Lessons learned from the porcine CARPA model: constant and variable responses to different nanomedicines and administration protocols
-
Rudolf Urbanics
János Szebeni, MD, PhD, DSc, MedHabil, immunologist, director of the Nanomedicine Research and Education Center at Semmelweis University, Budapest, Hungary. He is also founder and CEO of a contract research SME “SeroScience”, and full Professor of (immune) biology at Miskolc University, Hungary. He has held various guest professor and scientific positions in Hungary and abroad, mostly in the USA where he lived for 22 years. His research on various themes in hematology, membrane biology and immunology resulted in more than 120 scientific papers (citations: >4550, H index: 35), 14 book chapters, two granted patents, a book entitled “The Complement System: Novel Roles in Health and Disease” (Kluwer, 2004). Three fields stand out where he has been most active: artificial blood, liposomes and the complement system. His original works led to the “CARPA” concept, i.e. that complement activation underlies numerous drug-induced (pseudo)allergic (anaphylactoid) reactions.
Péter Bedőcs, MD, PhD graduated cum laude as a doctor of Medicine from Semmelweis University in 2004. He completed his PhD training at the same institution, focusing on pathophysiological effects and immunotoxicity of drug delivery systems utilizing nanotechnology. In collaboration with Semmelweis University, the Bay Zoltan Foundation in Hungary, and the Uniformed Services University of the Health Sciences (USUHS) in the United States, he participated in the development of an animal model for the testing of complement activation by intravenous therapeutic and diagnostic agents, which became an FDA-recommended preclinical test for nanopharmaceuticals. This also led to the establishment of an effective desensitization procedure to prevent immunotoxic reactions provoked by certain nanomedicines.
Currently he works at the Defense and Veterans Center for Integrative Pain Management as a senior scientist and as an Assistant Professor at the Department of Military and Emergency Medicine at USUHS in Bethesda, MD, overseeing multiple clinical and animal studies in the area of patient safety in anesthesia, developing new effective modalities for pain management. Additionally, he continues to serve as a collaborator and consultant for the Malignant Hyperthermia Diagnostic Center at USUHS.
Rudolf Urbanics, MD, PhD, is Head of the in vivo laboratory of Nanomedicine Research and Education Center of Semmelweis University, and SeroScience Ltd., an immunotoxicity CRO, since 2008 in Budapest, Hungary.
He obtained his MD diploma and the PhD degree at Semmelweis Medical School, Budapest, Hungary. He had various research/collaboration positions at MaxPlanck Institute of Systemphysiology, at University of Pennsylvania, Cerebrovascular Research Center, at Pennsylvania Muscle Institute, working in the field of CNS regulation of blood flow/metabolism, ischemic/hypoxic disorders, stroke and chronic neurodegenerative disease animal models.
He was the Deputy R&D Director and Head of CNS Pharmacology Department at Biorex R&D Co., and worked at IVAX/Drug Research Institute Budapest, as a leading researcher in safety and CNS pharmacology and later at the IVAX/Drug Research Institute, a subsidiary of TEVA as Head of the In Vivo Pharmacology Group.
Currently, he is working with in vivo models of nano drug-nano carrier induced, complement activation related pseudoallergic reactions (CARPA), clarifying their immune-toxicological and safety hazards.


Abstract
Pigs provide a sensitive and quantitative animal model of complement (C) activation-related pseudoallergy (CARPA) caused by liposomes and a wide range of nanoparticulate drugs or drug nanocarriers (nanomedicines). The tetrad of symptoms (hemodynamic, hematological, laboratory and skin changes) that arise within minutes after i.v. injection of reactogenic nanomedicines (RNMs) are highly reproducible among different pigs but the presence, direction and relative severity of symptoms are very different with different RNMs and their administration schedule. Bolus administration of RNMs usually trigger pulmonary hypertension with or without various degrees of systemic hyper- or hypotension, tachy-or bradycardia, arrhythmia, blood cell and inflammatory mediator changes and skin rash. These reactions can be rapid or protracted, and fully tachyphylactic, semi-tachyphylactic or non-tachyphylactic. Slow infusion usually diminishes the reactions and/or entail delayed, protracted and less severe hemodynamic and other changes. The goal of this review is to present some technical details of the porcine CARPA model, point out its constant and variable parameters, show examples of different reactions, highlight the unique features and capabilities of the model and evaluate its utility in preclinical safety assessment. The information obtained in this model enables the understanding of the complex pathomechanism of CARPA involving simultaneous anaphylatoxin and inflammatory mediator actions at multiple sites in different organs.
Introduction: CARPA as an immune-mediated stress reaction in blood triggered by nanomedicines
Nanotechnology has achieved remarkable success in improving the therapeutic index of numerous drugs and agents by the use drug carrier nanosystems (nanocarriers) that carry and target the active pharmaceutical ingredient (API) to its site of action and/or control its ADME properties. However, the resulting increase of efficacy or decrease of toxicity is in some cases not without a price, as many of such nanodrugs or agents can cause adverse effects that the API alone would not do. We refer to them as reactogenic nanomedicines (RNMs). One of such adverse effects is complement (C) activation, which can lead to a syndrome called C activation-related pseudoallergy (CARPA). It is an acute and reversible immune reaction, also known as infusion reaction (or anaphylactic/anaphylactoid or idiosyncratic reaction), or non-immune allergy, whose symptoms, clinical significance, mechanism and many other properties were detailed previously in numerous reviews (1–8). An apparently trivial observation 20 years ago, that C activation underlies the major hemodynamic disturbance in rats following the infusion of liposomes (9) launched the progress of the CARPA concept, whose latest milestone is a broad claim that CARPA is a manifestation of a chemical stress on blood (8). More explicitely, it is a universal defense process against the “threat” of nanomedicines, that can occur in any organisms that has blood and C proteins therein, which falsely perceive the nanoparticles as pathogenic viruses. It is a stress reaction because just like in classical stress, an external harm (“pseudovirus”) triggers a nonspecific, standard battery of physiological changes via a multiorgan “axis”, namely the “immuno-cardiovascular” axis, which corresponds to the “hypothalamo-pituitary-adrenal” axis involved in classical stress (8).
Depending on the features of RNMs, their administration speed and the use of anti-allergic premedication, the prevalence of CARPA in man can reach 30%–40% with mostly transient and mild symptoms. The reaction usually occurs unpredictably at the first use of the drug, as there is no known laboratory or any clinical test that could estimate the risk of a reaction. Because in rare cases CARPA may be severe, even fatal, the phenomenon is getting increasing regulatory attention. For example, the European Medicines Agency’s latest guidance on generic liposome development recommends CARPA assays for preclinical safety testing (10). However, it has not solidified to date which CARPA tests are appropriate, or best for regulatory purposes, which parameters need to be measured and under what conditions? In general, the use and utility of CARPA tests are in an early stage of scientific evaluation.
In vitro tests for CARPA
In vitro testing of CARPA can be non-cellular and cellular. The non-cellular testing is based on the measurement of C activation by the RNM in serum, plasma or whole blood, using C split product ELISAs. There are many C split products that serve as activation marker and have commercially available pathway specific ELISAs, the best known being C3a, C5a, iC3b, C4d, Bb and SC5b-9. One issue in this regard is the inter-laboratory variation of test results, which problem has been addressed recently by efforts to produce international C standards (11–13). The information that these in vitro assays provide on the risk of CARPA is limited, since they report only on the activity of afferent arm of CARPA, the extent of anaphylatoxin formation. The efferent arm, the body’s response to anaphylatoxins, remains unknown. The most relevant cellular assays that can measure anaphylatoxin sensitivity are various basophil assays, which quantitate basophil leukocyte activation and/or secretion to model the mast cells’ response to allergens or other RNMs (14–16). We reported some preliminary, promising results with a basophil assay measuring CD203c upregulation as a predictor of liposomal CARPA (6), however, a study dedicated for the evaluation of the predictive value of the basophil test in pseudoallergy remains to be done.
Animal models of immune toxicity: which is good for CARPA evaluation?
Animal models represent a major tool for the study of mechanisms in virtually all biomedical research. Hypersensitivity reactions result from a complex combination of genetic, environmental and temporal factors as well as complex interactions between the immune and other organ systems and the drug, making these reactions uniquely diverse. As delineated above, in vitro systems are unlikely able to mimic such complexity. There is a consensus in the field that HSRs can only be tested in whole animal models (17–19). The critical question in this regard whether the standard, already accepted tests are appropriate for CARPA evaluation, or not.
Among the established animal models of immune toxicity, some can a priori be ruled out as CARPA test, based on their mode of operation. These include the mouse popliteal lymph node assay (PLNA) (20–22), the mouse ear-swelling test (23–25), the guinea pig (GP) maximization test (GPMT), the GP occluded patch test (Buehler’s test) (26, 27) and the murine local lymph node assay (MLLA) (26). In fact, the standard guinea pig assays were shown to be useless in predicting systemic hypersensitivity (28). The measurement of blood lymphocyte counts and lymph node weight or assaying B, T or other immune cell proliferation or function evaluate long-term immune toxicity, and not CARPA, which is a short term toxicity. Taken together, to our best knowledge, none of the regulatory standard animal tests may be appropriate to assess CARPA.
Non-standard immunotoxicity tests of CARPA in different animals
As pointed out, CARPA may be considered as a universal stress reaction, which implies that it may be present in most, if not all levels of mammalian evolution. Accordingly, there is a great number of reports in the literature on C activation and its consequences in various animals, including rats (9, 29–33), mice (33), dogs (34–37), rabbits (38, 39) nonhuman primates (40–49) and pigs (50–55). Among these experimental systems, it is the pigs’ response to C activator liposomes that best mimics the human infusion reactions to liposomes in terms of kinetics and spectrum of symptoms and the conditions of reaction induction (Table 1). For Doxil, for example, it was calculated that the drug dose that triggers CARPA in pigs corresponds to the dose that triggers infusion reactions in hypersensitive man (56). These facts, taken together with the favorable ethical and financial aspects of working with pigs rather than dogs or primates, rationalizes the use of pigs as CARPA model. Thus, CARPA adds to the list of diseases that are successfully studied in pigs, as discussed below.
Identities, similarities and differences between CARPA in men and pigs.
| Comparison | Abnormalities | Symptoms | |
|---|---|---|---|
| Human | Pig | ||
| Identical | Some cardiopulmonary and hemodynamic alterations | Difficulty of breathing, hypo- or hypertension, arrhythmia, tachycardia, bradycardia, edema | |
| Body temperature | Fever | ||
| Blood cell changes | Leukopenia/leukocytosis, thrombocytopenia | ||
| Skin changes | Erythema, rash | ||
| Range of minimum reactogenic phospholipid dose in infusion or bolus | 1–10 μg/kg/s | ||
| Similar | Blood reaction markers | Rise of SC5b-9 | Rise of TXB2 |
| Cardiopulmonary and hemodynamic alterations | Shortness of breath, fatigue, dizziness, fainting, swelling of the ankles, abdomen or legs, cyanosis, chest pain, passing out or dizziness | Rise of PAP, fall of SAP | |
| Time course | Symptoms start within 10 min after infusion and subside within 30–60 min | PAP rise between 3 and 10 (10) min, returns to normal within 30–60 min | |
| Different | Reaction frequency | <10% | >90% |
PL, phospholipid.
The use of pigs as disease models
Pigs are widely used as large animal models in biomedical research (57–61), particularly in studies on cardiovascular diseases (62–67), trauma (68–71), sepsis (72–76), drug intoxications (54, 55), and, since 1999 (53), CARPA. As described in many previous experimental studies (50–53, 77) and a recent review (7), the porcine CARPA model represents a highly sensitive and reproducible model for the most serious, life threatening HSRs in man caused by RNM.
Technical details of the porcine CARPA model
Figure 1 illustrates the setup and instruments used in the model. In brief, domestic pigs (usually 2–3 months old) or miniature pigs are initially sedated (with Calypsol/Xilazine) and, after tracheal intubation, anesthesized with isoflurane while breathing spontaneously. The animals thereafter undergo surgery to place multiple catheters into their circulation for the measurement of different hemodynamic parameters, administration of test drugs and blood sampling. Namely, a Swan-Ganz catheter is placed to the pulmonary artery wedge, for the measurement of pulmonary arterial pressure (PAP), and (optionally) central venous pressure (CVP) and cardiac output (CO). Additional catheters are placed into the femoral artery to record the systemic arterial pressure (SAP) and, (optionally), left ventricular end-diastolic pressure (LVEDP). The left femoral vein is canulated for blood sampling, and the external jugular vein for the administration of test articles and to maintain a slow drop infusion of saline (∼3 mL/kg/h). For more sophisticated hemodynamic analysis to measure systemic vascular resistance (SVR) and pulmonary vascular resistance (PVR), additional catheters are placed and measurements and calculations are carried out. The hemodynamic, EKG and respiratory parameters are measured continuously, while blood cell counts, O2 saturation, blood analytes (inflammatory and vasoactive mediators) and temperature are measured at predetermined times, usually in 10–20 min intervals. EKG leads I-III are placed at the standard Einthoven positions.
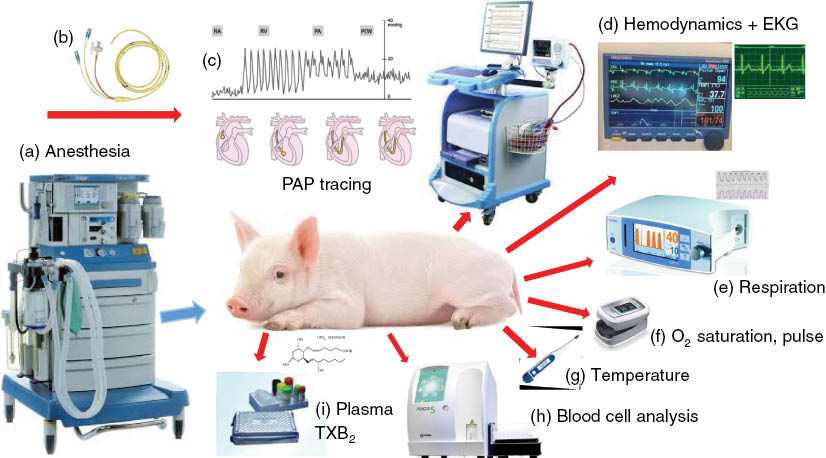
Instruments and endpoints measured in the porcine CARPA model. (a) anesthesia machine; (b) Swan-Ganz catheter; (c) blood pressure wave forms directing the passage of the tip of the Swan-Ganz catheter via the right atrium (RA), right ventricle (RV) and pulmonary artery (PA) until being wedged into the pulmonary capillary bed; (d) computerized multiple parameter hemodynamic monitoring system (1000 Hz sampling rate). From the continuous recording of SAP and PAP signals online averaging is performed and recorded, together with the heart rate, derived from SAP signal; (e) capnograph connected to the tracheal tube to measure respiratory rate, etCO2 and inCO2; (f) pulse oximeter (fixed on the tail) measures O2 saturation in blood and pulse rate; (g) temperature is measured with a thermometer placed in the rectum; (h) veterinary hematology analyzer measuring all blood cell counts and WBC differential; (i) ELISA for measuring biomarkers of allergic/inflammatory reactions, e.g. TXB2, histamine, leukotrienes, adenosine, tryptase, PAF and C3 levels, etc.
The symptom tetrad
Figure 2 illustrates the four types of symptoms observed during CARPA in pigs: hemodynamic (A), hematological (B); laboratory (C) and skin (D) changes. Among the hemodynamic symptoms, the rise of PAP is the most prominent and reproducible measure of CARPA in the porcine model, which is invariably present with all RNMs. The transient, massive pulmonary hypertension is most likely due to the presence of pulmonary intravascular macrophages (PIM cells) in the lung of pigs, a theory based on the speed (seconds) and prominence (maximal possible) of pulmonary changes in this species taken together with the known properties and functions of PIM cells (78). Namely, PIM cells are directly exposed to blood and their function is to screen out from blood particulate pathogens. They can be activated both by anaphylatoxins and via particle binding to their surface receptors, and they respond to activation with massive secretion of vasoactive mediators (78). These features are necessary and at the same time enough to explain the characteristic changes of PAP during CARPA in pigs.
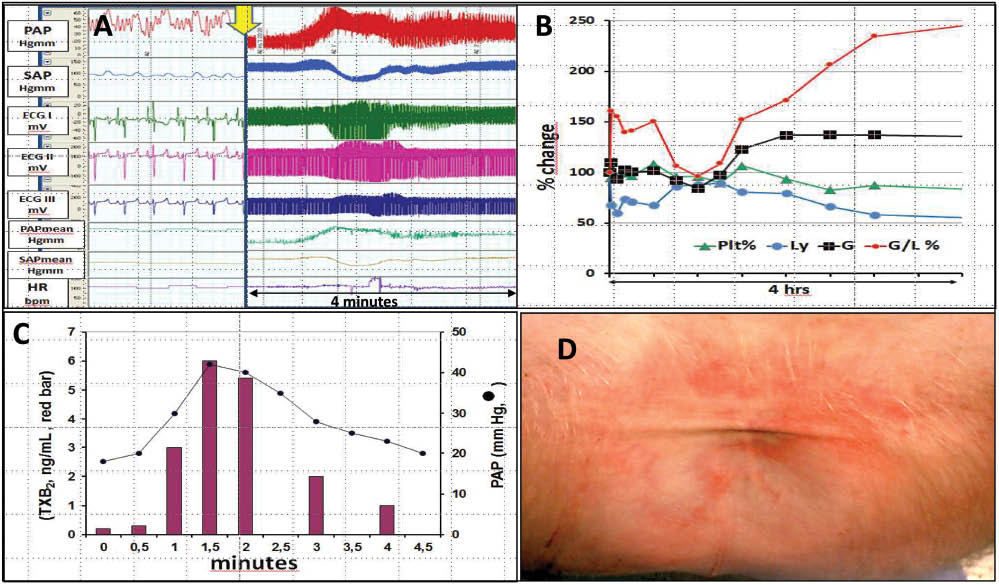
The symptom tetrad of CARPA in pigs. The hemodynamic (A) hematological (B), blood chemistry (C) and skin (D) changes are characteristic of porcine CARPA. Reproduced with permission from (6, 7).
The changes of SAP are more variable; it can drop, rise, display no change or undulate. The hematological changes typically include initial leukopenia followed by protracted leukocytosis and thrombocytopenia: among these the leukopenic effect is the most frequent. Among the laboratory changes the rise of TXB2 is measured most often as it was found to show massive alterations in CARPA (53). The exploration of changes of further allergy and inflammatory mediators (e.g. tryptase, leukotrienes, PAF, chemokines, cytokines) represents an unmet need in this field. Finally, the skin changes are rare and variable, they are seen only in case of very strong reactions.
Uniqueness of the porcine CARPA model
The uniqueness of the pig CARPA model lies in the identity, or close similarities of symptoms to the human CARPA reactions (Table 1) and the quantitative nature and reproducibility of measured endpoints, most significantly the rise of PAP. It should be stressed here again that the high reproducibility applies to the reactions to the same RNMs in different pigs, and NOT to the reactions to different RNMs.
Invariable parameters of porcine CARPA
As mentioned, the rise of PAP to i.v. bolus injection of certain RNMs is remarkably constant and reproducible if the dose and administration schedule is the same in all animals. This statement is demonstrated in Table 2, which summarizes the rise of PAP to 3 different RNMs in numerous independent experiments performed over many years.
Inter-experimental variation of pulmonary hypertensive responses to (first) bolus administration of reactogenic liposomal nanomedicines and to zymosan.
| 1st PAP change (% of baseline±S.E.M.) | n | |
|---|---|---|
| Zymosan | 368.9±57.32 | 15 |
| AmBisome | 236.81±100.91 | 7 |
| Doxil | 233.11±91.79 | 12 |
Data collected from n different experiments performed over years.
Yet another remarkable constancy of the model is the rise of PAP in the same animal, upon repetitive administration of a non-tachyphylactic RNM, such as large multilamellar liposomes (MLV) consisting of dimyristoyl phosphatidylcholine, dimyristoyl phosphatidylglycerol and cholesterol (50:5:45 mole ratios). Figure 3 illustrates this constancy: repeated injection of 5-mg lipid containing MLV in a pig at 30- to 60-min intervals eight times raised the PAP with negligible (6%) coefficient of variation (53).
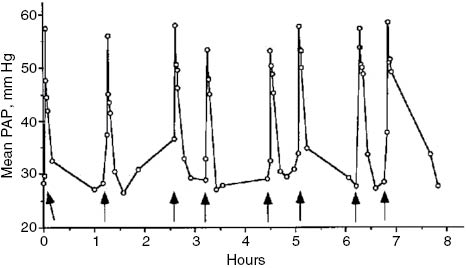
PAP responses to repetitive liposome injections. A pig was injected intravenously with 5-mg lipid-containing liposome boluses at indicated time points, and changes in PAP were recorded. Arrows indicate time points of injections. Figure reproduced from (53) with permission.
Variable parameters of porcine CARPA
Reaction kinetics
The standard pulmonary reaction in porcine CARPA, observed immediately after bolus injection of RNMs consists of a rapid rise of PAP within 2–5 min after the bolus, a plateau reached in 5–15 min and decline to baseline or near baseline within 15–60 min (Figures 2A and 5). However, there are unpublished examples for delayed start and/or protracted reactions as well, which we obtained with bolus administration of a surface conjugated liposomal nanomedicine (Figure 4A) and upon stepwise infusion of a PEGylated liposomal drug candidate (Figure 4B).
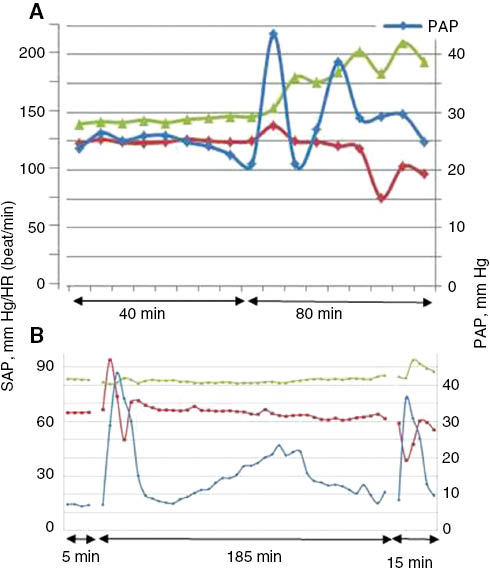
Examples of delayed and or protracted pulmonary hypertension caused by reactogenic nanomedicines. Excerpts from experiments performed in our laboratory, in collaboration with the suppliers of RNMs, which were conjugated (A) and PEGylated (B) small unilamellar liposomes (SUV). Blue, red and green indicate PAP, SAP and heart rate curves registered continuously. Unpublished data.
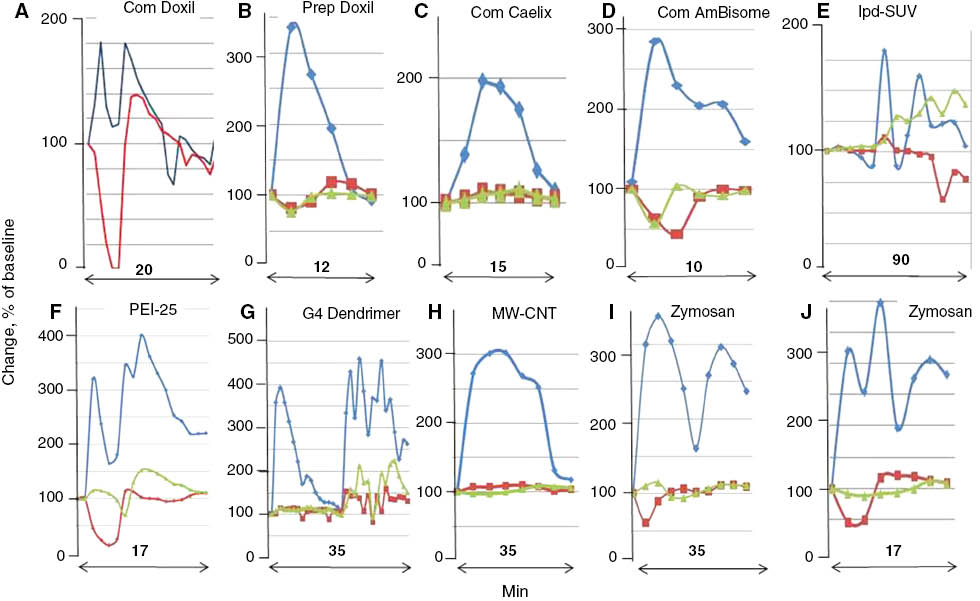
Variation of PAP and SAP waveforms. Panels were redrawn from different experiments wherein the CARPAgenic potential of nanoparticulate drugs or drug carriers were tested in pigs. Minutes indicate the timespan of reactions. Blue, red and green are PAP, SAP and heart rate curves respectively. Changes are shown in % of baseline. Abbreviations only here: com, commercial; prep, self-prepared; lpd, lipophilic prodrug-containing liposomes (79); PEI25, 25 kD pegylated poly(ethylene imine) (80); G4 dendrimer, 4th generation dendrimer; MW-CNT, multiwall carbon nanotube. Most curves were reconstituted from experiments in Refs. (7, 50, 51, 77, 81). The applied doses of different reaction triggers ranged in the 0.01–0.5 mg/kg range, for individual values see original references.
In the former experiment, testing the reactogenicity of a surface-conjugated SUV (Figure 4A), the rise of PAP started only 40 min after bolus administration of the sample and the curve showed 3 progressively diminishing peaks over ∼1.5 h. This observation remains to be understood, just as the late start of a protracted pulmonary hypertension observed during stepwise infusion of a PEGylated SUV drug candidate (Figure 4B). After the initial, usual sharp and rapid reaction another reaction started at 30 min, the PAP peaked at 110 min and returned to normal after 180 min. These complex curves reflect complex interactions underlying the reactions, whose understanding will require a lot more work. It should be noted though that in clinical practice reactions can also start late, even hours after the infusion, and may also extend over hours.
Blood pressure wave forms
Another feature of porcine CARPA is that the relative intensities and waveforms of PAP and SAP show substantial variation among different reaction inducers. Figure 5 shows a selection of different PAP/SAP/heart rate (HR) curves that were registered following bolus injection of different RNMs specified in the legend. The combination of different changes appears random at this time, as no known particle property can be associated with any particular time course or kinetic variables of PAP or SAP or HR. Nevertheless one observation can be explained: the initial splicing of PAP arises most likely from the coincidence of pulmonary hypertension with systemic hypotension, best discernible in Figure 5A. The example shown for extended reaction is in Panel E, wherein a highly negative SUV formulation of an anticancer lipophilic (pro)drug was tested. The roller-coaster swings of PAP extended over 1.5 h, which may be attributed to metabolism or short-term physiological effects of the liposomes or the released drug leading to varying blood levels of different vasoactive mediators.
Tachyphylaxis
Yet another property of CARPA showing significant variation depending on the RNM is the presence/absence, and duration of tachyphylaxis, i.e. self-induced tolerance. The importance of tachyphylactic CARPA lies in the capability it offers for tolerance induction, i.e. to develop safe administration protocols for RNMs. In tachyphylactic CARPA the first dose of the RNM induces tolerance to the second or later administration of the same RNM, and if the first tolerogenic dose is made harmless, the hypersensitivity reactions (HSRs) can be entirely prevented. The utilization of tachyphylaxis for the prevention of HSR was first described for Doxil, wherein the HSR could be prevented by prior slow infusion with Doxebo (82) (Figure 6A), a doxorubicine-free (placebo) Doxil that retains the tachyphylaxis-causing, self- tolerogenic effect of Doxil without causing major HSR (82). However, not all RNMs can cause full tachyphylaxis, as it is a variable and structure-dependent property of RNMs; and it can also be absent (like in Figure 3), partial (Figure 6B) or biphasic (Figure 6C).
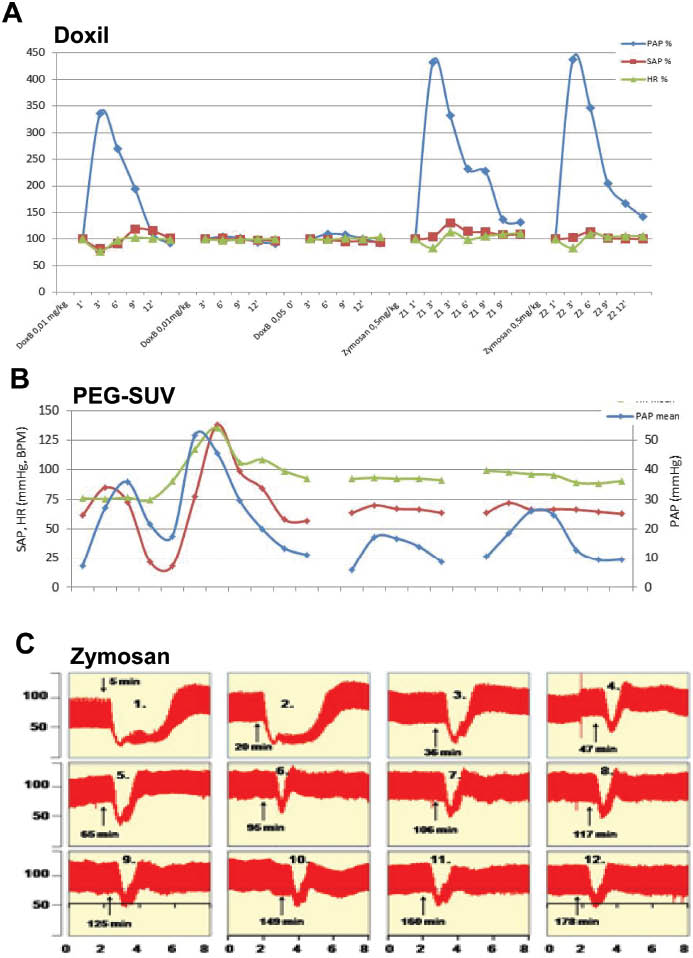
Full, partial and biphasic tachyphylaxis. (A) First 3 injections: repeated bolus administration of 0.01, 0.01 and 0.05 mg/kg Doxil, followed by 2 injections of 0.5 mg/kg zymosan. Figure shows the PAP (Blue), SAP (red) and HR (green) values as % of baseline. x-Axis: Minutes. Reproduced from (82). (B) Repeated bolus administration of comparable doses of another PEGylated reactogenic liposome preparation, containing an anti-inflammatory steroid. Unpublished data. (C) Repeated bolus administration of 0.01 mg/kg zymosan in a pig, Y values are SAP, mm Hg. Reproduced from (7) with permission.
To date, the absence of tachyphylaxis was observed with MLV (Figure 3), zymosan (Figure 6A), Ambisome (50), some PEI polymers (80) and, in general, highly charged nanoparticles (unpublished observations). Partial tachyphylaxis occurs more frequently with liposomes and it is a dose dependent phenomenon. The biphasic tachyphylaxis shown in Figure 6C, wherein a section of nontachyphylactic responses switches to another nontachyphylatic section with smaller, but equal peaks, was observed for SAP after repetitive administration of the same dose of zymosan (7).
The mechanism of full or partial tachyphylaxis is not understood; based on its rapid appearance (minutes), it is most likely a passive phenomenon rather than an immune learning-based, cell-mediated active process. One theory, referred to as “double hit” hypothesis, points to allergy-mediating cells (mast cells/basophils/PIM cells as anatomic sites of tachyphylaxis, and suggests that tachyphylactogenic RNMs trigger CARPA both via C activation and via direct binding to these allergy mediating cells, and the two signals together trigger these cells to release vasoactive mediators. If one of these mechanisms is not working for some reason at the time of repeated treatments, because a mediator gets consumed or a signal transduction pathway gets exhausted, tachyphylaxis will ensue (7, 78).
Summary and future directions
This review provided background information for placing the porcine CARPA model on the map of preclinical immunotoxicological tests for CARPA. It also focused, for the first time, on the constant and variable features of CARPA symptoms in pigs, particularly those seen in the hemodynamic response. The inter-animal and inter-experimental stability of PAP changes in response to a certain type of RNM, and switch to another type of response with another type of RNM represent remarkable, yet ill-understood features of the model that reflect the involvement of highly regulated, complex immunological and physiological processes.
Figure 7 illustrates this complexity by providing an overall scheme of molecular and cellular interactions in CARPA (8). It shows the involvement of several organ systems, numerous cells and highly active biomolecules and mediators and multiple redundant pathways in both the afferent (triggering of allergy-mediating cells) and efferent arms (mediation of trigger signal to the effector cells) of CARPA.
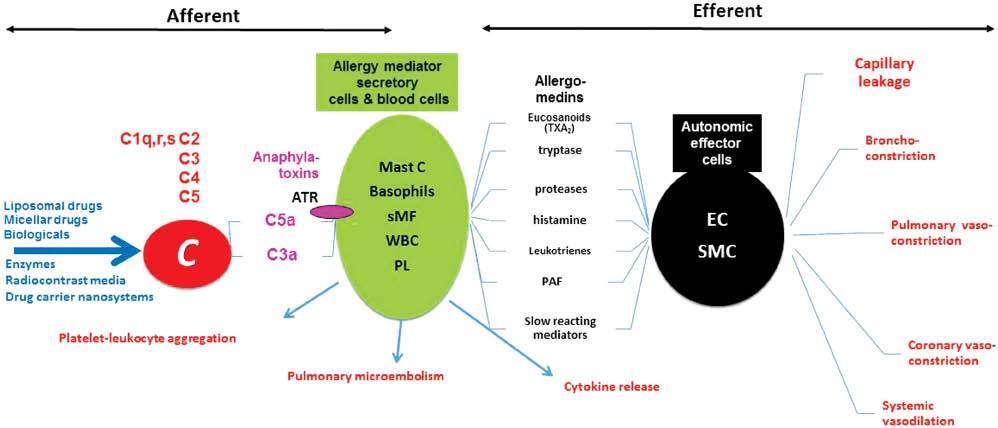
Afferent and efferent arms of CARPA. The hypothetical scheme illustrates the steps and interactions among a great number of cells and mediators involved in CARPA. AR, anaphylatoxin receptors; Mast C, mast cells; sMF, secretory macrophages; WBC, white blood cells; PL, platelets; EC, endothelial cells; SMC, smooth muscle cells. Different types of systems, cells, mediators and effects are color coded. Modified from (8) with permission.
Figure 8 provides further glimpses into the complex picture of CARPA pathogenesis, by highlighting the vicious cycle of hemodynamic and other organ derangements that may lead to the worst outcome of this immune toxicity; anaphylactic shock with circulatory collapse.
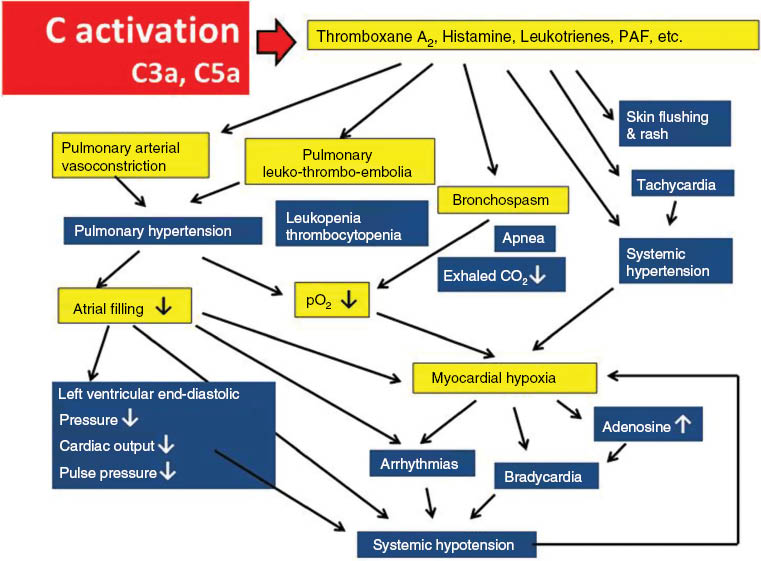
The vicious cycle of physiological changes in CARPA. The scheme illustrates the cause-effect relationships among different physiological changes that entail the activation of the CARPA cascade. The entries on blue and yellow background indicate clinical symptoms and underlying physiological changes, respectively. Reproduced from (8) with permission.
In summary, the constant and, at the same time highly variable symptoms of CARPA in pigs reflect the simplicity and at the same time the complexity of a “stress reaction in blood” (8), a novel immune phenomenon brought to light by the introduction of nanotechnology in pharmacotherapy. Further studies on the porcine CARPA model will hopefully reveal more details of the phenomenon and more understanding of its clinical use in the safety evaluation of novel nanomedicines.
About the authors

János Szebeni, MD, PhD, DSc, MedHabil, immunologist, director of the Nanomedicine Research and Education Center at Semmelweis University, Budapest, Hungary. He is also founder and CEO of a contract research SME “SeroScience”, and full Professor of (immune) biology at Miskolc University, Hungary. He has held various guest professor and scientific positions in Hungary and abroad, mostly in the USA where he lived for 22 years. His research on various themes in hematology, membrane biology and immunology resulted in more than 120 scientific papers (citations: >4550, H index: 35), 14 book chapters, two granted patents, a book entitled “The Complement System: Novel Roles in Health and Disease” (Kluwer, 2004). Three fields stand out where he has been most active: artificial blood, liposomes and the complement system. His original works led to the “CARPA” concept, i.e. that complement activation underlies numerous drug-induced (pseudo)allergic (anaphylactoid) reactions.

Péter Bedőcs, MD, PhD graduated cum laude as a doctor of Medicine from Semmelweis University in 2004. He completed his PhD training at the same institution, focusing on pathophysiological effects and immunotoxicity of drug delivery systems utilizing nanotechnology. In collaboration with Semmelweis University, the Bay Zoltan Foundation in Hungary, and the Uniformed Services University of the Health Sciences (USUHS) in the United States, he participated in the development of an animal model for the testing of complement activation by intravenous therapeutic and diagnostic agents, which became an FDA-recommended preclinical test for nanopharmaceuticals. This also led to the establishment of an effective desensitization procedure to prevent immunotoxic reactions provoked by certain nanomedicines.
Currently he works at the Defense and Veterans Center for Integrative Pain Management as a senior scientist and as an Assistant Professor at the Department of Military and Emergency Medicine at USUHS in Bethesda, MD, overseeing multiple clinical and animal studies in the area of patient safety in anesthesia, developing new effective modalities for pain management. Additionally, he continues to serve as a collaborator and consultant for the Malignant Hyperthermia Diagnostic Center at USUHS.

Rudolf Urbanics, MD, PhD, is Head of the in vivo laboratory of Nanomedicine Research and Education Center of Semmelweis University, and SeroScience Ltd., an immunotoxicity CRO, since 2008 in Budapest, Hungary.
He obtained his MD diploma and the PhD degree at Semmelweis Medical School, Budapest, Hungary. He had various research/collaboration positions at MaxPlanck Institute of Systemphysiology, at University of Pennsylvania, Cerebrovascular Research Center, at Pennsylvania Muscle Institute, working in the field of CNS regulation of blood flow/metabolism, ischemic/hypoxic disorders, stroke and chronic neurodegenerative disease animal models.
He was the Deputy R&D Director and Head of CNS Pharmacology Department at Biorex R&D Co., and worked at IVAX/Drug Research Institute Budapest, as a leading researcher in safety and CNS pharmacology and later at the IVAX/Drug Research Institute, a subsidiary of TEVA as Head of the In Vivo Pharmacology Group.
Currently, he is working with in vivo models of nano drug-nano carrier induced, complement activation related pseudoallergic reactions (CARPA), clarifying their immune-toxicological and safety hazards.
Acknowledgments
JS and UR acknowledge the financial support to the Nanomedicine Research and Education Center at Semmelweis University from Gedeon Richter NyRT and EU FP7 projects No: 309820 (NanoAthero) and 310337 (CosmoPhos). We also acknowledge the grants to SeroScience Ltd from EU FP7 projects No: 602923 (TheraGlio) and 281035 (TransInt).
References
1. Szebeni J. Complement activation-related pseudoallergy caused by liposomes, micellar carriers of intravenous drugs and radiocontrast agents. Crit Rev Ther Drug Carr Syst 2001;18:567–606.10.1615/CritRevTherDrugCarrierSyst.v18.i6.50Suche in Google Scholar
2. Szebeni J. Complement activation-related pseudoallergy: mechanism of anaphylactoid reactions to drug carriers and radiocontrast agents. In: Szebeni J, editor. The complement system: novel roles in health and disease. Boston: Kluwer, 2004: 399–440.Suche in Google Scholar
3. Szebeni J. Complement activation-related pseudoallergy: a new class of drug-induced immune toxicity. Toxicology 2005;216: 106–21.10.1016/j.tox.2005.07.023Suche in Google Scholar PubMed
4. Szebeni J, Alving CR, Rosivall L, Bunger R, Baranyi L, Bedocs P, et al. Animal models of complement-mediated hypersensitivity reactions to liposomes and other lipid-based nanoparticles. J Liposome Res 2007;17:107–17.10.1080/08982100701375118Suche in Google Scholar PubMed
5. Szebeni J, Muggia F, Gabizon A, Barenholz Y. Activation of complement by therapeutic liposomes and other lipid excipient-based therapeutic products: prediction and prevention. Adv Drug Deliv Rev 2011;63:1020–30.10.1016/j.addr.2011.06.017Suche in Google Scholar PubMed
6. Szebeni J. Hemocompatibility testing for nanomedicines and biologicals: predictive assays for complement mediated infusion reactions. Eur J Nanomed 2012;5:33–53.10.1515/ejnm-2012-0002Suche in Google Scholar
7. Szebeni J, Bedocs P, Csukas D, Rosivall L, Bunger R, Urbanics R. A porcine model of complement-mediated infusion reactions to drug carrier nanosystems and other medicines. Adv Drug Deliv Rev 2012;64:1706–16.10.1016/j.addr.2012.07.005Suche in Google Scholar PubMed
8. Szebeni J. Complement activation-related pseudoallergy: a stress reaction in blood triggered by nanomedicines and biologicals. Mol Immunol 2014;61:163–73.10.1016/j.molimm.2014.06.038Suche in Google Scholar PubMed
9. Szebeni J, Wassef NM, Spielberg H, Rudolph AS, Alving CR. Complement activation in rats by liposomes and liposome-encapsulated hemoglobin: evidence for anti-lipid antibodies and alternative pathway activation. Biochem Biophys Res Comm 1994;205:255–63.10.1006/bbrc.1994.2658Suche in Google Scholar PubMed
10. EMA-CHMP Reflection paper on the data requirements for intravenous liposomal products developed with reference to an innovator liposomal product. EMA/CHMP/806058/2009/Rev 02 2013;www.ema.europa.eu/docs/en_GB/document.../WC500140351.pdf.Suche in Google Scholar
11. Seelen MA, Roos A, Wieslander J, Mollnes TE, Sjoholm AG, Wurzner R, et al. Functional analysis of the classical, alternative, and MBL pathways of the complement system: standardization and validation of a simple ELISA. J Immunol Methods 2005;296:187–98.10.1016/j.jim.2004.11.016Suche in Google Scholar PubMed
12. Bergseth G, Ludviksen JK, Kirschfink M, Giclas PC, Nilsson B, Mollnes TE. An international serum standard for application in assays to detect human complement activation products. Mol Immunol 2013;56:232–9.10.1016/j.molimm.2013.05.221Suche in Google Scholar PubMed
13. Roos A, Wieslander J. Evaluation of complement function by ELISA. Methods Mol Biol 2014;1100:11–23.10.1007/978-1-62703-724-2_2Suche in Google Scholar
14. Ocmant A, Peignois Y, Mulier S, Hanssens L, Michils A, Schandené L. Flow cytometry for basophil activation markers: the measurement of CD203c up-regulation is as reliable as CD63 expression in the diagnosis of cat allergy. J Immunol Methods 2007;320:40–8.10.1016/j.jim.2006.12.002Suche in Google Scholar
15. Boumiza R, Debard AL, Monneret G. The basophil activation test by flow cytometry: recent developments in clinical studies, standardization and emerging perspectives. Clin Mol Allergy 2005;3:9.10.1186/1476-7961-3-9Suche in Google Scholar
16. Boumiza R, Monneret G, Forissier MF, Savoye J, Gutowski MC, Powell WS, et al. Marked improvement of the basophil activation test by detecting CD203c instead of CD63. Clin Exp Allergy 2003;33:259–65.10.1046/j.1365-2222.2003.01594.xSuche in Google Scholar
17. Uetrecht JP. New concepts in immunology relevant to idiosyncratic drug reactions: the “danger hypothesis” and innate immune system. Chem Res Toxicol 1999;12:387–95.10.1021/tx980249iSuche in Google Scholar
18. Choquet-Kastylevsky G, Descotes J. Value of animal models for predicting hypersensitivity reactions to medicinal products. Toxicology 1998;129:27–35.10.1016/S0300-483X(98)00060-2Suche in Google Scholar
19. Uetrecht J. Role of animal models in the study of drug-induced hypersensitivity reactions. AAPS J 2005;7:E914–21.10.1208/aapsj070489Suche in Google Scholar PubMed PubMed Central
20. Lin M, Sun W, Wang Y, Li X, Jin Y, Gong W, et al. An intravenous exposure mouse model for prediction of potential drug-sensitization using reporter antigens popliteal lymph node assay. J Appl Toxicol 2012;32:395–401.10.1002/jat.1696Suche in Google Scholar PubMed
21. Suda A, Iwaki Y, Kimura M. Differentiation of responses to allergenic and irritant compounds in mouse popliteal lymph node assay. J Toxicol Sci 2000;25:131–6.10.2131/jts.25.131Suche in Google Scholar PubMed
22. Shinkai K, Nakamura K, Tsutsui N, Kuninishi Y, Iwaki Y, Nishida H, et al. Mouse popliteal lymph node assay for assessment of allergic and autoimmunity-inducing potentials of low-molecular-weight drugs. J Toxicol Sci 1999;24:95–102.10.2131/jts.24.95Suche in Google Scholar PubMed
23. Gad SC. The mouse ear swelling test (MEST) in the 1990s. Toxicology 1994;93:33–46.10.1016/0300-483X(94)90194-5Suche in Google Scholar
24. Cornacoff JB, House RV, Dean JH. Mouse ear swelling test (MEST). Fundam Appl Toxicol 1992;19:157–8.10.1016/0272-0590(92)90040-OSuche in Google Scholar
25. Gad SC. Standard mouse ear swelling test (MEST). Fundam Appl Toxicol 1988;11:732–3.10.1016/0272-0590(88)90138-8Suche in Google Scholar
26. Kimber I, Hilton J, Botham PA. Identification of contact allergens using the murine local lymph node assay: comparisons with the Buehler occluded patch test in guinea pigs. J Appl Toxicol 1990;10:173–80.10.1002/jat.2550100306Suche in Google Scholar
27. Robinson MK, Nusair TL, Fletcher ER, Ritz HL. A review of the Buehler guinea pig skin sensitization test and its use in a risk assessment process for human skin sensitization. Toxicology 1990;61:91–107.10.1016/0300-483X(90)90012-6Suche in Google Scholar
28. Weaver JL, Staten D, Swann J, Armstrong G, Bates M, Hastings KL. Detection of systemic hypersensitivity to drugs using standard guinea pig assays. Toxicology 2003;193:203–17.10.1016/S0300-483X(03)00267-1Suche in Google Scholar
29. Rabinovici R, Rudolph AS, Vernick J, Feuerstein G. Lyophilized liposome-encapsulated hemoglobin: evaluation of hemodynamic, biochemical, and hematologic responses. Crit Care Med 1994;22:480–5.10.1097/00003246-199403000-00019Suche in Google Scholar
30. Rabinovici R, Rudolph AS, Ligler FS, Smith III EF, Feuerstein G. Biological responses to exchange transfusion with liposome-encapsulated hemoglobin. Circ Shock 1992;37:124–33.Suche in Google Scholar
31. Rabinovici R, Rudolph AS, Yue T-L, Feuerstein G. Biological responses to liposome-encapsulated hemoglobin (LEH) are improved by a PAF antagonist. Circ Shock 1990;31:431–45.Suche in Google Scholar
32. Dézsi L, Fülöp T, Mészáros T, Szénási G, Urbanics R, Vázsonyi C, et al. Features of complement activation-related pseudoallergy to liposomes with different surface charge and PEGylation: comparison of the porcine and rat responses. J Control Release 2014;195:2–10.10.1016/j.jconrel.2014.08.009Suche in Google Scholar PubMed
33. Dézsi L, Rosivall L, Hamar P, Szebeni J, Szénási G. Rodent models of complement activation-related pseudoallergy: inducers, symptoms, inhibitors and reaction mechanisms. Eur J Nanomedicine 2015;7:15–25.10.1515/ejnm-2015-0002Suche in Google Scholar
34. Qiu S, Liu Z, Hou L, Li Y, Wang J, Wang H, et al. Complement activation associated with polysorbate 80 in beagle dogs. Int Immunopharmacol 2013;15:144–9.10.1016/j.intimp.2012.10.021Suche in Google Scholar PubMed
35. Moghimi SM, Wibroe PP, Szebeni J, Hunter AC. Surfactant-mediated complement activation in beagle dogs. Int Immunopharmacol 2013;17:33–4.10.1016/j.intimp.2013.05.012Suche in Google Scholar
36. Summary JJ, Dubick MA, Zaucha GM, Kilani AF, Korte DW, Wade CE. Acute and subacute toxicity of 7.5% hypertonic saline/6% dextran-70 (HSD) in dogs. 1. Serum immunoglobulin and complement responses. J Appl Toxicol 1992;12:261–6.10.1002/jat.2550120409Suche in Google Scholar
37. Rossen RD, Michael LH, Kagiyama A, Savage HE, Hanson G, Reisberg MA, et al. Mechanism of complement activation after coronary artery occlusion: evidence that myocardial ischemia in dogs causes release of constituents of myocardial subcellular origin that complex with human C1q in vivo. Circ Res 1988;62:572–84.10.1161/01.RES.62.3.572Suche in Google Scholar
38. Awasthi VD, Garcia D, Goins BA, Phillips WT. Circulation and biodistribution profiles of long-circulating PEG-liposomes of various sizes in rabbits. Int J Pharm 2003;253:121–32.10.1016/S0378-5173(02)00703-2Suche in Google Scholar
39. Rudolph AS, Klipper RW, Goins B, Phillips WT. In vivo biodistribution of a radiolabeled blood substitute: 99mTc-labeled liposome-encapsulated hemoglobin in an anesthetized rabbit. Proc Natl Acad Sci USA 1991;88:10976–80.10.1073/pnas.88.23.10976Suche in Google Scholar
40. Acierno MJ, Labato MA, Stern LC, Mukherjee J, Jakowski RM, Ross LA. Serum concentrations of the third component of complement in healthy dogs and dogs with protein-losing nephropathy. Am J Vet Res 2006;67:1105–9.10.2460/ajvr.67.7.1105Suche in Google Scholar
41. Breathnach R, Donahy C, Jones BR, Bloomfield FJ. Increased leukotriene B(4) production, complement C3 conversion and acid hydrolase enzyme concentrations in different leucocyte sub-populations of dogs with atopic dermatitis. Vet J 2006;171:106–13.10.1016/j.tvjl.2004.09.012Suche in Google Scholar
42. Aganezov SA, Shcherbak IG, Galebskaia LV, Solovtsova IM, Riumina EV. [Activity of the complement system in blood serum of dogs after treatment of acute necrotic pancreatitis with boiled pancreatic juice]. Patologicheskaia fiziologiia i eksperimental’naia terapiia 1995:18–20.Suche in Google Scholar
43. Trail PA, Yang TJ, Cameron JA. Increase in the haemolytic complement activity of dogs affected with cyclic haematopoiesis. Vet Immunol Immunopathol 1984;7:359–68.10.1016/0165-2427(84)90093-XSuche in Google Scholar
44. Muller-Peddinghaus R, Schwartz-Porsche D. [Clinical significance of serum complement in dogs]. Zentralblatt fur Veterinarmedizin Reihe A 1983;30:698–711.Suche in Google Scholar
45. Imai T, Arii H, Sato T, Asakura Y, Sakuraya N, Sudo I, et al. [Change in complement titers of dogs under endotoxemia]. Masui Jpn J Anesthesiol 1979;28:564–9.Suche in Google Scholar
46. Tawara T, Hasegawa K, Sugiura Y, Harada K, Miura T, Hayashi S, et al. Complement activation plays a key role in antibody-induced infusion toxicity in monkeys and rats. J Immunol 2008;180:2294–8.10.4049/jimmunol.180.4.2294Suche in Google Scholar
47. Henry SP, Beattie G, Yeh G, Chappel A, Giclas P, Mortari A, et al. Complement activation is responsible for acute toxicities in rhesus monkeys treated with a phosphorothioate oligodeoxynucleotide. Int Immunopharmacol 2002;2: 1657–66.10.1016/S1567-5769(02)00142-XSuche in Google Scholar
48. Mugge A, Heistad DD, Densen P, Piegors DJ, Armstrong ML, Padgett RC, et al. Activation of leukocytes with complement C5a is associated with prostanoid-dependent constriction of large arteries in atherosclerotic monkeys in vivo. Atherosclerosis 1992;95:211–22.10.1016/0021-9150(92)90024-BSuche in Google Scholar
49. Hedin H, Smedegard G. Complement profiles in monkeys subjected to aggregate (immune complex) anaphylaxis, and following injection of soluble and particulate polysaccharides. Int Arch Allergy Appl Immunol 1979;60:286–94.10.1159/000232353Suche in Google Scholar PubMed
50. Szebeni J, Bedőcs P, Rozsnyay Z, Weiszhár Z, Urbanics R, Rosivall L, et al. Liposome-induced complement activation and related cardiopulmonary distress in pigs: factors promoting reactogenicity of Doxil and AmBisome. Nanomedicine NBM 2012;8:176–84.10.1016/j.nano.2011.06.003Suche in Google Scholar PubMed
51. Szebeni J, Baranyi L, Savay S, Bodo M, Milosevits J, Alving CR, et al. Complement activation-related cardiac anaphylaxis in pigs: role of C5a anaphylatoxin and adenosine in liposome-induced abnormalities in ECG and heart function. Am J Physiol Heart Circul Physiol 2006;290:H1050–8.10.1152/ajpheart.00622.2005Suche in Google Scholar PubMed
52. Bodo M, Szebeni J, Baranyi L, Savay S, Pearce F, Alving C, et al. Rheoencephalographic evidence of complement activation-related cerebrovascular changes in pigs. J Cerebr Blood Flow Met 2005;25:S550–S.10.1038/sj.jcbfm.9591524.0550Suche in Google Scholar
53. Szebeni J, Fontana JL, Wassef NM, Mongan PD, Morse DS, Dobbins DE, et al. Hemodynamic changes induced by liposomes and liposome-encapsulated hemoglobin in pigs: a model for pseudoallergic cardiopulmonary reactions to liposomes. Role of complement and inhibition by soluble CR1 and anti-C5a antibody. Circulation 1999;99:2302–9.10.1161/01.CIR.99.17.2302Suche in Google Scholar
54. Bedocs P, Capacchione J, Potts L, Chugani R, Weiszhar Z, Szebeni J, et al. Hypersensitivity reactions to intravenous lipid emulsion in swine: relevance for lipid resuscitation studies. Anesth Analg 2014;119:1094–101.10.1213/ANE.0000000000000396Suche in Google Scholar PubMed
55. Buckenmaier CC, Capacchione J, Mielke AR, Bina S, Shields C, Kwon KH, et al. The effect of lipid emulsion infusion on postmortem ropivacaine concentrations in swine: endeavoring to comprehend a soldier’s death. Anesth Analg 2012;114:894–900.10.1213/ANE.0b013e31822c10b8Suche in Google Scholar PubMed
56. Szebeni J, Baranyi B, Savay S, Lutz LU, Jelezarova E, Bunger R, et al. The role of complement activation in hypersensitivity to pegylated liposomal doxorubicin (Doxil®). J Liposome Res 2000;10:347–61.10.3109/08982100009031112Suche in Google Scholar
57. Swindle MM, Makin A, Herron AJ, Clubb FJ, Jr., Frazier KS. Swine as models in biomedical research and toxicology testing. Vet Pathol 2012;49:344–56.10.1177/0300985811402846Suche in Google Scholar PubMed
58. Schook L, Beattie C, Beever J, Donovan S, Jamison R, Zuckermann F, et al. Swine in biomedical research: creating the building blocks of animal models. Anim Biotechnol 2005;16:183–90.10.1080/10495390500265034Suche in Google Scholar PubMed
59. Azria M, Kiger JL. [Value of miniature swine in biomedical research]. Therapie 1972;27:723–32.Suche in Google Scholar
60. McClellan RO. Applications of swine in biomedical research. Lab Anim care 1968;18:120–6.Suche in Google Scholar
61. Bustad LK, McClellan RO. Swine in biomedical research. Science 1966;152:1526–30.10.1126/science.152.3728.1526Suche in Google Scholar PubMed
62. Elmadhun NY, Sabe AA, Robich MP, Chu LM, Lassaletta AD, Sellke FW. The pig as a valuable model for testing the effect of resveratrol to prevent cardiovascular disease. Ann NY Acad Sci 2013;1290:130–5.10.1111/nyas.12216Suche in Google Scholar PubMed
63. Gabler NK, Osrowska E, Imsic M, Eagling DR, Jois M, Tatham BG, et al. Dietary onion intake as part of a typical high fat diet improves indices of cardiovascular health using the mixed sex pig model. Plant Food Hum Nutr 2006;61:179–85.10.1007/s11130-006-0030-8Suche in Google Scholar PubMed
64. Lanoye L, Segers P, Tchana-Sato V, Rolin S, Dogne JM, Ghuysen A, et al. Cardiovascular haemodynamics and ventriculo-arterial coupling in an acute pig model of coronary ischaemia-reperfusion. Exp Physiol 2007;92:127–37.10.1113/expphysiol.2006.034983Suche in Google Scholar PubMed
65. Gross DR. Thromboembolic phenomena and the use of the pig as an appropriate animal model for research on cardiovascular devices. Int J Artif Organs 1997;20:195–203.10.1177/039139889702000402Suche in Google Scholar
66. Fletcher MP, Stahl GL, Longhurst JC. In vivo and in vitro assessment of porcine neutrophil activation responses to chemoattractants: flow cytometric evidence for the selective absence of formyl peptide receptors. J Leukocyte Biol 1990;47:355–65.10.1002/jlb.47.4.355Suche in Google Scholar PubMed
67. Suzuki Y, Yeung AC, Ikeno F. The representative porcine model for human cardiovascular disease. J Biomed Biotechnol 2011;2011:195483.10.1155/2011/195483Suche in Google Scholar PubMed PubMed Central
68. Couret D, de Bourmont S, Prat N, Cordier PY, Soureau JB, Lambert D, et al. A pig model for blunt chest trauma: no pulmonary edema in the early phase. Am J Emerg Med 2013;31: 1220–5.10.1016/j.ajem.2013.05.028Suche in Google Scholar PubMed
69. Kieser JA, Weller S, Swain MV, Neil Waddell J, Das R. Compressive rib fracture: peri-mortem and post-mortem trauma patterns in a pig model. Leg Med (Tokyo) 2013;15:193–201.10.1016/j.legalmed.2013.01.001Suche in Google Scholar PubMed
70. Byard RW, Cains GE, Gilbert JD. Use of a pig model to demonstrate vulnerability of major neck vessels to inflicted trauma from common household items. Am J Foren Med Path 2007;28:31–4.10.1097/01.paf.0000233530.18744.07Suche in Google Scholar
71. Zierold D, Perlstein J, Weidman ER, Wiedeman JE. Penetrating trauma to the diaphragm: natural history and ultrasonographic characteristics of untreated injury in a pig model. Arch Surg 2001;136:32–7.10.1001/archsurg.136.1.32Suche in Google Scholar
72. Sauer M, Altrichter J, Mencke T, Klohr S, Thomsen M, Kreutzer HJ, et al. Role of different replacement fluids during extracorporeal treatment in a pig model of sepsis. Ther Apher Dial 2013;17:84–92.10.1111/j.1744-9987.2012.01103.xSuche in Google Scholar
73. Sauer M, Altrichter J, Mencke T, Klohr S, Thomsen M, Kreutzer HJ, et al. Plasma separation by centrifugation and subsequent plasma filtration: impact on survival in a pig model of sepsis. Ther Apher Dial 2012;16:205–12.10.1111/j.1744-9987.2011.01055.xSuche in Google Scholar
74. Sauer M, Altrichter J, Kreutzer HJ, Logters T, Scholz M, Noldge-Schomburg G, et al. Extracorporeal cell therapy with granulocytes in a pig model of Gram-positive sepsis. Crit Care Med 2009;37:606–13.10.1097/CCM.0b013e318194aa77Suche in Google Scholar
75. Lobe TE, Woodall DL, Griffin MP. Early hemodynamic indicators of gram-negative sepsis and shock in an infant pig model. J Pediatr Surg 1991;26:1051–7.10.1016/0022-3468(91)90672-GSuche in Google Scholar
76. Dunn DL, Ferguson RM. Immunotherapy of gram-negative bacterial sepsis: enhanced survival in a guinea pig model by use of rabbit antiserum to Escherichia coli J5. Surgery 1982;92:212–9.Suche in Google Scholar
77. Szebeni J, Baranyi B, Savay S, Bodo M, Morse DS, Basta M, et al. Liposome-induced pulmonary hypertension: properties and mechanism of a complement-mediated pseudoallergic reaction. Am J Physiol 2000;279:H1319–28.10.1152/ajpheart.2000.279.3.H1319Suche in Google Scholar PubMed
78. Csukas D, Urbanics R, Weber G, Rosivall L, Szebeni J. Pulmonary intrvascular macrophages: prime suspects as cellular mediators of porcine CARPA. Eur J Nanomedicine 2015;7:27–36.10.1515/ejnm-2015-0008Suche in Google Scholar
79. Kuznetsova NR, Sevrin C, Lespineux D, Bovin NV, Vodovozova EL, Mészáros T, et al. Hemocompatibility of liposomes loaded with lipophilic prodrugs of methotrexate and melphalan in the lipid bilayer. J Control Release 2012;160:394–400.10.1016/j.jconrel.2011.12.010Suche in Google Scholar PubMed
80. Merkel OM, Urbanics R, Bedőcs P, Rozsnyay Z, Rosivall L, Toth M, et al. In vitro and in vivo complement activation and related anaphylactic effects associated with polyethylenimine and polyethylenimine-grafted-poly (ethylene glycol) block copolymers. Biomaterials 2011;32:4936–42.10.1016/j.biomaterials.2011.03.035Suche in Google Scholar PubMed
81. Szebeni J, Fontana J, Wassef N, Mongan P, Morse D, Stahl G, et al. Liposome-induced and complement-mediated cardiopulmonary distress in pigs as a model of pseudo-allergic reactions to liposomal drugs. Mol Immunol 1998;35:401.10.1016/S0161-5890(98)90814-8Suche in Google Scholar
82. Szebeni J, Bedőcs P, Urbanics R, Bunger R, Rosivall L, Tóth M, et al. Prevention of infusion reactions to PEGylated liposomal doxorubicin via tachyphylaxis induction by placebo vesicles: a porcine model. J Control Release 2012;160:382–7.10.1016/j.jconrel.2012.02.029Suche in Google Scholar PubMed
©2015 by De Gruyter
Artikel in diesem Heft
- Frontmatter
- In this issue
- News
- News from the European Foundation for Clinical Nanomedicine (CLINAM)
- What’s up in nanomedicine?
- Special Section Nanosafety
- Guest Editorial
- Sizing up the safety of nanomaterials
- Essay
- Keeping it small: towards a molecular definition of nanotoxicology
- Reviews
- The bio-corona and its impact on nanomaterial toxicity
- In vitro-ex vivo model systems for nanosafety assessment
- Meeting Report
- Nanosafety forum for young scientists: a meeting report
- Short Communication
- Nanomedicines in the European translational process
- Special Section CARPA Part 2
- Guest Editorial
- Complement activation-related pseudoallergy: an innate response to nanomedicines acting as pseudo-viruses
- Reviews
- Exosomes: potential model for complement-stealth delivery systems
- Lessons learned from the porcine CARPA model: constant and variable responses to different nanomedicines and administration protocols
- Mini Review
- Blood cell changes in complement activation-related pseudoallergy
- Short Communications
- Membrane attack complex formation on a supported lipid bilayer: initial steps towards a CARPA predictor nanodevice
- Paradoxical rise of hemolytic complement in the blood of mice during zymosan- and liposome-induced CARPA: a pilot study
- Commentary
- Insidious pathogen-mimicking properties of nanoparticles in triggering the lectin pathway of the complement system
Artikel in diesem Heft
- Frontmatter
- In this issue
- News
- News from the European Foundation for Clinical Nanomedicine (CLINAM)
- What’s up in nanomedicine?
- Special Section Nanosafety
- Guest Editorial
- Sizing up the safety of nanomaterials
- Essay
- Keeping it small: towards a molecular definition of nanotoxicology
- Reviews
- The bio-corona and its impact on nanomaterial toxicity
- In vitro-ex vivo model systems for nanosafety assessment
- Meeting Report
- Nanosafety forum for young scientists: a meeting report
- Short Communication
- Nanomedicines in the European translational process
- Special Section CARPA Part 2
- Guest Editorial
- Complement activation-related pseudoallergy: an innate response to nanomedicines acting as pseudo-viruses
- Reviews
- Exosomes: potential model for complement-stealth delivery systems
- Lessons learned from the porcine CARPA model: constant and variable responses to different nanomedicines and administration protocols
- Mini Review
- Blood cell changes in complement activation-related pseudoallergy
- Short Communications
- Membrane attack complex formation on a supported lipid bilayer: initial steps towards a CARPA predictor nanodevice
- Paradoxical rise of hemolytic complement in the blood of mice during zymosan- and liposome-induced CARPA: a pilot study
- Commentary
- Insidious pathogen-mimicking properties of nanoparticles in triggering the lectin pathway of the complement system