Plankton interaction model: Effect of prey refuge and harvesting
-
Poulomi Basak
 ,
Jai Prakash Tripathi
,
Vandana Tiwari
,
Jai Prakash Tripathi
,
Vandana Tiwari

Abstract
Harmful algal blooms are one of the major threats to aquatic ecosystem. Some phytoplankton species produce toxins during algal bloom and affect other aquatic species as well as human beings. Thus, for the conservation of aquatic habitat, it is much needed to control such phenomenon. In the present study, we propose a mathematical model of toxin-producing phytoplankton and zooplankton species, which follows the Holling Type III functional response. We consider the effect of prey refuge and harvesting on both the species. Boundedness of the proposed model, existence of equilibria, and their stability have been discussed analytically. We also discuss the optimal harvesting policy and existence of bionomic equilibrium. The numerical simulation has also been performed. We identify the control parameters that are responsible for the system dynamics of the model. The parameter prey refuge has a great impact on the dynamics of the model system. Higher value of prey refuge leads to the stable dynamics. Also, the growth rate of phytoplankton acts as a control parameter for the dynamics of the model. The higher value of growth rate of phytoplankton is responsible for oscillatory behavior.
1 Introduction
Planktons are the main supplier of dissolved oxygen in water and play an important role in the food chain of aquatic ecosystem [13]. Harmful algal blooms (HABs) are one of the major causes of the degradation of aquatic biodiversity. Human health, fisheries, and local economies are also affected by this global problem, “Harmful Algal Blooms” [23]. HABs occur when an algae population (e.g., Dinoflagellates) experiences explosive growth and releases toxins or has other negative effects on the environment [5]. River Ganga is one of the most significant rivers in India and provides the habitat to numerous aquatic species and microorganisms. However, the color of the river Ganga suddenly changed to green due to the rise of algal bloom in the Varanasi and the nearby regions during May–June 2021 [3]. Thus, the conservation of aquatic diversity from such threats is much needed for ecological as well as economical points of view. Many authors have developed different mathematical models addressing the issue of HABs and suggested proper management for biodiversity conservation [6–9,20–22]. Chattopadhayay et al. [8] observed that toxic substances released by toxin-producing phytoplankton (TPP) play a significant role in the termination of planktonic blooms. As a result, toxic chemicals serve as a biocontrol agent for other plankton species. A three-species model was studied by Roy and Chattopadhyay [21], and the authors observed that toxic or allelopathic compounds produced by TPP may assist in diminishing the competition coefficient among various phytoplankton species. Chakraborty et al. [6] developed a model with three interacting species, taking into account the impact of avoidance of zooplankton for the toxic phytoplankton in the presence of a non-toxic one. It has been noted that high avoidance plays a crucial role in facilitating the coexistence of toxic phytoplankton, non-toxic phytoplankton and zooplankton. A mathematical model of HABs was studied by Braselton and Braselton [5], and the authors have suggested that introducing additional prey or predators of the original predator may assist in its control. Sarkar et al. [22] studied the effect of HABs on zooplankton species due to TPP through a food chain model among TPP and zooplankton species. A three-species plankton model with optimal control strategy has been studied by Xiang et al. [29].
The dynamical behavior of toxic phytoplankton and zooplankton model with the impact of phytoplankton harvesting was analyzed by Zhang and Niu [30]. The authors found that phytoplankton and zooplankton populations oscillate periodically when the harvesting level is high. Li et al. [14] developed a phytoplankton–zooplankton interaction model with toxin effect and refuge. The authors suggested that when making decisions regarding predation and control of HABs in aquatic system, the toxin and refuge have a great impact. Pandey et al. [17] constructed and examined a stage-structured prey-predator model that incorporates refuge behavior for prey and gestational delay. The authors have shown that the immature prey refuge functions as a destabilizing factor in the system in the absence of time delay while it serves as a control parameter in the presence of delay. The detailed recent studies related to prey refuge can also be found in [25–27]. An optimal control of harvesting effort in phytoplankton-zooplankton model with the effect of toxicity was discussed by Agnihotri and Kaur [2]. Huda et al. [12] investigated the dynamic behavior of a prey-predator model in which harvesting effort is employed as a control variable. Their study revealed that the optimal solution can lead to the achievement of ecosystem sustainability. Panja et al. [18] suggested that for the stability of ecological system, we should control the environmental toxins released by natural or different human activities through a plankton-fish model with toxicant and harvesting. Lv et al. [15] studied the effect of harvesting on plankton dynamics and found that the zero discounting leads to the maximization of economic revenue and that an infinite discount rate leads to complete dissipation of economic rent. The effect of linear harvesting along with the Allee effect in a delayed phytoplankton zooplankton model was discussed by Meng et al. [16], and the authors obtained optimal harvesting policy using Pontryagin’s maximum principle. Recently, Agmour et al. [1] developed a mathematical model among the dinoflagellate phytoplankton and mytilid species with effect of harvesting in both the species and analyzed the bionomic equilibrium and optimal harvesting policy for their proposed model. In this article, we propose a mathematical model of the interaction of phytoplankton and zooplankton influenced by toxin release by phytoplankton. We also consider the impact of harvesting on both the species in the present model. The main objective of this article is to study the dynamics of the proposed model analytically as well as numerically.
The present article has been organized as follows. In Section 2, a two-species phytoplankton-zooplankton model is presented. The mathematical analysis and optimal harvesting policy are discussed in Section 3. Results of numerical simulations are presented and discussed in Section 4. Finally, Section 5 concludes the study.
2 Model formulation
In this study, we propose a mathematical model to elucidate the dynamics of phytoplankton and zooplankton populations. We consider the impact of toxins released by phytoplankton, which affects the zooplankton species. We assume that the interaction among the plankton species follows the Holling type-III functional response. As the liberation of toxin reduces the growth of zooplankton, it causes substantial mortality of zooplankton. During the toxin liberation period, TPP is not easily accessible; hence, a more common and intuitive choice may be the Holling type II or type III functional response to describe the grazing phenomena [8]. Holling [10] proposed Holling type-III function response of the form
Here,
with initial condition
Here,
3 Mathematical analysis of model system
Lemma 1
Proof
Obviously, the solution of
From the first equation of system
Then, by the standard comparison theorem, it can be shown that
Let us define a function
The time derivative of
Now, we have
By applying the theory of Birkhoff and Rota [4] of differential inequality, we obtain
When
3.1 Existence of equilibrium points and their local stability
In this section, we delve into the examination of the existence and stability of equilibrium points to analyze the dynamics of the temporal model (1).
The equilibrium points of the system (1) are as follows:
The trivial equilibrium point
The zooplankton-free equilibrium point
The coexistence equilibrium point
(3)(4)
Figure 1 shows the existence of equilibrium point

Existence of interior equilibrium point
3.2 Stability analysis
The Jacobian matrix of model system (1) at
The eigenvalues of
The Jacobian matrix of model system (1) at
The eigenvalues of
The Jacobian matrix corresponding to
where
The characteristic equation of the above matrix is given by
where
Theorem 1
The equilibrium point
Proof
Using Routh-Hurwitz criterion, equilibrium point
3.3 Bionomic equilibrium and optimal harvesting policy
In this section, we explore the concept of bionomic equilibrium and the optimal harvesting policy. Bionomic equilibrium is the combination of biological equilibrium and economic equilibrium. The bionomic equilibrium is a concept that integrates biological and economical aspects in the population dynamics. The biological equilibrium is found by solving the equations
3.3.1 Bionomic equilibrium
The net economic revenue at time
where
Now, we have
From equations (6) and (7), the following equation represents the non-trivial equilibrium solution that occurs at a point on the biological equilibrium curve:
Thus, from equation (8), the bionomic equilibrium
where
Equation (9) has unique positive root
3.3.2 Optimal harvesting policy
The objective function that we optimize is given by
where
The associated Hamiltonian function is
where
We aim to find an optimal equilibrium solution of the objective function
From equations (11) and (13), we have
From equations (12) and (13), we have
By eliminating
where
The shadow prices
Using the Hamiltonian function (10), the control constraint is given by
Now, by using the values of
which is equivalent to
By solving equations (6), (7), and (19), we obtain the optimal populations of phytoplankton
then
4 Numerical simulations
In this section, we illustrate the dynamical behavior of system (1) through numerical simulations. For the numerical simulation, we have taken the set of parametric values given in equation (5). Figure 2 depicts the impact of prey refuge



Bifurcation diagram of model system (1) with respect to (a)
In Figure 5, one can observe that higher value of harvesting effort leads to the extinction of zooplankton. At
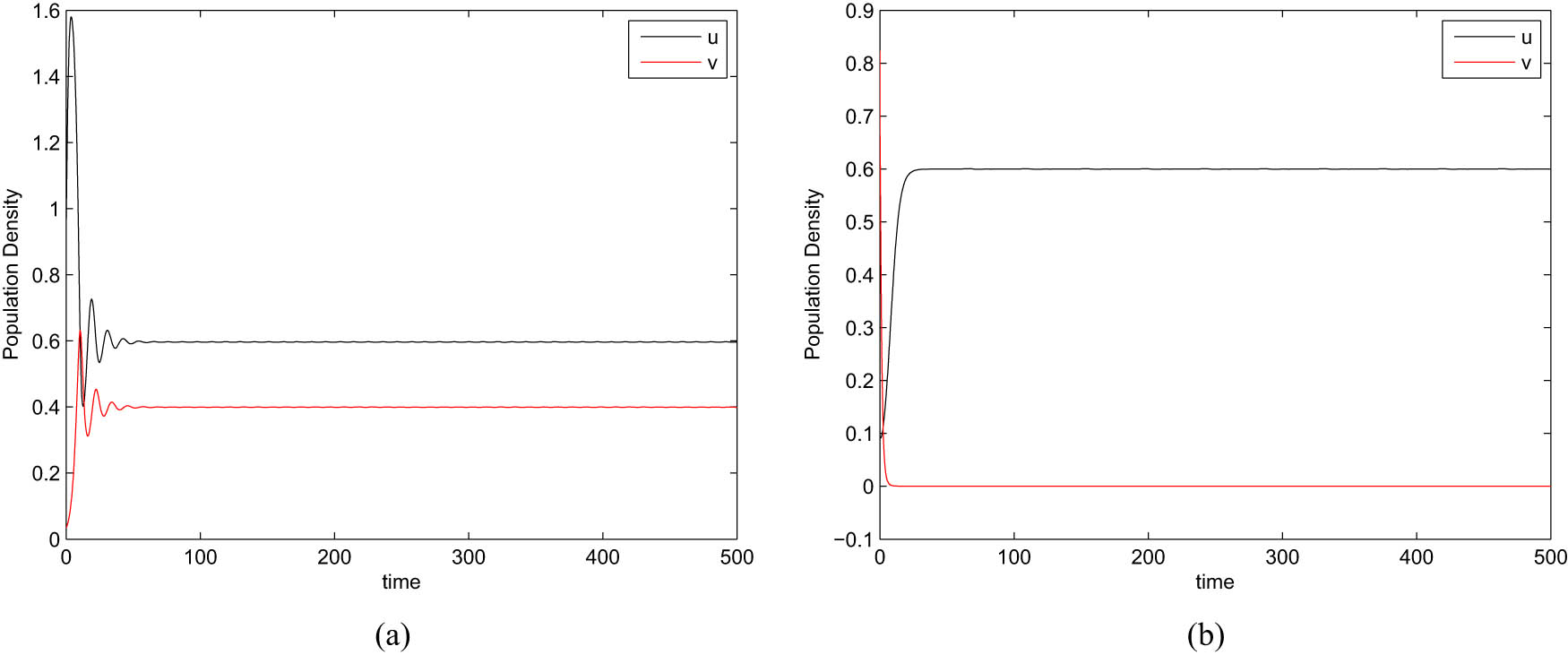
5 Summary and conclusion
The hiding behavior of prey is crucial in ensuring the survival of species. Seeking refuge allows prey to shield themselves from predators, thus avoiding excessive predation. This contributes to the overall stability of the ecosystem. Additionally, implementing an optimal harvesting policy is essential in preventing the overexploitation of species. The study of Tripathi et al. [28] revealed that harvesting effort acts as a mechanism of biological control for plankton species. In this article, we have proposed a nonlinear mathematical model among phytoplankton and zooplankton species with the combined effects of prey refuge and harvesting. The phytoplankton and zooplankton populations are harvested according to catch per unit effort hypothesis. We have analyzed the linear stability of the model system analytically. We have also derived conditions for bionomic equilibrium and optimal harvesting policy using Pontryagin’s maximum principle. It has been observed that zero discount rate gives the maximum value of the net revenue, while an infinite discount rate leads to zero economic revenue. Numerically, we investigated the dynamics of the plankton model by varying the parameters. We have found that prey refuge and growth rate of phytoplankton have a great impact on the dynamics of the system. The higher value of prey refuge leads to the stable dynamics, whereas the lower value of growth rate of phytoplankton is responsible for the stable behavior of the model system. We can also observe the same behavior from the bifurcation diagram of the model system (Figure 4). In [24], Tripathi and Singh observed that the occurrence of Hopf bifurcation guarantees the survival prospects of all species. Also, we have found the extinction scenario of zooplankton at a higher value of harvesting effort which indicates the bad health of biodiversity of aquatic ecosystem. Thus, management of harvesting effort is much needed for the coexistence of both species.
Acknowledgement
The authors are grateful for the reviewer’s valuable comments that improved the manuscript.
-
Funding information: This research work of Satish Kumar Tiwari is supported by RGIPT, Jais Amethi (P-2207). The research work of Jai Prakash Tripathi is supported by the Science and Engineering Research Board (SERB), India [File No. MTR/2022/001028].
-
Author contributions: All authors have accepted responsibility for the entire content of this manuscript and consented to its submission to the journal, reviewed all the results, and approved the final version of the manuscript. Poulomi Basak and Satish Kumar Tiwari proposed the problem, developed the model, and carried out mathematical and numerical analysis. Jai Prakash Tripathi supervised, reviewed, and corrected the manuscript. Vandana Tiwari and Ratnesh Kumar Mishra reviewed the manuscript.
-
Conflict of interest: There is no conflict of interest to declare.
-
Ethical statement: This research did not require ethical approval.
References
[1] Agmour, I., Baba, N., Bentounsi, M., Achtaich, N., El Foutayeni, Y. (2021). Mathematical study and optimal control of bioeconomic model concerning harmful dinoflagellate phytoplankton. Computational and Applied Mathematics, 40, 1–16. 10.1007/s40314-021-01509-3Search in Google Scholar
[2] Agnihotri, K., Kaur, H. (2021). Optimal control of harvesting effort in a phytoplankton-zooplankton model with infected zooplankton under the influence of toxicity. Mathematics and Computers in Simulation, 190, 946–964. 10.1016/j.matcom.2021.06.022Search in Google Scholar
[3] Bhattacharjee, R., Gupta, A., Das, N., Agnihotri, A. K., Ohri, A., Gaur, S. (2022). Analysis of algal bloom intensification in mid-ganga river, India, using satellite data and neural network techniques. Environmental Monitoring and Assessment, 194(8), 547. 10.1007/s10661-022-10213-6Search in Google Scholar PubMed PubMed Central
[4] Birkhoff, G., Rota, G. (1982). Ordinary Differential Equation, ginn and co. Boston.Search in Google Scholar
[5] Braselton, J., Braselton, L. (2004). A model of harmful algal blooms. Mathematical and computer modelling, 40(9–10), 923–934. 10.1016/j.mcm.2004.09.001Search in Google Scholar
[6] Chakraborty, S., Bhattacharya, S., Feudel, U., Chattopadhyay, J. (2012). The role of avoidance by zooplankton for survival and dominance of toxic phytoplankton. Ecological complexity, 11, 144–153. 10.1016/j.ecocom.2012.05.006Search in Google Scholar
[7] Chakraborty, S., Chattopadhyay, J. (2008). Nutrient-phytoplankton-zooplankton dynamics in the presence of additional food source – mathematical study. Journal of Biological Systems, 16(04), 547–564. 10.1142/S0218339008002654Search in Google Scholar
[8] Chattopadhayay, J., Sarkar, R. R., Mandal, S. (2002). Toxin-producing plankton may act as a biological control for planktonic blooms – field study and mathematical modelling. Journal of Theoretical Biology, 215(3), 333–344. 10.1006/jtbi.2001.2510Search in Google Scholar PubMed
[9] Chattopadhyay, J., Sarkar, R. R., El Abdllaoui, A. (2002). A delay differential equation model on harmful algal blooms in the presence of toxic substances. Mathematical Medicine and Biology: A Journal of the IMA, 19(2), 137–161. 10.1093/imammb19.2.137Search in Google Scholar
[10] Holling, C. S. (1965). The functional response of predators to prey density and its role in mimicry and population regulation. The Memoirs of the Entomological Society of Canada, 97(S45), 5–60. 10.4039/entm9745fvSearch in Google Scholar
[11] Huang, Y., Chen, F., Zhong, L. (2006). Stability analysis of a prey-predator model with holling type iii response function incorporating a prey refuge. Applied Mathematics and Computation, 182(1), 672–683. 10.1016/j.amc.2006.04.030Search in Google Scholar
[12] Huda, M. N., A’yun, Q. Q., Wigantono, S., Sandariria, H., Raming, I., Asmaidi, A. (2023). Effects of harvesting and planktivorous fish on bioeconomic phytoplankton-zooplankton models with ratio-dependent response functions and time delays. Chaos, Solitons & Fractals, 173, 113736. 10.1016/j.chaos.2023.113736Search in Google Scholar
[13] Ji, J., Lei, C., Yuan, Y. (2023). Qualitative analysis on a reaction-diffusion nutrient-phytoplankton model with toxic effect of holling-type ii functional. Discrete & Continuous Dynamical Systems-Series B, 28(4), 2745–2767. 10.3934/dcdsb.2022190Search in Google Scholar
[14] Li, J., Song, Y., Wan, H., Zhu, H. (2016). Dynamical analysis of a toxin-producing phytoplankton-zooplankton model with refuge. Mathematical Biosciences & Engineering, 14(2), 529–557. Search in Google Scholar
[15] Lv, Y., Pei, Y., Gao, S., Li, C. (2010). Harvesting of a phytoplankton-zooplankton model. Nonlinear Analysis: Real World Applications, 11(5), 3608–3619. 10.1016/j.nonrwa.2010.01.007Search in Google Scholar
[16] Meng, X., Li, J. (2020). Stability and hopf bifurcation analysis of a delayed phytoplankton-zooplankton model with allee effect and linear harvesting. Mathematical Biosciences and Engineering, 17(3), 1973–2002. 10.3934/mbe.2020105Search in Google Scholar PubMed
[17] Pandey, S., Ghosh, U., Das, D., Chakraborty, S., Sarkar, A. (2024). Rich dynamics of a delay-induced stage-structure prey-predator model with cooperative behavior in both species and the impact of prey refuge. Mathematics and Computers in Simulation, 216, 49–76. 10.1016/j.matcom.2023.09.002Search in Google Scholar
[18] Panja, P., Mondal, S. K., Jana, D. K. (2017). Effects of toxicants on phytoplankton-zooplankton-fish dynamics and harvesting. Chaos, Solitons & Fractals, 104, 389–399. 10.1016/j.chaos.2017.08.036Search in Google Scholar
[19] Pontryagin, L. S. (2018). Mathematical theory of optimal processes. London: Routledge. 10.1201/9780203749319Search in Google Scholar
[20] Roy, S., Alam, S., Chattopadhyay, J. (2006). Competing effects of toxin-producing phytoplankton on overall plankton populations in the Bay of Bengal. Bulletin of Mathematical Biology, 68, 2303–2320. 10.1007/s11538-006-9109-5Search in Google Scholar PubMed
[21] Roy, S., Chattopadhyay, J. (2007). Toxin-allelopathy among phytoplankton species prevents competitive exclusion. Journal of Biological Systems, 15(01), 73–93. 10.1142/S021833900700209XSearch in Google Scholar
[22] Sarkar, R. R., Pal, S., Chattopadhyay, J. (2005). Role of two toxin-producing plankton and their effect on phytoplankton-zooplankton system-a mathematical study supported by experimental findings. BioSystems, 80(1), 11–23. 10.1016/j.biosystems.2004.09.029Search in Google Scholar PubMed
[23] Shaika, N., Khan, S., Sultana, S. (2022). Harmful algal blooms in the coastal waters of Bangladesh: an overview. Journal of Aquaculture & Marine Biology, 11(3), 105–111. 10.15406/jamb.2022.11.00344Search in Google Scholar
[24] Tripathi, D., Singh, A. (2023). An eco-epidemiological model with predator switching behavior. Computational and Mathematical Biophysics, 11(1), 20230101. 10.1515/cmb-2023-0101Search in Google Scholar
[25] Tripathi, J. P., Abbas, S., Thakur, M. (2015). A density dependent delayed predator-prey model with Beddington-Deangelis type function response incorporating a prey refuge. Communications in Nonlinear Science and Numerical Simulation, 22, 427–450. 10.1016/j.cnsns.2014.08.018Search in Google Scholar
[26] Tripathi, J. P., Abbas, S., Thakur, M. (2015). Dynamical analysis of a prey-predator model with beddington-deangelis type function response incorporating a prey refuge. Nonlinear Dynamics, 80, 177–196. 10.1007/s11071-014-1859-2Search in Google Scholar
[27] Tripathi, J. P., Jana, D. D., Vyshnavi Devi, N., Tiwari, V., Abbas, S. (2020). Intraspecific competition of predator for prey with variable rates in protected areas. Nonlinear Dynamics, 102, 511–535. 10.1007/s11071-020-05951-6Search in Google Scholar
[28] Tripathi, J. P., Tripathi, D., Mandal, S., Shrimali, M. (2023). Cannibalistic enemy-pest model: effect of additional food and harvesting. Journal of Mathematical Biology, 87, 58. 10.1007/s00285-023-01991-9Search in Google Scholar PubMed
[29] Xiang, H., Liu, B., Fang, Z. (2018). Optimal control strategies for a new ecosystem governed by reaction-diffusion equations. Journal of Mathematical Analysis and Applications, 467(1), 270–291. 10.1016/j.jmaa.2018.07.001Search in Google Scholar
[30] Zhang, H., Niu, B. (2020). Dynamics in a plankton model with toxic substances and phytoplankton harvesting. International Journal of Bifurcation and Chaos, 30(2), 2050035. 10.1142/S0218127420500352Search in Google Scholar
© 2024 the author(s), published by De Gruyter
This work is licensed under the Creative Commons Attribution 4.0 International License.
Articles in the same Issue
- Special Issue: Recent Trends in Mathematical Biology – Theory, Methods, and Applications
- Editorial for the Special Issue: Recent trends in mathematical Biology – Theory, methods, and applications
- Behavior of solutions of a discrete population model with mutualistic interaction
- Influence of media campaigns efforts to control spread of COVID-19 pandemic with vaccination: A modeling study
- Optimal control and bifurcation analysis of SEIHR model for COVID-19 with vaccination strategies and mask efficiency
- A mathematical study of the adrenocorticotropic hormone as a regulator of human gene expression in adrenal glands
- On building machine learning models for medical dataset with correlated features
- Analysis and numerical simulation of fractional-order blood alcohol model with singular and non-singular kernels
- Stability and bifurcation analysis of a nested multi-scale model for COVID-19 viral infection
- Augmenting heart disease prediction with explainable AI: A study of classification models
- Plankton interaction model: Effect of prey refuge and harvesting
- Modelling the leadership role of police in controlling COVID-19
- Robust H∞ filter-based functional observer design for descriptor systems: An application to cardiovascular system monitoring
- Regular Articles
- Mathematical modelling of COVID-19 dynamics using SVEAIQHR model
- Optimal control of susceptible mature pest concerning disease-induced pest-natural enemy system with cost-effectiveness
- Correlated dynamics of immune network and sl(3, R) symmetry algebra
- Variational multiscale stabilized FEM for cardiovascular flows in complex arterial vessels under magnetic forces
- Assessing the impact of information-induced self-protection on Zika transmission: A mathematical modeling approach
- An analysis of hybrid impulsive prey-predator-mutualist system on nonuniform time domains
- Modelling the adverse impacts of urbanization on human health
- Markov modeling on dynamic state space for genetic disorders and infectious diseases with mutations: Probabilistic framework, parameter estimation, and applications
- In silico analysis: Fulleropyrrolidine derivatives against HIV-PR mutants and SARS-CoV-2 Mpro
- Tangleoids with quantum field theories in biosystems
- Analytic solution of a fractional-order hepatitis model using Laplace Adomian decomposition method and optimal control analysis
- Effect of awareness and saturated treatment on the transmission of infectious diseases
- Development of Aβ and anti-Aβ dynamics models for Alzheimer’s disease
- Compartmental modeling approach for prediction of unreported cases of COVID-19 with awareness through effective testing program
- COVID-19 transmission dynamics in close-contacts facilities: Optimizing control strategies
- Modeling and analysis of ensemble average solvation energy and solute–solvent interfacial fluctuations
- Application of fluid dynamics in modeling the spatial spread of infectious diseases with low mortality rate: A study using MUSCL scheme
Articles in the same Issue
- Special Issue: Recent Trends in Mathematical Biology – Theory, Methods, and Applications
- Editorial for the Special Issue: Recent trends in mathematical Biology – Theory, methods, and applications
- Behavior of solutions of a discrete population model with mutualistic interaction
- Influence of media campaigns efforts to control spread of COVID-19 pandemic with vaccination: A modeling study
- Optimal control and bifurcation analysis of SEIHR model for COVID-19 with vaccination strategies and mask efficiency
- A mathematical study of the adrenocorticotropic hormone as a regulator of human gene expression in adrenal glands
- On building machine learning models for medical dataset with correlated features
- Analysis and numerical simulation of fractional-order blood alcohol model with singular and non-singular kernels
- Stability and bifurcation analysis of a nested multi-scale model for COVID-19 viral infection
- Augmenting heart disease prediction with explainable AI: A study of classification models
- Plankton interaction model: Effect of prey refuge and harvesting
- Modelling the leadership role of police in controlling COVID-19
- Robust H∞ filter-based functional observer design for descriptor systems: An application to cardiovascular system monitoring
- Regular Articles
- Mathematical modelling of COVID-19 dynamics using SVEAIQHR model
- Optimal control of susceptible mature pest concerning disease-induced pest-natural enemy system with cost-effectiveness
- Correlated dynamics of immune network and sl(3, R) symmetry algebra
- Variational multiscale stabilized FEM for cardiovascular flows in complex arterial vessels under magnetic forces
- Assessing the impact of information-induced self-protection on Zika transmission: A mathematical modeling approach
- An analysis of hybrid impulsive prey-predator-mutualist system on nonuniform time domains
- Modelling the adverse impacts of urbanization on human health
- Markov modeling on dynamic state space for genetic disorders and infectious diseases with mutations: Probabilistic framework, parameter estimation, and applications
- In silico analysis: Fulleropyrrolidine derivatives against HIV-PR mutants and SARS-CoV-2 Mpro
- Tangleoids with quantum field theories in biosystems
- Analytic solution of a fractional-order hepatitis model using Laplace Adomian decomposition method and optimal control analysis
- Effect of awareness and saturated treatment on the transmission of infectious diseases
- Development of Aβ and anti-Aβ dynamics models for Alzheimer’s disease
- Compartmental modeling approach for prediction of unreported cases of COVID-19 with awareness through effective testing program
- COVID-19 transmission dynamics in close-contacts facilities: Optimizing control strategies
- Modeling and analysis of ensemble average solvation energy and solute–solvent interfacial fluctuations
- Application of fluid dynamics in modeling the spatial spread of infectious diseases with low mortality rate: A study using MUSCL scheme