Heat shock and titanium dioxide nanoparticles decrease superoxide dismutase and glutathione enzymes activities in Saccharomyces cerevisiae
-
Joana Capela-Pires
Joana Capela-Pires graduated in Biochemistry from the University of Évora, Évora, Portugal (2009). From the same university, she obtained a MSc in Biochemistry with a thesis on the antioxidant response of
Saccharomyces cerevisiae to titanium dioxide nanoparticles (2011). Since 2011, she has been a PhD student at the Department of Chemistry and Institute of Mediterranean Agrarian and Environmental Sciences, University of Évora, Évora, Portugal. Her field of research is: stress and metabolic regulation onS. cerevisiae cells by titanium dioxide nanoparticles; her field of specialization is: energetic and antioxidant systems of yeast cells.Rui Ferreira graduated in Biochemistry (1987) from the University of Lisbon, Portugal. He obtained a PhD (1999) in Biochemistry from the University of Évora, Portugal. He was an Assistant Professor (1999–2004) and has been an Associate Professor of Enzymology since 2004 at the Department of Chemistry, University of Évora. Since 1988, he has also been a researcher at ICAAM, University of Évora. His fields of research are stress and metabolic regulation by metals and pesticides. He specializes in energetics, biotransformation and antioxidant systems of animals and yeast.
 und
Isabel Alves-Pereira
und
Isabel Alves-Pereira
Isabel Alves-Pereira graduated as an agronomic engineer from the University of Évora, Portugal. She obtained an MSc in Microbial Molecular Genetics from the University of Minho, Portugal and a PhD in Biochemistry (2008) from the University of Extremadura, Spain. Since 2008, she has been an Assistant Professor of Biochemistry at the Department of Chemistry, University of Évora. Since 1994, she has also been a researcher at ICAAM, University of Évora. Her fields of research are the effects of metals and pesticides on antioxidant and biotransformation systems.


Abstract
The exposure of living organisms to metals can generate reactive oxygen species and failure in their antioxidant defences, triggering oxidative stress and oxidative damage. Despite the intensive use of engineered nanoparticles in numerous consumer and industrial products, data on their potential hazards in eukaryotic cells and their dependence on environmental factors such as temperature are still scarce. The aim of this study was to evaluate the antioxidant response of Saccharomyces cerevisiae, grown in presence of glycerol and glucose, to 5 μg/ml titanium dioxide nanoparticles (size<100 nm) under heat shock conditions. The results showed that biomass, levels of reactive oxygen species and glutathione reductase activity in respiratory/fermentative cells were higher than those detected in respiratory cells. Furthermore, respiratory/fermentative cells exhibited lower levels of glutathione, malondialdehyde, cytoplasmic catalase and glutathione peroxidase than those detected in the respiratory yeast. Saccharomyces cerevisiae grown in the presence of glycerol, glucose and titanium dioxide nanoparticles, under heat shock conditions, caused oxidative stress, due to a decrease in antioxidant defences such as superoxide dismutases or a slowdown of the glutathione cycle, relative to cells grown in presence of glycerol and glucose.
1 Introduction
The ubiquitous occurrence of nanoparticles (NPs) in the earth’s crust, as well as their geological origin suggests that living organisms have developed adaptive mechanisms to these materials throughout their evolution. However, several studies have reported that nano-sized particles, produced on a large scale for industrial and domestic use showed more serious toxicity than their bulk form, being involved in the inflammatory response and cytotoxic effects in mammals [1–5]. Thus, the mass production of nanomaterials has created new types of environmental contaminants whose interaction with living organisms is unclear. The size of NPs correlates directly with their properties, such as large specific surface area, solubility and chemical reactivity that may determine the biological behaviour of NPs in vivo [6]. In contrast to animal cells, plants and yeast possess cell walls that constitute a primary site for interaction and a barrier to the entry of NPs to their cells. Cell walls in yeast, mainly consisting of chitin, are semipermeable, allowing the passage of small molecules while limiting the passage of larger molecules. The diameter of pores across the cell wall, which has a thickness ranging from 5 to 20 nm, determines their sifting properties [7–10]. Thus, only NPs with a size smaller than that of the largest pore are able to pass through the cell wall and reach the plasma membrane. Despite the internalisation of NPs into yeast cells having not yet been studied, some authors have reported that Saccharomyces cerevisiae exhibits greater resistance to TiO2-NP than Gram-positive bacteria or animal cells, since the structural rigidity of the cell wall should prevent the direct uptake of NPs [5]. Moreover, interactions of cells with NPs may induce the formation of new pores, which may be larger than usual and thus increase the internalisation of the NPs through the cell wall. Once the NPs encounter the cell membrane, possible forms of interaction include specific or nonspecific forces, receptor ligand binding interactions or membrane wrapping of NPs [11]. During endocytic processes, the plasma membrane forms a cavity-like structure that surrounds the NPs and pulls it into the cell. NPs may also cross cell membranes using embedded transport carrier proteins or ion channels. Throughout this process, NPs may have a direct damaging effect on membrane composition, disrupting membrane composition and fluidity by means of a mechanism for adaptation to NPs which probably involves the regulation of specific desaturase activity [12, 13]. When inside cells, NPs may bind with different types of organelles (e.g., the endoplasmic reticulum or the Golgi apparatus) and interfere with their metabolic processes, possibly as a result of the production of reactive oxygen species (ROS) [13–15]. Furthermore, release of toxic ions mainly caused by physical restraints are among the indirect effects also assigned to metal NPs [14]. Some examples of NP-mediated effects in organisms include oxidative stress, genetic damage, inhibition of cell division and cell death [16, 17]. Most work to date has suggested that ROS generation (which can be either protective or harmful) during biological interactions and consequent oxidative stress are frequently observed with NP toxicity. Exposures to metal oxide NP of Ti, Cu and Fe were reported to induce oxidative stress via lipid peroxidation in experimental models such as A431, HEp-2 and MRC-5 cells [18–20]. In order to overcome excess of ROS, cells can activate enzymatic and nonenzymatic antioxidant systems [21, 22]. For example, superoxide dismutases (SOD1, SOD2) catalyse the dismutation of the superoxide radical (·O2-) into hydrogen peroxide (H2O2); catalases (CTT1, CTA1) detoxify hydrogen peroxide into water, and glutathione peroxidases (GPx) metabolise lipid hydroperoxides [23] in corresponding alcohol using glutathione (GSH) as a reducing agent. The resulting glutathione disulphide then regenerates to GSH by means of glutathione reductase (GR) [24] using reducing equivalents such as NADPH produced by means of glucose-6-phosphate dehydrogenase (G6PD), a key enzyme of the pentose phosphate pathway [25]. Thus, the healthy functioning of living cells is correlated with a set of events that maximise the GSH/GSSG pair, ensuring a stable and strongly reducing environment. Therefore, changes in this parameter are often used as an indicator of oxidative stress, which may trigger phenomena such as cell growth and differentiation, activation and apoptosis signalling pathways [26]. Heat stress affects a broad range of cellular processes, including transitory arrest of cell division [27–32], uncoupling of oxidative phosphorylation [33], damage to membranes and cytoskeletal structures and defective trafficking through the secretory pathway [34]. Heat stress is also associated with pronounced production of ROS [34–37]. Studies in Saccharomyces cerevisiae showed a transcription induction of a set of genes after a sudden shift in temperature from 30 to 50° [38] and a decrease in the rate of synthesis of some proteins produced at significant levels at normal growth temperatures [39]. After heat shock, cells focus on the synthesis of heat shock proteins as chaperones for assisting protein folding and disaggregation [40] cell-wall remodelling [41], metabolic enzymes, regulatory proteins, and others involved in their preservation. More than a thousand genes were identified in response to heat shock, after a rapid shift in temperature from 30 to 50°C [42]. In addition, there are a large number of yeast genes whose expression is higher in respirable (YEPG) as compared with fermentable (YEPD) media [43]. Genes include those involved in glycerol utilization, the citrate cycle, the electron transport chain, oxidative phosphorylation and stress resistance [43–46]. However, no studies associating heat shock with NP toxicity are described in the literature. Considering that S. cerevisiae is a simple model for the study of mechanisms of oxidative stress, studies on ROS and heat shock mediated toxicity of NPs on yeast may provide new scientific knowledge on NPs toxicity that could be transferable to more complex eukaryotic cells [5]. Although some authors consider that TiO2-NP are not toxic to S. cerevisiae as Mn2O3-NPs or Fe0-NPs [47], recent metabolic studies in S. cerevisiae showed that TiO2-NP increased the flux of the whole glycolysis pathway [48] and are cytotoxic for yeast cells [49]. The wild-type S. cerevisiae UE-ME3, used in this study, exhibits physiological characteristics that enable their survival in hostile environments as the musts of Alentejo wines where the following conditions are detected: high levels of carbohydrates (140–160 g/l) and alcohol (>15%, v/v), acidic pH (3.0–3.5) and a low content of nitrogen, fat and vitamins [50]. Despite the intensive use of engineered NPs in various consumer and industrial products, data on their potential hazards are still scarce and mechanisms of action only partially understood [51]. Consequently, the aim of this study was to evaluate the antioxidant response of S. cerevisiae, grown in presence of glycerol and glucose (a respirable/fermentable medium), to 5 μg/ml TiO2-NP (size<100 nm) in heat shock conditions.
2 Materials and methods
2.1 Preparation of TiO2 nanoparticle suspensions
TiO2-NP (size<100 nm, from Sigma-Aldrich, St. Louis, MO, USA) stock suspensions were prepared by sonication in membrane filtered (0.45 μm) deionized (DI) water (resistivity 18.2 MX cm-1), using a 130-watt ultrasonic processor for 30 min [52–54]. All other reagents were also purchased from Sigma Chemicals Co.
2.2 Microorganisms and growth conditions
Saccharomyces cerevisiae UE-ME3, a strain isolated during the process of making Regional Alentejo Wine (Portugal), which is housed at the Enology Laboratory of the University of Évora, provided the eukaryotic model used in this study [54]. Bioassays were performed in 250-ml Erlenmeyer flasks containing 100 ml of YEPG medium (1% w/v yeast extract, 2% w/v peptone and 3% w/v glycerol). Culture flasks were inoculated using a fresh culture of wild-type S. cerevisiae UE-ME3 and were shaken 150 rpm, at 28°C in a shaking water bath with controlled temperature Memmert, model 3200R. In the exponential growth phase (OD=0.8), 2% (w/v) glucose was added (YEPGD medium). After 100 min, TiO2-NP stock solution was added to obtain a final concentration of 5 μg/ml. Yeast was allowed to grow for 100 min at 28°C or 40°C (heat shock, ST). Flasks lacking glucose (YEPG) or NPs (YEPGD) with or without heat shock, served as control samples.
2.3 Determination of dry weight
At the end of the experiment, the amount of biomass was determined by the measurement of dry weight in a micro analytical balance mettler Toledo, model AX 205. The collected aliquots were dried in an oven Memmert, model BE400 at 80°C until constant weight was attained.
2.4 Preparation of post-12,000 g supernatant
In order to obtain cellular fractions, at the end of the TiO2-NP exposure test for 200 min at controlled temperature, the cells were harvested by centrifugation at 5000 g for 10 min, rinsed in sterile double distilled water, resuspended in 10 mm phosphate buffer pH 7.0 and disintegrated in the ultrasonic homogeniser Branson sonifier 450 (10 s 3×5 min, 0.1 kHz pulse). The homogenates were subjected to differential centrifugation at 12,000 g for 20 min and the supernatant was collected and the pellet resuspended in 10 mm phosphate buffer pH 7.0. The cell fractions obtained, using a refrigerated super centrifuge Hermle Z323 K, were divided into aliquots and stored at -20°C for the later determination of cell contents and enzyme activities [55].
2.5 Determination of protein concentration
Protein concentration was determined according to Lowry [56], in a double-beam spectrophotometer Hitachi-U2001 using bovine serum albumin as a standard.
2.6 Determination of GSH and GSSG
Glutathione and glutathione disulphide were assayed as previously described [57]. This method was based on the reaction of o-phthalaldehyde (OPT) as a fluorescent reagent with GSH at pH 8.0 and GSSG at pH 12.0. GSH was complexed to 0.04 mN-ethylmaleimide (NEM) to prevent interference of GSH with the measurement of GSSG. Fluorescence of products was determined at 420 nm with the excitation at 350 nm, at 25°C, using GSH or GSSG as a standard in a single-beam Shimadzu RF-5001PC spectrofluorophotometer.
2.7 Measurements of ROS
ROS level was determined according to LeBel [58] in a single-beam Shimadzu RF-5001PC spectrofluorophotometer, using hydrogen peroxide as a standard, since it reacts with 2’,7’-dichlorofluorescein (DCFH) to rapidly generate DCF, which was measured by means of fluorescence using an excitation wavelength of 488 nm and an emission wavelength of 525 nm at 25°C.
2.8 Measurement of MDA
Malondialdehyde (MDA) was assessed as an index of lipid peroxidation, based on the quantification of TBA oxidation products, which was measured by means of fluorescence in a single-beam Shimadzu RF-5001PC spectrofluorophotometer using an excitation wavelength of 515 nm and an emission wavelength of 553 nm at 25°C [59], using as a standard the MDA generated from 1,1,3,3-tetramethoxypropane by means of hydrolysis in an acid medium [60].
2.9 Enzyme assays
GR activity was determined using a reaction mixture containing 0.5 mm EDTA, 2.2 mm GSSG and a suitable concentration of post-12,000 g supernatant in 100 mm phosphate buffer pH 7.2, incubated at 37°C for 300 s. This reaction was followed by reading the absorbance at 340 nm for 180 s, after the addition of 0.17 mm NADPH [61].
GPx activity was determined in a reaction mixture containing 5.0 mm GSH, 0.24 U/ml GR, 5.0 mm EDTA, 1.60 mm NADPH and a suitable concentration of post-12,000 g supernatant in 100 mm phosphate buffer pH 7.0, incubated at 37°C for 5 min. The reaction was initiated by the addition of 1.0 mm t-butyl hydroperoxide, and NADPH oxidation was monitored for 10 min at 37°C, by reading the decrease in absorbance at 340 nm [62].
G6PD activity was determined according to Postma [63] in a reaction mixture containing 400 μm NADP+, 5 mm MgCl2.6H2O, and a suitable concentration of post-12,000 g supernatant in 50 mm Tris-HCl, pH 8. The reaction was initiated by the addition of 5 mm glucose-6-P and NADPH formation was monitored by reading the increase of absorbance at 340 nm for 180 s. The coefficient of molar absorptivity for NADPH of 6.22 mm-1 cm-1 was used to calculate GR, GPx and G6PD activity.
Catalase activity was determined in a reaction mixture consisting of 30 mm H2O2 and a suitable concentration of post-12,000 g supernatant (CTT1) or pellet (CTA1) in 50 mm phosphate buffer pH 7.5, by measuring the decrease in absorbance at 240 nm due to H2O2 consumption, according to Beers and Sizer [64].
SOD assay using a xanthine oxidase (XO)/nitroblue tetrazolium salt (NBT) system was determined in a reaction mixture consisting of 0.1 mm xanthine, 0.1 mm EDTA, 0.025 mm NBT, 0.5% (w/v) BSA, and a suitable concentration of post-12,000 g supernatant (SOD1, Cu-ZnSOD) or pellet (SOD2, MnSOD) in 50 mm sodium phosphate buffer pH 7.8. The reaction was initiated by the addition of 7 mU XO and the absorbance change at 560 nm was monitored at 25°C for 20 min. One unit of SOD was defined as the amount of protein that inhibits NBT reduction to 50% of the maximum (units: U/mg of protein) [65]. All enzyme activities are determined in a double-beam spectrophotometer Hitachi-U2001.
2.10 Statistics
All values were presented as the mean of five independent experiments±the standard error of the mean (SEM). The normality and homogeneity of variance were assessed by means of a P-plot and Levene test, respectively. The statistical analysis of the results was performed using ANOVA I and the Duncan test to determine significant differences (p<0.01) between treatments, using SPSS® statistical software, version 22.0 (SPSS Inc., Chicago, IL, USA) for Windows®, licenced to the University of Évora.
3 Results and discussion
The results showed that the biomass produced by S. cerevisiae UE-ME3 grown in the YEPG medium (respirable) was lower than determined in the cells grown in the YEPGD medium (respirable/fermentable) (Figure 1A) (p<0.01). This difference in biomass production is consistent with what has been observed in industrial wine yeast, in which there is a rapid rate of growth in rich medium with glucose (YEPD) [43]. Cells grown in YEPGD medium (respirable/fermentable) and 5 μg/ml TiO2-NP with heat shock or no heat shock presented biomass levels approximate to those determined in yeast cells grown in the YEPG medium (respirable) (p<0.01). However, the biomass produced by cells grown in YEPGD medium (respirable/fermentable) under heat shock conditions was significantly lower than the biomass determined in S. cerevisiae grown in the YEPG medium (respirable), but significantly higher than that detected under the other experimental conditions. Thus, TiO2-NP exposure may have countered the effect of glucose or heat shock on the biomass produced by S. cerevisiae grown in the YEPGD medium (respirable/fermentable). The content in ROS from S. cerevisiae UE-ME3 grown in the YEPG medium was lower than that determined in the cells grown in the YEPGD medium (Figure 1B) (p<0.01), a result that appears to counteract that which is described in the literature, in which the content of ROS is higher in respiratory cells [66]. This response may be due to the partial suppression of antioxidant defence pathways by glucose when they are still likely producing high levels of ROS via respiratory metabolism [43, 67]. Although the yeast cells grown in the YEPGD medium (respirable/fermentable) with NPs presented ROS levels which were higher than those found in yeast grown in the YEPG medium (respirable) with heat shock (control sample) or without heat shock, the combined exposure of 5 μg/ml TiO2-NP with heat shock significantly increased the ROS content of S. cerevisiae grown in the presence of glycerol and glucose (respirable/fermentable) (Figure 1B) (p<0.01). This response may explain in part the loss of biomass described in Figure 1A.
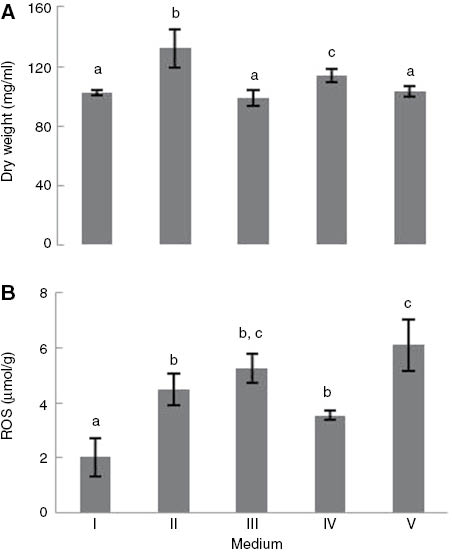
(A) Wet weight and (B) ROS content of Saccharomyces cerevisiae UE-ME3 grown in I- YEPG medium, II- YEPGD medium, III- YEPGD medium and 5 μg/ml TiO2-NP add at 100 min, IV-YEPGD medium with heat shock (28/40°C) at 100 min,V-YEPGD medium and 5 μg/ml TiO2-NP with heat shock (28/40°C) at 100 min. The results represent the mean of five independent experiments±standard error of mean. The bars marked with different letters (a-c) are significantly different, in accordance with the Duncan test (p<0.01).
The content of glutathione (GSH) and total glutathione (GSH+GGSG) of S. cerevisiae UE-ME3 grown in the YEPG medium (respirable) was higher than that measured in yeast cells grown in the YEPGD medium (respirable/fermentable) (Table 1) (p<0.01), results which suggest the repression of de novo biosynthesis of the antioxidant tripeptide when respiratory cells change their energetic metabolism using glucose as a carbon source [43]. Cells grown in the YEPGD medium (respirable/fermentable) with 5 μg/ml TiO2-NP contained levels of glutathione (GSH) and total glutathione (GSH+GSSG) lower than those detected in S. cerevisiae grown in the YEPG medium (respirable) (p<0.01). In addition, cells grown in the YEPGD medium (respirable/fermentable) under heat-shock conditions presented levels of GSH and GSH+GSSG which were lower than S. cerevisiae, which were only grown in YEPG (respirable) or YEPGD media (respirable/fermentable). In addition, the exposure of S. cerevisiae to 5 μg/ml TiO2-NP with heat shock accentuated the decrease in GSH levels to values below those determined in all other treatments, a response which may be due not only to the decrease in de novo biosynthesis but also to a decrease in cell ability to regenerate glutathione from glutathione disulfide, as suggested by increased levels of glutathione disulphide [24]. Yeast cells grown in YEPG (respirable) and YEPGD (respirable/fermentable) media showed no significant differences in intracellular redox environment estimated by the GSH/GSSG ratio (p<0.01). Yeast cells grown in the YEPG (respirable) and YEPGD (respirable/fermentable) media showed no significant differences in intracellular redox environment as estimated by the GSH/GSSG ratio (p<0.01), neither did the presence of 5 μg/ml TiO2-NP in the culture medium affect the redox GSH/GSSG pair (p<0.01), attaining, in all cases, values approximate to those presented in the literature for S. cerevisiae [25, 68, 69]. However, yeast cells grown in the YEPGD (respirable/fermentable) medium under heat shock conditions showed a significant decrease in the GSH/GSSG ratio, an effect that provided evidence for the occurrence of oxidative stress. This response was accentuated by the presence of 5 μg/ml TiO2-NP, a decrease in the GSH/GSSG ratio from 1.39 to 0.93 occurring.
GSH, GSSG, GSH+GSSG (calculated as [GSH]+2[GSSG]) contents and GSH/GSSG ratio of S.cerevisiae UE-ME3 grown in I-YEPG medium, II-YEPGD medium, III-YEPGD medium and 5 μg/ml TiO2-NP add at 100 min, IV-YEPGD medium with heat-shock (28/40°C) at 100 min,V-YEPGD medium and 5 μg/ml TiO2-NP with heat shock (28/40°C) at 100 min.
| Medium | GSH (nmol/gD.Wt) | GSSG (nmol/gD.Wt) | GSH+GSSG (nmol/gD.Wt) | GSH/GSSG |
|---|---|---|---|---|
| I | 131.01±20.19a | 44.85±9.03a | 220.71±38.11a | 3.13 0.25a |
| II | 78.42±9.73b | 26.04±6.29a | 139.16±17.99b | 2.36±0.91a |
| III | 86.43±4.29b | 33.97±6.73a | 166.00±9.98b | 2.34±0.33a |
| IV | 24.76±1.21c | 17.88±1.86b | 60.52±2.69c | 1.39±0.05b |
| V | 41.31±5.51d | 35.50±6.37a | 97.37±15.89d | 0.93±0.13c |
The results represent the mean of five independent experiments±standard error of mean. Means within a column with no common superscript letters (a–d) are significantly different, in accordance with the Duncan test (p<0.01).
The glutathione reductase activity of S. cerevisiae UE-ME3 grown in the YEPG medium (respirable) was below the level of this enzyme activity detected in cells grown in the YEPGD medium (respirable/fermentable) (Figure 2A) (p<0.01). In addition, the G6PD activity of cells grown in the YEPG medium (respirable) and the YEPGD medium (respirable/fermentable) showed no significant differences (Figure 2B). Thus, the regenerative capacity of GR for transformation GSSG into GSH seems to be enhanced by the presence of glucose, since the formation of reducing equivalents in the form of NADPH [25] required for the reduction of GSSG by the GR enzyme [24] were unaffected by the composition in carbon sources of the culture medium (p<0.01). However, the GPx activity of the yeast cells grown in the YEPGD medium (respirable/fermentable) was below the level of this enzyme activity detected in cells grown in the YEPG (respirable) without glucose as a carbon source was available (p<0.01). Therefore, the presence of glucose may partially suppress the antioxidant defence mechanism mediated by GPx against hydrogen peroxide and lipid hydroperoxides (Figure 2C), although the energetic mitochondrial metabolism may still have continued to generate high levels of ROS [67]. The GR activity of S. cerevisiae grown in the YEPGD medium (respirable/fermentable) with 5 μg/ml TiO2-NP was below the level of this enzyme activity detected in cells grown only in the YEPGD medium (respirable/fermentable), presenting values similar to those that was detected in yeast grown in the absence of glucose (YEPG medium, respirable) (p<0.01). However, the exposure of yeast cells to 5 μg/ml TiO2-NP, in the YEPGD medium did not disturb their G6PD and GPx enzyme activities (p<0.01). Furthermore, heat shock conditions did not affect the glutathione regeneration ability of S. cerevisiae grown in the YEPGD medium (respirable/fermentable), since there were no significant changes in GR enzyme activity. However, there was a significant decrease in antioxidant capacity mediated by GPx activity (Figure 2A–C) (p<0.01). Interestingly, the exposure of S. cerevisiae to TiO2-NP with heat shock caused a significant decrease in GR and G6PD enzyme activity, and GPx activity remained at similar levels to those detected in cells exposed to heat shock only (Figure 2A–C) (p<0.01), thereby contributing to a slowdown in the glutathione cycle. This response to TiO2-NP with heat shock by S. cerevisiae UE-ME3 seems have contributed to the increase in the intracellular ROS level and the consequent change in the redox environment (GSH/GSSG) [24, 25] discussed above.
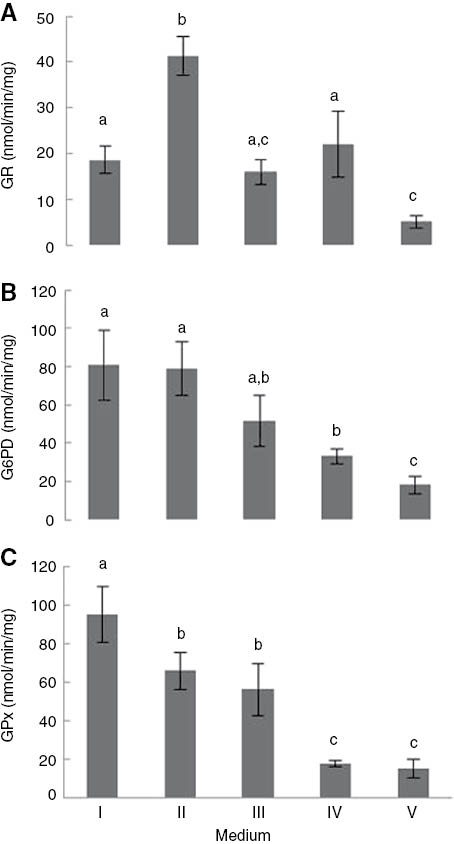
(A) GR, (B) G6PD, and (C) GPx activity of Saccharomyces cerevisiae UE-ME3 grown in I- YEPG medium, II- YEPGD medium, III- YEPGD medium and 5 μg/ml TiO2-NP add at 100 min, IV-YEPGD medium with heat shock (28/40°C) at 100 min,V-YEPGD medium and 5 μg/ml TiO2-NP with heat shock (28/40°C) at 100 min. The results represent the mean of five independent experiments±standard deviation. The bars marked with different letters (a–c) are significantly different, in accordance with the Duncan test (p<0.01).
Saccharomyces cerevisiae grown in the YEPG medium (respirable) showed cytoplasmic catalase activity levels (CTT1) higher than those detected in the yeast cells grown in YEPGD medium (respirable/fermentable) (Figure 3A) (p<0.01). This response to the presence of glucose in the culture medium was found to be consistent with the metabolic repression described by [43]. However, the antioxidant enzyme activities mediated by CTA1, SOD1 of yeast cells grown in the absence (YEPG) and presence of glucose (YEPGD) showed no significant differences (Figure 3B, C). The composition of the YEPGD medium (respirable/fermentable) or the exposure time was probably not enough to change the profile of such activities in the exclusively fermentative mode. Interestingly, the SOD2 activity levels of S. cerevisiae grown in the YEPGD medium (respirable/fermentable) were significantly higher than those detected in cells grown in the YEPG medium, an intriguing result (p<0.01). The cells grown in the YEPGD medium (respirable/fermentable) in the presence of NPs showed levels of CTT1 activity below those detected in the yeast grown in the YEPG and YEPGD media (Figure 3A) (p<0.01). However, there were no significant differences in the CTA1, SOD1 and SOD2 enzyme activities determined in cells grown under these three experimental conditions (Figure 3B–D). The maintenance of CTA1, SOD1 and SOD2 activity levels in the presence of 5 μg/ml TiO2-NP may have contributed to preventing the increase in intracellular ROS and the establishment of oxidative stress conditions, as mentioned above, although the NPs demonstrated the ability to reduce the antioxidant defences mediated by cytoplasmic catalase (CTT1).
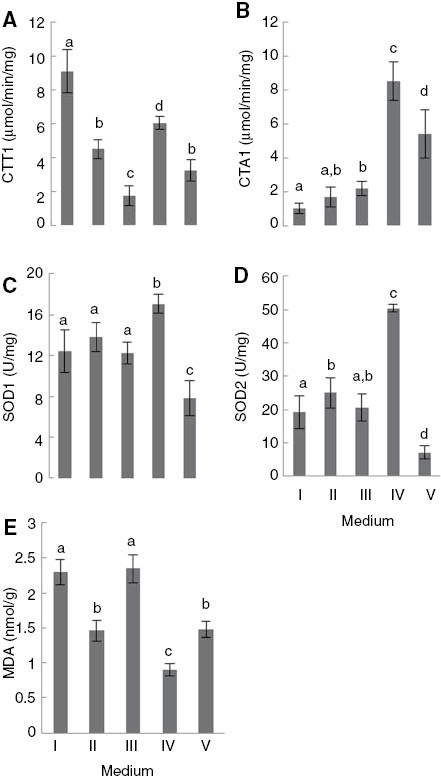
(A) CTT1, (B) CTA1, (C) SOD1, (D) SOD2 activity and (E) MDA level of Saccharomyces cerevisiae UE-ME3 grown in I- YEPG medium, II- YEPGD medium, III- YEPGD medium and 5 μg/ml TiO2-NP add at 100 min, IV-YEPGD medium with heat shock (28/40°C) at 100 min,V-YEPGD medium and 5 μg/ml TiO2-NP with heat shock (28/40°C) at 100 min. The results represent the mean of five independent experiments±standard deviation. The bars marked with different letters (a–d) are significantly different, in accordance with the Duncan test (p<0.01).
Furthermore, heat shock conditions caused an increase in the capacity of S. cerevisiae for scavenging superoxide anion radical and hydrogen peroxide when yeast cells are grown in the YEPGD (respirable/fermentable) medium, since there occurred an increase in CTT1, CTA1, SOD1 and SOD2 enzyme activity, achieving values which were higher than those found under the other test conditions, except for cytoplasmic catalase (CTT1), whose specific activity achieved values which were lower than those detected in yeast cells grown in the YEPG medium (respirable). The exposure of S. cerevisiae to 5 μg/ml TiO2-NP with heat shock produced an opposite effect in terms of CTT1 activity achieving values approximate to those which were determined in yeast grown only in the YEPGD medium (respirable/fermentable) and also causing a significant decrease in CTA1 activity to values lower than those detected in S. cerevisiae exposed to heat shock only but which were higher than those detected under the other test conditions (Figure 3A, B) (p<0.01). However the combined effect of 5 μg/ml TiO2-NP and heat shock on the YEPGD medium significantly decreased cytoplasmic (SOD1) and mitochondrial (SOD2) superoxide dismutase activities (p<0.01), a response that may have contributed to the increase of ROS and triggering the oxidative stress described above (Figure 3C, D). The malondialdehyde level determined in S. cerevisiae UE-ME3 grown in the YEPG medium (respirable) was higher than in yeast cells grown in the YEPGD medium (respirable/fermentable) (Figure 3E) (p<0.01), in which the protection against oxidative damage in respiratory/fermentative cells may have primarily depended on the response of superoxide dismutase (SOD2) and catalase (CTA1) located in cellular compartments surrounded by membranes, since cytoplasmic glutathione peroxidase (GPX) and catalase (CTT1) activities presented a significant decrease (p<0.01). Exposure of S. cerevisiae grown in the YEPGD medium to 5 μg/ml TiO2-NP reversed the significant decrease in the malondialdehyde level induced by the presence of glucose in the culture medium, presenting values approximate those detected in the yeast grown in the absence of glucose (YEPG, a respirable medium) (p<0.01). In this case, the loss of enzymatic antioxidant capacity, such as the decrease of GR and catalase (CTT1) activities, may have contributed to the observed increase in oxidative damage. The results also revealed that yeast cells grown in the YEPGD medium under heat shock conditions showed the lowest level of MDA. It is possible that the significant increase in CTA1, CTT1, SOD1 and SOD2 activity may have contributed to minimising cell damage resulting from heat shock exposure. Surprisingly, the exposure of S. cerevisiae to 5 μg/ml TiO2-NP with heat shock significantly reversed the effect observed in cells grown in the YEPGD medium containing NPs, the malondialdehyde level decreasing to values approximate to those which were determined in cells grown in the YEPGD medium only (p<0.01). In this case, the significant increase in CTA1 (≈3.0x) and CTT1 (≈1.5x) enzyme activity may have contributed to minimising the oxidative damage resulting from TiO2-NP exposure in the respirable/fermentable medium (YEPDG).
4 Conclusions
The biomass produced by S. cerevisiae grown in the YEPGD medium (respirable/fermentable) with 5 μg/ml TiO2-NP was similar to the biomass produced by the yeast cells in the YEPG medium (respirable), but less than that which was determined in the YEPGD medium (respirable/fermentable). The presence of TiO2-NP in YEPGD medium also caused a decrease in cell ability for regenerating glutathione (GSH) by means of GR and detoxifying hydrogen peroxide by means of catalase (CTT1), increasing cell damage (MDA level). The oxidation-reduction properties of the cell environment were disturbed (GSH/GSSG ratio decrease) in the S. cerevisiae grown in the YEPGD medium (respirable/fermentable) with 5 μg/ml TiO2-NP with heat shock, probably due to a decrease in their reserves of glutathione (GSH) and nonprotein thiols (GSH+GSSG). In addition, the decrease in GR activity detected in the yeast cells grown in the YEPGD medium (respirable/fermentable), but only exposed to 5 μg/ml TiO2-NP, was even less under heat shock conditions. Furthermore, the exposure of S. cerevisiae to 5 μg/ml titanium dioxide NPs with heat shock triggered a decrease in yeast cell capacity for detoxifying the superoxide anion radical, mediated by cytoplasmic (SOD1) and mitochondrial (SOD2) superoxide dismutase activities and the antioxidant power mediated by glutathione peroxidase (GPx), with an increase in intracellular ROS. In conclusion, under heat shock conditions TiO2-NP caused oxidative stress in S. cerevisiae grown in the presence of glycerol and glucose, decreasing antioxidant defences such as superoxide dismutases or a slowdown of the glutathione cycle.
About the authors

Joana Capela-Pires graduated in Biochemistry from the University of Évora, Évora, Portugal (2009). From the same university, she obtained a MSc in Biochemistry with a thesis on the antioxidant response of Saccharomyces cerevisiae to titanium dioxide nanoparticles (2011). Since 2011, she has been a PhD student at the Department of Chemistry and Institute of Mediterranean Agrarian and Environmental Sciences, University of Évora, Évora, Portugal. Her field of research is: stress and metabolic regulation on S. cerevisiae cells by titanium dioxide nanoparticles; her field of specialization is: energetic and antioxidant systems of yeast cells.

Rui Ferreira graduated in Biochemistry (1987) from the University of Lisbon, Portugal. He obtained a PhD (1999) in Biochemistry from the University of Évora, Portugal. He was an Assistant Professor (1999–2004) and has been an Associate Professor of Enzymology since 2004 at the Department of Chemistry, University of Évora. Since 1988, he has also been a researcher at ICAAM, University of Évora. His fields of research are stress and metabolic regulation by metals and pesticides. He specializes in energetics, biotransformation and antioxidant systems of animals and yeast.

Isabel Alves-Pereira graduated as an agronomic engineer from the University of Évora, Portugal. She obtained an MSc in Microbial Molecular Genetics from the University of Minho, Portugal and a PhD in Biochemistry (2008) from the University of Extremadura, Spain. Since 2008, she has been an Assistant Professor of Biochemistry at the Department of Chemistry, University of Évora. Since 1994, she has also been a researcher at ICAAM, University of Évora. Her fields of research are the effects of metals and pesticides on antioxidant and biotransformation systems.
Acknowledgments
This work is funded by FEDER Funds through the Operational Programme for Competitiveness Factors (COMPETE) and National Funds through Foundation for Science and Technology (FCT) under the Strategic Projects PEst-C/AGR/UI0115/2011 and PEst-OE/AGR/UI0115/2014.
References
[1] Hagfeldt A, Gratzel M. Chem. Rev. 1995, 95, 49–68.Suche in Google Scholar
[2] Chen H, Su S, Chien T, Lin W, Yu S, Chou C, Chen J, Yang P. The FASEB J. 2006, 20, 2393–2395.Suche in Google Scholar
[3] Warheit D, Hoke R, Finlay C, Donner E, Reed K, Sayes C. Toxicol. Lett. 2007, 171, 99–110.Suche in Google Scholar
[4] Zhu X, Chang Y, Chen Y. Chemosphere 2010, 78, 209–215.10.1016/j.chemosphere.2009.11.013Suche in Google Scholar PubMed
[5] Kasemets K, Ivask A, Dubourguier H-C, Kahru A. Toxicol. In Vitro. 2009, 23, 1116–1122.Suche in Google Scholar
[6] Chang Y-N, Zhang M, Xia L, Zhang J, Xing G. The Materials. 2012, 5, 2850–2871.Suche in Google Scholar
[7] Fleischer A, O’Neill MA, Ehwald R. Plant Physiol. 1999, 121, 829–838.Suche in Google Scholar
[8] Fujino T, Itoh T. Plant Cell Physiol. 1998, 39, 1315–1323.Suche in Google Scholar
[9] Madigan MT, Martinko JM, Parker J. Brock Biology of Microorganisms, Prentice Hall/Pearson Higher Education Group: Upper Saddle River, 2003.Suche in Google Scholar
[10] Zemke-White WL, Clements KD, Harris PJ. J. Exp. Mar Bio. Ecol. 2000, 245, 57–68.Suche in Google Scholar
[11] Nel AE, Madler L, Velegol D, Xia T, Hoek EM, Somasundaran P, Klaessig, F, Castranova V, Thompson M. Nat. Mater. 2009, 8, 543–557.Suche in Google Scholar
[12] Chen R, Huang G, Ke P-C. Appl. Phys. Lett. 2010, 97, ID 093706.Suche in Google Scholar
[13] Mortimer M, Kasemets K, Vodovnik M, Marinsek-Logar R, Kahru A. Environ. Sci. Technol. 2011, 45, 6617–6624.Suche in Google Scholar
[14] Navarro E, Baun A, Behra R, Hartmann NB, Filser J, Miao A-J, Quigg A, Santschi PH, Sigg L. Ecotoxicology 2008, 17, 372–386.10.1007/s10646-008-0214-0Suche in Google Scholar PubMed
[15] Moore VC, Strano MS, Haroz EH, Hauge RH, Smalley RE, Schmidt J, Talmon Y. Nano. Lett. 2003, 3, 1379–1382.Suche in Google Scholar
[16] Ju-Nam Y, Lead JR. Sci. Total. Environ. 2008, 400, 396–414.Suche in Google Scholar
[17] Li N, Xia T, Nel AE. Free Radic. Biol. Med. 2008, 44, 1689–1699.Suche in Google Scholar
[18] Shukla RK, Sharma V, Pandey AK, Singh S, Sultana S, Dhawan A. Toxicol. In Vitro. 2011 25, 231–241.Suche in Google Scholar
[19] Fahmy B, Cormier S A. Toxicol. In Vitro. 2009, 23, 1365–1371.Suche in Google Scholar
[20] Radu M, Munteanu M C, Petrache S, Serban A I, Dinu D, Hermenean A, Sima C and Dinischiotu A. Acta Biochimica Polonica. 2010, 57, 355–360.Suche in Google Scholar
[21] Manke A, Wang L, Rojanasakul Y. Biomed. Res. Int. 2013, ID 942916.10.1155/2013/942916Suche in Google Scholar PubMed PubMed Central
[22] Sies H. Oxidative Stress Oxidants and Antioxidants, Academic Press: London, 1991.Suche in Google Scholar
[23] Gilmore W, Kirby GJ. Pharmacol. Exp. Ther. 2004, 308, 600–608.Suche in Google Scholar
[24] Rice-Evans C, Miller N, Paganga G. Free Radic. Biol. Med. 1996, 20, 933–956.Suche in Google Scholar
[25] Espindola AS, Gomes DS, Panek AD, Eleutherio ECA. Cryobiology 2003, 47, 236–241.10.1016/j.cryobiol.2003.10.003Suche in Google Scholar PubMed
[26] Filomeni G, Rotilio G, Ciriolo M. Biochem. Pharmacol. 2002, 64, 1057–1064.Suche in Google Scholar
[27] Rowley A, Johnston GC, Butler B, Werner-Washburne M, Singer RA. Mol. Cell. Biol. 1993,13, 1034–1041.Suche in Google Scholar
[28] Mager WH, Ferriera PM. Biochem. J. 1993, 290, 1–13.Suche in Google Scholar
[29] Tissieres A, Mitchell HK, Tracy U. J. Mol. Biol. 1974, 84, 389–398.Suche in Google Scholar
[30] Miller MJ, Xuong NH, Geiduschek EP. J. Bacteriol. 1982, 151, 311–327.Suche in Google Scholar
[31] Lindquist S. Annu. Rev. Biochem. 1986, 55, 1151–1191.Suche in Google Scholar
[32] Trott A, Morano KA. Yeast Stress Response, 1st ed., Springer Press: New York, 2003.Suche in Google Scholar
[33] Patriarca EJ, Maresca B. Exp. Cell Res. 1990, 190, 57–64.Suche in Google Scholar
[34] Richter K, Haslbeck M, Buchner J. Mol. Cell. 2010, 40, 253–266.Suche in Google Scholar
[35] Sakaki K, Tashiro K, Kuhara S, Mihara K. J. Biochem, 2003. 134, 373–384.10.1093/jb/mvg155Suche in Google Scholar PubMed
[36] Davidson JF, Schiestl RH. Cell. Biol. 2001, 21, 8483–8489.Suche in Google Scholar
[37] Davidson JF, Whyte B, Bissinger PH, Schiestl RH. Proc. Natl. Acad. Sci. USA. 1996, 93, 5116–5121.Suche in Google Scholar
[38] Miller MJ, Xuong NH, Geiduschek EP. Proc. Natl. Acad. Sci. USA. 1979, 76: 5222–5225.10.1073/pnas.76.10.5222Suche in Google Scholar PubMed PubMed Central
[39] McAlister L, Strausberg S, Kulaga A, Finkelstein DB. Curr. Genet. 1979, 1, 63–74.Suche in Google Scholar
[40] Hendrick JP, Hartl FU. Annu. Rev. Biochem. 1993, 62, 349–384.Suche in Google Scholar
[41] Imazu H, Sakurai H. Cell. 2005, 4, 1050–1056.Suche in Google Scholar
[42] Jarolim S, Ayer A, Pillay B, Gee AC, Phrakaysone A, Perrone GG, Breitenbach M, Dawes IW. Genes, Genomics Genetics. 2013, 3, 2321–2323.Suche in Google Scholar
[43] Dueñas-Sánchez R, Gutiérrez G, Rincón AM, Codón AC, Benítez T. FEMSYeast Res. 2012, 12, 625–636.Suche in Google Scholar
[44] DeRisi JL, Iyer VR, Brown PO. Science 1997, 278, 680–686.10.1126/science.278.5338.680Suche in Google Scholar PubMed
[45] Roberts GG, Hudson AP. Mol. Genet. Genomics. 2006, 276, 170–186.Suche in Google Scholar
[46] Watanabe D, Wu H, Noguchi C, Zhou Y, Akao T, Shimoi H. Appl. Environ. Microbiol. 2010, 77, 934–941.Suche in Google Scholar
[47] Otero-González L, García-Saucedo C, Field JA, Sierra-Álvarez R. Chemosphere 2013, 93, 1201–1206.10.1016/j.chemosphere.2013.06.075Suche in Google Scholar PubMed
[48] Park S, Lee S, Kim B, Lee S, Lee J, Sim S, Gu M, Yi J, Lee J. Biotechnol. BioprocessEng. 2012, 17, 276–282.Suche in Google Scholar
[49] Bayat N, Rajapakse K, Marinsek-Logar R, Drobne D, Cristobal S. Nanotoxicology 2014, 8, 363–373.10.3109/17435390.2013.788748Suche in Google Scholar PubMed
[50] Carrasco P, Querol A, del Olmo M. Arch. Microbiol. 2001, 175, 450–457.Suche in Google Scholar
[51] Klaine S, Koelmans A, Horne N, Carley S, Handy R, Kapustka L, Nowack B, Kammer F. Environ. Toxicol. Chem. 2012, 31, 3–14.Suche in Google Scholar
[52] Federici G, Shaw B. Aquat. Toxico. 2007, 84, 415–430.Suche in Google Scholar
[53] Lovern S, Klape R. Environ. Toxicol. Chem. 2006, 25, 1132–1137.Suche in Google Scholar
[54] Rosado T, Conim A, Alves-Pereira I, Ferreira R. Ecotoxicology. 2009. 18, 1116–1122.Suche in Google Scholar
[55] Lake, B. Biochemical Toxicology – A Practical Approach, 1st ed., IRL Press: Oxford, 1987.Suche in Google Scholar
[56] Lowry O, Rosebrough G, Farr A. J. Biol. Chem. 1951, 193, 265–275.Suche in Google Scholar
[57] Hissin A, Hilf P. Anal. Biochem. 1976, 74, 214–226.Suche in Google Scholar
[58] LeBel P, Ischiropoulos H, Bondy C. Chem. Res Toxicol. 1992, 5, 227–231.Suche in Google Scholar
[59] Uchiyama M, Mihara M. Anal. Biochem. 1978, 86, 271–278.Suche in Google Scholar
[60] Turton H, Dawes I, Grant C. J. Bacteriol. 1997, 179, 1096–1101.Suche in Google Scholar
[61] Goldberg D, Spooner R. Methods of Enzymatic Analysis, 3rd ed., Bergmayer, VCH: New York, 1987.Suche in Google Scholar
[62] Inoue Y, Matsuda T, Sugiyama K, Izawa S, Kimura A. J. Biol. Chem, 1999. 17, 27002–27009.10.1074/jbc.274.38.27002Suche in Google Scholar PubMed
[63] Postma E, Verduyn C, Scheffers W, van Dijken J. Appl. Environ. Microbiol. 1989. 55, 468–477.Suche in Google Scholar
[64] Beers R, Sizer I. J. Biol. Chem. 1952, 195, 133–140.Suche in Google Scholar
[65] Oberley L, Spitz D. Meth. Enzymol. 1984, 105, 457–464.Suche in Google Scholar
[66] Kelley R, Ideker T. PLoS Genet. 2009, 5, e1000488.Suche in Google Scholar
[67] Kasemets K, Nisamedtinov I, Laht T-M, Abner K, Paalme T. Antonie Leeuwenhoek. 2007, 92, 109–128.Suche in Google Scholar
[68] Paumi CM, Pickin KA, Jarrar R, Herren CK, Cowley ST. FEBS Lett. 2012, 586, 847–853.Suche in Google Scholar
[69] Adamis PDB, Panek AD, Eleutherio ECA. Toxicol. Lett. 2007, 173, 1–7.Suche in Google Scholar
©2015 by De Gruyter
This article is distributed under the terms of the Creative Commons Attribution Non-Commercial License, which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Artikel in diesem Heft
- Frontmatter
- In this issue
- Editorial
- Has GPS landed with precision?
- Original articles
- Solar production of WO3: a green approach
- Sesbania sesban L. biomass as a novel adsorbent for removal of Pb(II) ions from aqueous solution: non-linear and error analysis
- Green methacrylated lignin model compounds as reactive monomers with low VOC emission for thermosetting resins
- Deposition behavior of TiB2 by microwave heating chemical vapor deposition (CVD)
- Interaction and thermodynamics of methylene blue adsorption on oxidized multi-walled carbon nanotubes
- Alkaline leaching of zinc from low-grade oxide zinc ore using ammonium citrate as complexing agent
- Heat shock and titanium dioxide nanoparticles decrease superoxide dismutase and glutathione enzymes activities in Saccharomyces cerevisiae
- Momordica charantia fruit mediated green synthesis of silver nanoparticles
- The continuous synthesis and application of graphene supported palladium nanoparticles: a highly effective catalyst for Suzuki-Miyaura cross-coupling reactions
- Conference announcements
- 5th Flow Chemistry Congress (San Diego, CA, USA, September 15–16, 2015)
- Conferences 2015–2017
- Book reviews
- Flow chemistry
- Biomass as a sustainable energy source for the future: fundamentals of conversion processes
Artikel in diesem Heft
- Frontmatter
- In this issue
- Editorial
- Has GPS landed with precision?
- Original articles
- Solar production of WO3: a green approach
- Sesbania sesban L. biomass as a novel adsorbent for removal of Pb(II) ions from aqueous solution: non-linear and error analysis
- Green methacrylated lignin model compounds as reactive monomers with low VOC emission for thermosetting resins
- Deposition behavior of TiB2 by microwave heating chemical vapor deposition (CVD)
- Interaction and thermodynamics of methylene blue adsorption on oxidized multi-walled carbon nanotubes
- Alkaline leaching of zinc from low-grade oxide zinc ore using ammonium citrate as complexing agent
- Heat shock and titanium dioxide nanoparticles decrease superoxide dismutase and glutathione enzymes activities in Saccharomyces cerevisiae
- Momordica charantia fruit mediated green synthesis of silver nanoparticles
- The continuous synthesis and application of graphene supported palladium nanoparticles: a highly effective catalyst for Suzuki-Miyaura cross-coupling reactions
- Conference announcements
- 5th Flow Chemistry Congress (San Diego, CA, USA, September 15–16, 2015)
- Conferences 2015–2017
- Book reviews
- Flow chemistry
- Biomass as a sustainable energy source for the future: fundamentals of conversion processes