Lebensbedingungen haben einen starken Einfluss auf die Plastizität des Gehirns
-
Siegrid Löwel
Siegrid Löwel studierte Biologie in Würzburg und Frankfurt/M., 1988 Promotion (Dr. phil. nat.) und 1995 Habilitation in Zoologie an der Johann-Wolfgang-Goethe-Universität in Frankfurt am Main, bis 1996 wissenschaftliche Mitarbeiterin am Max-Planck- Institut für Hirnforschung in Frankfurt am Main in der Neurophysiologischen Abteilung von Prof. Dr. Wolf Singer, 1997–2005 Leiterin der Forschergruppe „Visuelle Entwicklung und Plastizität“ am Leibniz- Institut für Neurobiologie in Magdeburg, 2002–2003 Research Associate Professor am Keck-Center der University of California in San Francisco, U.S.A., im Labor von Prof. Dr. Michael P. Stryker, 2003-2004 Dorothea-Erxleben-Gastprofessorin an der Otto-von-Guericke-Universität Magdeburg, 2004–2005 Stipendiatin im Hertie-Exzellenz-Programm „Neurowissenschaften“, 2005–2010 Universitätsprofessorin (W2) für Neurobiologie an der Friedrich-Schiller-Universität Jena, seit August 2010 Universitätsprofessorin (W3) und Leiterin der Abteilung „Systemische Neurobiologie“ an der Fakultät für Biologie und Psychologie, Georg-August-Universität Göttingen. Das Hauptinteresse von Prof. Löwel liegt darin, zu erforschen wie sich die Flexibilität unseres Gehirns im Alter und nach Läsionen erhöhen lässt und welche Mechanismen dieser Plastizität zugrunde liegen.
 ,
Evgenia Kalogeraki
,
Evgenia Kalogeraki
Evgenia Kalogeraki studierte Biologische Technologie an der Universität Ioannina, Griechenland. 2011: Master in Molekularbiologie und Biomedizin, Universität von Kreta. 2012: Beginn ihrer Promotion in der Arbeitsgruppe von Prof. Dr. Siegrid Löwel und Aufnahme in die Göttinger Graduiertenschule für Neurowissenschaften, Biophysik und Molekulare Biowissenschaften (GGNB). 2015: Promotion. Im Augenblick forscht sie als Postdoktorandin in der Abteilung „Systemische Neurobiologie“ der Universität Göttingen.
Susanne Dehmel studierte Biologie in Braunschweig und Leipzig und promovierte 2006 in der Arbeitsgruppe Neurobiologie von Prof. R. Rübsamen am Zoologischen Institut der Universität Leipzig. Von 2007 bis 2011 war sie Postdoktorandin in der Arbeitsgruppe Sensory Neurobiology von Prof. S. Shore am Kresge Hearing Research Institute, University of Michigan. Seit 2011 ist sie Postdoktorandin in der Abteilung Systemische Neurobiologie von Prof. Dr. Siegrid Löwel an der Universität Göttingen.
Kalina Makowiecki studierte Psychologie (2010, BA Hons) und promovierte zum PhD in Experimental and Regenerative Neurosciences (School of Biological Sciences) an der University of Western Australia (UWA) in Perth (2016), gefördert durch einen Preis und ein Stipendium (Australian Postgraduate Award und UWA Completion Scholarship). Im Jahr 2016 begann sie als Postdoktorandin in der Abteilung „Systemische Neurobiologie“ (Prof. Dr. Siegrid Löwel) an der Universität Göttingen. Sie wurde kürzlich mit einer Dorothea-Schlözer-Stelle für Postdoktorandinnen ausgezeichnet, die im Oktober 2017 begann.

Zusammenfassung
Während der Entwicklung führt das Zusammenwirken von Erfahrung und genetisch festgelegter Information zur Ausbildung und Optimierung neuronaler Schaltkreise und Verhaltensweisen. Daher üben Lebensbedingungen einen großen Einfluss auf das Gehirn aus. Bis heute häufen sich Nachweise dafür, dass die Haltung von Tieren in sogenannten „stimulierenden/angereicherten“ Käfigen erhebliche Auswirkungen auf das Gehirn auf molekularer, anatomischer und funktionaler Ebene hat, im Vergleich zu Tieren, die in „Standard“käfigen gehalten werden. In unserem Artikel geben wir einen kurzen Überblick dieses Forschungsgebietes und beschreiben die Unterschiede der Forschungsergebnisse auf dem Gebiet der Plastizität des visuellen Systems von Nagern, die in stimulierenden Käfigen gehalten werden im Vergleich zu Nagern aus Standardkäfigen. Außerdem gehen wir kurz auf Studien ein, die in vergleichbarer Weise die Auswirkungen von „angereicherten“ Lebensbedingungen bei Menschen untersuchen. Zusammenfassend ist festzustellen, dass Studienergebnisse immer im Kontext mit den Haltungsbedingungen der Versuchstiere gesehen werden müssen.
Einleitung und Hauptanliegen
Der kanadische Psychologe Donald Hebb ist bekannt durch seine einflussreiche Theorie darüber, wie sich Neurone des Gehirns während des Lernens anpassen, dargelegt in seinem klassischen Werk „The Organization of Behavior“ (1949). Hebbs Postulat wird oft mit der Redewendung „neurons wire together if they fire together“ (Löwel und Singer, 1992) zusammengefasst. Obwohl weniger bekannt, so ist er auch in unbeabsichtigter Weise ein Gründungsvater der Erforschung des Einflusses von „stimulierenden“ Lebensbedingungen auf das Verhalten von Tieren. In den 1940er Jahren nahm er einige Laborratten mit nach Hause und lies seine Kinder mit diesen als Haustiere spielen. Während der Zeit in Hebb’s Haus wurden die Haustierratten aus ihren Käfigen genommen und hatten so die Gelegenheit, mit den anderen Haustierratten zu spielen und sich zu sozialisieren. Hebb berichtete anekdotenhaft, dass die Haustierraten gegenüber den Laborratten bei der Lösungsfindung bestimmter Aufgaben überlegen waren (Hebb, 1947). In den 1960er Jahren zeigte der Psychologe Mark Rosenzweig, dass adulte Ratten aus angereicherten/stimulierenden Käfigen eine um 8% verdickte Großhirnrinde hatten (Rosenzweig et al., 1962). Ungeachtet dieses erstaunlichen Ergebnisses, entging die Vorstellung, dass das Gehirn adulter Ratten plastisch sein könnte (wachsen und sich ändern könnte) – eine Fähigkeit, die nur der Jugend zugestanden wurde – weiterhin der Aufmerksamkeit der Wissenschaft. Erst mit den grundlegenden Experimenten von William T. Greenough in den späten 1960er und 1970er Jahren, die ein erhöhtes Dendritenwachstum in der Sehrinde von Ratten zeigten, die in stimulierenden Käfigen, mit täglichem Austausch von Spielzeugen und wechselnder Position von hölzernen Kletterbrettern gehalten wurden, wurde der Einfluss der Lebensumwelt auf das Gehirn in den Fokus der Aufmerksamkeit gerückt. In der Tat bedeuteten diese bahnbrechenden Ergebnisse einen Paradigmenwechsel: Während man zuvor annahm, dass der Aufbau des Gehirns sehr früh in der Entwicklung und hauptsächlich auf genetischer Information basierend erfolgt, zeigten die neuen Ergebnisse, dass die Lebensumwelt einen tiefgreifenden Einfluss auf die Ausformung des Gehirns hat (eine kurze historische Übersicht findet man in Markham und Greenough, 2004).
Seit diesen ersten Untersuchungen, gibt es eine riesige und stetig wachsende Anzahl wissenschaftlicher Studien, die den Einfluss von angereicherten Lebensbedingungen auf das Gehirn untersuchen. Eine stimulierende/angereicherte Lebensumwelt (englisch: enriched environment) wird üblicherweise als „eine Kombination komplexer unbelebter und sozialer Reizung” definiert (Rosenzweig et al., 1962). Während die spezifischen Details einer angereicherten Lebensumwelt für verschiedene Spezies und verschiedene Arbeitsgruppen/Forschungsvorhaben unterschiedlich sind, ist ihnen gemeinsam, dass es erweiterte Möglichkeiten für freiwillige körperliche Betätigung und soziale und kognitive Stimulation gibt. In Bezug auf die Haltung von Nagetieren im Labor sind Standardkäfige relativ klein, üblicherweise transparent und besetzt mit einer kleinen Anzahl von Tieren (bis zu fünf) in einem ansonsten leeren Käfig, mit Holzeinstreu und Wasser und Futter ad libitum. Im Gegensatz dazu sind angereicherte/stimulierende Käfige (enriched environment, EE) größer, mit einer größeren Anzahl an Tieren besetzt und bieten eine Vielzahl von Stimulationsmöglichkeiten, wie zum Beispiel Laufräder, regelmäßig ausgetauschte Labyrinthe und Spielzeug (Abbildung 1; siehe auch van Praag et al., 2000).

Vergleich unterschiedlicher Maushaltungen aus dem Löwel-Labor: Standardkäfig, Standardkäfig mit Laufrad, stimulierender Käfig („enriched environment“).
Tiere, die in EE-Käfigen gezüchtet werden oder die dorthin umgesetzt werden, auch wenn es nur für eine kurze Zeitspanne ist, zeigen erhebliche Änderungen im Gehirn auf molekularer, anatomischer und funktionaler Ebene im Vergleich zu Tieren aus Standardkäfigen (ein Übersichtsartikel findet sich bei Sale et al., 2014). Bei Nagern führt die Anreicherung der Lebensumwelt zum Beispiel zu einer geänderten Expression von Signalmolekülen, die eine Schlüsselrolle bei der Regulation der Erregbarkeit und Plastizität des Gehirns spielen (Cancedda et al., 2004), zum Anstieg des Volumens vieler Gehirnareale (Diamond et al., 1964; Beaulieu und Colonnier, 1987), und zu verändertem mütterlichen Verhalten (Sale et al., 2004).
In unserem Artikel möchten wir einen kurzen Überblickt geben über Studien zum Einfluss einer angereicherten/stimulierenden Lebensumwelt auf den altersbedingten Rückgang der Plastizität. Früh im Leben ist das Gehirn außerordentlich plastisch, und Erfahrungen können die Organisation und Funktion neuronaler Schaltkreise leicht verändern. Mit zunehmendem Alter nimmt diese Fähigkeit zu plastischen Veränderungen ab. In unserem Übersichtsartikel werden wir anhand von Studien zum Einfluss einer angereicherten Lebensumwelt auf die Sehrinde von Nagetieren (Ratten, Mäuse), die Bedeutung der Haltungsbedingungen für die Plastizität in verschiedenen Altersabschnitten diskutieren. Dabei zeigen in Standardkäfigen gehaltene Tiere nicht nur eine stark reduzierte neuronale Plastizität, sondern auch einen mit dem Alter schnell fortschreitenden Abfall der Plastizität, der durch Haltung in EE-Käfigen verhindert werden kann.
Die Anreicherung der Lebensumwelt hatte außerdem positive Auswirkungen in Mausmodellen vieler Hirnerkrankungen, wie der Huntington-Krankheit, Multipler Sklerose, Epilepsie, Parkinson-Krankheit, Alzheimer-Krankheit, Schizophrenie, Autismus und Depression (Hannan, 2014; Mo et al., 2015; Fischer, 2015). Obgleich eine detaillierte Diskussion translationaler Aspekte und klinischer Studien über den Rahmen dieses Übersichtsartikels hinausgeht, möchten wir einige Beispiele vergleichbarer Befunde in Studien zur Anreicherung der Lebensumwelt bei gesunden Probanden vorstellen.
Wie auch schon von anderen Autoren festgestellt wurde, ist „angereichert/stimulierend“ offensichtlich ein relativer Begriff, und Standardkäfige für die Haltung von Labornagern entsprechen vielmehr einer verarmten, und nicht einer ‚normalen‘ Lebensumwelt (Hannan, 2014). Obwohl der Einfluss der Haltungsbedingungen in vorklinischen und translationalen medizinischen Studien üblicherweise berücksichtigt wird, insbesondere bei der Bewertung der Konstruktvalidität von Mausmodellen (ausführliche Übersichten dazu in Tkacs und Thompson, 2006; Burrows und Hannan, 2013; Burrows et al., 2015; Mo et al., 2015), wurde dagegen im Bereich der Grundlagenforschung den Haltungsbedingungen und anderen kontextuellen Faktoren erst in jüngster Zeit Beachtung geschenkt. Wir vertreten die Ansicht, dass es essenziell ist, den Einfluss der Haltungsbedingungen auf „Basis“- oder Kontrolldaten zu berücksichtigen, wenn Daten von Tierversuchsstudien sinnvoll interpretiert werden sollen.
Stimulierende Lebensbedingungen wirken sich auf die Fähigkeit und Schnelligkeit erfahrungsabhängiger Plastizität in der Sehrinde von Nagern aus
Okuläre Dominanz (englisch: ocular dominance) oder Augenpräferenz bezeichnet die relative Stärke neuronaler Antworten in der primären Sehrinde (V1) auf Eingänge eines Auges im Vergleich zu denen des anderen Auges. Die sogenannte „Augendominanzplastizität“ in V1 von Säugetieren ist eines der am besten untersuchten Modelle erfahrungsabhängiger Plastizität (Wiesel und Hubel, 1963; Espinosa und Stryker, 2012). Sie stellt auch ein gut charakterisiertes Paradigma dar, mit dem die Auswirkungen einer angereicherten Lebensumwelt auf plastische Veränderungen getestet werden können. Ähnlich wie bei Menschen ist auch die Sehrinde von Nagern aufgeteilt in einen monokularen Anteil, der Eingänge nur vom gegenüberliegenden (kontralateralen) Auge erhält, und in einen binokularen Anteil, der Eingänge von beiden Augen erhält. Die klassischen Experimente von Hubel und Wiesel in den 1960er Jahren bei Katzen zeigten, dass eine Störung der normalen binokularen Seheindrücke nach Verschluss eines Auges (monokulare Deprivation, ein Katarakt-Modell) während einer frühen Phase der postnatalen Entwicklung irreversible Veränderungen der V1 Schaltkreise verursacht. Ebenso ist die neuronale Aktivität in V1 von Nagern normalerweise vom kontralateralen Auge dominiert, jedoch verschiebt sich die Augendominanz nach monokularer Deprivation in Richtung des offenen Auges (Dräger, 1975; Dräger, 1978). Bei Mäusen, die in Standardkäfigen aufgezogen werden, ist die Augendominanzplastizität bei jungen Tieren maximal (postnataler Tag (P) 28; Dräger, 1978; Gordon und Stryker, 1996; Sawtell et al., 2003), und lässt mit zunehmendem Alter nach (Gordon und Stryker, 1996, Cang et al., 2005). Bei erwachsenen (adulten) Mäusen sind signifikante Veränderungen der Augendominanz bis zu einem Alter von etwa 110 Tagen noch möglich, benötigen dafür aber längere Phasen einer monokularen Deprivation (7 Tage im Vergleich zu nur 4 Tagen bei jungen Mäusen) (Sato und Stryker, 2008). Bei noch älteren Standardkäfigtieren konnte selbst eine 14 Tage andauernde monokulare Deprivation keine Veränderung der Augendominanz bewirken (Lehmann und Löwel, 2008; Espinosa und Stryker, 2012; Levelt und Hübener, 2012).
Die Verwendung von stimulierenden Käfigen beeinflusste diese durch Deprivation ausgelösten Veränderungen der Aktivierung von V1 in starkem Maße (Greifzu et al., 2014) (Abbildung 2). In einer ersten Studie verglichen wir Mäuse aus Standard- bzw. stimulierenden Käfigen bis zu einem Alter von ~200 Tagen. Im Unterschied zu Mäusen aus Standardkäfigen, die im Alter von >130 Tagen keine Augendominanzplastizität mehr zeigten, wiesen selbst die ältesten Mäuse aus den stimulierenden Käfigen noch deutliche Aktivitätsänderungen in V1 und damit Plastizität nach einer monokularen Deprivation von 7 Tagen auf. Da ein Alter von 200 Tagen für eine Maus nicht wirklich ‚betagt‘ ist, testeten wir im Anschluss noch ältere Tiere und stellten überraschenderweise fest, dass die mit optischen Methoden gemessene Augendominanzplastizität bei den Mäusen aus stimulierenden Käfigen lebenslang erhalten bleibt (Greifzu et al., 2016). Bemerkenswerterweise konnte auch ein Umsetzen von Mäusen aus einem Standardkäfig in einen stimulierenden Käfig in einem fortgeschrittenen Alter (nach 110 Tagen) die Fähigkeit zur Plastizität der Augendominanz wiederherstellen: Selbst die älteste untersuchte Maus (922 Tage alt) zeigte noch plastische Veränderungen der V1 – Aktivität (Abbildung 2, rechts unten) (Greifzu et al., 2014; Greifzu et al., 2016).

Die Plastizität des Gehirns nimmt mit dem Alter ab: Eine stimulierende Lebensumwelt kann sowohl der altersabhängigen Abnahme der Plastizität entgegenwirken als auch helfen, Hirnplastizität wiederherzustellen. A: Schema der altersabhängigen Abnahme der Hirnplastizität: Im jungen Gehirn gibt es Phasen stark erhöhter Plastizität. B: Eine stimulierende Lebensumwelt bewahrt eine lebenslange Augendominanzplastizität in der primären Sehrinde (V1) von Mäusen und kann Plastizität auch dann wiederherstellen, wenn die Anreicherung der Lebensbedingungen erst spät erfolgt (nach dem Tag 110). Aktivitätsabhängige Änderungen in der Aktivierung von V1 wurden durch optisches Ableiten intrinsischer Signale visualisiert, nach Reizung des gegenüberliegenden (kontralateralen) oder ipsilateralen Auges vor (obere Reihe) und nach Deprivation eines Auges (untere Reihe). Grauwertkodierte Aktivitätskarten aus dem binokularen Bereich von V1 sind dargestellt: Dunklere Bereiche entsprechen einer stärkeren Aktivierung der Sehrinde. Zusätzlich sind zweidimensionale Augendominanzkarten (OD-map) und das Histogramm der Augendominanzwerte (OD-index) abgebildet. Vor der Deprivation (der schwarze Punkt markiert das deprivierte Auge) ist die Aktivitätsregion in V1 sowohl bei Mäusen aus Standardkäfigen (links) als auch bei Tieren aus stimulierenden Käfigen (rechts) nach visueller Reizung des kontralateralen (contra) Auges dunkler als nach Reizung des ipsilateralen (ipsi) Auges, die zweidimensionale Augendominankarte zeigt warme Farben (rot repräsentiert positive, blau negative Augendominanzwerte) und der mittlere OD-index ist positiv, was zeigt, dass die Aktivität in der Sehriinde vom kontralateralen Auge dominiert wird. Nach Deprivation des kontralateralen Auges verändert sich die Augendominanz in Richtung des offenen (ipsilateralen) Auges nur bei den Tieren aus stimulierenden Käfigen, aber nicht bei adulten Standardkäfigmäusen: Kältere Farben herrschen jetzt in der Augendominanzkarte vor und der OD-index ist reduziert (blaue Pfeile). Bitte beachten Sie, dass die abgebildete Aktivitätskarte nach monokularer Deprivation (unten rechts) von einer 922 Tage alten Maus (aus einem stimulierenden Käfig) stammt, die trotz ihres hohen Alters immer noch Augendominanzplastizität zeigte. Verändert nach Greifzu et al. (2014; 2016).
Ein weiteres altersabhängiges Merkmal der Augendominanzplastizität ist, dass ältere Mäuse aus Standardkäfigen dafür eine länger anhaltende monokulare Deprivation benötigen im Vergleich zu jüngeren Tieren. Während junge Mäuse aus Standardkäfigen bereits nach 4 Tagen monokularer Deprivation eine Veränderung der Augendominanz zeigen, sind für ältere Tiere mindestens 7 Tage nötig, um eine gleichstarke Verschiebung der Augendominanz zu erzeugen, gemessen mit optischen und elektrophysiologischen Messmethoden (Gordon und Stryker, 1996; Lehmann und Löwel, 2008; Sato und Stryker, 2008). Und wie sieht es bei Mäusen aus stimulierenden Käfigen aus? Wir konnten kürzlich zeigen, dass die erfahrungsabhängigen Aktivitätsänderungen in V1 bei diesen Mäusen bereits nach zwei Tagen monokularer Deprivation visualisierbar waren, und dies sogar bei allen getesteten Altersstufen (bis zum Alter von 283 Tagen), was nahelegt, dass die Anreicherung der Lebensumwelt diese schnelle Plastizität in jedem Alter ermöglicht. Bei einigen jungen Mäusen aus stimulierenden Käfigen war eine Plastizität der Augendominanz sogar schon nach einem Tag monokularer Deprivation messbar (Kalogeraki et al., 2017). Diese Daten verdeutlichen, dass die Haltungsbedingungen nicht nur auf die Fähigkeit zur erfahrungsbedingten Plastizität in der Sehrinde einwirken, sondern auch deren Geschwindigkeit beeinflussen; es ist deshalb möglich, dass die Standardkäfighaltung die erfahrungsabhängige Plastizität neuronaler Schaltkreise stark verlangsamt.
Während die Plastizität generell mit dem Alter abnimmt, ist sie zusätzlich auch nach Gehirnverletzungen beeinträchtigt. Beispielsweise zeigen adulte Mäuse aus Standardkäfigen nach kleinen Schlaganfallläsionen in Hirnrindenregionen außerhalb der Sehrinde keine Augendominanzplastizität mehr: Dies gilt für Läsionen in der primären Fühlrinde (somatosensorischer Kortex) (Greifzu et al., 2011, Greifzu et al., 2012) oder auch für Läsionen im noch weiter von V1 entfernten motorischen Kortex (Pielecka-Fortuna et al., 2015). Interessanterweise war dies nicht der Fall, wenn wir Mäuse aus stimulierenden Käfigen untersuchten: Augendominanzplastizität blieb bei diesen Tieren nicht nur bis in ein höheres Alter erhalten, sondern auch nach einem Schlaganfall in der Fühlrinde (Greifzu et al., 2014). Im nächsten Schritt überprüften wir, ob auch junge Mäuse (aus Standardkäfigen) weniger durch einen Schlaganfall im somatosensorischen Kortex beeinträchtigt sind. Dies war in der Tat der Fall: junge Standardkäfigmäuse zeigten weiterhin Plastizität in ihrer Sehrinde, wie dies bei Tieren ohne Läsion der Fall ist. Somit sind Mäuse vor (zumindest einigen) schlaganfallbedingten Beeinträchtigungen ihrer Hirnplastizität geschützt, wenn sie entweder jung oder in stimulierenden Käfigen aufgewachsen sind. Mit anderen Worten zeigen adulte Mäuse eine beeinträchtigte Augendominanzplastizität nach Schlaganfall nur, wenn sie in der verarmten Lebensumwelt von Standardkäfigen leben.
Was können wir aus diesen Beobachtungen schlussfolgern? Da das Halten von Mäusen in weniger verarmten bzw. stimulierenden Käfigen deren Hirnplastizität bis ins hohe Alter und sogar nach Schlaganfallläsionen bewahrt, ist unserer Meinung nach die angemessenste Schlussfolgerung aus den vorhandenen Daten, dass sich Standardkäfighaltung stark nachteilig auf Hirnplastizität auswirkt und zu einer rapiden Abnahme der Augendominanzplastizität mit dem Alter führt.
Wie verstärkt eine stimulierende Lebensumwelt Plastizität?
Eine wichtige Frage ist es, ob der Augendominanzplastizität bei Tieren aus stimulierenden Käfigen im Vergleich zu jungen Tieren aus verarmten Standardkäfigen gemeinsame oder unterschiedliche Wirkmechanismen zugrunde liegen. Die Balance zwischen neuronaler Erregung und Hemmung ist ein Hauptunterschied zwischen juvenilen und adulten Tieren aus Standardkäfigen, und der Reifung inhibitorischer (hemmender) Schaltkreise wird eine wichtige Rolle bei den altersbedingten Veränderungen der Plastizität zugeschrieben (Hensch, 2005). So führt beispielweise bei jungen Mäusen aus Standardkäfigen eine pharmakologisch gesteigerte Hemmung zu einem vorzeitigem Verlust der Plastizität in der Sehrinde, wohingegen in umgekehrter Weise eine Reduktion der Hemmung die Augendominanzplastizität bei adulten Tieren begünstigt (Maya-Vetencourt et al., 2008; Harauzov et al., 2010; Morishita et al., 2010). Dies legt nahe, dass ein niedrigeres Niveau an Hemmung Plastizität erlauben könnte und dass eine Reduktion der Hemmung eine Grundvoraussetzung für die Expression von Plastizität bei Erwachsenen ist. Diese Hypothese wird durch den Befund gestützt, dass das Niveau der intrakortikalen Hemmung in V1 bei adulten Mäusen (>130 Tage) aus stimulierenden Käfigen genauso niedrig war wie in jungen Mäusen aus Standardkäfigen (Greifzu et al., 2014). Gleichermaßen ging bei Ratten eine durch eine stimulierende Lebensumwelt induzierte Plastizität (nach langanhaltender monokularer Deprivation) mit einer Erniedrigung der extrazellulären GABA-Konzentration einher, was auf eine Reduktion der Hemmung hinweist (Sale et al., 2007, Baroncelli et al., 2010).
Allerdings gibt es Hinweise darauf, dass die Effekte durch Anreicherung der Lebensumwelt nicht einfach eine Reaktivierung eines jugendlichen Status des Gehirns sind. So gibt es epigenetische und sogar generationsübergreifende Auswirkungen einer stimulierenden Lebensumwelt auf die Ausprägung der Plastizität: Beispielsweise haben Knockout-Mäuse (ras-grf-/-) eine gestörte Langzeitpotenzierung im Hippocampus, wenn sie in Standardkäfigen gehalten werden; dieser Phänotyp ist aber verschwunden nachdem die Mäuse für nur zwei Wochen in stimulierenden Käfigen gehalten wurden, und der Effekt der Anreicherung wurde sogar auf den wieder in Standardkäfigen herangewachsenen Nachwuchs übertragen (Arai et al., 2009; Arai und Feig, 2011). Weitere Veränderungen, die mit einer Anreicherung der Lebensumwelt einhergehen, wurden ebenso mit der Verstärkung der Plastizität adulter Nager in Zusammenhang gebracht: Eine stimulierende Lebensumwelt veränderte die Expression verschiedener, bedeutender Signalfaktoren, die bekanntermaßen kortikale Aktivität und Plastizität beeinflussen, darunter der brain-derived neurotrophic factor (Falkenberg et al., 1992; Ickes et al., 2000; Cancedda et al., 2004; Sale et al., 2004), Serotonin (Baroncelli et al., 2010), der nerve growth factor (Mohammed et al., 1993; Pham et al., 1999), und der insulin-like growth factor (IGF; Carro et al., 2000; Ciucci et al., 2007).
Eine angereicherte Lebensumwelt kombiniert motorische, soziale, kognitive, und multisensorische Stimulation und übt dadurch einen globalen Einfluss auf das Gehirn aus (van Praag et al., 2000). Immer mehr Befunde weisen darauf hin, dass sogenannte primär sensorische Hirnrindenareale multimodale Eingänge über kortiko-kortikale Netzwerke erhalten und verarbeiten (Ghazanfar und Schroeder, 2006; Kayser und Logothetis, 2007; Driver und Noesselt, 2008; Henschke et al., 2017). Folglich beeinflusst die Anreicherung der Lebensumwelt neuronale Aktivität und Plastizität in mehreren kortikalen und subkortikalen Gehirnbereichen gleichzeitig und verändert kortiko-kortikale Netzwerkinteraktionen auch über lokale Aktivitäten hinaus. Beispielsweise war bei in stimulierenden Käfigen gehaltenen Mäusen die Kopplung lokaler Feldpotenziale (LFPs) zwischen der Sehrinde und dem Motorkortex reduziert, während die Tiere einen Käfig erkundeten, und zwar in Zeitfenstern, die auf eine Dekorrelation der direkten monosynaptischen Verbindungen zwischen den beiden Arealen hinweist. Diese Dekorrelation könnte bedeuten, dass sich das Gehirn der bereicherten Mäuse während der Exploration in einem aktiveren und stärker stimulierten Zustand befindet (Di Garbo et al., 2011). Als weiterer Hinweis auf die Bedeutung von Netzwerkinteraktionen für Plastizität verhinderte eine angereicherte Lebensumwelt zum Beispiel Begleiterscheinungen des Alterungsprozesses, die bei Mäusen aus Standardkäfigen auftreten, wie eine Verschiebung des LFP-Spektrums und eine Dekorrelation zwischen LFPs des primären auditorischen Kortex und V1 (Mainardi et al., 2014).
Die durch eine Anreicherung der Lebensumwelt ausgelöste Plastizität ist nicht zwangsläufig modalitätsspezifisch. In der Tat benötigt Sehrindenplastizität keine visuelle Anreicherung per se. Ratten, die in Dunkelheit in einer angereicherten Lebensumwelt aufgezogen wurden, entwickelten normale Sehschärfe, während Ratten, die in Standardkäfigen in Dunkelheit aufgezogen wurden, eine verminderte Sehschärfe zeigten (Bartoletti et al., 2004). Des Weiteren zeigten Rattenwelpen, die eine Körpermassage erhielten (ohne weitere Formen der Lebensumweltanreicherung) eine beschleunigte Entwicklung der Sehschärfe, begleitet von einem Anstieg der IGF-Expression in verschiedenen Gehirnarealen, ein Effekt, der auch bei Mehrkomponentenanreicherung zu beobachten war (Guzzetta et al., 2009). Diese Ähnlichkeit könnte von direkten Verbindungen zwischen den Hirnarealen, die durch einzelne Komponenten der Anreicherung der Lebensumwelt (z. B. Körpermassage) stimuliert werden, herrühren, und/oder durch konvergente Effekte verschiedener Anreicherungskomponenten der Lebensumwelt auf gemeinsam benutzte molekulare Signalwege (Maya-Vetencourt and Origlia, 2012; Vivar et al., 2013).
Auswirkungen einer angereicherten Lebensumwelt wurden auch in anderen Sinnesmodalitäten beobachtet. Im auditorischen System bewirkte eine Anreicherung der Lebensumwelt gepaart mit passiver akustischer Stimulation einen Anstieg der Antwortstärke und ein Absenken der Antwortschwelle von Neuronen der Hörrinde (e.g. Dinse, 2004; Engineer et al., 2004), und eine veränderte zeitliche Informationsverarbeitung und räumliche Abbildung des Schalles (Percaccio et al., 2005; Kilgard et al., 2007; Cai et al.,2009; Cai et al., 2010; Jakkamsetti et al., 2012). Eine akustische Anreicherung der Lebensumwelt förderte außerdem eine Erholung von früher lärminduzierter auditorischer Fehlfunktion (Zhu et al., 2014; Jiang et al., 2015; Sturm et al., 2017), und es wurde kürzlich auch gezeigt, dass eine längerfristige körperliche Betätigung das Voranschreiten des altersbedingten Hörverlustes verlangsamen kann (Han et al., 2016). Desgleichen wurden auch plastizitätsfördernde Auswirkungen einer Anreicherung der Lebensumwelt für den somatosensorischen Kortex gezeigt (e.g. Coq und Xerri, 1998; Florence et al., 2001; Godde et al., 2002; Bourgeon et al., 2004; Polley et al., 2004; Landers et al., 2011).
Sind alle Komponenten der stimulierenden Käfige notwendig, um die Plastizität zu erhalten?
Wie oben bereits erwähnt, beinhaltet das Konzept der angereicherten Lebensumwelt die Kombination von physischer Aktivität, sozialer und kognitiver Stimulation. Die relative Bedeutung jeder einzelnen Komponente für die nach Anreicherung induzierten Veränderungen hängt vom jeweils betrachteten Ergebnis und dem spezifischen experimentellen Modell ab, was auf vielfache Wirkmechanismen und höchstwahrscheinlich kombinatorische Effekte hinweist (Hannan, 2014). Obwohl schwer abgrenzbar, scheinen die sozialen Komponenten der Lebensumweltanreicherung eine weniger bedeutende Rolle für die Plastizität der Sehrinde von Nagern zu spielen – im Vergleich zu visuellen Komponenten oder physischer Aktivität. Obwohl sich Einzelhaltung nachteilig auf Plastizität von Mäusen auswirkte (Balog et al., 2014), war eine Erhöhung der Gruppengröße (überwiegend eine soziale „Anreicherung“) nicht ausreichend, um die Auswirkungen der Plastizität bei Ratten zu verändern (Rosenzweig et al., 1978; Baroncelli et al., 2012). Im Gegensatz dazu stellte die Beschäftigung mit einer visuellen Lernaufgabe (was einer sensorischen, kognitiven und motorischen Reizung entspricht, ohne soziale Aspekte) die Sehschärfe amblyoper Ratten wieder her (Baroncelli et al., 2012). In ähnlicher Weise führte bei amblyopen Mäusen eine Kombination von körperlicher Aktivität und visueller Stimulation zu einer schnelleren Wiederherstellung der Sehschärfe (Kaneko und Stryker, 2014). Allerdings konnte auch alleinige visuelle Anreicherung der Lebensumwelt die Augendominanzplastizität bei in Standardkäfigen gehaltenen Mäusen fördern (Matthies et al., 2013).
Des Weiteren scheint auch freiwillige physische Aktivität einen entscheidenden Beitrag für die durch die stimulierenden Haltungsbedingungen verursachten Änderungen der Plastizität zu leisten: Das Hinzufügen eines Laufrades zu einem (leicht größeren) Standardkäfig reichte aus, um die Augendominanzplastizität sowohl bis ins hohe Alter zu bewahren, als auch sie bei gealterten Mäusen und nach einem Schlaganfall wiederherzustellen (Kalogeraki et al., 2014; Kalogeraki et al., 2016). Beachtenswert ist hierbei, dass das Benutzen eines Laufrades auch erst nach Auftreten des Schlaganfalles ausreichend war, um die Augendominanzplastizität in V1 der Mäuse zu reaktivieren. Demnach förderte freiwilliges Laufen alleine bereits effektiv die Plastizität in der Sehrinde adulter Mäuse, auch wenn erst spät damit begonnen wurde. Laufen ist ein besonders interessanter Parameter, weil es nicht nur die Plastizität und Bildung neuer Zellen im Gehirn fördert, sondern weil es auch nachweislich die Aufmerksamkeit zum Beispiel für visuelle Stimuli verändert (Stryker, 2014; Fu et al., 2015; Pakan et al., 2016; Cooper et al., 2017).
Die Plastizität des menschlichen Gehirns ähnelt den Effekten einer stimulierenden Lebensumwelt, die in Tierstudien gezeigt wurden
Die Förderung der Plastizität durch stimulierende Lebensbedingungen und durch freiwillige physische Aktivität in Tierstudien erinnert an die positiven Effekte von physischer Aktivität oder einem „aktiveren Lebensstil“ bei erwachsenen oder älteren Menschen. Obwohl es schwieriger ist, genau zu definieren, welche Komponenten einer angereicherten Lebensumwelt notwendig sind, um die Plastizität des menschlichen Gehirns zu fördern, gibt es eine große Anzahl von Ergebnissen aus Humanstudien, die in auffälliger Weise den Ergebnissen von Tierstudien ähneln (Übersichten in Hertzog et al., 2008; Hotting und Röder, 2013; Voss et al., 2013). Beispielsweise wurde für gesunde, 65-84 Jahre alte Probanden gezeigt, dass Tanzen, welches eine Kombination von physischer Aktivität, sozialer Interaktion, sensorischer und kognitiver Stimulation ist, die Leistungen in einer Reihe von kognitiven, taktilen und motorischen Aufgaben erhöht (Kattenstroth et al., 2013; Dinse, 2016).
Ebenso wurde gezeigt, dass bei älteren Erwachsenen Änderungen der körperlichen Fitness nach Training auf stationären Fahrrädern mit höherer reizspezifischer Aktivierung in der Sehrinde korrelierten, die mit kognitiven Fähigkeiten assoziiert ist und typischerweise während des Alterns abnimmt (Kleemeyer et al., 2017). Eine weitere Studie identifizierte regelmäßige physische Aktivität als eine präventive Maßnahme gegen eine altersbedingte Verschlechterung der Geruchsdetektionsschwelle (Schubert et al., 2017) und die Alzheimer-Krankheit (Erickson et al., 2012; Santos-Lozano et al., 2016). Schließlich ist zeitlich reglementiertes Videospielen eine Form der Anreicherung der Lebensumwelt für Menschen, da es intensive visuelle Stimulation und sensomotorische Integration mit kognitiver Reizung und einer Belohnung für das Erreichen des Spielziels kombiniert. Sowohl Action- als auch Nicht-Actionspiele verbesserten nicht nur visuelle Fähigkeiten wie Kontrastsensitivität, Sehschärfe und räumliches Sehen, sondern verbesserten auch Lernen, Aufmerksamkeit und kognitive Fähigkeiten bei nicht direkt mit dem Spiel verbundenen Aufgaben und sind daher ein vielversprechendes Instrument zur Steigerung der Plastizität bei gesunden, älteren Erwachsenen und zur Verbesserung des Sehens bei amblyopen Patienten (Li et al., 2009; Li et al., 2011; Bavelier et al., 2012; Stryker und Löwel, 2017). Maßnahmen basierend auf einer Anreicherung der Lebensumwelt haben sich auch als wirkungsvoll erwiesen bei der Behandlung vieler Krankheiten, wie Depression, Schizophrenie und Autismus, die mit einer gewissen ‚Selbst-Deprivation‘ einhergehen (das heißt, mit sozialem Rückzug, Überempfindlichkeit und Vermeidungsreaktionen gegenüber sensorischen Reizen, verminderter Neigung zur Suche nach Neuem) (Mabunga et al., 2015). Diese Befunde aus Tierstudien ergänzend, besteht in Altenpflegeeinrichtungen ein erhöhtes Bewusstsein für die positiven Auswirkungen auf die mentale und physische Gesundheit, die mit zusätzlichen Aktivitäten zur Förderung der sozialen Interaktion, physischen Aktivität und kognitiven/sensorischen Stimulation erzielt wird, im Gegensatz zur bloßen Befriedigung der Grundbedürfnisse.
Zusammenfassend verdeutlicht die bestehende Literatur den großen Einfluss der Lebensumwelt auf die Plastizität und Leistungsfähigkeit des gesunden und erkrankten oder alternden Gehirns sowohl bei Tiermodellen als auch beim Menschen. Deshalb ist es für die Interpretation und zum Vergleich von Tierstudien zur Plastizität des Gehirns von essenzieller Bedeutung, sämtliche Details der Haltung zu benennen. Angesichts der Tatsache, dass auch kurzzeitige und nur zeitweilige Anreicherung der Lebensumwelt einen deutlichen Einfluss auf Hirnplastizität haben, müssen des Weiteren Verfahren wie Habituation, Training und Verhaltensversuche als eine Form von Anreicherung bewertet und dementsprechend beschrieben werden. Es ist deutlich geworden, dass für die Untersuchung der ‚normalen‘ Prozesse der Plastizität in einem Tiermodell eines gesunden Gehirns, eine angereicherte/stimulierende Lebensumwelt den natürlichen Lebensbedingungen wesentlich näher kommt und damit eine überlegene Validität hat. Schließlich sollte die Haltung von Versuchstieren in Standardkäfigen bewusster als die stark verarmte Lebensumwelt behandelt werden, die sie darstellt, mit allen bekannten Konsequenzen von Deprivation auf die Funktionsfähigkeit und Plastizität des Gehirns.
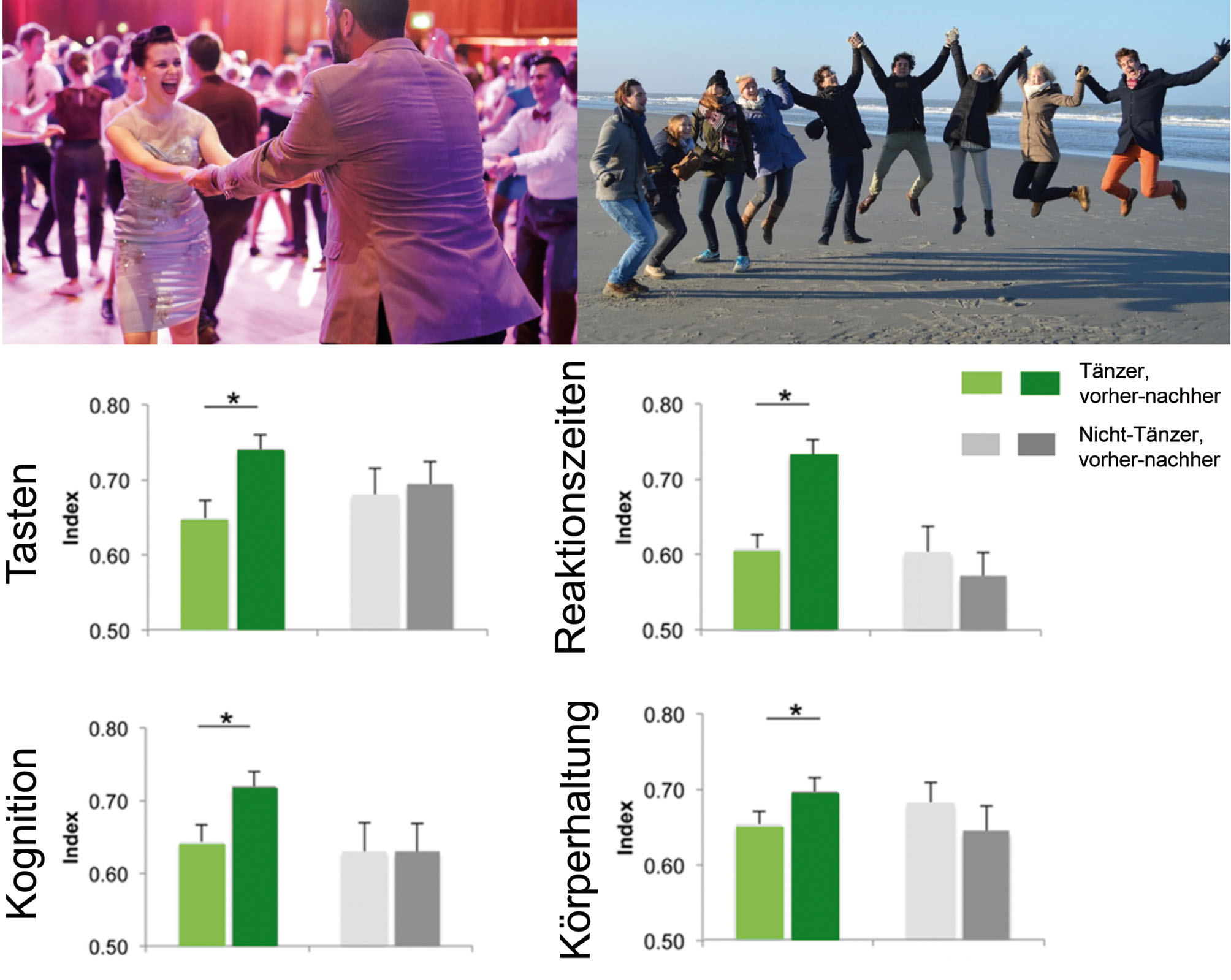
Treiben Sie Sport, treffen Sie Freunde: Es wurde wiederholt nachgewiesen, dass ein aktiver Lebensstil sowohl einem altersabhängigem Abbau kognitiver, sensorischer und motorischer Leistungen entgegenwirken, als auch therapeutisch wirksam sein kann, indem er positive Effekte bei einer Reihe von neurodegenerativen Erkrankungen bewirkt. Bildquelle (oben links): www.jazzmad.co.uk/learn-to-swing-dance/. Untere Grafiken: Sechs Monate Tanzen (eine Stunde/Woche) verbesserte die Körperhaltung, sensomotorische und kognitive Leistungen älterer Menschen (Abbildung verändert nach Kattenstroth et al., 2013): Gemittelte Indizes, die individuelle Leistungen der Probanden der Tanzgruppe (grün) und der Kontrollgruppe (grau) vor (vorher, hellere Farbe) und nach (nachher, dunklere Farbe) einer sechs-monatigen Zeitspanne mit oder ohne Tanzen charakterisieren. Um die Leistungen über alle Tests und Probanden vergleichen zu können, wurden die normalisierten Leistungsindizes für alle Probanden und jeden Test wie folgt berechnet: (wp-ip)/(wp-bp), wobei wp der schlechtesten Leistung aller Probanden, ip der individuellen Leistung, und bp der besten Leistung aller Probanden entspricht. Die beste ip ist 1, die schlechteste 0. Indizes wurden danach über alle zu einem Bereich gehörenden Tests gemittelt, wie oben beschrieben. Tasten (p≤0.001) umfasst Tastschwelle, Zweipunktdiskrimination und haptische Objekterkennung. Reaktionszeiten (p≤0.001) beinhaltet Multiple-Choice-Reaktionszeiten für die linke und rechte Hand und Reaktionszeitanalysen. Kognition (p≤0.001) umfasst den geriatrischen Konzentrationstest (AKT), Ravens Standard Progressiven Matrizentest (RSPM), das Frankfurter Aufmerksamkeitsinventar (FAIR), und den Non-Verbalen Lerntest (NVLT). Körperhaltung (p=0.001) beinhaltet Haltungs- und Gleichgewichtsleistungen bei sieben statischen und dynamischen Tests auf einer Messplattform. Die vertikalen Balken zeigen den Standardfehler des Mittelwerts. Sternchen markieren signifikante Unterschiede vor und nach der Tanzintervention oder nach sechs Monaten ohne Tanzen. Sogar eine moderate Tanzzeit konnte einer großen Menge altersabhängiger Verschlechterungen entgegenwirken. Mehr Details finden sich in Kattenstroth et al. (2013).
About the authors

Siegrid Löwel studierte Biologie in Würzburg und Frankfurt/M., 1988 Promotion (Dr. phil. nat.) und 1995 Habilitation in Zoologie an der Johann-Wolfgang-Goethe-Universität in Frankfurt am Main, bis 1996 wissenschaftliche Mitarbeiterin am Max-Planck- Institut für Hirnforschung in Frankfurt am Main in der Neurophysiologischen Abteilung von Prof. Dr. Wolf Singer, 1997–2005 Leiterin der Forschergruppe „Visuelle Entwicklung und Plastizität“ am Leibniz- Institut für Neurobiologie in Magdeburg, 2002–2003 Research Associate Professor am Keck-Center der University of California in San Francisco, U.S.A., im Labor von Prof. Dr. Michael P. Stryker, 2003-2004 Dorothea-Erxleben-Gastprofessorin an der Otto-von-Guericke-Universität Magdeburg, 2004–2005 Stipendiatin im Hertie-Exzellenz-Programm „Neurowissenschaften“, 2005–2010 Universitätsprofessorin (W2) für Neurobiologie an der Friedrich-Schiller-Universität Jena, seit August 2010 Universitätsprofessorin (W3) und Leiterin der Abteilung „Systemische Neurobiologie“ an der Fakultät für Biologie und Psychologie, Georg-August-Universität Göttingen. Das Hauptinteresse von Prof. Löwel liegt darin, zu erforschen wie sich die Flexibilität unseres Gehirns im Alter und nach Läsionen erhöhen lässt und welche Mechanismen dieser Plastizität zugrunde liegen.

Evgenia Kalogeraki studierte Biologische Technologie an der Universität Ioannina, Griechenland. 2011: Master in Molekularbiologie und Biomedizin, Universität von Kreta. 2012: Beginn ihrer Promotion in der Arbeitsgruppe von Prof. Dr. Siegrid Löwel und Aufnahme in die Göttinger Graduiertenschule für Neurowissenschaften, Biophysik und Molekulare Biowissenschaften (GGNB). 2015: Promotion. Im Augenblick forscht sie als Postdoktorandin in der Abteilung „Systemische Neurobiologie“ der Universität Göttingen.
Susanne Dehmel studierte Biologie in Braunschweig und Leipzig und promovierte 2006 in der Arbeitsgruppe Neurobiologie von Prof. R. Rübsamen am Zoologischen Institut der Universität Leipzig. Von 2007 bis 2011 war sie Postdoktorandin in der Arbeitsgruppe Sensory Neurobiology von Prof. S. Shore am Kresge Hearing Research Institute, University of Michigan. Seit 2011 ist sie Postdoktorandin in der Abteilung Systemische Neurobiologie von Prof. Dr. Siegrid Löwel an der Universität Göttingen.

Kalina Makowiecki studierte Psychologie (2010, BA Hons) und promovierte zum PhD in Experimental and Regenerative Neurosciences (School of Biological Sciences) an der University of Western Australia (UWA) in Perth (2016), gefördert durch einen Preis und ein Stipendium (Australian Postgraduate Award und UWA Completion Scholarship). Im Jahr 2016 begann sie als Postdoktorandin in der Abteilung „Systemische Neurobiologie“ (Prof. Dr. Siegrid Löwel) an der Universität Göttingen. Sie wurde kürzlich mit einer Dorothea-Schlözer-Stelle für Postdoktorandinnen ausgezeichnet, die im Oktober 2017 begann.
Danksagung
Die Autorinnen danken allen Kolleginnen und Kollegen, mit denen sie im Rahmen der hier vorgestellten Projekte zusammengearbeitet haben, insbesondere Drs. Franziska Greifzu and Justyna Pielecka-Fortuna. Besonderer Dank geht an Prof. Dr. Hubert Dinse für die Graphiken der Abbildung 3, an Simone Kleinhans für das Korrekturlesen der deutschen Fassung, und an Matthias Schink für seine Betreuung unserer Tierkolonie. Ein Teil der hier vorgestellten Arbeiten wurden gefördert durch das Bundesministerium für Bildung und Forschung (BMBF) 01GQ0921 und 01GQ0810 und durch Mittel der Deutschen Forschungsgemeinschaft (DFG) über den Sonderforschungsbereich 889 „Zelluläre Mechanismen sensorischer Verarbeitung“ an SL (Projekt B5); wir bedanken uns auch für Unterstützung durch das Dorothea Schlözer Programm der Universität Göttingen (KM).
Literatur
Arai, J. A., Li, S., Hartley, D. M. and Feig, L. A. (2009). Transgenerational rescue of a genetic defect in long-term potentiation and memory formation by luvenile enrichment. J. Neurosci. 29, 1496–1502.10.1523/JNEUROSCI.5057-08.2009Search in Google Scholar PubMed PubMed Central
Arai, J. A. and Feig, L. A. (2011) Long-lasting and transgenerational effects of an environmental enrichment on memory formation. Brain Res. Bull. 85, 30–5.10.1016/j.brainresbull.2010.11.003Search in Google Scholar PubMed PubMed Central
Balog, J., Matthies, U., Naumann, L., Voget, M., Winter, C. and Lehmann, K. (2014). Social experience modulates ocular dominance plasticity differentially in adult male and female mice. Neuroimage. 103, 454–461.10.1016/j.neuroimage.2014.08.040Search in Google Scholar PubMed
Baroncelli, L., Bonaccorsi, J., Milanese, M., Bonifacino, T., Giribaldi, F., Manno, I., Cenni, M. C., Berardi, N., Bonanno, G., Maffei, L. and Sale, A. (2012). Enriched experience and recovery from amblyopia in adult rats: Impact of motor, social and sensory components. Neuropharmacol. 62, 2388–2397.10.1016/j.neuropharm.2012.02.010Search in Google Scholar PubMed
Baroncelli, L., Sale, A., Viegi, A., Maya-Vetencourt, J. F., De Pasquale, R., Baldini, S. and Maffei, L. (2010). Experience-dependent reactivation of ocular dominance plasticity in the adult visual cortex. Exp. Neurol. 226, 100–109.10.1016/j.expneurol.2010.08.009Search in Google Scholar PubMed
Bartoletti, A., Medini, P., Berardi, N. and Maffei, L. (2004). Environmental enrichment prevents effects of dark-rearing in the rat visual cortex. Nat. Neurosci. 7, 215–216.10.1038/nn1201Search in Google Scholar PubMed
Bavelier, D., Green, C. S., Pouget, A. and Schrater, P. (2012). Brain plasticity through the life span: learning to learn and action video games. Annu. Rev. Neurosci. 35, 391–416.10.1146/annurev-neuro-060909-152832Search in Google Scholar PubMed
Beaulieu, C. and Colonnier, M. (1987). Effect of the richness of the environment on the cat visual cortex. J. Comp. Neurol. 266, 478–494.10.1002/cne.902660404Search in Google Scholar PubMed
Bourgeon, S., Xerri, C. and Coq, J. O. (2004). Abilities in tactile discrimination of textures in adult rats exposed to enriched or impoverished environments. Behav. Brain Res. 153, 217–231.10.1016/j.bbr.2003.12.002Search in Google Scholar PubMed
Burrows, E. L. and Hannan, A. J. (2013). Characterizing social behavior in genetically targeted mouse models of brain disorders. Methods Mol. Bio. 1017, 95–104.10.1007/978-1-62703-438-8_7Search in Google Scholar PubMed
Burrows, E. L., McOmish, C. E., Buret, L. S., Van den Buuse, M. and Hannan, A. J. (2015). Environmental enrichment ameliorates behavioral impairments modeling schizophrenia in mice lacking metabotropic glutamate receptor 5. Neuropsychopharmacol. 40, 1947–1956.10.1038/npp.2015.44Search in Google Scholar PubMed PubMed Central
Cai, R., Guo, F., Zhang, J., Xu, J., Cui, Y. and Sun, X. (2009). Environmental enrichment improves behavioral performance and auditory spatial representation of primary auditory cortical neurons in rat. Neurobiol. Learn. Mem. 91, 366–376.10.1016/j.nlm.2009.01.005Search in Google Scholar PubMed
Cai, R., Zhou, X., Guo, F., Xu, J., Zhang, J. and Sun, X. (2010). Maintenance of enriched environment induced changes of auditory spatial sensitivity and expression of GABAA, NMDA, and AMPA receptor subunits in rat auditory cortex. Neurobiol. Learn. Mem. 94, 452–460.10.1016/j.nlm.2010.08.008Search in Google Scholar PubMed
Cancedda, L., Putignano, E., Sale, A., Viegi, A., Berardi, N. and Maffei, L. (2004). Acceleration of visual system development by environmental enrichment. J. Neurosci. 24, 4840–4848.10.1523/JNEUROSCI.0845-04.2004Search in Google Scholar PubMed PubMed Central
Cang, J. H., Kalatsky, V. A., Löwel, S. and Stryker, M. P. (2005). Optical imaging of the intrinsic signal as a measure of cortical plasticity in the mouse. Vis. Neurosci. 22, 685–691.10.1017/S0952523805225178Search in Google Scholar PubMed PubMed Central
Carro, E., Nuñez, A., Busiguina, S. and Torres-Aleman, I. (2000). Circulating insulin-like growth factor I mediates effects of exercise on the brain. J. Neurosci. 20, 2926–2933.10.1523/JNEUROSCI.20-08-02926.2000Search in Google Scholar PubMed
Ciucci, F., Putignano, E., Baroncelli, L., Landi, S., Berardi, N. and Maffei, L. (2007). Insulin-like growth factor 1 (IGF-1) mediates the effects of enriched environment (EE) on visual cortical development. PLoS One 2, e475.10.1371/journal.pone.0000475Search in Google Scholar PubMed PubMed Central
Cooper, C., Moon, H. Y. and van Praag, H. (2017). On the run for hippocampal plasticity. Cold Spring Harb. Perspect. Med. 11. pii: a029736. doi: 10.1101/cshperspect.a029736. [Epub ahead of print].10.1101/cshperspect.a029736Search in Google Scholar PubMed PubMed Central
Coq, J. O. and Xerri, C. (1998). Environmental enrichment alters organizational features of the forepaw representation in the primary somatosensory cortex of adult rats. Exp. Brain Res. 121, 191–204.10.1007/s002210050452Search in Google Scholar PubMed
Di Garbo, A., Mainardi, M., Chillemi, S., Maffei, L. and Caleo, M. (2011). Environmental enrichment modulates cortico-cortical interactions in the mouse. PLoS One 6, e25285.10.1371/journal.pone.0025285Search in Google Scholar PubMed PubMed Central
Diamond, M. C., Krech, D. and Rosenzweig, M. R. (1964). The effects of an enriched environment on the histology of the rat cerebral cortex. J. Comp. Neurol. 123, 111–119.10.1002/cne.901230110Search in Google Scholar PubMed
Dinse, H. R. (2004). Sound case for enrichment. J. Neurophysiol. 92, 36–37.10.1152/jn.00213.2004Search in Google Scholar PubMed
Dinse, H. R. (2016). Bereichernde Umgebung – Tanzen im Alter aus Sicht der Neurowissenschaften. Kulturräume+: Kubia 10, 13–17.Search in Google Scholar
Dräger, U. C. (1975). Receptive fields of single cells and topography in mouse visual cortex. J. Comp. Neurol. 160, 269–289.10.1002/cne.901600302Search in Google Scholar PubMed
Dräger, U. C. (1978). Observations on monocular deprivation in mice. J. Neurophysiol. 41, 28–42.10.1152/jn.1978.41.1.28Search in Google Scholar PubMed
Driver, J. and Noesselt, T. (2008) Multisensory interplay reveals crossmodal influences on ‘sensory-specific’ brain regions, neural responses, and judgments. Neuron. 57, 11–23.10.1016/j.neuron.2007.12.013Search in Google Scholar PubMed
Engineer, N. D., Percaccio, C. R., Pandya, P. K., Moucha, R., Rathbun, D. L. and Kilgard, M. P. (2004). Environmental enrichment improves response strength, threshold, selectivity, and latency of auditory cortex neurons. J. Neurophysiol. 92, 73–82.10.1152/jn.00059.2004Search in Google Scholar PubMed
Erickson, K. I., Weinstein, A. M. and Lopez, O. L. (2012). Physical activity, brain plasticity, and Alzheimer‘s disease. Arch. Med. Res. 43, 615–621.10.1016/j.arcmed.2012.09.008Search in Google Scholar PubMed
Espinosa, J. S. and Stryker, M. P. (2012). Development and plasticity of the primary visual cortex. Neuron 75, 230–249.10.1016/j.neuron.2012.06.009Search in Google Scholar PubMed
Falkenberg, T., Mohammed, A. K., Henriksson, B., Persson, H., Winblad, B. and Lindefors, N. (1992). Increased expression of brain-derived neurotrophic factor mRNA in rat hippocampus is associated with improved spatial memory and enriched environment. Neurosci. Lett. 138, 153–156.10.1016/0304-3940(92)90494-RSearch in Google Scholar PubMed
Fischer, A., Sananbenesi, F., Wang, X., Dobbin, M. and Tsai, L.-H. (2007). Recovery of learning and memory is associated with chromatin remodelling. Nature 447, 178–182.10.1038/nature05772Search in Google Scholar PubMed
Fischer, A. (2016) Environmental enrichment as a method to improve cognitive function. What can we learn from animal models? Neuroimage 131, 42–7.10.1016/j.neuroimage.2015.11.039Search in Google Scholar PubMed
Florence, S. L., Boydston, L. A., Hackett, T. A., Lachoff, H. T., Strata, F. and Niblock, M. M. (2001). Sensory enrichment after peripheral nerve injury restores cortical, not thalamic, receptive field organization. Europ. J. Neurosci. 13, 1755–1766.10.1046/j.0953-816x.2001.01555.xSearch in Google Scholar
Fu, Y., Kaneko, M., Tang, Y., Alvarez-Buylla, A. and Stryker, M. P. (2015). A cortical disinhibitory circuit for enhancing adult plasticity. eLife 4, e05558.10.7554/eLife.05558Search in Google Scholar PubMed
Ghazanfar, A. A. and Schroeder, C. E. (2006) Is neocortex essentially multisensory? Trends Cogn. Sci. 10, 278–285.10.1016/j.tics.2006.04.008Search in Google Scholar
Godde, B., Berkefeld, T., David-Jürgens, M. and Dinse, H. R. (2002). Age-related changes in primary somatosensory cortex of rats: evidence for parallel degenerative and plastic-adaptive processes. Neurosci. Biobehav. Rev. 26, 743–752.10.1016/S0149-7634(02)00061-1Search in Google Scholar PubMed
Gordon, J. A. and Stryker, M. P. (1996). Experience-dependent plasticity of binocular responses in the primary visual cortex of the mouse. J. Neurosci. 16, 3274–3286.10.1523/JNEUROSCI.16-10-03274.1996Search in Google Scholar PubMed
Greifzu, F., Kalogeraki, E. and Löwel, S. (2016). Environmental enrichment preserved lifelong ocular dominance plasticity, but did not improve visual abilities. Neurobiol. Aging 41, 130–137.10.1016/j.neurobiolaging.2016.02.014Search in Google Scholar PubMed
Greifzu, F., Pielecka-Fortuna, J., Kalogeraki, E., Krempler, K., Favaro, P. D., Schlüter, O. M. and Löwel, S. (2014). Environmental enrichment extends ocular dominance plasticity into adulthood and protects from stroke-induced impairments of plasticity. Proc. Natl. Acad. Sci. U.S.A. 111, 1150–1155.10.1073/pnas.1313385111Search in Google Scholar PubMed PubMed Central
Greifzu, F., Schmidt, S., Schmidt, K. F., Kreikemeier, K., Witte, O. W. and Löwel, S. (2011). Global impairment and therapeutic restoration of visual plasticity mechanisms after a localized cortical stroke. Proc. Natl. Acad. Sci. U.S.A. 108, 15450–15455.10.1073/pnas.1016458108Search in Google Scholar PubMed PubMed Central
Greifzu, F., Wolf, F. and Löwel, S. (2012). Network influences on cortical plasticity. e-Neuroforum 3, 41–48.10.1007/s13295-012-0030-0Search in Google Scholar
Guzzetta, A., Baldini, S., Bancale, A., Baroncelli, L., Ciucci, F., Ghirri, P., Putignano, E., Sale, A., Viegi, A., Berardi, N., Boldrini, A., Cioni, G. and Maffei, L. (2009). Massage accelerates brain development and the maturation of visual function. J. Neurosci. 29, 6042–6051.10.1523/JNEUROSCI.5548-08.2009Search in Google Scholar PubMed PubMed Central
Han, C., Ding, D., Lopez, M. C., Manohar, S., Zhang, Y., Kim, M. J., Park, H. J., White, K., Kim, Y. H., Linser, P., Tanokura, M., Leeuwenburgh, C., Baker, H. V., Salvi, R. J. and Someya, S. (2016). Effects of long-term exercise on age-related hearing loss in mice. J. Neurosci. 36, 11308–11319.10.1523/JNEUROSCI.2493-16.2016Search in Google Scholar PubMed PubMed Central
Hannan, A. J. (2014). Environmental enrichment and brain repair: harnessing the therapeutic effects of cognitive stimulation and physical activity to enhance experience-dependent plasticity. Neuropathol. Appl. Neurobiol. 40, 13–25.10.1111/nan.12102Search in Google Scholar PubMed
Harauzov, A., Spolidoro, M., DiCristo, G., De Pasquale, R., Cancedda, L., Pizzorusso, T., Viegi, A., Berardi, N. and Maffei, L. (2010). Reducing intracortical inhibition in the adult visual cortex promotes ocular dominance plasticity. J. Neurosci. 30, 361–371.10.1523/JNEUROSCI.2233-09.2010Search in Google Scholar PubMed PubMed Central
Hebb, D. O. (1947). Spontaneous neurosis in chimpanzees; theoretical relations with clinical and experimental phenomena. Psychosom. Med. 9, 3–19.10.1097/00006842-194701000-00001Search in Google Scholar PubMed
Hebb, D. O. (1949). The organization of behavior. New York: Wiley & Sons.Search in Google Scholar
Hensch, T. K. (2005). Critical period plasticity in local cortical circuits. Nat. Rev. Neurosci. 6, 877–88.10.1038/nrn1787Search in Google Scholar PubMed
Henschke, J. U., Oelschlegel, A. M., Angenstein, F., Ohl, F. W., Goldschmidt, J., Kanold, P. O. and Budinger, E. (2017). Early sensory experience influences the development of multisensory thalamocortical and intracortical connections of primary sensory cortices. Brain Struct. Funct. Nov 1. doi: 10.1007/s00429-017-1549-1.10.1007/s00429-017-1549-1Search in Google Scholar PubMed PubMed Central
Hertzog, C., Kramer, A. F., Wilson, R. S. and Lindenberger, U. (2008). Enrichment effects on adult cognitive development: can the functional capacity of older adults be preserved and enhanced? Psychol. Sci. Pub. Interest. 9, 1–65.10.1111/j.1539-6053.2009.01034.xSearch in Google Scholar PubMed
Hotting, K. and Röder, B. (2013). Beneficial effects of physical exercise on neuroplasticity and cognition. Neurosci. Biobehav. Rev. 37, 2243–2257.10.1016/j.neubiorev.2013.04.005Search in Google Scholar PubMed
Ickes, B. R., Pham, T. M., Sanders, L. A., Albeck, D. S., Mohammed, A. H. and Granholm, A. C. (2000). Long-term environmental enrichment leads to regional increases in neurotrophin levels in rat brain. Exp. Neurol. 164, 45–52.10.1006/exnr.2000.7415Search in Google Scholar PubMed
Jakkamsetti, V., Chang, K. Q. and Kilgard, M. P. (2012). Reorganization in processing of spectral and temporal input in the rat posterior auditory field induced by environmental enrichment. J. Neurophysiol. 107, 1457–1475.10.1152/jn.01057.2010Search in Google Scholar PubMed PubMed Central
Jiang, C., Xu, X., Yu, L., Xu, J. and Zhang, J. (2015). Environmental enrichment rescues the degraded auditory temporal resolution of cortical neurons induced by early noise exposure. Europ. J. Neurosci. 42, 2144–2154.10.1111/ejn.12975Search in Google Scholar PubMed
Kalogeraki, E., Greifzu, F., Haack, F. and Löwel, S. (2014). Voluntary physical exercise promotes ocular dominance plasticity in adult mouse primary visual cortex. J. Neurosci. 34, 15476–15481.10.1523/JNEUROSCI.2678-14.2014Search in Google Scholar PubMed PubMed Central
Kalogeraki, E., Pielecka-Fortuna, J., Hüppe, J. M. and Löwel, S. (2016). Physical exercise preserves adult visual plasticity in mice and restores it after a stroke in the somatosensory cortex. Front. Aging Neurosci. 8, 212.10.3389/fnagi.2016.00212Search in Google Scholar PubMed PubMed Central
Kalogeraki, E., Pielecka-Fortuna, J. and Löwel, S. (2017). Environmental enrichment accelerates ocular dominance plasticity in mouse visual cortex whereas transfer to standard cages resulted in a rapid loss of increased plasticity. PloS One. 12, e0186999.10.1371/journal.pone.0186999Search in Google Scholar PubMed PubMed Central
Kaneko, M. and Stryker, M. P. (2014). Sensory experience during locomotion promotes recovery of function in adult visual cortex. eLife. 3, e02798.10.7554/eLife.02798Search in Google Scholar PubMed PubMed Central
Kattenstroth, J. C., Kalisch, T., Holt, S., Tegenthoff, M. and Dinse, H. R. (2013). Six months of dance intervention enhances postural, sensorimotor, and cognitive performance in elderly without affecting cardio-respiratory functions. Front. Aging Neurosci. 5, 5.10.3389/fnagi.2013.00005Search in Google Scholar PubMed PubMed Central
Kayser, C. and Logothetis, N. K. (2007). Do early sensory cortices integrate cross-modal information? Brain Struct. Funct. 212, 121–132.10.1007/s00429-007-0154-0Search in Google Scholar
Kilgard, M. P., Vazquez, J. L., Engineer, N. D. and Pandya, P. K. (2007). Experience dependent plasticity alters cortical synchronization. Hear Res. 229, 171–179.10.1016/j.heares.2007.01.005Search in Google Scholar PubMed PubMed Central
Kleemeyer, M. M., Polk, T. A., Schaefer, S., Bodammer, N. C., Brechtel, L. and Lindenberger, U. (2017). Exercise-induced fitness changes correlate with changes in neural specificity in older adults. Front. Hum. Neurosci. 11, 123.10.3389/fnhum.2017.00123Search in Google Scholar PubMed PubMed Central
Landers, M. S., Knott, G. W., Lipp, H. P., Poletaeva, I. and Welker, E. (2011). Synapse formation in adult barrel cortex following naturalistic environmental enrichment. Neuroscience 199, 143–152.10.1016/j.neuroscience.2011.10.040Search in Google Scholar PubMed
Lehmann, K. and Löwel, S. (2008). Age-dependent ocular dominance plasticity in adult mice. PLoS One 3, e3120.10.1371/journal.pone.0003120Search in Google Scholar PubMed PubMed Central
Levelt, C. N. and Hübener, M. (2012). Critical-period plasticity in the visual cortex. Ann. Rev. Neurosci. 35, 309–330.10.1146/annurev-neuro-061010-113813Search in Google Scholar PubMed
Li, R., Polat, U., Makous, W. and Bavelier, D. (2009). Enhancing the contrast sensitivity function through action video game training. Nat. Neurosci. 12, 549–551.10.1038/nn.2296Search in Google Scholar PubMed PubMed Central
Li, R. W., Ngo, C., Nguyen, J. and Levi, D. M. (2011). Video-game play induces plasticity in the visual system of adults with amblyopia. PLoS Biol. 9, 30.10.1371/journal.pbio.1001135Search in Google Scholar PubMed PubMed Central
Löwel, S. and Singer, W. (1992). Selection of intrinsic horizontal connections in the visual cortex by correlated neuronal activity. Science 255, 209–212.10.1126/science.1372754Search in Google Scholar PubMed
Mabunga, D. F., Gonzales, E. L., Kim, J. W., Kim, K. C. and Shin, C. Y. (2015). Exploring the validity of valproic acid animal model of autism. Exp. Neurobiol. 24, 285–300.10.5607/en.2015.24.4.285Search in Google Scholar PubMed PubMed Central
Mainardi, M., Di Garbo, A., Caleo, M., Berardi, N., Sale, A. and Maffei, L. (2014). Environmental enrichment strengthens corticocortical interactions and reduces amyloid-β oligomers in aged mice. Front. Aging Neurosci. 6, 1.10.3389/fnagi.2014.00001Search in Google Scholar PubMed PubMed Central
Markham, J. A. and Greenough, W. T. (2004). Experience-driven brain plasticity: beyond the synapse. Neuron Glia Biol. 1, 351–363.10.1017/S1740925X05000219Search in Google Scholar PubMed PubMed Central
Matthies, U., Balog, J. and Lehmann, K. (2013). Temporally coherent visual stimuli boost ocular dominance plasticity. J. Neurosci. 33, 11774–11778.10.1523/JNEUROSCI.4262-12.2013Search in Google Scholar PubMed PubMed Central
Maya-Vetencourt, J. F. and Origlia, N. (2012). Visual cortex plasticity: A complex interplay of genetic and environmental influences. Neural Plast. 2012, 631965. doi: 10.1155/2012/631965.10.1155/2012/631965Search in Google Scholar PubMed PubMed Central
Maya-Vetencourt, J. F., Sale, A., Viegi, A., Baroncelli, L., De Pasquale, R., O‘Leary, O. F., Castren, E. and Maffei, L. (2008). The antidepressant fluoxetine restores plasticity in the adult visual cortex. Science 320, 385–388.10.1126/science.1150516Search in Google Scholar PubMed
Mo, C., Hannan, A. J. and Renoir, T. (2015). Environmental factors as modulators of neurodegeneration: insights from gene-environment interactions in Huntington‘s disease. Neurosci. Biobehav. Rev. 52, 178–192.10.1016/j.neubiorev.2015.03.003Search in Google Scholar PubMed
Mohammed, A. H., Henriksson, B. G., Soderstrom, S., Ebendal, T., Olsson, T. and Seckl, J. R. (1993). Environmental influences on the central nervous system and their implications for the aging rat. Behav. Brain Res. 57, 183–191.10.1016/0166-4328(93)90134-CSearch in Google Scholar PubMed
Morishita, H., Miwa, J. M., Heintz, N. and Hensch, T. K. (2010). Lynx1, a cholinergic brake limits plasticity in adult visual cortex: (a cure for amblyopia through nicotinic receptor signaling). Science 330, 1238–1240.10.1126/science.1195320Search in Google Scholar
Pakan, J. M., Lowe, S. C., Dylda, E., Keemink, S. W., Currie, S. P., Coutts, C. A. and Rochefort, N. L. (2016). Behavioral-state modulation of inhibition is context-dependent and cell type specific in mouse visual cortex. Elife 5, e14985.10.7554/eLife.14985.013Search in Google Scholar PubMed
Percaccio, C. R., Engineer, N. D., Pruette, A. L., Pandya, P. K., Moucha, R., Rathbun, D. L. and Kilgard, M. P. (2005). Environmental enrichment increases paired-pulse depression in rat auditory cortex. J. Neurophysiol. 94, 3590–3600.10.1152/jn.00433.2005Search in Google Scholar PubMed
Percaccio, C. R., Pruette, A. L., Mistry, S. T., Chen, Y. H. and Kilgard, M. P. (2007). Sensory experience determines enrichment-induced plasticity in rat auditory cortex. Brain Res. 1174, 76–91.10.1016/j.brainres.2007.07.062Search in Google Scholar PubMed
Pham, T. M., Ickes, B., Albeck, D., Söderström, S., Granholm, A. C. and Mohammed, A. H. (1999). Changes in brain nerve growth factor levels and nerve growth factor receptors in rats exposed to environmental enrichment for one year. Neurosci. 94, 279–286.10.1016/S0306-4522(99)00316-4Search in Google Scholar
Pielecka-Fortuna, J., Kalogeraki, E., Greifzu, F. and Löwel, S. (2015). A small motor cortex lesion abolished ocular dominance plasticity in the adult mouse primary visual cortex and impaired experience-dependent visual improvements. PLoS One 10, e0137961.10.1371/journal.pone.0137961Search in Google Scholar PubMed
Polley, D. B., Kvasnak, E. and Frostig, R. D. (2004). Naturalistic experience transforms sensory maps in the adult cortex of caged animals. Nature 429, 67–71.10.1038/nature02469Search in Google Scholar PubMed
Rosenzweig, M. R., Bennett, E. L., Hebert, M. and Morimoto, H. (1978). Social grouping cannot account for cerebral effects of enriched environments. Brain Res. 153, 563–576.10.1016/0006-8993(78)90340-2Search in Google Scholar PubMed
Rosenzweig, M. R., Krech, D., Bennett, E. L. and Diamond, M. C. (1962). Effects of environmental complexity and training on brain chemistry and anatomy: a replication and extension. J. Comp. Physiol. Psychol. 55, 429–437.10.1037/h0041137Search in Google Scholar PubMed
Sale, A., Berardi, N. and Maffei, L. (2014). Environment and brain plasticity: towards an endogenous pharmacotherapy. Physiol. Rev. 94, 189–234.10.1152/physrev.00036.2012Search in Google Scholar PubMed
Sale, A., Maya-Vetencourt, J. F., Medinin, P., Cenni, M. C., Baroncelli, L., De Pasquale, R. and Maffei, L. (2007). Environmental enrichment in adulthood promotes amblyopia recovery through a reduction of intracortical inhibition. Nat. Neurosci. 10, 679–681.10.1038/nn1899Search in Google Scholar PubMed
Sale, A., Putignano, E., Cancedda, L., Landi, S., Cirulli, F., Berardi, N. and Maffei, L. (2004). Enriched environment and acceleration of visual system development. Neuropharmacol. 47, 649–660.10.1016/j.neuropharm.2004.07.008Search in Google Scholar
Santos-Lozano, A., Pareja-Galeano, H., Sanchis-Gomar, F., Quindos-Rubial, M., Fiuza-Luces, C., Cristi-Montero, C., Emanuele, E., Garatachea, N. and Lucia, A. (2016). Physical Activity and Alzheimer Disease: A Protective Association. Mayo Clin. Proc. 91, 999–1020.10.1016/j.mayocp.2016.04.024Search in Google Scholar PubMed
Sato, M. and Stryker, M. P. (2008). Distinctive features of adult ocular dominance plasticity. J. Neurosci. 28, 10278–10286.10.1523/JNEUROSCI.2451-08.2008Search in Google Scholar PubMed
Sawtell, N. B., Frenkel, M. Y., Philpot, B. D., Nakazawa, K., Tonegawa, S. and Bear, M. F. (2003). NMDA receptor-dependent ocular dominance plasticity in adult visual cortex. Neuron 38, 977–985.10.1016/S0896-6273(03)00323-4Search in Google Scholar PubMed
Schubert, C. R., Fischer, M. E., Pinto, A. A., Klein, B. E. K., Klein, R. and Cruickshanks, K. J. (2017). Odor detection thresholds in a population of older adults. Laryngoscope 127, 1257–1262.10.1002/lary.26457Search in Google Scholar PubMed PubMed Central
Stryker, M. P. (2014). A neural circuit that controls cortical state, plasticity, and the gain of sensory responses in mouse. Cold Spring Harb. Symp. Quant. Biol. 79, 1–9.10.1101/sqb.2014.79.024927Search in Google Scholar PubMed PubMed Central
Stryker, M. P. and Löwel, S. (2017). Amblyopia: New molecular/pharmacological and environmental approaches. Chapter 7. In: Lasker-Foundation Report on Amblyopia 2017, www.laskerfoundation.org/new-noteworthy/articles/amblyopia-challenges.10.1017/S0952523817000256Search in Google Scholar
Sturm, J. J., Zhang-Hooks, Y.-X., Roos, H., Nguyen, T. and Kandler, K. (2017). Noise trauma-induced behavioral gap detection deficits correlate with reorganization of excitatory and inhibitory local circuits in the inferior colliculus and are prevented by acoustic enrichment. J. Neurosci. 37, 6314–6330.10.1523/JNEUROSCI.0602-17.2017Search in Google Scholar PubMed PubMed Central
Tkacs, N. C. and Thompson, H. J. (2006). From bedside to bench and back again: research issues in animal models of human disease. Biol. Res. Nurs. 8, 78–88.10.1177/1099800406289717Search in Google Scholar PubMed PubMed Central
van Praag, H., Kempermann, G. and Gage, F. H. (2000). Neural consequences of environmental enrichment. Nat. Rev. Neurosci. 1, 191–198.10.1038/35044558Search in Google Scholar PubMed
Vivar, C., Potter, M. C. and van Praag, H. (2013). All about running: synaptic plasticity, growth factors and adult hippocampal neurogenesis. Curr. Top. Behav. Neurosci. 15, 189–210.10.1007/7854_2012_220Search in Google Scholar PubMed PubMed Central
Voss, M. W., Vivar, C., Kramer, A. F. and van Praag, H. (2013). Bridging animal and human models of exercise-induced brain plasticity. Trends Cogn. Sci. 17, 525–544.10.1016/j.tics.2013.08.001Search in Google Scholar PubMed PubMed Central
Wiesel, T. N. and Hubel, D. H. (1963). Effects of visual deprivation on morphology and physiology of cells in the cat‘s lateral geniculate body. J. Neurophysiol. 26, 978–993.10.1152/jn.1963.26.6.978Search in Google Scholar PubMed
Zhu, X., Wang, F., Hu, H., Sun, X., Kilgard, M. P., Merzenich, M. M. and Zhou, X. (2014). Environmental acoustic enrichment promotes recovery from developmentally degraded auditory cortical processing. J. Neurosci. 34, 5406–5415.10.1523/JNEUROSCI.5310-13.2014Search in Google Scholar PubMed PubMed Central
Anmerkung
Übersetzung der englischen Version des Artikels online verfügbar unter https://doi.org/10.1515/nf-2017-A050
© 2017 by De Gruyter
Articles in the same Issue
- Frontmatter
- Übersichtsartikel
- Selektive Degeneration dopaminerger Neurone beim Parkinson-Syndrom: die zunehmende Rolle von veränderter Kalziumhomöostase und nukleolärer Funktion
- Selective degeneration of dopamine neurons in Parkinson’s disease: emerging roles of altered calcium homeostasis and nucleolar function
- Altruismus aus Sicht der Sozialen Neurowissenschaften
- Altruism from the Perspective of the Social Neurosciences
- Lebensbedingungen haben einen starken Einfluss auf die Plastizität des Gehirns
- Environmental conditions strongly affect brain plasticity
- Die neuronalen Signale, die Wahrnehmung verändern
- The neural events that change perception
- Der Einfluss von Fortbewegung auf die sensorische Informationsverarbeitung und die zugrunde liegenden neuronalen Schaltkreise
- The influence of locomotion on sensory processing and its underlying neuronal circuits
- Forschungsförderung
- Sonderforschungsbereich (SFB/TRR 167) NeuroMac „Entwicklung, Funktion und Potenzial von myeloischen Zellen im zentralen Nervensystem“
- Rezension
- Michael Madeja/Joachim Müller-Jung (Hrsg.): Hirnforschung – was kann sie wirklich
- Nachrichten
- Neue NWG-Website
Articles in the same Issue
- Frontmatter
- Übersichtsartikel
- Selektive Degeneration dopaminerger Neurone beim Parkinson-Syndrom: die zunehmende Rolle von veränderter Kalziumhomöostase und nukleolärer Funktion
- Selective degeneration of dopamine neurons in Parkinson’s disease: emerging roles of altered calcium homeostasis and nucleolar function
- Altruismus aus Sicht der Sozialen Neurowissenschaften
- Altruism from the Perspective of the Social Neurosciences
- Lebensbedingungen haben einen starken Einfluss auf die Plastizität des Gehirns
- Environmental conditions strongly affect brain plasticity
- Die neuronalen Signale, die Wahrnehmung verändern
- The neural events that change perception
- Der Einfluss von Fortbewegung auf die sensorische Informationsverarbeitung und die zugrunde liegenden neuronalen Schaltkreise
- The influence of locomotion on sensory processing and its underlying neuronal circuits
- Forschungsförderung
- Sonderforschungsbereich (SFB/TRR 167) NeuroMac „Entwicklung, Funktion und Potenzial von myeloischen Zellen im zentralen Nervensystem“
- Rezension
- Michael Madeja/Joachim Müller-Jung (Hrsg.): Hirnforschung – was kann sie wirklich
- Nachrichten
- Neue NWG-Website