Altruismus aus Sicht der Sozialen Neurowissenschaften
-
Sabine Windmann
Sabine Windmann ist seit 2006 Professorin für Allgemeine Psychologie II an der Johann Wolfgang Goethe-Universität Frankfurt am Main. Sie studierte Psychologie in Gießen und Bonn, promovierte an der Universität Trier und habilitierte sich an der Ruhr-Universität Bochum. Zwei Jahre ihrer Post-Doc Forschungstätigkeit absolvierte sie an der University of California San Diego und zwei weitere an der University of Plymouth (UK). Ihre Forschungsinteressen beziehen sich auf die Auflösung von Unsicherheit, Ambiguität und Konflikt, zuletzt vor allem in sozialen Kontexten. Sie lebt mit ihrem Mann und ihren beiden Kindern in Weinheim an der Bergstraße.
 and
Grit Hein
and
Grit Hein
Grit Hein ist Psychologin, Neurowissenschaftlerin, und Professorin für Translationale Soziale Neurowissenschaften an der Universität Würzburg. Nach ihrem Psychologiestudium an der Humboldt-Universität zu Berlin und dem City College of New York promovierte sie am Max-Planck-Institut für Kognitions- und Neurowissenschaften in Leipzig. Nach Forschungsaufenthalten in Cambridge, England, Frankfurt am Main und Berkeley, USA, war sie als Wissenschaftlerin an der Universität Zürich und als Dozentin an der Universität Bern tätig. Schwerpunkte ihrer Forschung sind neuronale Grundlagen sozialer Motive, und ihre Auswirkung auf Verhalten, das Zusammenspiel zwischen sozialer Motivation und Lernen, und motivationale Defizite bei Patienten mit psychischen Störungen. Grit Hein ist verheiratet und Mutter von zwei Söhnen.

Zusammenfassung
Altruismus ist ein verblüffendes Phänomen, vor allem aus Sicht der Biologie und der Ökonomie. Warum geben Individuen anderen etwas von ihren Ressourcen ab und verringern damit ihre eigenen Möglichkeiten? Die Antwort auf diese Frage kann auf ultimater oder proximater Ebene gesucht werden. Die Sozialen Neurowissenschaften versuchen, die Gehirnmechanismen zu spezifizieren, die Menschen dazu antreiben, altruistisch zu handeln, denn äußerlich gleiches Verhalten kann durch unterschiedliche Motive bedingt sein. Aktivierungen und funktionelle Konnektivitäten der Anterioren Insula und der Temporoparietalen Junction spielen bestimmbare Rollen bei empathiebasiertem versus strategischem Altruismus, während der dorsolaterale präfrontale Kortex, neben anderen Regionen, bei punitiven Formen von Altruismus involviert ist. Zukünftige Forschungsarbeiten könnten sich auf die Verarbeitung von Ambiguität und Konflikt in der Verfolgung altruistischer Absichten beziehen.
Konzeptionen von Altruismus
Altruismus (lat. alter, the other) kann definiert werden als kostspieliges Verhalten, welches dem Wohlergehen anderer zu Gute kommt. Übereinstimmend mit unserem Alltagsverständnis wird es in der Wissenschaft mit prosozialem Verhalten und Kooperation in Verbindung gebracht (Nowak, 2006). Mit Altruismus kontrastiert werden können Egoismus/Individualismus (Maximierung der eigenen Ressourcen), Konkurrenz (Maximierung der Differenz zwischen eigenen Ressourcen und denjenigen anderer) und Boshaftigkeit (Minimierung der Ressourcen anderer; siehe Murphy und Ackermann, 2014).
In den letzten ein bis zwei Jahrzehnten wurden Altruismus und Kooperation verstärkt beforscht sowohl von theoretischen als auch empirischen Disziplinen innerhalb der Ökonomie, Biologie, Psychologie und den Neurowissenschaften. Einer der Gründe für das enorme Interesse ist die schwierige Vereinbarkeit von Altruismus mit klassischen theoretischen Annahmen. Wie kann Darwins Evolutionstheorie erklären, dass Altruismus aus natürlicher Selektion hervorging, wenn es kostspielig und fremdnutzend ist? Warum sollte sich aus Sicht der klassischen Ökonomie, die an die Maximierung subjektiver Nutzen glaubt, jemals jemand dafür entscheiden, Kosten auf sich zu nehmen, um damit anderen zu nutzen?
Solche theoretischen Konflikte entstehen vor allem, wenn Altruismus in Termini von Input-Output-Relationen bzw. in Termini reproduktiver Fitness definiert wird. Aus naturwissenschaftlicher Perspektive sind solche Definitionen notwendig, denn sie erlauben die rein objektive, beobachtungsbasierte Bestimmung des zu erklärenden Phänomens. Jedwedes Verhalten, das kostspielig ist für den Akteur, aber jemand anderem nützt, wird als altruistisch angesehen, gleichgültig, ob es von absichtsvollen Menschen gezeigt wird (Becker und Eagly, 2004) oder von Fischen (Daniels, 1981), Bakterien (Lee et al., 2010) oder Pflanzen (Murphy und Dudley, 2009).
Andere Forscher, vor allem Psychologen, finden ein solches „behavioristisches“ Verständnis von Altruismus unbefriedigend und verweisen daher auf innere Zustände wie die Intention, jemand anderem zu nutzen. Beispielsweise definieren Batson und Shaw (1991) Altruismus als „motivationalen Zustand mit dem ultimaten Ziel, das Wohlergehen eines anderen zu erhöhen“ (S. 108, Übersetzung der Verfasserinnen). Aus dieser Perspektive müssen Forscher nicht in erster Linie das Verhalten beobachten, um über das Vorliegen von Altruismus zu entscheiden, sondern die aktiven Zielvorstellungen des Akteurs bestimmen. Bemerkenswerterweise akzeptieren diejenigen Forscher, die solche Definitionen verwenden, oft auch proximate Variablen bei der Bewertung des Kosten-Nutzen-Verhältnisses für den Akteur. So werden Gefühle der „Herzerwärmung“, die der Akteur bei erfolgreich geleisteter Hilfe empfinden kann, für diesen als wertvoll (nützlich) eingeschätzt und daher als Evidenz gegen das Vorliegen von „echter“ Selbstlosigkeit gewertet (Batson und Shaw, 1991; Harbaugh et al., 2007).
In der vorliegenden Übersicht wollen wir nicht versuchen, metatheoretische Differenzen zwischen den Disziplinen zu diskutieren oder aufzulösen, obwohl wir betonen, dass man die Unterschiede im Hinterkopf behalten sollte beim Studium der Literatur. Stattdessen werden wir ultimate und proximate Erklärungen von Altruismus erläutern und auf den Beitrag der Sozialen Neurowissenschaften eingehen. Insgesamt finden wir, dass die Altruismus-Forschung einige der traditionellen Grenzen zwischen objektiv und subjektiv, biologisch und kulturell, proximat und ultimat in Frage stellt (Laland et al., 2011).
Ultimate Erklärungen
Schon lang ist bekannt, dass natürliche Selektion Altruismus fördern kann, wenn Geber und Rezipient des altruistischen Verhaltens miteinander verwandt sind und die Kosten für die altruistische Handlung geringer sind als der Grad der Verwandtschaft. Diese Verhältnisbildung pointierte John B. S. Haldane, als er scherzte: „Ich würde in den Fluss springen, um entweder zwei meiner Brüder zu retten oder acht meiner Cousins“ (Nowak, 2006).
Um jedoch Altruismus zwischen Nichtverwandten zu erklären, müssen evolutionäre Theorien von indirekten Nutzen für den Akteur ausgehen, bzw. für die Gruppe oder das Netzwerk, in dem der Akteur lebt (Nowak, 2006, 2012). Der Gedanke ist, kurz gesagt, dass altruistische Individuen, die durch ihre Handlungen das Wohlergehen ihrer sozialen Umgebung steigern, irgendwann „zurückprofitieren“, entweder persönlich oder indirekt durch die Stärkung ihrer Bezugsgruppe. Nowak spricht von einem „snuggle for survival“ (Nowak, 2012, p. 34).
Zwei zentrale Strategien sollen hier hervorgehoben werden aufgrund ihrer möglichen proximaten Implikationen. Die erste ist Reziprozität, ein Mechanismus, durch den gesichert wird, dass das Äquivalent des Wertes einer altruistischen Handlung zum Akteur zurückkehrt (Milinski, 2016). Reziprozität gibt es in zwei Formen: Bei „direkter Reziprozität“ erhält der altruistische Akteur selbst eine gleichwertige Belohnung zurück von dem Rezipienten; die beiden kooperieren also nach dem Prinzip „Eine Hand wäscht die andere“. Bei „indirekter Reziprozität“ hingegen sind mehr als zwei Parteien involviert. Sie gibt es in zwei Formen. Erstens, bei „abwärtsgerichteter“ indirekter Reziprozität erwirbt der altruistische Akteur durch seine Handlung einen guten Ruf, der gewährleistet, dass er in zukünftigen Situationen leichter Kooperationspartner findet (auch Sexualpartner), sodass seine reproduktive Fitness erhöht wird (Iredale und Vugt, 2009). Zweitens, bei „aufwärtsgerichteter“ indirekter Reziprozität werden die erhaltenen Belohnungen undifferenziert weitergeleitet zu anderen Gruppenmitgliedern, analog zu Kants Imperativ: „Handle nur nach derjenigen Maxime, durch die du zugleich wollen kannst, dass sie ein allgemeines Gesetz werde“.
Reziprozität als Prinzip macht Altruismus kompatibel mit Darwins Evolutionstheorie. Allerdings hat es einen schwerwiegenden Nachteil, besonders im Fall großer Gruppen, in denen der Beitrag des einzelnen zum Wohlergehen der Gruppe nicht gut verfolgt werden kann: die Gefahr des Trittbrettfahrens. Individuen, die vom Altruismus anderer Gruppenmitglieder profitieren, aber ihrerseits keinen fairen Beitrag zum Gruppenwohl leisten, weisen eine höhere Erfolgsbilanz auf als diejenigen, die kooperieren. Sie behalten von vorneherein mehr von ihren Ressourcen und profitieren zusätzlich von dem Investment der anderen Gruppenmitglieder. Unterm Strich sollte daher ihre reproduktive Fitness höher sein als die der anderen Gruppenmitglieder. Und je mehr Nachkommen sie produzieren, desto mehr bringen sie kooperierende Individuen zum Aussterben, insofern Selektionsdruck herrscht. Die Bedingungen solcher Dynamiken werden mit computationalen Modellierungsstudien untersucht (Le und Boyd, 2007; Nowak und Sigmund, 1998; Riolo et al., 2001).
Trittbrettfahrer stellen also Theorien zur Erklärung von Altruismus unter Nichtverwandten vor ein großes Problem. Es gibt jedoch Hilfe, und sie kommt aus der verhaltensökonomischen Forschung. Ernst Fehr und seine Kollegen haben in ökonomischen Spielen im Labor gezeigt, dass kooperative Gruppenmitglieder bereit sind, Kosten auf sich zu nehmen, um Trittbrettfahrer zu bestrafen (Fehr und Fischbacher, 2003; Fehr und Gachter, 2002). Durch die Bestrafung wird die Bilanz der Trittbrettfahrer reduziert, was sie zukünftig von unfairem Verhalten abhalten kann. Ohne die Möglichkeit einer solchen Bestrafung bricht die Kooperation von Gruppen zusammen, aber unter der Androhung von Strafe bleibt sie aufrechterhalten (Fehr und Fischbacher, 2003; Fehr und Gachter, 2002).
Kostspielige Bestrafung ist der zweite zentrale Mechanismus von Altruismus mit hohem Potenzial für die Sozialen Neurowissenschaften. Aus objektiver Perspektive ist die Bestrafung selbstlos, weil sie den Bestrafer kostet, während sie der Gemeinschaft nützt, indem sie Fairness-Normen sichert. Sie erfolgt in besonders effektiver und überzeugender Weise, wenn sie von Autoritäten oder unbeteiligten Institutionen vollzogen wird. Mindestens auf dieser hoch organisierten Ebene ist kostspielige Bestrafung wahrscheinlich ein Produkt genetisch-kultureller Coevolution (Bowles et al., 2003; Fehr und Fischbacher, 2003; Henrich et al., 2006; Nowak, 2006).
Proximate Erklärungen
Jedes evolvierte Verhalten muss einen proximaten Mechanismus einschließen, der den Organismus dazu bringt, das Verhalten zu zeigen. Wilson (1992, p. 62) fragt: „Sind Verhaltensweisen, die als altruistisch im evolutionären Sinn gelten, notwendigerweise verursacht durch proximate Mechanismen, die als altruistisch im psychologischen Sinn gelten?“ (Übersetzung der Verfasserinnen). Hier kommen die Möglichkeiten der Sozialen Neurowissenschaften ins Spiel. Mithilfe von Gehirnbildgebung und anderen Methoden können diese die proximaten Mechanismen, die altruistischem Verhalten unterliegen, spezifizieren und dissoziieren und in Beziehung setzen zu den dahintersteckenden Motiven (siehe Tabelle 1), und zwar ohne angewiesen zu sein auf Introspektion und Selbstbericht (Camerer, 2008; Hein et al., 2016). Objektive Korrelate für subjektive Motive zu identifizieren kann sehr wichtig sein bei Konzepten, die stark der sozialen Erwünschtheit unterliegen und daher anfällig sind für subjektive Verzerrungen. Natürlich muss man gegenüber der Idee eines „Mind-Reading“ kritisch bleiben aus methodologischen (Poldrack, 2011) sowie ethischen Gründen (Evers und Sigman, 2013). Aus einzelnen Studien allein (erst recht nicht aus Gehirndaten einzelner Probanden) lassen sich normalerweise keine subjektiven oder motivationalen Zustände ablesen, hauptsächlich wegen der hohen interindividuellen Variabilität der strukturellen und funktionellen Neuroanatomie sowie der hohen Vernetztheit und Multifunktionalität kortikaler Areale. Daher können die Sozialen Neurowissenschaften nicht einfach Gehirndaten erheben und direkt interpretieren; vielmehr müssen sie spezielle experimentelle Bedingungen realisieren und vergleichen bzw. die Motive der Probanden variieren oder bestimmen. Anschließend setzen sie diese in Beziehung zu den gemessenen neuronalen Korrelaten und Dynamiken.
Mögliche Erklärungen für die Entstehung und Aufrechterhaltung von Altruismus
| Typ Altruismus | Hilfeverhalten (belohnend, Kooperation anregend) | Kostspielige Bestrafung (konfrontativ, gegen Unfairness und Delinquenz gerichtet) |
|---|---|---|
| Ultimate Erklärung | Genetische Verwandtschaft Reziprozität, direkt oder indirekt Gruppenselektion Netzwerk-Selektion | Gruppenselektion Genetisch-kulturelle Coevolution |
| Proximate Erklärung | Empathie Perspektivenübernahme Erwartung von Gegenseitigkeit Unfairnessaversion | Unfairnessaversion Normatives Denken Wut, Rachegefühle, Dominanzstreben Moralische Überzeugungen |
Üblicherweise treffen die Probanden in solchen Experimenten Entscheidungen, die einer oder mehreren anderen Person zu Gute kommen, aber mit Kosten für sie selbst verbunden sind. In vielen Studien sind diese Kosten monetär (Hein et al., 2016; Hutcherson et al., 2015; Morishima et al., 2012); eine Übersicht experimenteller Paradigmen bieten Fehr und Camerer, 2007 und Sanfey, 2007. In einigen Studien ist eine altruistische Entscheidung aber auch an physische Kosten, z. B. an einen schmerzhaften Schock am Handrücken, gekoppelt (Hein et al., 2011; Hein et al., 2010). Während die Probanden ihre Entscheidungen treffen oder Einschätzungen abgeben, werden Korrelate ihrer Hirnaktivierung, Konnektivitäten zwischen verschiedenen Hirnarealen, hormoneller Status oder genetische Parameter erhoben, bzw. untersucht, ob diese Maße mit den Entscheidungen oder Präferenzen der Probanden in Verbindung stehen. Das Gesamtmuster der Daten kann über die neuronale Implementation der Motive informieren, die den altruistischen Entscheidungen zugrunde liegen.
Mittlerweile zeigen zahlreiche Befunde, dass altruistische Entscheidungen auf Veränderungen von neuronalen Aktivierungen bzw. neuronalen Konnektivitäten in Netzwerken von Hirnregionen basieren, die allgemein mit sozialen Prozessen in Verbindung gebracht werden (Adolphs, 2009; Rilling und Sanfey, 2011; Van Overwalle, 2009). Manchmal werden diese Hirnregionen auch als das „soziale Gehirn“ bezeichnet. Im Kontext von altruistischen Entscheidungen spielen unter anderem die Insel, das Striatum, der anteriore zinguläre Kortex (ACC), die sogenannte Temporoparietale Junction (TPJ), und der dorsolaterale präfrontale Kortex (DLPFC; Abbildung 1) eine Rolle. Diese Regionen sind jedoch auch bei nicht-sozialen Anforderungen aktiviert. Daher wird angenommen, dass sie mit übergeordneten Funktionen korrelieren, die dann auch in sozialen Kontexten benutzt werden. Die Insula wird beispielsweise mit der Verarbeitung interozeptiver (Craig, 2009) und viszeral-sensorischer Reize (z. B. Schmerz; (Critchley und Harrison, 2013)) in Verbindung gebracht und korreliert im sozialen Kontext mit der Beobachtung und Simulation solcher emotionaler und körperlicher Zustände bei anderen Personen (z. B. Empathie für den Schmerz eines anderen (Zaki et al., 2016)). Das Striatum ist reich an Dopamin, d. h. einer der wichtigsten Neurotransmitter für die Übermittlung von Belohnungssignalen. Darauf basierend spielt das Striatum eine zentrale Rolle bei der neuronalen Verarbeitung von Belohnungen in sozialen wie auch nicht-sozialen Kontexten (Schultz, 2017). Der anteriore zinguläre Kortex steht mit der Verarbeitung von Fehlern, Belohnungen und Konflikten (z. B. zwischen Stimuluskategorien) in Verbindung (Kolling et al., 2016), anhand derer flexibles, adaptives Verhalten in sozialen und nicht-sozialen Situationen initiiert wird (Shackman et al., 2011). Im Folgenden werden zwei Beispielstudien besprochen, bei denen Hirnaktivierungen und neuronale Konnektivitäten zur Untersuchung von altruistischen Verhalten und dessen Motiven eingesetzt wurden.
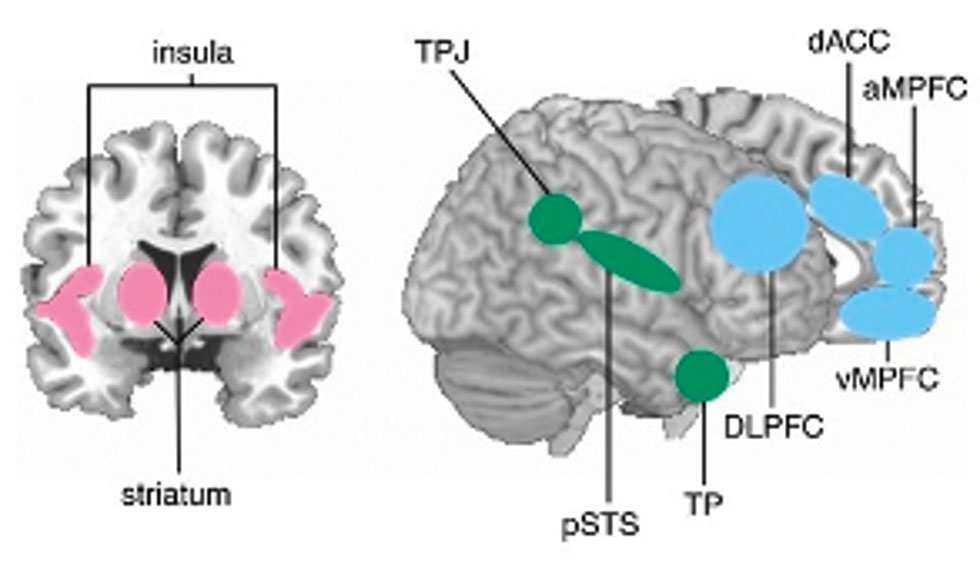
Übersicht von Hirnregionen, die mit der Verarbeitung sozialer Prozesse in Verbindung stehen. TPJ = Temporal Parietal Junction; dACC = dorsaler Teil des anterioren zingulären Kortex, DLPFC = dorsolateral präfrontaler Kortex, pSTS = posteriorer Teil des superioren temporären Sulcus, TP = temporaler Pol; vMPFC = ventraler Teil des medialen präfrontalen Kortex, aMPFC = anteriorer Teil des medialen präfrontalen Kortex.
In einer Beispielstudie (Hein et al., 2010) wurde das Motiv untersucht, welches dem Mangel an altruistischem Verhalten gegenüber Mitgliedern von Fremdgruppen zugrunde liegt. Die Probanden beobachteten, wie ein Mitglied ihrer eigenen Gruppe (ingroup member) oder ein Mitglied einer Fremdgruppe (outgroup member) Schmerz am Handrücken erhielt. Sie konnten entscheiden, die Hälfte dieses Schmerzes auf ihre eigene Hand umzuleiten und damit den Schmerz der anderen Person zu halbieren, d. h. der anderen Person zu helfen, indem sie physische Kosten (Schmerz) auf sich nahmen. Im Durchschnitt halfen die Probanden dem Mitglied der eigenen Gruppe signifikant häufiger als dem Mitglied der Fremdgruppe. Interessanterweise wurde das Ausmaß, in dem die eigene Gruppe bevorzugt wurde, von Unterschieden in der Aktivität der Anterioren Insula (AI) vorhergesagt, welche die Probanden vorher beim Beobachten des Schmerzes des Eigen- und Fremdgruppenmitglieds gezeigt hatten, und welche mit Empathie korrelierte. Je stärker ein Proband die AI beim Beobachten des Schmerzes in der Eigengruppe, aber nicht in der Fremdgruppe aktivierte, desto wahrscheinliches war es, dass er dem Eigengruppenmitglied, aber nicht dem Fremdgruppenmitglied später half (Abbildung 2). Probanden, die besonders negativ gegenüber dem Fremdgruppenmitglied eingestellt waren, aktivierten außerdem das ventrale Striatum, also einen Teil des Belohnungsverarbeitungssystems, wenn sie Schmerz in der Fremdgruppe beobachteten. Das Ausmaß dieser Aktivierung sagte fehlendes Helfen gegenüber dem Fremdgruppenmitglied vorher.
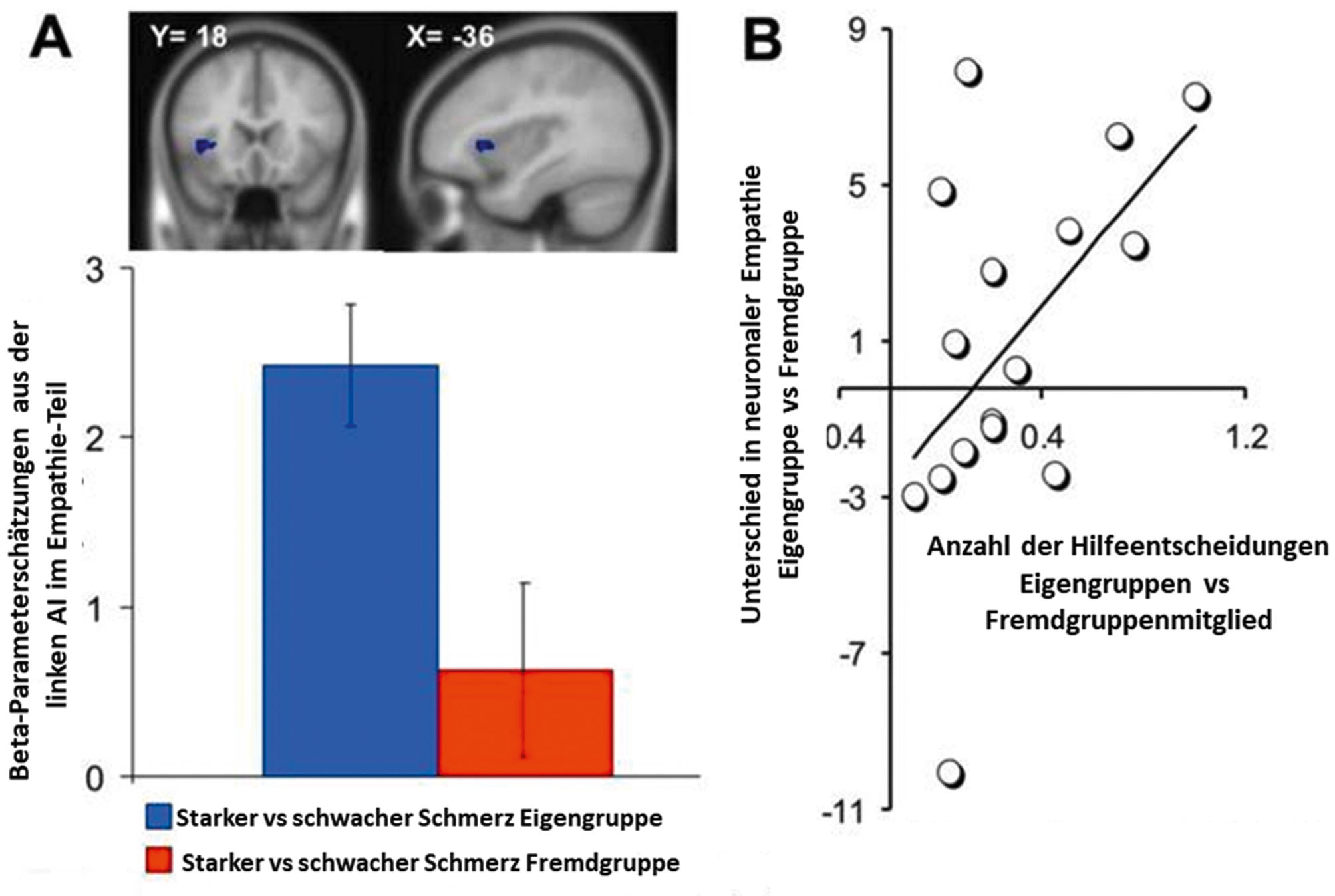
Befunde einer Beispielstudie. A) Die Ergebnisse zeigten eine stärkere durchschnittliche Aktivierung der linken anterioren Insel (AI) beim Beobachten des Schmerzes eines Mitglieds der Gruppe des Probanden (ingroup – IG) im Vergleich zum Beobachten des Schmerzes eines Mitglieds einer fremden Gruppe (outgroup – OG). B) Die Unterschiede in empathiebezogener Aktivierung in der AI zwischen der ingroup und der outgroup Bedingung sagten Unterschiede im späteren helfenden Verhalten voraus. Je stärker ein Proband die AI in der ingroup und nicht der outgroup Bedingung aktivierte, desto wahrscheinlicher war es, dass der Proband später dem Mitglied seiner Gruppe und nicht dem Mitglied der fremden Gruppe half. Originalabbildung von Hein et al., 2010, aus Neuron68, S. 149–160.
In einer zusätzlichen Analyse wurde verglichen, wie gut Hirnmaße (Aktivierung in der AI und im ventralen Striatum) und Fragebogenmaße (Skalen, die die Empathie und die Einstellung des Probanden bezüglich des Eigen- und Fremdgruppenmitglieds erfassten) späteres helfendes Verhalten gegenüber dem Eigengruppen- und Fremdgruppenmitglied vorhersagen. Es zeigte sich, dass Verhalten gegenüber dem Eigengruppenmitglied von Hirnmaßen und Fragebogenmaßen gleich gut vorhergesagt wurde, dass jedoch Hirnmaße deutlich besser zur Vorhersage des Verhaltens gegenüber der Fremdgruppe geeignet waren. Insgesamt weisen die Befunde darauf hin, dass Unterschiede in altruistischen Verhalten gegenüber Eigen- und Fremdgruppenmitgliedern durch Unterschiede in Empathie motiviert sein können, und dass Hirnmaße Verhalten in sozial sensitiven Situationen vorhersagen können, z. B. gegenüber Mitgliedern einer ungeliebten Fremdgruppe.
Eine weitere aktuelle Beispielstudie benutzte monetäre Kosten, um zwei Motive voneinander zu trennen, die zum gleichen altruistischen Verhalten führten (Hein et al., 2016). Im ersten Teil der Studie wurde entweder ein Empathie- oder ein Reziprozitätsmotiv aktiviert. Um das Empathiemotiv zu aktivieren, beobachten die Probanden, wie eine andere Person eine Schmerzstimulation am Handrücken erhielt. Das Reziprozitätsmotiv wurde aktiviert, indem eine andere Person dem Proband einen Gefallen tat. Basierend auf diesen beiden Motiven trafen die Probanden dann ökonomische Entscheidungen, in denen sie Punkte zwischen sich und einer anderen Person aufteilten, die später in Geld umgewandelt wurden. Die Aktivierung des Empathie- und des Reziprozitätsmotivs führten zu einer vergleichbaren Anzahl von altruistischen Entscheidungen (d. h. Entscheidungen, in denen der Proband die Punkte des anderen maximierte und dabei gleichzeitig seine Punkte minimierte). Das heißt, dass die beiden Motive nicht anhand des Verhaltens der Probanden unterschieden werden konnten. Auch bezüglich funktionaler Hirnaktivierungen zeigten sich keine Unterschiede. Empathie-basierte und reziprozitätsbasierte Entscheidungen aktivierten ein vergleichbares neuronales Netzwerk, welches aus der AI, dem ventralen Striatum und dem ACC bestand. In einem nächsten Schritt analysierten die Autoren die neuronalen Konnektivitäten zwischen diesen Regionen mithilfe von Dynamic Causal Modeling (DCM), ein Verfahren, durch das aufgrund von Annahmen über das Zustandekommen des gemessenen Signals effektive Verbindungen zwischen aktivierten Regionen geschätzt werden sollen. Es zeigte sich, dass empathiebasierte und reziprozitätsbasierte altruistische Entscheidungen zwar das gleiche Netzwerk aktivierten, aber verschiedene neuronale Konnektivitätsmuster innerhalb dieses Netzwerks hervorriefen (Abbildung 3). Anhand dieser Unterschiede in den neuronalen Konnektivitätsmustern konnte statistisch vorhergesagt werden, ob die altruistische Entscheidung einer Person von Empathie oder Reziprozität getrieben war.
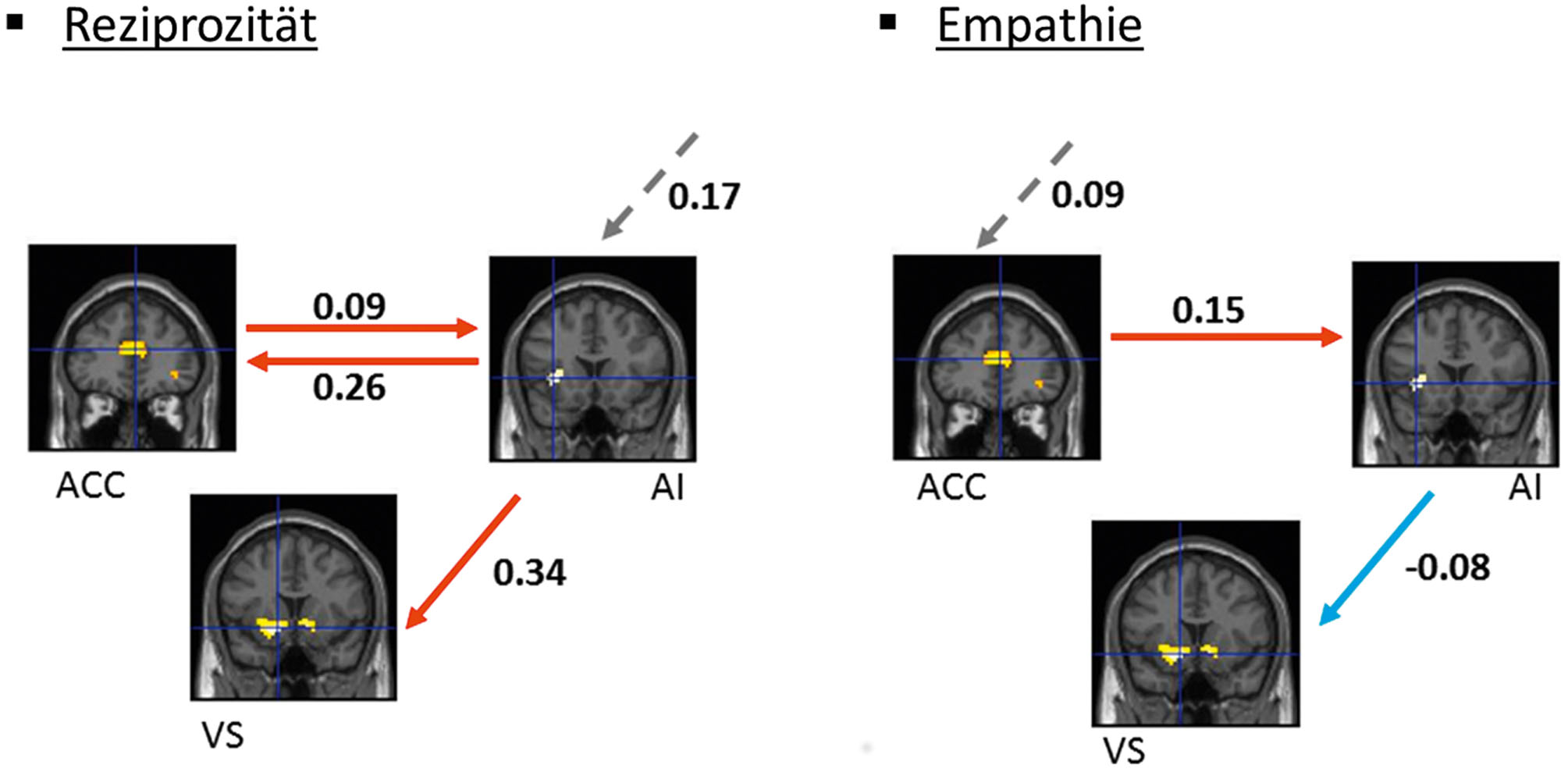
Befunde einer Beispielstudie, welche unterschiedliche Motive anhand von neuronalen Konnektivitäten klassifizierte. Durchgestrichene Pfeile zeigen an, wie stark das Ausmaß der Aktivierung einer Region die Aktivierung einer anderen Region verändert (Effektive Konnektivität). Gestrichelte Pfeile zeigen Veränderung der Aktivierung innerhalb einer Region in Abhängigkeit von der experimentellen Manipulation (Input) an. Die Zahlen sind Modell-Parameter, die die Stärke der effektiven Konnektivitäten oder Inputs reflektieren. ACC = anteriorer zingulärer Kortex, AI = anteriore Insel, VS = ventrales Striatum.
Als ein weiterer interessanter Befund wurde gezeigt, dass vorwiegend egoistische und vorwiegend prosoziale Personen unterschiedlich auf die Aktivierung des Empathie- und des Reziprozitätsmotivs reagierten. Überwiegend egoistische Personen zeigten mehr altruistische Entscheidungen nach der Aktivierung des Empathiemotivs. Im Gegensatz erhöhten Personen, die schon „von Hause“ aus prosozial eingestellt waren, ihre altruistischen Entscheidungen nur nach Aktivierung des Reiziprozitätsmotivs, und nicht nach der Aktivierung von Empathie.
Die beiden Studien von Hein et al. (2010) und Hein et al. (2016) zeigen exemplarisch die Methoden und Paradigmen des noch jungen Feldes der Sozialen Neurowissenschaften auf, die in weiteren Studien konsolidiert, optimiert und ständig weiterentwickelt werden müssen.
Belohnende versus punitive Formen von Altruismus
Die bislang beschriebene Forschung hat diverse Motive spezifiziert, die hinter kostspieligem Helfen stecken (Helfen im Sinne vom Teilen materieller oder physischer Ressourcen), also einem belohnenden Verhalten, das Kooperation fördert und so das Wohlergehen aller maximiert. Wie verhalten sich diese Befunde jedoch zum altruistischen Bestrafen, dem zweiten Schlüsselmechanismus von Kooperation? Interessanterweise haben Verhaltensexperimente mit ökonomischen Spielen gezeigt, dass diese beiden Typen von Verhaltensweisen, kostspieliges Belohnen einerseits und kostspieliges Bestrafen andererseits, im Grunde unkorreliert sind (Peysakhovich et al., 2014; Yamagishi et al., 2012). Erneut können die Sozialen Neurowissenschaften dabei helfen, die Unterschiede in Termini der proximaten Mechanismen zu spezifizieren, speziell der neuronalen Basis der zugrunde liegenden Motive.
Im Hinblick auf zugrunde liegende emotionale Prozesse hat eine Reihe von Studien gezeigt, dass kostspieliges Helfen/Belohnen von „Herzerwärmung“ und ähnlichen positiven Gefühlen begleitet ist (Harbaugh et al., 2007; Hu et al., 2016a; Rand et al., 2015). Demgegenüber ist kostspieliges Bestrafen von Wut und Racheglüsten geprägt (Crockett et al., 2014; Fehr und Gachter, 2002; Seip et al., 2009; Singer et al., 2006). Insofern wirkt letzteres eher kompetitiv und konfrontativ, ganz entgegen den fürsorglichen prosozialen Absichten, die bei belohnendem Hilfeverhalten eine Rolle spielen.
Bildgebende Verfahren bestätigen, dass die beiden Verhaltensweisen mit unterschiedlichen neuronalen Mechanismen verbunden sind. Obwohl beide ventrale und dorsale Teile des Striatums aktivieren als anreizvermittelnde Struktur (für Hilfeverhalten, (Genevsky et al., 2013; Harbaugh et al., 2007; Hein et al., 2010; Kuss et al., 2013), für Bestrafung (Buckholtz et al., 2008; de Quervain et al., 2004; Hu et al., 2015; Strobel et al., 2011), involviert speziell Bestrafung recht konsistent den (typischerweise rechten) DLPFC. Dies wurde nicht nur mit funktionellem MRI gezeigt (Buckholtz et al., 2008; Buckholtz et al., 2015; Sanfey et al., 2003), sondern auch mit repetitiver transkranialer Gehirnstimulation (Buckholtz et al., 2015; Knoch et al., 2008; Strang et al., 2015) und Enzephalografie im Ruhezustand (Knoch et al., 2010). Der DLPFC ist bekannt für seine Rolle in der Modulation automatischer Entscheidungsfindung und Handlungsauswahl; seine Rolle im Zusammenhang mit kostspieligem selbstlosen Verhalten könnte also in der Vermittlung von Information über soziale Normen, moralische Werte und ähnlichen abstrakten Gedanken liegen, wodurch dominante reflexive Impulse daran gehindert werden, das Verhalten zu bestimmen (Feng et al., 2015).Demgegenüber sollen fürsorgliche Tendenzen, die dem Helfen und Teilen zugrunde liegen, entweder durch affektive Empathie gegenüber der/n bedürftigen Person/en angetrieben werden, dabei die AI und den medialen PFC involvierend, oder durch kognitive Empathie und Perspektivenübernahme, basierend auf der TPJ, neben anderen Arealen (Haas et al., 2015; Hein et al., 2010; Morelli et al., 2014; Strombach et al., 2015; Tusche et al., 2016).
Schlussfolgernd scheinen Hilfeverhalten und kostspielige Bestrafung, zwei Verhaltensweisen, die ultimat beide der evolutionären Definition von Altruismus genügen, proximat auf unterschiedlichen neuronalen Netzwerken zu basieren. Zwar sind konkrete Interpretationen über die funktionale Rolle der beteiligten Strukturen zunächst vorläufig und spekulativ, teilweise auch simplifizierend, sie orientieren sich jedoch an der Gesamtschau der Befundlage, indem sie die Ergebnisse im Lichte von etablierten Befunden und Theorien aus den Kognitiven Neurowissenschaften betrachten. Über die Zeit soll auf diese Weise ein Puzzle zusammengesetzt werden, das die verschiedenen Facetten von Altruismus sowie die zugrunde liegenden Prinzipien und Prozesse auf neuronaler Ebenene aufzeigt und miteinander in Beziehung setzt.
Ausblick
Zusammenfassend können die Sozialen Neurowissenschaften dabei helfen, durch die Untersuchung neuronaler Korrelate die unterschiedlichen proximaten Motive zu spezifizieren, die Verhaltensweisen zugrunde liegen, welche die evolutionäre Definition von Altruismus erfüllen. Einige dieser Mechanismen basieren auf empathischer Fürsorge, andere rekrutieren strategisches Denken, um über den Weg der Kooperation eigene Vorteile zu erlangen. Wiederum andere Verhaltensweisen scheinen in der Motivation zu wurzeln, anderen Menschen Normen aufzuerlegen, um Fairness und andere abstrakte Ziele zu erreichen anstelle direkter Belohnungen. So können die Sozialen Neurowissenschaften Interpretationshinweise liefern, um die proximaten Motive für Altruismus mit grundlegenderen kognitiven, emotionalen und motivationalen Prozessen in Beziehung zu setzen. Um Kooperation im großen gesellschaftlichen Maßstab zu erreichen unter Menschen, die einander meist nicht näher kennen, ist anscheinend eine Bandbreite von proximaten Motiven erforderlich, von denen nur einige altruistisch sind im psychologischen Sinne.
Zukünftige Arbeiten könnten die proximaten Ursachen von belohnendem und bestrafendem Verhalten direkt kontrastieren (experimentelle Ansätze hierfür bieten (Hu et al., 2016b; Hu et al., 2015)). Sind es tatsächlich prosoziale moralische Werte und Kognitionen, die beim altruistischen Bestrafen aktiviert werden, unter Mitwirkung des DLPFC, oder spielen möglicherweise soziale Dominanzbestrebungen eine Rolle (z. B. das Bedürfnis nach Homogenisierung der Gruppe)? Wie sind zivilcouragierte Ambitionen einzuordnen (z. B. Whistleblowing), altruistische Akte, die über die Orientierung an sozialen Normen hinausgehen und hohe Risiken bergen, im Unterschied zum altruistischen Bestrafen? Welche Rolle spielen Empathie und Emotionskontrolle, wenn fürsorgliche Intentionen nur durch soziale Konfrontation oder durch die Induktion aversiver Zustände im Empfänger der Hilfe verfolgt werden können (e.g. Lopez-Perez et al., 2017)? Einige dieser Fragen zielen in die Richtung von Motivkonflikten, die das soziale Gehirn auflösen muss. Sie zeigen die Vielschichtigkeit des Phänomens Altruismus und betonen die Notwendigkeit interdisziplinärer Ansätze in der Forschung unter Mitwirkung der Sozialen Neurowissenschaften.
Funding source: Deutsche Forschungsgemeinschaft
Award Identifier / Grant number: DFG, HE 4566/5-1 an GH
Funding statement: Funding: Diese Arbeit wurde von der Deutschen Forschungsgemeinschaft unterstützt (DFG, HE 4566/5-1 an GH).
About the authors

Sabine Windmann ist seit 2006 Professorin für Allgemeine Psychologie II an der Johann Wolfgang Goethe-Universität Frankfurt am Main. Sie studierte Psychologie in Gießen und Bonn, promovierte an der Universität Trier und habilitierte sich an der Ruhr-Universität Bochum. Zwei Jahre ihrer Post-Doc Forschungstätigkeit absolvierte sie an der University of California San Diego und zwei weitere an der University of Plymouth (UK). Ihre Forschungsinteressen beziehen sich auf die Auflösung von Unsicherheit, Ambiguität und Konflikt, zuletzt vor allem in sozialen Kontexten. Sie lebt mit ihrem Mann und ihren beiden Kindern in Weinheim an der Bergstraße.

Grit Hein ist Psychologin, Neurowissenschaftlerin, und Professorin für Translationale Soziale Neurowissenschaften an der Universität Würzburg. Nach ihrem Psychologiestudium an der Humboldt-Universität zu Berlin und dem City College of New York promovierte sie am Max-Planck-Institut für Kognitions- und Neurowissenschaften in Leipzig. Nach Forschungsaufenthalten in Cambridge, England, Frankfurt am Main und Berkeley, USA, war sie als Wissenschaftlerin an der Universität Zürich und als Dozentin an der Universität Bern tätig. Schwerpunkte ihrer Forschung sind neuronale Grundlagen sozialer Motive, und ihre Auswirkung auf Verhalten, das Zusammenspiel zwischen sozialer Motivation und Lernen, und motivationale Defizite bei Patienten mit psychischen Störungen. Grit Hein ist verheiratet und Mutter von zwei Söhnen.
Literatur
Adolphs, R. (2009). The social brain: neural basis of social knowledge. Annu. Rev. Psychol. 60, 693–716.10.1146/annurev.psych.60.110707.163514Search in Google Scholar PubMed
Batson, C. D. and Shaw, L. L. (1991). Evidence for altruism: toward a pluralism of prosocial motives. Psychol. Inq. 2(2), 107–122.10.1207/s15327965pli0202_1Search in Google Scholar
Becker, S. W. and Eagly, A. H. (2004). The heroism of women and men. Am. Psychol. 59, 163–78.10.1037/0003-066X.59.3.163Search in Google Scholar PubMed
Bowles, S., Choi, J. K. and Hopfensitz, A. (2003). The co-evolution of individual behaviors and social institutions. J. Theor. Biol. 223, 135–47.10.1016/S0022-5193(03)00060-2Search in Google Scholar PubMed
Buckholtz, J. W., Asplund, C. L., Dux, P. E., Zald, D. H., Gore, J. C., Jones, O. D. and Marois, R. (2008). The neural correlates of third-party punishment. Neuron 60, 930–40.10.1016/j.neuron.2008.10.016Search in Google Scholar PubMed
Buckholtz, J. W., Martin, J. W., Treadway, M. T., Jan, K., Zald, D. H., Jones, O. and Marois, R. (2015). From Blame to Punishment: Disrupting Prefrontal Cortex Activity Reveals Norm Enforcement Mechanisms. Neuron 87, 1369–80.10.1016/j.neuron.2015.08.023Search in Google Scholar PubMed PubMed Central
Camerer, C. F. (2008). Neuroeconomics: opening the gray box. Neuron 60, 416–9.10.1016/j.neuron.2008.10.027Search in Google Scholar PubMed
Craig, A. D. (2009). How do you feel – now? The anterior insula and human awareness. Nat. Rev. Neurosci. 10, 59–70.10.1038/nrn2555Search in Google Scholar PubMed
Critchley, H. D. and Harrison, N. A. (2013). Visceral influences on brain and behavior. Neuron 77, 624–38.10.1016/j.neuron.2013.02.008Search in Google Scholar PubMed
Crockett, M. J., Ozdemir, Y. and Fehr, E. (2014). The value of vengeance and the demand for deterrence. J. Exp. Psychol. Gen. 143, 2279–86.10.1037/xge0000018Search in Google Scholar PubMed PubMed Central
Daniels, R. A. (1981). Altruism in an antarctic fish. Science 213, 1281.10.1126/science.213.4513.1281Search in Google Scholar PubMed
de Quervain, D. J., Fischbacher, U., Treyer, V., Schellhammer, M., Schnyder, U., Buck, A. and Fehr, E. (2004). The neural basis of altruistic punishment. Science 305, 1254–8.10.1126/science.1100735Search in Google Scholar PubMed
Evers, K. and Sigman, M. (2013). Possibilities and limits of mind-reading: a neurophilosophical perspective. Conscious. Cogn. 22, 887–97.10.1016/j.concog.2013.05.011Search in Google Scholar PubMed
Fehr, E. and Camerer, C. F. (2007). Social neuroeconomics: the neural circuitry of social preferences. Trends Cognit. Sci. 11, 419–27.10.1016/j.tics.2007.09.002Search in Google Scholar PubMed
Fehr, E. and Fischbacher, U. (2003). The nature of human altruism. Nature 425, 785–91.10.1038/nature02043Search in Google Scholar PubMed
Fehr, E. and Gachter, S. (2002). Altruistic punishment in humans. Nature 415, 137–40.10.1038/415137aSearch in Google Scholar PubMed
Feng, C., Luo, Y. J. and Krueger, F. (2015). Neural signatures of fairness-related normative decision making in the ultimatum game: a coordinate-based meta-analysis. Hum. Brain Mapp. 36, 591–602.10.1002/hbm.22649Search in Google Scholar PubMed PubMed Central
Genevsky, A., Vastfjall, D., Slovic, P. and Knutson, B. (2013). Neural underpinnings of the identifiable victim effect: affect shifts preferences for giving. J. Neurosci. 33, 17188–96.10.1523/JNEUROSCI.2348-13.2013Search in Google Scholar PubMed PubMed Central
Haas, B. W., Brook, M., Remillard, L., Ishak, A., Anderson, I. W. and Filkowski, M. M. (2015). I know how you feel: the warm-altruistic personality profile and the empathic brain. PLoS One 10, e0120639.10.1371/journal.pone.0120639Search in Google Scholar PubMed PubMed Central
Harbaugh, W. T., Mayr, U. and Burghart, D. R. (2007). Neural responses to taxation and voluntary giving reveal motives for charitable donations. Science 316, 1622–5.10.1126/science.1140738Search in Google Scholar PubMed
Hein, G., Lamm, C., Brodbeck, C. and Singer, T. (2011). Skin conductance response to the pain of others predicts later costly helping. PLoS One 6, e22759.10.1371/journal.pone.0022759Search in Google Scholar PubMed PubMed Central
Hein, G., Morishima, Y., Leiberg, S., Sul, S. and Fehr, E. (2016). The brain’s functional network architecture reveals human motives. Science 351, 1074–8.10.1126/science.aac7992Search in Google Scholar PubMed
Hein, G., Silani, G., Preuschoff, K., Batson, C. D. and Singer, T. (2010). Neural responses to ingroup and outgroup members’ suffering predict individual differences in costly helping. Neuron 68, 149–60.10.1016/j.neuron.2010.09.003Search in Google Scholar PubMed
Henrich, J., McElreath, R., Barr, A., Ensminger, J., Barrett, C., Bolyanatz, A., Cardenas, J. C., Gurven, M., Gwako, E., Henrich, N., Lesorogol, C., Marlowe, F., Tracer, D. and Ziker, J. (2006). Costly punishment across human societies. Science 312, 1767–70.10.1126/science.1127333Search in Google Scholar PubMed
Hu, T. Y., Li, J., Jia, H. and Xie, X. (2016a). Helping Others, Warming Yourself: Altruistic Behaviors Increase Warmth Feelings of the Ambient Environment. Front. Psychol. 7, 1349.10.3389/fpsyg.2016.01349Search in Google Scholar PubMed PubMed Central
Hu, Y., Scheele, D., Becker, B., Voos, G., David, B., Hurlemann, R. and Weber, B. (2016b). The Effect of Oxytocin on Third-Party Altruistic Decisions in Unfair Situations: An fMRI Study. Sci. Rep. 6, 20236.10.1038/srep20236Search in Google Scholar PubMed PubMed Central
Hu, Y., Strang, S. and Weber, B. (2015). Helping or punishing strangers: neural correlates of altruistic decisions as third-party and of its relation to empathic concern. Front. Behav. Neurosci. 9, 24.10.3389/fnbeh.2015.00024Search in Google Scholar PubMed PubMed Central
Hutcherson, C. A., Bushong, B. and Rangel, A. (2015). A Neurocomputational Model of Altruistic Choice and Its Implications. Neuron 87, 451–62.10.1016/j.neuron.2015.06.031Search in Google Scholar PubMed PubMed Central
Iredale, W. and Vugt, M. V. (2009). The peacock’s tail of altruism. The Psychologist 22, 4.Search in Google Scholar
Knoch, D., Gianotti, L. R., Baumgartner, T. and Fehr, E. (2010). A neural marker of costly punishment behavior. Psychol. Sci. 21, 337–42.10.1177/0956797609360750Search in Google Scholar PubMed
Knoch, D., Nitsche, M. A., Fischbacher, U., Eisenegger, C., Pascual-Leone, A. and Fehr, E. (2008). Studying the neurobiology of social interaction with transcranial direct current stimulation – the example of punishing unfairness. Cereb. Cortex 18, 1987–90.10.1093/cercor/bhm237Search in Google Scholar PubMed
Kolling, N., Wittmann, M. K., Behrens, T. E., Boorman, E. D., Mars, R. B. and Rushworth, M. F. (2016). Value, search, persistence and model updating in anterior cingulate cortex. Nat. Neurosci. 19, 1280–5.10.1038/nn.4382Search in Google Scholar PubMed PubMed Central
Kuss, K., Falk, A., Trautner, P., Elger, C. E., Weber, B. and Fliessbach, K. (2013). A reward prediction error for charitable donations reveals outcome orientation of donators. Soc. Cogn. Affect. Neurosci. 8, 216–23.10.1093/scan/nsr088Search in Google Scholar PubMed PubMed Central
Laland, K. N., Sterelny, K., Odling-Smee, J., Hoppitt, W. and Uller, T. (2011). Cause and effect in biology revisited: is Mayr’s proximate-ultimate dichotomy still useful? Science 334, 1512–6.10.1126/science.1210879Search in Google Scholar PubMed
Le, S. and Boyd, R. (2007). Evolutionary dynamics of the continuous iterated prisoner’s dilemma. J. Theor. Biol. 245, 258–67.10.1016/j.jtbi.2006.09.016Search in Google Scholar PubMed
Lee, H. H., Molla, M. N., Cantor, C. R. and Collins, J. J. (2010). Bacterial charity work leads to population-wide resistance. Nature 467, 82–5.10.1038/nature09354Search in Google Scholar PubMed PubMed Central
Lopez-Perez, B., Howells, L. and Gummerum, M. (2017). Cruel to Be Kind: Factors Underlying Altruistic Efforts to Worsen Another Person’s Mood. Psychol. Sci. 28, 862–871.10.1177/0956797617696312Search in Google Scholar PubMed
Milinski, M. (2016). Reputation, a universal currency for human social interactions. Philos. Trans. R. Soc., B. 371, 20150100.10.1098/rstb.2015.0100Search in Google Scholar PubMed PubMed Central
Morelli, S. A., Rameson, L. T. and Lieberman, M. D. (2014). The neural components of empathy: predicting daily prosocial behavior. Soc. Cogn. Affect. Neurosci. 9, 39–47.10.1093/scan/nss088Search in Google Scholar PubMed PubMed Central
Morishima, Y., Schunk, D., Bruhin, A., Ruff, C. C. and Fehr, E. (2012). Linking brain structure and activation in temporoparietal junction to explain the neurobiology of human altruism. Neuron 75, 73–9.10.1016/j.neuron.2012.05.021Search in Google Scholar PubMed
Murphy, G. P. and Dudley, S. A. (2009). Kin recognition: Competition and cooperation in Impatiens (Balsaminaceae). Am. J. Bot. 96, 1990–6.10.3732/ajb.0900006Search in Google Scholar PubMed
Murphy, R. O. and Ackermann, K. A. (2014). Social value orientation: theoretical and measurement issues in the study of social preferences. Pers. Soc. Psychol. Rev. 18, 13–41.10.1177/1088868313501745Search in Google Scholar PubMed
Nowak, M. A. (2006). Five rules for the evolution of cooperation. Science 314, 1560–3.10.1126/science.1133755Search in Google Scholar PubMed PubMed Central
Nowak, M. A. (2012). Why we help: Far from being a nagging exception to the rule of evolution, cooperation has been one of its primary architects. Sci. Am. 307, 34–9.10.1038/scientificamerican0712-34Search in Google Scholar PubMed
Nowak, M. A. and Sigmund, K. (1998). Evolution of indirect reciprocity by image scoring. Nature 393, 573–7.10.1038/31225Search in Google Scholar PubMed
Peysakhovich, A., Nowak, M. A. and Rand, D. G. (2014). Humans display a ‘cooperative phenotype’ that is domain general and temporally stable. Nat. Commun. 5, 4939.10.1038/ncomms5939Search in Google Scholar PubMed
Poldrack, R. A. (2011). Inferring mental states from neuroimaging data: from reverse inference to large-scale decoding. Neuron 72, 692–7.10.1016/j.neuron.2011.11.001Search in Google Scholar PubMed PubMed Central
Rand, D. G., Kraft-Todd, G. and Gruber, J. (2015). The collective benefits of feeling good and letting go: positive emotion and (dis)inhibition interact to predict cooperative behavior. PLoS One 10, e0117426.10.1371/journal.pone.0117426Search in Google Scholar PubMed PubMed Central
Rilling, J. K. and Sanfey, A. G. (2011). The neuroscience of social decision-making. Annu. Rev. Psychol. 62, 23–48.10.1146/annurev.psych.121208.131647Search in Google Scholar PubMed
Riolo, R. L., Cohen, M. D. and Axelrod, R. (2001). Evolution of cooperation without reciprocity. Nature 414, 441–3.10.1038/35106555Search in Google Scholar PubMed
Sanfey, A. G. (2007). Social decision-making: insights from game theory and neuroscience. Science 318, 598–602.10.1126/science.1142996Search in Google Scholar PubMed
Sanfey, A. G., Rilling, J. K., Aronson, J. A., Nystrom, L. E. and Cohen, J. D. (2003). The neural basis of economic decision-making in the Ultimatum Game. Science 300, 1755–8.10.1126/science.1082976Search in Google Scholar PubMed
Schultz, W. (2017). Reward prediction error. Curr. Biol. 27, R369-R371.10.1016/j.cub.2017.02.064Search in Google Scholar PubMed
Seip, E. C., van Dijk, W. W. and Rotteveel, M. (2009). On hotheads and Dirty Harries: the primacy of anger in altruistic punishment. Ann. N. Y. Acad. Sci. 1167, 190–6.10.1111/j.1749-6632.2009.04503.xSearch in Google Scholar PubMed
Shackman, A. J., Salomons, T. V., Slagter, H. A., Fox, A. S., Winter, J. J. and Davidson, R. J. (2011). The integration of negative affect, pain and cognitive control in the cingulate cortex. Nat. Rev. Neurosci. 12, 154–67.10.1038/nrn2994Search in Google Scholar PubMed PubMed Central
Singer, T., Seymour, B., O’Doherty, J. P., Stephan, K. E., Dolan, R. J. and Frith, C. D. (2006). Empathic neural responses are modulated by the perceived fairness of others. Nature 439, 466–9.10.1038/nature04271Search in Google Scholar PubMed PubMed Central
Strang, S., Gross, J., Schuhmann, T., Riedl, A., Weber, B. and Sack, A. T. (2015). Be nice if you have to – the neurobiological roots of strategic fairness. Soc. Cogn. Affect. Neurosci. 10, 790–6.10.1093/scan/nsu114Search in Google Scholar PubMed PubMed Central
Strobel, A., Zimmermann, J., Schmitz, A., Reuter, M., Lis, S., Windmann, S. and Kirsch, P. (2011). Beyond revenge: neural and genetic bases of altruistic punishment. Neuroimage 54, 671–80.10.1016/j.neuroimage.2010.07.051Search in Google Scholar PubMed
Strombach, T., Weber, B., Hangebrauk, Z., Kenning, P., Karipidis, II, Tobler, P. N. and Kalenscher, T. (2015). Social discounting involves modulation of neural value signals by temporoparietal junction. Proc. Natl. Acad. Sci. U.S.A. 112, 1619–24.10.1073/pnas.1414715112Search in Google Scholar PubMed PubMed Central
Tusche, A., Bockler, A., Kanske, P., Trautwein, F. M. and Singer, T. (2016). Decoding the Charitable Brain: Empathy, Perspective Taking, and Attention Shifts Differentially Predict Altruistic Giving. J. Neurosci. 36, 4719–32.10.1523/JNEUROSCI.3392-15.2016Search in Google Scholar PubMed PubMed Central
Van Overwalle, F. (2009). Social cognition and the brain: a meta-analysis. Hum. Brain Mapp. 30, 829–58.10.1002/hbm.20547Search in Google Scholar PubMed PubMed Central
Wilson, D. S. (1992). On the relationship between evolutionary and psychological definitions of altruism and selfishness. Biol. Philos. 7, 61–68.10.1007/BF00130164Search in Google Scholar
Yamagishi, T., Horita, Y., Mifune, N., Hashimoto, H., Li, Y., Shinada, M., Miura, A., Inukai, K., Takagishi, H. and Simunovic, D. (2012). Rejection of unfair offers in the ultimatum game is no evidence of strong reciprocity. Proc. Natl. Acad. Sci. U.S.A. 109, 20364–8.10.1073/pnas.1212126109Search in Google Scholar PubMed PubMed Central
Zaki, J., Wager, T. D., Singer, T., Keysers, C. and Gazzola, V. (2016). The Anatomy of Suffering: Understanding the Relationship between Nociceptive and Empathic Pain. Trends Cognit. Sci. 20, 249–59.10.1016/j.tics.2016.02.003Search in Google Scholar PubMed PubMed Central
Anmerkung:
Übersetzung der englischen Version des Artikels online verfügbar unter https://doi.org/10.1515/nf-2017-A047
© 2018 by De Gruyter
Articles in the same Issue
- Frontmatter
- Übersichtsartikel
- Selektive Degeneration dopaminerger Neurone beim Parkinson-Syndrom: die zunehmende Rolle von veränderter Kalziumhomöostase und nukleolärer Funktion
- Selective degeneration of dopamine neurons in Parkinson’s disease: emerging roles of altered calcium homeostasis and nucleolar function
- Altruismus aus Sicht der Sozialen Neurowissenschaften
- Altruism from the Perspective of the Social Neurosciences
- Lebensbedingungen haben einen starken Einfluss auf die Plastizität des Gehirns
- Environmental conditions strongly affect brain plasticity
- Die neuronalen Signale, die Wahrnehmung verändern
- The neural events that change perception
- Der Einfluss von Fortbewegung auf die sensorische Informationsverarbeitung und die zugrunde liegenden neuronalen Schaltkreise
- The influence of locomotion on sensory processing and its underlying neuronal circuits
- Forschungsförderung
- Sonderforschungsbereich (SFB/TRR 167) NeuroMac „Entwicklung, Funktion und Potenzial von myeloischen Zellen im zentralen Nervensystem“
- Rezension
- Michael Madeja/Joachim Müller-Jung (Hrsg.): Hirnforschung – was kann sie wirklich
- Nachrichten
- Neue NWG-Website
Articles in the same Issue
- Frontmatter
- Übersichtsartikel
- Selektive Degeneration dopaminerger Neurone beim Parkinson-Syndrom: die zunehmende Rolle von veränderter Kalziumhomöostase und nukleolärer Funktion
- Selective degeneration of dopamine neurons in Parkinson’s disease: emerging roles of altered calcium homeostasis and nucleolar function
- Altruismus aus Sicht der Sozialen Neurowissenschaften
- Altruism from the Perspective of the Social Neurosciences
- Lebensbedingungen haben einen starken Einfluss auf die Plastizität des Gehirns
- Environmental conditions strongly affect brain plasticity
- Die neuronalen Signale, die Wahrnehmung verändern
- The neural events that change perception
- Der Einfluss von Fortbewegung auf die sensorische Informationsverarbeitung und die zugrunde liegenden neuronalen Schaltkreise
- The influence of locomotion on sensory processing and its underlying neuronal circuits
- Forschungsförderung
- Sonderforschungsbereich (SFB/TRR 167) NeuroMac „Entwicklung, Funktion und Potenzial von myeloischen Zellen im zentralen Nervensystem“
- Rezension
- Michael Madeja/Joachim Müller-Jung (Hrsg.): Hirnforschung – was kann sie wirklich
- Nachrichten
- Neue NWG-Website