Insidious pathogen-mimicking properties of nanoparticles in triggering the lectin pathway of the complement system
-
S. Moein Moghimi
S. Moein Moghimi read Biochemistry at the University of Manchester (UK) and in 1989 earned his PhD in Biochemistry (Liposome Immunobiology) from Charing Cross Hospital Medical School (Imperial College, University of London). He is currently based at the University of Copenhagen (Denmark) where he serves as Professor of Nanomedicine at the Department of Pharmacy, Professor of Pharmaceutical Nanotechology at the NanoScience Centre, and Director of the Centre for Pharmaceutical Nanotechnology and Nanotoxiocology. He is also a full member and affiliate professor at the Department of Translational Imaging, Houston Methodist Research Institute (Weill Cornell Medical College), Houston Methodist Hospital Systems, Houston, Texas (USA), adjoint professor at the Department of Pharmaceutical Sciences, Skaggs School of Pharmacy and Pharmaceutical Sciences, University of Colorado-Denver Medical Center (USA), visiting professor at University of Padova (Italy) and the elected Fellow of the Institute of Nanotechnology (UK). Earlier, he served as the Honorary Professor of Nanomedicine at the Multidisciplinary Research Center, Shantou University (China). Before joining Copenhagen, Moein was Senior Lecturer in Biopharmacy and Molecular Pharmaceutics at the School of Pharmacy, University of Brighton (UK) and The University Research Fellow in Advanced Drug Delivery Systems at the Department of Pharmaceutical Sciences, University of Nottingham (UK). His research activities are focused on pharmaceutical nanoscience and fundamental nanomedicine and nanosafety. Moein is the Associate Editor of
Nanomedicine: Nanotechnology, Biology and Medicine and theJournal of Biomedical Nanotechnology and further serves on the editorial board of several peer-reviewed international journals, includingAdvanced Drug Delivery Reviews, Nanomedicine-UK ,Journal of Liposome Research andMolecular and Cellular Therapies.  ,
Peter P. Wibroe
,
Peter P. Wibroe
Peter P. Wibroe has a solid background at the interface of nanotechnology and pharmaceutical sciences. Peter is dedicating his research to the intriguing interplay between the physicochemical properties of synthetic nanoparticles and biological systems, with a special focus on the complement system. Peter is currently enrolled as a PhD student in the Nanomedicine Group led by Professor Moghimi, where he is developing immune-safe and efficient drug delivery vehicles based on understanding of the molecular mechanisms by which nanoparticle surface presentation modulates complement recognition and responses. In this respect, Peter has already published several papers and reviews about the complement system and the underlying mechanisms of material recognition.
Linping WU obtained a Master’s degree in Biomaterials from Shantou University (China) in 2008. He began his PhD studies under the direction of Professor Moghimi at the University of Copenhagen and obtained his PhD in Nanomedicine in 2014. Currently he is a senior postdoctoral scientist in the same group, focusing on design and engineering of immune-safe advanced biomaterials and drug delivery systems for specific targeting through an interdisciplinary approach.
Z. Shadi Farhangrazi, PhD, MS, MBA is a biochemist, neuroscientist, infectious diseases expert, strategist, entrepreneur, and an expert in the area of innovation and entrepreneurship. She is the Founder, President and Managing Partner of management consulting firm Biotrends International. Farhangrazi is also a faculty member at Daniels College of Business, and University College, University of Denver. She has been teaching workshops and classes on entrepreneurship, innovation, strategy and non-profit management and has worked with multinational companies, SMEs, non-profit organizations and governmental organizations internationally. She has an active interest in nanomedicine innovation and translation.

Abstract
The lectin pathway of the complement system is an integral component of the innate immune system recognizing pathogens through patterns of sugar moieties displayed on their surfaces and neutralizing them through an antibody-independent reaction cascade. Many engineered nanoparticles incite complement through the lectin pathway, but these nanoparticles inherently do not express surface-exposed sugars. However, the projected polymeric surface architecture of nanoparticles may transiently resemble structural motifs of peptidoglycan constituents of pathogens and trigger the lectin pathway. We discuss these issues in relation to nanomedicine design and immune safety.
Nanoparticles and the complement system
Many functional particles (e.g., iron oxide nanocrystals, gold nanoparticles, carbon nanotubes, graphene) and drug carriers (e.g., liposomes, polymeric nanospheres, oil-in-water emulsions, polymeric micelles) exhibit an array of ‘pathogen-mimicking’ properties, which arises from a combination of their nanoscale size and aspect ratio, morphology and surface display of repetitive epitopes (e.g., clusters of functional groups, surface projected polymers), making them susceptible for interception by the body’s defences (1). The complement system is a key component of the innate immunity, which can be triggered by a wide repertoire of invading particles (including pathogens) (2). Accordingly, the complement system provides critical immunoprotective and immunoregulatory functions comprising opsonization, lytic and inflammatory processes (e.g., chemotaxis and leukocyte activation) (2). There are three established pathways of complement activation: classical, lectin and alternative pathways (2). Each pathway is triggered differently, but they converge to generate the same set of effector molecules (2, 3).
Complement activation and fixation has been a central point for efficient clearance of nanoparticles and drug carriers by phagocytic cells (4, 5). On the other hand, inadvertent complement activation may initiate adverse reactions and this has been noted following infusion of many therapeutic agents including antibody, polymeric, micellar- and particulate-based formulations into human subjects (6–8). The exact role and contribution of the complement system in infusion-related reactions is not clear, but liberation of complement bioactive products (e.g., C3a, C5a and C5b-9) can potentially modulate the function of a variety of immune cells (e.g., macrophages, monocytes, polymorphonuclear cells, platelets, mast cells) and vascular endothelial cells either directly or in cross-talk with pattern-recognition receptors (e.g., Toll-like receptors 2, 4 and 9) to account for some of the observed adverse events (6, 7). Furthermore, complement activation is also of serious concern for successful development of intravenous anti-cancer nanomedicine initiatives (9, 10). Indeed, recent studies have indicated that intratumoral accumulation of complement activating long-circulating nanoparticles in immunocompetent mice can accelerate tumor growth through C5a liberation (9), presumably through recruitment of regulatory T cells (resulting in deregulation or suppression of CD8+ cytotoxic T cell activity), immunosuppressive monocytes and alternatively activated macrophages into malignant tumors as well as increasing tumor angiogenesis (9–11). Therefore, understanding of nanomaterial properties that incite complement is a prerequisite for design and engineering of immunologically safer nanomedicine and biomedical devices.
The interaction between particulate matters and the complement system is complex and regulated by inter-related factors comprising morphology, dimension, chemical make up and surface characteristics (3). The majority of complement activation studies with medically relevant nanoparticles have been predominantly qualitative with limited focus on mechanistic aspects (1, 3). In case of the latter, attention has been mostly towards the classical and alternative pathways (3). Compelling evidence now suggest that many such nanoparticles insidiously incite complement through the lectin pathway, which involve carbohydrate recognition, but these nanoparticles natively do not express surface-exposed sugars (12–21).
The lectin pathway
In humans, five pattern recognition molecules are capable of initiating the lectin pathway (22, 23). These include mannose-binding lectin (MBL), ficolins of M, L and H classes, and collectin 11 (CL11 or CL-K1) all in association with a complex set of serine protease zymogens. To date, three serine proteases, namely MBL-associated serine protease (MASP) 1, 2 and 3 have been identified (22). In addition, two non-enzymatic fragments of the MASPs (MAp44 and MAp19) have been found in the recognition complexes (22). MBL binds carbohydrates with 3- and 4-hydroxyl groups in the pyranose ring (d-mannose and N-acetyl-d-glucosamine, GlcNAc) in a calcium-dependent manner (22). Serum L- and H-ficolin has a common binding specificity for GlcNAc. The M-ficolin is not a serum protein, but its mRNA is found in leukocytes and lung (22). CL11 has preference for L-fucose and d-mannose and binding is calcium-dependent, which resembles the specificity of mouse MBL-A (23). Binding of the lectin pathway recognition molecules to the corresponding surface-exposed carbohydrate ligands activates MASPs, although the exact molecular details are still poorly understood (22, 24). Typically, MASP2 cleaves the fourth and the second complement proteins (C4 and C2, respectively) to form the C3 convertase C4bC2a (22). In the absence of MASP2, the lectin pathway is not functional as confirmed in both MASP2-depleted human serum and MASP2 knockout mouse (25, 26). A recent study (24), however, indicated that MASP2 activation strictly depends on MASP1, and inhibition of MASP1 prevents activation of MASP2. Furthermore, MASP1 was suggested to produce 60% of C2a responsible for C3 convertase formation (24). Accordingly, a new model of lectin pathway activation proposes that MASP1 when activated cleaves zymogen MASP2, where the active MASP2 then cleaves C4 and the associated C4b binds C2, which is then cleaved primarily by MASP1 and to lesser extent by MASP2 thus forming the C4bC2a complex (24). MASP1 is also able to cleave C3 directly, which results in activation of the alternative pathway (27). Others, based on a knockout mouse model, have also proposed a role for MASP1 and MASP3 in directly activating the alternative pathway factor D as well as limited factor B cleavage by MASP3 (28–30). However, there are ongoing debates on these findings (31, 32).
Examples of nanoparticle-mediated activation of lectin pathway
Nanoparticle surface camouflaging with materials such as poly(ethylene glycol)s, PEGs, and block copolymers of poloxamer and poloxamine series has long been shown to combat macrophage recognition and confer longevity in the blood (1, 33, 34). Although long believed that such strategies could suppress opsonization events in the blood (33, 34), recent studies have demonstrated that some of these engineered entities can, indeed, incite complement and depending on surface polymer configuration complement activation may proceed exclusively through the lectin pathway (12, 13, 17–19, 21). For example, alteration of poloxamine 908 copolymer architecture on polystyrene nanospheres of 220 nm in size from a flat to mushroom-brush configuration switched complement activation from the C1q-dependent classical pathway to a lectin pathway in human serum (12). This copolymer has repetitive recognition patterns of relative polarity and hydrophobicity, where the patterns may change with changes in surface density of the copolymer. Consequently, this may create new binding sites for complement recognition and shifts complement activation pathway from classical to lectin mode. Indeed, one intriguing aspect of PEG and block copolymers of poloxamer and poloxamine series is structural similarities between their terminal region and d-mannose/GlcNAc (Figure 1). In this respect, the surface projected poly(ethylene oxide) chains of the poloxamine 908 in close proximity (as in mushroom-brush configuration) may form dynamic ‘pathogen-mimicking’ clusters transiently resembling structural motifs of the d-mannose/GlcNAc, which serves as a platform for MBL/ficolin/CL11 docking (Figure 2). Accordingly, it is not surprising to see that block copolymers in different forms (e.g., micelles, gels) also trigger lectin pathway in a size- and shape-independent manner (14).
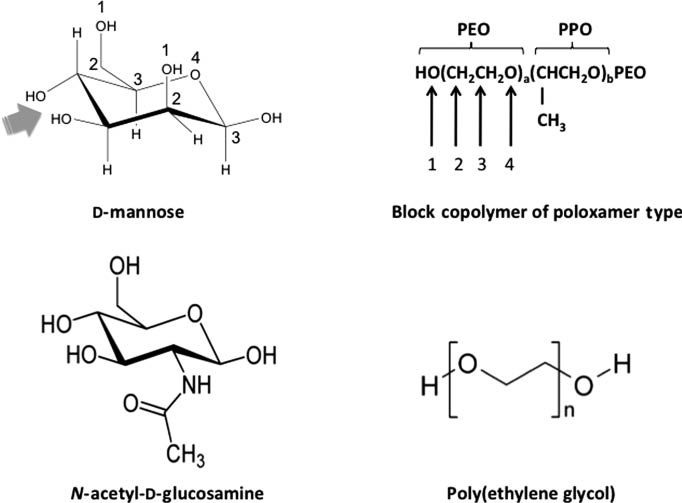
Structural similarities between d-mannose, N-acetyl-d-glucosamine and selected polymers used in nanoparticle engineering. Compare sequences numbered 1–4 between the sugars and polymers, which can be repeated in different ways on both sugars. The arrow indicates the MBL binding site (the equatorial OH groups) in d-mannose (arrow head). The binding sites for ficolins and collectin 11, however, remain unknown. PEO, polyethylene oxide; PPO, polypropylene oxide. The structure of poloxamines is not shown, but poloxamines are star-shaped copolymers comprising of four POP chains joined by a central ethylene diamine bridge, where each POP block is flanked at the other end by a POE chain.
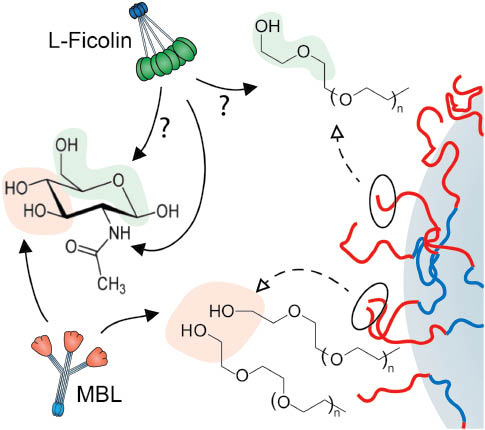
Possible mechanisms of nanoparticle-mediated activation of the complement lectin pathway. The right segment is a schematic representation of a polymer-coated nanoparticle. The dynamics of the terminal regions of surface projected polymers (e.g., poloxamers and poloxamines) in close proximity may transiently resemble structural motifs of N-acetyl-d-glucosamine/d-mannose recognizable by lectin pathway initiators (e.g., MBL, L-ficolin). See Figure 1 for polymer structures.
Similar observations have been reported with carbon nanotubes coated with PEG-conjugated molecules or covalently functionalized with PEG, where gross surface polymer architectural changes not only triggered lectin pathway, but further modulated the mode of lectin pathway activation through binding of different lectin pathway initiating molecules (e.g., MBL vs. L-ficolin) (13).
Non-specific adsorption of heavily glycosylated plasma proteins such as apolipoprotein B-100 and certain classes of antibodies to surface engineered nanoparticles may further contribute to lectin pathway activation (12, 14, 34). On binding and conformational transformation, the glycosylated modules of these proteins may act as template for binding of lectin pathway initiators.
Dextran-coated super paramagnetic iron oxide nanoparticles also incite complement through lectin pathway (21). In human sera, activation was MBL-dependent as well as proceeding directly through the alternative pathway. In mouse sera, activation was MBL-A/C dependent, but alternative pathway contributed via the amplification loop only (21). These observations highlight important differences and similarities in nanoparticle-mediated complement activation between humans and mice. Accordingly, complement activation and related adverse events studies in animals may not necessarily translate into a human system (10, 21).
Outlook
Evolution has afforded complement system a powerful tool to combat pathogens through lectin pattern recognition. It is not surprising to see that man-made materials such as various polymers [e.g., PEG, poloxamers, polxamines, poly(caprolactone), poly(glycolyic acid), poly(lactic acid), poly(lactide-co-glycolide), polyoxazoline, chitosan] and drug delivery systems based on such polymers trigger lectin pathway. Indeed, many such entities share structural motifs with peptidoglycan constituents of the pathogens that incite complement through MBL, ficolin and collectin 11 binding, and therefore needs revisiting. For example, chitosan is composed of repeating units of d-glucosamine, together with residual GlcNAc units. A repertoire of glycosylated plasma proteins may further aid lectin pathway activation on nanoparticle binding and conformational changes and also warrants clarification (34). Protein fingerprinting and mass spectrometry studies, however, reveal little in terms of surface-dependent and fluid-phase complement cascade and the precise role of lectin pathway initiators in nanoparticle-mediated complement activation (35, 36), where qualitative analysis could be misleading (1). Future studies should examine nanoparticle-mediated lectin pathway activation of the human complement system more carefully through integrated approaches and ‘structure-activity’ mapping, which may lead to identification of structural motifs and geometrical arrangements that dock lectin pathway initiators. Collectively, these approaches may initiate better approaches for safer nanoparticle design and engineering (an ‘immune safe-by-redesign’ concept). Indeed, a recent study demonstrated that MBL-binding kinetics are critically dependent on structural characteristics on the nanometer scale, including the dimensions of polyvalent MBL oligomers as well as the mode of ligand presentation on surfaces (37). Future developments in microstamping may further aid mechanistic understanding of spatial and architectural arrangements of immobilized polymers in controlling the binding kinetics of lectin pathway initiators. Through such studies and computational analysis it may be possible to simulate time-dependent nanoparticle surface changes in vivo (e.g., in the blood or on nanoparticle extravasation to interstitial spaces) that may subsequently shift complement activation from one pathway to another, thereby initiating delayed reactions. Finally, complement activation as whole seems to play an important role in nanomedicine-mediated infusion-related reactions and progression of cancer, but the exact contribution of the lectin pathway needs to be mapped out. Accordingly, we should be cautious in extrapolating the outcomes of animal studies to human safety pertaining intravenous nanomedicines and nanopharmaceuticals, since there are differences in complement activation processes between humans and other species and particularly in relation to lectin pathway activation.
About the authors

S. Moein Moghimi read Biochemistry at the University of Manchester (UK) and in 1989 earned his PhD in Biochemistry (Liposome Immunobiology) from Charing Cross Hospital Medical School (Imperial College, University of London). He is currently based at the University of Copenhagen (Denmark) where he serves as Professor of Nanomedicine at the Department of Pharmacy, Professor of Pharmaceutical Nanotechology at the NanoScience Centre, and Director of the Centre for Pharmaceutical Nanotechnology and Nanotoxiocology. He is also a full member and affiliate professor at the Department of Translational Imaging, Houston Methodist Research Institute (Weill Cornell Medical College), Houston Methodist Hospital Systems, Houston, Texas (USA), adjoint professor at the Department of Pharmaceutical Sciences, Skaggs School of Pharmacy and Pharmaceutical Sciences, University of Colorado-Denver Medical Center (USA), visiting professor at University of Padova (Italy) and the elected Fellow of the Institute of Nanotechnology (UK). Earlier, he served as the Honorary Professor of Nanomedicine at the Multidisciplinary Research Center, Shantou University (China). Before joining Copenhagen, Moein was Senior Lecturer in Biopharmacy and Molecular Pharmaceutics at the School of Pharmacy, University of Brighton (UK) and The University Research Fellow in Advanced Drug Delivery Systems at the Department of Pharmaceutical Sciences, University of Nottingham (UK). His research activities are focused on pharmaceutical nanoscience and fundamental nanomedicine and nanosafety. Moein is the Associate Editor of Nanomedicine: Nanotechnology, Biology and Medicine and the Journal of Biomedical Nanotechnology and further serves on the editorial board of several peer-reviewed international journals, including Advanced Drug Delivery Reviews,Nanomedicine-UK, Journal of Liposome Research and Molecular and Cellular Therapies.

Peter P. Wibroe has a solid background at the interface of nanotechnology and pharmaceutical sciences. Peter is dedicating his research to the intriguing interplay between the physicochemical properties of synthetic nanoparticles and biological systems, with a special focus on the complement system. Peter is currently enrolled as a PhD student in the Nanomedicine Group led by Professor Moghimi, where he is developing immune-safe and efficient drug delivery vehicles based on understanding of the molecular mechanisms by which nanoparticle surface presentation modulates complement recognition and responses. In this respect, Peter has already published several papers and reviews about the complement system and the underlying mechanisms of material recognition.

Linping WU obtained a Master’s degree in Biomaterials from Shantou University (China) in 2008. He began his PhD studies under the direction of Professor Moghimi at the University of Copenhagen and obtained his PhD in Nanomedicine in 2014. Currently he is a senior postdoctoral scientist in the same group, focusing on design and engineering of immune-safe advanced biomaterials and drug delivery systems for specific targeting through an interdisciplinary approach.

Z. Shadi Farhangrazi, PhD, MS, MBA is a biochemist, neuroscientist, infectious diseases expert, strategist, entrepreneur, and an expert in the area of innovation and entrepreneurship. She is the Founder, President and Managing Partner of management consulting firm Biotrends International. Farhangrazi is also a faculty member at Daniels College of Business, and University College, University of Denver. She has been teaching workshops and classes on entrepreneurship, innovation, strategy and non-profit management and has worked with multinational companies, SMEs, non-profit organizations and governmental organizations internationally. She has an active interest in nanomedicine innovation and translation.
Acknowledgments
SMM acknowledges financial support by the Danish Agency for Science, Technology and Innovation, reference 09-065736 (Det Strategiske Forskningsråd) and the European Community’s Seventh Framework Programme (FP7-NMP-2012-Large-6) under grant agreement No. 310337-2 CosmoPHOS-nano.
References
1. Moghimi SM, Hunter AC, Andresen TL. Factors controlling nanoparticle pharmacokinetics: an integrated approach and perspective. Annu Rev Pharmacol Toxicol 2012;52:481–503.10.1146/annurev-pharmtox-010611-134623Suche in Google Scholar PubMed
2. Ricklin D, Hajishengallis G, Yang K, Lambris JD. Complement – a key system for immune surveillance and homeostasis. Nat Immunol 2010;11:785–97.10.1038/ni.1923Suche in Google Scholar PubMed PubMed Central
3. Moghimi SM, Andersen A, Ahmadvand D, Wibroe PP, Andresen TL, Hunter AC. Material properties in complement activation. Adv Drug Deliv Rev 2011;63:1000–7.10.1016/j.addr.2011.06.002Suche in Google Scholar PubMed
4. Moghimi SM, Parhamifar L, Ahmadvand D, Wibroe PP, Andresen TL, Farhangrazi ZS, et al. Particulate systems for targeting of macrophages: basic and therapeutic concepts. J Innate Immun 2012;4:509–28.10.1159/000339153Suche in Google Scholar PubMed PubMed Central
5. Moghimi SM, Farhangrazi ZS. Nanoparticle in medicine: nanoparticle engineering for macrophage targeting and nanoparticles that avoid macrophage recognition. In: Boraschi D, Duschl A, editors. Nanoparticles and the immune system. Safety and effects. San Diego: Elsevier, 2014:77–89.Suche in Google Scholar
6. Moghimi SM, Wibroe PP, Helvig SY, Farhangrazi ZS, Hunter AC. Genomic perspectives in inter-individual adverse responses following nanomedicine administration: the way forward. Adv Drug Deliv Rev 2012;64:1385–93.10.1016/j.addr.2012.05.010Suche in Google Scholar PubMed
7. Moghimi SM, Farhangrazi ZS. Nanomedicine and complement paradigm. Nanomedicine: Nanotechnol Biol Med 2013;9:458–60.10.1016/j.nano.2013.02.011Suche in Google Scholar PubMed
8. Szebeni J. Complement activation-related pseudoallergy: a stress reaction in blood triggered by nanomedicines and biological. Mol Immunol 2014;61:163–73.10.1016/j.molimm.2014.06.038Suche in Google Scholar PubMed
9. Moghimi SM. Cancer nanomedicines and the complement system activation paradigm: anaphylaxis and tumour growth. J Control Rel 2014;190:556–62.10.1016/j.jconrel.2014.03.051Suche in Google Scholar PubMed
10. Moghimi SM, Farhangrazi ZS. Just so stories: random acts of anti-cancer nanomedicine performance. Nanomedicine: Nanotechnol Biol Med 2014;10:1661–66.10.1016/j.nano.2014.04.011Suche in Google Scholar PubMed
11. Sabnani MK, Rajan R, Rowland B, Mavinkurve V, Wood LM, Gabizon AA, et al. Liposome promotion of tumor growth is associated with angiogenesis and inhibition of antitumor immune responses. Nanomedicine: Nanotechnol Biol Med 2015;11:259–62.10.1016/j.nano.2014.08.010Suche in Google Scholar PubMed
12. Hamad I, Al-Hanbali O, Hunter AC, Rutt KJ, Andresen TL, Moghimi SM. Distinct polymer architecture mediates switching of complement activation pathways at nanosphere-serum interface: implications for stealth nanoparticles engineering. ACS Nano 2010;4:6629–38.10.1021/nn101990aSuche in Google Scholar PubMed
13. Andersen AJ, Robinson JT, Dai H, Hunter AC, Andresen TL, Moghimi SM. Single-walled carbon nanotubes surface control of complement sensing and activation. ACS Nano 2013;7:1108–19.10.1021/nn3055175Suche in Google Scholar PubMed
14. Hamad I, Hunter AC, Moghimi SM. Complement activation by Pluronic 127 gel and micelles: suppression of copolymer-mediated complement activation by elevated serum levels of HDL, LDL, and apolipoproteins A-I and B-100. J Control Rel 2013;170:167–74.10.1016/j.jconrel.2013.05.030Suche in Google Scholar PubMed
15. Andersen AJ, Windschiegl B, Ilbasmis-Tamer S, Degim IT, Hunter AC, Andresen TL, et al. Complement activation by PEG-functionalized MWCNTs is independent of PEG molecular mass and surface density. Nanomedicine: Nanotechnol Biol Med 2013;9:469–73.10.1016/j.nano.2013.01.011Suche in Google Scholar PubMed
16. Wibroe PP, Moghimi SM. Complement sensing of nanoparticles and nanomedicines. In: Hepel M, Zhong CJ, editors. Functional nanoparticles for bioanalysis, nanomedicine and bioelectronic devices, ACS Symposium Series, American Chemical Society, 2012;2:365–82.Suche in Google Scholar
17. Moghimi SM, Andersen AJ, Hashemi SH, Lettiero B, Ahmadvand D, Hunter AC, et al. Complement activation cascade triggered by PEG-PL engineered nanomedicines and carbon nanotubes: the challenges ahead. J Control Rel 2010;146:175–81.10.1016/j.jconrel.2010.04.003Suche in Google Scholar PubMed
18. Moghimi SM, Hunter AC. Complement monitoring of carbon nanotubes. Nat Nanotechnol 2010;5:382.10.1038/nnano.2010.110Suche in Google Scholar
19. Hamad I, Hunter AC, Rutt KJ, Liu Z, Dai H, Moghimi SM. Complement activation by PEGylated single-walled carbon nanotubes is independent of C1q and alternative pathway turnover. Mol Immunol 2008;45:3797–803.10.1016/j.molimm.2008.05.020Suche in Google Scholar PubMed PubMed Central
20. Hamad I, Hunter AC, Szebeni J, Moghimi SM. Poly(ethylene glycol)s generate complement activation products in human serum through increased alternative pathway turnover and a MASP-2-dependent process. Mol Immunol 2008;46:225–32.10.1016/j.molimm.2008.08.276Suche in Google Scholar PubMed
21. Banda NK, Mehta G, Chao Y, Wang G, Inturi S, Fossati-Jimak L, et al. Mechanisms of complement activation by superparamagentic dextran iron oxide (SPIO) nanoworms in mouse versus human sera. Particle Fibre Toxicol 2014;11:64.10.1186/s12989-014-0064-2Suche in Google Scholar PubMed PubMed Central
22. Fujita T. Evolution of the lectin-complement pathway and its role in innate immunity. Nat Rev Immunol 2002;2:346–53.10.1038/nri800Suche in Google Scholar PubMed
23. Hansen S, Selman L, Palaniyar N, Ziegler K, Brandt J, Kliem A, et al. Collectin 11 (CL-11, CL-K1) is a MASP-1/3-associated plasma collectin with microbial-binding activity. J Immunol 2010;185:6096–104.10.4049/jimmunol.1002185Suche in Google Scholar
24. Heja D, Kocsis A, Dobo J, Szilagyi K, Szasz R, Zavodszky P, et al. Revised mechanism of complement lectin-pathway activation revealing the role of serine protease MASP1 as the exclusive activator of MASP-2. Proc Natl Acad Sci USA 2012;109:10498–503.10.1073/pnas.1202588109Suche in Google Scholar
25. Møller-Kristensen M, Thiel S, Sjöholm A, Matsushita M, Jensenius JC. Co-operation between MASP-1 and MASP-2 in the generation of C3 convertase through the MBL pathway. Int Immunol 2007;19:141–49.10.1093/intimm/dxl131Suche in Google Scholar
26. Schwaeble WJ, Lynch NJ, Clark JE, Marber M, Samani NJ, Ali YM, et al. Targeting of mannan-binding lectin-associated serine protease-2 confers protection from myocardial and gastrointestinal ischemia/re-perfusion injury. Proc Natl Acad Sci USA 2011;108:7523–28.10.1073/pnas.1101748108Suche in Google Scholar
27. Matsushita M, Fujita T. Cleavage of the third component of complement (C3) by mannose-binding-protein-associated serine protease (MASP) with subsequent complement activation. Immunobiology 1995;194:443–48.10.1016/S0171-2985(11)80110-5Suche in Google Scholar
28. Sekine H, Takahashi M, Iwaki D, Fukita T. The role of MASP-1/3 in complement activation. Adv Exp Med Biol 2013;735:41–53.10.1007/978-1-4614-4118-2_3Suche in Google Scholar PubMed
29. Takahashi M, Ishida Y, Iwaki D, Kanno K, Suzuki T, Endo Y, et al. Essential role of mannose-binding lectin-associated serine protease-1 in activation of the complement factor D. J Exp Med 2010;207:29–37.10.1084/jem.20090633Suche in Google Scholar PubMed PubMed Central
30. Banda NK, Takahashi M, Levitt B, Glogowska M, Nicholas J, Takahashi K, et al. Essential role of complement mannose-binding lectin-associated serine proteases-1/3 in the murine collagen antibody-induced model of inflammatory arthritis. J Immunol 2010;185:5598–606.10.4049/jimmunol.1001564Suche in Google Scholar PubMed PubMed Central
31. Degn SE, Jensenius JC, Thiel S. The pro-factor D cleaving activity of MASP-1/-3 is not required for alternative pathway function. J Immunol 2014;192:5447–8.10.4049/jimmunol.1400777Suche in Google Scholar PubMed
32. Takahashi M, Sekine H, Fujita T. Comment on “The pro-factor D cleaving activity of MASP-1/-3 is not required for alternative pathway function”. J Immunol 2014;192:5448–9.10.4049/jimmunol.1400766Suche in Google Scholar PubMed
33. Moghimi SM, Hunter AC, Murray JC. Long-circulating and target-specific nanoparticles; theory to practice. Pharmacol Rev 2001;53:283–318.Suche in Google Scholar
34. Moghimi SM, Szebeni J. Stealth liposomes and nanoparticles: critical issues on protein-binding properties, activation of proteolytic blood cascades and intracellular fate. Prog Lipid Res 2003;42:463–78.10.1016/S0163-7827(03)00033-XSuche in Google Scholar
35. Tenzer S, Docter D, Kuharev J, Musyanovych A, Fetz V, Hecht R, et al. Rapid formation of plasma protein corona critically affects nanoparticle pathophysiology. Nat Nanotechnol 2013;8:772–81.10.1038/nnano.2013.181Suche in Google Scholar PubMed
36. Cedervall T, Lynch I, Lindman S, Berggard T, Thulin E, Nilsson H, et al. Understanding the nanoparticle-protein corona using methods to quantify exchange rates and affinities of proteins for nanoparticles. Proc Natl Acad Sci USA 2007;104:2050–5.10.1073/pnas.0608582104Suche in Google Scholar PubMed PubMed Central
37. Gjelstrup LC, Kaspersen LD, Behrens MA, Pedersen JS, Thiel S, Kingshott P, et al. The role of nanometer-scaled ligand patterns in polyvalent binding by large mannan-binding lectin oligomers. J Immunol 2012;188:1292–306.10.4049/jimmunol.1103012Suche in Google Scholar PubMed
©2015 by De Gruyter
Artikel in diesem Heft
- Frontmatter
- In this issue
- News
- News from the European Foundation for Clinical Nanomedicine (CLINAM)
- What’s up in nanomedicine?
- Special Section Nanosafety
- Guest Editorial
- Sizing up the safety of nanomaterials
- Essay
- Keeping it small: towards a molecular definition of nanotoxicology
- Reviews
- The bio-corona and its impact on nanomaterial toxicity
- In vitro-ex vivo model systems for nanosafety assessment
- Meeting Report
- Nanosafety forum for young scientists: a meeting report
- Short Communication
- Nanomedicines in the European translational process
- Special Section CARPA Part 2
- Guest Editorial
- Complement activation-related pseudoallergy: an innate response to nanomedicines acting as pseudo-viruses
- Reviews
- Exosomes: potential model for complement-stealth delivery systems
- Lessons learned from the porcine CARPA model: constant and variable responses to different nanomedicines and administration protocols
- Mini Review
- Blood cell changes in complement activation-related pseudoallergy
- Short Communications
- Membrane attack complex formation on a supported lipid bilayer: initial steps towards a CARPA predictor nanodevice
- Paradoxical rise of hemolytic complement in the blood of mice during zymosan- and liposome-induced CARPA: a pilot study
- Commentary
- Insidious pathogen-mimicking properties of nanoparticles in triggering the lectin pathway of the complement system
Artikel in diesem Heft
- Frontmatter
- In this issue
- News
- News from the European Foundation for Clinical Nanomedicine (CLINAM)
- What’s up in nanomedicine?
- Special Section Nanosafety
- Guest Editorial
- Sizing up the safety of nanomaterials
- Essay
- Keeping it small: towards a molecular definition of nanotoxicology
- Reviews
- The bio-corona and its impact on nanomaterial toxicity
- In vitro-ex vivo model systems for nanosafety assessment
- Meeting Report
- Nanosafety forum for young scientists: a meeting report
- Short Communication
- Nanomedicines in the European translational process
- Special Section CARPA Part 2
- Guest Editorial
- Complement activation-related pseudoallergy: an innate response to nanomedicines acting as pseudo-viruses
- Reviews
- Exosomes: potential model for complement-stealth delivery systems
- Lessons learned from the porcine CARPA model: constant and variable responses to different nanomedicines and administration protocols
- Mini Review
- Blood cell changes in complement activation-related pseudoallergy
- Short Communications
- Membrane attack complex formation on a supported lipid bilayer: initial steps towards a CARPA predictor nanodevice
- Paradoxical rise of hemolytic complement in the blood of mice during zymosan- and liposome-induced CARPA: a pilot study
- Commentary
- Insidious pathogen-mimicking properties of nanoparticles in triggering the lectin pathway of the complement system