mTOR modulates CD8+ T cell differentiation in mice with invasive pulmonary aspergillosis
-
 and
and

Abstract
CD8+ T cells are a vital component of the adaptive immune system and important for eliminating intracellular pathogens. Notably, mTOR activity is associated with CD8+ T effector memory (Tem) cell differentiation in fungal infections. This study investigates the molecular mechanisms of CD8+ Tem cell proliferation and differentiation mediated by the mTOR pathway in immunosuppressed mice with invasive pulmonary aspergillosis (IPA). We first established the immunosuppressed IPA mouse model, then mice were subjected to rapamycin treatment daily or interleukin (IL)-12 treatment every other day. Lung tissues and blood samples were obtained seven days later. Aspergillus fumigatus was cultured from the lung tissue of mice inoculated with A. fumigatus spores. After IL-12 treatment, the expression of mTOR and its downstream signaling molecule S6 kinase, number of CD8+ Tem cells and interferon-γ expression were significantly increased, while they were significantly decreased after treatment with rapamycin. Additionally, IL-12 treatment induced T-bet but inhibited Eomesodermin expression, while the opposite was seen when the mTOR pathway was blocked by rapamycin. In conclusion, we found that the mTOR pathway induced CD8+ T cell proliferation and differentiation by regulating T-bet and Eomesodermin expression, which significantly influenced immune regulation during IPA and enhanced the immune response against fungal infection.
1 Introduction
The number of community and hospitalized patients with compromised host defenses has increased dramatically in recent years. This increase is due mainly to the significant progress in transplantation procedures, the rapid development of cancer chemotherapy and immunotherapy, and the broader use of corticosteroids [1]. Invasive pulmonary aspergillosis (IPA) is one of the devastating comorbidities of immunosuppression, especially in patients with immune dysfunction [2]. CD8+ T cells are a vital component of the adaptive immune system and important for eliminating intracellular pathogens [3]. The effector-phenotype CD8+ memory T cells (Tem) are responsible for protecting the host from subsequent reinfection with the same pathogen by rapidly expanding, engaging cytolytic activity and expressing effector cytokines [4, 5]. In an earlier clinical study, we found that the number of CD8+ T cell subsets during the early stage of IPA decreased significantly and the CD8+ T cell differentiation phenotype is closely related to the patient prognosis [1]. Therefore, focusing on the molecular mechanism of CD8+ T cell differentiation in the early stage of infection may provide a new opportunity to reverse the abnormal immune response in severely infected patients and effectively control the infection.
The mammalian target of rapamycin (mTOR) signaling pathway plays an important role in regulating cell metabolism, survival, growth and proliferation. The ribosomal S6 kinase (S6K) is one of the key substrates downstream of the mTOR signaling pathway. Activated mTOR phosphorylates S6K, thereby starting the process of transcription to synthesize proteins for cell proliferation [6]. Recent studies indicated that mTOR contributes to proliferation and differentiation of both CD4+ and CD8+ T cells in response to antigen stimulation [7, 8], but the mechanisms by which mTOR integrates extracellular signals with intracellular molecule expression and activity to achieve distinct functions in CD8+ T cells, especially in immunocompromised hosts with IPA, remains unknown.
The transcription factors T-box expressed in T cells (T-bet) and Eomesodermin (Eomes) are important transcription factors which regulate effector T cells and participate in the differentiation and development of CD4+ effector I type T cells or type 1 helper T (Th1) cells, NK cells, and CD8+ cytotoxic T lymphocytes [9, 10]. Recent studies indicate that interleukin (IL)-12 upregulates T-bet but inhibits Eomes expression to favor the generation of effector CD8+ T cells versus memory CD8+ T cells [11]. Moreover, IL-12 can also enhance the activity of mTOR kinase in naïve CD8+ T cells, and the activation of the mTOR pathway was significantly associated with CD8+ Tem cell differentiation in fungal infections [12, 13]. Therefore, it is important for us to understand the cell-intrinsic factors which regulate the expression of T-bet and Eomes and generate required CD8+ T cell functions during the immune response in IPA.
In patients with IPA, IL-6 functions as a proinflammatory cytokine and restores antifungal activity, while IL-10 is an anti-inflammatory cytokine and has the opposite effect through down-regulating Th1 and macrophage responses [14, 15]. The plasma level of IL-6 and IL-10 can thus reflect the immune response of the host. Galactomannan, a polysaccharide composed of units of D-galactose and D-mannose, is one component of the Aspergillus cell wall. Plasma galactomannan levels can represent the burden of fungal infection, so has been used to diagnose IPA in the clinic [16].
Based on the experimental results and theories mentioned above, we hypothesized that the role of mTOR during the immune response in immunosuppressed individuals with IPA is regulating CD8+ Tem cell proliferation and differentiation by modifying the expression of T-bet and Eomes. Increasing mTOR activity induced T-bet-mediated CD8+ Tem cell differentiation, higher levels of inflammatory cytokines, and greater antifungal efficacy, which could be a new strategy to modulate immune responses in IPA infection.
2 Materials and Methods
2.1 Pathogen preparation
The Aspergillus fumigatus used in this study was obtained from a case of pulmonary aspergillosis. We cultured the viable conidia by growing on Sabouraud dextrose agar at 35°C for 5 days. Conidia were harvested with Tween-80/PBS and then filtered through five layers of gauze. We used the turbidity method to adjust the concentration of conidia to 1×108 CFU/mL.
2.2 Invasive pulmonary aspergillosis model preparation
Healthy 6-8 weeks old female BALB/c mice weighing 20 ± 5 g, were obtained from the Animal Facility Center, PUMCH. All animals were housed in a pathogen-free environment. Mice were randomly divided into the following groups (6 mice per group): (1) Control group: no treatment; (2) CTX + IPA: after intraperitoneal injection of cyclophosphamide (CTX) (200 mg kg-1 d-1, Jiangsu Hengrui Medicine Co. Ltd., Lianyungang, China) for 5 days, the mice were inoculated with 0.1 ml A. fumigatus spore suspension intranasally; (3) CTX + IPA + IL-12: mice were given 5 μg/kg IL-12 intraperitoneally every other day for seven days after the injection of CTX and inoculation with A. fumigatus spore suspension. IL-12 concentration was chosen based on the common concentration that is used to favor the generation of effector CD8+ T cells (8); (4) CTX + IPA + RAPA: Mice were given 2 mg/kg rapamycin intraperitoneally for seven days after injection of CTX and inoculation with A. fumigatus spore suspension. The concentration of rapamycin was chosen to inhibit the mTOR pathway (8). Blood samples and lung tissue were collected seven days after the injection of IL-12 or rapamycin. One part of the lung tissue was minced and used for A. fumigatus culture, the other part of the lung tissue was fixed with 4% formaldehyde. All experiments were repeated twice.
Ethical approval
The research related to animals use has been complied with all the relevant national regulations and institutional policies for the care and use of animals.
2.3 Lung tissue histology
The lung tissue was first treated with 4% formaldehyde for histological examination, then lung tissue sections of 3 μm in thickness were affixed to slides, deparaffinized, and stained with hematoxylin-eosin (H&E), masson trichrome (Masson), and periodic Schiff-methenamine (PASM) stain to evaluate the structure of lung tissue in different groups. 10 fields (× 200, × 400, × 600) were randomly selected on each slide.
2.4 CD8+ Tem cell counts and IFN-γ, mTOR, S6K, T-bet, and EOMES expression levels
We used flow cytometry to isolate and count CD8+ Tem cells. Cells were labeled with the following fluorescently conjugated monoclonal antibodies: anti-mouse CD45-PE (12045181, eBioscience, San Diego, CA, USA), anti-mouse CD8a-APC (17008181, eBioscience), anti-mouse CD44-PE (25044181, eBioscience), and anti-mouse CD62L (104432, BioLegend, San Diego, CA, USA). After the CD8+ Tem cells were sorted by flow cytometry, they were stained with antibodies against mTOR (ab87540, Abcam, Cambridge, MA, USA), interferon γ (IFN-γ) (11731181, eBioscience), T-bet (ab91109, Abcam), S6K (ab32529, Abcam) and Eomes (53-4875-82, eBioscience).
2.5 Cytokine quantification
We used enzyme-linked immunosorbent assay (ELISA) kits to quantify cytokines and severity of fungal infection. The kits were used to detect IL-6 (Abcam), IL-10 (Abcam), and galactomannan (Affymetrix Bioscience, Santa Clara, CA, USA).
2.6 Statistical analysis
Data were analyzed with SPSS 18.0 (IBM SPSS, Chicago, IL, USA). Student’s t-test or analysis of variance (ANOVA) were used to determine the statistical significance of differences. A P-value of p < 0.05 was considered statistically significant.
3 Results
3.1 A. fumigates Culture and Lung Histology
We detected the viability of the conidia through tissue culture. Viable A. fumigatus was seen in lung from the CTX + IPA, CTX + IPA + IL-12 and CTX + IPA + RAPA groups (Figure 1A–C), but we did not observe any viable A. fumigatusin the cultured lung tissue from the control group. Histological examination showed that the structure of the lung tissue was intact in the lungs of control mice (Figure 1D). In contrast, inflammatory cell infiltration, blood congestion, and interstitial lung tissue injury were observed in the mice from the CTX + IPA, CTX + IPA + IL-12 and CTX + IPA + RAPA groups (Figure 1E–G).
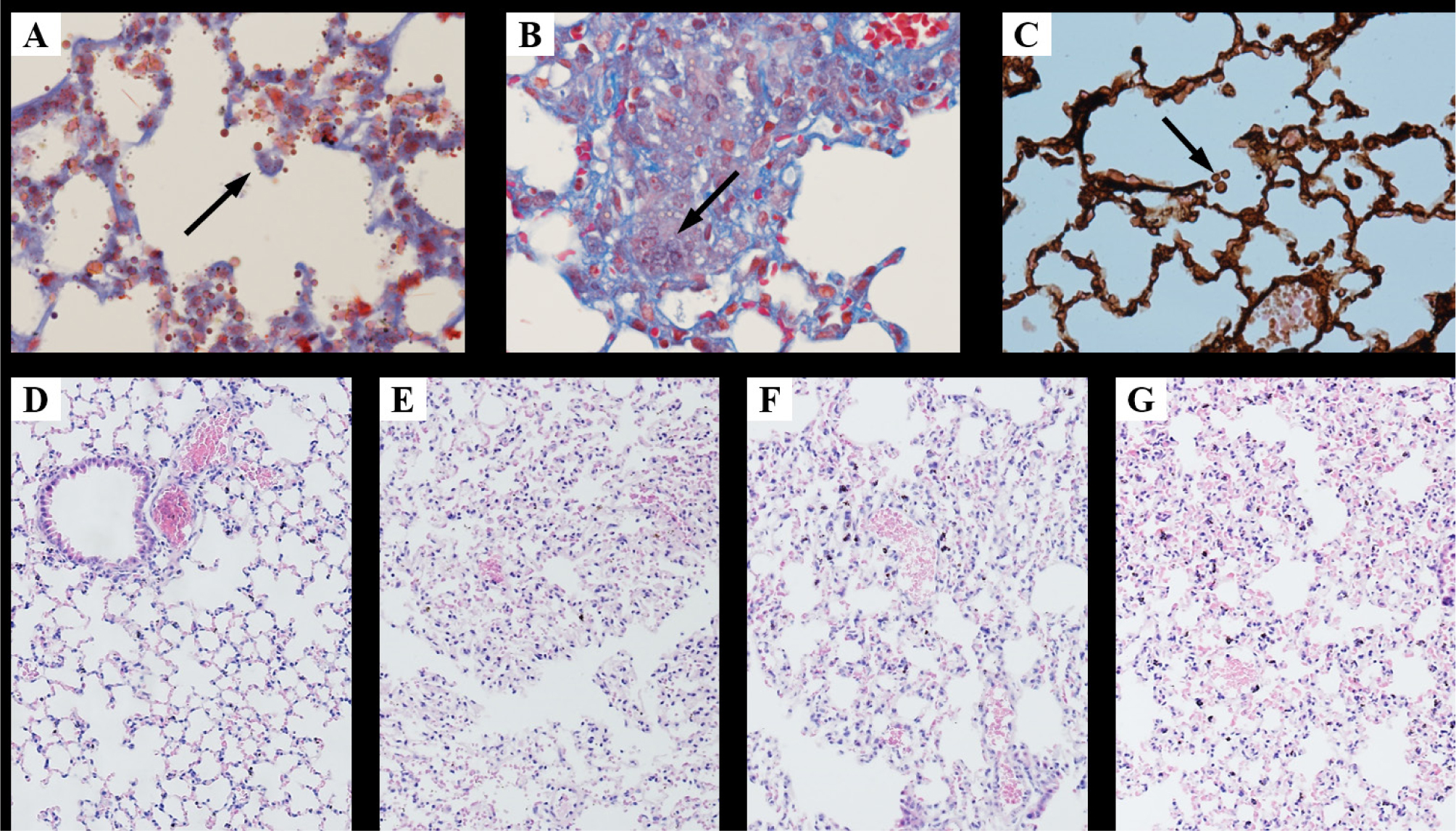
Histology of lung tissue stained with hematoxylin and eosin, Masson trichrome, and PASM. A–C: Fungal spores in the lung tissue of a mouse infected with A. fumigatus stained of Masson trichrome at magnifications of 200× (A), 400× (B), and PASM staining at 600× (C). D–G: HE-stained lung sections at a magnification of 100× from an animal in the control group (D), the cyclophosphamide immunosuppression plus invasive pulmonary aspergillosis (CTX + IPA) group (E), the immunosuppression plus IPA plus IL-12 treatment (CTX + IPA + IL-12) group (F), or the immunosuppression plus IPA plus rapamycin treatment (CTX + IPA + RAPA) group (G).
3.2 mTOR modulates the differentiation of CD8+ T cells
To determine the role of the mTOR pathway in CD8+ Tem cell differentiation, we upregulated the activity of mTOR using IL-12. As shown in Figure 2, there was a significant increase in the proportion of CD8+ Tem cells as well as their level of IFN-γ production in the CTX + IPA + IL-12 group (CD8+ Tem cells: 47% ± 6%; IFN-γ: 0.09 ± 0.03) compared with the CTX + IPA group (CD8+ Tem cells: 35% ± 10%, p = 0.024; IFN-γ: 0.05 ± 0.04, p = 0.032). In addition to this, we found compared with the control group (0.25 ± 0.04) and CTX + IPA group (0.29 ± 0.04) mTOR expression was significantly increased in the CTX + IPA + IL-12 group (0.39 ± 0.12; p = 0.004 and p = 0.034, respectively). We also detected the expression of S6K, a well-known downstream molecule of the mTOR pathway. As expected, correlated with mTOR expression, IL-12 also significantly enhanced the expression of S6K in CD8+ Tem cells (0.13 ± 0.03) compared with the control group (0.06 ± 0.04, p < 0.001) and the CTX + IPA group (0.09 ± 0.01, p = 0.032).

The proportion of CD8+ effector memory T (Tem) cells and levels of IFN-γ, mTOR and S6K in CD8+ Tem cells were significantly increased by IL-12 stimulation but significantly decreased by rapamycin treatment. The peripheral blood mononuclear cells obtained from the cyclophosphamide plus invasive pulmonary aspergillosis (CTX + IPA) mice, CTX + IPA mice treated with IL-12 (CTX + IPA+IL-12) or rapamycin (CTX + IPA+RAPA), and control animals were assessed using flow cytometry seven days after A. fumigatus infection. The side scatter (SSC) and CD8a were applied to gate on CD8+ T lymphocytes, CD44+ CD45+ CD62+/– were considered to represent the Tem cells. Representative dot plots are shown, and the percentages of Tem cells and of Tem cells expressing IFNγ, mTOR, and S6K are expressed as the mean ± S.D. *p < 0.05; **p < 0.01; ***p< 0.001.
As shown in Figure 2, when the mTOR pathway was blocked by rapamycin, the expression of mTOR was significantly decreased (p < 0.05). S6K expression was likewise decreased in the CTX + IPA + RAPA group (0.04 ± 0.04) compared with the CTX + IPA group (0.09 ± 0.01, p = 0.01). More importantly, we also found that rapamycin significantly decreased the number of CD8+ Tem cells and the production of IFN-γ (CD8+ Tem cells: 0.22 ± 0.05; IFN-γ: 0.02 ± 0.01) compared with the CTX + IPA group (CD8+ Tem cells: 0.35 ± 0.10, p = 0.017; IFN-γ: 0.05 ± 0.04, p = 0.039) or the CTX + IPA + IL-12 group (CD8+ Tem cells: 0.47 ± 0.06, p < 0.001; IFN-γ: 0.09 ± 0.03, p < 0.001). These results show that the mTOR pathway was essential for the differentiation of CD8+ Tem cells and the production of type I effector functions.
3.3 T-bet and Eomesodermin expression was regulated by the mTOR pathway
IL-12 can promote the generation of effector CD8+ T cells versus memory CD8+ T cells by increasing T-bet and inhibiting Eomes expression. To determine the role of the mTOR pathway in the expression of T-bet and Eomes during the immune response in IPA, we examined the expression of T-bet and Eomes in CD8+ Tem cells from IL-12-treated IPA mice. As shown in Figure 3, we found that after treating mice with IL-12, T-bet expression was increased (p < 0.05) while the expression of Eomes was inhibited (p < 0.05) in CD8+ Tem cells, which favors the generation of effector cells over memory cells. We also blocked the mTOR pathway with rapamycin to examine the effect on expression of T-bet and Eomes. In contrast to the effect of IL-12, the expression of T-bet was decreased, while the expression of Eomes was increased, in CD8+ Tem cells after treating mice with rapamycin (both p < 0.001). These observations show that mTOR-regulated expression of T-bet and Eomes may modulate CD8+ T cell differentiation during immune responses in IPA.

IL-12 significantly increased the expression of T-bet but significantly decreased the expression of Eomesodermin in CD8+ effector memory T (Tem) cells, which was blocked by rapamycin treatment. The peripheral blood mononuclear cells obtained from the cyclophosphamide plus invasive pulmonary aspergillosis (CTX + IPA) mice, CTX + IPA mice treated with IL-12(CTX + IPA+IL-12) or rapamycin (CTX + IPA+RAPA), and control animals were assessed using flow cytometry seven days after A. fumigatus infection. The side scatter (SSC) and CD8a were applied to gate on CD8+ T lymphocytes, CD44+ CD45+ CD62+/– were considered to represent the Tem cells. Representative dot plots are shown, and the percentages of Tem cells and of Tem cells expressing T-bet and Eomesodermin are expressed as the mean ± S.D. *p < 0.05; **p < 0.01; ***p < 0.001.
3.4 Changes in the inflammatory response and fungal infection severity
As shown in Figure 4, compared with the control group (245.65 ± 85.78 pg/ml), the CTX + IPA group (1426.57 ± 488.03 pg/ml), and the CTX + IPA + RAPA group (822.74 ± 211.30 pg/ml), the CTX + IPA + IL-12 group had the highest level of IL-6 (2053.91 ± 402.37 pg/ml; p < 0.001, p = 0.036 and p < 0.001, respectively), which reflected the inflammatory response in IPA mice. In contrast, the level of IL-10, which reflects the anti-inflammatory response, showed the opposite trend. The IL-10 concentration in CTX + IPA + IL-12 and the control group was significantly lower than in the CTX + IPA group (p < 0.001 for both) and the CTX + IPA + RAPA group (p < 0.001 for both).
Galactomannan is used to determine the severity of fungal infection in the clinic. In this study, as shown in Figure 4, after intranasal inoculation with A. fumigatus, the galactomannan level was significantly increased in the CTX + IPA, CTX + IPA + IL-12, and CTX + IPA + RAPA groups compared with the control group (p < 0.001 for all). Interestingly, compared with the CTX + IPA and CTX + IPA + RAPA groups, galactomannan level was significantly lower in the CTX + IPA + IL-12 group (p < 0.001, respectively).

Inflammatory responses and severity of fungal infection in invasive pulmonary aspergillosis. ELISAs were performed to assess the levels of IL-6 (left), IL-10 (middle), and galactomannan (right) in the plasma samples obtained from the cyclophosphamide plus invasive pulmonary aspergillosis (CTX + IPA) mice, CTX + IPA mice treated with IL-12 (CTX + IPA+IL-12) or rapamycin (CTX + IPA+RAPA), and control animals seven days after A. fumigatus infection. The data are presented as the mean ± S.D. *p < 0.05; **p < 0.01; ***p < 0.001.
4 Discussion
Our study shows that IL-12 increased CD8+ Tem cells number and type I effector maturation (IFN-γ production) in immunocompromised hosts with IPA. This occurred along with increased mTOR expression and activity, enhanced T-bet levels, and decreased Eomes levels in the CD8+ Tem cells. These changes coincided with a significant increase in the level of IL-6 and a significant decrease in the level of IL-10, as well as a decrease in the fungal infection load (as measured by galactomannan levels). All these effects could be blocked by the inhibition of the mTOR pathway with rapamycin.
CD8+ T cells play an important role in the adaptive immune system. The ratio and number of the different phenotypes of CD8+ T cells determines the overall function of CD8+ T cells. After infection, some CD8+ T cells take on a memory cell phenotype and can survive for a long time after their induction. The CD8+ Tem cells, a sub-population of memory T cells that are differentiated with an effector cell phenotype, are the cellular basis for the rapid host immune response that occurs in the early stage of infection [4, 5]. Recent evidence has shown that various kinds of pathogens can promote CD8+ T cell differentiation, which is closely related to the effectiveness of the host immune response and the occurrence of immune disorders in a septic host [17, 18]. Immunosuppressed patients are at high risk of repeated infections. It is thus crucial for these patients to have an effective immune response in the early stage of infection to avoid more severe infection in the later. Differentiation of CD8+ T cells is regulated by several transcription factors and signal transduction pathways [22, 23]. Therefore, targeting molecular mechanisms that mediate the differentiation of CD8+ T cells offers new ways to promote CD8+ T cell regulated immune responses in immunocompromised hosts with IPA.
The energy-sensitive mTOR signaling pathway integrates environmental stimuli to regulate important cellular biochemical metabolic pathways and processes. Recent evidence has demonstrated the role of the mTOR pathway in memory CD8+ T cell differentiation [19]. Studies have indicated that inhibiting the mTOR pathway with rapamycin could block the differentiation of CD8+ T cells, which demonstrates that the mTOR pathway is required for CD8+ T cell differentiation [20, 21]. Furthermore, a recent study indicated that IL-12 could enhance T-bet and inhibit Eomes expression to favor the generation of effector CD8+ T cells versus memory CD8+ T cells [11]. Moreover, IL-12 could also enhance the activity of mTOR kinase in naïve CD8+ T cells. Therefore, we hypothesized that the upregulation of CD8+ T cell differentiation by IL-12 may be mediated via the mTOR pathway. To confirm our hypothesis, we stimulated mice with IL-12 following CTX-mediated immunosuppression and A. fumigatus infection and found increased mTOR activity, and enhanced CD8+ Tem proliferation and type I effector maturation. The effector functions induced by IL-12 treatment were significantly attenuated by the mTOR specific inhibitor rapamycin, confirming that enhancing and maintaining the activity of the mTOR pathway is essential for IL-12-induced proliferation and type I effector function in CD8+ Tem in response to a fungal infection.
Several transcription factors have been shown to regulate the differentiation of CD8+ T cells during antigen stimulation such as T-bet and Eomes. T-bet and Eomes belong to the T-box gene family and both are essential in regulating the differentiation of T cells. T-bet plays an important role in modulation of the differentiation of Th1-type effector cells and is required for the synthesis of inflammatory factors, while the expression Eomes increases in the later stages of an infection and accelerates the effect of CD8+ Tem cell transformation [22, 23]. Recent studies have found that mTOR activity is essential for T-bet mediated effector functions and blockade of mTOR activity promotes Eomes expression and memory generation [11]. Our results indicated that promoting the expression and activity of the mTOR pathway could increase the proliferation rate of CD8+ Tem cells and IFN-γ expression. In contrast, the mTOR inhibitor rapamycin blocked the expression of T-bet and increased expression of Eomes in CD8+ Tem cells. This result showed that the mTOR signaling pathway can influence the proliferation and differentiation of memory and effector CD8+ T cells by regulating T-bet and Eomes activity, however the mechanism by which mTOR regulates T-bet and Eomes expression deserves further study.
The current study also elucidated the overall outcome on the immunosuppressed host with IPA. Our results showed that, compared with the control and CTX + IPA groups, IL-12 treatment increased plasma proinflammatory cytokines (IL-6), decreased anti-inflammatory factors (IL-10), and reduced the fungal infection load, as assessed by galactomannan level, which is used clinically as a diagnostic indicator of IPA. The mTOR inhibitor rapamycin caused completely the opposite effects. These results suggest that the mTOR signaling pathway plays a vital role in CD8+ T cell differentiation and function, which may result in changes in the immune response and fungal load in immunosuppressed patients with IPA infection.
In summary, we found that mTOR regulates CD8+ T cell proliferation and differentiation by modifying T-bet and Eomesodermin expression, and activating this pathway with IL-12 enhanced the immune response against fungal infection in IPA. Whether this mechanism offers a new target to modulate CD8+ T cell responses in antifungal immunotherapy requires further study.
Conflict of interest: Authors state no conflict of interest.
Acknowledgments
The work was supported by the National Natural Science Foundation of China (no. 81601657)
References
[1] Cui N, Wang H, Long Y, Liu D. CD8 (+) T-cell counts: an early predictor of risk and mortality in critically ill immunocompromised patients with invasive pulmonary aspergillosis. Critical care (London, England). 2013; 17 (4):R157.10.1186/cc12836Search in Google Scholar PubMed PubMed Central
[2] Patterson TF, Thompson GR, 3rd, Denning DW, Fishman JA, Hadley S, Herbrecht R, et al. Practice Guidelines for the Diagnosis and Management of Aspergillosis: 2016 Update by the Infectious Diseases Society of America. Clinical infectious diseases: an official publication of the Infectious Diseases Society of America. 2016; 63 (4): e1-e60.10.1093/cid/ciw326Search in Google Scholar PubMed PubMed Central
[3] Zhang N, Bevan MJ. CD8(+) T cells: foot soldiers of the immune system. Immunity. 2011; 35 (2): 161-8.10.1016/j.immuni.2011.07.010Search in Google Scholar PubMed PubMed Central
[4] Williams MA. Instant recall: a key role for effector-phenotype CD8 (+) memory T cells in immune protection. Immunity. 2013; 38 (6): 1090-1.10.1016/j.immuni.2013.06.007Search in Google Scholar PubMed
[5] Plumlee CR, Sheridan BS, Cicek BB, Lefrancois L. Environmental cues dictate the fate of individual CD8+ T cells responding to infection. Immunity. 2013; 39 (2): 347-56.10.1016/j.immuni.2013.07.014Search in Google Scholar PubMed PubMed Central
[6] Cobbold SP. The mTOR pathway and integrating immune regulation. Immunology. 2013; 140 (4): 391-8.10.1111/imm.12162Search in Google Scholar PubMed PubMed Central
[7] Chang JT, Palanivel VR, Kinjyo I, Schambach F, Intlekofer AM, Banerjee A, et al. Asymmetric T lymphocyte division in the initiation of adaptive immune responses. Science (New York, NY). 2007; 315 (5819): 1687-91.10.1126/science.1139393Search in Google Scholar PubMed
[8] Cui N, Su LX, Wang H, Xiao M, Yang F, Zheng M, et al. mTOR Modulates Lymphocyte Differentiation through T-bet and Eomesodermin in Response to Invasive Pulmonary Aspergillosis in Rats. Chinese medical journal. 2016; 129 (14): 1704-10.10.4103/0366-6999.185858Search in Google Scholar PubMed PubMed Central
[9] Delgoffe GM, Kole TP, Zheng Y, Zarek PE, Matthews KL, Xiao B, et al. The mTOR kinase differentially regulates effector and regulatory T cell lineage commitment. Immunity. 2009; 30 (6): 832-44.10.1016/j.immuni.2009.04.014Search in Google Scholar PubMed PubMed Central
[10] McLane LM, Banerjee PP, Cosma GL, Makedonas G, Wherry EJ, Orange JS, et al. Differential localization of T-bet and Eomes in CD8 T cell memory populations. Journal of immunology (Baltimore, Md: 1950). 2013; 190 (7): 3207-15.10.4049/jimmunol.1201556Search in Google Scholar PubMed PubMed Central
[11] Rao RR, Li Q, Odunsi K, Shrikant PA. The mTOR kinase determines effector versus memory CD8+ T cell fate by regulating the expression of transcription factors T-bet and Eomesodermin. Immunity. 2010; 32 (1): 67-78.10.1016/j.immuni.2009.10.010Search in Google Scholar PubMed PubMed Central
[12] Takemoto N, Intlekofer AM, Northrup JT, Wherry EJ, Reiner SL. Cutting Edge: IL-12 inversely regulates T-bet and eomesodermin expression during pathogen-induced CD8+ T cell differentiation. Journal of immunology (Baltimore, Md : 1950). 2006; 177 (11): 7515-9.10.4049/jimmunol.177.11.7515Search in Google Scholar PubMed
[13] Buggert M, Tauriainen J, Yamamoto T, Frederiksen J, Ivarsson MA, Michaelsson J, et al. T-bet and Eomes are differentially linked to the exhausted phenotype of CD8+ T cells in HIV infection. PLoS pathogens. 2014; 10 (7): e1004251.10.1371/journal.ppat.1004251Search in Google Scholar PubMed PubMed Central
[14] Cenci E, Mencacci A, Casagrande A, Mosci P, Bistoni F, Romani L. Impaired antifungal effector activity but not inflammatory cell recruitment in interleukin-6-deficient mice with invasive pulmonary aspergillosis. The Journal of infectious diseases. 2001; 184 (5): 610-7.10.1086/322793Search in Google Scholar PubMed
[15] Clemons KV, Grunig G, Sobel RA, Mirels LF, Rennick DM, Stevens DA. Role of IL-10 in invasive aspergillosis: increased resistance of IL-10 gene knockout mice to lethal systemic aspergillosis. Clinical and experimental immunology. 2000; 122 (2):186-91.10.1046/j.1365-2249.2000.01382.xSearch in Google Scholar PubMed PubMed Central
[16] Ambasta A, Carson J, Church DL. The use of biomarkers and molecular methods for the earlier diagnosis of invasive aspergillosis in immunocompromised patients. Medical mycology. 2015; 53(6): 531-57.10.1093/mmy/myv026Search in Google Scholar PubMed
[17] Duong S, Condotta SA, Rai D, Martin MD, Griffith TS, Badovinac VP. Polymicrobial sepsis alters antigen-dependent and -independent memory CD8 T cell functions. Journal of immunology (Baltimore, Md: 1950). 2014; 192(8): 3618-25.10.4049/jimmunol.1303460Search in Google Scholar PubMed PubMed Central
[18] Lertmemongkolchai G, Cai G, Hunter CA, Bancroft GJ. Bystander activation of CD8+ T cells contributes to the rapid production of IFN-gamma in response to bacterial pathogens. Journal of immunology (Baltimore, Md : 1950). 2001; 166 (2): 1097-105.10.4049/jimmunol.166.2.1097Search in Google Scholar PubMed
[19] Araki K, Turner AP, Shaffer VO, Gangappa S, Keller SA, Bachmann MF, et al. mTOR regulates memory CD8 T-cell differentiation. Nature. 2009;460(7251):108-12.10.1038/nature08155Search in Google Scholar PubMed PubMed Central
[20] Ouyang W, Beckett O, Ma Q, Paik JH, DePinho RA, Li MO. Foxo proteins cooperatively control the differentiation of Foxp3+ regulatory T cells. Nature immunology. 2010; 11 (7): 618-27.10.1038/ni.1884Search in Google Scholar PubMed
[21] Sinclair LV, Finlay D, Feijoo C, Cornish GH, Gray A, Ager A, et al. Phosphatidylinositol-3-OH kinase and nutrient-sensing mTOR pathways control T lymphocyte trafficking. Nature immunology. 2008; 9 (5): 513-21.10.1038/ni.1603Search in Google Scholar PubMed PubMed Central
[22] Intlekofer AM, Takemoto N, Wherry EJ, Longworth SA, Northrup JT, Palanivel VR, et al. Effector and memory CD8+ T cell fate coupled by T-bet and eomesodermin. Nature immunology. 2005; 6 (12): 1236-44.10.1038/ni1268Search in Google Scholar PubMed
[23] Mullen AC, High FA, Hutchins AS, Lee HW, Villarino AV, Livingston DM, et al. Role of T-bet in commitment of TH1 cells before IL-12-dependent selection. Science (New York, NY). 2001; 292 (5523): 1907-10.10.1126/science.1059835Search in Google Scholar PubMed
Abbreviations
- CTX
cyclophosphamide
- Eomes
Eomesodermin
- H&E
hematoxylin-eosin
- IL
interleukin
- IPA
invasive pulmonary aspergillosis
- Masson
masson trichrome
- mTOR
the mammalian target of rapamycin
- PASM
periodic Schiff-methenamine
- RAPA
Rapamycin
- S6K
the ribosomal S6 kinase
- T-bet
T-box expressed in T cells
- Tem
CD8+ T effector memory
- Th1
type 1 helper T cells
© 2018 Hao Wang et al.
This work is licensed under the Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 License.
Articles in the same Issue
- Research Article
- Purification of Tea saponins and Evaluation of its Effect on Alcohol Dehydrogenase Activity
- Runt-related transcription factor 3 promoter hypermethylation and gastric cancer risk: A meta-analysis
- Risk Factors for Venous Thromboembolism in Hospitalized Patients in the Chinese Population
- Value of Dual-energy Lung Perfusion Imaging Using a Dual-source CT System for the Pulmonary Embolism
- A new combination of substrates: biogas production and diversity of the methanogenic microorganisms
- mTOR modulates CD8+ T cell differentiation in mice with invasive pulmonary aspergillosis
- Direct Effects on Seed Germination of 17 Tree Species under Elevated Temperature and CO2 Conditions
- Role of water soluble vitamins in the reduction diet of an amateur sportsman
- Aberrant DNA methylation involved in obese women with systemic insulin resistance
- 16S ribosomal RNA-based gut microbiome composition analysis in infants with breast milk jaundice
- Characterization of Haemophilus parasuis Serovar 2 CL120103, a Moderately Virulent Strain in China
- MiRNA-145 induces apoptosis in a gallbladder carcinoma cell line by targeting DFF45
- Telmisartan induces osteosarcoma cells growth inhibition and apoptosis via suppressing mTOR pathway
- Optimizing the Formulation for Ginkgolide B Solid Dispersion
- Determination of the In Vitro Gas Production and Potential Feed Value of Olive, Mulberry and Sour Orange Tree Leaves
- Factors Influencing the Successful Isolation and Expansion of Aging Human Mesenchymal Stem Cells
- The Value of Diffusion-Weighted Magnetic Resonance Imaging in Predicting the Efficacy of Radiation and Chemotherapy in Cervical Cancer
- Chemical profile and antioxidant activity of Trollius europaeus under the influence of feeding aphids
- SSR Markers Suitable for Marker Assisted Selection in Sunflower for Downy Mildew Resistance
- A Fibroblast Growth Factor Antagonist Peptide Inhibits Breast Cancer in BALB/c Mice
- Antihyperglycemic and antihyperlipidemic effects of low-molecular-weight carrageenan in rats
- Microbial indicators and environmental relationships in the Umhlangane River, Durban, South Africa
- TUFT1 promotes osteosarcoma cell proliferation and predicts poor prognosis in osteosarcoma patients
- Long non-coding RNA-2271 promotes osteogenic differentiation in human bone marrow stem cells
- The prediction of cardiac events in patients with acute ST segment elevation myocardial infarction: A meta–analysis of serum uric acid
- Risk expansion of Cr through amphibious clonal plant from polluted aquatic to terrestrial habitats
- Overexpression of Zinc Finger Transcription Factor ZAT6 Enhances Salt Tolerance
- Sini decoction intervention on atherosclerosis via PPARγ-LXRα-ABCA1 pathway in rabbits
- Soluble myeloid triggering receptor expressed on myeloid cell 1 might have more diagnostic value for bacterial ascites than C-reactive protein
- A Preliminary Study on the Newly Isolated High Laccase-producing Fungi: Screening, Strain Characteristics and Induction of Laccase Production
- Hydrolytic Enzyme Production by Thermophilic Bacteria Isolated from Saudi Hot Springs
- Analysis of physiological parameters of Desulfovibrio strains from individuals with colitis
- Emodin promotes apoptosis of human endometrial cancer through regulating the MAPK and PI3K/ AKT pathways
- Down-regulation of miR-539 indicates poor prognosis in patients with pancreatic cancer
- Inhibitory activities of ethanolic extracts of two macrofungi against eggs and miracidia of Fasciola spp.
- PAQR6 expression enhancement suggests a worse prognosis in prostate cancer patients
- Characterization of a potential ripening regulator, SlNAC3, from Solanum lycopersicum
- Expression of Angiopoietin and VEGF in cervical cancer and its clinical significance
- Umbilical Cord Tissue Derived Mesenchymal Stem Cells Can Differentiate into Skin Cells
- Isolation and Characterization of a Phage to Control Vancomycin Resistant Enterococcus faecium
- Glycogen Phosphorylase Isoenzyme Bb, Myoglobin and BNP in ANT-Induced Cardiotoxicity
- BAG2 overexpression correlates with growth and poor prognosis of esophageal squamous cell carcinoma
- Relationship between climate trends and grassland yield across contrasting European locations
- Review Articles
- Mechanisms of salt tolerance in halophytes: current understanding and recent advances
- Salivary protein roles in oral health and as predictors of caries risk
- Nanoparticles as carriers of proteins, peptides and other therapeutic molecules
- Survival mechanisms to selective pressures and implications
- Up-regulation of key glycolysis proteins in cancer development
- Communications
- In vitro plant regeneration of Zenia insignis Chun
- DNA barcoding of online herbal supplements: crowd-sourcing pharmacovigilance in high school
- Case Reports
- Management of myasthenia gravis during pregnancy: A report of eight cases
- Three Cases of Extranodal Rosai-Dorfman Disease and Literature Review
- Letters to the Editor
- First report of Chlamydia psittaci seroprevalence in black-headed gulls (Larus ridibundus) at Dianchi Lake, China
- Special Issue on Agricultural and Biological Sciences - Part II
- Chemical composition of essential oil in Mosla chinensis Maxim cv. Jiangxiangru and its inhibitory effect on Staphylococcus aureus biofilm formation
- Secondary metabolites of Antarctic fungi antagonistic to aquatic pathogenic bacteria
- Study of Seizure-Manifested Hartnup Disorder Case Induced by Novel Mutations in SLC6A19
- Transcriptome analysis of Pinus massoniana Lamb. microstrobili during sexual reversal
- Mechanism of oxymatrine-induced human esophageal cancer cell apoptosis by the endoplasmic reticulum stress pathway
- Methylation pattern polymorphism of cyp19a in Nile tilapia and hybrids
- A Method of Biomedical Information Classification based on Particle Swarm Optimization with Inertia Weight and Mutation
- A novel TNNI3 gene mutation (c.235C>T/ p.Arg79Cys) found in a thirty-eight-year-old women with hypertrophic cardiomyopathy
- Remote Sensing-Based Extraction and Analysis of Temporal and Spatial Variations of Winter Wheat Planting Areas in the Henan Province of China
- Topical Issue on Precision Medicine
- Serum sTREM-1, PCT, CRP, Lac as biomarkers for death risk within 28 days in patients with severe sepsis
- IL-17 gene rs3748067 C>T polymorphism and gastric cancer risk: A meta-analysis
- Efficacy of Danhong injection on serum concentration of TNF-α, IL-6 and NF-κB in rats with intracerebral hemorrhage
- An ensemble method to predict target genes and pathways in uveal melanoma
- Evaluation of the quality of CT images acquired with smart metal artifact reduction software
- NPM1A in plasma is a potential prognostic biomarker in acute myeloid leukemia
- Arterial infusion of rapamycin in the treatment of rabbit hepatocellular carcinoma to improve the effect of TACE
- New progress in understanding the cellular mechanisms of anti-arrhythmic drugs
Articles in the same Issue
- Research Article
- Purification of Tea saponins and Evaluation of its Effect on Alcohol Dehydrogenase Activity
- Runt-related transcription factor 3 promoter hypermethylation and gastric cancer risk: A meta-analysis
- Risk Factors for Venous Thromboembolism in Hospitalized Patients in the Chinese Population
- Value of Dual-energy Lung Perfusion Imaging Using a Dual-source CT System for the Pulmonary Embolism
- A new combination of substrates: biogas production and diversity of the methanogenic microorganisms
- mTOR modulates CD8+ T cell differentiation in mice with invasive pulmonary aspergillosis
- Direct Effects on Seed Germination of 17 Tree Species under Elevated Temperature and CO2 Conditions
- Role of water soluble vitamins in the reduction diet of an amateur sportsman
- Aberrant DNA methylation involved in obese women with systemic insulin resistance
- 16S ribosomal RNA-based gut microbiome composition analysis in infants with breast milk jaundice
- Characterization of Haemophilus parasuis Serovar 2 CL120103, a Moderately Virulent Strain in China
- MiRNA-145 induces apoptosis in a gallbladder carcinoma cell line by targeting DFF45
- Telmisartan induces osteosarcoma cells growth inhibition and apoptosis via suppressing mTOR pathway
- Optimizing the Formulation for Ginkgolide B Solid Dispersion
- Determination of the In Vitro Gas Production and Potential Feed Value of Olive, Mulberry and Sour Orange Tree Leaves
- Factors Influencing the Successful Isolation and Expansion of Aging Human Mesenchymal Stem Cells
- The Value of Diffusion-Weighted Magnetic Resonance Imaging in Predicting the Efficacy of Radiation and Chemotherapy in Cervical Cancer
- Chemical profile and antioxidant activity of Trollius europaeus under the influence of feeding aphids
- SSR Markers Suitable for Marker Assisted Selection in Sunflower for Downy Mildew Resistance
- A Fibroblast Growth Factor Antagonist Peptide Inhibits Breast Cancer in BALB/c Mice
- Antihyperglycemic and antihyperlipidemic effects of low-molecular-weight carrageenan in rats
- Microbial indicators and environmental relationships in the Umhlangane River, Durban, South Africa
- TUFT1 promotes osteosarcoma cell proliferation and predicts poor prognosis in osteosarcoma patients
- Long non-coding RNA-2271 promotes osteogenic differentiation in human bone marrow stem cells
- The prediction of cardiac events in patients with acute ST segment elevation myocardial infarction: A meta–analysis of serum uric acid
- Risk expansion of Cr through amphibious clonal plant from polluted aquatic to terrestrial habitats
- Overexpression of Zinc Finger Transcription Factor ZAT6 Enhances Salt Tolerance
- Sini decoction intervention on atherosclerosis via PPARγ-LXRα-ABCA1 pathway in rabbits
- Soluble myeloid triggering receptor expressed on myeloid cell 1 might have more diagnostic value for bacterial ascites than C-reactive protein
- A Preliminary Study on the Newly Isolated High Laccase-producing Fungi: Screening, Strain Characteristics and Induction of Laccase Production
- Hydrolytic Enzyme Production by Thermophilic Bacteria Isolated from Saudi Hot Springs
- Analysis of physiological parameters of Desulfovibrio strains from individuals with colitis
- Emodin promotes apoptosis of human endometrial cancer through regulating the MAPK and PI3K/ AKT pathways
- Down-regulation of miR-539 indicates poor prognosis in patients with pancreatic cancer
- Inhibitory activities of ethanolic extracts of two macrofungi against eggs and miracidia of Fasciola spp.
- PAQR6 expression enhancement suggests a worse prognosis in prostate cancer patients
- Characterization of a potential ripening regulator, SlNAC3, from Solanum lycopersicum
- Expression of Angiopoietin and VEGF in cervical cancer and its clinical significance
- Umbilical Cord Tissue Derived Mesenchymal Stem Cells Can Differentiate into Skin Cells
- Isolation and Characterization of a Phage to Control Vancomycin Resistant Enterococcus faecium
- Glycogen Phosphorylase Isoenzyme Bb, Myoglobin and BNP in ANT-Induced Cardiotoxicity
- BAG2 overexpression correlates with growth and poor prognosis of esophageal squamous cell carcinoma
- Relationship between climate trends and grassland yield across contrasting European locations
- Review Articles
- Mechanisms of salt tolerance in halophytes: current understanding and recent advances
- Salivary protein roles in oral health and as predictors of caries risk
- Nanoparticles as carriers of proteins, peptides and other therapeutic molecules
- Survival mechanisms to selective pressures and implications
- Up-regulation of key glycolysis proteins in cancer development
- Communications
- In vitro plant regeneration of Zenia insignis Chun
- DNA barcoding of online herbal supplements: crowd-sourcing pharmacovigilance in high school
- Case Reports
- Management of myasthenia gravis during pregnancy: A report of eight cases
- Three Cases of Extranodal Rosai-Dorfman Disease and Literature Review
- Letters to the Editor
- First report of Chlamydia psittaci seroprevalence in black-headed gulls (Larus ridibundus) at Dianchi Lake, China
- Special Issue on Agricultural and Biological Sciences - Part II
- Chemical composition of essential oil in Mosla chinensis Maxim cv. Jiangxiangru and its inhibitory effect on Staphylococcus aureus biofilm formation
- Secondary metabolites of Antarctic fungi antagonistic to aquatic pathogenic bacteria
- Study of Seizure-Manifested Hartnup Disorder Case Induced by Novel Mutations in SLC6A19
- Transcriptome analysis of Pinus massoniana Lamb. microstrobili during sexual reversal
- Mechanism of oxymatrine-induced human esophageal cancer cell apoptosis by the endoplasmic reticulum stress pathway
- Methylation pattern polymorphism of cyp19a in Nile tilapia and hybrids
- A Method of Biomedical Information Classification based on Particle Swarm Optimization with Inertia Weight and Mutation
- A novel TNNI3 gene mutation (c.235C>T/ p.Arg79Cys) found in a thirty-eight-year-old women with hypertrophic cardiomyopathy
- Remote Sensing-Based Extraction and Analysis of Temporal and Spatial Variations of Winter Wheat Planting Areas in the Henan Province of China
- Topical Issue on Precision Medicine
- Serum sTREM-1, PCT, CRP, Lac as biomarkers for death risk within 28 days in patients with severe sepsis
- IL-17 gene rs3748067 C>T polymorphism and gastric cancer risk: A meta-analysis
- Efficacy of Danhong injection on serum concentration of TNF-α, IL-6 and NF-κB in rats with intracerebral hemorrhage
- An ensemble method to predict target genes and pathways in uveal melanoma
- Evaluation of the quality of CT images acquired with smart metal artifact reduction software
- NPM1A in plasma is a potential prognostic biomarker in acute myeloid leukemia
- Arterial infusion of rapamycin in the treatment of rabbit hepatocellular carcinoma to improve the effect of TACE
- New progress in understanding the cellular mechanisms of anti-arrhythmic drugs