The influence of locomotion on sensory processing and its underlying neuronal circuits
-
Laura Busse
Laura Busse has been professor for “Organismic Neurobiology” at the Division of Neurobiology of the Department of Biology II at the LMU Munich since 2016 and is affiliated with the Bernstein Center for Computational Neuroscience. She obtained her PhD from the University of Göttingen for work at the German Primate Center, was a postdoctoral fellow at the Smith Kettlewell Eye Research Institute in San Francisco and at the UCL Institute of Ophthalmology in London. Between 2010 and 2015 Laura Busse was a Junior Research Group Leader at the Centre for Integrative Neuroscience (CIN) at the University of Tübingen.


Abstract
Processing of sensory information can be modulated in both cortex and thalamus by behavioral context, such as locomotion. During active behaviors, coding of sensory stimuli and perception are improved, in particular during physical activity of moderate intensity. These locomotion-related modulations seem to arise from a combination of mechanisms, including neuromodulation, the recruitment of inhibitory interneurons, and specific top-down or motor-related inputs. The application of new experimental methods in mice during walking under head-fixation on treadmills made it possible to study the circuit and cellular basis underlying modulations by behavioral context with unprecedented detail. This article reviews the current state of these studies and highlights some important open questions.
Introduction
The study of the neural basis of sensation has a long tradition of powerful experimental approaches, where anesthetized or awake, head-fixed animals are presented with highly controlled sensory stimuli. It was this exquisite experimental control that allowed detailed characterization of receptive fields (RFs) in the visual, auditory, somatosensory and other sensory systems. In addition, training head-fixed non-human primates (Roelfsema et al., 1998; Spitzer et al., 1988; Treue and Maunsell, 1996), ferrets (Atiani et al., 2009; David et al., 2012) or rodents (Histed et al., 2012; Schwarz et al., 2011) in carefully designed behavioral tasks, which ensure a constant sensory drive under different cognitive demands, enabled in sensory areas the isolation of non-sensory, contextual modulations of neuronal responses, such as effects of attention, reward expectancies, or decision making. Performing these experiments in stationary animals was crucial, as it allowed stability of both sensory stimulation and neural recordings.
In general, studying perception under stationary conditions might particularly well capture sensory processes in primates, where a substantial portion of sensory experience involves focused information processing in quiet wakeful states, such as during communication, reading (in case of humans), and other forms of attentive information gathering. For many animals, however, important sensory processing happens mainly during physical activity, where sensation serves for navigation, finding food and mates, and avoiding predation, whereas periods of quietness are often associated with rest. Remarkably, some of the hallmarks of neural processing during active behaviors and focused attention seem shared, opening the possibility that some of the underlying circuits might be evolutionarily conserved (Harris and Thiele, 2011; Maimon, 2011).
The idea to investigate sensory processing during active behaviors is not new: For many years, research on visuo-motor integration has relied on an experimental approach, which combines the advantages of head-fixation with the ability of animals to move freely. This approach has been pioneered at the Max-Planck Institute for Biological Cybernetics (Tübingen), where it was first applied to the fly visual system (Buchner, 1976) (Figure 1a). It consists of an air-cushioned, floating ball, on which the fly can rest or move, and the surrounding stimulation system, where visual stimuli can be displayed dependently (closed loop) or independently (open loop) from the animal’s locomotion behavior. Recognizing the power of this spherical treadmill approach, Mallot and colleagues (University of Tübingen) (Holscher et al., 2005) more than 10 years ago have adapted the floating-ball-system for rats, and could demonstrate that rats are able to navigate in virtual environments displayed in the system. The application of two photon, single-cell-resolution population Ca2+ imaging (Dombeck et al., 2007), intracellular recordings (Harvey et al., 2009) and high-density extracellular recordings (Niell and Stryker, 2010), in mice actively behaving in this system, lead to the final break-through of the paradigm (Figure 1b–c). Remarkably, research on effects of active behaviors on neural processing in flies and rodents has also inspired human cognitive psychology, where new setups have now been developed, which allow measurement of task performance and brain activity during controlled physical activity, such as cycling on a stationary exercise bike (Bullock et al., 2016).

Locomotion during head fixation on treadmill systems. a) One of the first “floating ball setups” developed for the fly Drosophila, suspended on the ball via a micromanipulator. A fiber optics based system registers the movement of the animal by measuring the displacement of the patterns on the ball. Modified with permission from Buchner (1976). b) A similar system applied to mice. Modified with permission from Carandini and Churchland (2013). c) When combined with a large-scale immersive stimulation system, display of visual stimuli can occur in closed loop (matching the animal’s locomotion) or open loop configuration (mismatching the animal’s locomotion). Modified with permission from Fiser et al. (2016).
Modulations of sensory information processing
When observing head-fixed mice during locomotion with a close-up camera for tracking of eye movements, several groups have realized that locomotion goes along with prominent modulations of pupil size: while pupil size tends to be smaller during stationary periods, pupil size can be dramatically enlarged during locomotion (Erisken et al., 2014; Vinck et al., 2015; McGinley et al., 2015; Reimer et al., 2014). Pupil size has long been known from cognitive psychology as an excellent proxy of arousal (Bradshaw, 1967); hence, pupil dilation under constant illumination in mice during stationary periods has been explored as a marker to isolate effects of arousal (Vinck et al., 2015; McGinley et al., 2015; Reimer et al., 2014), which are likely mixed with more specific locomotion-related signals during running (Keller et al., 2012; Saleem et al., 2013).
The ecological relevance of pupil dilation with arousal or, more generally, demanding cognitive processes, is still an open question: on the one hand, pupil dilation increases the amount of light entering the eye, which can, e.g. in mice, amount to a more than 20-fold increase in retinal brightness (Pennesi et al., 1998). To which degree such increase in retinal brightness translates into increases in sensitivity is unclear, however, because there are neural circuits as early as in the retina, which exert gain control and normalization for overall luminance (Carandini and Heeger, 2012). On the other hand, pupil size under different illumination conditions is optimized for providing highest visual acuity (Laughlin, 1992), so further dilating the pupil in the context of arousal might hurt the quality of the image due to optical aberrations. In summary, the functional relevance of pupil dilation under constant illumination is still unclear, as direct evidence for a contribution of pupil size to enhanced visual sensitivity is still missing.
Visual cortex
Building on a long tradition of work on state-dependent neuronal processing in sensory systems (Harris and Thiele, 2011; Gilbert and Sigman, 2007; Alonso and Swadlow, 2017; Zagha and McCormick, 2014), Niell & Stryker (Niell and Stryker, 2010) used the spherical treadmill paradigm to demonstrate in extracellular recordings that neural responses in mouse primary visual cortex (V1) dramatically depend on whether the animal is sitting or running. More specifically, during locomotion broad-spiking neurons in V1 layer 2/3 can show a more than 2-fold increase in responsivity. This increase is typically an additive and multiplicative enhancement of gain (Figure 2a, b): V1 orientation tuning curves, for instance, are up-scaled during locomotion without exhibiting concomitant changes in tuning width, at least on the overall population level. Later studies revealed that the subthreshold mechanism underlying these state-dependent differences in V1 processing is a depolarization of the membrane potential of pyramidal cells (Niell and Stryker, 2010; Polack et al., 2013). This depolarization of membrane potential is accompanied by a remarkable reduction of trial-to-trial variability (Bennett et al., 2013), improving the reliability of visual evoked responses.
Besides increasing the gain of orientation tuning (Niell and Stryker, 2010; Polack et al., 2013) and contrast response functions (Erisken et al., 2014; Lee et al., 2014), locomotion can also change several aspects of V1 stimulus selectivity. First, in experiments probing V1 neurons with gratings of varying diameter, locomotion increases preferred stimulus size and markedly reduces surround suppression, thus controlling spatial integration and contextual modulation (Figure 2c) (Erisken et al., 2014; Ayaz et al., 2013). Second, locomotion-related gain changes seem largest in neurons tuned for high spatial frequencies, resulting in a relative enhancement of spatial resolution during locomotion (Mineault et al., 2016). Third, during locomotion, temporal frequency tuning of neurons in V1 and extrastriate area AL (anterolateral) shifts towards higher frequencies (Andermann et al., 2011). Forth, high arousal in the absence of locomotion indexed by pupil dilation leads to a sharpening of orientation tuning (Reimer et al., 2014). These changes in neuronal tuning properties make simple explanations for the observed effects of locomotion unlikely, such as those based on increases in brain temperature (Moser et al., 1993). In addition, since cortical temperature fluctuations with voluntary locomotion have been found to be small (0.1 °C), some authors conclude that the contribution of temperature changes during locomotion on neural excitability are minimal (Shirey et al., 2015).

Effects of locomotion on several V1 response properties. a) Locomotion increases orientation tuning curves. b) Locomotion exerts response gain control on contrast response functions. c) Locomotion alters spatial integration by increasing preferred RF center size and decreasing surround suppression. (a-c) Modified with permission from Erisken et al. (2014). d) Locomotion suppresses LFP power at low frequencies and enhances power at higher frequencies, including a narrow-band gamma oscillation. Fiorini & Busse, unpublished. e) Example V1 neurons with tuning to running speed. Erisken & Busse, unpublished (see also Saleem et al., 2013). f) Full-field and local mismatch responses of a population of V1 neurons. Modified with permission from Zmarz and Keller (2016).
Beyond strengthening of responses, locomotion improves stimulus encoding by enhancing the mutual information between visual stimuli and single neuron responses (Dadarlat and Stryker, 2017). The locomotion-dependent shift of the V1 population into more informative processing regimes does not only result from the increased amount of activity (Dadarlat and Stryker, 2017), but also from changes in the population response pattern, where noise correlations decrease (Erisken et al., 2014; Dadarlat and Stryker, 2017).
At least in the mouse visual system, the effects of locomotion seem to be layer-dependent. Locomotion-dependent increases of mutual information (Dadarlat and Stryker, 2017) and firing rates (Erisken et al., 2014; Dadarlat and Stryker, 2017) have been found largest and most consistent in supragranular cortical layers, while changes in firing rate with locomotion in infragranular layers seem more diverse and often of opposite sign, thus resulting in a smaller overall modulation (Erisken et al., 2014). The mechanisms and underlying neural circuits for the diversity of locomotion-related response modulations in lower layers are currently not well understood, but at least two proposals have been made: A cellular resolution, two-photon calcium imaging study using a microprism approach to reach all V1 layers revealed an intriguing population of neurons whose activity was strongly suppressed by locomotion (Andermann et al., 2013). Based on their location in the imaging plane, these locomotion-suppressed were attributed to L6 (Andermann et al., 2013). Interestingly, previous studies have shown that a particular class of L6 neurons (the Ntsr1-positive L6 neurons) can regulate the gain across the V1 column (Olsen et al., 2012) via reduced driving of a transcolumnar inhibitory interneuron (Bortone et al., 2014). Hence, a locomotion-related reduction of activity of the L6 Ntsr1+ neurons might be a potential mechanism for increased gain in the superficial layers. Other results point towards the fact that locomotion-suppressed neurons in deep layers might instead be located in L5 (Erisken et al., 2014); if these locomotion-suppressed neurons were somatostatin-positive (SOM+) inhibitory interneurons, they could provide gain modulations of the cortical column via disinhibition of pyramidal cell activity. Future experiments are needed to distinguish between these and other alternatives, and identify the deep-layer, locomotion-suppressed neurons.
Consistent with a change in overall brain state, periods of locomotion are accompanied by a marked change in the power distribution of the local field potential (LFP) (Figure 2d). Specifically, locomotion decreases the power of low frequencies (< 20 Hz) and increases power in the LFP gamma band range (> 40 Hz) (Niell and Stryker, 2010; Vinck et al., 2015), changing the LFP from exhibiting synchronized low frequency, high amplitude fluctuations during stationary periods to more desynchronized patterns of activity with high frequency, low amplitude fluctuations during locomotion. Interestingly, there is in mouse primary visual cortex, a remarkably narrow gamma band oscillation with a 2–5 Hz bandwidth peaking close to 60 Hz, whose amplitude increases with locomotion (Niell and Stryker, 2010; Lee et al., 2014; Saleem et al., 2017). This narrowband gamma oscillation is highest in L4 of primary visual cortex and is present in the spiking output of neurons in the dorsolateral geniculate nucleus (dLGN) of the thalamus, even in the absence of cortical feedback (Saleem et al, 2017), suggesting that the narrowband gamma oscillation is generated upstream from cortex, i.e in the thalamus or potentially in the retina (Storchi et al., 2017). While the exact function of this narrow-band gamma oscillation remains unknown, it might represent a specific channel for feedforward retino-geniculo-cortico communication (Saleem et al, 2017).
In addition to global changes in brain state, locomotion can also exert more specific influences in primary visual cortex. First, a sizeable fraction of the neurons in mouse V1 seem to be tuned for running speed, even when the animals are running in complete darkness (Erisken et al., 2014; Saleem et al., 2013): Running speed can control smoothly the firing rate of V1 neurons, with run speed tuning being monotonically increasing, monotonically decreasing or peaking at intermediate running speeds (Figure 2e). Such run speed tuning is consistent with the idea that V1 can inform navigation, as run speed is represented in V1, even in the absence of visual flow. Second, 5-10% of neurons in mouse primary visual cortex signal mismatches between self-motion and optic flow (Keller et al., 2012; Zmarz and Keller, 2016), by strongly responding to the sudden uncoupling of optic flow patterns from self motion (Figure 2f). In the context of theories of predictive coding (Friston, 2005; Rao and Ballard, 1999), mismatch responses can be interpreted to signal deviations from a prediction based on a learned relationship between motor output and sensory feedback (Attinger t al., 2017). Interestingly, mismatch responses can also be triggered from local changes in optic flow (Figure 2f), making these responses a candidate mechanism for detecting deviations from self-motion, such as generated by moving objects during locomotion (Zmarz and Keller, 2016).
Other sensory cortices
Modulations of cortical responses by locomotion have been most extensively investigated in the visual system, but they also occur in the somatosensory and auditory system. In the primary somatosensory cortex (S1), locomotion’s effects seem to be generally similar to those observed in primary visual cortex (Fu et al., 2014; Sofroniew et al., 2015; Pluta et al., 2015), and also resemble those typically found during whisking, which has been used extensively in the context of somatosensation to index active behavioral states (Crochet and Petersen, 2006; Gentet et al., 2010; Poulet and Petersen, 2008). In the primary auditory cortex (A1), however, the observed effects seem to be markedly different. Instead of enhancing responses, locomotion seems to decrease A1 responses (McGinley et al., 2015; Zhou et al., 2014; Schneider et al., 2014), both during spontaneous as well as stimulus-driven activity. This suppression of A1 spiking activity is mediated by a scaling down of both excitation and inhibition of subthreshold membrane potentials (McGinley et al., 2015; Schneider et al., 2014). Since this suppression is relatively stronger during spontaneous background activity than during sensory evoked responses, signal-to-noise ratio during auditory processing in the active state seems improved (McGinley et al., 2015; Zhou et al., 2014), in particular in regimes of moderate arousal (McGinley et al., 2015). Interestingly, not only locomotion, but also active task engagement seems to generally hyperpolarize primary auditory cortex (Otazu et al., 2009; Kuchibhotla et al., 2017).
Locomotion effects in thalamus
Effects of locomotion, similar to more general effects of arousal (Cano et al., 2006; Bezdudnaya et al., 2006), are not restricted to cortex, but already occur at the level of the sensory thalamic nuclei (Erisken et al., 2014; McGinley et al., 2015; Williamson et al., 2015). In the visual system, locomotion shifts the firing mode of neurons in the dorsolateral geniculate nucleus (dLGN) of the thalamus from burst to tonic mode (Niell and Stryker, 2010; Erisken et al., 2014), enhances firing rates (Erisken et al., 2014; Roth et al., 2016) and controls spatial integration (Erisken et al., 2014). Similar to neurons in area V1, thalamic neurons can signal speed-related information (Erisken et al., 2014; Roth et al., 2016): dLGN neurons show tuning for running speed (Erisken et al., 2014), while neurons in the higher-order visual thalamic lateral posterior (LP) nucleus, a rodent homologue of the pulvinar, can represent discrepancies between self motion and optic flow (Hirata and Castro-Alamancos, 2010). Accordingly, in the auditory system, locomotion decreases responses already in the medial geniculate body (McGinley et al., 2015; Williamson et al., 2015).
Given that locomotion modulates sensory processing across multiple stages, an important open question is to which degree these modulations are carried over across processing stages via glutamatergic connections or generated de novo. In the auditory system, Schneider et al. (Hirata and Castro-Alamancos, 2010) estimated the cortical contribution to locomotion-related response modulation by using a powerful optogenetic stimulation approach: they found that locomotion effects on responses to optogenetic stimulation of afferent thalamo-cortical connections from the medial geniculate body over A1 were around 60% of the locomotion effects on responses to tones, pointing towards the importance of both cascading effects between processing stages and cortical contributions. Focused on more general state-dependent processing, similar results have been found in the somatosensory system, where stimulation of ventral posterior medial (VPM) thalamus can drive neuronal activity in barrel cortex typically associated with active whisking (Hirata and Castro-Alamancos, 2010; Poulet et al., 2012; but see, Constantinople and Bruno, 2011).
Effects of locomotion on behavioral performance
What are the effects of the locomotion-related improvements in sensory representations on behavior? In the visual system, the enhanced encoding during locomotion translates into improved behavioral sensitivity for stimulus detection, in particular during challenging low-contrast conditions (Bennett et al., 2013). For perceptual studies in the visual system, so far, the animal’s activity has largely been considered a binary variable, where performance has been measured during either stationary or locomotion conditions. Work in the auditory system, to date, has quantified behavioral performance as a function of behavioral state in a much more fine-grained way, and concluded that auditory detection performance follows the classically described, inverted U-shaped dependence on arousal (McGinley et al., 2015): performance is optimal for intermediate levels of arousal as indexed by pupil diameter, which occur in a sub-state of quiet wakefulness; in contrast, auditory performance during locomotion, i.e. a high-arousal state, or during drowsiness, is degraded. Intriguingly, a similar, inverted U-shaped relationship between visual feature-selective coding and exercise-level has recently been observed in the human visual system (Bullock et al., 2016).
Neural circuits mediating locomotion effects on sensory processing
Two mechanisms have recently received particular attention in the study of the neural circuits mediating locomotion effects on sensory processing: neuromodulation and inhibitory interneurons. These mechanisms are likely related, as inhibitory interneurons seem to be particularly prone to the influences of neuromodulation (Fanselow et al., 2008; Chen et al., 2015; Kruglikov and Rudy, 2008).
There is a substantial body of converging work demonstrating the role of acetylcholine in the expression of state-dependent neural processing (Thiele, 2013). Stimulation of basal forebrain can mimic in V1 some of the key effects of locomotion, alongside improving visual performance (Pinto et al., 2013). In fact, to evoke the V1 response enhancements typically associated with locomotion, it is sufficient to activate the axons terminating in basal forebrain from the mesencephalic locomotor region (MLR) (Lee et al., 2014), a brain stem region involved in the execution of locomotor patterns. Importantly, this recapitulation of V1 locomotion effects is possible even in subtle stimulations, where the descending locomotor outputs are not recruited (Lee et al., 2014). Consistent with the similarity of effects of locomotion with those of basal forebrain stimulation, activity of cholinergic axons imaged over V1 and A1 is correlated to locomotion (Reimer et al., 2016). Other work, however, has found that application of cholinergic antagonists could not abolish in L2/3 the tonic depolarization associated with locomotion (Polack et al., 2013), suggesting the involvement of additional neuromodulatory pathways. Given the prominent effects of locomotion on pupil size, another candidate neuromodulator is norepinephrine, which is well known to contribute to controlling pupil size via tonic activity in the locus coeruleus (Aston-Jones and Cohen, 2005; Samuels and Szabadi, 2008). Indeed, pharmacological blockade of noradrenergic inputs in V1 superficial layers seems to reduce locomotion-related depolarization by around 50% (Polack et al., 2013), and activity of noradrenergic axons over V1 seems well correlated to the fast changes of pupil size (Reimer et al., 2016). While it is sometimes implicitly assumed that the links between neuromodulatory brain stem centers and cortex is direct, neuromodulation also has substantial impact on thalamus and thalamocortical transmission: First, cholinergic projections from the parabrachial brainstem region, which also encompasses the MLR, terminate in dLGN, as well as noradrenergic projections from the locus coeruleus (reviewed in Lee and Dan, 2012). Second, GABAergic basal forebrain projections terminate in the TRN, where they exert hyperpolarizing influences (Bickford et al., 1994), potentially releasing sensory thalamic nuclei from inhibition. In summary, while it has become clear that several different circuits are involved in the modulation of sensory processing, it remains an open question how these circuits concert their actions during locomotion (Figure 3a).
Remarkably, neuromodulation via the noradrenergic system during locomotion combined with local synaptic activity, e.g. during visual stimulation, do not only affect neuronal responses, but also promote slower and long-lasting Ca2+ responses in the V1 astroglia network via the alpha-adrenergic receptor (Paukert et al., 2014; Ding et al., 2013). Increased Ca2+ signaling in astrocytes during coincident synaptic activity and arousal might enhance the ability of astrocytes to uptake K+ and glutamate from the extracellular space, thereby contributing to the control of excitability of neural circuits and the coordination of the broad effects of neuromodulators on neuronal activity (Paukert et al., 2014; Ding et al., 2013; Kjaerby et al., 2017).
What could be the possible routes of neuromodulatory action? In primary visual cortex, acetylcholine might have preferential impact on inhibitory interneurons (Fanselow, 2008; Chen et al., 2015; Kruglikov and Rudy, 2008), and could thereby change the recruitment of GABAergic inhibition and the function of the cortical circuit. According to a well-established circuit motif (Pfeffer et al., 2013), which seems to be conserved across different cortical areas and recruited in different behavioral contexts from gain control to sensorimotor integration and plasticity (Fu et al., 2014; Pi et al., 2013; Lee et al., 2013; Fu et al., 2015), cholinergic activation of vasoactive intestinal peptide positive (VIP+) inhibitory interneurons leads to inhibition of somatostatin positive (SOM+) inhibitory interneurons, which ultimately results in a disinhibition of pyramidal cell activity (Figure 3b). In relatively close accordance with the hypothesized circuit, during running in darkness, VIP+ interneurons in area V1 are generally excited, while SOM+ interneurons can be inhibited (Reimer et al., 2014; Fu et al., 2014; Pakan et al., 2016) (Figure 3c, left). Note, however, that while the VIP->SOM->pyramidal cell disinhibitory circuit can account during running in darkness for some of the average V1 locomotion effects, the distribution of locomotion effects in both interneuron types are broad (Fu et al., 2014; Pakan et al., 2016). Furthermore, the disinhibitory circuit model is challenged by novel findings that while running during visual stimulation, activity of both VIP+ and SOM+ interneurons is enhanced (Pakan et al., 2016) (Figure 3c, right).
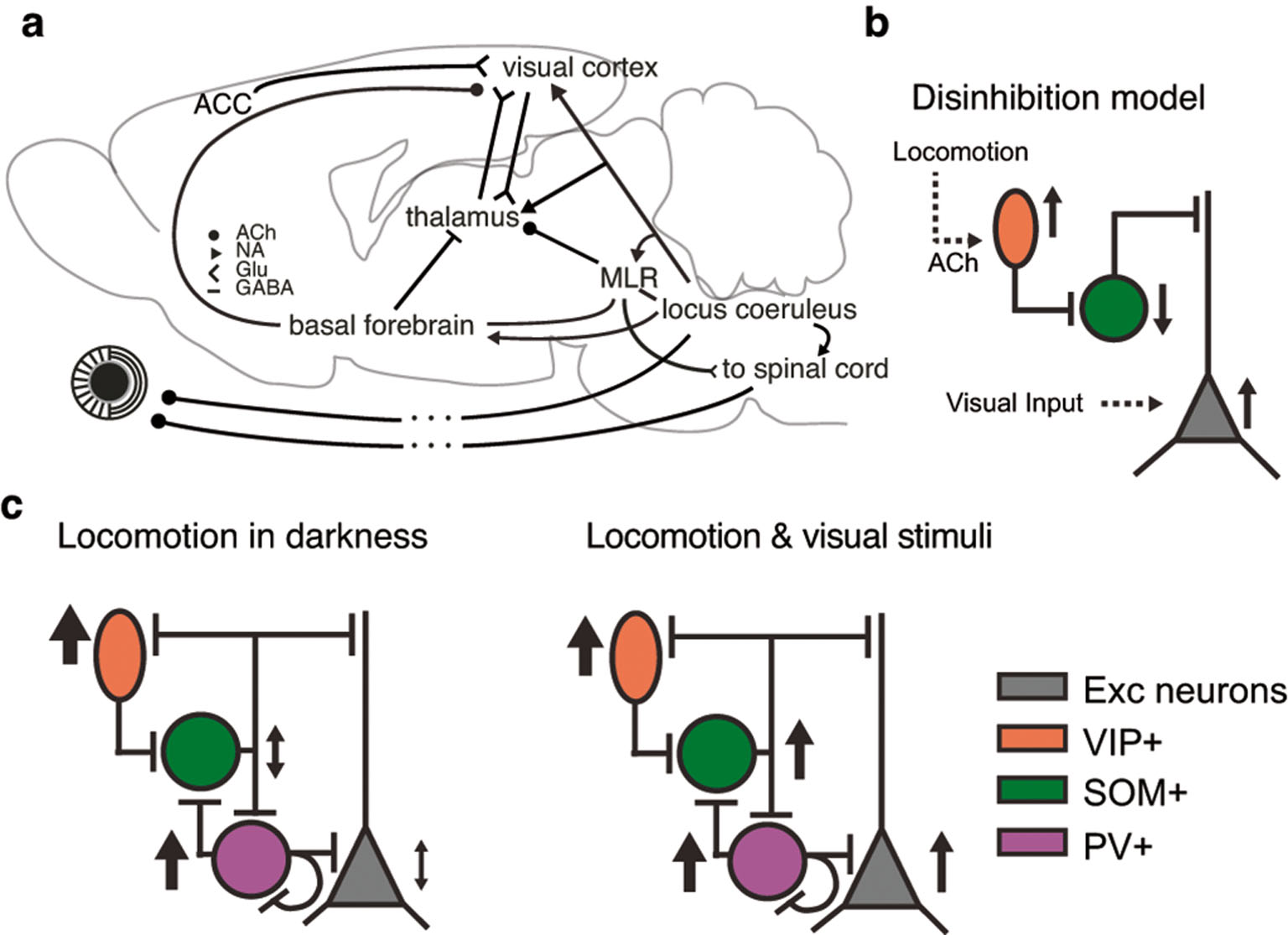
Disinhibitory circuit proposed to account for locomotion effects. a) It is likely that multiple neural circuits contribute to the modulation of sensory information processing by locomotion. ACh: Acetylcholine, NA: norepinephrine, Glu: glutamate, MLR: mesencephalic locomotor region, ACC: anterior cinguiate cortex. Modified with permission from Erisken et al. (2014) b) A proposed disinhibitory pathway: ACh released during locomotion activates VIP+ inhibitory interneurons, which in turn inhibit SOM+ inhibitory interneurons. This leads to disinhibition of pyramidal cell activity. c) Experimental observations during running in darkness (left) and during visual stimulation (right) conflicting with the proposed disinhibitory circuit. Modified with permission from Pakan et al. (2016).
A detailed assessment of the action of the VIP->SOM->pyramidal neuron disinhibitory circuit and an account of the observed variability within interneuron types has been recently provided in the somatosensory system (Muñoz et al., 2017). Here, Munoz et al. (Muñoz et al., 2017) could show that a particular subtype of SOM+ interneurons, with axonal arborizations in L1 and L5a, is specifically inhibited during whisking, while other SOM+ interneuron subtypes show opposite effects. Intriguingly, L5a of S1 is one of the major targets of L2/3 VIP+ interneurons (Zhang et al., 2016), and whisking-suppressed SOM+ interneurons receive specifically strong inhibition via VIP+ interneurons (Muñoz et al., 2017).
Neuromodulatory influences during locomotion might be accompanied by more specific top-down feedback modulations (Zhang et al., 2016; Zhang et al., 2014)and influences from motor efference copies (Schneider et al., 2014; Leinweber et al., 2017). Indeed, primary visual cortex receives strong input from a subdivision of the anterior cingulate (ACC) called A24b, and secondary motor cortex (M2), which provide motor-related signals dependent on visual experience (Ding et al., 2013) and can be interpreted as representing a prediction signal based on expected visual flow.
Future perspectives
The consideration of locomotion in studies of sensory processing has allowed the discovery of signals that can not only modulate neural responses but also serve active behaviors, like navigation. Yet, locomotion under head fixation still is a relatively reduced preparation compared to locomotion under real-life conditions. For instance, vestibular influences, which are known to have widespread influences throughout the rodent cortex (Rancz et al., 2015) are missing under head fixation. A combination of technological advances, including miniaturization of recording equipment (Kerr and Nimmerjahn., 2012), head-mounted eye tracking (Wallace et al., 2013), low-latency immersive virtual reality systems (Grosso et al., 2017; Stowers et al., 2017), and methods from machine learning for automatic animal tracking and detailed and accurate segmentation of behavior (Valletta et al., 2017), promises to allow in the future the combination of controlled sensory stimulation under complex, natural, and evolutionary relevant behaviors.
About the author

Laura Busse has been professor for “Organismic Neurobiology” at the Division of Neurobiology of the Department of Biology II at the LMU Munich since 2016 and is affiliated with the Bernstein Center for Computational Neuroscience. She obtained her PhD from the University of Göttingen for work at the German Primate Center, was a postdoctoral fellow at the Smith Kettlewell Eye Research Institute in San Francisco and at the UCL Institute of Ophthalmology in London. Between 2010 and 2015 Laura Busse was a Junior Research Group Leader at the Centre for Integrative Neuroscience (CIN) at the University of Tübingen.
References
Alonso, J. M., Swadlow, H. A. (2017). Thalamocortical Interactions for Sensory Processing. doi: 10.1093/acrefore/9780190264086.013.11210.1093/acrefore/9780190264086.013.112Suche in Google Scholar
Andermann, M. L., Gilfoy, N. B., Goldey, G. J., et al. (2013). Chronic cellular imaging of entire cortical columns in awake mice using microprisms. Neuron 80, 900–13, doi: 10.1016/j.neuron.2013.07.05210.1016/j.neuron.2013.07.052Suche in Google Scholar PubMed PubMed Central
Andermann, M. L., Kerlin, A. M., Roumis, D. K., et al. (2011). Functional Specialization of Mouse Higher Visual Cortical Areas. Neuron 72, 1025–103910.1016/j.neuron.2011.11.013Suche in Google Scholar PubMed PubMed Central
Aston-Jones, G., Cohen, J. D. (2005). AN INTEGRATIVE THEORY OF LOCUS COERULEUS-NOREPINEPHRINE FUNCTION: Adaptive Gain and Optimal Performance. Annu. Rev. Neurosci. 28, 403–450, doi: 10.1146/annurev.neuro.28.061604.13570910.1146/annurev.neuro.28.061604.135709Suche in Google Scholar PubMed
Atiani, S., Elhilali, M., David, S. V., et al. (2009). Task Difficulty and Performance Induce Diverse Adaptive Patterns in Gain and Shape of Primary Auditory Cortical Receptive Fields. Neuron 61, 467–48010.1016/j.neuron.2008.12.027Suche in Google Scholar PubMed PubMed Central
Attinger, A., Wang, B., Keller, G. B. (2017). Visuomotor Coupling Shapes the Functional Development of Mouse Visual Cortex. Cell 169, 1291–1302.e14, doi: 10.1016/j.cell.2017.05.02310.1016/j.cell.2017.05.023Suche in Google Scholar PubMed
Ayaz, A., Saleem, A. B., Schölvinck, M. L., Carandini, M. (2013). Locomotion Controls Spatial Integration in Mouse Visual Cortex. Curr Biol 23, 890–894, doi: 10.1016/j.cub.2013.04.01210.1016/j.cub.2013.04.012Suche in Google Scholar PubMed PubMed Central
Bennett, C., Arroyo, S., Hestrin, S. (2013). Subthreshold Mechanisms Underlying State-Dependent Modulation of Visual Responses. Neuron 80, 350–35710.1016/j.neuron.2013.08.007Suche in Google Scholar PubMed PubMed Central
Bezdudnaya, T., Cano, M., Bereshpolova, Y., et al. (2006). Thalamic Burst Mode and Inattention in the Awake LGNd. Neuron 49, 421–432, doi: 10.1016/j.neuron.2006.01.01010.1016/j.neuron.2006.01.010Suche in Google Scholar PubMed
Bickford, M. E., Günlük, A. E., Van Horn, S. C., Sherman, S. M. (1994). GABAergic projection from the basal forebrain to the visual sector of the thalamic reticular nucleus in the cat. J. Comp. Neurol. 348, 481–510, doi: 10.1002/cne.90348040210.1002/cne.903480402Suche in Google Scholar PubMed
Bortone, D. S., Olsen, S. R., Scanziani, M. (2014). Translaminar inhibitory cells recruited by layer 6 corticothalamic neurons suppress visual cortex. Neuron 82, 474–85, doi: 10.1016/j.neuron.2014.02.02110.1016/j.neuron.2014.02.021Suche in Google Scholar PubMed PubMed Central
Bradshaw, J. (1967). Pupil Size as a Measure of Arousal during Information Processing. Nature 216, 515–516, doi: 10.1038/216515a010.1038/216515a0Suche in Google Scholar PubMed
Buchner, E. (1976). Elementary movement detectors in an insect visual system. Biol Cybern 24, 85–101, doi: 10.1007/BF0036064810.1007/BF00360648Suche in Google Scholar
Bullock, T., Elliott, J. C., Serences, J. T., Giesbrecht, B. (2016). Acute Exercise Modulates Feature-selective Responses in Human Cortex. J. Cogn. Neurosci. 1–14, doi: 10.1162/jocn_a_0108210.1162/jocn_a_01082Suche in Google Scholar PubMed
Cano, M., Bezdudnaya, T., Swadlow, H. A., Alonso, J.-M. (2006). Brain state and contrast sensitivity in the awake visual thalamus. Nat. Neurosci. 9, 1240–124210.1038/nn1760Suche in Google Scholar PubMed
Carandini, M., Churchland, A. K. (2013). Probing perceptual decisions in rodents. Nat. Neurosci. 16, 824–31, doi: 10.1038/nn.341010.1038/nn.3410Suche in Google Scholar PubMed PubMed Central
Carandini, M., Heeger, D. J. (2012). Normalization as a canonical neural computation. Nat. Rev. Neurosci. 13, 51–6210.1038/nrn3136Suche in Google Scholar PubMed PubMed Central
Chen, N., Sugihara, H., Sur, M. (2015). An acetylcholine-activated microcircuit drives temporal dynamics of cortical activity. Nat. Neurosci. 18, 892–902, doi: 10.1038/nn.400210.1038/nn.4002Suche in Google Scholar PubMed PubMed Central
Constantinople, C. M., Bruno, R. M. (2011). Effects and mechanisms of wakefulness on local cortical networks. Neuron 69, 1061–8, doi: 10.1016/j.neuron.2011.02.04010.1016/j.neuron.2011.02.040Suche in Google Scholar PubMed PubMed Central
Crochet, S., Petersen, C. C. H. (2006). Correlating whisker behavior with membrane potential in barrel cortex of awake mice. Nat. Neurosci. 9, 608–610, doi: 10.1038/nn169010.1038/nn1690Suche in Google Scholar PubMed
Dadarlat, M. C., Stryker, M. P. (2017). Locomotion Enhances Neural Encoding of Visual Stimuli in Mouse V1. J. Neurosci. 37, 3764–3775, doi: 10.1523/JNEUROSCI.2728-16.201710.1523/JNEUROSCI.2728-16.2017Suche in Google Scholar PubMed PubMed Central
David, S. V., Fritz, J. B., Shamma, S. A. (2012). Task reward structure shapes rapid receptive field plasticity in auditory cortex. Proc. Natl. Acad. Sci. 109, 2144–214910.1073/pnas.1117717109Suche in Google Scholar PubMed PubMed Central
Ding, F., O’Donnell, J., Thrane, A. S., et al. (2013). α1-Adrenergic receptors mediate coordinated Ca2+ signaling of cortical astrocytes in awake, behaving mice. Cell Calcium 54, 387–394, doi: 10.1016/j.ceca.2013.09.00110.1016/j.ceca.2013.09.001Suche in Google Scholar PubMed PubMed Central
Dombeck, D. A., Khabbaz, A. N., Collman, F., et al. (2007). Imaging Large-Scale Neural Activity with Cellular Resolution in Awake, Mobile Mice. Neuron 56, 43–5710.1016/j.neuron.2007.08.003Suche in Google Scholar PubMed PubMed Central
Erisken, S., Vaiceliunaite, A., Jurjut, O., et al. (2014). Effects of Locomotion Extend throughout the Mouse Early Visual System. Curr Biol 24, 2899–2907, doi: 10.1016/j.cub.2014.10.04510.1016/j.cub.2014.10.045Suche in Google Scholar PubMed
Fanselow, E. E., Richardson, K. A., Connors, B. W. (2008). Selective, State-Dependent Activation of Somatostatin-Expressing Inhibitory Interneurons in Mouse Neocortex. J. Neurophysiol. 100, 2640–2652, doi: 10.1152/jn.90691.200810.1152/jn.90691.2008Suche in Google Scholar PubMed PubMed Central
Fiser, A., Mahringer, D., Oyibo, H. K., et al. (2016). Experience-dependent spatial expectations in mouse visual cortex. Nat. Neurosci. 19, 1658–1664, doi: 10.1038/nn.438510.1038/nn.4385Suche in Google Scholar PubMed
Friston, K. (2005). A theory of cortical responses. Philos Trans R Soc B Biol Sci 360, 815–83610.1098/rstb.2005.1622Suche in Google Scholar PubMed PubMed Central
Fu, Y., Kaneko, M., Tang, Y., et al. (2015). A cortical disinhibitory circuit for enhancing adult plasticity. eLife 4, e05558, doi: 10.7554/eLife.0555810.7554/eLife.05558Suche in Google Scholar PubMed PubMed Central
Fu, Y., Tucciarone, J. M., Espinosa, J. S., et al. (2014). A cortical circuit for gain control by behavioral state. Cell 156, 1139–52, doi: 10.1016/j.cell.2014.01.05010.1016/j.cell.2014.01.050Suche in Google Scholar PubMed PubMed Central
Gentet, L. J., Avermann, M., Matyas, F., et al. (2010). Membrane Potential Dynamics of GABAergic Neurons in the Barrel Cortex of Behaving Mice. Neuron 65, 422–43510.1016/j.neuron.2010.01.006Suche in Google Scholar PubMed
Gilbert, C. D., Sigman, M. (2007). Brain States: Top-Down Influences in Sensory Processing. Neuron 54, 677–69610.1016/j.neuron.2007.05.019Suche in Google Scholar PubMed
Grosso, N. A. D., Graboski, J. J., Chen, W., et al. (2017). Virtual Reality system for freely-moving rodents. bioRxiv 161232, doi: 10.1101/16123210.1101/161232Suche in Google Scholar
Harris, K. D., Thiele, A. (2011). Cortical state and attention. Nat. Rev. Neurosci. 12, 509–512, doi: 10.1038/nrn308410.1038/nrn3084Suche in Google Scholar PubMed PubMed Central
Harvey, C. D., Collman, F., Dombeck, D. A., Tank, D. W. (2009). Intracellular dynamics of hippocampal place cells during virtual navigation. Nature 461, 941–94610.1038/nature08499Suche in Google Scholar PubMed PubMed Central
Hirata, A., Castro-Alamancos, M. A. (2010). Neocortex Network Activation and Deactivation States Controlled by the Thalamus. J. Neurophysiol. 103, 1147–1157, doi: 10.1152/jn.00955.200910.1152/jn.00955.2009Suche in Google Scholar PubMed PubMed Central
Histed, M. H., Carvalho, L. A., Maunsell, J. H. R. (2012). Psychophysical measurement of contrast sensitivity in the behaving mouse. J. Neurophysiol. 107, 758–65, doi: 10.1152/jn.00609.201110.1152/jn.00609.2011Suche in Google Scholar PubMed PubMed Central
Holscher, C., Schnee, A., Dahmen, H., et al. (2005). Rats are able to navigate in virtual environments. J. Exp. Biol. 208, 561–56910.1242/jeb.01371Suche in Google Scholar PubMed
Keller, G. B., Bonhoeffer, T., Hübener, M. (2012). Sensorimotor mismatch signals in primary visual cortex of the behaving mouse. Neuron 74, 809–15, doi: 10.1016/j.neuron.2012.03.04010.1016/j.neuron.2012.03.040Suche in Google Scholar PubMed
Kerr, J. N., Nimmerjahn, A. (2012). Functional imaging in freely moving animals. Curr Opin Neurobiol 22, 45–53, doi: 10.1016/j.conb.2011.12.00210.1016/j.conb.2011.12.002Suche in Google Scholar PubMed
Kjaerby, C., Rasmussen, R., Andersen, M., Nedergaard, M. (2017). Does Global Astrocytic Calcium Signaling Participate in Awake Brain State Transitions and Neuronal Circuit Function? Neurochem Res 42, 1810–1822, doi: 10.1007/s11064-017-2195-y10.1007/s11064-017-2195-ySuche in Google Scholar PubMed PubMed Central
Kruglikov, I., Rudy, B. (2008). Perisomatic GABA Release and Thalamocortical Integration onto Neocortical Excitatory Cells Are Regulated by Neuromodulators. Neuron 58, 911–924, doi: 10.1016/j.neuron.2008.04.02410.1016/j.neuron.2008.04.024Suche in Google Scholar PubMed PubMed Central
Kuchibhotla, K. V., Gill, J. V., Lindsay, G. W., et al. (2017). Parallel processing by cortical inhibition enables context-dependent behavior. Nat. Neurosci. 20, 62–7110.1038/nn.4436Suche in Google Scholar PubMed PubMed Central
Laughlin, S. B. (1992). Retinal information capacity and the function of the pupil. Ophthalmic Physiol Opt 12, 161–164, doi: 10.1111/j.1475-1313.1992.tb00281.x10.1111/j.1475-1313.1992.tb00281.xSuche in Google Scholar PubMed
Lee, S.-H., Dan, Y. (2012). Neuromodulation of Brain States. Neuron 76, 209–222, doi: 10.1016/j.neuron.2012.09.01210.1016/j.neuron.2012.09.012Suche in Google Scholar PubMed PubMed Central
Lee, A. M., Hoy, J. L., Bonci, A., et al. (2014). Identification of a Brainstem Circuit Regulating Visual Cortical State in Parallel with Locomotion. Neuron 83, 455–466, doi: 10.1016/j.neuron.2014.06.03110.1016/j.neuron.2014.06.031Suche in Google Scholar PubMed PubMed Central
Lee, S., Kruglikov, I., Huang, Z. J., et al. (2013). A disinhibitory circuit mediates motor integration in the somatosensory cortex. Nat. Neurosci. 16, 1662–1670, doi: 10.1038/nn.354410.1038/nn.3544Suche in Google Scholar PubMed PubMed Central
Leinweber, M., Ward, D. R., Sobczak, J. M., et al. (2017). A Sensorimotor Circuit in Mouse Cortex for Visual Flow Predictions. Neuron 95, 1420–1432.e5, doi: 10.1016/j.neuron.2017.08.03610.1016/j.neuron.2017.08.036Suche in Google Scholar PubMed
Maimon, G. (2011). Modulation of visual physiology by behavioral state in monkeys, mice, and flies. Curr Opin Neurobiol 21, 559–64, doi: 10.1016/j.conb.2011.05.00110.1016/j.conb.2011.05.001Suche in Google Scholar PubMed
McGinley, M. J., David, S. V., McCormick, D. A. (2015). Cortical Membrane Potential Signature of Optimal States for Sensory Signal Detection. Neuron 87, 179–192, doi: 10.1016/j.neuron.2015.05.03810.1016/j.neuron.2015.05.038Suche in Google Scholar PubMed PubMed Central
Mineault, P. J., Tring, E., Trachtenberg, J. T., Ringach, D. L. (2016). Enhanced Spatial Resolution During Locomotion and Heightened Attention in Mouse Primary Visual Cortex. J. Neurosci. 36, 6382–6392, doi: 10.1523/JNEUROSCI.0430-16.201610.1523/JNEUROSCI.0430-16.2016Suche in Google Scholar PubMed PubMed Central
Moser, E., Mathiesen, I., Andersen, P. (1993). Association between brain temperature and dentate field potentials in exploring and swimming rats. Science 259, 1324–1326, doi: 10.1126/science.844690010.1126/science.8446900Suche in Google Scholar PubMed
Muñoz, W., Tremblay, R., Levenstein, D., Rudy, B. (2017). Layer-specific modulation of neocortical dendritic inhibition during active wakefulness. Science 355, 954–959, doi: 10.1126/science.aag259910.1126/science.aag2599Suche in Google Scholar PubMed
Niell, C. M., Stryker, M. P. (2010). Modulation of Visual Responses by Behavioral State in Mouse Visual Cortex. Neuron 65, 472–47910.1016/j.neuron.2010.01.033Suche in Google Scholar PubMed PubMed Central
Olsen, S. R., Bortone, D. S., Adesnik, H., Scanziani, M. (2012). Gain control by layer six in cortical circuits of vision. Nature 483, 47–52, doi: 10.1038/nature1083510.1038/nature10835Suche in Google Scholar PubMed PubMed Central
Otazu, G. H., Tai, L.-H., Yang, Y., Zador, A. M. (2009). Engaging in an auditory task suppresses responses in auditory cortex. Nat. Neurosci. 12, 646–65410.1038/nn.2306Suche in Google Scholar PubMed PubMed Central
Pakan, J. M., Lowe, S. C., Dylda, E., et al. (2016). Behavioral-state modulation of inhibition is context-dependent and cell type specific in mouse visual cortex. eLife 5, e14985, doi: 10.7554/eLife.1498510.7554/eLife.14985Suche in Google Scholar PubMed PubMed Central
Paukert, M., Agarwal, A., Cha, J., et al. (2014). Norepinephrine Controls Astroglial Responsiveness to Local Circuit Activity. Neuron 82, 1263–1270, doi: 10.1016/j.neuron.2014.04.03810.1016/j.neuron.2014.04.038Suche in Google Scholar PubMed PubMed Central
Pennesi, M. E., Lyubarsky, A. L., Pugh, E. N. (1998). Extreme responsiveness of the pupil of the dark-adapted mouse to steady retinal illumination. Invest Ophthalmol Vis Sci 39, 2148–2156Suche in Google Scholar PubMed
Pfeffer, C. K., Xue, M., He, M., et al. (2013). Inhibition of inhibition in visual cortex: the logic of connections between molecularly distinct interneurons. Nat. Neurosci. 16, 1068–76, doi: 10.1038/nn.344610.1038/nn.3446Suche in Google Scholar PubMed PubMed Central
Pi, H.-J., Hangya, B., Kvitsiani, D., et al. (2013). Cortical interneurons that specialize in disinhibitory control. Nature 503, 521–524, doi: 10.1038/nature1267610.1038/nature12676Suche in Google Scholar PubMed PubMed Central
Pinto, L., Goard, M. J., Estandian, D., et al. (2013). Fast modulation of visual perception by basal forebrain cholinergic neurons. Nat. Neurosci. 16, 1857–1863, doi: 10.1038/nn.355210.1038/nn.3552Suche in Google Scholar PubMed PubMed Central
Pluta, S., Naka, A., Veit, J., et al. (2015). A direct translaminar inhibitory circuit tunes cortical output. Nat. Neurosci. 18, 1631–1640, doi: 10.1038/nn.412310.1038/nn.4123Suche in Google Scholar PubMed PubMed Central
Polack, P.-O., Friedman, J., Golshani, P. (2013). Cellular mechanisms of brain state-dependent gain modulation in visual cortex. Nat. Neurosci. 16, 1331–9, doi: 10.1038/nn.346410.1038/nn.3464Suche in Google Scholar PubMed PubMed Central
Poulet, J. F. A., Fernandez, L. M. J., Crochet, S., Petersen, C. C. H. (2012). Thalamic control of cortical states. Nat. Neurosci. 15, 370–37210.1038/nn.3035Suche in Google Scholar PubMed
Poulet, J. F. A., Petersen, C. C. H. (2008). Internal brain state regulates membrane potential synchrony in barrel cortex of behaving mice. Nature 454, 881–88510.1038/nature07150Suche in Google Scholar PubMed
Prönneke, A., Scheuer, B., Wagener, R. J., et al. (2015). Characterizing VIP Neurons in the Barrel Cortex of VIPcre/tdTomato Mice Reveals Layer-Specific Differences. Cereb Cortex 25, 4854–4868, doi: 10.1093/cercor/bhv20210.1093/cercor/bhv202Suche in Google Scholar PubMed PubMed Central
Rancz, E. A., Moya, J., Drawitsch, F., et al. (2015). Widespread Vestibular Activation of the Rodent Cortex. J. Neurosci. 35, 5926–5934, doi: 10.1523/JNEUROSCI.1869-14.201510.1523/JNEUROSCI.1869-14.2015Suche in Google Scholar PubMed PubMed Central
Rao, R. P. N., Ballard, D. H. (1999). Predictive coding in the visual cortex: a functional interpretation of some extra-classical receptive-field effects. Nat. Neurosci. 2, 79–87, doi: 10.1038/458010.1038/4580Suche in Google Scholar PubMed
Reimer, J., Froudarakis, E., Cadwell, C. R., et al. (2014). Pupil Fluctuations Track Fast Switching of Cortical States during Quiet Wakefulness. Neuron 84, 355–362, doi: 10.1016/j.neuron.2014.09.03310.1016/j.neuron.2014.09.033Suche in Google Scholar PubMed PubMed Central
Reimer, J., McGinley, M. J., Liu, Y., et al. (2016). Pupil fluctuations track rapid changes in adrenergic and cholinergic activity in cortex. Nat Commun 7, 13289, doi: 10.1038/ncomms1328910.1038/ncomms13289Suche in Google Scholar PubMed PubMed Central
Roelfsema, P. R., Lamme, V. A., Spekreijse, H. (1998). Object-based attention in the primary visual cortex of the macaque monkey. Nature 395, 376–38110.1038/26475Suche in Google Scholar PubMed
Roth, M. M., Dahmen, J. C., Muir, D. R., et al. (2016). Thalamic nuclei convey diverse contextual information to layer 1 of visual cortex. Nat. Neurosci. 19, 299–307, doi: 10.1038/nn.419710.1038/nn.4197Suche in Google Scholar PubMed PubMed Central
Saleem, A. B., Ayaz, A., Jeffery, K. J., et al. (2013). Integration of visual motion and locomotion in mouse visual cortex. Nat. Neurosci. 16, 1864–9, doi: 10.1038/nn.356710.1038/nn.3567Suche in Google Scholar PubMed PubMed Central
Saleem, A. B., Lien, A. D., Krumin, M., et al. (2017). Subcortical Source and Modulation of the Narrowband Gamma Oscillation in Mouse Visual Cortex. Neuron 93, 315–322, doi: 10.1016/j.neuron.2016.12.02810.1016/j.neuron.2016.12.028Suche in Google Scholar PubMed PubMed Central
Samuels, E. R., Szabadi, E. (2008). Functional neuroanatomy of the noradrenergic locus coeruleus: its roles in the regulation of arousal and autonomic function part II: physiological and pharmacological manipulations and pathological alterations of locus coeruleus activity in humans. Curr Neuropharmacol 6, 254–285, doi: 10.2174/15701590878577719310.2174/157015908785777193Suche in Google Scholar PubMed PubMed Central
Schneider, D. M., Nelson, A., Mooney, R. (2014). A synaptic and circuit basis for corollary discharge in the auditory cortex. Nature 513, 189–194, doi: 10.1038/nature1372410.1038/nature13724Suche in Google Scholar PubMed PubMed Central
Schwarz, C., Hentschke, H., Butovas, S. et al. (2011). The head-fixed behaving rat – Procedures and pitfalls. Somatosensory & Motor Research 27, 131–148, doi: 10.3109/08990220.2010.51311110.3109/08990220.2010.513111Suche in Google Scholar PubMed PubMed Central
Shirey, M. J., Smith, J. B., Kudlik, D. E., et al. (2015). Brief anesthesia, but not voluntary locomotion, significantly alters cortical temperature. J. Neurophysiol. 114, 309–322, doi: 10.1152/jn.00046.201510.1152/jn.00046.2015Suche in Google Scholar PubMed PubMed Central
Sofroniew, N. J., Vlasov, Y. A., Hires, S. A., et al. (2015). Neural coding in barrel cortex during whisker-guided locomotion. eLife 4, e12559, doi: 10.7554/eLife.1255910.7554/eLife.12559Suche in Google Scholar PubMed PubMed Central
Spitzer, H., Desimone, R., Moran, J. (1988). Increased attention enhances both behavioral and neuronal performance. Science 240, 338–34010.1126/science.3353728Suche in Google Scholar PubMed
Storchi, R., Bedford, R. A., Martial, F. P., et al. (2017). Modulation of Fast Narrowband Oscillations in the Mouse Retina and dLGN According to Background Light Intensity. Neuron 93, 299–307, doi: 10.1016/j.neuron.2016.12.02710.1016/j.neuron.2016.12.027Suche in Google Scholar PubMed
Stowers, J. R., Hofbauer, M., Bastien, R., et al. (2017). Virtual reality for freely moving animals. Nat Methods advance online publication:, doi: 10.1038/nmeth.439910.1038/nmeth.4399Suche in Google Scholar PubMed PubMed Central
Thiele, A. (2013). Muscarinic Signaling in the Brain. Annu. Rev. Neurosci. 36, 271–294, doi: 10.1146/annurev-neuro-062012-17043310.1146/annurev-neuro-062012-170433Suche in Google Scholar PubMed
Treue, S., Maunsell, J. H. R. (1996). Attentional modulation of visual motion processing in cortical areas MT and MST. Nature 382, 539–54110.1038/382539a0Suche in Google Scholar PubMed
Valletta, J. J., Torney, C., Kings, M., et al. (2017). Applications of machine learning in animal behaviour studies. Anim Behav 124, 203–220, doi: 10.1016/j.anbehav.2016.12.00510.1016/j.anbehav.2016.12.005Suche in Google Scholar
Vinck, M., Batista-Brito, R., Knoblich, U., Cardin, J. A. (2015). Arousal and Locomotion Make Distinct Contributions to Cortical Activity Patterns and Visual Encoding. Neuron 86, 740–754, doi: 10.1016/j.neuron.2015.03.02810.1016/j.neuron.2015.03.028Suche in Google Scholar PubMed PubMed Central
Wallace, D. J., Greenberg, D. S., Sawinski, J., et al. (2013). Rats maintain an overhead binocular field at the expense of constant fusion. Nature 498, 65–69, doi: 10.1038/nature1215310.1038/nature12153Suche in Google Scholar PubMed
Williamson, R. S., Hancock, K. E., Shinn-Cunningham, B. G., Polley, D. B. (2015). Locomotion and Task Demands Differentially Modulate Thalamic Audiovisual Processing during Active Search. Curr Biol 25, 1885–1891, doi: 10.1016/j.cub.2015.05.04510.1016/j.cub.2015.05.045Suche in Google Scholar PubMed PubMed Central
Zagha, E., McCormick, D. A. (2014). Neural control of brain state. Curr Opin Neurobiol 29, 178–186, doi: 10.1016/j.conb.2014.09.01010.1016/j.conb.2014.09.010Suche in Google Scholar PubMed PubMed Central
Zhang, S., Xu, M., Chang, W.-C., et al. (2016). Organization of long-range inputs and outputs of frontal cortex for top-down control. Nat. Neurosci. 19, 1733–1742, doi: 10.1038/nn.441710.1038/nn.4417Suche in Google Scholar PubMed PubMed Central
Zhang, S., Xu, M., Kamigaki, T., et al. (2014). Selective attention. Long-range and local circuits for top-down modulation of visual cortex processing. Science 345, 660–665, doi: 10.1126/science.125412610.1126/science.1254126Suche in Google Scholar PubMed PubMed Central
Zhou, M., Liang, F., Xiong, X. R., et al. (2014). Scaling down of balanced excitation and inhibition by active behavioral states in auditory cortex. Nat. Neurosci. 17, 841–850, doi: 10.1038/nn.370110.1038/nn.3701Suche in Google Scholar PubMed PubMed Central
Zmarz, P., Keller, G. B. (2016). Mismatch Receptive Fields in Mouse Visual Cortex. Neuron 92, 766–772, doi: 10.1016/j.neuron.2016.09.05710.1016/j.neuron.2016.09.057Suche in Google Scholar PubMed
Article note:
German version available at https://doi.org/10.1515/nf-2017-0046
© 2017 by De Gruyter
Artikel in diesem Heft
- Frontmatter
- Übersichtsartikel
- Selektive Degeneration dopaminerger Neurone beim Parkinson-Syndrom: die zunehmende Rolle von veränderter Kalziumhomöostase und nukleolärer Funktion
- Selective degeneration of dopamine neurons in Parkinson’s disease: emerging roles of altered calcium homeostasis and nucleolar function
- Altruismus aus Sicht der Sozialen Neurowissenschaften
- Altruism from the Perspective of the Social Neurosciences
- Lebensbedingungen haben einen starken Einfluss auf die Plastizität des Gehirns
- Environmental conditions strongly affect brain plasticity
- Die neuronalen Signale, die Wahrnehmung verändern
- The neural events that change perception
- Der Einfluss von Fortbewegung auf die sensorische Informationsverarbeitung und die zugrunde liegenden neuronalen Schaltkreise
- The influence of locomotion on sensory processing and its underlying neuronal circuits
- Forschungsförderung
- Sonderforschungsbereich (SFB/TRR 167) NeuroMac „Entwicklung, Funktion und Potenzial von myeloischen Zellen im zentralen Nervensystem“
- Rezension
- Michael Madeja/Joachim Müller-Jung (Hrsg.): Hirnforschung – was kann sie wirklich
- Nachrichten
- Neue NWG-Website
Artikel in diesem Heft
- Frontmatter
- Übersichtsartikel
- Selektive Degeneration dopaminerger Neurone beim Parkinson-Syndrom: die zunehmende Rolle von veränderter Kalziumhomöostase und nukleolärer Funktion
- Selective degeneration of dopamine neurons in Parkinson’s disease: emerging roles of altered calcium homeostasis and nucleolar function
- Altruismus aus Sicht der Sozialen Neurowissenschaften
- Altruism from the Perspective of the Social Neurosciences
- Lebensbedingungen haben einen starken Einfluss auf die Plastizität des Gehirns
- Environmental conditions strongly affect brain plasticity
- Die neuronalen Signale, die Wahrnehmung verändern
- The neural events that change perception
- Der Einfluss von Fortbewegung auf die sensorische Informationsverarbeitung und die zugrunde liegenden neuronalen Schaltkreise
- The influence of locomotion on sensory processing and its underlying neuronal circuits
- Forschungsförderung
- Sonderforschungsbereich (SFB/TRR 167) NeuroMac „Entwicklung, Funktion und Potenzial von myeloischen Zellen im zentralen Nervensystem“
- Rezension
- Michael Madeja/Joachim Müller-Jung (Hrsg.): Hirnforschung – was kann sie wirklich
- Nachrichten
- Neue NWG-Website