Membrane attack complex formation on a supported lipid bilayer: initial steps towards a CARPA predictor nanodevice
-
Saziye Yorulmaz
Saziye Yorulmaz received her Bachelor of Engineering degree in Materials Science and Engineering from Anadolu University (Turkey) in 2012. She was awarded a postgraduate scholarship from TEV-SINGA& A*STAR. Since 2013, she has been a postgraduate student in the Translational Science Group under the supervision of Prof. Nam Joon Cho at Nanyang Technological University (Singapore). Currently; her research project focuses on the design of protein-containing model lipid bilayer platforms for biological applications.
Seyed R Tabaei is currently a postdoctoral fellow at the Translational Science Group at Nanyang Technological University. Dr. Tabaei received a PhD in Bioscience from Chalmers University of Technology, Goteborg, Sweden in 2013. His research interests include lipid self-assembly and application of lipid-based platforms for membrane-protein interaction analysis and biomedical applications. He has published 16 original research papers.
Myung Hee Kim is currently a postdoctoral fellow at the Translational Science Group at Nanyang Technological University. Dr. Kim received a PhD in Biochemistry from ChungNam National University, DaeJeon, Korea in 2009. During her PhD studies, she focused on chemical genomics and bioactive small molecules from natural product. Currently, her research interests include drug development and liver tissue engineering using biomaterials.
Jeongeun Seo received B.E. and Ph.D. degrees in Chemistry from Hanyang University, Seoul, Korea, in 2008 and 2013, respectively. She joined the Translational Science Group at Nanyang Technological University, in 2013, where she is currently a postdoctoral fellow. Her research interests are three-dimensional network of carbon materials and their applications.
Walter Hunziker obtained his PhD from the Swiss Federal Institute of Technology in 1986 for the cloning of sucrase-isomaltase. His postdoctoral work at Yale University identified signals for basolateral sorting in epithelial cells. In 1992 he joined the Biochemistry Institute, University of Lausanne, as an Assistant Professor. In 2000 he moved to the Institute of Molecular and Cell Biology (IMCB), Singapore, where he currently is a Research Director and Deputy Director, with adjunct appointments at the National University of Singapore and the Singapore Eye Research Institute. Besides membrane traffic in epithelial cells, he is interested in the structure function relationship of tight junction proteins and how tight junctions regulating epithelial homeostasis.
János Szebeni MD, PhD, DSc, Med. Habil., immunologist, director of the Nanomedicine Research and Education Center at Semmelweis University, Hungary. He is also founder and CEO of a contract research SME “SeroScience”, and full professor of (immune) biology at Miskolc University. He has held various guest professor and scientific positions in Hungary and abroad, mostly in the USA where he lived for 22 years. His research on various themes in hematology, membrane biology and immunology resulted >120 scientific papers (citations: >4550, H index: 35), 14 book chapters, 2 granted patents, a book entitled “The Complement System: Novel Roles in Health and Disease” (Kluwer, 2004). Three fields stand out where he has been most active: artificial blood, liposomes and the complement system. His original works led to the “CARPA” concept, i.e., that complement activation underlies numerous drug-induced (pseudo)allergic (anaphylactoid) reactions.
 and
Nam-Joon Cho
and
Nam-Joon Cho
Nam-Joon Cho is Nanyang Associate Professor in the School of Materials Science and Engineering at Nanyang Technological University in Singapore and Deputy Director of the Nanyang Institute of Technology in Health and Medicine. In addition, he is a Principal Investigator at the Singapore-MIT Alliance for Research and Technology and Visiting Associate Professor at the Stanford University School of Medicine. His group’s research focuses on engineering approaches to solve important biomedical problems and to translate these capabilities into practical applications for global health. He also leads a multi-institution tissue engineering collaboration involving NTU, Singapore General Hospital, and the Stanford University School of Medicine, which focuses on developing an artificial liver platform for regenerative medicine applications. He is a graduate of Stanford University and the University of California, Berkeley.

Abstract
The rapid advance of nanomedicines and biologicals in pharmacotherapy gives increasing importance to a common adverse effect of these modern therapeutics: complement (C) activation-related pseudoallergy (CARPA). CARPA is a relatively frequent and potentially lethal acute immune toxicity of many intravenous drugs that contain nanoparticles or proteins, whose prediction by laboratory or in vivo testing has not yet been solved. Preliminary studies suggest that proneness of the drug to cause C activation in the blood of patients may predict the individual risk of CARPA, thus, a sensitive and rapid bedside assay for individualized assessment of a drug’s C activating potential could alleviate the CARPA problem. The goal of the present study was to lay down the foundations of a novel approach for real-time sensing of C activation on a supported lipid bilayer platform. We utilized the quartz crystal microbalance with dissipation (QCM-D) monitoring technique to measure the self-assembly of C terminal complex (or membrane attack complex [MAC]) on supported lipid bilayers rapidly assembled by the solvent-assisted lipid bilayer (SALB) formation method, as an immediate measure of C activation. By measuring the changes in frequency and energy dissipation of deposited protein, the technique allows extremely sensitive real-time quantification of the sequential assembly of MAC from its molecular components (C5b-6, C7, C8 and C9) and hence, measure C activation in the ambient medium. The present paper delineates the technique and our initial evidence with purified C proteins that the approach enables sensitive and rapid (real-time) quantification of MAC formation on a silicon-supported planar (phospho) lipid bilayer, which can be used as an endpoint in a clinically useful bedside C activation assay.
Introduction
Complement (C) activation-related pseudoallergy (CARPA) is an acute allergy syndrome triggered by intravenous (i.v.) injection or infusion of (nano)medicines, biologicals and many other drugs (1–8). The various aspects of this phenomenon, including the features of triggering drugs and agents, its symptoms, kinetics, occurrence, mechanism, prediction, prevention and many other aspects are amply detailed in earlier reviews (3–8), as well as in the present special papers on CARPA in the European Journal of Nanomedicine. To reiterate its clinical relevance; CARPA occasionally manifests itself in severe, potentially lethal anaphylactic reaction whose rise is unpredictable, prevention is not fully effective and treatment is nonspecific, empirical at this time. Apart from the medicinal aspects, the problem impacts the pharmaceutical industry as well, because severe reactions and deaths usually draw attention only in late phase clinical trials or in the post-marketing period of reactogenic drugs, at a stage where implementation of changes are not easy. Also, there are already a large number of “CARPA-genic” drugs in clinical use (3), and prevention of the reactions caused by those drugs has not been solved to-date.
The existing approaches of CARPA prediction and prevention include consideration of drug- and other allergies, the use of premedication protocols with massive doses of steroids, non-steroidal anti-inflammatory agents and antihistamines, and slow infusion of reactogenic drugs (9, 10). It is clear, however, that these approaches are neither fully effective, nor optimal. A previous clinical study has shown significant correlation between strong C activation in the blood of cancer patients treated with a reactogenic anticancer drug, Doxil, and the rise of CARPA in these patients (11). Furthermore, a study in the present issue suggests that proneness for C activation by the reactogenic drug or by zymosan in the blood of cancer patients tested in vitro correlates with the symptoms of CARPA in vivo. These findings taken together suggest that pretreatment measurements of C activation in the blood of patients might serve as a predictive biomarker for CARPA.
However, a practical obstacle of realizing this approach is the slow and labor intensive, complicated measurement of C activation today, mostly with ELISA. Thus, there is a yet unfulfilled need for individualized prediction of the risk of CARPA with a rapid bedside bioassay. Despite wide interest and substantial industrial R&D for such a testing kit or device, to our best knowledge, the idea has not been realized to date. One of the problems lies in the use of relatively slow ELISA-based quantitation of C cleavage products as measures of C activation. The present study was conducted to explore the possibility of an alternative approach: quantitation of C activation via real-time tracing of MAC formation on SiO2-supported planar bilayer membranes (12–17) as first step in the translation of this signal into a diagnostic platform.
Materials and methods
Reagents
Human complement proteins C5b-6, C7, C8, and C9 were purchased from Complement Tech (TX, USA). Table 1 provides background information about these proteins and the concentrations used in the experimental protocols of this study.
Complement proteins and concentrations used in the experiments.
| Protein | C5b-6 | C7 | C8 | C9 |
|---|---|---|---|---|
| Molecular weight, kDa | 285 | 92.40 | 151 | 71 |
| Stock concentration, mg/mL | 0.2 | 1 | 1 | 1 |
| Concentration, μg/mL | 5 | 5 | 5 | 60 |
| Mole, pmol | 17.54 | 54.11 | 33.11 | 845.07 |
| Molarity, nM | 17.54 | 54.11 | 33.11 | 845.07 |
| Isoelectric point | ND | 6.0–6.5 | 6.2–7.5 | 4.7 |
ND, not determined.
An aqueous buffer containing 10 mM Tris, 150 mM NaCl [pH 7.5] was used in all experiments. 1,2-dioleoyl-sn-glycero-3-phosphocholine (DOPC), 1,2-Dioleoyl-sn-glycero-3-ethylphosphocholine (chloride salt) (DOEPC), and 1-palmitoyl-2-oleoyl-sn-glycero-3-phospho-(1′-rac-glycerol) (sodium salt) (POPG) were purchased either in chloroform or powder form from Avanti Polar Lipids (Alabaster, AL, USA). Immediately before experiment, zwitterionic DOPC lipid powder was dissolved in isopropanol at the final concentration of 10 mg/mL. POPG and DOEPC lipid molecules were received in chloroform. First, the chloroform was evaporated with a stream of nitrogen air in order to create dried lipid films. Then, the dried lipid films were dissolved in appropriate organic solvent, e.g. ethanol or isopropanol. The negatively charged POPG lipids were dissolved to 5 mg/mL in ethanol at elevated temperature (at or above 40°C) in order to improve solubility. Prior to experiment, the desired lipid compositions were diluted to the experimental lipid concentration of 0.5 mg/mL.
Quartz crystal microbalance with dissipation (QCM-D) monitoring
QCM-D measurements were conducted using a Q-Sense E4 instrument equipped with flow modules (Q-Sense AB, Gothenburg, Sweden) in order to track the changes in resonance frequency (ΔF) and energy dissipation (ΔD) of an oscillating silicon oxide-coated quartz crystal. The crystal was excited at its fundamental resonance frequency of 5 MHz and odd overtones (n=3, 5, 7, and 11) in order to generate the thickness shear mode. The reported data were recorded at the third overtone and all the experiments were performed under continuous flow conditions at a flow rate of 100 μL/min for bilayer formation and 41.8 μL/min for injection of MAC components, as controlled by a peristaltic pump.
Results
Charged lipid bilayer formation by solvent-assisted lipid bilayer (SALB) formation method
To study the effect of membrane electrostatic surface charge on MAC formation, we created planar bilayer membrane platforms with different surface charges by way of tuning the lipid composition. The synthetic lipid molecules used in the experiments included zwitterionic DOPC, cationic DOEPC and anionic POPG lipid. More specifically, 30% molar fraction of POPG or DOEPC lipid were mixed with the remaining fraction of DOPC lipids in order to make negatively or positively charged lipid bilayer membranes, respectively. Experiments were also conducted with 100% DOPC lipid composition.
Figure 1B shows the representative QCM-D kinetic measurements for supported lipid bilayer formation on SiO2 by the SALB formation method. Initially, the aqueous buffer solution (10 mM Tris, 150 mM NaCl, pH 7.5) was injected into the QCM-D measurement cell (Step 1) in order to establish stable baseline signals for the frequency and energy dissipation responses. After establishing the baselines, isopropanol solution was added into the measurement cell (Step 2) which led to drastic changes in the frequency and energy dissipation signals during mixing isopropanol with buffer. During the mixing stage, the obtained dramatic changes mainly arise from the differences in density and viscosity of the buffer and isopropanol solutions, respectively. Afterwards, a freshly prepared 0.5 mg/mL lipid solution composed of DOPC or DOEPC:DOPC mixture (30:70) was dissolved in only isopropanol, or POPG:DOPC (30:70) dissolved in a mixture of ethanol and isopropanol was injected into the measurement cell (Step 3). There were small changes in frequency of around –6 Hz and no changes in energy dissipation during DOPC and DOEPC: DOPC (30:70) lipid deposition on SiO2. On the other hand, deposition of the POPG:DOPC (30:70) lipid mixture on SiO2 corresponded to a –4.7 Hz frequency shift and a 1.9×10–6 dissipation shift. In the final step, the isopropanol solution was gradually exchanged with buffer solution (Step 4) in order to form a complete planar lipid bilayer. Additionally, in order to check the completeness of the formed bilayer, 0.1 mg/mL BSA protein was finally added to detect nonspecific adhesion (Step 5). There was no significant change due to BSA protein adsorption in all cases, suggesting that high-quality lipid bilayers were formed in order to investigate MAC assembly.
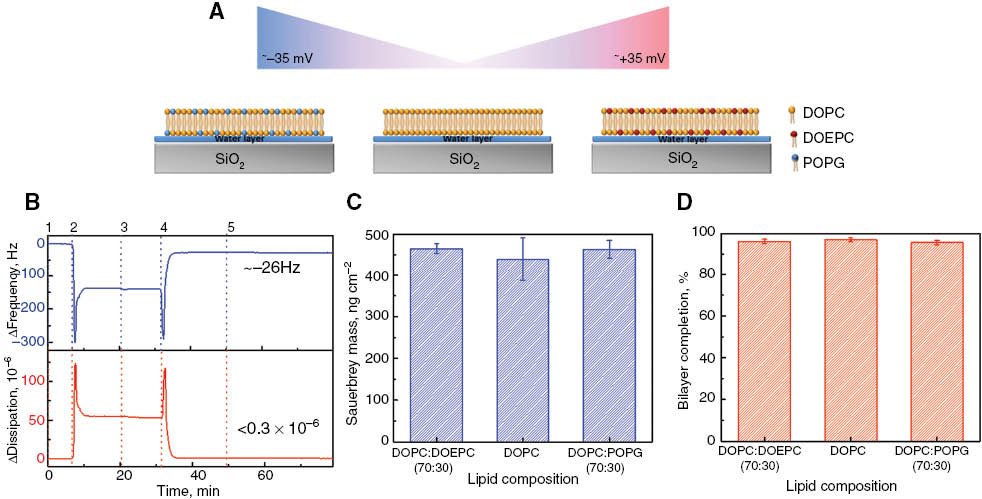
Design of charged supported lipid bilayer platforms by the SALB formation method. QCM-D monitoring of the supported lipid bilayer formed on SiO2 by the SALB formation method. (A) The schematic display charged lipids used to form a planar bilayer as follows: Left) DOPC:POPG (70:30), Middle) DOPC Right) DOPC:DOEPC (70:30). Color key: yellow: DOPC; red: DOEPC; blue: POPG. (B) Changes in frequency (ΔF, blue) and energy dissipation (ΔD, red) were recorded as a function of time at the third overtone, n=3. QCM-D signatures of planar bilayer formation by the SALB formation method were obtained by the following experimental sequence: (Step 1) Add Tris buffer solution; (Step 2) Inject isopropanol solution; (Step 3) Add 0.5 mg/mL lipid molecules in isopropanol; (Step 4) Exchange isopropanol to Tris buffer solution; (Step 5) add BSA protein (0.1 mg/mL) to check the completeness of the bilayers. (C) Adsorbed lipid mass surface density was calculated by the Sauerbrey equation. (D) Percentage of bilayer completion was calculated based on BSA adsorption to bare SiO2 substrate (control) and the bilayer after completion of the SALB procedure.
As quantitative measures, the changes in final frequency and dissipation values after bilayer formation and BSA adsorption are summarized in Table 2. The obtained final frequency and dissipation values are consistent with the literature (16–18). For rigid films such as a planar lipid bilayer, the sauerbrey equation is used to calculate the adsorbed mass on the substrate based on frequency shifts (19). The attached mass on SiO2 was approximately 450 ng/cm2 after bilayer formation and was independent of lipid composition as shown in Figure 1C. In order to quantify the bilayer completion depending on lipid composition, the frequency shifts obtained by BSA adsorption on bare SiO2 are normalized according the frequency shifts obtained by BSA adsorption on a planar lipid bilayer. The calculation indicated that bilayer completion was more than ∼97% complete irrespective of the lipid composition used to form the bilayer as shown in Figure 1D.
QCM-D measurement responses obtained for various SALB platforms and their nonspecific interactions with BSA protein.
| Platforms | Bilayer | BSA | ||
|---|---|---|---|---|
| ΔF(Hz) | ΔD(10−6) | ΔF(Hz) | ΔD(10−6) | |
| DOPC | 24.79±2.924 | 0.04±0.031 | 25.45±0.5 | 0.83±0.2 |
| DOPC:DOEPC (70:30) | 26.28±0.692 | 0.49±0.002 | 26.91±0.95 | 0.59±0.05 |
| DOPC:POPG (70:30) | 26.16±1.20 | 0.55±0.18 | 27.25±1.76 | 0.71±0.35 |
Initial strategy to form MAC assembly on SLBs
We first investigate the effect of membrane surface charge on MAC self-assembly on the planar lipid bilayer. The QCM-D signatures of the self-assembly of MAC components from the sequential addition of C5b-7, C8 and C9 proteins on three different types of lipid bilayer membranes are presented in Figure 2A and 2B. The initial measurement baseline was recorded during the validation of lipid bilayer quality via BSA adsorption and then normalized. Immediately before the injection of C5b-7 precomplex, 5 μg/mL of C5b-6 and 5 μg/mL of C7 protein precursors were incubated for 30 min in order to generate the C5b-7 precomplexed adduct at room temperature. Once the obtained precomplex was added onto the zwitterionic (DOPC), positively (DOEPC: DOPC, 30:70) and negatively charged (POPG: DOPC, 30:70) bilayers (Figure 2, arrow 1), changes in frequency and dissipation values were recorded as a function of time. The QCM-D signatures varied depending on the lipid compositions. Adsorption of the C5b-7 precomplex on the negatively charged SALB resulted in frequency and dissipation values of –16.2±2.6 Hz and 2.3±0.5×10–6, respectively. However, in the case of the positive charged lipid bilayer, the changes in frequency and dissipation values were –3.4±2.2 Hz and 0.2±0.2×10–6, respectively, whereas addition of the C5b-7 precomplex onto zwitterionic lipid bilayer membranes resulted in almost no binding based on the frequency and energy dissipation values. It has been previously reported that C5b-7 interacts with only negatively charged liposomes through a hydrophobic interaction (20). Therefore, our find findings are consistent with the scientific literature.
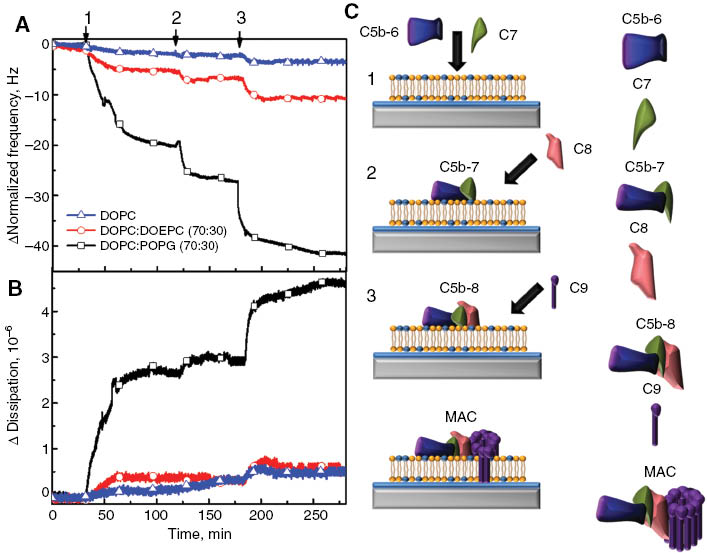
Assembly of MAC components on supported lipid bilayer platforms. MAC formation is initiated by the incubation of hydrophilic C5b-6 and C7 precursors (1). During the incubation of these two protein precursors, C7 interacts with C5b-6 and causes a hydrophilic-amphiphilic transition to result in precomplex C5b-7 formation and its membrane association. Addition of C8 proteins to membrane-bound C5b-7 precomplex leads both formation of the C5b-8 intermediate complex and its insertion into the hydrophobic core of the membrane (2). Subsequent interaction of C9 (3) with the membrane-inserted C5b-8 intermediate complex causes significant changes in C9 protein conformation and this makes 12–18 copies of C9 polymerize circularly to generate the final MAC complex. Using QCM-D tracking, changes in frequency (A) and dissipation (B) were monitored in order to validate MAC formation as a function of time. Three different kinds of SLB platforms composed of DOPC: POPG (70:30) (black square); DOPC: DOEPC (70:30) (red circle) and DOPC (blue triangle) were used to determine the effects of membrane surface charge on MAC formation. After completion of the SALB procedure and validation of bilayer completeness, the previously incubated C5b-6 and C7 proteins were added in the form of a precomplex C5b-7 to the SLB platform. Then, C8 and C9 were added to the membrane-associated C5b-7 precomplex, respectively. (C) Schematic representation of MAC formation on the negatively charged SALB platform.
After association of the C5b-7 precomplex into the lipid bilayers, 5 μg/mL of C8 protein was added onto the C5b-7 precomplex containing lipid bilayers (Figure 2, arrow 2). The variations in frequency and dissipation responses depended on the lipid composition, reflecting C5b-7 precomplex association. On negatively charged lipid bilayer membranes, the frequency and dissipation shifts due to the presence of C8 proteins were –6.1±1.0 Hz and 0.05±0.20×10–6, respectively. Likewise, the frequency and dissipation shifts were negligible for both the positively charged and zwitterionic lipid bilayers (<2 Hz and ∼0.5×10–6). The interaction between the C5b-7 precomplex and C8 leads to formation of a C5b-8 intermediate complex on negatively charged lipid bilayers (21). This interaction is crucial for further association of C9 proteins in order to make the MAC self-assembly.
Finally, 60 μg/mL C9 protein – the final MAC component – was injected to the membrane bound C5b-8 intermediate complex (Figure 2, arrow 3). The absolute frequency and dissipation shifts on the negatively charged bilayer were –11.0±2.3 and 1.6±0.4×10–6, respectively. However, there were no significant changes in frequency and dissipation responses obtained from the assembly of C9 proteins on both zwitterionic and positively charged lipid bilayers which is quite similar to the changes obtained from the association of the C5b-7 precomplex and C8 proteins. These small changes in frequency and dissipation of both positively charged and zwitterionic lipid bilayer membranes were caused by non-specific interactions between protein and the bilayer. The detailed schematic of MAC formation is presented in Figure 2C. The changes in absolute frequency, energy dissipation, and mass binding responses during the self-assembly of MAC components are presented in Table 3. As a result, the obtained frequency and dissipation shifts were quite appreciable on negatively charged lipid bilayers. Therefore, negatively charged lipid bilayers were utilized for further studies in order to investigate the molecular assembly of MAC components.
QCM-D measurement responses obtained for the addition of individual MAC components on the SLB platform.
| Platforms | C5b-7 | C8 | C9 | |||
|---|---|---|---|---|---|---|
| ΔF(Hz) | ΔD(10–6) | ΔF(Hz) | ΔD(10–6) | ΔF(Hz) | ΔD(10–6) | |
| DOPC | 1.52 | 0.077 | 0.39 | 0.00023 | 1.35 | 0.0972 |
| DOPC:DOEPC (70:30) | 3.44±2.16 | 0.20±0.21 | 1.62±0.60 | 0.33±0.188 | 3.11±0.46 | 0.11±0.04 |
| DOPC:POPG (70:30) | 16.21±2.59 | 2.34±0.49 | 6.17±0.96 | 0.049±0.202 | 10.95±2.27 | 1.55±0.37 |
The changes in final frequency and dissipation values were reported after the addition of individual MAC components in the order of C5b-7, C8 and C9, respectively. The C5b-7 precomplex was formed during a 30 min incubation of MAC initial precursors of C5b-6 and C7. The average and standard deviation of each measurement value are reported for three independent experiments.
C5b-6 and C7 in MAC assembly
Individual C5b-6 and C7 protein precursors and the C5b-7 precomplex were utilized in order to further investigate the preferential association of these proteins on negatively charged lipid bilayers in order to initiate MAC self-assembly formation as shown in Figure 3. First of all, after formation of the bilayer, 5 μg/mL of C5b-6 protein was added to the negatively charged lipid bilayer as shown in Figures 3A and 3B. Adsorption of C5b-6 onto the bilayer did not induce remarkable variations in both frequency and dissipation values. The association of C5b-6 to the cell surface is still unclear to date (22, 23). It has been reported that C5b-6 loosely interacts with the cell membrane but the specific membrane molecules modulating this interaction are still unknown (22). However, Silversmith et al. has reported that the composition of lipid vesicles determines the interaction of C5b-6 and C5b-7 complexes with liposomes based on light scattering intensity measurements (23, 24). It was observed that liposomes formed by 100% phosphatidic acid (PA) or phosphatidylglycerol (PG) interacted with C5b-6, unlike liposomes composed phosphatidylcholine (PC), phosphatidylserine (PS), or phosphatidylinositol (PI) (23).
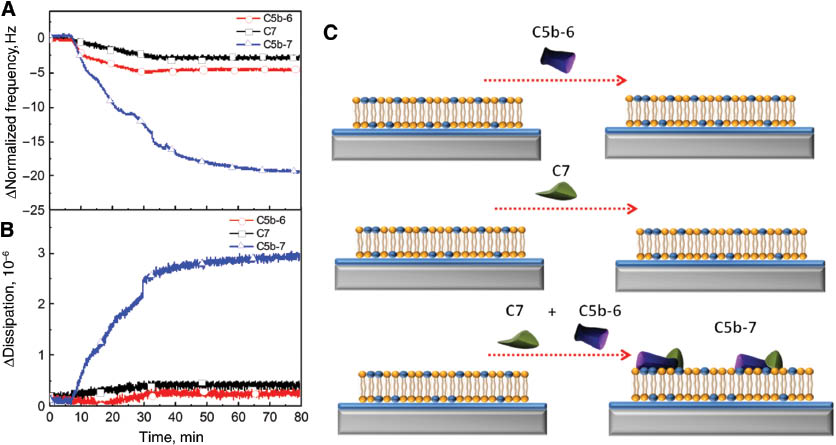
Complement components C5b-6 and C7 create a C5b-7 precomplex which associates on the negatively charged lipid bilayer in order to initiate MAC formation. Changes in normalized frequency (A) and dissipation (B) are presented as a function of time. The initial MAC precursors including C5b-6 (red circle), C7 (black square), and C5b-7 (blue up triangle) were individually injected onto the negatively charged bilayers after a stable baseline obtained from BSA addition. C5b-7 was formed at the end of a 30 min incubation period of C5b-6 and C7. (C) Schematic diagram depicts the interaction of initial MAC precursors with the negatively charged bilayer.
When C7 protein was added onto the negatively charged lipid bilayer, there were only minor changes, with frequency and energy dissipation shifts of only –2.5 Hz and less than 0.5×10–6, respectively. However, when C5b-6 interacts with C7, the C5b-7 precomplex spontaneously transferred from the solution to the targeted negatively charged SALB where it attaches to the lipid bilayer. Therefore, the frequency values increased almost 10-fold while changes in energy dissipation increased almost 6-fold compared to the individual association of C5b-6 and C7 proteins. The reported mechanism for interaction of the C5b-7 precomplex with the negatively charged lipid bilayer is the presence of an irreversible hydrophilic-amphiphilic transition during the interaction between C5b-6 and C7 protein precursors (21). In other words, a conformational alteration occurs due to binding of C7 to C5b-6 which causes the exposure of an already existing internal hydrophobic domain with the surrounding environment. This alteration creates a bilayer targeting site in the C5b-7 precomplex which appears to involve the C7 protein even though the binding of individual C7 protein to negatively charged lipid bilayers is negligible (25). In terms of affinity to the negatively charged lipid bilayer surface, it has been shown that the deposition level of the C5b-7 precomplex can also be enhanced by the presence of two sialylated molecules of erythrocyte membranes such as gangliosides and glycoporin through ionic interactions (26).
C8 in MAC assembly
The preferential association of C8 protein in the presence and absence of the C5b-7 complex on the lipid bilayer membrane surface was examined in order to demonstrate the formation of the C5b-8 intermediate complex as shown in Figure 4. Once 5 μg of C8 protein was added to the bare lipid bilayer and the C5b-7 precomplex-containing lipid bilayer, the changes in frequency and dissipation responses were quite comparable (Figure 4A, B). The interaction of C8 with the bare lipid bilayer led to small changes in final frequency and dissipation responses (–5.1 Hz and 0.09×10–6, respectively). In the case of the presence of the C5b-7 precomplex on the lipid bilayer, the variations in frequency and dissipation responses were slightly different than in the absence of the C5b-7 precomplex. They were –6.8 Hz and 0.5×10–6, respectively. It can be inferred that the changes in QCM-D responses were independent of the presence of the C5b-7 precomplex on the lipid bilayer. It has reported that the positively charged C8 protein interacts with the negatively charged C5b-7 precomplex. In other words, the interaction between C8 and the C5b-7 precomplex is driven by electrostatic interactions (27). Therefore, it is not possible to confirm the formation of the C5b-8 intermediate complex based on obtained results because C8 would interact with the C5b-7 precomplex or the lipid bilayer itself as depicted in the schematic in Figure 4C. In order to prove the existence of C5b-8 precomplex, further investigation is necessary using C9 proteins.
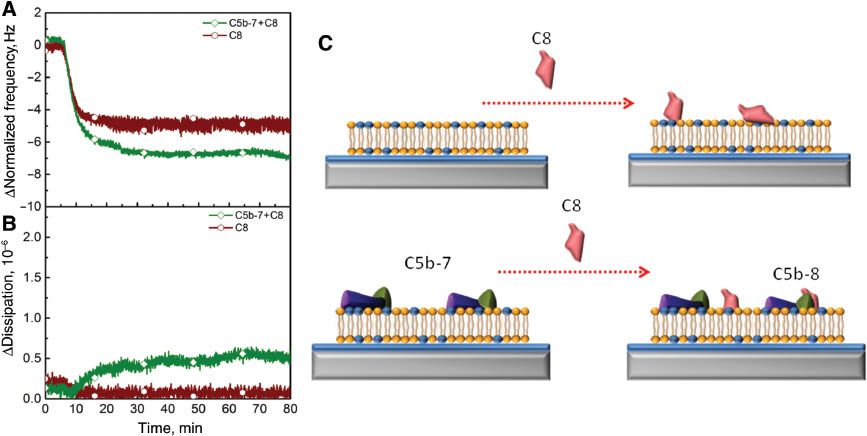
Electrostatic interaction is essential for the activity of C8 proteins regardless of the presence of the precomplex C5b-7. The changes in frequency (A) and energy dissipation (B) are tracked as a function of time. C8 protein was added right after stabilization of BSA (wine hexagon) and association of the C5b-7 precomplex (green tetragon). The binding of C8 to membrane-associated C5b-7 precomplex creates a C5b-8 complex. (C) Schematic diagram presents the interaction of C8 proteins with a bilayer in the presence or absence of the C5b-7 precomplex.
Transformation of C9 monomer to MAC self-assembly complex
As a final test, 60 μg/mL of C9 protein was added to the negatively charged lipid bilayer in order to investigate the association of C9 in the presence and absence of C5b-8 intermediate complex. The changes in frequency and dissipation shifts are presented in Figure 5A,B. In the absence of all MAC precursor proteins, C9 proteins interacted non-specifically with the lipid bilayers so the changes in frequency and energy shifts were negligible and the obtained frequency and dissipation shifts were –3.1 Hz and 0.1×10–6, respectively. These small changes indicated that the nonspecific interaction of C9 did not affect the viscoelastic properties and integrity of the lipid bilayers. However, when the same concentration of C9 proteins was injected into the lipid bilayer which contains the C5b-8 intermediate complex, the variations in frequency and dissipation responses were about –14.0 Hz and 1.7×10–6, respectively. These appreciable changes designate the importance of the C5b-8 intermediate complex for association of C9 proteins. For example, the light scattering measurements were previously performed and the obtained light scattering intensity data showed that C9 did not interact with lipid vesicles unless the C5b-8 intermediate complex was present on the lipid vesicles (23). In addition, C9 primarily interacts with C8 within the C5b-8 intermediate complex and this creates a C9 polymerization site which is essential for further MAC formation. 12–18 copies of C9 are necessary to polymerize circularly to create the MAC (28, 29). If the concentration of C9 is lower, then C5b-9 forms due to the fact that the C9 polymerization stops before ring closure occurs (30). The final MAC formation schematic is presented in Figure 5C. The presence of the C5b-8 intermediate complex on the lipid bilayer decreases the time required for C9 polymerization because it lowers the activation energy required for spontaneous polymerization of C9 (31, 32). During this process, the hydrophilic-amphiphilic transition occurs from the monomeric C9 to polymeric C9 complex which is able to insert the whole polymeric C9 complex itself into the hydrophobic core of the lipid bilayer. Based on our results, the initial association of C9 proteins to C5b-8 bound membrane occurred within 10 min, which might be preliminary evidence of C9 polymerization.
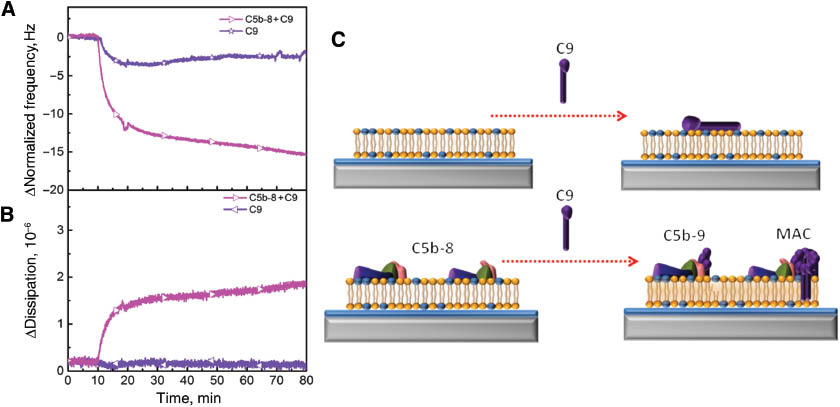
Polymerization of C9 proteins depends on the existence of the membrane-inserted C5b-8 complex in order to form MAC. The changes in frequency (A) and energy dissipation (B) are presented as a function of time. C9 proteins were injected after the stabilization of BSA (purple star) and association of C5b-8 complex (pink left triangle). (C) Schematic representation demonstrates the interaction of C9 with a bilayer and membrane inserted C5b-8.
Discussion
Supported lipid bilayers constitute a two-dimensional membrane-mimicking surface that is compatible with a wide range of surface-sensitive measurement techniques, including acoustic and optical biosensors of various kinds (12, 13). Supported lipid bilayers can be readily formed on silica-based materials by vesicle fusion (14) or solvent-assisted lipid bilayer (16) (SALB) formation methods, and the chemical composition of the bilayer can be adjusted, including electrical surface charge, membrane fluidity, and cholesterol content. For vesicle fusion, a variety of experimental conditions, including vesicle size (15), lipid composition (33), solution pH (34), osmotic pressure (35), and freeze-thaw pretreatment (36) need to be optimized for reproducible bilayer fabrication which is difficult for non-expert users. On the other hand, the SALB process is a simpler and more versatile approach that involves lipid deposition in alcohol, followed by solvent-exchange with aqueous buffer. Its optimization is straightforward and supported lipid bilayers can be formed on a wide range of substrates, including gold, aluminum oxide, and titanium oxide in addition to silicon oxide (16, 37–40). Straightforward operation and high reproducibility lends strong potential for SALB fabrication approaches to serve as the basis for scale-up production of supported lipid bilayer platforms for clinical applications.
Importantly, in conjunction with label-free biosensing measurements, supported lipid bilayers enable quantitative kinetic tracking of molecular processes occurring on membrane interfaces. The quartz crystal microbalance with dissipation (QCM-D) monitoring technique has demonstrated significant promise for bioanalytical measurements by simultaneously recording changes in resonance frequency and energy dissipation of an oscillating quartz crystal due to a mass adsorbate (41). The concept has been extended to molecular diagnostics in biological fluids, motivating the exploration of this measurement technique towards clinical applications, including high-sensitivity predictive detection of C activation.
Outlook
Herein, as an initial effort to design a CARPA predictor nanodevice, we demonstrated the assembly of MAC on silicon-oxide-supported, negatively charged lipid bilayer by the SALB formation method. By utilizing an acoustic sensor device, we were able to monitor MAC formation in real-time, thus obviating the need for ELISA measurement of C scission products as an endpoint of C activation. By doing so, we provided a promising new approach for real-time monitoring of C activation, one of the desired basic features of a potential bedside CARPA predictor nanodevice. Obviously however, intense further R&D will be needed to translate this advance into a clinically useful device which will differentiate between plasma proteins for selective response to MAC deposition only. An important question remains to verify whether detection of MAC formation requires sequential addition of C proteins or the simultaneous deposition of the implicated proteins is possible for more rapid measurement readout. Progress in this direction is currently underway in our laboratory.
About the authors

Saziye Yorulmaz received her Bachelor of Engineering degree in Materials Science and Engineering from Anadolu University (Turkey) in 2012. She was awarded a postgraduate scholarship from TEV-SINGA& A*STAR. Since 2013, she has been a postgraduate student in the Translational Science Group under the supervision of Prof. Nam Joon Cho at Nanyang Technological University (Singapore). Currently; her research project focuses on the design of protein-containing model lipid bilayer platforms for biological applications.

Seyed R Tabaei is currently a postdoctoral fellow at the Translational Science Group at Nanyang Technological University. Dr. Tabaei received a PhD in Bioscience from Chalmers University of Technology, Goteborg, Sweden in 2013. His research interests include lipid self-assembly and application of lipid-based platforms for membrane-protein interaction analysis and biomedical applications. He has published 16 original research papers.

Myung Hee Kim is currently a postdoctoral fellow at the Translational Science Group at Nanyang Technological University. Dr. Kim received a PhD in Biochemistry from ChungNam National University, DaeJeon, Korea in 2009. During her PhD studies, she focused on chemical genomics and bioactive small molecules from natural product. Currently, her research interests include drug development and liver tissue engineering using biomaterials.

Jeongeun Seo received B.E. and Ph.D. degrees in Chemistry from Hanyang University, Seoul, Korea, in 2008 and 2013, respectively. She joined the Translational Science Group at Nanyang Technological University, in 2013, where she is currently a postdoctoral fellow. Her research interests are three-dimensional network of carbon materials and their applications.

Walter Hunziker obtained his PhD from the Swiss Federal Institute of Technology in 1986 for the cloning of sucrase-isomaltase. His postdoctoral work at Yale University identified signals for basolateral sorting in epithelial cells. In 1992 he joined the Biochemistry Institute, University of Lausanne, as an Assistant Professor. In 2000 he moved to the Institute of Molecular and Cell Biology (IMCB), Singapore, where he currently is a Research Director and Deputy Director, with adjunct appointments at the National University of Singapore and the Singapore Eye Research Institute. Besides membrane traffic in epithelial cells, he is interested in the structure function relationship of tight junction proteins and how tight junctions regulating epithelial homeostasis.

János Szebeni MD, PhD, DSc, Med. Habil., immunologist, director of the Nanomedicine Research and Education Center at Semmelweis University, Hungary. He is also founder and CEO of a contract research SME “SeroScience”, and full professor of (immune) biology at Miskolc University. He has held various guest professor and scientific positions in Hungary and abroad, mostly in the USA where he lived for 22 years. His research on various themes in hematology, membrane biology and immunology resulted >120 scientific papers (citations: >4550, H index: 35), 14 book chapters, 2 granted patents, a book entitled “The Complement System: Novel Roles in Health and Disease” (Kluwer, 2004). Three fields stand out where he has been most active: artificial blood, liposomes and the complement system. His original works led to the “CARPA” concept, i.e., that complement activation underlies numerous drug-induced (pseudo)allergic (anaphylactoid) reactions.

Nam-Joon Cho is Nanyang Associate Professor in the School of Materials Science and Engineering at Nanyang Technological University in Singapore and Deputy Director of the Nanyang Institute of Technology in Health and Medicine. In addition, he is a Principal Investigator at the Singapore-MIT Alliance for Research and Technology and Visiting Associate Professor at the Stanford University School of Medicine. His group’s research focuses on engineering approaches to solve important biomedical problems and to translate these capabilities into practical applications for global health. He also leads a multi-institution tissue engineering collaboration involving NTU, Singapore General Hospital, and the Stanford University School of Medicine, which focuses on developing an artificial liver platform for regenerative medicine applications. He is a graduate of Stanford University and the University of California, Berkeley.
Acknowledgments
This work was supported by the National Research Foundation (NRF-NRFF2011-01) and the National Medical Research Council (NMRC/CBRG/0005/2012). J.S. is a recipient of the Tan Chin Tuan Exchange Fellowship at Nanyang Technological University.
References
1. Cheifetz A, Mayer L. Monoclonal antibodies, immunogenicity, and associated infusion reactions. Mt Sinai J Med 2005;72:250–6.Search in Google Scholar
2. Maggi E, Vultaggio A, Matucci A. Acute infusion reactions induced by monoclonal antibody therapy. Expert Rev Clin Immunol 2011;7:55–63.10.1586/eci.10.90Search in Google Scholar PubMed
3. Szebeni J. Complement activation-related pseudoallergy: a stress reaction in blood triggered by nanomedicines and biologicals. Mol Immunol 2014;61:163–73.10.1016/j.molimm.2014.06.038Search in Google Scholar PubMed
4. Szebeni J. Hemocompatibility testing for nanomedicines and biologicals: predictive assays for complement mediated infusion reactions. Eur J Nanomed 2012;5:33–53.10.1515/ejnm-2012-0002Search in Google Scholar
5. Szebeni J, Muggia F, Gabizon G, Barenholz Y. Activation of complement by therapeutic liposomes and other lipid excipient-based therapeutic products: Prediction and prevention. Advanced Drug Delivery Rev 2011;63:1020–30.10.1016/j.addr.2011.06.017Search in Google Scholar PubMed
6. Szebeni J, Alving CR, Baranyi L, Bunger R. Interactions of liposomes with complement leading to adverse reactions. Liposome technology, Volume III: Interactions of Liposomes with the Biological Milieu 2006;3:1.Search in Google Scholar
7. Szebeni J. Complement activation-related pseudoallergy: a new class of drug-induced immune toxicity. Toxicology 2005;216:106–21.10.1016/j.tox.2005.07.023Search in Google Scholar PubMed
8. Szebeni J. Complement activation-related pseudoallergy: Mechanism of anaphylactoid reactions to drug carriers and radiocontrast agents. In: Szebeni J, editor. The Complement System: Novel Roles in Health and Disease. Boston: Kluwer; 2004. p. 399–440.10.1007/1-4020-8056-5_17Search in Google Scholar
9. Mayer L, Young Y. Infusion reactions and their management. GastroenterolClinNorth Am. 2006;35:857–66.10.1016/j.gtc.2006.09.006Search in Google Scholar PubMed
10. Vogel WH. Infusion reactions: diagnosis, assessment, and management. Clin J Oncol Nursing 2010;14:E10–21.10.1188/10.CJON.E10-E21Search in Google Scholar PubMed
11. Chanan-Khan A, Szebeni J, Savay S, Liebes L, Rafique NM, Alving CR, Complement activation following first exposure to pegylated liposomal doxorubicin (Doxil): possible role in hypersensitivity reactions. Ann Oncol 2003;14:1430–7.10.1093/annonc/mdg374Search in Google Scholar PubMed
12. Sackmann, E. Supported membranes: scientific and practical applications. Science 1996;271:43–8.10.1126/science.271.5245.43Search in Google Scholar PubMed
13. Jackman JA, Cho N-J. Model membrane platforms for biomedicine: case study on antiviral drug development. Biointerphases 2012;7:18.10.1007/s13758-011-0018-2Search in Google Scholar
14. Tamm LK, McConnell HM. Supported phospholipid bilayers. Biophys J 1985;47:105.10.1016/S0006-3495(85)83882-0Search in Google Scholar
15. Reimhult E, Höök F, Kasemo B. Intact vesicle adsorption and supported biomembrane formation from vesicles in solution: influence of surface chemistry, vesicle size, temperature, and osmotic pressure. Langmuir 2003;19:1681–91.10.1021/la0263920Search in Google Scholar
16. Tabaei SR, Choi J-H, Haw Zan G, Zhdanov VP, Cho N-J. Solvent-assisted lipid bilayer formation on silicon dioxide and gold. Langmuir 2014;30:10363–73.10.1021/la501534fSearch in Google Scholar
17. Zan GH, Jackman JA, Cho N-J. AH peptide-mediated formation of charged planar lipid bilayers. J Phys Chem B 2014;118:3616–21.10.1021/jp411648sSearch in Google Scholar
18. Keller C, Kasemo B. Surface specific kinetics of lipid vesicle adsorption measured with a quartz crystal microbalance. Biophys J 1998;75:1397–402.10.1016/S0006-3495(98)74057-3Search in Google Scholar
19. Sauerbrey G. Verwendung von Schwingquarzen zur Wägung dünner Schichten und zur Mikrowägung. Zeitschrift für physik 1959;155:206–22.10.1007/BF01337937Search in Google Scholar
20. Liu C, Marshall P, Schreibman I, Vu A, Gai W, Whitlow M. Interaction between terminal complement proteins C5b-7 and anionic phospholipids. Blood 1999;93:2297–301.10.1182/blood.V93.7.2297Search in Google Scholar
21. Podack ER, Esser AF, Biesecker G, Müller-Eberhard H. Membrane attack complex of complement: a structural analysis of its assembly. J Exp Med 1980;151:301–13.10.1084/jem.151.2.301Search in Google Scholar PubMed PubMed Central
22. Hu VW, Esser A, Podack E, Wisnieski B. The membrane attack mechanism of complement: photolabeling reveals insertion of terminal proteins into target membrane. J Immunol 1981;127:380–6.10.4049/jimmunol.127.1.380Search in Google Scholar
23. Silversmith RE, Nelsestuen GL. Interaction of complement proteins C5b-6 and C5b-7 with phospholipid vesicles: effects of phospholipid structural features. Biochemistry 1986;25:7717–25.10.1021/bi00371a065Search in Google Scholar PubMed
24. Silversmith RE, Nelsestuen GL. The assembly of the membrane attack complex of complement on small unilamellar phospholipid vesicles. Biochemistry 1986;25:852–60.10.1021/bi00352a017Search in Google Scholar
25. Preissner KT, Podack E, Müller-Eberhard H. The membrane attack complex of complement: relation of C7 to the metastable membrane binding site of the intermediate complex C5b-7. J Immunol 1985;135:445–51.10.4049/jimmunol.135.1.445Search in Google Scholar
26. Marshall P, Hasegawa A, Davidson EA, Nussenzweig V, Whitlow M. Interaction between complement proteins C5b-7 and erythrocyte membrane sialic acid. J Exp Med 1996;184:1225–32.10.1084/jem.184.4.1225Search in Google Scholar
27. Podack ER, Biesecker G, Kolb WP, Müller-Eberhard HJ. The C5b-6 complex: reaction with C7, C8, C9. J Immunol 1978;121:484–90.10.4049/jimmunol.121.2.484Search in Google Scholar
28. Hadders MA, Bubeck D, Roversi P, Hakobyan S, Forneris F, Morgan BP et al. Assembly and regulation of the membrane attack complex based on structures of C5b6 and sC5b9. Cell Reports 2012;1:200–07.10.1016/j.celrep.2012.02.003Search in Google Scholar
29. Bubeck D. The making of a macromolecular machine: Assembly of the membrane attack complex. Biochemistry 2014;53:1908–15.10.1021/bi500157zSearch in Google Scholar
30. Tschopp J. Ultrastructure of the membrane attack complex of complement. Heterogeneity of the complex caused by different degree of C9 polymerization. J Biol Chem 1984;259:7857–63.10.1016/S0021-9258(17)42872-9Search in Google Scholar
31. Tschopp J, Podack E, Müller-Eberhard H. The membrane attack complex of complement: C5b-8 complex as accelerator of C9 polymerization. J Immunol 1985;134:495–9.10.4049/jimmunol.134.1.495Search in Google Scholar
32. Podack ER, Tschoop J, Muller-Eberhard H. Molecular organization of C9 within the membrane attack complex of complement. Induction of circular C9 polymerization by the C5b-8 assembly. J Exp Med 1982;156:268–82.10.1084/jem.156.1.268Search in Google Scholar PubMed PubMed Central
33. Jackman JA, Cho N-J, Duran RS, Frank CW. Interfacial binding dynamics of bee venom phospholipase A2 investigated by dynamic light scattering and quartz crystal microbalance. Langmuir 2009;26:4103–12.10.1021/la903117xSearch in Google Scholar PubMed
34. Cho N-J, Jackman JA, Liu M, Frank CW. pH-Driven assembly of various supported lipid platforms: a comparative study on silicon oxide and titanium oxide. Langmuir 2011;27:3739–48.10.1021/la104348fSearch in Google Scholar PubMed
35. Jackman JA, Choi J-H, Zhdanov VP, Cho N-J. Influence of osmotic pressure on adhesion of lipid vesicles to solid supports. Langmuir 2013;29:11375–84.10.1021/la4017992Search in Google Scholar PubMed
36. Jackman JA, Zhao Z, Zhdanov VP, Frank CW, Cho N-J. Vesicle adhesion and rupture on silicon oxide: Influence of freeze–thaw pretreatment. Langmuir 2014;30:2152–60.10.1021/la404582nSearch in Google Scholar PubMed
37. Jackman JA, Tabaei SR, Zhao Z, Yorulmaz S, Cho N-J. Self-assembly formation of lipid bilayer coatings on bare aluminum oxide: overcoming the force of interfacial water. ACS Appl Mater Inter 2015;7:959–68.10.1021/am507651hSearch in Google Scholar PubMed
38. Tabaei SR, Jackman JA, Kim S-O, Zhdanov VP, Cho N-J. Solvent-assisted lipid self-assembly at hydrophilic surfaces: factors influencing the formation of supported membranes. Langmuir 2015;31:3125–34.10.1021/la5048497Search in Google Scholar PubMed
39. Tabaei SR, Jackman JA, Kim S-O, Liedberg B, Knoll W, Parikh AN, et al. Formation of cholesterol-rich supported membranes using solvent-assisted lipid self-assembly. Langmuir 2014;30:13345–52.10.1021/la5034433Search in Google Scholar PubMed
40. Tabaei SR, Jackman JA, Liedberg B, Parikh AN, Cho N-J. Observation of stripe superstructure in the ß-two-phase coexistence region of cholesterol–phospholipid mixtures in supported membranes. J Am Chem Soc 2014;136:16962–5.10.1021/ja5082537Search in Google Scholar PubMed
41. Cho N-J, Frank CW, Kasemo B, Höök F. Quartz crystal microbalance with dissipation monitoring of supported lipid bilayers on various substrates. Nature Protocols 2010;5:1096–1106.10.1038/nprot.2010.65Search in Google Scholar PubMed
©2015 by De Gruyter
Articles in the same Issue
- Frontmatter
- In this issue
- News
- News from the European Foundation for Clinical Nanomedicine (CLINAM)
- What’s up in nanomedicine?
- Special Section Nanosafety
- Guest Editorial
- Sizing up the safety of nanomaterials
- Essay
- Keeping it small: towards a molecular definition of nanotoxicology
- Reviews
- The bio-corona and its impact on nanomaterial toxicity
- In vitro-ex vivo model systems for nanosafety assessment
- Meeting Report
- Nanosafety forum for young scientists: a meeting report
- Short Communication
- Nanomedicines in the European translational process
- Special Section CARPA Part 2
- Guest Editorial
- Complement activation-related pseudoallergy: an innate response to nanomedicines acting as pseudo-viruses
- Reviews
- Exosomes: potential model for complement-stealth delivery systems
- Lessons learned from the porcine CARPA model: constant and variable responses to different nanomedicines and administration protocols
- Mini Review
- Blood cell changes in complement activation-related pseudoallergy
- Short Communications
- Membrane attack complex formation on a supported lipid bilayer: initial steps towards a CARPA predictor nanodevice
- Paradoxical rise of hemolytic complement in the blood of mice during zymosan- and liposome-induced CARPA: a pilot study
- Commentary
- Insidious pathogen-mimicking properties of nanoparticles in triggering the lectin pathway of the complement system
Articles in the same Issue
- Frontmatter
- In this issue
- News
- News from the European Foundation for Clinical Nanomedicine (CLINAM)
- What’s up in nanomedicine?
- Special Section Nanosafety
- Guest Editorial
- Sizing up the safety of nanomaterials
- Essay
- Keeping it small: towards a molecular definition of nanotoxicology
- Reviews
- The bio-corona and its impact on nanomaterial toxicity
- In vitro-ex vivo model systems for nanosafety assessment
- Meeting Report
- Nanosafety forum for young scientists: a meeting report
- Short Communication
- Nanomedicines in the European translational process
- Special Section CARPA Part 2
- Guest Editorial
- Complement activation-related pseudoallergy: an innate response to nanomedicines acting as pseudo-viruses
- Reviews
- Exosomes: potential model for complement-stealth delivery systems
- Lessons learned from the porcine CARPA model: constant and variable responses to different nanomedicines and administration protocols
- Mini Review
- Blood cell changes in complement activation-related pseudoallergy
- Short Communications
- Membrane attack complex formation on a supported lipid bilayer: initial steps towards a CARPA predictor nanodevice
- Paradoxical rise of hemolytic complement in the blood of mice during zymosan- and liposome-induced CARPA: a pilot study
- Commentary
- Insidious pathogen-mimicking properties of nanoparticles in triggering the lectin pathway of the complement system