Die Analyse synaptischer Plastizität auf Einzelzellebene mit Hilfe der STDP
-
Elke Edelmann
Elke Edelmann studierte Biologie an den Universitäten Hohenheim und Tübingen. Nach dem Abschluss ihrer Doktorarbeit in Gravitationsbiologie an der Universität Hohenheim, fokussierte sie sich auf lern- und gedächtnisrelevante Funktionen des Hippokampus. Im Institut für Physiologie der Universität in Kiel wurde sie zur Elektrophysiologin ausgebildet. Nach einem Wechsel an das Institut für Physiologie der Otto-von-Guericke-Universität in Magdeburg, etablierte sie dort die Spike timing-dependent plasticity-Methode im Hippokampus. 2016 übernahm sie die Arbeitsgruppe „Hippocampale Synaptische Plastizität“ am Institut für Physiologie, wo ihr Team und sie auf dem Gebiet „Synaptische Plastizität und deren Regulation durch Neuromodulatoren und Mediatoren“ forschen. Ihre derzeitige Arbeit wird durch Drittmittel der DFG, dem Land Sachsen-Anhalt und dem Europäischen Regionalen Entwicklungsfond unterstützt.
 and
Volkmar Leßmann
and
Volkmar Leßmann
Volkmar Leßmann studierte Biochemie an der Leibniz – Universität Hannover (bis 1990). Er führte seine Doktorarbeit am Max-Planck-Institut für Psychiatrie in Martinsried und an der Ruhr-Universität Bochum (RUB, 1993) durch. 2002 habilitierte er in Neurobiochemie an der RUB und war bis 2007 Hochschuldozent an der Johannes-Gutenberg-Universität (JoGU) Mainz. In 2003 erfolgte die Habilitation für Physiologie an der JoGU in Mainz. 2007: Ruf-Annahme auf die W3-Professur für Physiologie an der Otto-von-Guericke Universität Magdeburg, wo er die Leitung des Instituts für Physiologie übernahm (seit 2007). Seine Forschung konzentriert sich auf lern- und gedächtnisrelevante Mechanismen der synaptischen Plastizität, die im Institut auf zellulärer Ebene, auf neuronaler Netzwerkebene und auf Verhaltensebene untersucht werden. Ein besonderer Schwerpunkt liegt dabei auf Mechanismen der BDNF-Sekretion und Neuromodulation der synaptischen Plastizität, sowie auf zellulären Ursachen der Alzheimer-Demenz. Seine Arbeiten werden derzeit durch Drittmittel der DFG, des BMBF und des EU Horizon2020-Programms unterstützt.

Zusammenfassung
Mithilfe des Patch clamp-Verfahrens können die molekularen Prozesse, die der Langzeitpotentierung (LTP) und der Langzeitdepression (LTD) zugrunde liegen, auf der Ebene eines einzelnen postsynaptischen Neurons (Nervenzelle) in akut isolierten Gehirnschnitten untersucht werden. Über die in der Ableitelektrode enthaltene Pipettenlösung können dabei Inhibitoren intrazellulärer Prozesse in das postsynaptische Neuron eingebracht werden, um so die an der synaptischen Plastizität beteiligten Signalwege zu identifizieren. Zur Untersuchung der synaptischen Plastizität wurden in den letzten Jahren zunehmend Protokolle herangezogen, die durch eine minimale Anzahl von synaptischen Stimulationen mit niedriger Frequenz robuste LTP oder LTD auslösen. Zu diesen Stimulationsmustern gehört die sogenannte Spike timing-dependent plasticity (STDP). Sie kann durch wiederholtes nahezu gleichzeitiges Feuern von Aktionspotenzialen (APs) im präsynaptischen und im nachgeschalteten postsynaptischen Neuron induziert werden, wenn diese APs mit einer kurzen zeitlichen Verzögerung von ca. 5–20 ms ausgelöst werden. Während „Vorwärtspaarungen“ mit kurzem positiven Zeitversatz (erst prä- dann postsynaptisches AP) LTP auslösen, führt „Rückwärtspaarung“ (erst post- dann präsynaptisches AP) zu LTD. Zusätzlich zur Abfolge (vorwärts oder rückwärts) und der zeitlichen Verzögerung der APs, ist die Wirkung von neuromodulatorischen Transmittern (z. B. Dopamin, Acetylcholin, Noradrenalin) und die synaptische Freisetzung von intrazellulären Mediatoren der synaptischen Plastizität (z. B. BDNF, Endocannabinoide) kritisch an der Regulation von STDP-Protokollen beteiligt. In diesem Übersichtsartikel fokussieren wir uns auf die Rolle dieser Mediatoren und Modulatoren bei durch STDP ausgelösten synaptischen Plastizitätsphänomenen.
Einleitung
Es wird angenommen, dass Gedächtnisspuren durch zeitlich korreliertes Feuern von APs in einer Gruppe wechselseitig miteinander verbundener Neurone eines synaptischen Netzwerkes gebildet werden. Durch LTP oder LTD können bestimmte synaptische Verbindungen des Netzwerkes entweder verstärkt oder abgeschwächt werden und legen so die Anzahl und die Reihenfolge des Feuerns von APs in den synaptisch verknüpften Neuronen fest. Verschiedene Neuronentypen (erregende und hemmende) sind an diesem aktivitätsabhängigen Netzwerkprozess beteiligt und tragen mit ihren spezifischen Funktionen zum finalen „Erinnerungsbild“ (Engramm) bei (zu Grundlagen der LTP siehe Eingangskapitel). Um die genaue Funktion der unterschiedlichen Zelltypen bei der Gedächtnisbildung zu verstehen, ist es notwendig, synaptische Plastizitätsmechanismen auf der Ebene einzelner Zellen zu untersuchen. LTP kann dabei mit verschiedenen Protokollen ausgelöst werden, die auf hochfrequenter synaptischer Aktivierung (z. B. 100 Hz für mehrere Sekunden) oder auf künstlich herbeigeführter langanhaltender postsynaptischer Depolarisation beruhen (vgl. Meis et al., 2012; Schildt et al., 2013, Edelmann et al., 2014). LTD dagegen wird experimentell typischerweise durch eine langanhaltende niederfrequente (1 Hz) synaptische Stimulation (>500 Wiederholungen über 15 min) induziert. Obwohl die so ausgelösten Prozesse exzellente Modelle zur Untersuchung synaptischer Plastizität darstellen, treten solch langanhaltende und sehr robuste Aktivitätsmuster nur sehr selten unter physiologischen Bedingungen in vivo auf. Im Gegensatz dazu kann die STDP sowohl LTP als auch LTD durch niederfrequente Paarung einer relativ geringen Anzahl von APs in prä- und postsynaptischen Neuronen auslösen (Markram et al., 1997; Bi und Poo, 1998; Campanac und Debanne, 2008). Diese elektrischen Aktivitätsmuster ähneln Rhythmen, die unter anderem im Hippokampus während Lernprozessen in vivo zu beobachten sind (vgl. z. B. Otto et al., 1991). Es erscheint deshalb sinnvoll, die zellulären Mechanismen der eingangs beschriebenen Standard-LTP- und LTD-Modelle mit den entsprechenden Resultaten von STDP-Experimenten zu vergleichen, um Gemeinsamkeiten zu identifizieren.
STDP wurde nicht nur im Hippokampus, sondern auch in vielen anderen Gehirnregionen beobachtet. STDP kann die synaptische Transmissionsstärke von erregenden und hemmenden Synapsen beeinflussen, und sie ist sowohl in vitro als auch in vivo zu beobachten (vgl. Übersichtsartikel Bi und Poo, 2001; Caporale und Dan, 2008; Markram et al., 2011; Feldman, 2012).

Messung von Spike timing-dependent plasticity (STDP) in akut isolierten Schnitten des Hippokampus in An- bzw. Abwesenheit von endogenem Dopamin
A: STDP kann in akuten Hippokampusschnitten an Schaffer-Kollateral-CA1 Synapsen mit der Patch clamp-Technik untersucht werden. Um t-LTP zu induzieren (orange), werden pre before post-Paarungen mit kurzem Zeitintervall ausgelöst. Das Axon des präsynaptischen Neurons wird dabei durch eine extrazelluläre Stimulation der Schaffer-Kollateralen (①) zum Feuern eines AP gebracht, im postsynaptischen Neuron werden Aktionspotenziale über die Patch pipette am Soma ausgelöst (②). T-LTD kann durch post before pre-Paarungen ausgelöst werden (②→①, blaue Box). B: Grafische Darstellung der Ergebnisse verschiedener t-LTP- und t-LTD-Versuche einzelner Zellen in Abhängigkeit des zeitlichen Versatzes (Δt, ms; spike timing: Zeit zwischen prä- und postsynaptischer Aktivierung). Geschlossene Symbole zeigen t-LTP und t-LTD Ergebnisse von einzelnen Patch Clamp-Experimenten bei Vorhandensein von Dopamin (grüne Symbole) oder in Abwesenheit von Dopamin (rote Symbole). Die typische asymmetrische und bidirektionale STDP-Kurve mit t-LTP und t-LTD kann nur in Gegenwart von Dopamin beobachtet werden. Längere Zeitintervalle (>20ms) führen zu einer Abschwächung der t-LTP (für Vorwärtspaarungen) und der t-LTD (für Rückwärtspaarungen) auch in der Anwesenheit von Dopamin.
An erregenden, glutamatergen Synapsen kann STDP durch fast simultane Aktivierung des prä- und des postsynaptischen Neurons induziert werden. „Vorwärtspaarung“ mit kurzem positiven zeitlichen Versatz (d. h. das präsynaptisches Neuron feuert vor dem postsynaptischen Neuron, ∆t= +5 bis + 20 ms) führt zur zeitabhängigen LTP (engl. t-LTP), während „Rückwärtspaarungen“ mit kurzem zeitlichen negativen Versatz (postsynaptisches Neuron feuert vor dem präsynaptischen Neuron, ∆t= –5 bis –20 ms; vgl. Abb. 1) zu t-LTD führt. T-LTP und t-LTD können entweder durch Veränderung der postsynaptischen Empfindlichkeit (z. B. bei t-LTP: Einbau zusätzlicher, bei t-LTD: Wegnahme vorhandener Glutamat-Rezeptoren in der postsynaptischen Membran) oder durch erleichterte (bei t-LTP) bzw. verminderte Transmitterfreisetzung (bei t-LTD) aus der Präsynapse verwirklicht werden. Eine Verlängerung der zeitlichen Verzögerung zwischen prä- und postsynaptischer Aktivierung führt zu einer raschen Abnahme der t-LTP oder t-LTD Amplitude (vgl. Abb. 1). Schon eine geringfügige Abwandlung des STDP-Protokolls (z. B. hinsichtlich der Anzahl der prä- und postsynaptischen Aktivierungen oder dem Wert für ∆t) kann die Effizienz und den Mechanismus der Expression der t-LTP oder t-LTD gravierend verändern (siehe Abb. 2). Hieraus resultiert eine Fülle an Möglichkeiten zur Erzeugung synaptischer Verstärkungen oder Abschwächungen mittels STDP (vgl. Edelmann et al., 2015; Costa et al., 2017). Auf der Ebene eines einzelnen Neurons bedeutet dies, dass unterschiedliche präsynaptische Eingänge (bestehend aus jeweils 5–10 ultrastrukturellen Synapsen) eines Neurons (mit insgesamt bis zu 10.000 Synapsen) zu mehreren Gedächtnisspuren beitragen können, die durch jeweils andere Gruppen synchron aktivierter Neurone eines Netzwerkes repräsentiert werden. Ein einzelnes Neuron kann zur Integration in verschiedene solcher synchronisiert feuernder Sub-Netzwerke durch LTP- oder LTD-Prozesse an bestimmten synaptischen Eingängen herangezogen werden (Abb. 3). In dieser Hinsicht sind also individuelle Neurone integraler Bestandteil mehrerer Gedächtnisspuren, die innerhalb eines neuronalen Netzwerks durch synchrones Feuern verschiedener Subgruppen von Neuronen abgelegt werden können. Dieser Mechanismus erklärt die enorme Speicherkapazität des Hippokampus und anderen Hirnareale (vgl. Edelmann et al., 2017).
Sowohl Acetylcholin (ACh) sekretierende Neurone aus dem Nucleus basalis Meynert (NBM), als auch Dopamin (DA) und Noradrenalin (NA) sekretierende Neurone aus verschiedenen Mittelhirnkernen, wie dem Locus coeruleus (LC) oder dem ventralen tegmentalen Areal (VTA) projizieren mit ihren Axonen in den Hippokampus und regulieren dort die Effizienz von t-LTP und t-LTD. Diese neuromodulatorischen Transmitter (Neuromodulatoren) sind extrazelluläre Botenstoffe, die u. a. auch in der Nähe von glutamatergen Neuronen freigesetzt werden und die Effizienz der t-LTP- oder t-LTD über prä- oder postsynaptische Mechanismen beeinflussen. Im Gegensatz dazu können Neuromediatoren wie Brain-derived neurotrophic factor (BDNF), Stickstoffmonooxid (NO) oder Endocannabinoide (eCBs) in prä- oder postsynaptischen glutamatergen Synapsen selber produziert und als Antwort auf den LTP- oder LTD-Stimulus sezerniert werden. Solche Neuromediatoren initiieren dann prä- und/oder postsynaptische Prozesse, die die Generierung von LTP oder LTD ermöglichen. Gemeinsam regulieren Neuromodulatoren und Neuromediatoren somit die Ausbildung synaptischer Plastizität sowohl auf Netzwerkniveau, als auch auf der Ebene einzelner Synapsen einer Zelle. Für STDP wurde gezeigt, dass entweder ein konstanter Tonus oder eine phasische Erhöhung der Konzentration von Neuromodulatoren im extrazellulären Raum t-LTP und t-LTD regulieren (z. B. Zhang et al., 2009; Lisman et al., 2010; Edelmann und Lessmann, 2011). Während einige Neuromodulatoren das Auftreten von t-LTP begünstigen, unterstützen andere Neuromodulatoren die t-LTD (vgl. Huang et al., 2012). Interessanterweise können diese Neuromodulatoren t-LTP oder t-LTD auch pro- oder retroaktiv beeinflussen. Wird das entsprechende neuromodulatorische System z. B. kurzfristig und transient vor der eigentlichen STDP-Induktion aktiviert (proaktive Wirkung; Huang und MacNamara, 2012), wird die Entstehung und der Erhalt der t-LTP- erleichtert. In ähnlicher Weise führt die Freisetzung eines Neuromodulators mehrere Minuten nach dem STDP-Stimulus an einer Synapse (retroaktive Wirkung) ebenfalls zu einer verbesserten t-LTP (Brzosko et al., 2015). Diese neuromodulatorischen Prozesse werden häufig als eine „Plastizität höherer Ordnung“ oder „Metaplastizität“ bezeichnet (Abraham 2008). Vergleichbare pro- und retroaktive Wirkungen von Neuromodulatoren konnten auch in Lernversuchen in vivo beobachtet werden (z. B. Kempadoo et al., 2016; Takeuchi et al., 2016; Moncada, 2017). Es erscheint daher plausibel anzunehmen, dass die oben beschriebenen zellulären Mechnismen der Metaplastizität ein verändertes Lernvermögen bedingen.
Der Einfluss von Dopamin auf STDP
Es ist seit Langem bekannt, dass Dopamin (DA) synaptische Plastizität im Hippokampus moduliert (Frey et al., 1991; Huang und Kandel, 1995; zusammengefasst in Edelmann und Lessmann, 2018). Die DA-Wirkung wird dabei durch verschiedene G-Protein-gekoppelte Rezeptoren (D1- bzw. D2-Rezeptoren) des prä- oder postsynaptischen Neurons vermittelt (z. B. Yao et al., 2008, Tritsch und Sabatini, 2012). Die Aktivierung von D1-Rezeptoren führt u. a. zu einer Stimulation des cAMP/PKA-Signalwegs, wohingegen D2-Rezeptoren den cAMP/PKA – Signalweg hemmen (Beaulieu und Gainetdinov, 2012). Durch die Aktivierung unterschiedlicher DA-Rezeptor-gekoppelter Signalwege kann DA verschiedene t-LTP-Formen, die durch unterschiedliche Protokolle induziert werden, unterstützen (Zhang et al., 2009; Edelmann und Lessmann, 2011, 2013; vgl. Abb. 1). Ob dabei bestimmte t-LTP-Formen nur durch D1- oder D2-Rezeptoren oder aber durch gleichzeitige Aktivierung beider Rezeptorsysteme zusammen unterstützt werden, ist derzeit noch unklar. Im Gegensatz zum gut dokumentierten Effekt der DA-Sekretion auf synaptische Plastizität, wird sowohl das Ursprungsgebiet der dopaminergen Axone (z. B. VTA, LC) als auch die Verteilung dieser DA-Fasern auf die verschiedenen Subregionen (z. B. CA1, Stratum radiatum, Stratum oriens, CA3) entlang der longitudinalen Achse des Hippokampus noch sehr kontrovers diskutiert (vgl. Übersichtsartikel Edelmann und Lessmann, 2018).
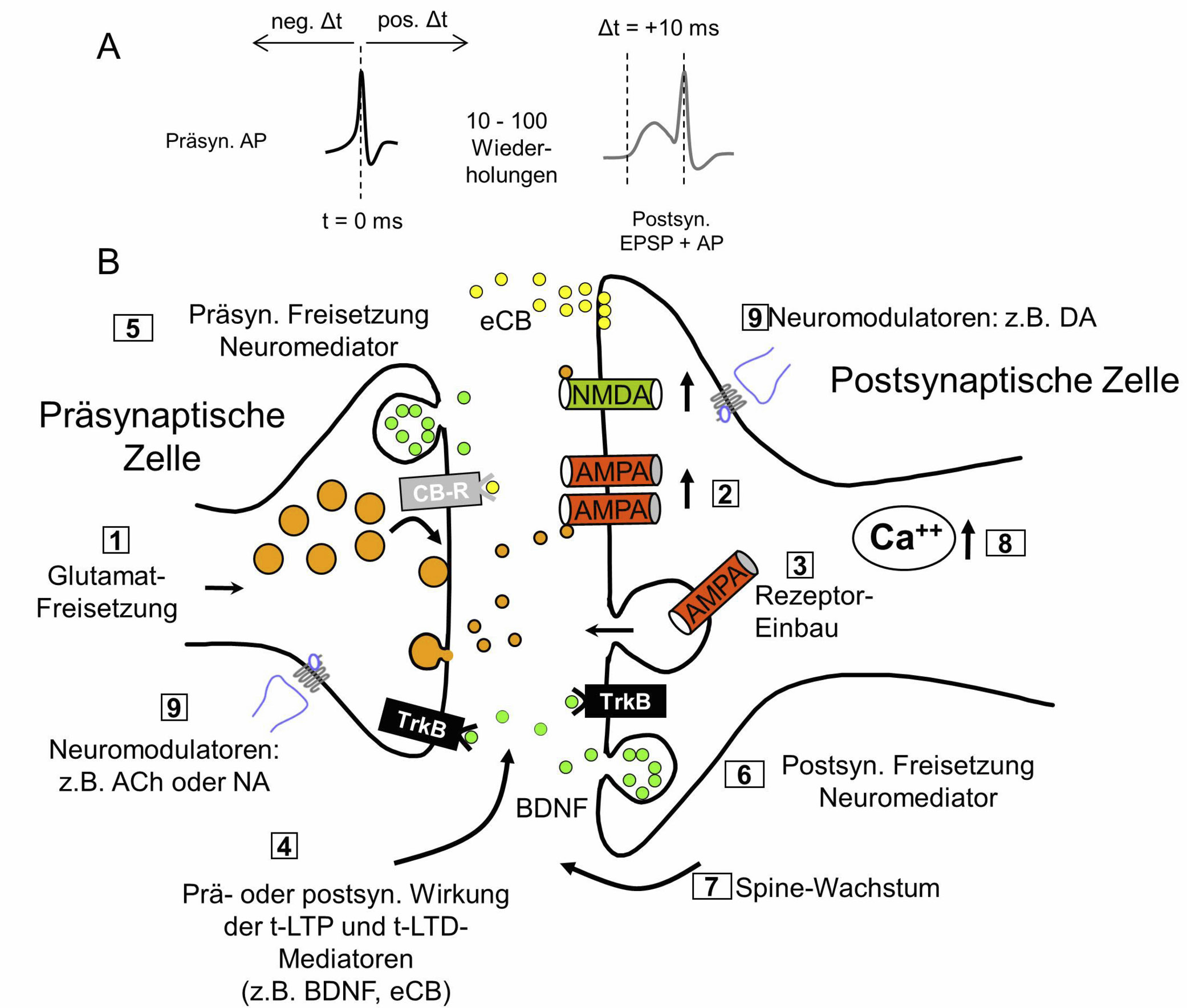
Prä- und Postsynaptische Effekte in t-LTP und t-LTD
A: Typisches STDP-Paradigma für die Induktion von t-LTP. B: Die Zahlen beziehen sich auf die molekularen Mechanismen der prä- oder postsynaptischen Veränderungen während t-LTP oder t-LTD. Präsynaptische Effekte: Erhöhte oder verminderte Freisetzung von Glutamat (①). Postsynaptische Effekte: vermehrte/verminderte AMPA-Rezeptorfunktion (②) oder veränderter AMPA-Rezeptor-Einbau (③). LTP/LTD-Induktion durch aktivitätsabhängige Sekretion von Neuromediatoren (④)(⑤)(⑥). Spine-Wachstum (⑦). Zentrale Rolle des postsynaptischen Ca2+-Anstiegs für die Induktion von t-LTP oder t-LTP (⑧). Die Sekretion von Neuromodulatoren, die relativ weit entfernt von der untersuchten Synapse stattfindet (Volume transmission), kann das Auslösen von t-LTP oder t-LTD begünstigen (⑨). Minimale Änderungen der Wiederholungen synaptischer Stimulationen oder im zeitlichen Versatz zwischen prä- und postsynaptischen APs haben starken Einfluss auf die Lokalisation (prä- oder postsynaptisch) der Expression von t-LTP oder t-LTD, und auf die zugrundeliegenden molekularen Mechanismen.
STDP und Endocannabinoide
Endocannabinoide (eCBs; körpereigene Lipidmediatoren) sind bedeutsame Regulatoren für die Netzwerkaktivität im Hippokampus. Ursprünglich wurden eCBs als Neuromediatoren identifiziert, die nach langanhaltender Depolarisation Ca2+-abhängig von postsynaptischen Strukturen glutamaterger Neurone freigesetzt werden. Folgerichtig können eCBs auch als sogenannte retrograde Botenstoffe auf das präsynaptische Neuron wirken und dort – je nach Typ der Synapse – entweder die Transmittersekretion GABAerger Interneurone oder glutamaterger Neurone reduzieren (Ohno-Shosaku et al., 2002; siehe Übersichtsartikel Xu und Cheng, 2015; Lupica et al., 2017). Der erstgenannte Mechanismus wird als DSI (depolarisationsinduzierte Verminderung der Hemmung, engl. depolarisation induced suppression of inhibition), der zweite Effekt als DSE (depolarisationsinduzierte Verminderung der Erregung, engl. depolarisation induced suppression of excitation) bezeichnet. Dieser präsynaptische Effekt der eCBs wird über G-protein gekoppelte Cannabinoid-Rezeptoren (CB1 und CB2) vermittelt. Aktuell ist nur wenig über den Einfluss von eCB-Signalkaskaden auf die STDP im Hippokampus bekannt. Bisher wurde lediglich eine CB-Rezeptor-abhängige Expression der t-LTD an CA3-CA1 – Synapsen juveniler Mäuse gezeigt (Andrade-Talavera et al., 2016). Die Resultate dieser Studie legen nahe, dass eCBs postsynaptisch auch durch niederfrequente Salven von APs freigesetzt werden können. Durch die retrograde Botenstofffunktion werden dann präsynaptische CB1-Rezeptoren aktiviert, die – durch eine Verringerung der Glutamatfreisetzung – zur Expression der t-LTD führen. Cui und Kollegen konnten auch eine Beteiligung von freigesetztem eCB an der Expression von t-LTP infolge nur weniger Wiederholungen eines STDP-Protokolls an cortico-striatalen Synapsen nachweisen (Cui et al., 2015). Dies zeigt, dass CB-Rezeptoren-Signalwege – je nach verwendetem STDP-Protokoll – tatsächlich auch zur synaptischen Verstärkung (t-LTP) oder zur Abschwächung (t-LTD) der glutamatergen synaptischen Transmission führen können. Während die eCB-abhängige t-LTD durch eine verringerte Glutamat-Sekretion gesteuert wird, wird die eCB-abhängige t-LTP vermutlich durch andere präsynaptische oder sogar postsynaptische Mechanismus vermittelt. So zeigten Dubruc und Kollegen z. B. eine positive Modulation des CA1-Feuerverhaltens durch EPSPs (Dubruc et al., 2013). Derzeit ist allerdings noch ungeklärt, ob diese Effekte indirekt durch eine reduzierte GABAerge Hemmung der synaptischen Netzwerke zustande kommt, oder ob eCBs als direkte Mediatoren an glutamatergen Synapsen t-LTP auslösen können.
Modulation von STDP durch Brain-derived neurotrophic factor über TrkB-Rezeptoren
Im Gegensatz zu den eCBs, wurde für den Brain-derived neurotrophic factor (BDNF)-Signalweg über den TrkB-Tyrosinkinase-Rezeptor eine Beteiligung an klassischer LTP im Hippokampus eindeutig nachgewiesen (vgl. erste Kapitel; für BDNF-vermittelte Plastizität siehe z. B. Gottmann et al., 2009, Park und Poo, 2013; Edelmann et al., 2017). Über die Rolle von BDNF für STDP ist aber nur wenig bekannt. Eine BDNF-abhängige t-LTP wurde allerdings bereits für glutamaterge Synapsen im präfrontalen Kortex, Neokortex und Hippokampus juveniler Mäuse und Ratten gezeigt. Diese Ergebnisse wurden entweder durch chronische Reduktion von BDNF in ko-Mäusen, exogene Applikation von BDNF, Hemmung der BDNF-abhängigen TrkB-Rezeptor-Aktivierung oder durch extrazelluläres Abfangen von sekretiertem körpereigenen BDNF erhalten (siehe z. B. Gordon et al., 2006; Lu et al., 2014; Edelmann et al., 2015). Dabei is zu beachten, dass die zugrunde liegenden elektrophysiologisch messbaren synaptischen Veränderungen in der t-LTP zu einer BDNF-abhängigen strukturellen Modifikation von Synapsen wie z. B. dem Wachstum von postsynaptischen Spines führen (Tanaka et al., 2008; Harward et al., 2016). Diese Befunde unterstreichen, dass synaptisch sekretiertes BDNF die Funktion eines Mediators der t-LTP erfüllt, der auch nachfolgende proteinsyntheseabhängige strukturelle Umbauprozesse an Synapsen vermittelt, die zur Ausbildung langanhaltender Erinnerungen in vivo notwendig sind.
Bemerkenswerterweise werden nicht alle Typen von t-LTP durch DA, eCBs oder BDNF moduliert bzw. mediiert. Vielmehr hängt die Einflussnahme dieser Modulatoren und Mediatoren der t-LTP stark von dem für die STDP-Induktion benutzten Aktivitätsmuster und der genauen dendritischen Lokalisation der jeweils betrachteten Synapse ab (für eine Übersicht siehe Edelmann et al., 2017).
Zusammenfassung und Schlussfolgerung
Spike timing-dependent Plasticity (STDP) dient zur Untersuchung Gedächtnis-relevanter Mechanismen der synaptischen Plastizität auf Einzelzellebene. Je nach gewähltem Stimulationsmuster kann entweder t-LTP oder t-LTD untersucht werden. Durch die Wahl eines spezifischen Protokolls (z. B. mit einer bestimmten Anzahl gepaarter prä- und postsynaptischer Aktivierungen, der Einbeziehung hochfrequenter Aktionspotenzialsalven oder der Wiederholungsrate) können t-LTP und t-LTD mit unterschiedlicher Effizienz und unter Beteiligung verschiedener Signalkaskaden induziert werden. In einem gegebenen postsynaptischen Neuron können verschiedene Gedächtnisspuren entweder unter Beteiligung unterschiedlicher synaptischer Eingänge koexistieren, oder sich gegenseitig beim Einbau hinzukommender neuer Information unterstützend verstärken.
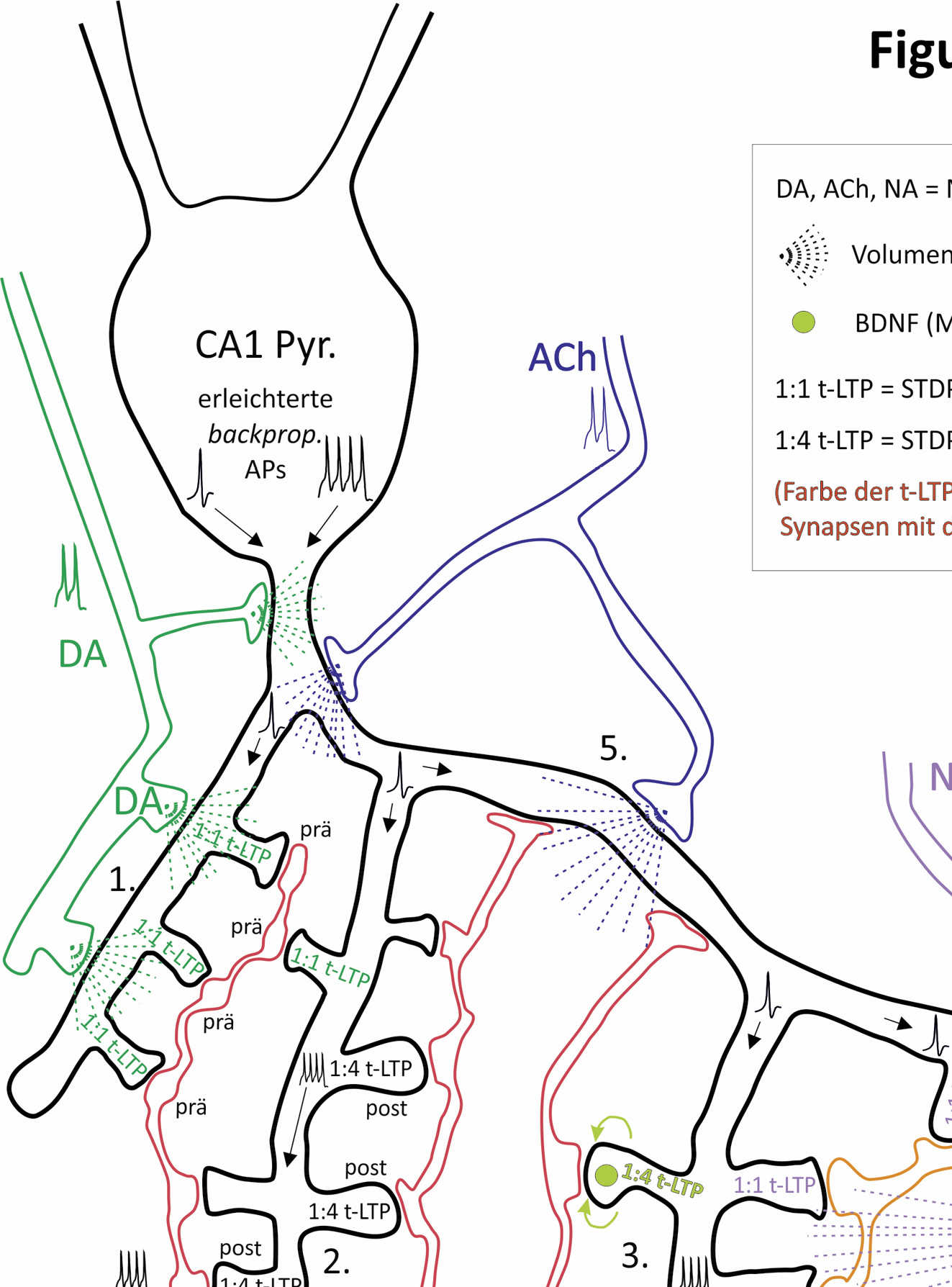
Ausprägung von unterschiedlichen t-LTP-Typen an individuelle Synapsen entlang des Dendritenbaums eines postsynaptischen CA1-Neurons.
CA1-Pyramidenzelle mit (verzweigten) Axonausläufern (Schaffer-Kollateralen, SC) von zwei verschiedenen präsynaptischen, glutamatergen CA3-Neuronen. Hypothese: Die genaue dendritische Lokalisation der Synapse, Aktivität von Neuromodulatoren und das verwendete Stimulationsparadigma bestimmen die Effizienz der t-LTP an dieser synaptischen Kontaktstelle. Synaptische Transmission zwischen jedem präsynaptischen CA3-Neuron und dem CA1-Neuron findet in unserem Beispiel an 7 (SC2) und 16 (SC1) ultrastrukturellen Synapsen statt, die jeweils aus einer präsynaptischen Terminale und einem postsynaptischen Spine bestehen. STDP Paradigmen: 1:1 (1 präsynaptisches: 1 postsynaptisches Aktionspotenzial, AP) oder 1:4 (1 präsynaptisches: 4 postsynaptische APs) gepaart mit entweder hoher (>35) oder niedriger (<15) Anzahl von Wiederholungen. Abhängig vom gewählten STDP-Paradigma wird an diskreten ultrastrukturellen Synapsen entweder prä- oder postsynaptisch exprimierte t-LTP induziert (zur Zuordnung siehe Farbe des Textes in den Spines; leere Spines repräsentieren nicht-potenzierte Synapsen). Unterschiedliche Typen von t-LTP können, durch Volume transmission- vermittelte Wirkung der Neuromodulatoren Dopamin (DA), Noradrenalin (NA) und Acetylcholin (ACh) entweder erleichtert, verstärkt oder gehemmt werden. Durch synaptische BDNF-Freisetzung wird ein spezifischer Typ von t-LTP vermittelt, der durch eine kurze Salve von postsynaptischen Aktionspotenzialen entsteht. Gezeigte Beispiele: (①) Erleichterte 1:1 t-LTP durch DA (präsynaptische Expression). (②) Nicht-modulierte 1:4 t-LTP mit geringer Anzahl von Wiederholungen (postsynaptische Expression). (③) BDNF-vermittelte 1:4 t-LTP mit hoher Wiederholungszahl (postsynaptische Expression). (④) Erleichterte 1:1 t-LTP durch NA. (⑤) Inhibition der 1:1 t-LTP durch ACh. (⑥) Assoziative t-LTP nach Stimulation mit einem 1:4 Paradigma mit wenigen Wiederholungen am synaptischen Eingang SC 2 bei zeitgleichem „Überschwappen“ (Spillover) von ausgeschüttetem BDNF an einer benachbarten Synapse (ausgelöst durch 1:4 Ko-Stimulation des synaptischen Eingangs SC 1 mit hoher Wiederholungszahl). Proximaler Dendrit eines CA1 Neurons: Das Zurückwandern von APs wird durch DA und ACh reguliert. Insgesamt folgt, dass in Abhängigkeit von a) der Wiederholungsrate der synaptischen Stimulation (repräsentiert hohe und niedrige Aktivitätszustände des Gehirns), b) den Eigenschaften des aktivierten glutamatergen Eingangs und c) dem verwendeten STDP-Paradigma, spezifische ultrastrukturelle Synapsen in ihrer Plastizität feinjustiert werden (abgewandelt nach Edelmann et al., Frontiers in Synaptic Neuroscience, 2017).
Die durch STDP induzierte synaptische Plastizität ist deshalb ein vielfältig einsetzbares Instrument zur Untersuchung grundlegender zellulärer Mechanismen von Lern- und Gedächtnisvorgängen auf Einzelzellebene. Dabei gewährt die STDP-Methodik nicht nur einen Einblick in die physiologischen Grundlagen dieser Prozesse im gesunden Zustand, sondern ermöglicht auch die Untersuchung und Entwicklung von Therapieansätzen gegen pathophysiologische synaptische Veränderungen, die z. B. Demenz-Erkrankungen zugrunde liegen.
Glossar
- Demenz
Verschlechterung des mentalen Status mit organischem oder funktionalem Ursprung
- In vitro
in einer physiologischen Umgebung, außerhalb des lebenden Körpers
- In vivo
im lebenden Körper eines Tieres oder eines Menschen
- Gedächtnisengramm
Mittel zur Speicherung eines Gedächtnisinhalts im Gehirn (theoretisches Konzept)
- Synaptische Plastizität
Verstärkung oder Abschwächung der synaptischen Übertragung zwischen verbundenen Nervenzellen (Neuronen)
- Neuromediator
intrazellulär synthetisierter biochemischer Botenstoff, der aus prä- oder postsynaptischen Strukturen bei Auslösen von LTP oder LTD freigesetzt wird und je nach Botenstoff entweder LTP oder LTD auslöst
- Neuromodulator
zellulär synthetisierter Botenstoff (Transmitter, Protein oder Lipid), der von Zellen freigesetzt wird und elektrische oder biochemische Signale von Zellen moduliert, die an der Ausprägung von LTP oder LTD beteiligt sind.
- Patch Clamp-Technik
Elektrophysiologische Methode, um Ionenströme einer Einzelzelle (z. B. eines Neurons) zu messen
- Retrograder Botenstoff
Eine Substanz, die vom postsynaptischen (2.) Neuron sekretiert wird und auf das präsynaptische (1.) Neuron wirkt um synaptische Plastizität auszulösen
- Spike timing-dependent plasticity (STDP)
Ein physiologisch relevanter Prozess, der zur Verstärkung oder Abschwächung der Kommunikation an einer Synapse führt. Die Stärke der Veränderung ist von der relativen Zeit der elektrischen Erregung im prä- und postsynaptischen Neuron abhängig.
- Synaptische Transmission
Signalweitergabe zwischen synaptisch verbundenen Neuronen. Die Signalübertragung erfolgt chemisch durch Transmittersubstanzen, die an spezifische Rezeptoren der postsynaptischen Membran binden und dort eine elektrische Antwort auslösen.
About the authors

Elke Edelmann studierte Biologie an den Universitäten Hohenheim und Tübingen. Nach dem Abschluss ihrer Doktorarbeit in Gravitationsbiologie an der Universität Hohenheim, fokussierte sie sich auf lern- und gedächtnisrelevante Funktionen des Hippokampus. Im Institut für Physiologie der Universität in Kiel wurde sie zur Elektrophysiologin ausgebildet. Nach einem Wechsel an das Institut für Physiologie der Otto-von-Guericke-Universität in Magdeburg, etablierte sie dort die Spike timing-dependent plasticity-Methode im Hippokampus. 2016 übernahm sie die Arbeitsgruppe „Hippocampale Synaptische Plastizität“ am Institut für Physiologie, wo ihr Team und sie auf dem Gebiet „Synaptische Plastizität und deren Regulation durch Neuromodulatoren und Mediatoren“ forschen. Ihre derzeitige Arbeit wird durch Drittmittel der DFG, dem Land Sachsen-Anhalt und dem Europäischen Regionalen Entwicklungsfond unterstützt.

Volkmar Leßmann studierte Biochemie an der Leibniz – Universität Hannover (bis 1990). Er führte seine Doktorarbeit am Max-Planck-Institut für Psychiatrie in Martinsried und an der Ruhr-Universität Bochum (RUB, 1993) durch. 2002 habilitierte er in Neurobiochemie an der RUB und war bis 2007 Hochschuldozent an der Johannes-Gutenberg-Universität (JoGU) Mainz. In 2003 erfolgte die Habilitation für Physiologie an der JoGU in Mainz. 2007: Ruf-Annahme auf die W3-Professur für Physiologie an der Otto-von-Guericke Universität Magdeburg, wo er die Leitung des Instituts für Physiologie übernahm (seit 2007). Seine Forschung konzentriert sich auf lern- und gedächtnisrelevante Mechanismen der synaptischen Plastizität, die im Institut auf zellulärer Ebene, auf neuronaler Netzwerkebene und auf Verhaltensebene untersucht werden. Ein besonderer Schwerpunkt liegt dabei auf Mechanismen der BDNF-Sekretion und Neuromodulation der synaptischen Plastizität, sowie auf zellulären Ursachen der Alzheimer-Demenz. Seine Arbeiten werden derzeit durch Drittmittel der DFG, des BMBF und des EU Horizon2020-Programms unterstützt.
Literatur
Abraham, W. C. (2008). Metaplasticity: tuning synapses and networks for plasticity. Nat. Rev. Neurosci. 9(5), 387.10.1038/nrn2356Search in Google Scholar PubMed
Andrade-Talavera, Y., Duque-Feria, P., Paulsen, O. und A. Rodriguez-Moreno, A. (2016). Presynaptic Spike Timing-Dependent Long-Term Depression in the Mouse Hippocampus. Cereb. Cortex 26(8), 3637–3654.10.1093/cercor/bhw172Search in Google Scholar PubMed PubMed Central
Bi, G.Q. und Poo, M.M. (1998). Synaptic modifications in cultured hippocampal neurons: dependence on spike timing, synaptic strength, and postsynaptic cell type. J. Neurosci. 18(24), 10464–10472.10.1523/JNEUROSCI.18-24-10464.1998Search in Google Scholar PubMed PubMed Central
Bi, G.Q. und Poo, M.M. (2001). Synaptic modification by correlated activity: Hebb’s postulate revisited. Annu. Rev. Neurosci. 24, 139–166.10.1146/annurev.neuro.24.1.139Search in Google Scholar PubMed
Brzosko, Z., Schultz, W. und Paulsen, O. (2015). Retroactive modulation of spike timing-dependent plasticity by dopamine. eLife 4.10.7554/eLife.09685Search in Google Scholar PubMed PubMed Central
Campanac, E. und Debanne, D. (2008). Spike timing-dependent plasticity: a learning rule for dendritic integration in rat CA1 pyramidal neurons. J. Physiol. 586(3), 779–793.10.1113/jphysiol.2007.147017Search in Google Scholar PubMed PubMed Central
Caporale, N. und Dan, Y. (2008). Spike timing-dependent plasticity: a Hebbian learning rule. Annu. Rev. Neurosci. 31, 25–46.10.1146/annurev.neuro.31.060407.125639Search in Google Scholar PubMed
Costa, R. P., Mizusaki, B.E., Sjostrom, P.J. und van Rossum,M.C. (2017). Functional consequences of pre- and postsynaptic expression of synaptic plasticity. Philos. Trans. R. Soc. Lond. B Biol. Sci. 372(1715).10.1098/rstb.2016.0153Search in Google Scholar PubMed PubMed Central
Cui, Y., Paille, V., Xu, H., Genet, S., Delord, B., Fino, E.; Berry, H. und Venance, L. (2015). Endocannabinoids mediate bidirectional striatal spike-timing dependent plasticity. J. Physiol.10.1113/JP270324Search in Google Scholar PubMed PubMed Central
Dubruc, F., Dupret, D. und Caillard, O. (2013): Self-tuning of inhibition by endocannabinoids shapes spike-time precision in CA1 pyramidal neurons. J. Neurophysiol. 110(8), 1930–1944.10.1152/jn.00099.2013Search in Google Scholar PubMed PubMed Central
Edelmann, E., Cepeda-Prado, E., Franck, M., Lichtenecker, P., Brigadski, T. und Lessmann, L. (2015). Theta Burst Firing Recruits BDNF Release and Signaling in Postsynaptic CA1 Neurons in Spike-Timing-Dependent LTP. Neuron 86(4), 1041–1054.10.1016/j.neuron.2015.04.007Search in Google Scholar PubMed
Edelmann, E., Cepeda-Prado, E. und Lessmann, V. (2017). Coexistence of Multiple Types of Synaptic Plasticity in Individual Hippocampal CA1 Pyramidal Neurons. Front. Synaptic Neurosci. 9, 7.10.3389/fnsyn.2017.00007Search in Google Scholar PubMed PubMed Central
Edelmann, E. and Lessmann, V. (2011). Dopamine Modulates Spike Timing-Dependent Plasticity and Action Potential Properties in CA1 Pyramidal Neurons of Acute Rat Hippocampal Slices. Front. Synaptic Neurosci. 3, 6.10.3389/fnsyn.2011.00006Search in Google Scholar PubMed PubMed Central
Edelmann, E. und Lessmann, V. (2013). Dopamine regulates intrinsic excitability thereby gating successful induction of spike timing-dependent plasticity in CA1 of the hippocampus. Front. Neurosci. 7, 25.10.3389/fnins.2013.00025Search in Google Scholar PubMed PubMed Central
Edelmann, E. und Lessmann, V. (2018). Dopaminergic innervation and modulation of hippocampal networks. Cell Tissue Res. doi: 10.1007/s00441-018-2800-710.1007/s00441-018-2800-7Search in Google Scholar PubMed
Edelmann, E., Lessmann, V. und Brigadski, T. (2014). Pre- and postsynaptic twists in BDNF secretion and action in synaptic plasticity. Neuropharmacology 76 Pt C, 610–627.10.1016/j.neuropharm.2013.05.043Search in Google Scholar PubMed
Feldman, D.E. (2012). The spike-timing dependence of plasticity. Neuron 75(4), 556–571.10.1016/j.neuron.2012.08.001Search in Google Scholar PubMed PubMed Central
Frey, U., Matthies, H., Reymann, K.G. and Matthies, H. (1991). The effect of dopaminergic D1 receptor blockade during tetanization on the expression of long-term potentiation in the rat CA1 region in vitro. Neurosci. Lett. 129(1), 111–114.10.1016/0304-3940(91)90732-9Search in Google Scholar PubMed
Gordon, U., Polsky, A. und Schiller, J. (2006). Plasticity compartments in basal dendrites of neocortical pyramidal neurons. J. Neurosci. 26(49), 12717–12726.10.1523/JNEUROSCI.3502-06.2006Search in Google Scholar PubMed PubMed Central
Gottmann, K., Mittmann, T. und Lessmann, V. (2009). BDNF signaling in the formation, maturation and plasticity of glutamatergic and GABAergic synapses. Exp. Brain Res. 199(3–4), 203–234.10.1007/s00221-009-1994-zSearch in Google Scholar PubMed
Hansen, N. (2017). The Longevity of Hippocampus-Dependent Memory Is Orchestrated by the Locus Coeruleus-Noradrenergic System. Neural Plast. 2017, 2727602.10.1155/2017/2727602Search in Google Scholar PubMed PubMed Central
Hansen, N. und Manahan-Vaughan, D. (2014). Dopamine D1/D5 receptors mediate informational saliency that promotes persistent hippocampal long-term plasticity. Cereb. Cortex 24(4), 845–858.10.1093/cercor/bhs362Search in Google Scholar PubMed PubMed Central
Huang, S., Trevino, M., He, K., Ardiles, A., Pasquale, R., Guo, Y., Palacios, A., Huganir, R. und Kirkwood, A. (2012). Pull-push neuromodulation of LTP and LTD enables bidirectional experience-induced synaptic scaling in visual cortex. Neuron 73(3), 497–510.10.1016/j.neuron.2011.11.023Search in Google Scholar PubMed PubMed Central
Huang, Y.Y. und Kandel, E.R. (1995). D1/D5 receptor agonists induce a protein synthesis-dependent late potentiation in the CA1 region of the hippocampus. Proc. Natl. Acad. Sci. U.S.A. 92(7): 2446–2450.10.1073/pnas.92.7.2446Search in Google Scholar PubMed PubMed Central
Huang, Y.Z. und McNamara, J.O. (2012). Neuroprotective effects of reactive oxygen species mediated by BDNF-independent activation of TrkB. J. Neurosci. 32(44), 15521–15532.10.1523/JNEUROSCI.0755-12.2012Search in Google Scholar PubMed PubMed Central
Kempadoo, K.A., Mosharov, E.V., Choi, S.J., Sulzer, D. und Kandel, E.R. (2016). Dopamine release from the locus coeruleus to the dorsal hippocampus promotes spatial learning and memory. Proc. Natl. Acad. Sci. U.S.A. 113(51): 14835–14840.10.1073/pnas.1616515114Search in Google Scholar PubMed PubMed Central
Lisman, J., Grace, A.A. und Duzel, E. (2011). A neoHebbian framework for episodic memory; role of dopamine-dependent late LTP. Trends Neurosci. 34(10), 536–547.10.1016/j.tins.2011.07.006Search in Google Scholar PubMed PubMed Central
Lu, H., Park, H. und Poo, M.M. (2014). Spike-timing-dependent BDNF secretion and synaptic plasticity. Philos. Trans. R. Soc. Lond. B Biol. Sci. 369(1633), 20130132.10.1098/rstb.2013.0132Search in Google Scholar PubMed PubMed Central
Lupica, C.R., Hu, Y., Devinsky, O. und Hoffman, A.F. (2017). Cannabinoids as hippocampal network administrators. Neuropharmacology 124, 25–37.10.1016/j.neuropharm.2017.04.003Search in Google Scholar PubMed
Markram, H., Gerstner, W. und Sjostrom, P.J. (2011). A history of spike-timing-dependent plasticity. Front. Synaptic Neurosci. 3, 4.10.3389/fnsyn.2011.00004Search in Google Scholar PubMed PubMed Central
Markram, H., Lubke, J., Frotscher, M. und Sakmann, B. (1997). Regulation of synaptic efficacy by coincidence of postsynaptic APs and EPSPs. Science 275(5297), 213–215.10.1126/science.275.5297.213Search in Google Scholar PubMed
Meis, S., Endres, T. und V. Lessmann, V. (2012). Postsynaptic BDNF signalling regulates long-term potentiation at thalamo-amygdala afferents. J. Physiol. 590(Pt 1), 193–208.10.1113/jphysiol.2011.220434Search in Google Scholar PubMed PubMed Central
Moncada, D. (2017). Evidence of VTA and LC control of protein synthesis required for the behavioral tagging process. Neurobiol. Learn. Mem. 138, 226–237.10.1016/j.nlm.2016.06.003Search in Google Scholar PubMed
Ohno-Shosaku T, Tsubokawa, H., Mizushima, I., Yoneda, N., Zimmer, A. und Kano, M. (2002). Presynaptic cannabinoid sensitivity is a major determinant of depolarization-induced retrograde suppression at hippocampal synapses. J. Neurosci. 15;22(10), 3864–3872.10.1523/JNEUROSCI.22-10-03864.2002Search in Google Scholar PubMed PubMed Central
Otto, T., Eichenbaum, H., Wiener, S.I. und Wible, C.G. (1991). Learning-related patterns of CA1 spike trains parallel stimulation parameters optimal for inducing hippocampal long-term potentiation. Hippocampus 1(2), 181–192.10.1002/hipo.450010206Search in Google Scholar PubMed
Park, H. und Poo, M.M. (2013). Neurotrophin regulation of neural circuit development and function. Nat. Rev. Neurosci. 14(1), 7–23.10.1038/nrn3379Search in Google Scholar PubMed
Schildt, S., Endres, T., Lessmann, V. und Edelmann, E. (2013). Acute and chronic interference with BDNF/TrkB-signaling impair LTP selectively at mossy fiber synapses in the CA3 region of mouse hippocampus. Neuropharmacology 71, 247–254.10.1016/j.neuropharm.2013.03.041Search in Google Scholar PubMed
Takeuchi, T., Duszkiewicz, A.J., Sonneborn, A., Spooner, P.A., Yamasaki, M., Watanabe, M., Smith, C.C., Fernandez, G., Deisseroth, K., Greene, R.W. und Morris, R.G. (2016). Locus coeruleus and dopaminergic consolidation of everyday memory. Nature 537(7620), 357–362.10.1038/nature19325Search in Google Scholar PubMed PubMed Central
Tanaka, J., Horiike, Y., Matsuzaki, M., Miyazaki, T., Ellis-Davies, G.C. und Kasai, H. (2008). Protein synthesis and neurotrophin-dependent structural plasticity of single dendritic spines. Science 319(5870), 1683–1687.10.1126/science.1152864Search in Google Scholar PubMed PubMed Central
Tritsch, N.X. und Sabatini, B.L. (2012). Dopaminergic modulation of synaptic transmission in cortex and striatum. Neuron 76(1), 33–50.10.1016/j.neuron.2012.09.023Search in Google Scholar PubMed PubMed Central
Xu, J.Y. und Chen, C. (2015). Endocannabinoids in synaptic plasticity and neuroprotection. Neuroscientist 21(2), 152–168.10.1177/1073858414524632Search in Google Scholar PubMed PubMed Central
Yao, W.D., Spealman, R.D. und Zhang, J. (2008). Dopaminergic signaling in dendritic spines. Biochem. Pharmacol. 75(11), 2055–2069.10.1016/j.bcp.2008.01.018Search in Google Scholar PubMed PubMed Central
Zhang, J.C., Lau, P.M. und Bi, G.Q. (2009). Gain in sensitivity and loss in temporal contrast of STDP by dopaminergic modulation at hippocampal synapses. Proc. Natl. Acad. Sci. U.S.A. 106(31): 13028–13033.10.1073/pnas.0900546106Search in Google Scholar PubMed PubMed Central
Anmerkung
Übersetzung der englischen Version des Artikels online verfügbar unter https://doi.org/10.1515/nf-2017-A064
© 2018 Walter de Gruyter GmbH, Berlin/Boston
Articles in the same Issue
- Titelseiten
- Übersichtsartikel
- Hippokampale Langzeitpotenzierung (LTP) – Geschichte, Gegenwart und Zukunft
- Hippocampal long-term potentiation (LTP) – past, present and future
- Langzeitpotenzierung im Hippokampus: Entdeckung, Mechanismen und Funktion
- Long-term potentiation in the hippocampus: discovery, mechanisms and function
- Die Regulation der hippokampalen Informationsenkodierung durch metabotrope Glutamatrezeptoren
- Regulation of hippocampal information encoding by metabotopic glutamate receptors
- Geclusterte Plastizität bei Langzeitpotenzierung: Wie starke Synapsen bestehen bleiben, um Langzeitgedächtnis aufrechtzuerhalten
- Clustered plasticity in Long-Term Potentiation: How strong synapses persist to maintain long-term memory
- Hippocampale synaptische Plastizität bei neurodegenerativen Erkrankungen: Aβ, Tau und darüber hinaus
- Hippocampal synaptic plasticity in neurodegenerative diseases: Aß, tau and beyond
- Die Analyse synaptischer Plastizität auf Einzelzellebene mit Hilfe der STDP
- Analyzing synaptic plasticity at the single cell level with STDP
- Institutsvorstellung
- Die Neurowissenschaftliche Gesellschaft ist Gründungsmitglied des German Brain Council
- DFG-Graduiertenkolleg 1957 „Adipocyte-Brain Crosstalk“
- Nachrichten
- Jugend forscht – Sonderpreis der Neurowissenschaftlichen Gesellschaft 2018
Articles in the same Issue
- Titelseiten
- Übersichtsartikel
- Hippokampale Langzeitpotenzierung (LTP) – Geschichte, Gegenwart und Zukunft
- Hippocampal long-term potentiation (LTP) – past, present and future
- Langzeitpotenzierung im Hippokampus: Entdeckung, Mechanismen und Funktion
- Long-term potentiation in the hippocampus: discovery, mechanisms and function
- Die Regulation der hippokampalen Informationsenkodierung durch metabotrope Glutamatrezeptoren
- Regulation of hippocampal information encoding by metabotopic glutamate receptors
- Geclusterte Plastizität bei Langzeitpotenzierung: Wie starke Synapsen bestehen bleiben, um Langzeitgedächtnis aufrechtzuerhalten
- Clustered plasticity in Long-Term Potentiation: How strong synapses persist to maintain long-term memory
- Hippocampale synaptische Plastizität bei neurodegenerativen Erkrankungen: Aβ, Tau und darüber hinaus
- Hippocampal synaptic plasticity in neurodegenerative diseases: Aß, tau and beyond
- Die Analyse synaptischer Plastizität auf Einzelzellebene mit Hilfe der STDP
- Analyzing synaptic plasticity at the single cell level with STDP
- Institutsvorstellung
- Die Neurowissenschaftliche Gesellschaft ist Gründungsmitglied des German Brain Council
- DFG-Graduiertenkolleg 1957 „Adipocyte-Brain Crosstalk“
- Nachrichten
- Jugend forscht – Sonderpreis der Neurowissenschaftlichen Gesellschaft 2018