Mesolithic Freshwater Fishing: A Zooarchaeological Case Study
-
Anne Bridault
 ,
Annelise Binois-Roman
,
Annelise Binois-Roman

Abstract
In this article we investigate the importance of freshwater fishing during the Mesolithic in France, in inland sites where fishing was always associated with big game hunting. We present a review of Mesolithic sites that have yielded freshwater fish remains and focus on a case study from an occupation dated to the eighth millennium BC at Les Cabônes rock shelter, located near the Doubs River, that yielded over 9,300 fish remains. Cyprinid (including bream, roach, and minnow) remains are predominant; other taxa, such as salmonids (probably trout), grayling, burbot, perch, and eel, are represented by only a few remains each. Although small fish (about 15 cm) constitute the majority of the catch, we provide evidence that their accumulation in the shelter is mostly of human origin. These catches constituted a source of food throughout the year, particularly during the lean season. According to the ecological preferences of the fish species identified, it is highly unlikely that they were caught in a single location. The animals were probably caught with passive fishing equipment in shallow banks or side channels where large numbers of specimens could be captured. Fishing appears to have been optimised to maximise yield-to-effort ratios, together with other wetland resources. Finally, we question the presumed causal links between site micro-setting, site occupation, and increased dependence on aquatic resources and reduced group mobility.
1 Introduction
Early on, Binford postulated that throughout the Old World, human societies became increasingly dependent on aquatic resources for food in the Terminal Palaeolithic and Mesolithic (Binford, 1968). He associated this with a higher degree of sedentism, population growth, and food storage. Since then, research has focused on discussing two economic characteristics of “post-Pleistocene” societies: a broadening of the range of exploited species and a markedly increased dependence on small, highly productive species. In France, investigation of prehistoric fishing was initiated by precursors of archaeo-ichthyology through the analysis of fish remains (Desse & Desse, 1976, 1983; Le Gall, 1982, 1984). Work by Le Gall, mainly in continental environments and in the South of France, has shown that systematic fishing for large migratory species (salmon and sea trout) developed during the Late Palaeolithic (Le Gall, 1991, 1992, 1993, 1999). Then, during the Azilian period, freshwater fishing tended to focus more on abundant perennial species, such as cyprinids and pike, though studies were limited to regions south of the Loire and in the Doubs basin (Cravinho & Desse-Berset, 2005; Cravinho, 2011; Frontin, 2017; Le Gall & Pannoux, 1994). The situation is not as clear for the Mesolithic: mixed documentation and a very limited number of archaeo-ichthyological analyses limit our understanding of fishing during the period. A decrease in fishing has even been suggested due to the scarcity of fish bones found in some cultural contexts. Even on the scale of Western Europe, with the exception of southern Scandinavia and northern Germany, there are few studies of large Mesolithic assemblages of fish bone from inland sites (Dauphin, 1989; Niekus, Brinkhuizen, Kerkhoven, Huisman, & Velthuizen, 2012; Wierer et al., 2018; Wijngaarden-Bakker, 1989). Stable isotope analyses of the Mesolithic diet tend to highlight a diversity of situations: diets that are exclusively terrestrial, with a low contribution of freshwater fish, or with mixed terrestrial/aquatic resources (Drucker et al., 2016; Drucker, Bridault, Ducrocq, Baumann, & Valentin, 2020; Schulting, Blockley, Bocherens, Drucker, & Richards, 2008), in both the Early and the Late Mesolithic. The question of freshwater fishing and its contribution to the economy of Mesolithic groups certainly needs to be examined on a case-by-case basis. The aim of this article is to discuss the importance of freshwater fishing in an economy largely based on large game hunting, using unpublished data from the study[1] of an assemblage of more than 9,300 fish remains from an occupation of the Ranchot site (Jura), dated to the eighth millennium BC.
2 Archaeological Context
2.1 Freshwater Fishing in Mesolithic France
A bibliographical search enabled us to identify 21 Mesolithic sites in France in which bones of freshwater ichthyofauna were identified. Six additional sites only mention the presence of unspecified fish remains, that were most probably of freshwater or migratory species given their geographic location. This corpus thus comprises a total of 27 sites, distributed mainly inland over a large part of the country, and 32 assemblages for the entire Mesolithic period (Figure 1). More than half of these (15 out of 27) contain less than 25 remains and only 3 assemblages comprise more than 1,000 remains: the Cabônes shelter at Ranchot (Jura), the open-air sites of Noyen-sur-Seine “Le Haut-des-Nachères” (Seine-et-Marne), and La Chaussée-Tirancourt “le Petit Marais” (Somme), and probably also the Montclus site (Gard) (Table S1). This disparity is striking, especially as fine mesh sieving was carried out on most of these sites, albeit not always exhaustively (Table S1). In the valleys of northern France, where Mesolithic groups regularly settled on dry land near rivers, the presence of fish remains is not systematic even though the mammalian fauna is well preserved and exhaustive sieving was carried out (e.g. Ducrocq et al., 2019). Fishing is only clearly attested at a small number of sites. This is the case at Noyen-sur-Seine where, in addition to eel and pike remains, a dugout log-boat and fish traps have been preserved (Deseine, Guéret, Vigne, Mordant, & Valentin, 2019; Marinval-Vigne et al., 1989; Mordant, Valentin, & Vigne, 2013), and stable isotope analysis demonstrates that freshwater aquatic resources made a significant contribution to the human diet (Drucker et al., 2016).
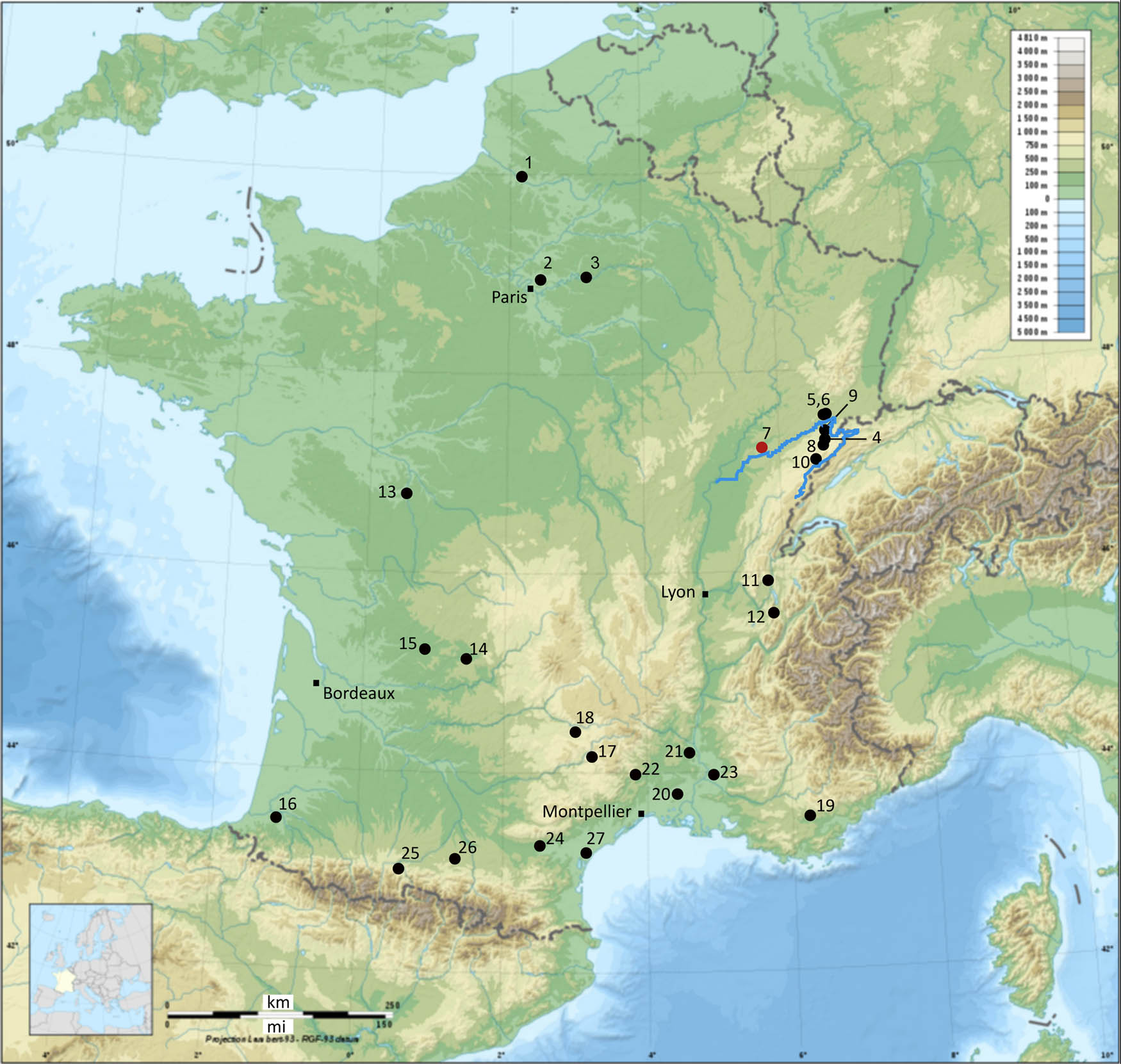
Location of French Mesolithic sites having yielded freshwater fish remains (map of France from cartesFrance.fr). (1) Le Petit Marais, (2) La Haute-Île, (3) Le Haut-des-Nachères, (4) Baume de Montandon, (5) Bavans, (6) Châtaillon, (7) Les Cabônes, (8) Gigot, (9) Rochedane, (10) La Roche-aux-Pêcheurs, (11) Sous Balme, (12) Jean-Pierre 1, (13) Abri des Rocs, (14) La Doue, (15) Le Peyrat, (16) Bourouilla, (17) La Poujade, (18) Roquemissou, (19) La Baume Fontbrégoua, (20) Le Plaisir, (21) La Baume de Montclus, (22) Grotte du Salpêtre, (23) Le Mourre de Sève, (24) Balma de l’Abeurador, (25) Moulin, (26) Buholoup, and (27) La Crouzade. The Doubs River is highlighted in blue. Please refer to Table S1 in the supporting information for the data.
Relative abundance (Numer of Identified Specimens -NISP- values) of fish taxa from layer 3 of Les Cabônes
| Taxa | NISP | % | |
|---|---|---|---|
| Cyprinidae | Cyprinids | 8,843 | 96.56 |
| Abramis brama | Common bream | 18 | |
| Rutilus rutilus | Roach | 2 | |
| Phoxinus phoxinus | Minnow | 1 | |
| Salmo cf. trutta fario | Trout | 243 | 2.65 |
| Thymallus thymallus | Grayling | 22 | 0.24 |
| Lota lota | Burbot | 20 | 0.22 |
| Perca fluviatilis | Perch | 23 | 0.25 |
| Anguilla anguilla | Eel | 7 | 0.08 |
| Total identified | 9,158 | 100 | |
| Unidentified | 170 | 1.82 | |
| Total | 9,328 |
These findings motivated a detailed study centred on the ichthyofaunal assemblage of layer 3 of the Cabônes shelter at Ranchot in the Jura (Frontin, 2017), a region rich in Mesolithic sites with well-preserved faunal remains.
2.2 Regional Context
Settlement intensified during the Early Mesolithic, from 8400 BC onwards. Then, during the Middle Mesolithic, between 7600 and 7000 BC, an increase in the number of sites is recorded at all altitudes, including large open-air sites (Cupillard et al., 2015); occupation levels are denser, richer, and attest to diversified activities. At the beginning of the Late Mesolithic, the number of sites in the region decreases, possibly due to a climatic deterioration (i.e. the 8200 cal. BP event), after which the number of occupations increases again from 6000 BC onwards (Cupillard et al., 2015).
Mesolithic sites with ichthyofaunal remains are distributed along the entire course of the Doubs. This river flows across the Central Jura for 453 km, crossing the Bourgogne Franche-Comté region in France as well as the cantons of Neuchâtel and Jura in Switzerland. It is supplied by some 15 tributaries, and is the main tributary of the Saône, which is itself the main tributary of the Rhône. The flow of the Doubs is irregular, of the pluvial to pluvial-nival type with severe low water in the summer; the flood period extends from September to the end of May due to feeding by the heavy rainfall that affects the Jura massif. While the extent of the Weichselian alluvial deposits is well established (Fleury, Farjanel, & Collin, 1982), our knowledge of the postglacial evolution of the river is incomplete (Rotillon, 2000; Séara, Rotillon, & Cupillard, 2002). It is known that the rapid warming phases of the Lateglacial–Holocene transition impacted hydrosystems all over Europe, increasing river bed incision and changing systems from laterally active braided flows to meandering single channels (Macklin & Lewin, 2003; Magny & Bravard, 2002; Pastre et al., 2000). According to a study conducted at Choisey near Dole (Rotillon, 2000), the Doubs first incised the valley at the beginning of the Lateglacial, then river dynamics slackened in the Preboreal and Boreal periods. The development of riparian vegetation may also have played a role in fixing river courses.
The impact of climate change can also be seen on fish populations. During the Last Glacial, only cryophilic and low-temperature breeding species, such as salmonids, burbot (Lota lota), or pike (Esox lucius) and, to a lesser extent, rheophilic cyprinids such as common dace (Leuciscus leuciscus), were able to survive north of refuge areas (Mediterranean fringe and Black Sea basin) (Le Gall, 2010). Opportunities for recolonisation arose during the deglaciation process, and fish populations attained their maximum expansion during the Lateglacial period, expanding along the natural channels formed by the Danube and the Rhone. Cryophilic species then dispersed rapidly, followed by thermophilic species (Le Gall, 2010; Persat & Keith, 1997, 2002; Persat, 2003).
2.3 The Cabônes Site at Ranchot
Les Cabônes rock shelter (or “abri du Colonel Martin”) is situated in the last ranges of the Jura front. It is located on the right bank of the Doubs River at an altitude of 216 m, near the town of Ranchot (Jura), halfway between Besançon and Dole, in the lower Doubs valley (Figure 2). In this sector, the bed of the Doubs is wide and gently sloped (0.45%). The site is a small cave dug into a Lower Sequanian limestone ledge. It was probably formed by the joint action of the Doubs and a former karstic flow (Campy, David, & Cupillard, 1989; Cupillard & David, 1995). The opening of the cave forms a shelter about ten metres long. Following surveys carried out in the 1950s and 1960s, the site was extensively excavated from 1978 to 1989 under the direction of A. Thévenin, M. Campy, S. David, and C. Cupillard (Campy et al., 1989; Cupillard, 1998b; Cupillard & David, 1995).

General view of Les Cabônes site (Ranchot, Jura, France). The site is on the left, and the Doubs River is on the right. © C. Cupillard.
The stratigraphy is fairly well preserved in the front part of the site, where three complexes can be identified (Campy et al., 1989) (Figure 3):
A lower sterile complex consisting of pebbles lying on the limestone bedrock, corresponding to the Würmian alluvial deposits of the Doubs (layer 7).
An intermediate complex consisting of Doubs overflow silts incorporating angular cryoclastic fragments and blocks from the dismantlement of the porch (layers 4, 5, and 6). The top part of this complex (layer 4a) marks the end of the lateglacial allochtonous contributions of the Doubs. These layers are contemporaneous with Upper-Final Magdalenian occupations (David, 1993). The fauna consists mainly of reindeer, with a date for one bone of 13965 ± 101 BP, while a cave lion remain was dated to 12565 ± 50 BP, i.e. 12620–13201 cal. BC (Cupillard et al., 2015; Stuart & Lister, 2007).
An upper complex composed of clayey matrix with cryoclastic fragments, which is a local expression of the “groizes litées” resulting from the dismantling of the shelter. This complex is subdivided into two layers:
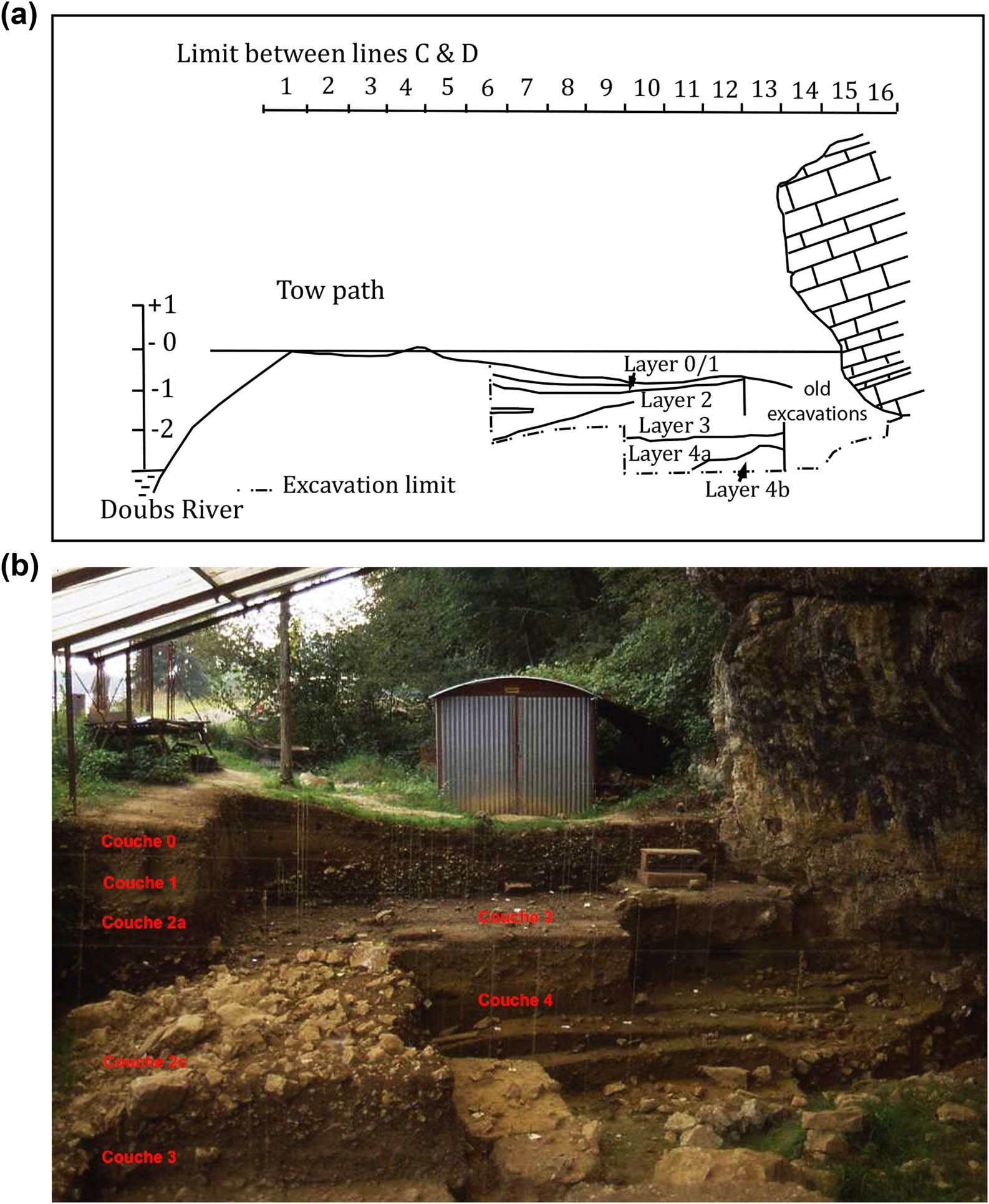
(a) Stratigraphy of Les Cabônes rock shelter. After S. Roué (2000). © N. Carquigny and (b) view of the excavation to the east with annotated stratigraphy. © C. Cupillard.
Layer 3 (formerly AOC, for Cryoclastic Organic Clay) is black and highly organic, rich in micro-charcoals of anthropic origin. It is 60–70 cm thick, thinning towards the Doubs around squares 6–8. It was excavated over 50 m2 for a potential surface area of 80 m2. The disturbances (burrows and ancient excavations) are generally located at the bottom of the shelter up to the porch. The outer zone of the site where material is very abundant probably corresponds to one or more discarding areas on the periphery of a dwelling area. However, little evidence of the latter subsists, apart from a hearth marked by a rubified zone in squares Z-A 14 (Figure 4). A lithic industry attributed to the Middle Sauveterrian forms most of the corpus; Early Mesolithic microliths such as crescents and oblique blunted points are also present at the base of the layer (Crotti & Cupillard, 2013; Cupillard et al., 2015; Roué, 2000). Isolated human remains were collected in this level, notably a skull fragment from square D9 dated to 8985 ± 40 BP (GrA-38019) belonging to a female individual related to haplogroup U5b1 (Posth et al., 2016) and the Villabruna cluster (Fu et al., 2016). Evidence from wild boar, red deer, and roe deer age-at-death data indicate a multi-seasonal use of the shelter (Leduc, Bridault, & Cupillard, 2015; Trémolières, 2020). The site might have been used (and reused) as a base camp as indicated by the abundance and diversity of the material recovered (Crotti & Cupillard, 2013; Cupillard, 2003). Layer 3 probably corresponds to an accumulation of several occupations dated between 8290 and 7060 cal. BC (2 sigma) (Table S2).

Excavation map of Les Cabônes shelter at Ranchot and sampling location of analysed fish remains from Layer 3; scale: a square represents 1 m2. CAD A. Bridault, modified from Leduc and Carquigny (Leduc et al., 2015).
Layer 2 (formerly AC, for Cryoclastic Clay) is light-brown in colour and thickens towards the Doubs. It is largely disturbed by burrows and contains rare Mesolithic flints as well as Neolithic, Protohistoric, and Gallo-Roman remains. Radiocarbon analyses of two red deer bones yielded dates between 7570 and 7180 cal BC (2 sigma) (Cupillard et al., 2015; Drucker, Bridault, Hobson, Szuma, & Bocherens, 2008).
A sterile grey clay layer (Layer 1) and a surface layer (Layer 0) seal this complex.
It is noteworthy that the sedimentary nature of the upper complex (layers 0–3) points to a very weak or non-existent influence of the river in the formation of the Mesolithic archaeological levels. This contrasts with the lower silty levels (4, 5, and 6) which, however, did not yield any fish bones.
3 Materials and Methods
3.1 Materials
The ichthyofaunal remains were collected in layer 3, after fine-mesh sieving (nested sieves of 10–1 mm) of all the sediments extracted during the excavation. The studied sample comprises a total of 9,328 remains grouped into 2 differently sorted subsets:
A first sample consists of 3,361 remains collected over a surface of 40 m2 and sorted with the naked eye and/or under a magnifying glass (2–5×) by the archaeologists in the 1980s and 1990s. A preliminary study was carried out on 438 of these by P. Morel who identified the presence of at least five species (Morel in Cupillard, Auguste, Cupillard-Perrenoud, David, & Morel, 1988): brown trout (Salmo trutta), chub (Squalius cephalus), barbel (Barbus barbus), perch (Perca fluviatilis), grayling (Thymallus thymallus) and, possibly, burbot (Lota lota) and pike (Esox lucius). This sample also contains remains collected on sight, mostly large Salmonidae remains.
The second sample was sorted by D. Frontin, a fish specialist, under a stereomicroscope (10–40×). It comprised 5,967 remains from five squares (H8/9, I9/10, and J10), equivalent to a volume of 13.63 L of sieving refuse resulting from the combined fractions of the sieving column (effective screen size 1 mm) (Figure 4). This area was selected because of the good preservation of the remains (Cupillard & David, 1995).
A small quantity of recent remains discarded on the site by a pair of kingfishers (Alcedo atthis) nesting in one of the shelter walls were mixed with these archaeological remains, but could be excluded from the sample because of their “fresh” appearance. Because of this, particular attention has been focused on determining the origin – anthropic or otherwise – of the archaeological ichthyofaunal remains.
3.2 Methodology
3.2.1 Taxonomic Identification
Prior to identification, a collection of reference skeletons of 22 freshwater species was created, selected for their historical (Bruslé & Quignard, 2006; Keith & Allardi, 2001) and current (ONEMA, unpublished 2008) presence in the Doubs basin. Only indigenous species were retained. 1–5 specimens were prepared per species for a total of 53 individuals, the vast majority of which were obtained from scientific fishing carried out by the Jura departmental service of the National Office for Water and Aquatic Environments (ONEMA). Three additional species possibly present in the study area at the Lateglacial–Holocene transition complete the list of potential species, but do not appear in the reference collection as it was not possible to obtain adequate specimens. The spectrum of potential species thus amounts to 26 taxa (Table S3), of which no less than 16 belong to the Cyprinidae family.
3.2.1.1 Cyprinidae
The Cyprinidae family is widespread in French rivers and includes an exceptional diversity of species with varied biological and ecological characteristics. Identification at species level is therefore crucial for understanding the fishing economies of ancient societies. Unfortunately, the skeletal morphology of cyprinids is particularly constant from one species to another, and the existence of natural hybrids between species further complicates determination (Libois & Hallet-Libois, 1988). These species are therefore notoriously difficult to distinguish by morphoscopical analysis on archaeological remains. Radiographic analysis can, in some cases, facilitate identification (Desse & Desse, 1976, 1981, 1983), but the small size of the vertebrae found at Ranchot (85% of these elements measure less than 3 mm in diameter) rules out this option.
We therefore limited our identifications to the morphological analysis of the three most diagnostic bone elements present in our material: the pharyngeal arch, the basioccipital, and the second vertebra, applied only to elements over 50% completeness. Identification was based on the protocols and identification keys defined by Le Gall (1982, 1984), and supplemented with subsequently published observations (Bruslé & Quignard, 2006; Keith & Allardi, 2001) and a careful examination of our reference skeletons and individuals from photographic atlases (pharyngeal arch only) (Froese & Pauly, 2021; University of Nottingham, 2021). This allowed us to enrich the published keys with seven new species for the pharyngeal arch and basioccipital, and six new species for the second vertebra. The resulting identification keys are provided in the appendix of this article (Figures S1 and S2).
Due to the fragmented state of the cranial bones, it was however often impossible to observe, on a given bone, every criterion required for identification. A probabilistic approach was therefore implemented: remains not identifiable to species were classified by degree of morphological proximity to different groups of species, according to a resemblance gradient. Data were then summarised by skeletal part, and the three lists of species obtained were compared with each other, in order to obtain a final list of the cyprinid species present, probably present, and possibly present at Ranchot.
3.2.1.2 Salmonidae
Only one indigenous salmonid species,[2] the brown trout (Salmo trutta), is currently present in the Doubs basin today. It has, however, been hypothesised that salmon (Salmo salar) populations may have found refuge in the Mediterranean at the end of the Last Glacial, which implies the species might have reproduced in our study area. This hypothesis is based on the identification of salmon bone remains at a dozen French, Italian, and Spanish Palaeolithic and Mesolithic sites in the Mediterranean basin (Kettle, Morales-Muñiz, Roselló-Izquierdo, Heinrich, & Vøllestad, 2011), and on the presence of a salmon depiction at the Grande Grotte de Bize, in Aude (Le Gall, 2001). These identifications are controversial, and the remains have been alternatively attributed to trout or to specimens transported from the Atlantic basin (Desse & Desse-Berset, 2002; Desse-Berset, 1994; Kettle et al., 2011; Le Gall, 1999). However, recent ecological niche modelling tends to confirm that conditions in the Mediterranean during the Lateglacial period may have been favourable for the establishment of salmon (Kettle et al., 2011). Salmon could thus be potentially present at Ranchot.
Brown trout and salmon both belong to the Salmo genus, and present such skeletal similarities that it is only possible to distinguish them morphologically on two cranial elements (premaxilla and glossohyal) and on the first vertebra (Le Gall, 1982), bones which are almost absent from the Ranchot material. Researchers have adopted different approaches to address this long-standing difficulty: vertebral radiography (Desse & Desse, 1983), geometric morphometry (Guillaud, Cornette, & Béarez, 2016; Huber, Jorgensen, Butler, Baker, & Stevens, 2011; Moss, Judd, & Kemp, 2014), ancient DNA (Moss et al., 2014; Yang, Cannon, & Saunders, 2004). In addition, stable isotopes have been successfully used to differentiate sea-run from landlocked salmon (Sr/Ca ratio; Robinson, Jacobson, Yates, Spiess, & Cowie, 2009) and salmonid migratory ecotypes (δ13C and δ15N; (Guiry et al., 2020) on archaeological material. For our part, we decided to approach trout/salmon determination by combining vertebral radiographs with 13C and 15N isotope analyses. Salmons are anadromous migrants, which, after spending part of their lives at sea, return to spawn in the river where they were born. As stable carbon and nitrogen isotope analyses can discriminate between marine and freshwater diets, spawning salmon[3] are expected to have a different isotopic signature than non-migratory trout (Bocherens, Polet, & Toussaint, 2007; Drucker & Bocherens, 2004; Robson et al., 2016). Isotopic analyses were conducted on ten salmonid vertebrae at the Laboratory of Isotopic Biogeochemistry of the University of Tübingen, Germany, by D. Drucker. However, the collagen in the remains was too degraded to obtain reliable results, and frontal X-rays of the vertebrae did not yield better results. Given that the presence of salmon in the Doubs basin during the Mesolithic has not been confirmed to date, and that the only two first vertebrae of the Salmo sp. of the collection were both attributable to trout, we chose to assign all of the Ranchot salmonid bones to this latter species. However, the presence of salmon remains in our sample cannot be strictly excluded.
3.2.1.3 Other Species
The ranges of potential taxa for the other fish families present in Ranchot are limited to one or two species. Their identification was based on our reference collection and on reference publications (Desse, Desse-Berset, & Rocheteau, 1987, 1990; Le Gall, 1982, 1984; Libois, Hallet, & Rosoux, 1987; Picquès, 1998; Thieren, Wouters, Neer, & Ervynck, 2012).
3.2.2 Quantification
Teleost assemblages are typically quantified using indices initially developed for the study of terrestrial vertebrates. Some of these indices (number of specimens (NSP), number of identified specimens (NISP), Weight, etc.) are applicable to ichthyofaunal assemblages, whereas others, such as the minimum number of individuals (MNI), are more problematic. Indeed, many fish bone assemblages are dominated, sometimes very largely, by vertebral remains, and the most commonly used calculation for the MNI is based on counting the minimum number of elements of the most frequent single skeletal part (lateralised if necessary) in the assemblage. Yet teleost vertebral remains cannot be attributed to a single skeletal part. Not only do vertebra characteristics, which are not very variable along the vertebral column, limit anatomical determination to an anatomical type (“thoracic B”) and not to a precise rank (“12th thoracic vertebra”), but the very number of vertebrae within a type is variable for a given species, as are the numbers of vertebrae between individuals of the same species. However, the MNI is a useful index for understanding assemblages in terms of the acquisition and exploitation of individuals. An eel vertebra, a species with between 110 and 120 vertebrae, will not have the same significance in terms of the number of individuals caught as a roach vertebra, which has between 39 and 41 vertebrae (Froese & Pauly, 2021).
To overcome this difficulty, we chose not to carry out MNI calculations, as these would be based on a very limited number of cranial bones, and would only have yielded very low and biased results. We therefore opted for a weighted number of vertebrae relating all species to the same theoretical standard. This method was initially developed by Sternberg (Sternberg, 1989) and is based, for a given species, on the weighting of the number of its independent vertebrae (centrums with more than 50% preserved, i.e. more than 90% of the vertebrae at Ranchot) by a multiplier index representing the ratio between the average number of vertebrae of the most common species in the assemblage, taken as reference species, and the average number of vertebrae of the examined species. This weighted number of vertebrae, called Corrected Expected Number of Vertebrae (NVcor), makes it possible to establish a direct proportional relationship with the number of captured individuals, while avoiding the MNI. Furthermore, unlike cranial elements, which present inconsistent archaeological preservation depending on species, vertebrae are only slightly affected by differential preservation, and are thus ideal for assessing specific representation in the collection.
At Ranchot, the choice of standard species was complicated by the fact that while cyprinids represented over 96% of the assemblage, only a very small proportion of the remains could be attributed to a specific species. We therefore decided to calculate an average vertebra count for a standard “generic” cyprinid, based on the average of the minimum and maximum vertebra counts observed for each species in the assemblage. The calculated multiplier indices are presented in Supporting Information (Table S4). The abbreviations NV (number of vertebrae over 50% completeness) and NVcor will be used to present the results of the method.
3.2.3 Evaluation of Catch Size and Weight
The so-called continuous growth of teleosts makes it possible to establish a mathematically modelled relationship between bone dimensions and the size and weight of the specimen for most species. In order to reconstruct the size distribution of catches at Ranchot, each single vertebra was determined to the species, classified according to its anatomical type along the rachis, and then measured with an electronic calliper to an accuracy of one hundredth of a millimetre, in keeping with the conventional measurements defined by Morales and Rosenlund (Morales & Rosenlund, 1979). The size and weight of the individual were then estimated by applying linear or polynomial equations defined according to the species, vertebral type, and chosen vertebral measurement. The following references were used: Cravinho (2009) for cyprinids, Desse et al., (1990) for burbot, Desse et al., (1987) for perch, Picquès (1998); Thieren et al., (2012) for eel, and Hajkova, Roche, & Kocian (2003) for trout and grayling. The equations for roach, which is definitely present in the collection, were retained for the estimation of the size and weight of indeterminate cyprinids.
It should be noted that, depending on the species, the entire rachis or only certain sections can be used for these size reconstructions. In the case of eel and burbot, in particular, only the thoracic vertebrae can be used. This implies that catch weights calculated per species cannot be directly compared with each other. Again, a weighting of the results was applied: the total weight obtained for a given species was divided by the number of vertebrae used for the reconstruction, and this ratio was then multiplied by the NV of the species, and then related to the standard species (NVcor) by the use of the previously defined multiplier indices.
The catch weights obtained with this mathematical correction have no tangible certainty. On the other hand, they are proportional to the real weight of the catch, and this is appreciable, even if the latter remains inaccessible to us. The relative contributions of each fish species to the total amount of fish eaten on site, for example, can thus be reliably assessed.
3.2.4 Taphonomic Analysis and Nature of the Assemblage
Before interpreting the results of a faunal analysis in terms of past human behaviour, it is always necessary to ascertain the anthropogenic nature of the collection studied. Indeed, various natural phenomena can also lead to the formation of bone accumulations in rock shelters, namely, the action of fish-eating animal predators and trapping phenomena following a flood (Russ & Jones, 2011). The aim here is thus to compare the characteristics of the archaeological assemblage in terms of specific composition, size of remains, anatomical distribution, and taphonomic status with those typically observed in naturally occurring accumulations. Six piscivorous predators likely to produce rock shelter bone accumulations were identified in the study area, two mammals, the domestic dog Canis familiaris and the otter Lutra lutra, and four birds, the eagle owl Bubo bubo, the barn owl Tyto alba, the kingfisher Alcedo atthis, and the dipper Cinclus cinclus. References for the osteological characteristics of accumulations produced by the first four species existed in the literature and were used here, respectively, (Jones, 1986, 1984; Nicholson, 1993), (Guillaud, Béarez, Denys, & Raimond, 2014; Guillaud, Bearez, Denys, & Raimond, 2017; Nicholson, 2000), (Guillaud, Lebreton, & Béarez, 2018; Laroulandie, 2000; Russ, 2010), and (Broughton, Cannon, Arnold, Bogiatto, & Dalton, 2006). However, to our knowledge, no such references existed for kingfishers and dippers, so we set out to create our own. The bone remains accumulated during a season by a pair of kingfishers breeding in the Meuse basin in Belgium were collected and sieved. A 10 mL sample of sieve refusal was analysed, yielding 5,868 ichthyofaunal bone elements. Specific diagnosis was carried out on the 10 most diagnostic cranial elements of the assemblage, i.e. 552 remains, and the metric was carried out on a random sample of 500 vertebrae. The surface conditions of the latter were also examined in keeping with the work of Butler & Schroeder, 1998; Nicholson, 2000. A summary of the obtained results is presented later in this article. The detailed results of this work will be the subject of a specialist publication, in preparation. On the other hand, no bone from dippers’ pellets could be obtained. In view of the very similar biological and ethological characteristics of the dipper to those of the kingfisher (size, feeding and dumping habits, nesting sites, etc.), we considered that the data from the latter species could constitute an acceptable proxy.
In order to undertake a comparison with the data of the accumulating agents, and to gain a better understanding of the taphonomic history of the collection, a survey of the surface conditions of all 9,328 remains of the Ranchot collection was carried out, noting for each the possible presence of pitting, rounding, deformation, and staining, as well as the presence of traces of fire marks and cutting marks, following the descriptions of Asmussen (2009); Butler & Schroeder (1998); Willis, Eren, & Rick (2008); Willis & Boehm (2014); Zohar, Ovadia, & Goren-Inbar (2016).
3.2.5 Seasonality of Catches
Finally, an assessment of fishing seasons was conducted through a skeleto-chronological analysis of a sample of 500 complete vertebrae selected for their excellent state of preservation. The method consists of the observation of growth rings on the articular faces of the vertebrae, which are presented in the form of alternating broad light-coloured annuli, corresponding to the growth of the individual during the warm season, and thin dark bands, corresponding to the cold season. The development stage of the ring on the outer edge of the vertebra allows us to assess the season of death of the individual. The protocol followed was adapted from the recommendations of Desse & Desse-Berset (1992); Le Gall (2003); Meunier (1988); Wheeler & Jones (1989). A comparative reference framework was established by analysing 183 vertebrae from the reference collection from the study area and from known capture seasons. Each archaeological vertebra was then examined three times, at intervals of several months, without taking note of the previously acquired results. Only data from elements that gave consistent results at all three appraisals were retained in our final results.
4 Results
4.1 Taxonomic Composition of the Studied Assemblage
The comparison of the faunal spectra resulting from the two sorting methods (by archaeologists and by the specialist) showed no difference in the representation of the identified species (Frontin, 2017). The results are therefore presented altogether.
Six families and seven species make up the Ranchot Layer 3 spectrum (Table 1). Cyprinid remains are largely predominant (96.6%), of which three species are attested with certainty – common bream, roach, and minnow.
The latter were identified using several diagnostic elements (basioccipital, second vertebrae, and cranial elements), only slightly affected by fragmentation.
The other families are represented by only one species each - trout with 243 remains (i.e. 2.6% NISP), grayling, burbot, perch, and eel, with only a few remains each (i.e. less than 1% NISP).
The species making up this group have fairly similar ecological habits. They are mostly gregarious and diurnal, usually live close to the bottom or in open water, and circulate in rather cool, well-oxygenated, and relatively running waters. The eel, the only migratory species, also adapts to these diverse environments. The predominance of cyprinids in the assemblage undoubtedly reflects an environmental reality. It is the most represented family in the environment in terms of number of species (Keith & Allardi, 2001).
4.2 Fragmentation Rate of the Material
The state of fragmentation of the material is estimated using a percentage of completeness assigned to each specimen (Butler & Schroeder, 1998; Nicholson, 2000). For fish vertebrae, which are the most abundant bones in the assemblage, the condition of the centrum is recorded, as spines are often absent in archaeological material. Less than 10% of a sample of 1,184 vertebrae are less than 50% complete (Table 2). Consequently, fragmentation is low and has little effect on identification. On the other hand, the opposite configuration was observed for the pharyngeal arch, an element of a masticatory apparatus in cyprinids, which is quite resistant but often found fragmented. The presence or absence of teeth has not been taken into account, as they are easily detached. Only complete or well-preserved arches, which represent a minority (11 are 50% complete or more), were used in order to produce a reliable diagnosis.
Completeness of cyprinid pharyngeal arches and vertebrae; sample of 1,971 layer 3 specimens
| 1–24% | 25–49% | 50–74% | 75–100% | N | |
|---|---|---|---|---|---|
| Pharyngeal arch – cyprinids | 390 | 308 | 68 | 21 | 787 |
| Vertebrae – all taxa | 58 | 51 | 97 | 978 | 1,184 |
4.3 Bone Modifications
Overall, bone surfaces are well preserved and only 13% of the remains show modifications (Table 3). No pitting marks, characteristic of the attack of bone surfaces by digestive juices, were recorded in the assemblage. Bone rounding caused by sediment abrasion is the most common modification found on any bone element, for all species and all sizes of specimens. It is clearly differentiated from rounding due to transit, on account of the absence of the characteristic rounded and glossy appearance of the latter (Llona & Andrews, 1999), which is more often confined to the extremities of elements and does not extend over entire surfaces. Only two types of deformations were found on vertebrae: “compression” and “undulations.” Compression of the articular faces, or even more rarely of the entire vertebral body, was only observed on small specimens (N = 46), mainly cyprinids and on a few salmonids. Two vertebrae presented crushing. The first shows crushing associated with a torsion of the vertebral body and the other displays crushing associated with a deformation of the lateral decoration. All of these deformations are related to traces of mastication described in the literature (Butler & Schroeder, 1998). These bones were chewed, but not ingested; instead, they were spat out after the flesh was consumed. This would therefore indicate that small fish were of dietary interest. The second type of deformation takes the form of “ripples” on the edges of articular faces (N = 44). This was only observed on white-coloured bones, which seem to result from heating to very high temperatures (calcination). Finally, changes in bone colour (from black to white) indicate different stages of heating, from carbonisation to calcination. These burn marks are observed on about 4% of the remains of all types of bones, all species, and all sizes of individuals taken together. They are probably not related to the cooking of fish but more logically to the burning of bone waste.
Bone modification values recorded on fish remains in the Ranchot Layer 3 assemblage (complete sample)
| Pitting | Rounding | Deformation | Staining | All | NSP C3 | |
|---|---|---|---|---|---|---|
| N | 0 | 727 | 90 | 368 | 9,328 | |
| % NSP C3 | 0 | 7.79 | 0.96 | 3.95 | 12.7 |
4.4 Representation of Skeletal Parts
The distribution of bones of all taxa into three anatomical units shows that axial skeleton bones are predominant (77.2%), mainly vertebrae. The presence of elements attached to the vertebral column (ribs, Weberian ossicles) is very marginal (Table 4). The appendicular skeleton is only documented by a few fin rays (0.1%). The cranial elements (basioccipital, pharyngeal arches, and teeth) account for 22.7%.
Relative abundance (NISP) of fish skeletal elements grouped by anatomic unit (%) – Ranchot Layer 3 assemblage (complete sample, all taxa)
| Anat. unit | Element | NISP | % |
|---|---|---|---|
| Head | Basioccipital | 17 | |
| Frontal | 1 | ||
| Dentary | 1 | ||
| Quadrate | 1 | ||
| Premaxilla | 1 | ||
| Hyomandibular | 1 | ||
| Opercular | 1 | ||
| Pharyngeal arch | 787 | ||
| Teeth | 1,304 | ||
| Total | 2,114 | 22.69 | |
| Appendicular skeleton | Pterygiophore | 5 | |
| Acanthotriche | 1 | ||
| Total | 6 | 0.06 | |
| Axial skeleton | Vertebrae | 7,121 | |
| Ribs | 62 | ||
| Weberian ossicles | 13 | ||
| Total | 9,316 | 77.22 | |
| Scales | 3 | 0.03 | |
| Total identified | 9,319 | 100.00 | |
| Unidentified element | 9 |
Only cyprinids are represented by different skeletal parts, including head bones. The perch is only represented by a few cranial remains, while the other species are only represented by their axial skeleton. The vertebrae are significantly more abundant at Les Cabônes (76.4% of NISP) than expected in a complete cyprinid skeleton (23%);[4] head and appendicular skeleton are significantly under-represented (χ 2 = 987.81, p < 0.0001). How can this distribution of anatomical units be interpreted? As no traces of incision were observed on the bones, it is impossible to assess whether the fish were cut up before consumption. The under-representation of the cranial elements could reflect heading before being brought to the site, but in view of the small size of the catches (see below), this seems unlikely. On the other hand, their destruction in situ by trampling and/or weathering is plausible. Most of the cranial elements have a lower bone density than post-cranial elements (vertebrae in particular), and are not very resistant to destructive factors (Butler, 1996; Butler & Chatters, 1994). Teeth, the basioccipital, or pharyngeal arch are the most resistant cranial elements, and often survive best in archaeological material (Le Gall, 1982, 1984; Wheeler & Jones, 1989). The differences observed in the degree of fragmentation of pharyngeal arches and (more complete) vertebrae are consistent with this (Table 3). Moreover, the remains of salmonids and thymallids are more degraded here than the remains of other taxa, which is in keeping with taphonomic logic.
At Ranchot, the most resistant elements are thus the most abundant: vertebrae for all species, basioccipitals and pharyngeal arches for cyprinids, and teeth for cyprinids and percids. Consequently, it is likely that the differential representation of the skeletal parts of this assemblage results mainly from the in situ post-depositional destruction of preparation and/or consumption waste.
4.5 Characterisation of Captured Individuals
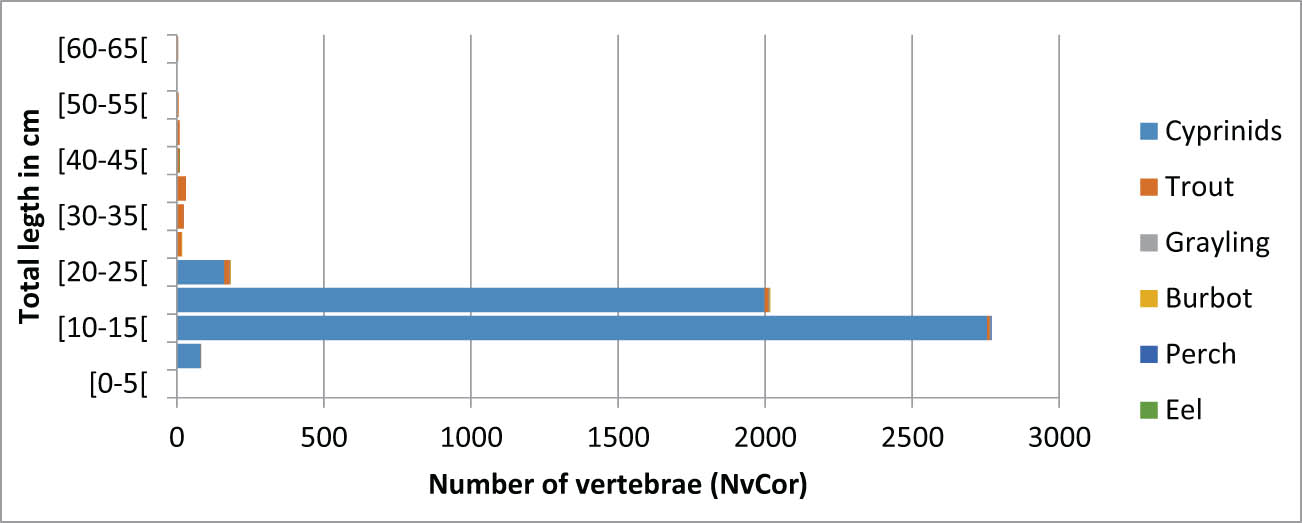
The relative representation of species in terms of the number of individuals caught was not assessed in MNI, due to the characteristics of the assemblage, but through %NVcor (refer methodology section) (Table 5). The method confirms the strong predominance of cyprinids (97%) established on the basis of the count in number of remains. In comparison, the other species, even the best represented species such as trout, represent an anecdotal number of catches.
Relative abundance of fish taxa represented in the Ranchot Layer 3 assemblage: Nvert = number of vertebrae, NVcor = corrected number of vertebrae
| Nvert | % Nvert | NVcor | % NVcor | |
|---|---|---|---|---|
| Cyprinids | 4,997 | 95.7% | 4,997 | 96.9% |
| Trout | 192 | 3.7% | 140 | 2.7% |
| Grayling | 19 | 0.4% | 14 | 0.3% |
| Burbot | 7 | 0.1% | 5 | 0.1% |
| Eel | 5 | 0.1% | 2 | 0.0% |
| Total | 5,220 | 100.0% | 5,158 | 100.0% |
This very large number of cyprinids is somewhat tempered by the small catch size. The reconstruction of the total length (TL) of individuals on a sample of 185 cyprinid vertebrae shows a distribution of values between 9.9 and 21.1 cm, with a median of 14.5 cm. Small cyprinids therefore appear to have been targeted. According to Fishbase (Luna, 2021b), the common size of a roach is 25 cm, and the average size of common cyprinid species potentially present in the assemblage is around 20 cm. The individuals from Ranchot therefore appear significantly smaller than expected. The size distribution of catches (Figure 5) forms a quasi-symmetrical bell curve, suggesting random recruitment of individuals.
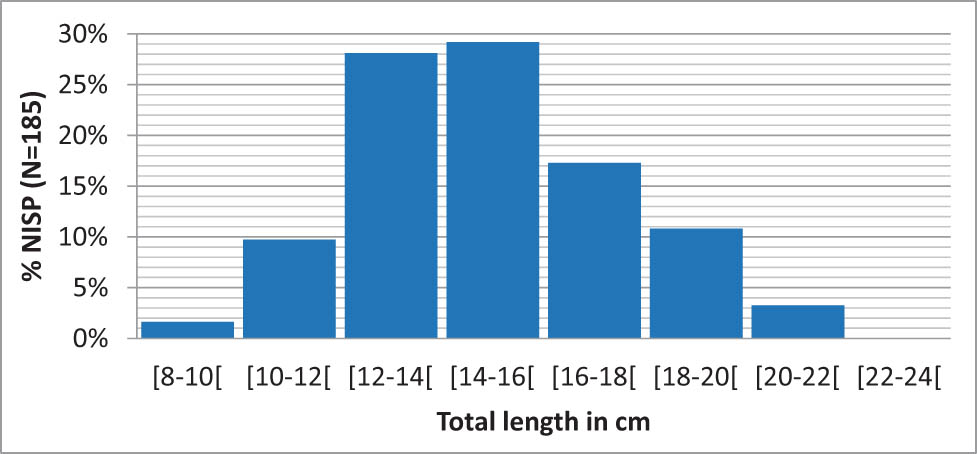
Reconstructed size distribution of cyprinids from layer 3 (N = 185 vertebrae).
This pattern also seems to be observed among certain rare species in the assemblage. Grayling shows a median size of 17.5 cm at Ranchot, for 17 vertebrae, whereas individuals caught today are commonly 30 cm in length (Luna, 2021c). Due to the small sample size, the distribution is not very precise, but shows a predominance of individuals between 10 and 25 cm, which is consistent with data for cyprinids (Figure 6). Similarly, the four reconstructed remains of burbot show sizes between 17.4 and 25.4 cm, for a median of 20.1 cm, well below the commonly expected size of 40 cm (Luna, 2021a). Finally, the only reconstructed perch remain is also from a small individual, estimated at 14.5 cm for a common species size of 25 cm.
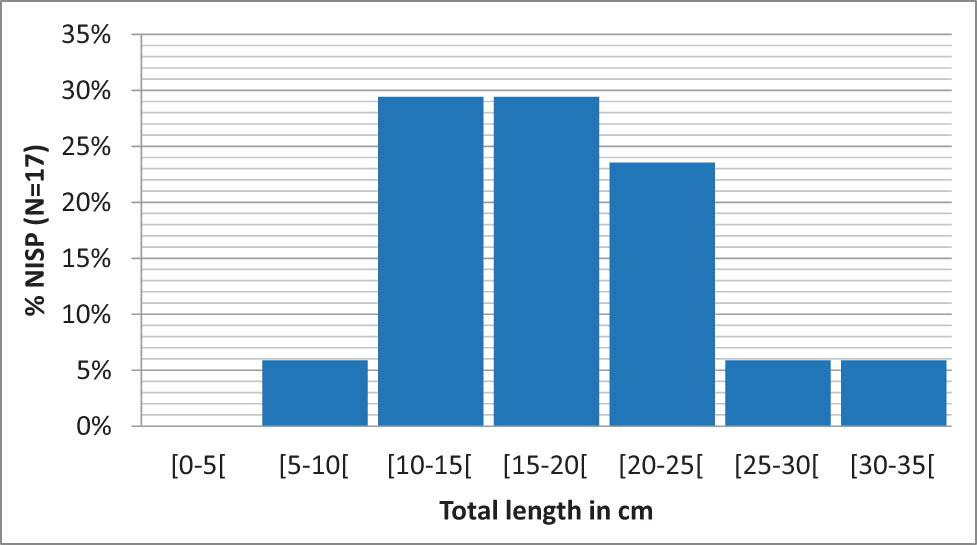
Reconstructed size distribution of grayling from layer 3 (N = 17 vertebrae).
Salmonids show a different pattern. On the one hand, reconstructed individuals (165 vertebrae) are larger and much closer to expected sizes, with TLs ranging from 6.6 to 64.2 cm, for a median of 33.5 cm. Today, the common size of a brown trout is 35 cm, even if record-breaking individuals can reach 1 m (Rochefort & Corolla, 2019). The reconstructed individuals are therefore consistent with these observations. On the other hand, size distribution here is anything but symmetrical (Figure 7). The curve gradually increases from 5 to 40 cm, reaches a mode in the [35–40 cm] class, which alone accounts for 22% of the catches, and then falls sharply beyond that, with only 6% of the catches measuring between 40 and 45 cm. There thus seems to be a threshold effect limiting the presence of large catches.
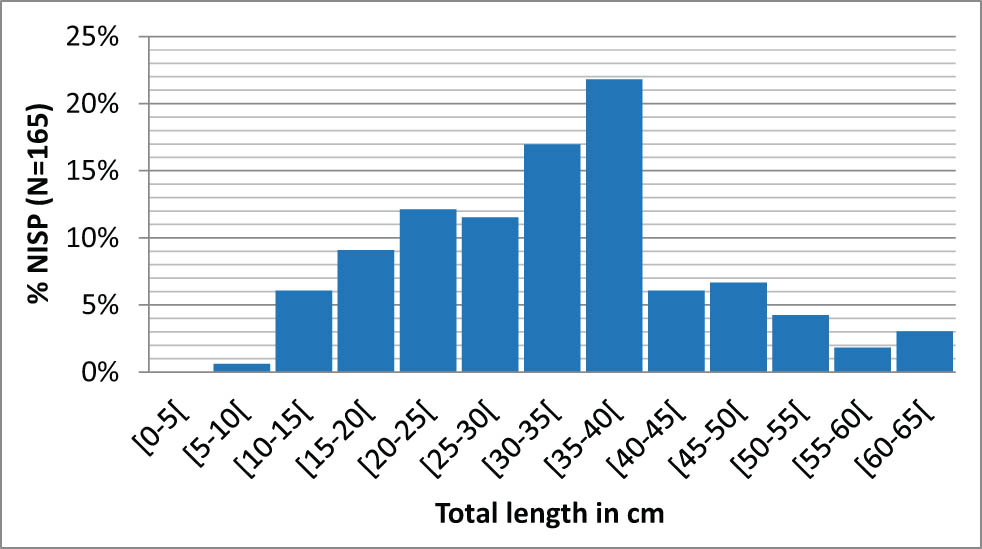
Reconstructed size distribution of salmonids from layer 3 (N = 165 vertebrae).
The eel could show a similar pattern. The sizes of the only two reconstructed individuals are 40.5 and 41.5 cm, in line with the common size of the species, 40 cm (Froese, 2021).
Based on data obtained for each species, we can also propose a reconstruction of catch sizes for the collection as a whole. To do this, we divided the NVcor of each species into size classes proportional to those found in the samples used for size estimates, and then added them together. The resulting projection, shown below in Figure 8, clearly illustrates the predominance of small cyprinids in the assemblage: almost 94.5% of the catches are less than 20 cm in total length, 99.3% of which are cyprinids. Medium to large fish are thus very poorly represented, with only 2.1% of catches exceeding 25 cm, 95.4% of which are salmonids.
Projected size distribution of reconstructed fish sizes, all taxa, from layer 3, N = 5,158 (size distribution patterns observed for each species have been applied to total corrected number of vertebrae from the species).
4.6 Food Contribution of Species
Using the measurements taken to reproduce catch size, it is also possible, on the same sample, to estimate weight. We were thus in a position to calculate an average specimen weight for each species in the assemblage, varying between 35.4 g for cyprinids and 495.7 g for trout, as presented in Table 6. In order to assess the relative contribution of species to the diet of the Ranchot occupants, these values were then multiplied by the NVcor of each species. The results of this transformation being strictly proportionate to the number of captured individuals, they allow the unbiased estimation of the part of each species in the fish eaten on site. The exact figures for the weight provided by each species are nonetheless theoretical constructs, and must not be taken at face value.
Estimated relative amount of each fish species in kilograms based on corrected number of vertebrae (NVcor); average weight of specimens calculated on vertebral measurements (samples in Figures 5–7)
| Average weight (g) | NVcor | % NVcor | Weight (kg) | Weight (%) | |
|---|---|---|---|---|---|
| Cyprinids | 35.4 | 4,997 | 96.88% | 177.14 | 71.3 |
| Trout | 495.7 | 140 | 2.71% | 69.39 | 28.0 |
| Grayling | 82.9 | 14 | 0.27% | 1.16 | 0.5 |
| Burbot | 65.6 | 5 | 0.10% | 0.33 | 0.1 |
| Eel | 124.0 | 2 | 0.04% | 0.25 | 0.1 |
| Total | 5,158 | 1 | 248.26 | 100.0 |
The results show that although cyprinids still dominate, trout made a significant contribution to the diet, accounting for more than a quarter (28%) of the consumed fish, despite the small number of remains (2.71% of NVcor).
4.7 Fishing Seasonality
Due to the very small size and state of preservation of remains, only 82 vertebrae had the required characteristics for a skeletal-chronological analysis of the season of capture. Of these, only 57 (70%) provided the three concordant readings required to ensure analysis reliability (sample details in Supporting Information, Table S4). Our study of catch seasonality is therefore based on a relatively limited sample. The results nevertheless show a strong structuring of the data (Figure 9): more than 57% of the catches occurred at the beginning of the good season. The number of catches then decreased rapidly as the summer season progressed, and only 7% of individuals appear to have been caught in the bad season. Thus, although fish could be caught all year round in Ranchot, it seems that this resource was mainly exploited in early spring, a period of food shortage known to be difficult and poor in resources.
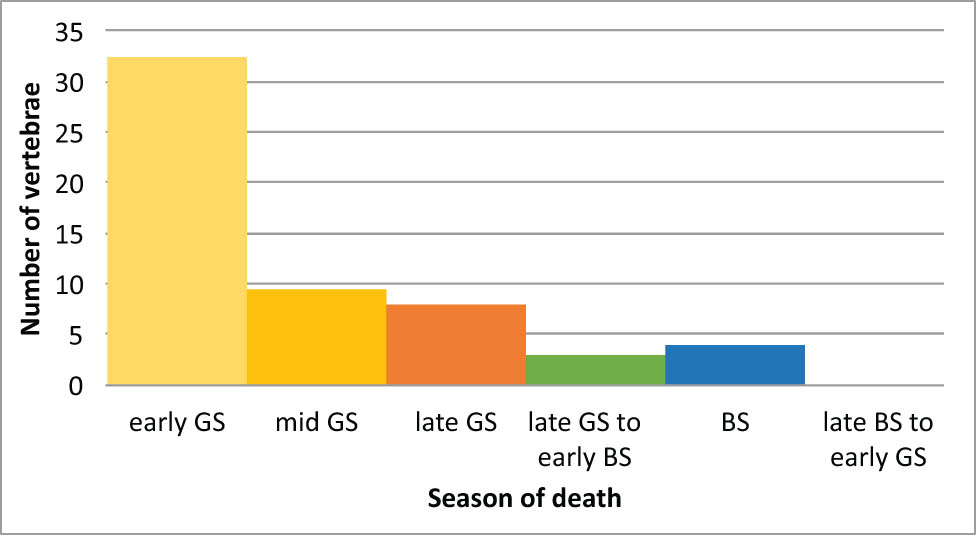
Distribution of identified season of death on a sample of 57 fish vertebrae from layer 3; GS = good season, BS = bad season.
No structuring of the data by species was observed.
5 Discussion
5.1 Origin of the Bone Accumulation
The question of the origin of the bone accumulation is central to any ichthyofaunal study, but was particularly central at Ranchot. The observation of present-day kingfisher pellets during the excavation immediately raised the possibility of a natural accumulator, and the unusually small size of the remains did not help to dismiss this hypothesis.
Specific attention was therefore paid to this problem during our study, in particular, through the analysis of a sample of remains from kingfisher pellets, leading to a better characterisation of this type of accumulation. A summary of the main results of our observations is presented in Table 7 below.
Criteria for identifying a Kingfisher pellets assemblage
| Criteria | Kingfisher accumulation characteristics |
|---|---|
| Skeletal part representation | Presence of complete skeleton, body-part representation similar to that of complete fish |
| Species identification | Broad spectrum, variable with species availability opportunistic fishing, favouring gregarious, benthic, and crepuscular/nocturnal species |
| Size of the remains | Centrum diameter always under 2 mm |
| Range 0.41–1.94 mm; average 1.16 mm; | |
| 77.4% of remains between 0.8 and 1.4 mm | |
| Prey size reconstruction (Cyprinids) | Range 5.4–13.5 cm TL; average 9.2 cm |
| Most preys under 10 cm TL, but variable according to species (head diameter and body shape) | |
| Vertebrae completeness | Excellent preservation, minimal breakage >90% of fully complete remains |
| Surface modifications | No modification of bone surfaces: no pitting, staining, rounding, or visible deformation of bones |
Fortunately, these data exclude the intervention of this accumulating agent with almost complete certainty. If we consider the Ranchot assemblage as a whole, it is clear that all the larger individuals, with vertebrae larger than 2 mm in diameter and/or with a reconstructed size larger than 15 cm, could not have been the prey of such a small bird. However, a mixture of kingfisher regurgitation with another accumulation process remains possible, and we thus carried out a more detailed comparison between the characteristics of the cyprinids from Ranchot (which overwhelmingly dominate the smaller remains) and those of the fish consumed by the kingfisher. This comparison shows that although the cyprinid remains from Ranchot are small, their average diameter is significantly greater than those from the kingfisher pellets (Student’s t = 8.42; p ≪ 0.001), and it is statistically unlikely that they are of the same origin. Above all, the reconstruction of sizes based on this sample shows that the vast majority of cyprinids captured at Ranchot, with an average size of 14.9 cm, exceed the consumption capacity of the kingfisher. Out of a total of 185 fish for which size could be estimated, all but one[5] exceed 10 cm, the maximum size of prey listed in the literature (Hallet, 1985; Libois & Hallet-Libois, 1995), and 61.4% exceed 13.5 cm, the maximum size observed in the reference assemblage. Finally, the unimodal, symmetrical bell-shaped size distribution curve of the Ranchot cyprinids (Figure 5) is more suggestive of a single origin of the population than a mixture of two populations resulting from distinct selection processes.
It is also possible to exclude most of the other potential accumulative agents of bone concentrations. Relatively few piscivorous predators can be found in caves and rock shelters, and mammals give rise to very different assemblages from those of Les Cabônes. The catch size observed at Ranchot is not incompatible with otter habits (typically from 10 to 30 g for cyprinids and percids (∼10–20 cm), and from 100 to 200 g for salmonids (∼30 cm) Guillaud et al., 2014), but the remains consumed by this species are significantly marked by digestive processes, with 90% of the remains bearing acid attack marks, resulting in polishing, pitting or surface dissolution, and tooth marks and deformations affecting 3% of the remains (Guillaud et al., 2014). As no digestion marks are observed at Les Cabônes, otter action can therefore be excluded. The dog produces assemblages that are even more marked by taphonomic degradation, with very intense fragmentation of remains, and often unrecognisable surviving bones, reduced to corroded fragments and marked by traces of digestion (Jones, 1986, 1984; Nicholson, 1993). Again, this appears to be incompatible with the state of preservation of the Ranchot remains.
Fish bone assemblages generated by the Eurasian eagle-owl (Bubo bubo) similarly display features inconsistent with that of the Ranchot assemblage. First, the bones ingested by owls are marked by digestive processes, though to a lesser extent than by mammals: about 9% of the remains display moderate to intense digestive alterations (Guillaud et al., 2018). Second, the eagle owl tends to select prey significantly larger than those found in our archaeological assemblage: Guillaud notes that the predominant weight class in the eagle owl sample included fish estimated between 200 and 300 g (Guillaud et al., 2018), whereas the average weight in the Cabônes sample was no higher than 48 g. Finally, no piscivorous specialisation of the eagle owl has to our knowledge been described (Russ, 2010), and assemblages generated by the species should therefore include many remains of birds and small mammals, mostly absent from the archaeological sample.
Finally, there remains the case of the barn owl (Tyto alba). This nocturnal raptor has an opportunistic diet that can in specific circumstances become primarily piscivorous. The bird shows a general preference for prey weighing 10 and 70 g (Broughton et al., 2006), consistent with most of the fish consumed at Ranchot. Pitting and deformations are observed on less than 5% of the remains, but 19% show traces of rounding, which has not been observed in the archaeological collection. Moreover, the piscivorous specialisation of the barn owl has only been recorded in North America, in the case of the seasonal abundance of dead or dying fish stranded in the pools of an intermittent stream (Broughton et al., 2006). These environmental conditions are far different from those found on the banks of the Doubs, a deep and wide permanent river, and the estimated season of capture (early spring) is similarly conflicting. Therefore, a broader-spectrum diet is to be expected for the barn owl, and, as for the eagle owl, the associated presence of small mammals should be expected were the accumulation produced by this species.
The inconsistencies between fish bone assemblages produced by animal accumulators and that of Les Cabônes appear therefore significant enough to exclude an animal origin of the archaeological collection. A natural origin can likewise be rejected: the stratigraphy of the shelter shows almost no influence of the Doubs River in the formation of the sediments of the Mesolithic levels having yielded fish remains. It seems therefore difficult to argue in favour of natural inputs of fish carried into the shelter during seasonal river flooding. Less importantly, the presence of the bream possibly upstream of its natural habitat also contradicts a natural fluvial origin. Finally, the size distribution pattern of salmonids, displaying a relatively brutal threshold for fish over 40 cm, appears inconsistent with a natural accumulation.
We therefore consider that an anthropogenic origin of the archaeological fish assemblage of Ranchot can be accepted, even though a marginal contribution by other accumulators cannot be ruled out entirely.
The spatial distribution analysis of fish bones shows a fairly homogeneous overall distribution of skeletal elements with, however, denser accumulation zones (Z-A-B 10–12, C10, H-I-J 8–10) coinciding with the preferential accumulation zones of lithic remains (Roué, 2000). Nevertheless, it is noteworthy that Salmonidae remains seem to be preferentially discarded near the hearth. Masticated and burnt remains are also preferentially located near the hearth.
5.2 Prey Acquisition
5.2.1 Fishing Environments
Fish are particularly sensitive to the physico-chemical and biological conditions of their living environment, and each species has specific environmental requirements. The slightest variation in these conditions influences its presence and demography in a given place. In a river environment, we observe a longitudinal organisation from the source to the mouth, in zones associating environmental conditions and well-defined fish populations. The most commonly used typology, proposed by Huet in 1949, defines five ecological zones with their associated species (Huet, 1949). Based on this range of species, it is thus possible to evaluate the capture zone(s) of individuals with an ichthyofaunal spectrum.
We thus plotted in Figure 10 the ecological preferences of the fish species identified at the site on the ecological zones of the Doubs (terminal zone not represented) (Verneaux, 1973).
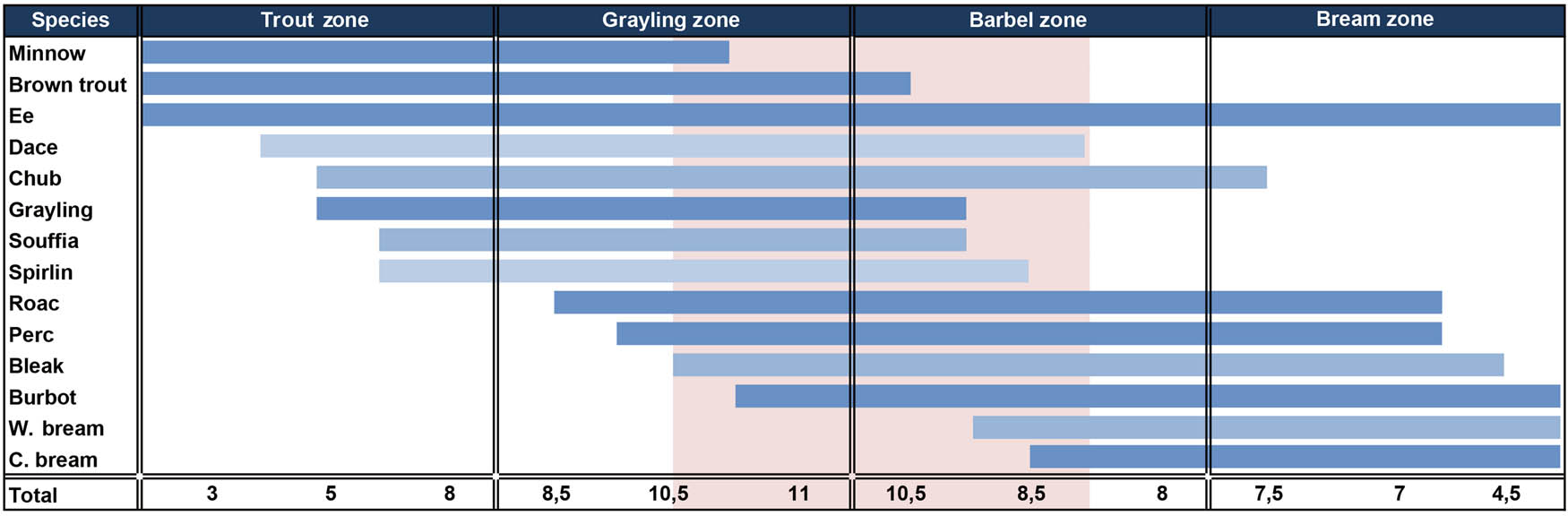
Ecological requirements of the fish species identified at Ranchot, plotted along the longitudinal zonation defined by Huet (1949), from upstream (Trout zone) to downstream (Bream zone). Total: number of Ranchot species present in sub-zone; pink area: minimal river area including all of the species documented at Ranchot.
The first observation is that, according to these data, it is highly unlikely that all the fish in the shelter were caught in a single location. The requirements of minnow and brown trout are incompatible with those of the two breams in terms of current, oxygen, and temperature. Fishing therefore seems to have been organised into river sections, with catches in different places. At the very least, for the capture of all the identified species, the fished area had to cover the downstream half of the grayling area and the upstream two-thirds of the barbel area. In addition, incursions into more distant fishing grounds cannot be excluded. Nevertheless, species composition remains consistent: 11 of the 14 determined species are typically present at the transition between the grayling and barbel zones, which could indicate a preferential fishing area.
The species identified in the corpus, and in particular cyprinids, generally prefer shallow waters with moderate currents. These species are therefore mainly found on the banks of the main channel or in secondary arms. Only grayling and the perch show divergent ecological preferences, favouring, respectively, the benthic zone of the main channel and slow-moving backwaters. These two minority species in the corpus may however move to secondary arms during the reproduction period. This points to the existence of a preferential catching environment for the whole assemblage.
5.2.2 Fishing Techniques
In France, during the Mesolithic, “obvious” fishing apparatus is rare. In continental and fluvial environments, only the “Haut-des-Nachères” deposit at Noyen-sur-Seine yielded exceptionally well-preserved remains of basketry traps made of privet and a wickerwork container in an anaerobic wetland environment (Mordant et al., 2013). In the European Mesolithic, numerous evidence of passive fishing securely dated exists. Fish traps, stakes, weirs, and wattle fences are recorded from Late Mesolithic contexts (e.g. McQuade and O’Donnell, 2007; Koivisto, 2017). At Ranchot, no such xylological remains were preserved, on account of deposition and conservation conditions, and the use of fishing tackle remains hypothetical. The Ranchot site did not yield any deer antler harpoons either, although these are known regionally during the Late Mesolithic (Cupillard, 1998a,c; Marquebielle, 2014). Some authors have suggested that certain types of microliths and armatures were used as arrow barbs for fishing (Rozoy, 1978), but there is no evidence for this at Ranchot.
The results of the ichthyological analysis described above shed light on the different fishing methods that may have been used at Ranchot. The use of traps (or nets) designed to capture small individuals is the most likely fishing technique to explain the observed cyprinid size (normal distribution around 14.5 cm). We cannot rule out the possibility that the graylings and burbots of the assemblage, which are slightly larger but more elongated than cyprinids, could have been caught in these same traps. Such traps could have been set in more or less deep water in diverse environments and could have captured both diurnal (bream, roach, minnow, and grayling) and nocturnal and crepuscular species (burbot).
However, larger trout could only have been caught with different traps and adapted baits, or by active fishing with harpoons, bows or arrows, or even by hand, which only targets specific individuals.
6 Conclusion
At Les Cabônes, small fish (around 15 cm) make up the majority of the catch, and provided food throughout the year, particularly during the lean season. Fishing seems to have been optimised to maximise yield-effort ratios. Passive fishing equipment could have captured large numbers of specimens “in shallow areas of rivers, perhaps those backwaters that gradually formed in large valleys in the early Holocene” (Marchand, 2020, p. 341, after Frontin, 2017, p. 274). There seems to have been little or no preparatory cutting. The food intake, which could have reached 258 kg of fish (mainly cyprinids and trout), at least, appears minor compared to food from ungulate hunting. Indeed, a minimum of 50 wild boar, 110 red deer, and 31 roe deer have been estimated from dental remains in layer 3 (Leduc et al., 2015; Trémolières, 2020). As these remains have been collected over the whole site, while fish remains have only been sampled from a selected number of contexts, a direct comparison of meat weight would not be relevant. Nevertheless, fishing in Ranchot could represent a key activity in the economy of the group(s), as it could be practised for most of the year, on a small scale, individually or by part of the group, in most river areas, and unlike hunting, could be combined with other activities.
No fish preparation activities, such as heading or smoking for later consumption, were detected by our analysis. The very small amount of burnt bones found may indicate that the fish were eaten raw, stewed, or boiled, as soon as they were captured. “Immediate” consumption of mostly small catches on the site would explain the presence of abundant vertebrae scattered over the entire studied surface, especially around the hearth.
The fish bone assemblage from Les Cabônes is remarkable, due to both the quantity of remains and the quality of the information it has yielded. Indeed, fishing does not seem to have been practised with the same intensity at all the sites in the Doubs River catchment (Abri Gigot 1, La Roche aux Pêcheurs, Rochedane, cf. Table S1) (Frontin, 2017). However, in the absence of a standardised protocol for the systematic collection of bones and the same volume of sediment study for all sites, this is difficult to demonstrate. But as we noted in the introduction, this is a pattern also observed at sites in the valleys of northern France. This suggests that the archaeological visibility of this activity is to some extent related to the settlement-system and/or site-function.
Interestingly, a fishing pattern remarkably similar to that at Les Cabônes is found at Galgenbühel in the Adige valley (south Tyrol, Italy), during the first two phases of the Sauveterrian occupation of the site, dated from 8600 to 7700 cal. BC: a fish spectrum dominated by cyprinids with a variety of species that live in slack and slow-moving streams; a similar dichotomy in the size of the catches, i.e. an abundance of medium to small sized individuals, and, for one species, larger individuals, all of which implies the use of non-selective passive fishing techniques (Wierer et al., 2018).
Thus, whether in the Adige, Doubs, or Seine valleys, analyses of the largest Mesolithic fish bone assemblages from inland sites provide converging information, namely, the capture of large quantities of small fish, through the use of traps set in areas adjacent to the main channel, and/or highly productive habitats. Moreover, in each of these sites, the exploitation of other wetland resources is well documented: pond turtle (Emys orbicularis) and beaver (Castor fiber), plus otter (Lutra lutra), and waterfowl in Noyen and Galgenbühel (Leduc et al., 2015; Marinval-Vigne et al., 1989; Wierer et al., 2018).
We interpret these results as evidence of Mesolithic occupations focused towards the exploitation of resource-rich wetlands. Such micro-settings offer the possibility of an embedded procurement of terrestrial and aquatic resources within a relatively circumscribed territory. If sites such as Les Cabônes were used repeatedly over many years, this may explain why they are large in size, include large assemblages of lithic and organic remains and display multi-seasonal signatures. It remains to be established whether there is a causal link between increasing dwelling sizes during some periods of the Mesolithic, multi-seasonal occupation, increased dependence on aquatic resources, and decreased group mobility.
The current state of the data does not yet allow us to assess whether the diet of Mesolithic groups from this period in the northern half of France is marked by an increased consumption of aquatic resources. Investigating subsistence strategies requires us to keep working at several scales of resolution and to enrich our database with complementary methods.
Acknowledgments
We wish to thank the professionals and individuals who provided us with valuable data and fish specimens, in particular to the staff of the SD39 of ONEMA (G. Durand, P. Gindre, P. Chanteloube, and S. Forgeois DR2 ONEMA Bourg-Achard). Thanks also to M. El Bettah, in charge of the Jura Federation for Fishing and the Protection of the Aquatic Environment, to P. Falcy of the Joux Valley Sports Fishermen’s Association and to O. Sommen, a fish farmer and member of the Salmon 2000 programme in the Petite Camargue Alsace Nature Reserve. We also thank L. Byrne for the translation. Finally, the manuscript benefited from the comments of two anonymous reviewers, who are thanked.
-
Funding information: This research was supported by regional funding through the project Des derniers Néandertaliens aux premiers agriculteurs dans le massif du Jura et ses marges. Projet Collectif de Recherche (PCR) 2013–2015. (coord. Christophe Cupillard) and by the University of Paris1 Panthéon-Sorbonne (PhD contract of D. Frontin). Financial support for the excavation of the Les Cabônes rockshelter (1978 to 1989) and for AMS radiocarbon dates (2005 to 2008) was provided by the French Ministère de la Culture et de la Communication (Sous-Direction de l’Archéologie) under different research programs.
-
Author contributions: AB and DF designed the experiments and DF carried them out. ABR, AB and DF analysed the data. AB and ABR wrote the manuscript and prepared the figures with contributions from all co-authors. The authors applied the SDC approach for the sequence of authors.
-
Conflict of interest: Authors state no conflict of interest.
-
Data availability statement: All data generated or analysed during this study are included in this published article and its supplementary information files.
References
Asmussen, B. (2009). Intentional or incidental thermal modification? Analysing site occupation via burned bone. Journal of Archaeological Science, 36(2), 528–536.10.1016/j.jas.2008.10.005Search in Google Scholar
Binford, L. R. (1968). Post-Pleistocene adaptations. In S. R. Binford & L. R. Binford (Eds.), New perspectives in archaeology (pp. 313–341). Chicago: Aldine.10.4324/9781315082165-19Search in Google Scholar
Bocherens, H., Polet, C., & Toussaint, M. (2007). Palaeodiet of Mesolithic and Neolithic populations of Meuse Basin (Belgium): Evidence from stable isotopes. Journal of Archaeological Science, 34(1), 10–27. doi: 10.1016/j.jas.2006.03.009.Search in Google Scholar
Broughton, J. M., Cannon, V. I., Arnold, S., Bogiatto, R. J., & Dalton, K. (2006). The taphonomy of owl-deposited fish remains and the origin of the Homestead Cave ichthyofauna. Journal of Taphonomy, 4(2), 69–95.Search in Google Scholar
Bruslé, J., & Quignard, J.-P. (2006). Biologie des poissons d’eau douce européens. Paris: Lavoisier.Search in Google Scholar
Butler, V. L. (1996). Tui Chub Taphonomy and the Importance of Marsh Resources in the Western Great Basin of North America. American Antiquity, 61(4), 699–717. doi: 10.2307/282012.Search in Google Scholar
Butler, V. L., & Chatters, J. C. (1994). The role of bone density in structuring Prehistoric Salmon bone assemblages. Journal of Archaeological Science, 21(3), 413–424. doi: 10.1006/jasc.1994.1039.Search in Google Scholar
Butler, V. L., & Schroeder, R. A. (1998). Do digestive processes leave diagnostic traces on fish bones? Journal of Archaeological Science, 25(10), 957–971.10.1006/jasc.1997.0725Search in Google Scholar
Campy, M., David, S., & Cupillard, C. (1989). « Ranchot – Abri des Cabônes » [notice archéologique]. ADLFI. Archéologie de la France – Informations, Bourgogne-Franche-Comté 27073.Search in Google Scholar
Cravinho, S. (2009). Les poissons d’eau douce à la fin du Paléolithique supérieur en France: Réexamen et étude complémentaire du site de Pont d’Ambon (Bourdeilles, Dordogne). (PhD Thesis). Aix-Marseille: Université de Provence.Search in Google Scholar
Cravinho, S. (2011). La taphonomie des poissons paléolithiques. Présentation des outils d’analyse appliqués au site azilien de Pont d’Ambon (Bourdeilles, Dordogne). In C. Denys, V. Laroulandie, J.-B. Mallye (Eds.), Taphonomie des petits vertébrés: Référentiels et transferts aux fossiles (pp. 141–152) Oxford: BAR Publishing.Search in Google Scholar
Cravinho, S., & Desse-Berset, N. (2005). Les poissons du Bois-Ragot (Gouex, Vienne). Mémoires de la Société Préhistorique Française, 38, 355–371.Search in Google Scholar
Crotti, P., & Cupillard, C. (2013). Le Mésolithique dans le Jura franco-suisse. In A. Richard, F. Schifferdecker, J.-P. Mazimann, & C. Bélet-Gonda (Eds.), Le peuplement de l’Arc jurassien de la Préhistoire au Moyen-Age (pp. 127–142). Besançon: Presses Universitaires de Franche-Comté et Porrentruy, Office de la Culture et Société Jurassienne D’émulation.Search in Google Scholar
Cupillard, C. (1998a). La pêche durant l’Epipaléolithique et le Mésolithique dans le Jura. In C. Cupillard & A. Richard (Eds.), Les derniers chasseurs-cueilleurs du massif jurassien et de ses marges (13000-5500 avant Jésus-Christ) (pp. 84–87). Lons-le-Saunier: Centre Jurassien du Patrimoine.Search in Google Scholar
Cupillard, C. (1998b). L’abri des Cabônes à Ranchot (Jura). In C. Cupillard & A. Richard (Eds.), Les derniers chasseurs-cueilleurs du massif jurassien et de ses marges (13000-5500 avant Jésus-Christ) (pp. 112–113). Lons-le-Saunier: Centre Jurassien du Patrimoine.Search in Google Scholar
Cupillard, C. (1998c). L’industrie en matière dure animale. In C. Cupillard & A. Richard (Eds.), Les derniers chasseurs-cueilleurs du massif jurassien et de ses marges (13000-5500 avant Jesus-Christ) (pp. 158–161). Lons-le-Saunier: Centre Jurassien du Patrimoine.Search in Google Scholar
Cupillard, C. (2003). Les derniers Chasseurs-Cueilleurs de l’Est de la France: L’exemple de la Franche-Comté entre 10000 et 6000 BP. In L. Bevan & J. Moore (Eds.), Peopling the Mesolithic in a Northern environment (pp. 129–158). Oxford: BAR Publishing.Search in Google Scholar
Cupillard, C., Auguste, P., Cupillard-Perrenoud, N., David, S., & Morel, P. (1988). Abri des Cabônes, Ranchot (Jura) (non paginé) [Rapport de fouille]. Service régional de l’Archéologie de Franche-Comté.Search in Google Scholar
Cupillard, C., & David, S. (1995). La prédation au Magdalénien final et au Mésolithique. In A. Richard & C. Munier (Eds.), Eclats d’Histoire, 25000 ans d’héritages, 10 ans d’archéologie en Franche-Comté (pp. 104–109). Besançon: Cêtre.Search in Google Scholar
Cupillard, C., Magny, M., Bocherens, H., Bridault, A., Bégeot, C., Bichet, V., … Walter-Simonnet, A.-V. (2015). Changes in ecosystems, climate and societies in the Jura Mountains between 40 and 8 ka cal BP. Quaternary International, 378, 40–72. doi: 10.1016/j.quaint.2014.05.032.Search in Google Scholar
Dauphin, C. (1989). L’ichtyofaune de Noyen-sur-Seine. In L’homme et l’eau au temps de la préhistoire. Actes du 112ème Congrès National des Sociétés Savantes, Lyon, 1987, Commission de Pré et Protohistoire. (pp. 11–31). Paris: CTHS.Search in Google Scholar
David, S. (1993). Magdalénien et Azilien en Franche-Comté: Contribution à l’étude des cultures du Tardiglaciaire dans l’Est de la France (Thèse d’Etat). Besançon: Université de Franche-Comté.Search in Google Scholar
Deseine, A., Guéret, C., Vigne, J.-D., Mordant, D., & Valentin, B. (2019). Nouveau regard sur les occupations du second Mésolithique du “ Haut des Nachères” à Noyen-sur-Seine (Seine et Marne). In R.-M. Arbogast, S. Griselin, … F. Séara (Eds.), Le second Mésolithique des Alpes à l’Atlantique (7e–5e millénaire), Mémoires d’archéologie du Grand-Est (Vol. 3, pp. 189–234). https://hal-mnhn.archives-ouvertes.fr/mnhn-02102894.Search in Google Scholar
Desse, G., & Desse, J. (1976). Diagnostic des pièces rachidiennes des téléostéens et des Chondrichthyens. III Téléostéens d’eau douce. Paris: L’Expansion Scientifique.Search in Google Scholar
Desse, G., & Desse, J. (1981). Diagnostic des pièces rachidiennes de poissons: Applications au matériel issu de sites archéologiques. Centre de Recherches Archéologiques Notes Internes, 28.Search in Google Scholar
Desse, G., & Desse, J. (1983). L’identification des vertèbres de poissons; application au matériel issu des sites archéologiques et paléontologiques. Archives des Sciences, 36(2), 291–296.Search in Google Scholar
Desse, J., & Desse-Berset, N. (1992). Age et saison de mort des poissons: Applications à l’archéologie. In J.-L. Baglinière, J. Castanet, F. Conand, & F.J. Meunier (Eds.), Tissus durs et age individuel des vertébrés (pp. 341–353). Paris: ORSTOM.Search in Google Scholar
Desse, J., & Desse-Berset, N. (2002). Le cortège de Neptune: Les poissons de la Méditerranée durant l’Holocène. In A. Gardeisen (Ed.), Mouvements ou déplacements de populations animales en Méditerranée au cours de l’Holocène, séminaire, 29 septembre 2000 (pp. 83–96). Oxford: BAR Publishing.Search in Google Scholar
Desse, J., Desse-Berset, N., & Rocheteau, M. (1987). Contribution à l’ostéométrie de la perche (Perca fluviatilis Linné, 1758). Juan-les-Pins: APDCA.Search in Google Scholar
Desse, J., Desse-Berset, N., & Rocheteau, M. (1990). L’ostéométrie de la lote d’eau douce, Lola lota (Linné, 1758). Juan-les-Pins: APDCA.Search in Google Scholar
Desse-Berset, N. (1994). Les poissons. In Les grottes Jean-Pierre I et II à Saint-Thibaud-de-Couz (Savoie) (pp. 218–224), Gallia Préhistoire, 36. doi: 10.3406/galip.1994.2122.Search in Google Scholar
Drucker, D., & Bocherens, H. (2004). Carbon and nitrogen stable isotopes as tracers of change in diet breadth during Middle and Upper Palaeolithic in Europe. International Journal of Osteoarchaeology, 14(3–4), 162–177. doi: 10.1002/oa.753.Search in Google Scholar
Drucker, D., Bridault, A., Ducrocq, T., Baumann, C., & Valentin, F. (2020). Environment and human subsistence in Northern France at the Late Glacial to early Holocene transition. Archaeological and Anthropological Sciences, 12(8), 1–16. doi: 10.1007/s12520-020-01149-4.Search in Google Scholar
Drucker, D., Bridault, A., Hobson, K. A., Szuma, E., & Bocherens, H. (2008). Can carbon-13 in large herbivores reflect the canopy effect in temperate and boreal ecosystems? Evidence from modern and ancient ungulates. Palaeogeography, Palaeoclimatology, Palaeoecology, 266(1–2), 69–82. doi: 10.1016/j.palaeo.2008.03.020.Search in Google Scholar
Drucker, D., Valentin, F., Thevenet, C., Mordant, D., Cottiaux, R., Delsate, D., & Van Neer, W. (2016). Aquatic resources in human diet in the Late Mesolithic in Northern France and Luxembourg: Insights from carbon, nitrogen and sulphur isotope ratios. Archaeological and Anthropological Sciences, 10(2), 351–368. doi: 10.1007/s12520-016-0356-6.Search in Google Scholar
Ducrocq, T., Leduc, C., Guéret, C., Muriel, B., Chombart, J., François, S., … Coutard, S. (2019). Premières données sur le site mésolithique du « Collège Belrem » à Beaurainville (Pas-de-Calais). In R.-M. Arbogast, S. Griselin, C. Jeunesse, & F. Séara (Eds.), Le second Mésolithique des Alpes à l’Atlantique (7e–5e millénaire) (Vol. 3, pp. 153–178). Strasbourg: Mémoires d’Archéologie du Grand-Est.Search in Google Scholar
Fleury, R., Farjanel, G., & Collin, J. (1982). Notice explicative, Carte géologique de la France (1/50 000), feuille Seurre (527). BRGM.Search in Google Scholar
Froese, R. (2021). Anguilla anguilla (Linnaeus, 1758), European eel: Fisheries, aquaculture, gamefish. FishBase. World Wide Web Electronic Publication. https://fishbase.mnhn.fr/Summary/SpeciesSummary.php?ID=35&AT=eel.Search in Google Scholar
Froese, R., & Pauly, D. (2021). FishBase. FishBase. World Wide Web Electronic Publication. www.fishbase.org.Search in Google Scholar
Frontin, D. (2017). Economie de pêche au Mésolithique et diversité piscicole à l’Holocène ancien dans le bassin hydrographique du Doubs. (PhD Thesis). Paris: Université de Paris 1 Panthéon-Sorbonne.Search in Google Scholar
Fu, Q., Posth, C., Hajdinjak, M., Petr, M., Mallick, S., Fernandes, D., … Mittnik, A. (2016). The genetic history of ice age Europe. Nature, 534(7606), 200–205.10.1038/nature17993Search in Google Scholar
Guillaud, E., Béarez, P., Denys, C., & Raimond, S. (2014). Taphonomy of a fish accumulation by the European Otter (Lutra lutra) in central France. Journal of Taphonomy, 12(1), 69–83.Search in Google Scholar
Guillaud, E., Bearez, P., Denys, C., & Raimond, S. (2017). New data on fish diet and bone digestion of the Eurasian otter (Lutra lutra) (Mammalia: Mustelidae) in central France. The European Zoological Journal, 84(1), 226–237. doi: 10.1080/24750263.2017.1315184.Search in Google Scholar
Guillaud, E., Cornette, R., & Béarez, P. (2016). Is vertebral form a valid species-specific indicator for salmonids? The discrimination rate of trout and Atlantic salmon from archaeological to modern times. Journal of Archaeological Science, 65, 84–92. doi: 10.1016/j.jas.2015.11.010.Search in Google Scholar
Guillaud, E., Lebreton, L., & Béarez, P. (2018). Taphonomic signature of Eurasian eagle owl (Bubo bubo) on fish remains. Folia Zoologica, 67(3–4), 143. doi: 10.25225/fozo.v67.i3-4.a1.2018.Search in Google Scholar
Hajkova, P., Roche, K., & Kocian, L. (2003). On the use of diagnostic bones of brown trout, Salmo trutta m. fario, grayling, Thymallus thymallus and Carpathian sculpin, Cottus poecilopus in Eurasian otter, Lutra lutra diet analysis. Folia Zoologica, 52(4), 389–398.Search in Google Scholar
Hallet, C. (1985). Modulations de la stratégie alimentaire d’un prédateur: Éco-éthologie de la prédation chez le martin-pêcheur, Alcedo atthis (L., 1758) en période de reproduction. Cahiers d’Ethologie Appliquée, 5(4), 1–206.Search in Google Scholar
Huber, H. R., Jorgensen, J. C., Butler, V. L., Baker, G., & Stevens, R. (2011). Can salmonids (Oncorhynchus spp.) be identified to species using vertebral morphometrics? Journal of Archaeological Science, 38(1), 136–146. doi: 10.1016/j.jas.2010.08.020.Search in Google Scholar
Huet, M. (1949). Aperçu des relations entre la pente et les populations piscicoles des eaux courantes. Schweizerische Zeitschriftt für Hydrologie, 11, 332–351.10.1007/BF02503356Search in Google Scholar
Jones, A. K. (1986). Fish bone survival in the digestive systems of the pig, dog and man: Some experiments. In D. C. Brinkhuizen, & A. T. Clason (Eds.), Fish and archaeology: Studies in osteometry, taphonomy, seasonality and fishing methods (53–61). Oxford: BAR Publishing.Search in Google Scholar
Jones, A. K. (1984). Some effects of the mammalian digestive system on fish bones. 2nd Fish Osteoarchaeology Meeting (pp. 61–65).Search in Google Scholar
Keith, P., & Allardi, J. (2001). Atlas des poissons d’eau douce de France. Paris: Muséum National d’Histoire Naturelle.Search in Google Scholar
Kettle, A. J., Morales-Muñiz, A., Roselló-Izquierdo, E., Heinrich, D., & Vøllestad, L. A. (2011). Refugia of marine fish in the northeast Atlantic during the last glacial maximum: Concordant assessment from archaeozoology and palaeotemperature reconstructions. Climate of the Past, 7(1), 181–201. doi: 10.5194/cp-7-181-2011.Search in Google Scholar
Koivisto, S. (2017). Archaeology of Finnish wetlands: with special reference to studies of Stone Age stationary wooden fishing structures. (PhD Thesis). Helsinki: University of Helsinki.Search in Google Scholar
Laroulandie, V. (2000). Taphonomie et archéozoologie des Oiseaux en grotte: Applications aux sites paléolithiques du Bois-Ragot (Vienne), de Combe Saunière (Dordogne) et de La Vache (Ariège). (PhD Thesis). Bordeaux: Université Sciences et Technologies-Bordeaux I.Search in Google Scholar
Le Gall, O. (1982). Les poissons de quelques gisements prehistoriques du Sud-Ouest de la France, Etude ostéologique. Intérêt paléoécologique et palethnologique. (Thèse de 3ème cycle). Géologie, Bordeaux.Search in Google Scholar
Le Gall, O. (1984). L’Ichtyofaune d’eau douce dans les sites préhistoriques. Ostéologie, paléoécologie, palethnologie. Cahiers du Quaternaire, 8, 196.Search in Google Scholar
Le Gall, O. (1991). Pêcheurs mésolithiques et néolithiques: Similitudes ou différences de comportement? In Coll (Ed.), Mésolithisation et Néolithisation en France et dans les régions limitrophes (Vol. 113, pp. 99–104). Paris: CTHS.Search in Google Scholar
Le Gall, O. (1992). Poissons et pêches au Paléolithique: Quelques données de l’Europe occidentale. L’Anthropologie, 96(1), 121–134.Search in Google Scholar
Le Gall, O. (1993). Evolution des pêches de l’Epipaléolithique au Néolithique ancien. Préhistoire Anthropologie Méditerranéennes, 2, 135–142.Search in Google Scholar
Le Gall, O. (1999). Éléments de réflexion sur la pêche dans le bassin méditerranéen nord-occidental pendant le développement des faciès leptolithiques. In Les faciès leptolithiques du nord-ouest méditerranéen: Milieux naturels et culturels, XXIVe Congrès Préhistorique de France – Carcassonne 26–30 Septembre 1994 (pp. 251–265). Société Préhistorique Française.Search in Google Scholar
Le Gall, O. (2001). Les représentations de poissons dans l’art mobilier magdalénien: Une expression de l’importance culturelle de la pêche. Préhistoire du Sud-Ouest, 8, 55–69.Search in Google Scholar
Le Gall, O. (2003). La Squelettochronologie appliquée aux poissons: Une méthode de reconnaissance des saisons de capture. Préhistoire du Sud-Ouest, 10, 9–24.Search in Google Scholar
Le Gall, O., & Pannoux, P. (1994). Annexes II. Les poissons: L’abri sous roche de Pont d’Ambon à Bourdeilles (Dordogne). Gallia Préhistoire, 36(1), 113–126. doi: 10.3406/galip.1994.2102 Search in Google Scholar
Le Gall, O. (2010). Influences des glaciaires-interglaciaires sur les ichtyofaunes des eaux douces européennes. Quaternaire, 21/3, 203–214. doi: 10.4000/quaternaire.5570.Search in Google Scholar
Leduc, C., Bridault, A., & Cupillard, C. (2015). Wild boar (Sus scrofa scrofa) hunting and exploitation strategies during the Mesolithic at Les Cabônes (Ranchot Jura, France), layer 3. Journal of Archaeological Science: Reports, 2, 473–484. doi: 10.1016/j.jasrep.2015.05.010.Search in Google Scholar
Libois, R., Hallet, C., & Rosoux, R. (1987). Éléments pour l’identification des restes crâniens des poissons dulçaquicoles de Belgique et du Nord de la France. 1. Anguilliformes, Gastérostéiformes, Cyprinodontiformes et Perciformes. Fiches d’Ostéologie Animale pour l’Archéologie. Série A, Poissons, 3, 1–15. Juan-les-Pins: ADPCA.Search in Google Scholar
Libois, R., & Hallet-Libois, C. (1988). Eléments pour l’identification des restes crâniens des poissons dulçaquicoles de Belgique et du nord de la France: 2. Cypriniformes. Juan-les-Pins: APDCA.Search in Google Scholar
Libois, R., & Hallet-Libois, C. (1995). Martin-pêcheur d’Europe, Alcedo atthis (L. 1758). In D. Yeatman-Berthelot & G. Jarry (Eds.), Nouvel atlas des oiseaux nicheurs de France, 1985–1989 (pp. 418–421). Société Ornithologique de France.Search in Google Scholar
Llona, A. C. P., & Andrews, P. J. (1999). Amphibian taphonomy and its application to the fossil record of Dolina (middle Pleistocene, Atapuerca, Spain). Palaeogeography, Palaeoclimatology, Palaeoecology, 149(1–4), 411–429.10.1016/S0031-0182(98)00215-6Search in Google Scholar
Luna, S. (2021a). Lota lota (Linnaeus, 1758), Burbot: Fisheries, aquaculture, gamefish, aquarium. FishBase. World Wide Web Electronic Publication. https://fishbase.mnhn.fr/Summary/SpeciesSummary.php?ID=310&AT=burbotSearch in Google Scholar
Luna, S. (2021b). Rutilus rutilus (Linnaeus, 1758), Roach: Fisheries, aquaculture, gamefish. FishBase. World Wide Web Electronic Publication. https://fishbase.mnhn.fr/Summary/SpeciesSummary.php?ID=272&AT=roachSearch in Google Scholar
Luna, S. (2021c). Thymallus thymallus (Linnaeus, 1758), Grayling: Fisheries, aquaculture, gamefish, aquarium. FishBase. World Wide Web Electronic Publication. https://fishbase.mnhn.fr/Summary/SpeciesSummary.php?ID=4605&AT=graylingSearch in Google Scholar
Machino, Y. (2003). L’Omble chevalier: Salvelinus alpinus (Linné, 1758). In M. Pascal, O. Lorvelec, J.-D. Vigne, P. Keith, & P. Clergeau (Eds.), Évolution holocène de la faune de Vertébrés de France: Invasions et disparitions. Paris: Institut National de la Recherche Agronomique, Centre National de la Recherche Scientifique, Muséum National d’Histoire Naturelle, Ministère de l’Aménagement du Territoire et de l’Environnement.Search in Google Scholar
Macklin, M. G., & Lewin, J. (2003). River sediments, great floods and centennial-scale Holocene climate change. Journal of Quaternary Science: Published for the Quaternary Research Association, 18(2), 101–105.10.1002/jqs.751Search in Google Scholar
McQuade, M., & O’Donnell, L. (2007). Late Mesolithic fish traps from the Liffey estuary,Dublin, Ireland. Antiquity, 81(313), 569–584.10.1017/S0003598X00095594Search in Google Scholar
Magny, M., & Bravard, J.-P. (2002). Variations paléohydrologiques en France depuis 15000 ans. Une introduction. In J.-P. Bravard & M. Magny (Eds.), Les fleuves ont une histoire. Paléo-environnement des rivières et des lacs français depuis 15 000 ans (pp. 9–14). Paris: Errance.Search in Google Scholar
Marchand, G. (2020). Degrés d’hydrosolubilité du Mésolithique en France. Bulletin de la Société Archéologique Champenoise, 113(2/3), 333–357.Search in Google Scholar
Marinval-Vigne, M.-C., Mordant, D., Auboire, G., Augereau, A., Bailon, S., Dauphin, C., … Vigne, J.-D. (1989). Noyen-sur-Seine, site stratifié en milieu fluviatile: Une étude multidisciplinaire intégrée. Bulletin de la Société Préhistorique Française, 86(10/12), 370–379.10.3406/bspf.1989.9894Search in Google Scholar
Marquebielle, B. (2014). Le travail des matières osseuses au Mésolithique. Caractérisation technique et économique à partir des séries du sud et de l’est de la France. (PhD Thesis). Toulouse: Université de Toulouse 2 le Mirail.Search in Google Scholar
Meunier, F. J. (1988). Détermination de l’âge individuel chez les Ostéichthyens à l’aide de la squelettochronologie: Historique et méthodologie. Acta Oecologica. Oecologia Generalis, 9(3), 299–329.Search in Google Scholar
Morales, A., & Rosenlund, K. D. (1979). Fish bone measurements. An attempt to standardize the Measuring of Fish Bones from Archaeological sites. Copenhagen: Steenstrupia.Search in Google Scholar
Mordant, D., Valentin, B., & Vigne, J.-D. (2013). Noyen-sur-Seine, vingt cinq ans après. In B. Valentin, B. Souffi, T. Ducrocq, J.-P. Fagnart, F. Séara, & C. Verjux (Eds.), Palethnographie du Mésolithique Recherches sur les habitats de plein air entre Loire et Neckar (pp. 37–49). Paris: Société Préhistorique Française.Search in Google Scholar
Moss, M. L., Judd, K. G., & Kemp, B. M. (2014). Can salmonids (Oncorhynchus spp.) be identified to species using vertebral morphometrics? A test using ancient DNA from Coffman Cove, Alaska. Journal of Archaeological Science, 41, 879–889. doi: 10.1016/j.jas.2013.10.017.Search in Google Scholar
Nicholson, R. A. (1993). An investigation into the effects on fish bone of passage through the human gut: Some experiments and comparisons with archaeological material. Circaea, 10(1), 38–51.Search in Google Scholar
Nicholson, R. A. (2000). Otter (Lutra lutra L.) spraint: An investigation into possible sources of small fish bones at coastal archaeological sites. Taphonomy and Interpretation, 120, 55.Search in Google Scholar
Niekus, M. J. L. T., Brinkhuizen, D. C., Kerkhoven, A. A., Huisman, J. J., & Velthuizen, D. E. (2012). An Early Atlantic Mesolithic site with micro-triangles and fish remains from Almere (the Netherlands). In D. C. M. Raemaekers, E. Esser, R. C. G. M. Lauwerier, & J. T. Zeile (Eds.), A bouquet of archaeozoological studies, essays in Honour of Wietske Prummel (pp. 61–76), Groningen: Barkhuis.10.2307/j.ctt227285m.8Search in Google Scholar
Pastre, J. F., Leroyer, C., Limondin-Lozouet, N., Chaussé, C., Fontugne, M., Gebhardt, A., … Krier, V. (2000). Le Tardiglaciaire des fonds de vallée du Bassin Parisien (France) [The Late-Glacial from the Paris basin floodplains (France)]. Quaternaire, 11(2), 107–122. doi: 10.3406/quate.2000.1660.Search in Google Scholar
Persat, H. (2003). Recolonisation postglaciaire de la façade externe de l’arc alpin par les poissons d’eau douce: Opportunités et réalités. Cymbium, 27(1), 27–64.Search in Google Scholar
Persat, H., & Keith, P. (1997). La répartition géographique des poissons d’eau douce en France: Qui est autochtone et qui ne l’est pas? Bulletin Français de la Pêche et de la Pisciculture, 344–345, 15–32.10.1051/kmae:1997007Search in Google Scholar
Persat, H., & Keith, P. (2002). La Reconquête du réseau hydrographique français par le poissons d’eau douce à l’Holocène: À la nage ou à pied? In J.-P. Bravard & M. Magny (Eds.), Les fleuves ont une histoire: Paléo-environnement des rivières et des lacs français depuis 15 000 ans (pp. 295–301). Paris: Errance.Search in Google Scholar
Picquès, G. (1998). Contribution à la connaissance de l’anguille (Anguilla anguilla, Linné, 1758) des sites archéologiques (Mémoire de DEA Archéologie et environnement). Paris: Université Paris 1 Panthéon-Sorbonne.Search in Google Scholar
Posth, C., Renaud, G., Mittnik, A., Drucker, D., Rougier, H., Cupillard, C., … Krause, J. (2016). Pleistocene mitochondrial genomes suggest a single major dispersal of non-Africans and a Late glacial population turnover in Europe. Current Biology, 26(6), 827–833. doi: 10.1016/j.cub.2016.01.037.Search in Google Scholar
Robinson, B. S., Jacobson, G. L., Yates, M. G., Spiess, A. E., & Cowie, E. R. (2009). Atlantic salmon, archaeology and climate change in New England. Journal of Archaeological Science, 36(10), 2184–2191. doi: 10.1016/j.jas.2009.06.001.Search in Google Scholar
Robson, H. K., Andersen, S. H., Clarke, L., Craig, O. E., Gron, K. J., Jones, A. K., … Ritchie, K. (2016). Carbon and nitrogen stable isotope values in freshwater, brackish and marine fish bone collagen from Mesolithic and Neolithic sites in central and northern Europe. Environmental Archaeology, 21(2), 105–118.10.1179/1749631415Y.0000000014Search in Google Scholar
Rochefort, G., & Corolla, J-P. (2019). Salmo trutta fario (Linnaeus, 1758)–DORIS. https://doris.ffessm.fr/Especes/Salmo-trutta-fario-Truite-de-riviere-388.Search in Google Scholar
Rotillon, S. (2000). L’ajustement des systèmes fluviaux de la fin du Tardiglaciaire au début de l’Holocène en tant qu’indicateur des changements environnementaux: La Seille à l’aval d’Arlay (Jura, France). In A. Richard, C. Cupillard, H. Richard, & A. Thévenin (Eds.), Les derniers chasseurs-cueilleurs d’Europe occidentale (13000-5500 av. J.-C.) (pp. 83–92). Besançon: Presses Universitaires de Franche-Comté.Search in Google Scholar
Roué, S. (2000). Les armatures mésolithiques de l’abri des Cabônes à Ranchot (Jura). In A. Richard, C. Cupillard, H. Richard, & A. Thévenin (Eds.), Les derniers chasseurs—Cueilleurs d’Europe occidentale (13000–5500 av. J.-C.) (pp. 133–141). Besançon: Presse Universitaires de Franche-Comté.Search in Google Scholar
Rozoy, J.-G. (1978). Les derniers chasseurs: L’Epipaléolithique en France et en Belgique: Essai de synthèse (Vol. n° spécial juin 1978). Bulletin de la Société Archéologique Champenoise.Search in Google Scholar
Russ, H. (2010). The Eurasian eagle owl (Bubo bubo): A fish bone accumulator on Pleistocene cave sites. Journal of Taphonomy, 8(4), 281–290.Search in Google Scholar
Russ, H., & Jones, A. K. (2011). Fish remains in cave deposits; how did they get there. Cave and Karst Science, 38(3), 117–120.Search in Google Scholar
Schulting, R. J., Blockley, S. M., Bocherens, H., Drucker, D., & Richards, M. (2008). Stable carbon and nitrogen isotope analysis on human remains from the Early Mesolithic site of La Vergne (Charente-Maritime, France). Journal of Archaeological Science, 35(3), 763–772. doi: 10.1016/j.jas.2007.06.008.Search in Google Scholar
Séara, F., Rotillon, S., & Cupillard, C. (2002). Campements mésolithiques en Bresse jurassienne: Choisey et Ruffey-sur-Seille. Paris: Editions de la Maison des Sciences de l’Homme.10.4000/books.editionsmsh.21353Search in Google Scholar
Sternberg, M. (1989). La consommation du poisson à Lattes (IIIe-Ier s. Av. N. È.). Méthodes d’étude et premiers résultats in Introduction à l’étude de l’environnement de Lattes antique. Lattara, 2, 101–120.Search in Google Scholar
Stuart, A. J., & Lister, A. M. (2007). Patterns of Late Quaternary megafaunal extinctions in Europe and northern Asia. Courier-Forschungsinstitut Senckenberg, 259, 287.Search in Google Scholar
Thieren, E., Wouters, W., Neer, W. V., & Ervynck, A. (2012). Body length estimation of the European eel Anguilla anguilla on the basis of isolated skeletal elements. Cybium, 36(4), 551–562.Search in Google Scholar
Trémolières, M. (2020). La chasse du cerf (Cervus elaphus) et du chevreuil (Capreolus capreolus) à l’abri des Cabônes à Ranchot (Jura). Contribution à l’étude de l’exploitation des grands gibiers au Mésolithique [Mémoire de Master 2, Anthropologie spécialité Préhistoire). Nanterre: Université de Paris Nanterre.Search in Google Scholar
University of Nottingham. (2021). Archaeological fish resource. Archaeological Fish Resource. http://fishbone.nottingham.ac.uk/.Search in Google Scholar
Verneaux, J. (1973). Recherches écologiques sur le réseau hydrographique du Doubs: Essai de biotypologie. Annales Scientifiques de L’université de Besançon 3e Série (Zoologie, Physiologie et Biologie Animale), Fascicule 9.Search in Google Scholar
Wheeler, A., & Jones, A. (1989). Fishes. Cambridge: Cambridge University Press.Search in Google Scholar
Wierer, U., Betti, L., Boscato, P., Bazzanella, M., Boschin, F., Crezzini, J., … Tagliacozzo, A. (2018). Vivre près de l’eau. Environnement, économie du milieu humide y techniques de pêche du site mésolithique Galgenbühel/Dos de la Forca dans la vallée de l’Adige (Tyrol du Sud, Italie). In C. Cupillard, F. Séara, S. Griselin (Dir.) Au cœur des sites mésolithiques: Entre processus taphonomiques et données archéologiques, Actes de la table-ronde internationale de Besançon (Doubs, France) « Hommages au Professeur André Thévenin » 29–30 octobre 2013 (pp. 241–257). Besançon: Presses Universitaires de Franche-Comté.Search in Google Scholar
Wijngaarden-Bakker, L. H. (1989) Faunal Remains and the Irish Mesolithic. In C. Bonsall (Ed.) The Mesolithic in Europe, Papers presented at the Third International Symposium, (Edinburgh) (pp. 125–133). Edinburgh: John Donald.Search in Google Scholar
Willis, L. M., & Boehm, A. R. (2014). Fish bones, cut marks, and burial: Implications for taphonomy and faunal analysis. Journal of Archaeological Science, 45, 20–25.10.1016/j.jas.2014.01.026Search in Google Scholar
Willis, L. M., Eren, M. I., & Rick, T. C. (2008). Does butchering fish leave cut marks? Journal of Archaeological Science, 35(5), 1438–1444.10.1016/j.jas.2007.10.010Search in Google Scholar
Yang, D. Y., Cannon, A., & Saunders, S. R. (2004). DNA species identification of archaeological salmon bone from the Pacific Northwest Coast of North America. Journal of Archaeological Science, 31(5), 619–631. doi: 10.1016/j.jas.2003.10.008 Search in Google Scholar
Zohar, I., Ovadia, A., & Goren-Inbar, N. (2016). The cooked and the raw: A taphonomic study of cooked and burned fish. Journal of Archaeological Science: Reports, 8, 164–172. doi: 10.1016/j.jasrep.2016.06.005.Search in Google Scholar
Zohar, I., & Biton, R. (2011). Land, lake, and fish: Investigation of fish remains from Gesher Benot Ya’aqov (paleo-Lake Hula). Journal of Human Evolution, 60(4), 343–356. doi: 10.1016/j.jhevol.2010.10.007.Search in Google Scholar
© 2022 Anne Bridault et al., published by De Gruyter
This work is licensed under the Creative Commons Attribution 4.0 International License.
Articles in the same Issue
- Editorial
- Editorial: Open Archaeology in Challenging Times
- Regular Articles
- Caves, Senses, and Ritual Flows in the Iberian Iron Age: The Territory of Edeta
- Tutankhamun’s Polychrome Wooden Shawabtis: Preliminary Investigation for Pigments and Gilding Characterization and Indirect Dating of Previous Restorations by the Combined Use of Imaging and Spectroscopic Techniques
- When TikTok Discovered the Human Remains Trade: A Case Study
- Nuraghi as Ritual Monuments in the Sardinian Bronze and Iron Ages (circa 1700–700 BC)
- A Pilot Study in Archaeological Metal Detector Geophysical Survey
- A Blocked-Out Capital from Berenike (Egyptian Red Sea Coast)
- The Winery in Context: The Workshop Complex at Ambarçay, Diyarbakır (SE Turkey)
- Tracing Maize History in Northern Iroquoia Through Radiocarbon Date Summed Probability Distributions
- Faunal Remains Associated with Human Cremations: The Chalcolithic Pits 16 and 40 from the Perdigões Ditched Enclosures (Reguengos de Monsaraz, Portugal)
- A Multi-Method Study of a Chalcolithic Kiln in the Bora Plain (Iraqi Kurdistan): The Evidence From Excavation, Micromorphological and Pyrotechnological Analyses
- Potters’ Mobility Contributed to the Emergence of the Bell Beaker Phenomenon in Third Millennium BCE Alpine Switzerland: A Diachronic Technology Study of Domestic and Funerary Traditions
- From Foragers to Fisher-Farmers: How the Neolithisation Process Affected Coastal Fisheries in Scandinavia
- Enigmatic Bones: A Few Archaeological, Bioanthropological, and Historical Considerations Regarding an Atypical Deposit of Skeletonized Human Remains Unearthed in Khirbat al-Dusaq (Southern Jordan)
- Who Was Buried at the Petit-Chasseur Site? The Contribution of Archaeometric Analyses of Final Neolithic and Bell Beaker Domestic Pottery to the Understanding of the Megalith-Erecting Society of the Upper Rhône Valley (Switzerland, 3300–2200 BC)
- Erratum
- Erratum to “Britain In or Out of Europe During the Late Mesolithic? A New Perspective of the Mesolithic–Neolithic Transition”
- Review Article
- Archaeological Practices and Societal Challenges
- Special Issue Published in Cooperation with Meso’2020 – Tenth International Conference on the Mesolithic in Europe, edited by Thomas Perrin, Benjamin Marquebielle, Sylvie Philibert, and Nicolas Valdeyron - Part I
- Animal Teeth and Mesolithic Society
- A Matter of Scale: Responses to Landscape Changes in the Oslo Fjord, Norway, in the Mesolithic
- Chipped Stone Assemblage of the Layer B of the Kamyana Mohyla 1 Site (South-Eastern Ukraine) and the Issue of Kukrek in the North Meotic Steppe Region
- Rediscovered Mesolithic Rock Art Collection from Kamyana Mohyla Complex in Eastern Ukraine
- Mesolithic Montology
- A Little Mystery, Mythology, and Romance: How the “Pigmy Flint” Got Its Name
- Preliminary Results and Research Perspectives on the Submerged Stone Age Sites in Storstrømmen, Denmark
- Techniques and Ideas. Zigzag Motif, Barbed Line, and Shaded Band in the Meso-Neolithic Bone Assemblage at Zamostje 2, Volga-Oka Region (Russia)
- Modelling Foraging Cultures According to Nature? An Old and Unfortunately Forgotten Anthropological Discussion
- Mesolithic and Chalcolithic Mandibular Morphology: Using Geometric Morphometrics to Reconstruct Incomplete Specimens and Analyse Morphology
- Britain In or Out of Europe During the Late Mesolithic? A New Perspective of the Mesolithic–Neolithic Transition
- Non-Spatial Data and Modelling Multiscale Systems in Archaeology
- Living in the Mountains. Late Mesolithic/Early Neolithic Settlement in Northwest Portugal: Rock Shelter 1 of Vale de Cerdeira (Vieira do Minho)
- Enculturating Coastal Environments in the Middle Mesolithic (8300–6300 cal BCE) – Site Variability, Human–Environment Relations, and Mobility Patterns in Northern Vestfold, SE-Norway
- Why Mesolithic Populations Started Eating Crabs on the European Atlantic Façade Only Over the Past 15 Years?
- “Ain’t No Mountain High Enough” – Mesolithic Colonisation Processes and Landscape Usage of the Inner-Alpine Region Kleinwalsertal (Prov. Vorarlberg, Western Austria)
- Mesolithic Freshwater Fishing: A Zooarchaeological Case Study
- Consumers, not Contributors? The Study of the Mesolithic and the Study of Hunter-Gatherers
- Fish Processing in the Iron Gates Region During the Transitional and Early Neolithic Period: An Integrated Approach
- Hunting for Hide. Investigating an Other-Than-Food Relationship Between Stone Age Hunters and Wild Animals in Northern Europe
- Changing the Perspective, Adapting the Scale: Macro- and Microlithic Technologies of the Early Mesolithic in the SW Iberian Peninsula
- Fallen and Lost into the Abyss? A Mesolithic Human Skull from Sima Hedionda IV (Casares, Málaga, Iberian Peninsula)
- Evolutionary Dynamics of Armatures in Southern France in the Late Mesolithic and Early Neolithic
- Combining Agent-Based Modelling and Geographical Information Systems to Create a New Approach for Modelling Movement Dynamics: A Case Study of Mesolithic Orkney
- Pioneer Archaeologists and the Influence of Their Scientific Relationships on Mesolithic Studies in North Iberia
- Neolithisation in the Northern French Alps: First Results of the Lithic Study of the Industries of La Grande Rivoire Rockshelter (Isère, France)
- Late Mesolithic Individuals of the Danube Iron Gates Origin on the Dnipro River Rapids (Ukraine)? Archaeological and Bioarchaeological Records
- Special Issue on THE EARLY NEOLITHIC OF EUROPE, edited by F. Borrell, I. Clemente, M. Cubas, J. J. Ibáñez, N. Mazzucco, A. Nieto-Espinet, M. Portillo, S. Valenzuela-Lamas, & X. Terradas - Part II
- Early Neolithic Large Blades from Crno Vrilo (Dalmatia, Croatia): Preliminary Techno-Functional Analysis
- The Neolithic Flint Quarry of Pozarrate (Treviño, Northern Spain)
- From Anatolia to Algarve: Assessing the Early Stages of Neolithisation Processes in Europe
- What is New in the Neolithic? – A Special Issue Dedicated to Lech Czerniak, edited by Joanna Pyzel, Katarzyna Inga Michalak & Marek Z. Barański
- What is New in the Neolithic? – Celebrating the Academic Achievements of Lech Czerniak in Honour of His 70th Birthday
- Do We Finally Know What the Neolithic Is?
- Intermarine Area Archaeology and its Contribution to Studies of Prehistoric Europe
- Households and Hamlets of the Brześć Kujawski Group
- Exploiting Sheep and Goats at the Late Lengyel Settlement in Racot 18
- Colonists and Natives. The Beginning of the Eneolithic in the Middle Warta Catchment. 4500–3500 BC
- Is It Just the Location? Visibility Analyses of the West Pomeranian Megaliths of the Funnel Beaker Culture
- An Integrated Zooarchaeological and Micromorphological Perspective on Midden Taphonomy at Late Neolithic Çatalhöyük
- The Neolithic Sequence of the Middle Dunajec River Basin (Polish Western Carpathians) and Its Peculiarities
- Great Transformation on a Microscale: The Targowisko Settlement Region
- Special Issue on Digital Methods and Typology, edited by Gianpiero Di Maida, Christian Horn & Stefanie Schaefer-Di Maida
- Digital Methods and Typology: New Horizons
- Critique of Lithic Reason
- Unsupervised Classification of Neolithic Pottery From the Northern Alpine Space Using t-SNE and HDBSCAN
- A Boat Is a Boat Is a Boat…Unless It Is a Horse – Rethinking the Role of Typology
- Quantifying Patterns in Mortuary Practices: An Application of Factor Analysis and Cluster Analysis to Data From the Taosi Site, China
- Reexamining Ceramic Standardization During Agricultural Transition: A Geometric Morphometric Investigation of Initial – Early Yayoi Earthenware, Japan
- Statistical Analysis of Morphometric Data for Pottery Formal Classification: Variables, Procedures, and Digital Experiences of Medieval and Postmedieval Greyware Clustering in Catalonia (Twelfth–Nineteenth Centuries AD)
Articles in the same Issue
- Editorial
- Editorial: Open Archaeology in Challenging Times
- Regular Articles
- Caves, Senses, and Ritual Flows in the Iberian Iron Age: The Territory of Edeta
- Tutankhamun’s Polychrome Wooden Shawabtis: Preliminary Investigation for Pigments and Gilding Characterization and Indirect Dating of Previous Restorations by the Combined Use of Imaging and Spectroscopic Techniques
- When TikTok Discovered the Human Remains Trade: A Case Study
- Nuraghi as Ritual Monuments in the Sardinian Bronze and Iron Ages (circa 1700–700 BC)
- A Pilot Study in Archaeological Metal Detector Geophysical Survey
- A Blocked-Out Capital from Berenike (Egyptian Red Sea Coast)
- The Winery in Context: The Workshop Complex at Ambarçay, Diyarbakır (SE Turkey)
- Tracing Maize History in Northern Iroquoia Through Radiocarbon Date Summed Probability Distributions
- Faunal Remains Associated with Human Cremations: The Chalcolithic Pits 16 and 40 from the Perdigões Ditched Enclosures (Reguengos de Monsaraz, Portugal)
- A Multi-Method Study of a Chalcolithic Kiln in the Bora Plain (Iraqi Kurdistan): The Evidence From Excavation, Micromorphological and Pyrotechnological Analyses
- Potters’ Mobility Contributed to the Emergence of the Bell Beaker Phenomenon in Third Millennium BCE Alpine Switzerland: A Diachronic Technology Study of Domestic and Funerary Traditions
- From Foragers to Fisher-Farmers: How the Neolithisation Process Affected Coastal Fisheries in Scandinavia
- Enigmatic Bones: A Few Archaeological, Bioanthropological, and Historical Considerations Regarding an Atypical Deposit of Skeletonized Human Remains Unearthed in Khirbat al-Dusaq (Southern Jordan)
- Who Was Buried at the Petit-Chasseur Site? The Contribution of Archaeometric Analyses of Final Neolithic and Bell Beaker Domestic Pottery to the Understanding of the Megalith-Erecting Society of the Upper Rhône Valley (Switzerland, 3300–2200 BC)
- Erratum
- Erratum to “Britain In or Out of Europe During the Late Mesolithic? A New Perspective of the Mesolithic–Neolithic Transition”
- Review Article
- Archaeological Practices and Societal Challenges
- Special Issue Published in Cooperation with Meso’2020 – Tenth International Conference on the Mesolithic in Europe, edited by Thomas Perrin, Benjamin Marquebielle, Sylvie Philibert, and Nicolas Valdeyron - Part I
- Animal Teeth and Mesolithic Society
- A Matter of Scale: Responses to Landscape Changes in the Oslo Fjord, Norway, in the Mesolithic
- Chipped Stone Assemblage of the Layer B of the Kamyana Mohyla 1 Site (South-Eastern Ukraine) and the Issue of Kukrek in the North Meotic Steppe Region
- Rediscovered Mesolithic Rock Art Collection from Kamyana Mohyla Complex in Eastern Ukraine
- Mesolithic Montology
- A Little Mystery, Mythology, and Romance: How the “Pigmy Flint” Got Its Name
- Preliminary Results and Research Perspectives on the Submerged Stone Age Sites in Storstrømmen, Denmark
- Techniques and Ideas. Zigzag Motif, Barbed Line, and Shaded Band in the Meso-Neolithic Bone Assemblage at Zamostje 2, Volga-Oka Region (Russia)
- Modelling Foraging Cultures According to Nature? An Old and Unfortunately Forgotten Anthropological Discussion
- Mesolithic and Chalcolithic Mandibular Morphology: Using Geometric Morphometrics to Reconstruct Incomplete Specimens and Analyse Morphology
- Britain In or Out of Europe During the Late Mesolithic? A New Perspective of the Mesolithic–Neolithic Transition
- Non-Spatial Data and Modelling Multiscale Systems in Archaeology
- Living in the Mountains. Late Mesolithic/Early Neolithic Settlement in Northwest Portugal: Rock Shelter 1 of Vale de Cerdeira (Vieira do Minho)
- Enculturating Coastal Environments in the Middle Mesolithic (8300–6300 cal BCE) – Site Variability, Human–Environment Relations, and Mobility Patterns in Northern Vestfold, SE-Norway
- Why Mesolithic Populations Started Eating Crabs on the European Atlantic Façade Only Over the Past 15 Years?
- “Ain’t No Mountain High Enough” – Mesolithic Colonisation Processes and Landscape Usage of the Inner-Alpine Region Kleinwalsertal (Prov. Vorarlberg, Western Austria)
- Mesolithic Freshwater Fishing: A Zooarchaeological Case Study
- Consumers, not Contributors? The Study of the Mesolithic and the Study of Hunter-Gatherers
- Fish Processing in the Iron Gates Region During the Transitional and Early Neolithic Period: An Integrated Approach
- Hunting for Hide. Investigating an Other-Than-Food Relationship Between Stone Age Hunters and Wild Animals in Northern Europe
- Changing the Perspective, Adapting the Scale: Macro- and Microlithic Technologies of the Early Mesolithic in the SW Iberian Peninsula
- Fallen and Lost into the Abyss? A Mesolithic Human Skull from Sima Hedionda IV (Casares, Málaga, Iberian Peninsula)
- Evolutionary Dynamics of Armatures in Southern France in the Late Mesolithic and Early Neolithic
- Combining Agent-Based Modelling and Geographical Information Systems to Create a New Approach for Modelling Movement Dynamics: A Case Study of Mesolithic Orkney
- Pioneer Archaeologists and the Influence of Their Scientific Relationships on Mesolithic Studies in North Iberia
- Neolithisation in the Northern French Alps: First Results of the Lithic Study of the Industries of La Grande Rivoire Rockshelter (Isère, France)
- Late Mesolithic Individuals of the Danube Iron Gates Origin on the Dnipro River Rapids (Ukraine)? Archaeological and Bioarchaeological Records
- Special Issue on THE EARLY NEOLITHIC OF EUROPE, edited by F. Borrell, I. Clemente, M. Cubas, J. J. Ibáñez, N. Mazzucco, A. Nieto-Espinet, M. Portillo, S. Valenzuela-Lamas, & X. Terradas - Part II
- Early Neolithic Large Blades from Crno Vrilo (Dalmatia, Croatia): Preliminary Techno-Functional Analysis
- The Neolithic Flint Quarry of Pozarrate (Treviño, Northern Spain)
- From Anatolia to Algarve: Assessing the Early Stages of Neolithisation Processes in Europe
- What is New in the Neolithic? – A Special Issue Dedicated to Lech Czerniak, edited by Joanna Pyzel, Katarzyna Inga Michalak & Marek Z. Barański
- What is New in the Neolithic? – Celebrating the Academic Achievements of Lech Czerniak in Honour of His 70th Birthday
- Do We Finally Know What the Neolithic Is?
- Intermarine Area Archaeology and its Contribution to Studies of Prehistoric Europe
- Households and Hamlets of the Brześć Kujawski Group
- Exploiting Sheep and Goats at the Late Lengyel Settlement in Racot 18
- Colonists and Natives. The Beginning of the Eneolithic in the Middle Warta Catchment. 4500–3500 BC
- Is It Just the Location? Visibility Analyses of the West Pomeranian Megaliths of the Funnel Beaker Culture
- An Integrated Zooarchaeological and Micromorphological Perspective on Midden Taphonomy at Late Neolithic Çatalhöyük
- The Neolithic Sequence of the Middle Dunajec River Basin (Polish Western Carpathians) and Its Peculiarities
- Great Transformation on a Microscale: The Targowisko Settlement Region
- Special Issue on Digital Methods and Typology, edited by Gianpiero Di Maida, Christian Horn & Stefanie Schaefer-Di Maida
- Digital Methods and Typology: New Horizons
- Critique of Lithic Reason
- Unsupervised Classification of Neolithic Pottery From the Northern Alpine Space Using t-SNE and HDBSCAN
- A Boat Is a Boat Is a Boat…Unless It Is a Horse – Rethinking the Role of Typology
- Quantifying Patterns in Mortuary Practices: An Application of Factor Analysis and Cluster Analysis to Data From the Taosi Site, China
- Reexamining Ceramic Standardization During Agricultural Transition: A Geometric Morphometric Investigation of Initial – Early Yayoi Earthenware, Japan
- Statistical Analysis of Morphometric Data for Pottery Formal Classification: Variables, Procedures, and Digital Experiences of Medieval and Postmedieval Greyware Clustering in Catalonia (Twelfth–Nineteenth Centuries AD)