Geclusterte Plastizität bei Langzeitpotenzierung: Wie starke Synapsen bestehen bleiben, um Langzeitgedächtnis aufrechtzuerhalten
-
Marina Mikhaylova
Dr. Marina Mikhaylova (geb. 1981) hat Biologie an der Bashkir State Universität in Ufa, Russische Föderation, studiert. Sie hat ihre Promotion in der Forschungsgruppe ‚Neuroplastizität‘ am Leibniz – Institut für Neurobiologie in Magdeburg 2010 verteidigt. Nach einer zweijährigen PostDoc – Zeit in der gleichen Arbeitsgruppe war sie dann von 2012–2015 als Wissenschaftlerin in der ‚Division of Cell Biology‘ an der Universität Utrecht in den Niederlanden tätig. Seit 2015 leitet sie eine durch das Emmy Noether – Programm finanzierte Arbeitsgruppe in Hamburg.
Michael R. Kreutz hat Psychologie, Philosophie und Linguistik an der Universität Münster studiert. Die Doktorarbeit fertigte er an der Ruhr-Universität Bochum an und nach Aufenthalten am MIT in Cambridge, USA, und am Max-Planck-Institut für experimentelle Medizin in Göttingen ist er seit 1993 in Magdeburg tätig, wo er seit 1998 eine Forschungsgruppe am Leibniz-Institut für Neurobiologie leitet. Seit 2015 ist auch Leiter einer Leibniz – Gastgruppe Zentrum für Molekulare Neurobiologie in Hamburg.

Zusammenfassung
Die Gedächtnisspeicherung erfordert, zumindest teilweise, die Langzeitpotenzierung (LTP) in den Synapsen der dendritischen Dornfortsätze aufrechtzuerhalten. Benachbarte Synapsen bilden häufig funktionelle Cluster. Gegenwärtig ist noch unklar, wie sich Cluster entwickeln, warum sie für längere Zeitabschnitte stabil sind, und wie Dornfortsätze innerhalb eines Clusters interagieren. In diesem Review werden wir einen Überblick über gegenwärtige Konzepte der geclusterten Plastizität geben, und wir werden die zellulären sowie die molekularen Mechanismen diskutieren, welche für die Stabilität der Dornfortsätze und die damit verbundenen Funktionen im Kontext mit LTP relevant sein können. Wir werden den Vorschlag machen, dass die Dynamik der initial gebildeten Cluster von der Kompartimentierung der Dendriten abhängt und dass die aktivitätsabhängige Genexpression dazu kommt, um die unterschiedlichen synaptischen Gewichtungen aufrechtzuerhalten. Wir werden diskutieren, wie für das Aufrechterhalten der geclusterten Plastizität eine Interaktion erfolgt zwischen den Mechanismen des synaptischen „Tagging“ (Etikettieren), der Anwesenheit sekretorischer Organellen in den Dendriten und dem Einbau der synaptischen Skalierungsfaktoren, welche durch unmittelbar-aktivierte Gene („immediate early genes, IEG) codiert werden.
Einleitung
Spezifität, Kapazität und Dauer der Gedächnisspeicherung hängen vermutlich sowohl von der Plastizität als auch der Stabilität der synaptischen Kontakte ab (Pozo und Goda, 2010). Insbesondere die dendritischen Dornfortsätze, ein spezialisierter Typ der glutamatergen Synapsen im Großhirn, sind mit höheren kognitiven Funktionen verbunden. Diese enthalten elektronendichte Proteinnetzwerke, welche postsynaptische Verdichtung (PSD) genannt werden, die dazu dienen, die Neurotransmitterrezeptoren, Ionenkanäle und synaptischen Zelladhäsionsmoleküle ebenso wie Signalbestandteile der Dornfortsätze zu verankern. Dendritische Dornfortsätze können in Form, Größe und Stabilität über die Zeit hinweg unterschiedlich sein. Reife Dornfortsätze haben häufig eine pilzartige Form mit einem breiten Fortsatzkopf (bis zu 0,8–1 µm Durchmesser) mit der PSD und einen dünnen Fortsatzhals (0,1–0,2 µm), welcher den Fortsatz an den dendritischen Schaft anbindet und als Diffusionsbarriere dient (Bosch und Hayashi, 2012). Aktin-Filamente (F-Actin) stellen die hauptsächliche Zytoskelettkomponente der Dornfortsätze dar und sind wesentlich beteiligt bei struktureller Plastizität; sie verankern die mRNA – Granula und Organellen und vermitteln den Transport in die Dornfortsätze und aus diesen heraus (Konietzny et al., 2017). Dendritische Dornfortsätze enthalten hochdynamisches verzweigtes F-Actin im Kopf der Dornfortsätze nahe bei der PSD. Dieser stabile Pool an F-Actin, welches wesentlich für das Aufrechterhalten der Struktur des Dornfortsatzes ist, ist an der Basis des Dornfortsatzes lokalisiert. So werden geradlinige Bündel wie auch ein periodisches Actin-Netz im Fortsatzhals gefunden. Eine solche Nano-Domänen-Organisation des F-Actin in den Dornfortsätzen erlaubt auf der einen Seite rasche Antworten auf extrazelluläre Reize und bietet auf der anderen Seite die Möglichkeit, möglichst optimal über lange Zeitperioden die Form zu stabilisieren. Größere Dornfortsätze können zusätzlich verschiedene Organellen enthalten, wie zum Beispiel den Dornfortsatzapparat, Polyribosomen und andere. Die synaptische Übertragung an erregenden Synapsen beruht auf der Aktivierung der N-Methyl-D-Aspartat-Rezeptoren (NMDARs) und der a Amino-3-hydroxy-5-methyl-4-isocazolepropionsäure-Rezeptoren (AMPARs). Aktivitätsabhängige Veränderungen der synaptischen Übertragung korrelieren streng mit Veränderungen in der Rezeptoranzahl sowie der Form und der Größe der dendritischen Dornfortsätze (Carlisle und Kennedy, 2005).
Mit bildgebenden Verfahren in Zeitraffermethode durchgeführte Untersuchungen der Dornfortsätze weisen darauf hin, dass die Lebensdauer synaptischer Verbindungen sich deutlich zwischen apikalen und basalen Dendriten unterscheidet und auch verschieden ist, je nach unterschiedlicher Hirnregion. In der CA1 – Region des Hippokampus zum Beispiel ist die Population der Dornfortsätze an basalen Dendriten hoch dynamisch. Die mittlere Lebenszeit der basalen CA1-Dornfortsätze (diese erhalten vor allem von den CA3-Zellen einen Input) wird auf zehn Tage geschätzt, und dies ermöglicht einen vollständigen Umbau der Verschaltung innerhalb von drei bis sechs Wochen (Attardo et al., 2015). Interessanterweise zeigte Langzeitbildgebung der apikalen Büscheldendriten der Pyramidenneuronen des CA1 (sie erhalten Input vom entorhinalen Kortex) oder des Neokortex, dass die meisten Dornfortsätze über mehr als drei Monate persistieren können (> 50 %) (Gu et al., 2014; Holtmaat et al., 2005). Diese Dornfortsätze sind gewöhnlich größer, was darauf hinweist, dass sie auch stärker sein könnten, wohingegen kleine Dornfortsätze häufiger auftauchen und verschwinden (Holtmaat et al., 2005). Experimentelle Daten weisen ferner darauf hin, dass die Größe der PSD mit der Stabilität der Dornfortsätze in vivo korreliert (Cane et al., 2014). Unterschiede der Stabilität der Dornfortsätze unterstützen die Idee, dass das transiente Vorhandensein des hippokampusabhängigen Gedächtnisses direkt die erhöhte Umsatzrate der hippocampalen Synapsen reflektiert. Die Situation kann jedoch noch viel komplexer sein. Stabilität und Plastizität der Dornfortsätze scheinen kompartimentiert zu sein, und zwar nicht nur in den apikalen und basalen Dendriten, sondern auch innerhalb eines gegebenen dendritischen Segments. Mehrere Argumente unterstützen die „geclusterte Plastizitätshypothese“, welche vorschlägt, dass Cluster, viel mehr als nur die einzelnen synaptischen Kontakte, eine fundamentale Grundeinheit für die Speicherung des Langzeitgedächtnisses bilden können (Abbildung 1A-B). Pyramidenneurone im Kortex und Hippokampus beherbergen bis zu 10.000 Dornfortsätze, und das gleichzeitige Wachstum und Verschwinden von Synapsen muss nicht nur an jedem einzelnen excitatorischen Input sondern auch auf der Ebene der funktionellen Cluster reguliert sein. Insgesamt gesehen werfen diese Beobachtungen folgende bedeutende Frage auf. In Anbetracht des molekularen Umsatzes ist zu fragen: Wie können geclusterte Synapsen, welche LTP erlebt haben, für so lange Zeitperioden stark bleiben, wie das Gedächtnis aufrechterhalten bleibt?
Geclusterte Plastizität in dendritischen Segmenten
Man weiß relativ wenig darüber, wie LTP die geclusterte Plastizität an sich beeinflusst und welches die zugrunde liegenden Mechanismen sind. Ausreichend starke synaptische Aktivierung kann in individuellen Dornfortsätzen LTP induzieren (Harvey und Svoboda, 2007; Matsuzaki et al., 2004). Jedoch ist die für die Potenzierung erforderliche Reizstärke vermindert, wenn ein benachbarter Dornfortsatz potenziert wird. Dieses Phänomen tritt während der Aktivierung von synaptischen Clustern auf (Govindarajan et al., 2011; Harvey und Svoboda, 2007). Der dendritische Ast stellt ein ideales Segment dar, durch das Signalmoleküle durchlaufen können. In der Tat ist es so, dass die Induktion der NMDAR–abhängigen LTP an individuellen dendritischen Dornfortsätzen Signalkaskaden aktiviert, welche sich über 5–10 µm bis in die Elterndendriten ausbreiten. Darüber hinaus enthalten die dendritischen Äste die Translationsmaschinerie für die Synthese neuer Proteine und sekretorische Transportorganellen, welche das richtige Falten, die Modifikation und die Zustellung der plastizitätsbezogenen Membranproteine sicherstellen (Hanus und Ehlers, 2016; Mikhaylova et al., 2016). Daher ermöglicht die dendritische Kompartimentierung auf der Ebene individueller Äste möglicherweise einen autonomen Aufbau und Unterhalt von geclusterten Synapsen.
Mehrere Untersuchungen haben in der Tat die Existenz einer dendritischen Kompartimentierung in vivo und in vitro bewiesen (Govindarajan et al., 2011; Kleindienst et al., 2011; Makino und Malinow, 2011; Takahashi et al., 2012). Interessanterweise scheinen die molekularen Mechanismen des synaptischen Clustering bei jungen und erwachsenen Gehirnen unterschiedlich zu sein. Bei synaptischer Aktivierung in der altersabhängigen Entwicklung breitet sich Kalzium in Dendriten aus und hilft dabei, andere gleichzeitig aktive Dornfortsätze zu stärken, indem die Reizschwelle, welche für die Potenzierung erforderlich ist, erniedrigt wird. Räumlich geclusterte und zeitlich korrelierte synaptische Eingänge zeigen lokal kooperative Plastizität, und Synapsenreifung ist zusammen mit dem Clustern der synaptischen Wichtungen in den sich entwickelnden Dendritenbäumen räumlich reguliert (Lee et al., 2016; van Bommel und Mikhaylova, 2016). In erwachsenen Neuronen beobachtet man eine zunehmende Dichte der synaptischen Cluster während der Lernvorgänge. In diesem Fall ist die Kalziumerhöhung hauptsächlich auf den Kopf des Dornfortsatzes begrenzt und die Signalübertragung zwischen benachbarten Dornfortsätzen hängt von lokaler Depolarisation, Aktivierung und Diffusion von Signalmolekülen ebenso wie von Translation der dendritischen mRNA ab. Studien, die das Ziel hatten, zu verstehen, wie synaptisches Clustering sich zu LTP und Synaptic tagging (Adressieren) verhält, zeigen, dass einige Schlüsselfaktoren auch bei der Induktion der LTP an Einzelsynapsen eine Rolle spielen (van Bommel und Mikhaylova, 2016).
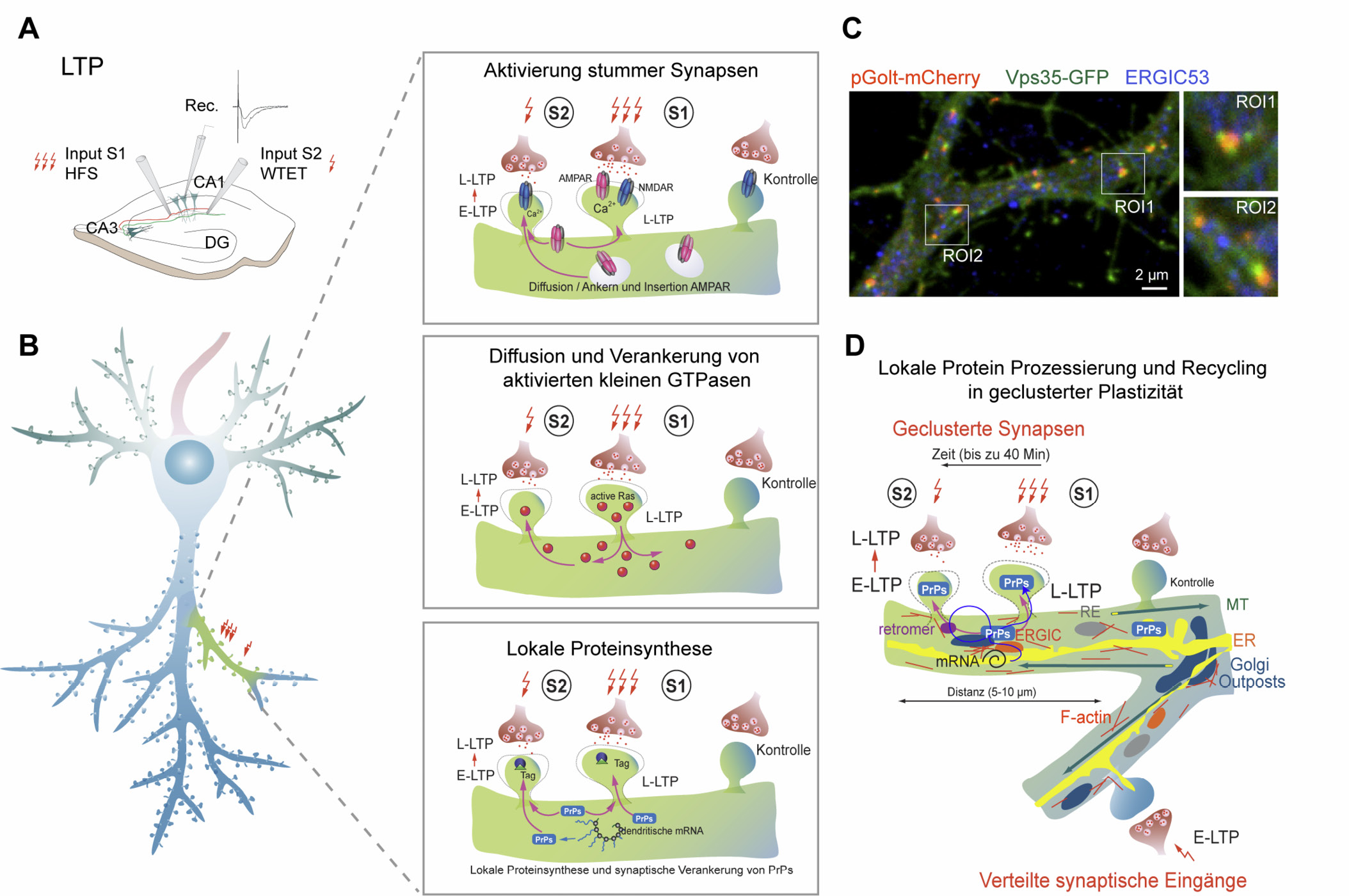
Molekulare Mechanismen der geclusterten Plastizität.
(A) Induktion von LTP und synaptischem Etikettieren in Hippokampuss-Schnitten in vitro. HFS – Hochfrequenzstimulation, WTET – schwache tetanische Stimulation mithilfe von Stimulationselektroden (Input S1 und S2), Rec. – Ableitelektrode.
(B) Mögliche Mechanismen, welche während des Aufbaus der geclusterten Plastizität mit jeweils verschiedener Zeitskala aktiv sind. Die Schwelle für die Induktion von LTP ist niedriger in Dornfortsätzen, welche in Nachbarschaft zu einem Dornfortsatz gelegen sind, bei dem LTP erfolgreich induziert wurde. Somit werden Reize, welche normalerweise nur eine frühe Form der LTP (E-LTP) auslösen, eine Transformation in eine späte Form verursachen (L-LTP). Mögliche molekulare Mechanismen werden dargestellt (siehe auch den Haupttext).
(C) Konfokales Bild eines Hippokampus-Neurons, welches mit einem Marker für Golgi Satelliten (pGolt-mCherry) und einem Retromer Marker (Vps35-GFP) transfiziert wurde, wobei das endogene ERGIC (ERGIC53) angefärbt wurde. Es zeigt die nahe beieinander liegende räumliche Verteilung der post-ER sekretorischen Organellen. Rechtes Bild: Hochauflösende Vergrößerung der Regionen von Interesse (ROI) 1 und 2. Abgedruckt mit Erlaubnis aus (Mikhaylova et al., 2016).
(D) Dendritische sekretorische Organellen können einen Beitrag liefern zur dendritischen Kompartimentierung von geclusterten Synapsen. ER –Endoplasmatisches Retikulum, MT – Mikrotubuli, RE – recycling endosome, PrPs – plasticity-related proteins, ERGIC – ER to Golgi intermediate compartment.
Zurzeit werden drei Mechanismen vorgeschlagen, welche in verschiedenen Zeitskalen wirken, (Figur 1B) (Winnubst und Lohmann, 2012):
Initial wird ein „aktiver“ dendritischer Cluster erzeugt, indem stille Synapsen aktiviert werden. „Stille Synapsen“ enthalten NMDRs aber keine AMPARs (Hanse et al., 2009). Induktion der LTP löst das Aufheben des Mg2+-Blocks an NMDRs aus und erhöht die Oberflächenexpression der AMPARs. Dieser Vorgang läuft in Sekunden ab.
Die gleichzeitige Aktivierung der beiden kleinen GTPasen Ras und RhoA während Hochfrequenzstimulation erzeugt eine Wechselwirkung mit benachbarten Dornfortsätzen (Harvey et al., 2008; Murakoshi et al., 2011). Aktivität von Ras breitet sich über circa 10 µm in Dendriten aus und dringt durch Diffusion in benachbarte Dornfortsätze ein (Harvey et al., 2008). Ras ist dann in der Lage, den Signalweg der mitogenaktivierten Proteinkinasen (MAPK) zu aktivieren, welcher die für LTP erforderliche Proteinsynthese stimuliert (Kelleher et al., 2004), wohingegen RhoA den Rock-Signalweg aktiviert, welcher für die Reorganisation des Aktin wichtig ist, um die Dornfortsätze zu vergrößern (Murakoshi et al., 2011). Dieser Prozess läuft innerhalb von Minuten ab.
Die Hypothese des „synaptischen Adressierens und Einfangens“ schlägt vor, dass die Induktion der LTP aktive Synapsen mit Adressen („tags“) versieht, und zwar unabhängig von der Aktivierungsstärke (Frey und Morris, 1997). Hochfrequenzstimulation, welche die Proteinsynthese-abhängige späte Phase der LTP (L-LTP) induziert, verursacht die Herstellung von plastizitätsbezogenen Proteinen (PrP), welche später durch eine jeweils aktive Synapse eingefangen werden, nicht notwendigerweise diejenige Synapse, welche ursprünglich den Hochfrequenzreiz erhalten hatte (Frey und Morris, 1997). Ein plausibles Modell verbindet synaptisches Adressieren mit synaptischer Clusterbildung, weil benachbarte Synapsen, die auf einem Ast lokalisiert sind, eine höhere Wahrscheinlichkeit haben, das „synaptische Etikett“ zu erwischen (Govindarajan et al., 2006). Infolge der Induktion der LTP ist die Expression von PrPs erhöht, was das synaptische Clustern in benachbarten Synapsen innerhalb von Stunden verstärken wird.
Jedoch gibt es andere Faktoren, wie zum Beispiel die Lokalisation der dendritischen Proteintranslation und der -prozessierungsmaschinerie, Signalwege von Synapse zu Zellkern und zurück, synaptisches Adressieren und invers-Adressieren, die eine wichtige Rolle spielen könnten bei Kompartimentierung der potenzierten Synapsen. Weiter unten werden wir die möglichen Beiträge dieser Faktoren diskutieren.
Spielen dendritische mikrosekretorische Systeme eine Rolle bei der dendritischen Kompartimentierung?
Neurone sind hoch polarisierte Zellen mit einem komplexen dendritischen Baum. Diese komplexe Zytoarchitektur stellt einzigartige Herausforderungen an die Proteostase (Dieterich und Kreutz, 2016; Rosenberg et al., 2014), wobei der Hauptteil der Maschinerie für Synthese und Abbau von Proteinen im Soma lokalisiert ist. Zu circa 20 % erfolgt de novo Proteinsynthese lokal in Dendriten, wo die Maschinerien sowohl für Proteinsynthese als auch – abbau vorhanden sind. Von diesen wurde gezeigt, dass sie die Verfügbarkeit an Protein während der synaptischen Übertragung regulieren. In den letzten Jahren wurde klar, dass es Satelliten-Mikrosekretionssysteme in neuronalen Fortsätzen gibt, welche die lokale Synthese und das Prozessieren der synaptischen Transmembranproteine ermöglichen. Das Endoplasmatische Retikulum in Pyramidenneuronen des Hippokampus ist zwischen Dornfortsätzen und der äußeren Kernmembran durchlaufend. Dendriten enthalten ERGIC, Golgi-Satelliten, Retromere, dendritische mRNA, wobei Polyribosomen überall gefunden werden können (Dieterich und Kreutz, 2016; Hanus und Schuman, 2013) (Abbildung 1C-D). Es wurde gezeigt, dass synaptische Plastizität abhängt von differenziellem Sortieren, Zustellen und Festhalten von Neurotransmitterrezeptoren, und dass NMDAR und AMPAR durch dendritisches ER, ERGIC, GS und Retromere prozessiert werden (Mikhaylova et al., 2016). Die Räumliche Eingrenzung für die Potenzierung geclusterter Synapsen beruht wahrscheinlich auf der Anwesenheit der lokalen mikrosekretorischen Maschinerie, welche den Bedarf nach Membranproteinen befriedigt und welche den verfügbaren Pool definiert. Das dendritische mikrosekretorische System, welches Satelliten-Golgi-Apparat enthält, existiert überall im dendritischen Baum der Pyramidenneurone, aber es ermöglicht die Verfügbarkeit von Proteinen für die Membranen nur in räumlich begrenzten dendritischen Segmenten. Es wird interessant sein zu überprüfen, ob die Anwesenheit von mikrosekretorischen Systemen in Dendriten einen Beitrag liefert zur geclusterten Plastizität (Abbildung 1D).
Integration der lokalen Vorgänge und der aktivitätsabhängigen Genexpression: Ist da eine Balance zwischen verteilter und geclusterter Plastizität?
Faszinierender Weise wird die Tendenz, die potenzierten Dornfortsätze in einem Ast der Dendriten zu akkumulieren, im Gegenzug durch nukleäre ERK-Signale und nachfolgende Genexpression ausgeglichen, welche wiederum durch räumlich verteilte Eingänge induziert werden. Dies mag von Bedeutung sein für die Entwicklung einer ausgewogenen räumlichen Verteilung der synaptischen Wichtungen (Zhai et al., 2013). Was könnte der zugrunde liegende Mechanismus für diese Art verteilter Plastizität sein? Es wurde vorgeschlagen, dass aktivitätsabhängige Genexpression auf die synaptische Funktionalität rückkoppelt, damit Langzeitgedächtnis aufrechterhalten wird (Kaushik et al., 2014; Rosenberg et al., 2014). Indes ist der spezifische Beitrag der Gentranskription zur Bildung des Langzeitgedächtnisses zum großen Teil noch schwer fassbar. Eine zentrale Herausforderung bezüglich der geclusterten Plastizität ist es, die synaptischen Verbindungen aufrechtzuhalten, damit die stromaufwärts und stromabwärts gelegenen Verschaltungen innerhalb des Engramm-Zellensembles bewahrt werden. Computermodellierung legt nahe, dass eine unimodale synaptische Wichtungsverteilung wesentlich für die synaptische Stabilität ist (Smolen, 2015). Die Stabilität dieser Verteilung erfordert, dass es zwischen den Synapsen, welche innerhalb kleiner Cluster organisiert sind, einen Wettbewerb um Ressourcen gibt. Unter diesem Wettbewerb sind die Cluster über Jahre hinaus stabil (Smolen, 2015).
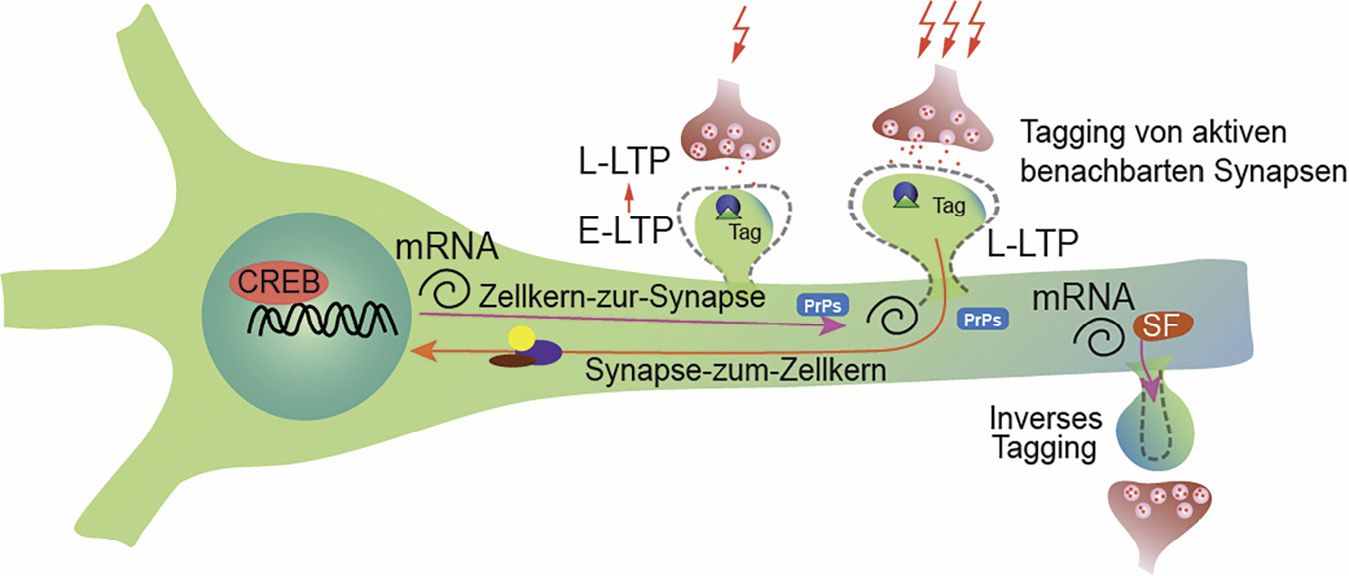
Synaptisches Etikettieren („tagging“) und Einfangen und inverses Etikettieren („inverse tagging“) bei geclusterter Plastizität. Das „Sushi Transportband Modell“ ermöglicht PrPs einzufangen, um LTP an potenzierten Synapsen aufrechtzuerhalten, wohingegen Inkorporation der synaptischen Skalierungsfaktoren an inaktiven Synapsen dazu beiträgt, die Unterschiede der synaptischen Wichtungen aufrechtzuerhalten. SF – Skalierungsfaktoren. CREB – cAMP response element-binding protein.
Unter dem Blickwinkel des Wettbewerbs um Ressourcen ist das inverse synaptische Etikettieren („inverse tagging“) (Okuno et al., 2012) eine faszinierende Möglichkeit, welche die aktivitätsabhängige Genexpression mit der Stabilität der Verteilung der synaptischen Wichtungen verbindet. In einer bahnbrechenden Studie konnten Okuno und Kollegen zeigen, dass die kurz vorher inaktiven Dornfortsätze das IEG-Produkt Arc infolge der Anhäufung von inaktiver CaMKII-β einfangen. Dies resultiert daraufhin in AMPAR-Endocytose und einem weiteren Abschwächen der synaptischen Antworten im Vergleich zu den benachbarten kürzlich vorher aktiven Dornfortsätzen (Okuno et al., 2012). Wir finden die Vorstellung sehr ansprechend, dass Proteinprodukte der IEG, welche beim Herunterskalieren der synaptischen Wichtungen beteiligt sind, mittels Endocytose der AMPAR selektiv auf eine Untergruppe der inaktiven Synapsen wirken (Abbildung 2). In der Tat wurde für IEG, wie zum Beispiel Arc, PLK2 oder Homer 1A, welche alle an Synapsen lokalisiert sind, eine Mitwirkung beim Herunterskalieren der synaptischen Antworten beschrieben (Hayashi et al., 2012). Eine Vorbedingung ist die Anwesenheit eines „tag“, welches IEG – Proteine an inaktiven Dornfortsätzen einfängt, wie es zum Beispiel für CaMKII-β gezeigt wurde. Nachdem das Aufrechterhalten der Unterschiede der synaptischen Wichtungen wesentlich für die geclusterte Plastizität und dem entsprechend für die Stabilität der LTP ist, liegt es nahe zu spekulieren, dass das inverse „tagging“ von IEG eine Verbindung herstellt zwischen aktivitätsabhängiger Genexpression und dem Aufrechterhalten von Langzeitgedächtnis. Parallel dazu kann die lokale Häufigkeit verschiedener PrPs räumlich und zeitlich kontrolliert werden, indem spezifische dendritische mRNAs innerhalb der Ribonukleinsäure-Protein Partikel (RNPs) gezielt in die dendritischen Kompartimente mit hoher oder niedriger synaptischer Aktivität gebracht werden. Dafür hat man den Namen „Sushi Transportband Modell“ geprägt, wo RNPs von multiplen Synapsen gefasst oder freigesetzt werden, wodurch ein weiteres molekulares Hilfsmittel für synaptisches Etikettieren geschaffen wird (Doyle and Kiebler, 2011).
Danksagung: M.M. wird unterstützt durch Beihilfen der Deutschen Forschungsgemeinschaft (DFG Emmy Noether – Programm (Ml 1923/1-1), FOR2419 (MI 1923/2-1), DFG MI1923/3-1). M.R.K wird unterstützt durch Beihilfen der DFG (Kr1879 3-1, 5-1, 6-1, SFB 779 TPB8), Bundesministerium für Bildung und Forschung ‚Energi‘ FKZ: 01GQ1421B, The EU Joint Programme – Neurodegenerative Disease Research (JPND) project STAD, People Programme (Marie Curie Actions) der European Union’s Seventh Framework Programme FP7/2007-2013/ unter REA grant agreement n° [289581] und Leibniz Foundation (Pakt für Forschung 2014, 2015, 2017).
About the authors

Dr. Marina Mikhaylova (geb. 1981) hat Biologie an der Bashkir State Universität in Ufa, Russische Föderation, studiert. Sie hat ihre Promotion in der Forschungsgruppe ‚Neuroplastizität‘ am Leibniz – Institut für Neurobiologie in Magdeburg 2010 verteidigt. Nach einer zweijährigen PostDoc – Zeit in der gleichen Arbeitsgruppe war sie dann von 2012–2015 als Wissenschaftlerin in der ‚Division of Cell Biology‘ an der Universität Utrecht in den Niederlanden tätig. Seit 2015 leitet sie eine durch das Emmy Noether – Programm finanzierte Arbeitsgruppe in Hamburg.

Michael R. Kreutz hat Psychologie, Philosophie und Linguistik an der Universität Münster studiert. Die Doktorarbeit fertigte er an der Ruhr-Universität Bochum an und nach Aufenthalten am MIT in Cambridge, USA, und am Max-Planck-Institut für experimentelle Medizin in Göttingen ist er seit 1993 in Magdeburg tätig, wo er seit 1998 eine Forschungsgruppe am Leibniz-Institut für Neurobiologie leitet. Seit 2015 ist auch Leiter einer Leibniz – Gastgruppe Zentrum für Molekulare Neurobiologie in Hamburg.
Literatur
Attardo, A., Fitzgerald, J.E. und Schnitzer, M.J. (2015). Impermanence of dendritic spines in live adult CA1 hippocampus. Nature 523, 592–596.10.1038/nature14467Search in Google Scholar PubMed PubMed Central
Bosch, M. und Hayashi, Y. (2012). Structural plasticity of dendritic spines. Curr. Opin. Neurobiol. 22, 383–388.10.1016/j.conb.2011.09.002Search in Google Scholar PubMed PubMed Central
Cane, M., Maco, B., Knott, G. und Holtmaat, A. (2014). The relationship between PSD-95 clustering and spine stability in vivo. J. Neurosci. 34, 2075–2086.10.1523/JNEUROSCI.3353-13.2014Search in Google Scholar PubMed PubMed Central
Carlisle, H.J. und Kennedy, M.B. (2005). Spine architecture and synaptic plasticity. Trends Neurosci. 28, 182–187.10.1016/j.tins.2005.01.008Search in Google Scholar PubMed
Dieterich, D.C. und Kreutz, M.R. (2016). Proteomics of the Synapse--A Quantitative Approach to Neuronal Plasticity. Mol. Cell. Proteomics 15, 368–381.10.1074/mcp.R115.051482Search in Google Scholar PubMed PubMed Central
Doyle, M. und Kiebler, M.A. (2011). Mechanisms of dendritic mRNA transport and its role in synaptic tagging. EMBO J. 30, 3540–3552.10.1038/emboj.2011.278Search in Google Scholar PubMed PubMed Central
Frey, U. und Morris, R.G. (1997). Synaptic tagging and long-term potentiation. Nature 385, 533–536.10.1038/385533a0Search in Google Scholar PubMed
Govindarajan, A., Israely, I., Huang, S.Y. und Tonegawa, S. (2011). The dendritic branch is the preferred integrative unit for protein synthesis-dependent LTP. Neuron 69, 132–146.10.1016/j.neuron.2010.12.008Search in Google Scholar PubMed PubMed Central
Govindarajan, A., Kelleher, R.J. und Tonegawa, S. (2006). A clustered plasticity model of long-term memory engrams. Nat. Rev. Neurosci. 7, 575–583.10.1038/nrn1937Search in Google Scholar PubMed
Gu, L., Kleiber, S., Schmid, L., Nebeling, F., Chamoun, M., Steffen, J., Wagner, J. und Fuhrmann, M. (2014). Long-term in vivo imaging of dendritic spines in the hippocampus reveals structural plasticity. J. Neurosci. 34, 13948–13953.10.1523/JNEUROSCI.1464-14.2014Search in Google Scholar PubMed PubMed Central
Hanse, E., Taira, T., Lauri, S. und Groc, L. (2009). Glutamate synapse in developing brain: an integrative perspective beyond the silent state. Trends Neurosci. 32, 532–537.10.1016/j.tins.2009.07.003Search in Google Scholar PubMed
Hanus, C. und Ehlers, M.D. (2016). Specialization of biosynthetic membrane trafficking for neuronal form and function. Curr. Opin. Neurobiol. 39, 8–16.10.1016/j.conb.2016.03.004Search in Google Scholar PubMed
Hanus, C. und Schuman, E.M. (2013). Proteostasis in complex dendrites. Nat. Rev. Neurosci. 14, 638–648.10.1038/nrn3546Search in Google Scholar PubMed
Harvey, C.D. und Svoboda, K. (2007). Locally dynamic synaptic learning rules in pyramidal neuron dendrites. Nature 450, 1195–1200.10.1038/nature06416Search in Google Scholar PubMed PubMed Central
Harvey, C.D., Yasuda, R., Zhong, H. und Svoboda, K. (2008). The spread of Ras activity triggered by activation of a single dendritic spine. Science 321, 136–140.10.1126/science.1159675Search in Google Scholar PubMed PubMed Central
Hayashi, Y., Okamoto, K., Bosch, M. und Futai, K. (2012). Roles of neuronal activity-induced gene products in Hebbian and homeostatic synaptic plasticity, tagging, and capture. Adv. Exp. Med. Biol. 970, 335–354.10.1007/978-3-7091-0932-8_15Search in Google Scholar PubMed
Holtmaat, A.J., Trachtenberg, J.T., Wilbrecht, L., Shepherd, G.M., Zhang, X., Knott, G.W. und Svoboda, K. (2005). Transient and persistent dendritic spines in the neocortex in vivo. Neuron 45, 279–291.10.1016/j.neuron.2005.01.003Search in Google Scholar PubMed
Kaushik, R., Grochowska, K.M., Butnaru, I. und Kreutz, M.R. (2014). Protein trafficking from synapse to nucleus in control of activity-dependent gene expression. Neuroscience 280, 340–350.10.1016/j.neuroscience.2014.09.011Search in Google Scholar PubMed
Kelleher, R.J., 3rd, Govindarajan, A., Jung, H.Y., Kang, H. und Tonegawa, S. (2004). Translational control by MAPK signaling in long-term synaptic plasticity and memory. Cell 116, 467–479.10.1016/S0092-8674(04)00115-1Search in Google Scholar
Kleindienst, T., Winnubst, J., Roth-Alpermann, C., Bonhoeffer, T. und Lohmann, C. (2011). Activity-dependent clustering of functional synaptic inputs on developing hippocampal dendrites. Neuron 72, 1012–1024.10.1016/j.neuron.2011.10.015Search in Google Scholar PubMed
Konietzny, A., Bär, J. und Mikhaylova, M. (2017). Dendritic Actin Cytoskeleton: Structure, Functions, and Regulations. Front. Cell. Neurosci. 11, 147.10.3389/fncel.2017.00147Search in Google Scholar PubMed PubMed Central
Lee, K.F., Soares, C., Thivierge, J.P. und Beique, J.C. (2016). Correlated Synaptic Inputs Drive Dendritic Calcium Amplification and Cooperative Plasticity during Clustered Synapse Development. Neuron 89, 784–799.10.1016/j.neuron.2016.01.012Search in Google Scholar PubMed
Makino, H. und Malinow, R. (2011). Compartmentalized versus global synaptic plasticity on dendrites controlled by experience. Neuron 72, 1001–1011.10.1016/j.neuron.2011.09.036Search in Google Scholar PubMed PubMed Central
Matsuzaki, M., Honkura, N., Ellis-Davies, G.C. und Kasai, H. (2004). Structural basis of long-term potentiation in single dendritic spines. Nature 429, 761–766.10.1038/nature02617Search in Google Scholar PubMed PubMed Central
Mikhaylova, M., Bera, S., Kobler, O., Frischknecht, R. und Kreutz, M.R. (2016). A Dendritic Golgi Satellite between ERGIC and Retromer. Cell Rep 14, 189–199.10.1016/j.celrep.2015.12.024Search in Google Scholar PubMed
Murakoshi, H., Wang, H. und Yasuda, R. (2011). Local, persistent activation of Rho GTPases during plasticity of single dendritic spines. Nature 472, 100–104.10.1038/nature09823Search in Google Scholar PubMed PubMed Central
Okuno, H., Akashi, K., Ishii, Y., Yagishita-Kyo, N., Suzuki, K., Nonaka, M., Kawashima, T., Fujii, H., Takemoto-Kimura, S., Abe, M. et al. (2012). Inverse synaptic tagging of inactive synapses via dynamic interaction of Arc/Arg3.1 with CaMKIIbeta. Cell 149, 886–898.10.1016/j.cell.2012.02.062Search in Google Scholar PubMed PubMed Central
Pozo, K. und Goda, Y. (2010). Unraveling mechanisms of homeostatic synaptic plasticity. Neuron 66, 337–351.10.1016/j.neuron.2010.04.028Search in Google Scholar PubMed PubMed Central
Rosenberg, T., Gal-Ben-Ari, S., Dieterich, D.C., Kreutz, M.R., Ziv, N.E., Gundelfinger, E.D. und Rosenblum, K. (2014). The roles of protein expression in synaptic plasticity and memory consolidation. Front. Mol. Neurosci. 7, 86.10.3389/fnmol.2014.00086Search in Google Scholar PubMed PubMed Central
Smolen, P. (2015). Modeling maintenance of long-term potentiation in clustered synapses: long-term memory without bistability. Neural Plast. 2015, 185410.10.1155/2015/185410Search in Google Scholar PubMed PubMed Central
Takahashi, N., Kitamura, K., Matsuo, N., Mayford, M., Kano, M., Matsuki, N. und Ikegaya, Y. (2012). Locally synchronized synaptic inputs. Science 335, 353–356.10.1126/science.1210362Search in Google Scholar PubMed
van Bommel, B. und Mikhaylova, M. (2016). Talking to the neighbours: The molecular and physiological mechanisms of clustered synaptic plasticity. Neurosci. Biobehav. Rev. 71, 352–361.10.1016/j.neubiorev.2016.09.016Search in Google Scholar PubMed
Winnubst, J. und Lohmann, C. (2012). Synaptic clustering during development and learning: the why, when, and how. Front. Mol. Neurosci. 5, 70.10.3389/fnmol.2012.00070Search in Google Scholar PubMed PubMed Central
Zhai, S., Ark, E.D., Parra-Bueno, P. und Yasuda, R. (2013). Long-distance integration of nuclear ERK signaling triggered by activation of a few dendritic spines. Science 342, 1107–1111.10.1126/science.1245622Search in Google Scholar PubMed PubMed Central
Anmerkung
Übersetzung der englischen Version des Artikels online verfügbar unter https://doi.org/10.1515/nf-2018-A006
© 2018 Walter de Gruyter GmbH, Berlin/Boston
Articles in the same Issue
- Titelseiten
- Übersichtsartikel
- Hippokampale Langzeitpotenzierung (LTP) – Geschichte, Gegenwart und Zukunft
- Hippocampal long-term potentiation (LTP) – past, present and future
- Langzeitpotenzierung im Hippokampus: Entdeckung, Mechanismen und Funktion
- Long-term potentiation in the hippocampus: discovery, mechanisms and function
- Die Regulation der hippokampalen Informationsenkodierung durch metabotrope Glutamatrezeptoren
- Regulation of hippocampal information encoding by metabotopic glutamate receptors
- Geclusterte Plastizität bei Langzeitpotenzierung: Wie starke Synapsen bestehen bleiben, um Langzeitgedächtnis aufrechtzuerhalten
- Clustered plasticity in Long-Term Potentiation: How strong synapses persist to maintain long-term memory
- Hippocampale synaptische Plastizität bei neurodegenerativen Erkrankungen: Aβ, Tau und darüber hinaus
- Hippocampal synaptic plasticity in neurodegenerative diseases: Aß, tau and beyond
- Die Analyse synaptischer Plastizität auf Einzelzellebene mit Hilfe der STDP
- Analyzing synaptic plasticity at the single cell level with STDP
- Institutsvorstellung
- Die Neurowissenschaftliche Gesellschaft ist Gründungsmitglied des German Brain Council
- DFG-Graduiertenkolleg 1957 „Adipocyte-Brain Crosstalk“
- Nachrichten
- Jugend forscht – Sonderpreis der Neurowissenschaftlichen Gesellschaft 2018
Articles in the same Issue
- Titelseiten
- Übersichtsartikel
- Hippokampale Langzeitpotenzierung (LTP) – Geschichte, Gegenwart und Zukunft
- Hippocampal long-term potentiation (LTP) – past, present and future
- Langzeitpotenzierung im Hippokampus: Entdeckung, Mechanismen und Funktion
- Long-term potentiation in the hippocampus: discovery, mechanisms and function
- Die Regulation der hippokampalen Informationsenkodierung durch metabotrope Glutamatrezeptoren
- Regulation of hippocampal information encoding by metabotopic glutamate receptors
- Geclusterte Plastizität bei Langzeitpotenzierung: Wie starke Synapsen bestehen bleiben, um Langzeitgedächtnis aufrechtzuerhalten
- Clustered plasticity in Long-Term Potentiation: How strong synapses persist to maintain long-term memory
- Hippocampale synaptische Plastizität bei neurodegenerativen Erkrankungen: Aβ, Tau und darüber hinaus
- Hippocampal synaptic plasticity in neurodegenerative diseases: Aß, tau and beyond
- Die Analyse synaptischer Plastizität auf Einzelzellebene mit Hilfe der STDP
- Analyzing synaptic plasticity at the single cell level with STDP
- Institutsvorstellung
- Die Neurowissenschaftliche Gesellschaft ist Gründungsmitglied des German Brain Council
- DFG-Graduiertenkolleg 1957 „Adipocyte-Brain Crosstalk“
- Nachrichten
- Jugend forscht – Sonderpreis der Neurowissenschaftlichen Gesellschaft 2018