Der Einfluss von Fortbewegung auf die sensorische Informationsverarbeitung und die zugrunde liegenden neuronalen Schaltkreise
-
Laura Busse
Laura Busse ist seit 2016 Professorin für „Organismische Neurobiologie” in der Abteilung Neurobiologie des Departments für Biologie II an der LMU München und ist mit dem Bernstein Zentrum für Computational Neuroscience affiliert. Sie promovierte an der Universität Göttingen im Rahmen ihrer Arbeit am Deutschen Primatenzentrum, war Postdoktorandin am Smith Kettlewell Eye Research Institute in San Francisco, USA, und am UCL Institute of Ophthalmology in London. Zwischen 2010 and 2015 leitete Laura Busse eine Juniorgruppe am Zentrum für Integrative Neurowissenschaften (CIN) der Universität Tübingen.


Zusammenfassung
Die Verarbeitung sensorischer Information kann sowohl im Kortex als auch im Thalamus durch den Verhaltenskontext, z. B. durch aktive Fortbewegung, moduliert werden. Solch aktives Verhalten verbessert die Kodierung sensorischer Reize und die Wahrnehmung, besonders während Aktivitäten von moderater Intensität. Der Modulation sensorischer Verarbeitung durch Fortbewegung scheint eine Kombination von Mechanismen zugrunde zu liegen, unter anderem neuromodulatorische Einflüsse, die Aktivität spezifischer, inhibitorischer Interneurone, sowie top-down- oder motorische Rückprojektionen. Neue experimentelle Ansätze, die es Mäusen erlauben, sich trotz Kopffixation auf Laufbällen oder –bändern fortzubewegen, ermöglichte es in den letzten Jahren, die neuronalen Schaltkreise und zellulären Elemente, die der Modulation durch Verhaltenskontext zugrunde liegen, eingehend zu untersuchen. Dieser Übersichtsartikel fasst den momentanen Stand dieser Studien zusammen und beleuchtet wichtige offenen Fragen.
Einleitung
Die Untersuchung der neuronalen Grundlagen sensorischer Wahrnehmung beruht auf einer lange Tradition von wichtigen experimentellen Ansätzen, in denen anästhesierten oder wachen, kopffixierten Tiere hochkontrollierte sensorische Reize dargeboten werden. Es war genau diese außerordentliche experimentelle Kontrolle, die es ermöglichte, rezeptive Felder (RFs) in visuellen, auditorischen, somatosensorischen und anderen sensorischen Systemen im Detail zu charakterisieren. Die Entwicklung von solch hochkontrollierten Verhaltensaufgaben für kopffixierte, nicht-humane Primaten (Roelfsema et al., 1998; Spitzer et al., 1988; Treue und Maunsell, 1996), Frettchen (Atiani et al., 2009; David et al., 2012) oder Nager (Histed et al., 2012; Schwarz et al., 2011) erlaubte es, in sensorischen Arealen nicht-sensorische, kontextabhängige Modulationen der neuronalen Antworten zu isolieren, indem identische sensorische Stimulation unter verschiedenen kognitiven Bedingungen dargeboten wurde. Zur Isolierung dieser Modulationen durch kognitiven Kontext, z. B. der Effekte von Aufmerksamkeit, Erwartung von Belohnung, oder Entscheidungsfindung, was es entscheidend, diese Experimente in unbeweglichen Tieren durchzuführen, da dies die Stabilität sowohl der sensorischen Stimulation als auch der neuronalen Messungen gewährleistete.
Ganz generell ist es plausibel, dass die Untersuchung von Wahrnehmungsprozessen in stationärem Zustand besonders gut Prozesse in Primaten abbildet, wo ein beachtlicher Anteil der sensorischen Eindrücke mit fokussierter Informationsverarbeitung einhergeht, z. B. während der Kommunikation, Lesen (im Falle von Menschen), und anderen Formen von Informationsaufnahme unter gerichteter Aufmerksamkeit. Für viele Lebewesen jedoch findet wichtige sensorische Informationsverarbeitung hauptsächlich während körperlicher Aktivität statt. Die Sinnesverarbeitung dient hier der Navigation, trägt dazu bei, Futter und Paarungspartner zu finden, und hilft, Fressfeinden zu entkommen; Ruheperioden werden hingegen oft mit Erholung und Regeneration in Verbindung gebracht. Bemerkenswerterweise scheinen sich einige Aspekte von neuronaler Verarbeitung während aktiven Verhaltens und gerichteter Aufmerksamkeit zu ähneln (Harris und Thiele, 2011; Maimon, 2011), was die Möglichkeit offen lässt, dass zumindest einige der zugrunde liegenden neuronalen Schaltkreise im Verlauf der Evolution konserviert sein könnten.
Die Idee, sensorische Verarbeitung während der Fortbewegung zu untersuchen, ist nicht neu. Im Bereich der Sensomotorik wird seit vielen Jahren mit einem experimentellen Ansatz gearbeitet, der die oben erwähnten Vorteile der Kopffixation mit der Möglichkeit verbindet, sich relativ frei zu bewegen. Der Weg für diesen Ansatz wurde im Max-Plank-Institut für Biologische Kybernetik in Tübingen bereitet, wo er zunächst für bahnbrechende Arbeiten am visuellen System der Fliege benutzt wurde (Buchner, 1976) (Abbildung 1a). Das experimentelle Setup bestand hier aus einem luftgepolsterten Ball, auf dem die Fliege sitzen oder laufen kann, umgeben von einer kuppelförmigen Leinwand, auf der visuelle Reize in Abhängigkeit vom Fortbewegungsverhalten (closed loop) oder unabhängig vom Fortbewegungsverhalten des Tieres (open loop) präsentiert werden. Als Mallot und Kollegen an der Universität Tübingen die experimentellen Möglichkeiten erkannten, die dieser Laufball ermöglichte, passten sie dieses System vor über zehn Jahren an die Arbeit mit Ratten an (Holscher et al., 2005) und konnten zeigen, dass Ratten erfolgreich in einer virtuellen Umgebung innerhalb dieses Systems navigieren können. Die spätere Kombination des Systems mit Zwei-Photonen Ca2+ Imaging von neuronalen Populationen mit Einzelzellauflösung (Dombeck et al., 2007), intrazellulären Messungen (Harvey et al., 2009) oder extrazellulären Ableitungen mit Multi-Kanal-Elektroden (Niell und Stryker, 2010) in Mäusen, die sich aktiv in dem Setup verhielten, führte letztlich zum Durchbruch des Paradigmas (Abbildung 1b-c). Bemerkenswerterweise hat diese Forschungsrichtung zu der Frage, wie aktives Verhalten die neuronale Verarbeitung in Fliegen und Mäusen moduliert, neue Untersuchungen im Bereich der kognitiven Psychologie inspiriert, wo nun Setups entwickelt werden, die es erlauben, Leistungen in Wahrnehmungsaufgaben und Gehirnaktivität während kontrollierter körperlicher Aktivität zu messen, z. B. EEG-Messungen während des Radfahrens auf einem stationären Trainingsrad (Bullock et al., 2016).
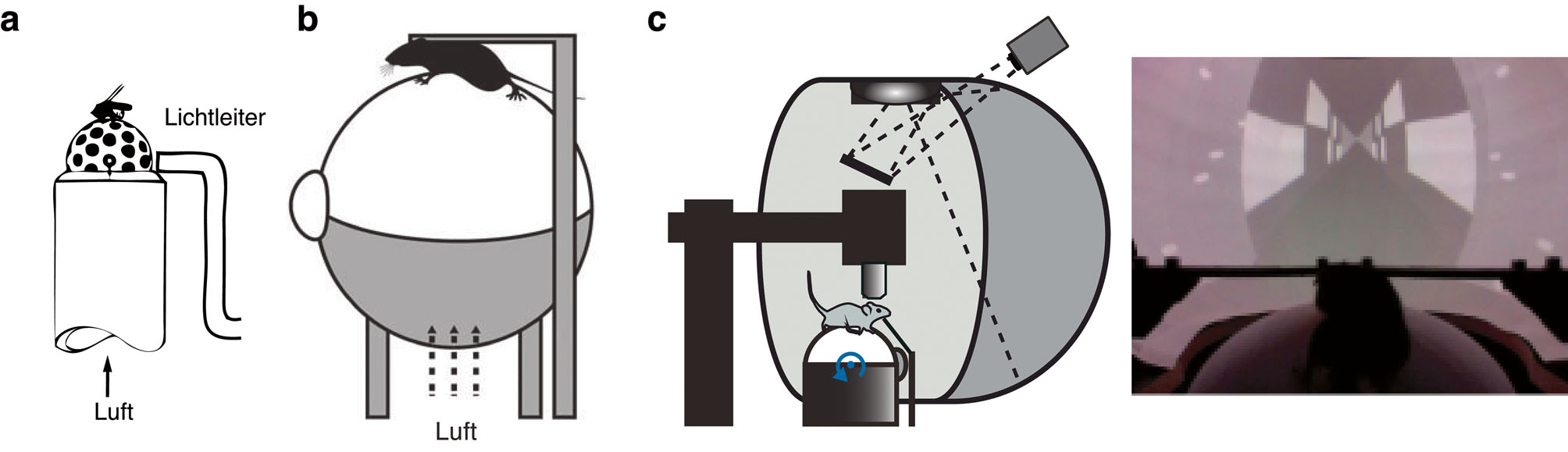
Fortbewegung während der Kopffixation auf Laufball- oder -bandsystemen. a) Eines der ersten “luftgepolsterten Ball-Setups”, das für die Fliege Drosophila entwickelt wurde, die hier auf dem Ball über einen Mikromanipulator fixiert ist. Ein System basierend auf Lichtleitfasern nimmt die Bewegung des Tieres auf, die über die Verschiebung der Muster auf dem Ball gemessen wird – mit freundlicher Genehmigung modifiziert aus Buchner (1976). b) Ein ähnliches System für Mäuse – mit freundlicher Genehmigung modifiziert aus Carandini und Churchland (2013). c) In Kombination mit einem großflächigen, lebensechtem Stimulationssystem kann der visuelle Stimulus mit geschlossener Rückkopplung (closed loop, entsprechend der Bewegung des Tieres) oder mit offener Rückkopplung (open loop, nicht in Übereinstimmung mit der Bewegung des Tieres) präsentiert werden – mit freundlicher Genehmigung modifiziert aus Fiser et al. (2016).
Modulation sensorischer Informationsverarbeitung
Mehrere Forschungsgruppen haben beobachtet, als sie kopffixierte Mäuse während der Fortbewegung mit einer Kamera zur Bestimmung von Augenbewegungen überwacht haben, dass Fortbewegung in Mäusen mit einer deutlichen Modulation der Pupillengröße einhergeht: Während die Pupille tendenziell kleiner ist, wenn das Tier stationär ist, kann die Pupille während der Fortbewegung dramatisch vergrößert sein (Erisken et al., 2014; Vinck et al., 2015; McGinley et al., 2015; Reimer et al., 2014). Die Pupillengröße ist seit Langem aus der kognitiven Psychologie als ein hervorragendes Maß für Erregung (engl. „arousal“) bekannt (Bradshaw, 1967); daher wird seit einiger Zeit die Erweiterung der Pupille bei konstanter Beleuchtung in Mäusen dazu benutzt, die Effekte von Arousal zu isolieren (Vinck et al., 2015; McGinley et al., 2015; Reimer et al., 2014), die normalerweise mit spezifischeren, fortbewegungsbezogenen Signalen während des Laufens vermischt sind (Keller et al., 2012).
Die ökologische Relevanz der Pupillenerweiterung mit Arousal, oder mit anspruchsvollen kognitiven Prozessen im Generellen, ist eine offene Frage: einerseits führt die Pupillenerweiterung zu einer Erhöhung der Lichtmenge, die in das Auge einfällt. Dies kann, z. B. bei Mäusen, zu einer mehr als 20-fachen Steigerung der retinalen Helligkeit führen (Pennesi et al., 1998). Zu welchem Grad eine solche Erhöhung der retinalen Helligkeit zu einer Erhöhung der visuellen Sensitivität führt, ist jedoch unklar, da es bereits auf der Verarbeitungsebene der Netzhaut neuronale Schaltkreise gibt, die für die Kontrolle der Signalverstärkung (engl. „gain“) und Normalisierung der Aktivität anhand der Gesamt-Luminanz zuständig sind (Carandini und Heeger, 2012). Andererseits ist die Pupillengröße für unterschiedliche Beleuchtungsintensitäten bereits dafür optimiert, die größtmögliche Sehschärfe zu gewährleisten (Laughlin, 1992); eine zusätzliche Erweiterung der Pupille im Kontext von Arousal könnte daher sogar die Bildqualität aufgrund von optischen Aberrationen verschlechtern. Zusammenfassend lässt sich sagen, dass die funktionelle Relevanz der Pupillenerweiterung mit Arousal für die visuelle Wahrnehmung noch unklar ist, da direkte Evidenz für einen Beitrag der Pupillengröße auf die visuelle Sensitivität noch aussteht.
Visueller Kortex
Niell und Stryker (2010) haben als erste den Laufball benutzt, um – aufbauend auf einer langen Tradition von Arbeiten über zustandsabhängige, neuronale Verarbeitung in sensorischen Systemen (Harris und Thiele, 2011; Gilbert und Sigman, 2007; Alonso und Swadlow, 2017; Zagha und McCormick, 2014) – in extrazellulären Ableitungen zu zeigen, dass neuronale Aktivität im primären visuellen Kortex (V1) der Maus stark davon abhängt, ob das Tier sitzt oder rennt. Während der Fortbewegung können Nervenzellen in Schicht 2/3 des primären visuellen Kortex ihre Antwortempfindlichkeit mehr als verdoppeln. Diese Steigerung besteht typischerweise aus einer additiven und multiplikativen Komponente (Abbildung 2a, b): Tuningkurven für Orientierung in V1 werden beispielsweise während der Fortbewegung hochskaliert, ohne dass sich die Tuningbreite, zumindest im Populationsdurchschnitt, ändert. Spätere Untersuchungen konnten zeigen, dass der Mechanismus auf Ebene des Membranpotenzials, der diesen aktivitätsabhängigen Unterschieden in der V1 – Verarbeitung zugrunde liegt, eine Depolarisation der Pyramidenzellen ist (Bennett et al., 2013; Polack et al., 2013). Diese Depolarisation des Membranpotenzials wird von einer bemerkenswerten Reduktion der Trial-zu-Trial Variabilität begleitet (Bennett et al., 2013), die dazu beiträgt, die Zuverlässigkeit der visuell evozierten Antwort zu verbessern.
Neben der Erhöhung des Orientierungstunings (Niell und Stryker, 2010; Polack et al., 2013) und der kontrastabhängigen Antworten (Erisken et al., 2014; Lee et al., 2014) kann Fortbewegung auch verschiedene Aspekte der Reizselektivität in V1 verändern. Erstens, in Experimenten, in denen die Antworten von V1 – Nervenzellen mit unterschiedlich großen Gittermustern charakterisiert werden, erhöht die Fortbewegung die bevorzugte Stimulusgröße und reduziert merklich die suppressiven Einflüsse der Stimulusumgebung („surround suppression“); Fortbewegung steuert daher die räumliche Integration und Modulation durch Stimuluskontext (Abbildung 2c) (Erisken et al., 2014; Ayaz et al., 2013). Zweitens scheinen die fortbewegungsinduzierten Erhöhungen der Antwortstärke für die Nervenzellen am größten zu sein, die hohe Raumfrequenzen präferieren, was zu einer relativen Steigerung der räumlichen Auflösung während der Fortbewegung führt (Mineault et al., 2016). Drittens, verschiebt sich während der Fortbewegung das zeitliche Tuning von Nervenzellen in V1 und dem extrastriatären Areal AL (anterolateral) zu höheren zeitlichen Frequenzen (Andermann et al., 2011). Viertens führt ein hohes „Arousal“ (gemessen als Pupillenerweiterung) in Abwesenheit von Fortbewegung zu einer Verschärfung des Orientierungstunings (Reimer et al., 2014). Diese Veränderungen in der V1 – Stimulus-Selektivität machen einfache Erklärungen für die beobachteten Effekte von Fortbewegung unwahrscheinlich, wie z. B. Erklärungen basierend auf einer Erhöhung der Gehirntemperatur (Moser et al., 1993). Da die Schwankungen der kortikalen Temperatur mit willentlicher Fortbewegung darüber hinaus gering zu sein scheinen (0.1 °C), haben einige Autoren geschlossen, dass der Beitrag von Temperaturveränderungen während der Fortbewegung auf neuronale Erregbarkeit minimal sind (Shirey et al., 2015).
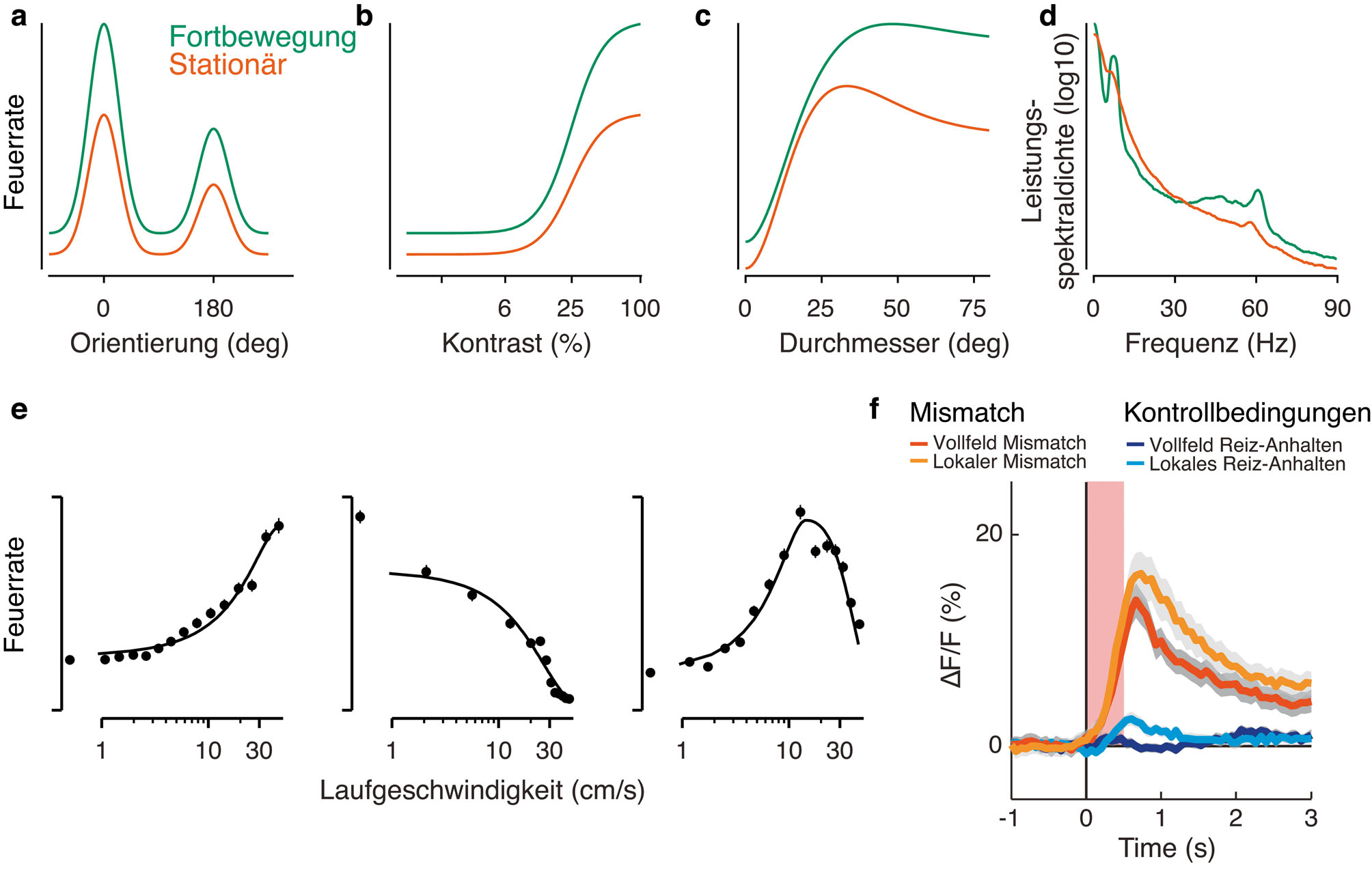
Einflüsse von Fortbewegung auf einige V1 – Antworteigenschaften. a) Fortbewegung erhöht Orientierungstuningkurven. b) Fortbewegung erhöht den Response-Gain von kontrastabhängigen Antworten. c) Fortbewegung verändert die räumliche Integration: Die bevorzugte Größe der rezeptiven Felder wird erhöht und die Einflüsse von Surround Suppression reduziert (a-c) – mit freundlicher Genehmigung modifiziert aus Erisken et al. (2014). d) Fortbewegung unterdrückt die Amplitude der niedrigen LFP – Frequenzen und erhöht die Amplitude der höheren Frequenzen, unter anderem eines schmalen Frequenzbandes im Gamma – Bereich (Fiorini und Busse, unpublished). e) Beispiel Nervenzellen in V1 mit Tuning für Laufgeschwindigkeit (Erisken und Busse, unpublished; Saleem et al., 2013). f) Vollfeld und lokale Mismatch-Antworten von einer Population von V1 – Nervenzellen – mit freundlicher Genehmigung modifiziert aus Zmarz und Keller (2016).
Über die Verstärkung der Antworten hinaus verbessert Fortbewegung die Kodierung von Reizen, indem die gegenseitige Information (engl. „mutual information“) zwischen visuellen Reizen und Einzelzellantworten verbessert wird (Dadarlat und Stryker, 2017). Die bewegungsabhängige Verschiebung der V1 – Population hin zu informativeren Verarbeitungsregimes ergibt sich nicht nur aus der Erhöhung der Antwortstärke (Dadarlat und Stryker, 2017), sondern auch durch Veränderungen der Antwortmuster in der Population, wo vom visuellen Reiz unabhängige, gemeinsame Fluktuationen der Antworten (engl. „noise correlations“) verringert werden (Erisken et al., 2014; Dadarlat und Stryker, 2017).
Zumindest im visuellen System der Maus scheinen die Effekte von Fortbewegung schichtabhängig zu sein. Es konnte gezeigt werden, dass die bewegungs-abhängige Erhöhung von gegenseitiger Information (Dadarlat und Stryker, 2017) und der Feuerraten (Erisken et al., 2014; Dadarlat und Stryker, 2017) am größten und konsistentesten in den supragranulären kortikalen Schichten ist. Veränderungen der Feuerrate mit Fortbewegung in infragranulären Schichten hingegen, sind diverser und haben oftmals sogar unterschiedliche Vorzeichen, was zu einer geringeren Gesamtmodulation führt (Erisken et al., 2014). Die Mechanismen und die der Diversität zugrunde liegenden Schaltkreise für bewegungsabhängige Modulation in tiefen kortikalen Schichten sind derzeit nicht gut verstanden. Zwei Erklärungsansätze wurden gemacht: Eine 2-Photonen-Kalzium-Imaging-Studie mit zellulärer Auflösung, in der mithilfe eines implantierten Mikroprismas auch die tiefen Schichten von V1 für das Imaging zugänglich gemacht wurden, konnte eine interessante Population von Nervenzellen zeigen, deren Aktivität stark durch Fortbewegung unterdrückt wurde (Andermann et al., 2013). Diese Zellen wurden aufgrund ihrer Lage in der Imaging-Ebene der Schicht 6 zugeschrieben (Andermann et al., 2013). Interessanterweise hatten frühere Studien gezeigt, dass eine bestimmte Klasse von L6 – Nervenzellen (die Ntsr1-positiven L6 – Nervenzellen) den Gain innerhalb der V1 – Kolumne (Olsen et al., 2012) durch reduziertes Antreiben eines vermutlich transkolumnaren inhibitorischen Interneurons regulieren können (Bortone et al., 2014). Eine Unterdrückung dieser L6 – Zellen durch Fortbewegung könnte somit zu einer Erhöhung der Aktivität in den oberen V1 – Schichten führen. Andere Befunde deuten darauf hin, dass die Nervenzellen in den tiefen kortikalen Schichten, deren Aktivität durch Bewegung unterdrückt wird, in Schicht 5 (L5) liegen (Erisken et al., 2014); falls es sich bei diesen Nervenzellen um Somatostatin-positive (SOM+) inhibitorische Interneurone handeln würde, könnte ihre Unterdrückung zu Erhöhung von Nervenzellaktivität in den anderen Schichten via Disinhibition von Pyramidenzellenaktivität führen. Zukünftige Experimente werden benötigt, um zwischen diesen und weiteren Alternativen zu unterscheiden, und die Nervenzellen in den tiefen Schichten, deren Aktivität durch Bewegung unterdrückt wird, eindeutig zu identifizieren.
Übereinstimmend mit einer Erhöhung des generellen Aktivierungszustand des Gehirns gehen Zeitabschnitte mit Fortbewegung mit einer deutlichen Veränderung in der Leistungsspektraldichte des lokalen Feldpotenzials (LFP) einher (Abbildung 2d). Insbesondere verringert die Fortbewegung die Leistungsspektraldichte des LFPs in niedrigen Frequenzen (< 20 Hz) und erhöht die Leistungsspektraldichte im Gammabandbereich (> 40 Hz) (Niell und Stryker, 2010; Vinck et al., 2015). Dadurch verändert sich das LFP von einem Signal, das von Fluktuationen mit niedrigen Frequenzen und hohen Amplituden während stationärer Perioden bestimmt wird, zu einem Signal während der Bewegung, das mehr desynchronisierte Aktivitätsmuster aufweist. Interessanterweise tritt im primären visuellen Kortex der Maus auch eine auffällig schmale Gammaband-Oszillation mit einer Bandbreite von 2–5 Hz auf, die nahe bei 60 Hz liegt und deren Amplitude durch Fortbewegung erhöht wird (Niell und Stryker, 2010; Lee et al., 2014; Saleem et al., 2017). Diese schmalbandige Gamma-Oszillation ist in Schicht 4 (L4) des primären visuellen Kortex am höchsten und kann auch in den Ausgangsimpulsen der Nervenzellen des Corpus geniculatum laterale des Thalamus (dLGN) gemessen werden, selbst wenn direkte Feedback-Verbindungen aus dem Kortex ausgeschaltet werden (Saleem et al., 2017). Dies deutet darauf hin, dass die schmalbandige Gamma-Oszillation vorgelagert vor dem Kortex generiert wird, also im Thalamus oder möglicherweise bereits in der Netzhaut (Storchi et al., 2017). Während die genaue Funktion der schmalbandigen Gamma-Oszillation noch unbekannt ist, könnte sie möglicherweise einen spezifischen Kanal für die retino-genikulo-kortikale Kommunikation darstellen (Saleem et al., 2017).
Zusätzlich zu globalen Veränderungen im Erregungszustand des Gehirns kann die Fortbewegung auch sehr spezifische Einflüsse im primären visuellen Kortex ausüben. Erstens scheint ein beträchtlicher Teil der V1 – Nervenzellen in der Maus ein Tuning für die Laufgeschwindigkeit zu haben, selbst wenn die Tiere in völliger Dunkelheit laufen (Erisken et al., 2014; Saleem et al., 2017): Die Laufgeschwindigkeit kann die Feuerrate von V1 – Nervenzellen regulieren, wobei das Tuning für Laufgeschwindigkeit monoton ansteigend oder monoton abfallend sein kann, oder bei mittleren Laufgeschwindigkeiten gipfeln kann (Abbildung 2e). Eine solche Selektivität für Laufgeschwindigkeit ist konsistent mit der Hypothese, dass V1 zur Navigation beitragen kann, sogar in Abwesenheit von optischem Fluss. Zweitens können 5-10% der Nervenzellen im primären visuellen Kortex der Maus Diskrepanzen zwischen der Eigenbewegung und dem optischen Fluss signalisieren (Keller et al., 2012; Zmarz und Keller, 2016): Sie antworten stark auf eine plötzliche Entkopplung des optischen Flussmusters von der Selbstbewegung (Abbildung 2f). Im Rahmen von Theorien der prädiktiven Kodierung (engl. „predictive coding“) (Friston, 2005; Rao und Ballard, 1999) können solche „mismatch“ Antworten dahingehend interpretiert werden, dass sie Abweichungen von einer Vorhersage signalisieren, die auf einer gelernten Beziehung zwischen motorischem Output und sensorischem Feedback beruht (Attinger et al., 2017). Interessanterweise können solche „mismatch“ Antworten auch von lokalen Veränderungen im optischen Fluss ausgelöst werden (Abbildung 2f), wodurch diese Antworten einen interessanten Mechanismus darstellen könnten, Abweichungen von der Selbstbewegung festzustellen, die z. B. durch sich bewegende Objekte während der Eigenbewegung entstehen (Zmarz und Keller, 2016).
Andere sensorische Kortizes
Modulationen von kortikalen Antworten durch Fortbewegung wurden bislang am intensivsten im visuellen System untersucht, aber sie treten auch im somatosensorischen und auditorischen System auf. Im primären somatosensorischen Kortex (S1) scheinen die Effekte von Fortbewegung im Allgemeinen mit denen vergleichbar zu sein, die im primären visuellen Kortex beobachtet werden können (Fu et al., 2014; Sofroniew et al., 2015; Pluta et al., 2015). Außerdem ähneln sie den Effekten, die typischerweise während der aktiven Bewegung der Vibrissen bei Nagern beobachtet werden, ein Maß, das in Untersuchungen des Tastsinns häufig als Index für aktive Verhaltenszustände benutzt wurde (Crochet und Petersen, 2006; Gentet et al., 2010; Poulet und Petersen, 2008). Im primären Hörkortex (A1) scheinen die beobachteten Effekte hingegen deutlich unterschiedlich zu sein. Anstatt die Antworten zu verstärken, scheint die Fortbewegung Antworten in A1 zu verringern (McGinley et al., 2015; Zhou et al., 2014; Schneider et al., 2014), und zwar sowohl während der Spontanaktivität als auch während der reizgetriebenen Aktivität. Diese Unterdrückung der A1 Feuerrate scheint durch eine Herunterregelung sowohl der erregenden als auch der hemmenden Einflüsse auf das Membranpotential vermittelt zu werden (McGinley et al., 2015; Zhou et al., 2014). Da die relative Unterdrückung während der spontanen Hintergrundaktivität stärker als während der sensorisch evozierten Antworten ist, ist das Signal-Rausch-Verhältnis während der auditorischen Verarbeitung im aktiven Zustand verbessert (McGinley et al., 2015; Zhou et al., 2014), insbesondere in Regimen von moderater Erregung (McGinley et al., 2015). Interessanterweise scheint nicht nur die Fortbewegung, sondern auch aktive Einbindung in eine auditorische Aufgabe den primären auditorischen Kortex im Allgemeinen zu hyperpolarisieren (Otazu et al., 2009; Kuchibhotla et al., 2017).
Effekte von Fortbewegung im Thalamus
Effekte von Fortbewegung, genau wie die Effekte von Arousal im Generellen (Cano et al., 2006; Bezdudnaya et al., 2006), sind nicht auf den Kortex beschränkt, sondern treten bereits auf der Ebene der sensorischen thalamischen Kerngebiete auf (Erisken et al., 2014, McGinley et al., 2015; Williamson et al., 2015). Im visuellen System verschiebt die Fortbewegung den Feuermodus der Nervenzellen im Corpus geniculatum laterale (dLGN) des Thalamus von einem „Burst“-Modus zum „Tonic“-Modus (Niell und Stryker, 2010; Erisken et al., 2014), erhöht die Feuerraten (Erisken et al., 2014; Williamson et al., 2015) und steuert die räumliche Integration (Erisken et al., 2014). Ähnlich wie Nervenzellen in V1 können thalamische Nervenzellen geschwindigkeitsbezogene Informationen signalisieren (Erisken et al., 2014, Roth et al., 2016): dLGN Neurone signalisieren mit ihrer Feuerrate die Laufgeschwindigkeit des Tieres (Erisken et al., 2014), während Nervenzellen in dem höheren visuellen Thalamuskern Nucleus lateralis posterior, ein Nager-Homolog des Pulvinars, Diskrepanzen zwischen Selbstbewegung und optischem Fluss repräsentieren können (Roth et al., 2016). Im auditorischen System verringert Fortbewegung die neuronalen Antworten bereits im Corpus geniculatum mediale des Thalamus (McGinley et al., 2015; Williamson et al., 2015).
Angesichts der Tatsache, dass Fortbewegung die sensorische Verarbeitung über mehrere Verarbeitungsstufen hinweg moduliert, ist es eine wichtige offene Frage, inwieweit diese Modulationen über Verarbeitungsstufen hinweg durch glutamaterge Verbindungen übertragen oder denovo erzeugt werden. Im auditorischen System haben Schneider et al. (2014) den kortikalen Beitrag zu fortbewegungsbezogener Antwortmodulation mithilfe eines leistungsstarken optogenetischen Stimulationsansatzes geschätzt: Sie stellten fest, dass die Modulation durch Fortbewegung während der optogenetischen Stimulation von afferenten thalamo-kortikalen Verbindungen aus dem thalamischen Korpus geniculatum mediale in A1 nur etwa 60% der Gesamtmodulation der Antworten auf Töne ausmachte. Dies ist starke Evidenz dafür, dass sowohl hintereinandergeschaltete Kaskadeneffekte zwischen Verarbeitungsstufen als auch kortikalen Beiträge eine bedeutsame Rolle für bewegungsabhängige Modulationen in A1 spielen. Ähnliche Ergebnisse wurden in Studien des somatosensorischen Systems gefunden, die sich allgemeiner mit erregungsabhängiger Verarbeitung beschäftigt haben, und in denen die Stimulation des Nucleus ventroposteriomedialis thalami (VPM) neuronale Aktivitätsmuster in S1 hervorrufen konnte, die normalerweise mit aktiver Bewegung der Vibrissen gekoppelt sind (Hirata und Castro-Alamancos, 2010; Poulet et al., 2012; siehe jedoch Constantinople und Bruno, 2011).
Effekte von Fortbewegung auf Verhaltensleistungen
Welche Auswirkungen haben die fortbewegungsbezogenen Verbesserungen der sensorischen Repräsentationen auf das Verhalten? Im visuellen System führt die verbesserte neuronale Kodierung während der Fortbewegung zu einer verbesserten Verhaltenssensitivität für Reizdetektion, insbesondere bei anspruchsvollen Bedingungen mit niedrigem visuellen Kontrast (Bennett et al., 2013). Bisher wurde die Aktivität des Tieres für Wahrnehmungsstudien im visuellen System weitgehend als eine binäre Variable betrachtet, bei der die Verhaltensleistung entweder im stationären Zustand oder im Fortbewegungszustand gemessen wurde. Arbeiten zur auditiven Wahrnehmung haben die Verhaltensleistungen auf viel feinkörnigere Weise quantifiziert und sind zum Schluss gekommen, dass die auditorische Detektionsleistungen der klassisch beschriebenen, umgekehrt-U-förmigen Abhängigkeit von der Erregung folgen (McGinley et al., 2015): Die Verhaltensleistung ist optimal für mittlere Erregungsstufen, definiert durch einen mittleren Pupillendurchmesser, die während eines Subzustands von ruhiger Wachheit (engl. „quiet wakefulness“) auftreten; im Gegensatz dazu ist die Hörleistung während des Rennens, also einem Zustand mit hoher Erregung, oder während der Schläfrigkeit, verschlechtert. Interessanterweise wurde kürzlich im menschlichen Sehsystem eine ähnliche, umgekehrt U-förmige Beziehung zwischen der Orientierungskodierung und der Bewegungsintensität beobachtet (Bullock et al., 2016).
Neuronale Schaltkreise für Modulationen von sensorischer Verarbeitung durch Fortbewegung
Zwei Mechanismen haben in jüngster Zeit besondere Aufmerksamkeit bei der Untersuchung der neuronalen Schaltkreise erhalten, die für die Fortbewegungseffekte auf die sensorische Verarbeitung zuständig sind: Neuromodulation und inhibitorische Interneurone. Diese Mechanismen sind wahrscheinlich verwandt, da inhibitorische Interneurone besonders sensitiv für Einflüsse von Neuromodulation zu sein scheinen (Fanselow et al., 2008; Chen et al., 2015; Kruglikov und Rudy, 2008).
Es gibt eine Vielzahl von zusammenlaufenden Arbeiten, welche die Rolle von Acetylcholin bei der Expression von zustandsabhängiger, neuronaler Verarbeitung demonstrieren (Thiele, 2013). Die Stimulation des basalen Vorderhirns kann in V1 einige der Haupteffekte von Fortbewegung imitieren, inklusive der Verbesserung der Sehleistung (Pinto et al., 2013). Für die Verbesserungen der V1 – Antworten, die typischerweise mit der Fortbewegung assoziiert sind, ist es tatsächlich ausreichend, die Axone zu aktivieren, die von der mesencephalischen lokomotorischen Region im Hirnstamm ausgehend (engl. „mesencephalic locomotor region“, MLR) im basalen Vorderhirn terminieren (Lee et al., 2014). Entscheidend ist, dass diese Rekapitulation von V1-Fortbewegungseffekten auch bei subtiler Stimulation möglich ist, bei der die absteigenden, lokomotorischen Ausgänge der mesencephalen Bewegungsregion nicht rekrutiert werden (Lee et al., 2014). In Übereinstimmung mit der Beobachtung, dass die Effekte von Fortbewegung sehr große Ähnlichkeit zu denen der basalen Vorderhirnstimulation aufweisen, korreliert die Aktivität der an der Oberfläche von V1 und A1 gemessenen cholinergen Axone mit dem Fortbewegungsmuster (Reimer et al., 2016). Andere Arbeiten konnten jedoch zeigen, dass die Anwendung von cholinergen Antagonisten in Schicht 2/3 die tonische Depolarisation, die in Zusammenhang mit der Fortbewegung auftritt, nicht aufheben konnte (Polack et al., 2013), was auf die Beteiligung zusätzlicher neuromodulatorischer Signalwege hindeutet. Angesichts der deutlichen Auswirkungen von Fortbewegung auf die Pupillengröße ist Norepinephrin ein weiterer, möglicherweise beteiligter Neuromodulator, der bekannt dafür ist, über tonische Aktivität im Locus coeruleus zur Kontrolle der Pupillengröße beizutragen (Aston-Jones und Cohen, 2005; Samuels und Szabadi, 2008). In der Tat scheint eine pharmakologische Blockade von noradrenergen Eingängen in oberflächlichen Schichten von V1 die fortbewegungsabhängige Depolarisation um etwa 50% zu reduzieren (Polack et al., 2013), und die Aktivität von noradrenergen Axonen an der Oberfläche von V1 scheint gut mit schnellen Veränderungen der Pupillengröße korreliert zu sein (Reimer et al., 2016). Während manchmal implizit davon ausgegangen wird, dass die Verbindungen zwischen neuromodulatorischen Zentren und Kortex direkt sind, hat die Neuromodulation auch erhebliche Auswirkungen auf den Thalamus und die thalamokortikale Reizübertragung: Erstens enden cholinerge Projektionen aus der parabrachialen Hirnstammregion, die auch den MLR umfasst, sowie noradrenerge Projektionen aus dem Locus coeruleus im dLGN [zusammengefasst in (Lee und Dan, 2012)]. Zweitens enden GABAerge basale Vorderhirnprojektionen im Nucleus reticularis thalami (TRN), wo sie hyperpolarisierende Einflüsse ausüben (Blickford et al., 1994), und dadurch möglicherweise sensorische Thalamuskerne disinhibiert werden. Zusammenfassend lässt sich feststellen, dass zwar in den letzten Jahren klar geworden ist, dass verschiedene Schaltkreise an der Modulation von sensorischer Informationsverarbeitung beteiligt sind, dass es aber eine bislang ungelöste Frage bleibt, wie die Einflüsse dieser Schaltkreise während der Fortbewegung koordiniert werden (Abbildung 3a).
Bemerkenswerterweise verändert Neuromodulation über das noradrenerge System während der Fortbewegung nicht nur neuronale Antworten, sondern führt auch zu langsameren und länger anhaltenden Ca2+ – Antworten im V1 – Astroglia-Netzwerk (Paukert et al., 2014; Ding et al., 2013). Erhöhte Kalziumsignale in Astrozyten während gleichzeitiger synaptischer Aktivität und Arousal könnte die Aufnahmefähigkeit der Astrozyten für K+ und Glutamat aus dem extrazellulären Raum erhöhen. Dies wiederum könnte zur Kontrolle der Erregbarkeit neuronalen Schaltkreise und der Koordination der breitgestreuten Einflüsse der Neuromodulatoren auf neuronale Aktivität beitragen (Paukert et al., 2014; Ding et al., 2013; Kjaerby et al., 2017).
Welches könnten mögliche Ziele dieser neuromodulatorischen Einflüsse sein? Im primären visuellen Kortex könnte Acetylcholin bevorzugt auf inhibitorische Interneurone wirken (Fanselow et al., 2008; Chen et al., 2015; Kruglikov und Rudy, 2008) und dadurch die Rekrutierung der GABAergen Hemmung und die Funktion des kortikalen Schaltkreises verändern. Ein besonders relevantes Verschaltungsmotiv scheint die cholinerge Aktivierung von VIP-positiven inhibitorischen Interneuronen sein, was über eine Hemmung von SOM-positiven inhibitorischen Interneuronen letztlich zu einer Disinhibierung der Pyramidenzellaktivität führt (Abbildung 3b). Dieses Verschaltungsmotiv wurde detailliert untersucht (Pfeffer et al., 2013) und mit unterschiedlichen Aspekten neuronaler Verarbeitung in Verbindung gebracht: von der Kontrolle der neuronalen Antwortstärke bis zur sensomotorischen Integration und erfahrungsabhängigen visuellen Plastizität (Fu et al., 2014; Pi et al., 2013; Lee et al., 2013; Fu et al., 2015).
In relativ guter Übereinstimmung mit der angenommenen Verschaltung werden VIP+ Interneurone im Areal V1 während der Dunkelheit angeregt, während SOM+ Interneurone inhibiert werden (Reimer et al., 2014; Fu et al., 2014; Pakan et al., 2016) (Abbildung 3c, links). Obwohl der disinhibitorische Signalweg VIP->SOM->Pyramidenzelle die durchschnittlichen Effekte von Fortbewegung in V1 erklären kann, gilt es jedoch zu beachten, dass die Verteilung der Fortbewegungseffekte bei beiden Interneurontypen breit ist (Fu et al., 2014; Pakan et al., 2016). Darüber hinaus wird das disinhibitorische Schaltungsmodell durch neuere Befunde in Frage gestellt, die zeigen, dass während der visuellen Stimulation die Aktivität von sowohl VIP+ als auch SOM+ Interneuronen erhöht ist (Pakan et al., 2016) (Abbildung 3c, rechts).
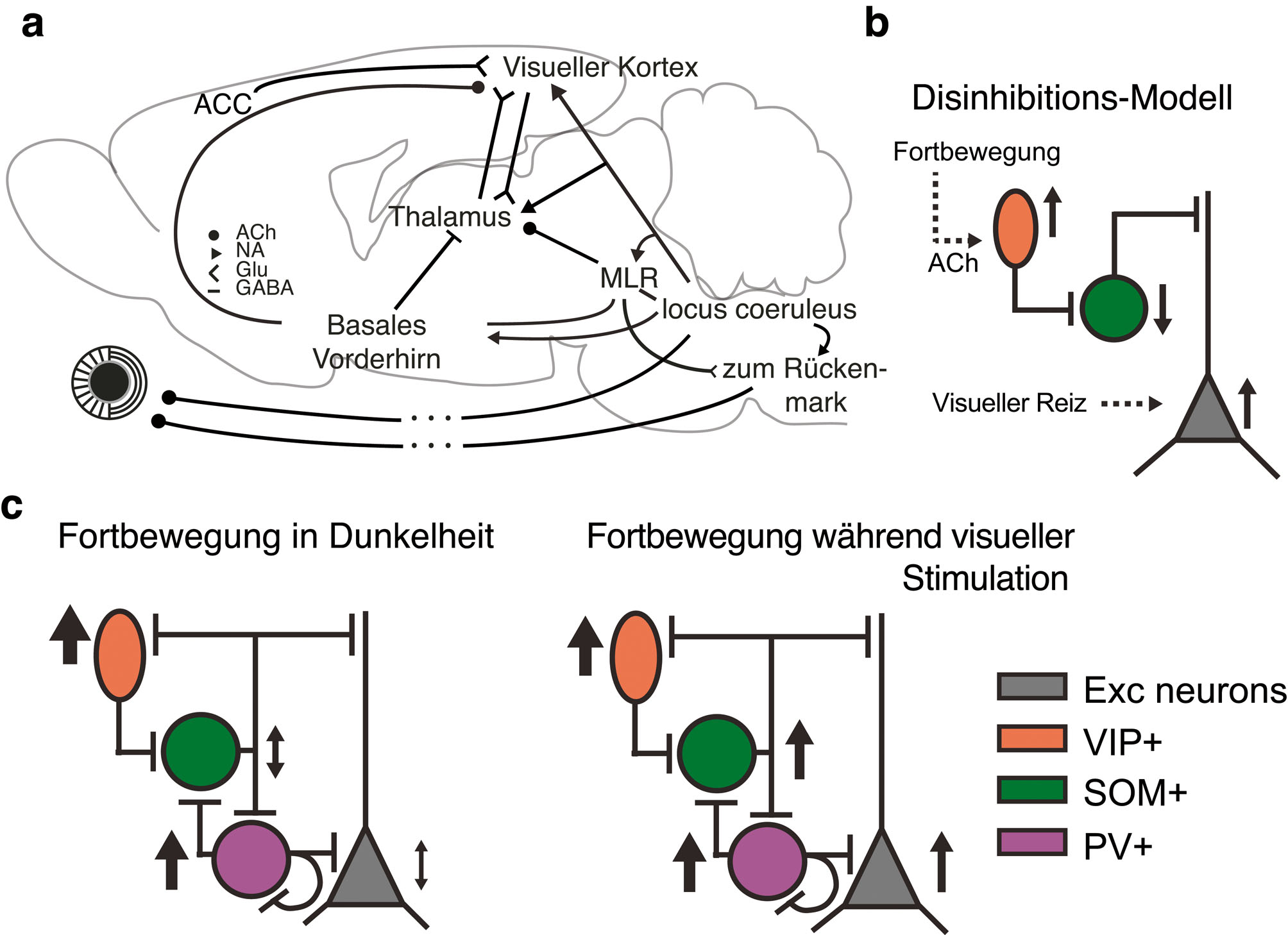
Neuronale Schaltkreise, die zur Modulation visueller Aktivität durch Bewegung beitragen können. a) Es ist wahrscheinlich, dass eine Vielzahl verschiedener neuronaler Schaltkreise zur Modulation der sensorischen Informationsverarbeitung durch Fortbewegung beitragen. ACh: Acetylcholin, NA: Norepinephrine, Glu: Glutamat, MLR: mesencephalic locomotor region, ACC: anteriorer cingulärer Kortex – mit freundlicher Genehmigung modifiziert aus Erisken et al. (2014). b) Ein vorgeschlagener disinhibitorischer Verarbeitungspfad: Acetylcholin, das während der Fortbewegung ausgeschüttet wird, aktiviert VIP+ inhibitorische Interneurone, die wiederum SOM+ inhibitorische Interneurone hemmen. Dies führt zu einer Disinhibition von Pyramidenzellaktivität. c) Experimentelle Beobachtungen während der Fortbewegung in Dunkelheit (links) und während der visuellen Stimulation (rechts), die mit dem vorgeschlagenen disinhibitorischen Schaltkreis nur bedingt übereinstimmen – mit freundlicher Genehmigung modifiziert aus Pakan et al. (2016).
Eine detaillierte Untersuchung des disinhibitorischen VIP->SOM->Pyramidenzellen-Signalwegs und eine Erklärung für die beobachtete Variabilität innerhalb von Interneurontypen wurde kürzlich im somatosensorischen System geliefert (Muñoz et al., 2017). Hier konnten Muñoz und Kollegen zeigen, dass ein spezifischer Subtyp von SOM+ Interneuronen mit axonalen Verzweigungen in Schicht 1 und Schicht 5a während der Bewegung der Vibrissen unterdrückt wird, während andere SOM+ Interneuron-Subtypen entgegengesetzte Effekte zeigen. Interessanterweise ist S1 – Schicht 5a eines der Hauptzielgebiete von Schicht 2/3 VIP+ Interneuronen (Prönneke et al., 2015), und SOM+ Interneurone, die durch Bewegung der Vibrissen unterdrückt werden, erhalten eine besonders starke Hemmung über VIP+ Interneurone (Muñoz et al., 2017).
Neuromodulatorische Einflüsse während der Fortbewegung können von spezifischeren Top-down-Feedback-Modulationen (Zhang et al., 2016; Zhang et al., 2014) und Einflüssen von motorischen Efferenzkopien begleitet werden (Schneider et al., 2014; Leinweber et al., 2017). Tatsächlich erhält der primäre visuelle Kortex einen starken Eingang von einer speziellen Region des anterioren Cingulums (ACC), genannt A24b, und dem sekundären motorischen Kortex (M2). Beide Strukturen senden motorbezogene Signale an V1, die von der spezifischen visuellen Erfahrung abhängen (Ding et al., 2013) und dahingehend interpretiert werden können, dass sie Vorhersagen repräsentieren, die auf dem erwarteten visuellen Fluss basieren.
Perspektiven für die Zukunft
Die Berücksichtigung der Fortbewegung in Studien zur sensorischen Informationsverarbeitung hat die Entdeckung von Signalen ermöglicht, die nicht nur die neuronalen Antworten modulieren können, sondern auch aktivem Verhalten, wie der Navigation, dienen. Dennoch unterliegt die Fortbewegung unter Kopffixierung im Vergleich zur Fortbewegung unter realen Bedingungen einem relativ reduzierten Einfluss von anderen Variablen. Zum Beispiel fehlen beim Laufen unter Kopffixierung vestibuläre Einflüsse, von denen bekannt ist, dass sie weitverbreitete Einflüsse im gesamten Nagetierkortex (Rancz et al., 2015) haben. Die Kombination technischer Fortschritte, unter anderem die Miniaturisierung der neurophysiologischen Messinstrumente (Kerr und Nimmerjahn, 2012), Eye Tracking, das direkt auf dem Kopf angebracht werden kann (Wallace et al., 2013), realistische virtuelle Realitätssysteme mit geringer Antwortlatenz (Grosso et al., 2017; Stowers et al., 2017) und Methoden des maschinellen Lernens zum automatischen Tracking von Tieren und detaillierter und genauer Segmentierung von Verhalten (Valetta et al., 2017) verspricht, in Zukunft kontrollierte sensorische Stimulation während komplexen, natürlichen und im Hinblick auf die Evolution wichtigen Verhaltens zu ermöglichen.
About the author

Laura Busse ist seit 2016 Professorin für „Organismische Neurobiologie” in der Abteilung Neurobiologie des Departments für Biologie II an der LMU München und ist mit dem Bernstein Zentrum für Computational Neuroscience affiliert. Sie promovierte an der Universität Göttingen im Rahmen ihrer Arbeit am Deutschen Primatenzentrum, war Postdoktorandin am Smith Kettlewell Eye Research Institute in San Francisco, USA, und am UCL Institute of Ophthalmology in London. Zwischen 2010 and 2015 leitete Laura Busse eine Juniorgruppe am Zentrum für Integrative Neurowissenschaften (CIN) der Universität Tübingen.
Literatur
Alonso, J. M. and Swadlow, H. A. (2017). Thalamocortical Interactions for Sensory Processing, doi: 10.1093/acrefore/9780190264086.013.112.10.1093/acrefore/9780190264086.013.112Suche in Google Scholar
Andermann, M. L., Gilfoy, N. B., Goldey, G. J. et al. (2013). Chronic cellular imaging of entire cortical columns in awake mice using microprisms. Neuron 80, 900–913, doi: 10.1016/j.neuron.2013.07.05210.1016/j.neuron.2013.07.052Suche in Google Scholar PubMed PubMed Central
Andermann, M. L., Kerlin, A. M., Roumis, D. K. et al. (2011). Functional Specialization of Mouse Higher Visual Cortical Areas. Neuron 72, 1025–1039.10.1016/j.neuron.2011.11.013Suche in Google Scholar PubMed PubMed Central
Aston-Jones, G. and Cohen, J. D. (2005). AN INTEGRATIVE THEORY OF LOCUS COERULEUS-NOREPINEPHRINE FUNCTION: Adaptive Gain and Optimal Performance. Annu. Rev. Neurosci. 28, 403–450, doi: 10.1146/annurev.neuro.28.061604.135709.10.1146/annurev.neuro.28.061604.135709Suche in Google Scholar PubMed
Atiani, S., Elhilali, M., David, S. V. et al. (2009). Task Difficulty and Performance Induce Diverse Adaptive Patterns in Gain and Shape of Primary Auditory Cortical Receptive Fields. Neuron 61, 467–480.10.1016/j.neuron.2008.12.027Suche in Google Scholar PubMed PubMed Central
Attinger, A., Wang, B. and Keller, G. B. (2017). Visuomotor Coupling Shapes the Functional Development of Mouse Visual Cortex. Cell 169, 1291–1302.e14, doi: 10.1016/j.cell.2017.05.023.10.1016/j.cell.2017.05.023Suche in Google Scholar PubMed
Ayaz, A., Saleem, A. B., Schölvinck, M. L. and Carandini, M. (2013). Locomotion Controls Spatial Integration in Mouse Visual Cortex. Curr. Biol. 23, 890–894, doi: 10.1016/j.cub.2013.04.012.10.1016/j.cub.2013.04.012Suche in Google Scholar PubMed PubMed Central
Bennett, C., Arroyo, S. and Hestrin, S. (2013). Subthreshold Mechanisms Underlying State-Dependent Modulation of Visual Responses. Neuron 80, 350–357.10.1016/j.neuron.2013.08.007Suche in Google Scholar PubMed PubMed Central
Bezdudnaya, T., Cano, M., Bereshpolova, Y. et al. (2006). Thalamic Burst Mode and Inattention in the Awake LGNd. Neuron 49, 421–432, doi: 10.1016/j.neuron.2006.01.010.10.1016/j.neuron.2006.01.010Suche in Google Scholar PubMed
Bickford, M. E., Günlük, A. E., Van Horn, S. C. and Sherman, S. M. (1994). GABAergic projection from the basal forebrain to the visual sector of the thalamic reticular nucleus in the cat. J. Comp. Neurol. 348, 481–510, doi: 10.1002/cne.903480402.10.1002/cne.903480402Suche in Google Scholar PubMed
Bortone, D. S., Olsen, S. R. and Scanziani, M. (2014). Translaminar inhibitory cells recruited by layer 6 corticothalamic neurons suppress visual cortex. Neuron 82, 474–485, doi: 10.1016/j.neuron.2014.02.021.10.1016/j.neuron.2014.02.021Suche in Google Scholar PubMed PubMed Central
Bradshaw, J. (1967). Pupil Size as a Measure of Arousal during Information Processing. Nature 216, 515–516, doi: 10.1038/216515a0.10.1038/216515a0Suche in Google Scholar PubMed
Buchner, E. (1976). Elementary movement detectors in an insect visual system. Biol. Cybern. 24, 85–101, doi: 10.1007/BF00360648.10.1007/BF00360648Suche in Google Scholar
Bullock, T., Elliott, J. C., Serences, J. T. and Giesbrecht, B. (2016). Acute Exercise Modulates Feature-selective Responses in Human Cortex. J. Cogn. Neurosci. 1–14, doi: 10.1162/jocn_a_01082.10.1162/jocn_a_01082Suche in Google Scholar PubMed
Cano, M., Bezdudnaya, T., Swadlow, H. A. and Alonso, J.-M. (2006). Brain state and contrast sensitivity in the awake visual thalamus. Nat. Neurosci. 9, 1240–1242.10.1038/nn1760Suche in Google Scholar PubMed
Carandini, M. and Churchland, A. K. (2013). Probing perceptual decisions in rodents. Nat. Neurosci. 16, 824–831, doi: 10.1038/nn.3410.10.1038/nn.3410Suche in Google Scholar PubMed PubMed Central
Carandini, M. and Heeger, D. J. (2012). Normalization as a canonical neural computation. Nat. Rev. Neurosci. 13, 51–62.10.1038/nrn3136Suche in Google Scholar PubMed PubMed Central
Chen, N., Sugihara, H. and Sur, M. (2015). An acetylcholine-activated microcircuit drives temporal dynamics of cortical activity. Nat. Neurosci. 18, 892–902, doi: 10.1038/nn.4002.10.1038/nn.4002Suche in Google Scholar PubMed PubMed Central
Constantinople, C. M. and Bruno, R. M. (2011). Effects and mechanisms of wakefulness on local cortical networks. Neuron 69, 1061–1068, doi: 10.1016/j.neuron.2011.02.040.10.1016/j.neuron.2011.02.040Suche in Google Scholar PubMed PubMed Central
Crochet, S. and Petersen, C. C. H. (2006). Correlating whisker behavior with membrane potential in barrel cortex of awake mice. Nat. Neurosci. 9, 608–610, doi: 10.1038/nn1690.10.1038/nn1690Suche in Google Scholar PubMed
Dadarlat, M. C. and Stryker, M. P. (2017). Locomotion Enhances Neural Encoding of Visual Stimuli in Mouse V1. J. Neurosci. 37, 3764–3775, doi: 10.1523/JNEUROSCI.2728-16.2017.10.1523/JNEUROSCI.2728-16.2017Suche in Google Scholar PubMed PubMed Central
David, S. V., Fritz, J. B. and Shamma, S. A. (2012). Task reward structure shapes rapid receptive field plasticity in auditory cortex. Proc. Natl. Acad. Sci. U.S.A. 109, 2144–2149.10.1073/pnas.1117717109Suche in Google Scholar PubMed PubMed Central
Ding, F., O’Donnell, J., Thrane, A. S. et al. (2013). α1-Adrenergic receptors mediate coordinated Ca2+ signaling of cortical astrocytes in awake, behaving mice. Cell Calcium 54, 387–394, doi: 10.1016/j.ceca.2013.09.001.10.1016/j.ceca.2013.09.001Suche in Google Scholar PubMed PubMed Central
Dombeck, D. A., Khabbaz, A. N., Collman, F. et al. (2007). Imaging Large-Scale Neural Activity with Cellular Resolution in Awake, Mobile Mice. Neuron 56, 43–57.10.1016/j.neuron.2007.08.003Suche in Google Scholar PubMed PubMed Central
Erisken, S., Vaiceliunaite, A., Jurjut, O. et al. (2014). Effects of Locomotion Extend throughout the Mouse Early Visual System. Curr. Biol. 24, 2899–2907, doi: 10.1016/j.cub.2014.10.045.10.1016/j.cub.2014.10.045Suche in Google Scholar PubMed
Fanselow, E. E., Richardson, K. A. and Connors, B. W. (2008). Selective, State-Dependent Activation of Somatostatin-Expressing Inhibitory Interneurons in Mouse Neocortex. J. Neurophysiol. 100, 2640–2652, doi: 10.1152/jn.90691.2008.10.1152/jn.90691.2008Suche in Google Scholar PubMed PubMed Central
Fiser, A., Mahringer, D., Oyibo, H. K. et al. (2016). Experience-dependent spatial expectations in mouse visual cortex. Nat. Neurosci. 19, 1658–1664, doi: 10.1038/nn.4385.10.1038/nn.4385Suche in Google Scholar PubMed
Friston, K. (2005). A theory of cortical responses. Philos. Trans. R. Soc. B Biol. Sci. 360, 815–836.10.1098/rstb.2005.1622Suche in Google Scholar PubMed PubMed Central
Fu, Y., Kaneko, M., Tang, Y. et al. (2015). A cortical disinhibitory circuit for enhancing adult plasticity. eLife 4, e05558, doi: 10.7554/eLife.05558.10.7554/eLife.05558Suche in Google Scholar PubMed PubMed Central
Fu, Y., Tucciarone, J. M., Espinosa, J. S. et al. (2014). A cortical circuit for gain control by behavioral state. Cell 156, 1139–1152, doi:10.1016/j.cell.2014.01.050.10.1016/j.cell.2014.01.050Suche in Google Scholar PubMed PubMed Central
Gentet, L. J., Avermann, M., Matyas, F. et al. (2010). Membrane Potential Dynamics of GABAergic Neurons in the Barrel Cortex of Behaving Mice. Neuron 65, 422–435.10.1016/j.neuron.2010.01.006Suche in Google Scholar PubMed
Gilbert, C. D. and Sigman, M. (2007). Brain States: Top-Down Influences in Sensory Processing. Neuron 54, 677–696.10.1016/j.neuron.2007.05.019Suche in Google Scholar PubMed
Grosso, N. A. D., Graboski, J. J., Chen, W. et al. (2017). Virtual Reality system for freely-moving rodents. BioRxiv 161232, doi: 10.1101/161232.10.1101/161232Suche in Google Scholar
Harris, K. D. and Thiele, A. (2011). Cortical state and attention. Nat. Rev. Neurosci. 12, 509–512, doi: 10.1038/nrn3084.10.1038/nrn3084Suche in Google Scholar PubMed PubMed Central
Harvey, C. D., Collman, F., Dombeck, D. A. and Tank, D. W. (2009). Intracellular dynamics of hippocampal place cells during virtual navigation. Nature 461, 941–946.10.1038/nature08499Suche in Google Scholar PubMed PubMed Central
Hirata, A. and Castro-Alamancos, M. A. (2010). Neocortex Network Activation and Deactivation States Controlled by the Thalamus. J. Neurophysiol. 103, 1147–1157, doi: 10.1152/jn.00955.2009.10.1152/jn.00955.2009Suche in Google Scholar PubMed PubMed Central
Histed, M. H., Carvalho, L. A. and Maunsell, J. H. R. (2012). Psychophysical measurement of contrast sensitivity in the behaving mouse. J. Neurophysiol. 107, 758–765, doi: 10.1152/jn.00609.2011.10.1152/jn.00609.2011Suche in Google Scholar PubMed PubMed Central
Holscher, C., Schnee, A., Dahmen, H. et al. (2005). Rats are able to navigate in virtual environments. J. Exp. Biol. 208, 561–569.10.1242/jeb.01371Suche in Google Scholar PubMed
Keller, G. B., Bonhoeffer, T. and Hübener, M. (2012). Sensorimotor mismatch signals in primary visual cortex of the behaving mouse. Neuron 74, 809–815, doi: 10.1016/j.neuron.2012.03.040.10.1016/j.neuron.2012.03.040Suche in Google Scholar PubMed
Kerr, J. N. and Nimmerjahn, A. (2012). Functional imaging in freely moving animals. Curr. Opin. Neurobiol. 22, 45–53, doi: 10.1016/j.conb.2011.12.002.10.1016/j.conb.2011.12.002Suche in Google Scholar PubMed
Kjaerby, C., Rasmussen, R., Andersen, M. and Nedergaard, M. (2017). Does Global Astrocytic Calcium Signaling Participate in Awake Brain State Transitions and Neuronal Circuit Function? Neurochem. Res. 42, 1810–1822, doi: 10.1007/s11064-017-2195-y.10.1007/s11064-017-2195-ySuche in Google Scholar PubMed PubMed Central
Kruglikov, I. and Rudy, B. (2008). Perisomatic GABA Release and Thalamocortical Integration onto Neocortical Excitatory Cells Are Regulated by Neuromodulators. Neuron 58, 911–924, doi: 10.1016/j.neuron.2008.04.024.10.1016/j.neuron.2008.04.024Suche in Google Scholar PubMed PubMed Central
Kuchibhotla, K. V., Gill, J. V., Lindsay, G. W. et al. (2017). Parallel processing by cortical inhibition enables context-dependent behavior. Nat. Neurosci. 20, 62–71.10.1038/nn.4436Suche in Google Scholar PubMed PubMed Central
Laughlin, S. B. (1992). Retinal information capacity and the function of the pupil. Ophthalmic Physiol. Opt. 12, 161–164, doi: 10.1111/j.1475-1313.1992.tb00281.x.10.1111/j.1475-1313.1992.tb00281.xSuche in Google Scholar PubMed
Lee, A. M., Hoy, J. L., Bonci, A. et al. (2014). Identification of a Brainstem Circuit Regulating Visual Cortical State in Parallel with Locomotion. Neuron 83, 455–466, doi: 10.1016/j.neuron.2014.06.031.10.1016/j.neuron.2014.06.031Suche in Google Scholar PubMed PubMed Central
Lee, S.-H. and Dan, Y. (2012). Neuromodulation of Brain States. Neuron 76, 209–222, doi: 10.1016/j.neuron.2012.09.012.10.1016/j.neuron.2012.09.012Suche in Google Scholar PubMed PubMed Central
Lee, S., Kruglikov, I., Huang, Z. J. et al. (2013). A disinhibitory circuit mediates motor integration in the somatosensory cortex. Nat. Neurosci. 16, 1662–1670, doi: 10.1038/nn.3544.10.1038/nn.3544Suche in Google Scholar PubMed PubMed Central
Leinweber, M., Ward, D. R., Sobczak, J. M. et al. (2017). A Sensorimotor Circuit in Mouse Cortex for Visual Flow Predictions. Neuron 95, 1420–1432.e5, doi: 10.1016/j.neuron.2017.08.036.10.1016/j.neuron.2017.08.036Suche in Google Scholar PubMed
Maimon, G. (2011). Modulation of visual physiology by behavioral state in monkeys, mice, and flies. Curr. Opin. Neurobiol. 21, 559–564, doi: 10.1016/j.conb.2011.05.001.10.1016/j.conb.2011.05.001Suche in Google Scholar PubMed
McGinley, M. J., David, S. V. and McCormick, D. A. (2015). Cortical Membrane Potential Signature of Optimal States for Sensory Signal Detection. Neuron 87, 179–192, doi: 10.1016/j.neuron.2015.05.038.10.1016/j.neuron.2015.05.038Suche in Google Scholar PubMed PubMed Central
Mineault, P. J., Tring, E., Trachtenberg, J. T. and Ringach, D. L. (2016). Enhanced Spatial Resolution During Locomotion and Heightened Attention in Mouse Primary Visual Cortex. J. Neurosci. 36, 6382–6392, doi: 10.1523/JNEUROSCI.0430-16.2016.10.1523/JNEUROSCI.0430-16.2016Suche in Google Scholar PubMed PubMed Central
Moser, E., Mathiesen, I. and Andersen, P. (1993). Association between brain temperature and dentate field potentials in exploring and swimming rats. Science 259, 1324–1326, doi: 10.1126/science.8446900.10.1126/science.8446900Suche in Google Scholar PubMed
Muñoz, W., Tremblay, R., Levenstein, D. and Rudy, B. (2017). Layer-specific modulation of neocortical dendritic inhibition during active wakefulness. Science 355, 954–959, doi: 10.1126/science.aag2599.10.1126/science.aag2599Suche in Google Scholar PubMed
Niell, C. M. and Stryker, M. P. (2010). Modulation of Visual Responses by Behavioral State in Mouse Visual Cortex. Neuron 65, 472–479.10.1016/j.neuron.2010.01.033Suche in Google Scholar PubMed PubMed Central
Olsen, S. R., Bortone, D. S., Adesnik, H. and Scanziani, M. (2012). Gain control by layer six in cortical circuits of vision. Nature 483, 47–52, doi: 10.1038/nature10835.10.1038/nature10835Suche in Google Scholar PubMed PubMed Central
Otazu, G. H., Tai, L.-H., Yang, Y. and Zador, A. M. (2009). Engaging in an auditory task suppresses responses in auditory cortex. Nat. Neurosci. 12, 646–654.10.1038/nn.2306Suche in Google Scholar PubMed PubMed Central
Pakan, J. M., Lowe, S. C., Dylda, E. et al. (2016). Behavioral-state modulation of inhibition is context-dependent and cell type specific in mouse visual cortex. eLife 5, e14985, doi: 10.7554/eLife.1498510.7554/eLife.14985Suche in Google Scholar PubMed PubMed Central
Paukert, M., Agarwal, A., Cha, J. et al. (2014). Norepinephrine Controls Astroglial Responsiveness to Local Circuit Activity. Neuron 82, 1263–1270, doi: 10.1016/j.neuron.2014.04.038.10.1016/j.neuron.2014.04.038Suche in Google Scholar PubMed PubMed Central
Pennesi, M. E., Lyubarsky, A. L. and Pugh, E. N. (1998). Extreme responsiveness of the pupil of the dark-adapted mouse to steady retinal illumination. Invest. Ophthalmol. Vis. Sci. 39, 2148–2156.Suche in Google Scholar PubMed
Pfeffer, C. K., Xue, M., He, M. et al. (2013). Inhibition of inhibition in visual cortex: the logic of connections between molecularly distinct interneurons. Nat. Neurosci. 16, 1068–1076, doi: 10.1038/nn.3446.10.1038/nn.3446Suche in Google Scholar PubMed PubMed Central
Pi, H.-J., Hangya, B., Kvitsiani, D. et al. (2013). Cortical interneurons that specialize in disinhibitory control. Nature 503, 521–524, doi: 10.1038/nature12676.10.1038/nature12676Suche in Google Scholar PubMed PubMed Central
Pinto, L., Goard, M. J., Estandian, D. et al. (2013). Fast modulation of visual perception by basal forebrain cholinergic neurons. Nat. Neurosci. 16, 1857–1863, doi: 10.1038/nn.3552.10.1038/nn.3552Suche in Google Scholar PubMed PubMed Central
Pluta, S., Naka, A., Veit, J. et al. (2015). A direct translaminar inhibitory circuit tunes cortical output. Nat. Neurosci. 18, 1631–1640, doi: 10.1038/nn.4123.10.1038/nn.4123Suche in Google Scholar PubMed PubMed Central
Polack, P.-O., Friedman, J. and Golshani, P. (2013). Cellular mechanisms of brain state-dependent gain modulation in visual cortex. Nat. Neurosci. 16, 1331–1339, doi: 10.1038/nn.3464.10.1038/nn.3464Suche in Google Scholar PubMed PubMed Central
Poulet, J. F. A. and Petersen, C. C. H. (2008). Internal brain state regulates membrane potential synchrony in barrel cortex of behaving mice. Nature 454, 881–885.10.1038/nature07150Suche in Google Scholar PubMed
Poulet, J. F. A., Fernandez, L. M. J., Crochet, S. and Petersen, C. C. H. (2012). Thalamic control of cortical states. Nat. Neurosci. 15, 370–372.10.1038/nn.3035Suche in Google Scholar PubMed
Prönneke, A., Scheuer, B., Wagener, R. J. et al. (2015). Characterizing VIP Neurons in the Barrel Cortex of VIPcre/tdTomato Mice Reveals Layer-Specific Differences. Cereb. Cortex 25, 4854–4868, doi: 10.1093/cercor/bhv202.10.1093/cercor/bhv202Suche in Google Scholar PubMed PubMed Central
Rancz, E. A., Moya, J., Drawitsch, F. et al. (2015). Widespread Vestibular Activation of the Rodent Cortex. J. Neurosci. 35, 5926–5934, doi: 10.1523/JNEUROSCI.1869-14.2015.10.1523/JNEUROSCI.1869-14.2015Suche in Google Scholar PubMed PubMed Central
Rao, R. P. N. and Ballard, D. H. (1999). Predictive coding in the visual cortex: a functional interpretation of some extra-classical receptive-field effects. Nat. Neurosci. 2, 79–87, doi: 10.1038/4580.10.1038/4580Suche in Google Scholar PubMed
Reimer, J., Froudarakis, E., Cadwell, C. R. et al. (2014). Pupil Fluctuations Track Fast Switching of Cortical States during Quiet Wakefulness. Neuron 84, 355–362, doi: 10.1016/j.neuron.2014.09.033.10.1016/j.neuron.2014.09.033Suche in Google Scholar PubMed PubMed Central
Reimer, J., McGinley, M. J., Liu, Y. et al. (2016). Pupil fluctuations track rapid changes in adrenergic and cholinergic activity in cortex. Nat. Commun. 7, 13289, doi: 10.1038/ncomms13289.10.1038/ncomms13289Suche in Google Scholar PubMed PubMed Central
Roelfsema, P. R., Lamme, V. A. and Spekreijse, H. (1998). Object-based attention in the primary visual cortex of the macaque monkey. Nature 395, 376–381.10.1038/26475Suche in Google Scholar PubMed
Roth, M. M., Dahmen, J. C., Muir, D. R. et al. (2016). Thalamic nuclei convey diverse contextual information to layer 1 of visual cortex. Nat. Neurosci. 19, 299–307, doi: 10.1038/nn.4197.10.1038/nn.4197Suche in Google Scholar PubMed PubMed Central
Saleem, A. B., Ayaz, A., Jeffery, K. J. et al. (2013). Integration of visual motion and locomotion in mouse visual cortex. Nat. Neurosci. 16, 1864–1869, doi: 10.1038/nn.3567.10.1038/nn.3567Suche in Google Scholar PubMed PubMed Central
Saleem, A. B., Lien, A. D., Krumin, M. et al. (2017). Subcortical Source and Modulation of the Narrowband Gamma Oscillation in Mouse Visual Cortex. Neuron 93, 315–322, doi: 10.1016/j.neuron.2016.12.028.10.1016/j.neuron.2016.12.028Suche in Google Scholar PubMed PubMed Central
Samuels, E. R. and Szabadi, E. (2008). Functional neuroanatomy of the noradrenergic locus coeruleus: its roles in the regulation of arousal and autonomic function part II: physiological and pharmacological manipulations and pathological alterations of locus coeruleus activity in humans. Curr. Neuropharmacol. 6, 254–285, doi: 10.2174/157015908785777193.10.2174/157015908785777193Suche in Google Scholar PubMed PubMed Central
Schneider, D. M., Nelson, A. and Mooney, R. (2014). A synaptic and circuit basis for corollary discharge in the auditory cortex. Nature 513, 189–194, doi: 10.1038/nature13724.10.1038/nature13724Suche in Google Scholar PubMed PubMed Central
Schwarz, C., Hentschke, H., Butovas, S. et al. (2011). The head-fixed behaving rat – Procedures and pitfalls. Somatosensory & Motor Research 27, 131–148, doi: 10.3109/08990220.2010.51311110.3109/08990220.2010.513111Suche in Google Scholar PubMed PubMed Central
Shirey, M. J., Smith, J. B., Kudlik, D. E. et al, (2015). Brief anesthesia, but not voluntary locomotion, significantly alters cortical temperature. J. Neurophysiol. 114, 309–322, doi: 10.1152/jn.00046.2015.10.1152/jn.00046.2015Suche in Google Scholar PubMed PubMed Central
Sofroniew, N. J., Vlasov, Y. A., Hires, S. A. et al. (2015). Neural coding in barrel cortex during whisker-guided locomotion. eLife 4, e12559, doi: 10.7554/eLife.12559.10.7554/eLife.12559Suche in Google Scholar PubMed PubMed Central
Spitzer, H., Desimone, R. and Moran, J. (1988). Increased attention enhances both behavioral and neuronal performance. Science 240, 338–340.10.1126/science.3353728Suche in Google Scholar PubMed
Storchi, R., Bedford, R. A., Martial, F. P. et al. (2017). Modulation of Fast Narrowband Oscillations in the Mouse Retina and dLGN According to Background Light Intensity. Neuron 93, 299–307, doi: 10.1016/j.neuron.2016.12.027.10.1016/j.neuron.2016.12.027Suche in Google Scholar PubMed
Stowers, J. R., Hofbauer, M., Bastien, R. et al. (2017). Virtual reality for freely moving animals. Nat. Methods advance online publication, doi: 10.1038/nmeth.4399.10.1038/nmeth.4399Suche in Google Scholar PubMed PubMed Central
Thiele, A. (2013). Muscarinic Signaling in the Brain. Annu. Rev. Neurosci. 36, 271–294, doi: 10.1146/annurev-neuro-062012-170433.10.1146/annurev-neuro-062012-170433Suche in Google Scholar PubMed
Treue, S. and Maunsell, J. H. R. (1996). Attentional modulation of visual motion processing in cortical areas MT and MST. Nature 382, 539–541.10.1038/382539a0Suche in Google Scholar PubMed
Valletta, J. J., Torney, C., Kings, M. et al. (2017). Applications of machine learning in animal behaviour studies. Anim. Behav. 124, 203–220, doi: 10.1016/j.anbehav.2016.12.005.10.1016/j.anbehav.2016.12.005Suche in Google Scholar
Vinck, M., Batista-Brito, R., Knoblich, U. and Cardin, J. A. (2015). Arousal and Locomotion Make Distinct Contributions to Cortical Activity Patterns and Visual Encoding. Neuron 86, 740–754, doi: 10.1016/j.neuron.2015.03.028.10.1016/j.neuron.2015.03.028Suche in Google Scholar PubMed PubMed Central
Wallace, D. J., Greenberg, D. S., Sawinski, J. et al. (2013). Rats maintain an overhead binocular field at the expense of constant fusion. Nature 498, 65–69, doi: 10.1038/nature12153.10.1038/nature12153Suche in Google Scholar PubMed
Williamson, R. S., Hancock, K. E., Shinn-Cunningham, B. G. and Polley, D. B. (2015). Locomotion and Task Demands Differentially Modulate Thalamic Audiovisual Processing during Active Search. Curr. Biol. 25, 1885–1891, doi: 10.1016/j.cub.2015.05.045.10.1016/j.cub.2015.05.045Suche in Google Scholar PubMed PubMed Central
Zagha, E. and McCormick, D. A. (2014). Neural control of brain state. Curr. Opin. Neurobiol. 29, 178–186, doi: 10.1016/j.conb.2014.09.010.10.1016/j.conb.2014.09.010Suche in Google Scholar PubMed PubMed Central
Zhang, S., Xu, M., Chang, W.-C. et al. (2016). Organization of long-range inputs and outputs of frontal cortex for top-down control. Nat. Neurosci. 19, 1733–1742, doi: 10.1038/nn.441710.1038/nn.4417Suche in Google Scholar PubMed PubMed Central
Zhang, S., Xu, M., Kamigaki, T. et al. (2014). Selective attention. Long-range and local circuits for top-down modulation of visual cortex processing. Science 345, 660–665, doi: 10.1126/science.1254126.10.1126/science.1254126Suche in Google Scholar PubMed PubMed Central
Zhou, M., Liang, F., Xiong, X. R. et al. (2014). Scaling down of balanced excitation and inhibition by active behavioral states in auditory cortex. Nat. Neurosci. 17, 841–850, doi: 10.1038/nn.3701.10.1038/nn.3701Suche in Google Scholar PubMed PubMed Central
Zmarz, P. and Keller, G. B. (2016). Mismatch Receptive Fields in Mouse Visual Cortex. Neuron 92, 766–772, doi: 10.1016/j.neuron.2016.09.057.10.1016/j.neuron.2016.09.057Suche in Google Scholar PubMed
Anmerkung
Übersetzung der englischen Version des Artikels online verfügbar unter https://doi.org/10.1515/nf-2017-A046
© 2017 by De Gruyter
Artikel in diesem Heft
- Frontmatter
- Übersichtsartikel
- Selektive Degeneration dopaminerger Neurone beim Parkinson-Syndrom: die zunehmende Rolle von veränderter Kalziumhomöostase und nukleolärer Funktion
- Selective degeneration of dopamine neurons in Parkinson’s disease: emerging roles of altered calcium homeostasis and nucleolar function
- Altruismus aus Sicht der Sozialen Neurowissenschaften
- Altruism from the Perspective of the Social Neurosciences
- Lebensbedingungen haben einen starken Einfluss auf die Plastizität des Gehirns
- Environmental conditions strongly affect brain plasticity
- Die neuronalen Signale, die Wahrnehmung verändern
- The neural events that change perception
- Der Einfluss von Fortbewegung auf die sensorische Informationsverarbeitung und die zugrunde liegenden neuronalen Schaltkreise
- The influence of locomotion on sensory processing and its underlying neuronal circuits
- Forschungsförderung
- Sonderforschungsbereich (SFB/TRR 167) NeuroMac „Entwicklung, Funktion und Potenzial von myeloischen Zellen im zentralen Nervensystem“
- Rezension
- Michael Madeja/Joachim Müller-Jung (Hrsg.): Hirnforschung – was kann sie wirklich
- Nachrichten
- Neue NWG-Website
Artikel in diesem Heft
- Frontmatter
- Übersichtsartikel
- Selektive Degeneration dopaminerger Neurone beim Parkinson-Syndrom: die zunehmende Rolle von veränderter Kalziumhomöostase und nukleolärer Funktion
- Selective degeneration of dopamine neurons in Parkinson’s disease: emerging roles of altered calcium homeostasis and nucleolar function
- Altruismus aus Sicht der Sozialen Neurowissenschaften
- Altruism from the Perspective of the Social Neurosciences
- Lebensbedingungen haben einen starken Einfluss auf die Plastizität des Gehirns
- Environmental conditions strongly affect brain plasticity
- Die neuronalen Signale, die Wahrnehmung verändern
- The neural events that change perception
- Der Einfluss von Fortbewegung auf die sensorische Informationsverarbeitung und die zugrunde liegenden neuronalen Schaltkreise
- The influence of locomotion on sensory processing and its underlying neuronal circuits
- Forschungsförderung
- Sonderforschungsbereich (SFB/TRR 167) NeuroMac „Entwicklung, Funktion und Potenzial von myeloischen Zellen im zentralen Nervensystem“
- Rezension
- Michael Madeja/Joachim Müller-Jung (Hrsg.): Hirnforschung – was kann sie wirklich
- Nachrichten
- Neue NWG-Website