-destructive evaluation of soluble solid content in fruits with various skin thicknesses using visible–shortwave near-infrared spectroscopy
- Special Issue on FCEM - International Web Conference on Food Choice & Eating Motivation - Part I
- Traditional agri-food products and sustainability – A fruitful relationship for the development of rural areas in Portugal
- Consumers’ attitudes toward refrigerated ready-to-eat meat and dairy foods
- Breakfast habits and knowledge: Study involving participants from Brazil and Portugal
- Food determinants and motivation factors impact on consumer behavior in Lebanon
- Comparison of three wine routes’ realities in Central Portugal
- Special Issue on Agriculture, Climate Change, Information Technology, Food and Animal (ACIFAS 2020)
- Environmentally friendly bioameliorant to increase soil fertility and rice (Oryza sativa) production
- Enhancing the ability of rice to adapt and grow under saline stress using selected halotolerant rhizobacterial nitrogen fixer