Analyzing synaptic plasticity at the single cell level with STDP
-
Elke Edelmann
Elke Edelmann studied biology at the Universities of Hohenheim and Tübingen. After receiving her PhD in Gravitational biology at the University of Hohenheim, she changed her focus from vestibular function in motion sickness to learning and memory related functions of the hippocampus. She was trained as an electrophysiologist at the Institute of Physiology of the University Kiel. Later on, she moved from Kiel to the Otto-von-Guericke University in Magdeburg starting to establish spike timing-dependent plasticity (STDP) to elicit hippocampal synaptic plasticity. In 2016, she became head of the research group “Hippocampal synaptic plasticity” at the Institute of Physiology, where she and her team focus on regulation of synaptic plasticity (e. g. STDP) by dopamine and BDNF. Her work is currently supported by grants from the DFG, the State Saxony-Anhalt and the European Regional Developmental Fund.
 und
Volkmar Leßmann
und
Volkmar Leßmann
Volkmar Leßmann studied biochemistry at the Leibniz University Hannover (until 1990). He completed his PhD thesis at the Max Planck Institute for Psychiatry in Martinsried and at the Ruhr University in Bochum (RUB, 1993). In 2002, he habilitated in Neurobiochemistry at RUB, before becoming an associate professor at the Johannes Gutenberg-University in Mainz, where he further qualified as professor for Physiology. In 2007, he accepted a W3 professorship position at the Otto-von-Guericke-University Magdeburg where he is appointed as head of the Institute of Physiology. His research is focused on learning and memory relevant mechanisms of synaptic plasticity which are investigated at the cellular, network and behavioral level, with a special focus on processes depending on BDNF secretion. His work is currently funded by grants from the DFG, BMBF, and EU Horizon2020.

Abstract
Using patch clamp recordings in acutely isolated brain slices allows to investigate molecular processes that are involved in long-term potentiation (LTP) and long-term depression (LTD) at the level of a single postsynaptic neuron. Via the pipette solution in the recording pipette, it is possible to apply inhibitors of signaling cascades selectively into the postsynaptic neuron to unravel the molecular mechanisms of synaptic plasticity. In recent years, LTP research has been increasingly focused on induction protocols for LTP and LTD that rely on a minimal number of repeated synaptic stimulations at low frequency to induce synaptic plasticity. This is where spike timing-dependent plasticity (STDP) comes into play. STDP can be induced by repetitive pairings of action potential firing in presynaptic (first neuron) – and postsynaptic (second synaptically connected) neurons, that occurs with a delay of roughly 5–20 ms in forward or backward manner. While forward pairing with short positive time delays (“pre before post”) leads to LTP, backward pairing (“post before pre”) leads to LTD. In addition to the exact sequence and timing of pre- and postsynaptic spiking, the presence of neuromodulatory transmitters in the extracellular space (e. g., dopamine, acetylcholine, noradrenaline) and the synaptic release of intercellular mediators of synaptic plasticity (e. g., BDNF, endocannabinoids) critically regulate the outcome of STDP protocols. In this review we focus on the role of these mediators and modulators of synaptic plasticity in STDP.
Introduction
Storage of memory traces is thought to rely on correlated firing of a specific set of simultaneously activated cells in networks of interconnected groups of neurons. The connecting synapses are either strengthened or weakened by LTP or LTD mechanisms. Different types of neurons (excitatory and inhibitory neurons) are involved in these network processes and contribute distinct aspects to the final memory engram formed by activity dependent changes of synaptic transmission (for LTP, see first chapters). To distinguish and examine the different contributions of the diverse neuron types to these learning processes, we need to take a deeper look and analyze synaptic plasticity mechanisms at the level of single neurons. LTP is commonly induced with various induction protocols that rely on high frequency synaptic activation or rather artificial long-lasting postsynaptic depolarization (see e. g., Meis et al. 2012, Schildt et al. 2013, Edelmann et al. 2014), while LTD is typically induced with a very high repeat number of (>500 over 15 min) synaptic activation at low frequency (1 Hz). Although these protocols are excellent models to test molecular mechanisms of synaptic plasticity, such patterns and rhythms of activity are seldom observed under physiological conditions in vivo. In contrast, spike timing-dependent plasticity (STDP) protocols lead to synaptic plasticity after eliciting modest numbers of action potentials in pre- and postsynaptic neurons delivered at low frequency (Markram et al., 1997; Bi and Poo, 1998; Campanac and Debanne, 2008), thereby more closely resembling the rhythms or patterns of electrical activity observed e. g. in hippocampal neurons during learning in vivo (e. g. Otto et al., 1991). It seems therefore adequate to correlate cellular mechanisms of standard LTP and LTD protocols with the respective results of STDP experiments to test whether they recruit common pathways. Besides the hippocampus, STDP was observed in many other brain regions. It can affect synaptic strength of excitatory and inhibitory synapses, and has been described to exist in vitro as well as in vivo (reviewed in e. g., Bi and Poo 2001, Caporale and Dan 2008, Markram et al., 2011; Feldman 2012).
STDP at glutamatergic synapses (compare Fig. 1) can be evoked by nearly coincident activation of the pre- and the postsynaptic partner neuron, with either short positive intervals (i. e. presynaptic before postsynaptic firing; ∆t= +5 to +20 ms) leading to timing-dependent LTP (t-LTP), or negative delays (i. e. post- before presynaptic firing; ∆t= –5 to –20 ms) yielding timing-dependent long-term depression (t-LTD). t-LTP and t-LTD can be either expressed by changed postsynaptic responsiveness (through incorporation of additional (for t-LTP) or removal of present (for t-LTD) glutamate receptors) or enhanced/reduced transmitter release. If only slightly longer time delays between firing in pre- and postsynaptic neurons are reached LTP and LTD rapidly decline in magnitude (compare Fig. 1). Even subtle changes in the pattern and number of repetitions of coincident synaptic stimulation have been described to change the magnitude and to switch expression mechanisms of t-LTP and t-LTD (Fig. 2) thereby representing a plethora of possible mechanisms how strengthening or weakening of synaptic transmission can be accomplished (compare e. g., Edelmann et al. 2015, Costa et al. 2017). At the single cell level this means that different groups (5–10 synapses) of an individual neuron’s synapses (overall up to 10000) can contribute to several memory engrams stored in distinct networks of synchronously activated neurons, via induction of t-LTP at these sets of synapses of this neuron (Fig. 3). In this way, individual neurons are an integral part of multiple “memory traces” in a network thereby significantly increasing the storage capacity of the hippocampus and other brain areas (discussed in Edelmann et al. 2017).
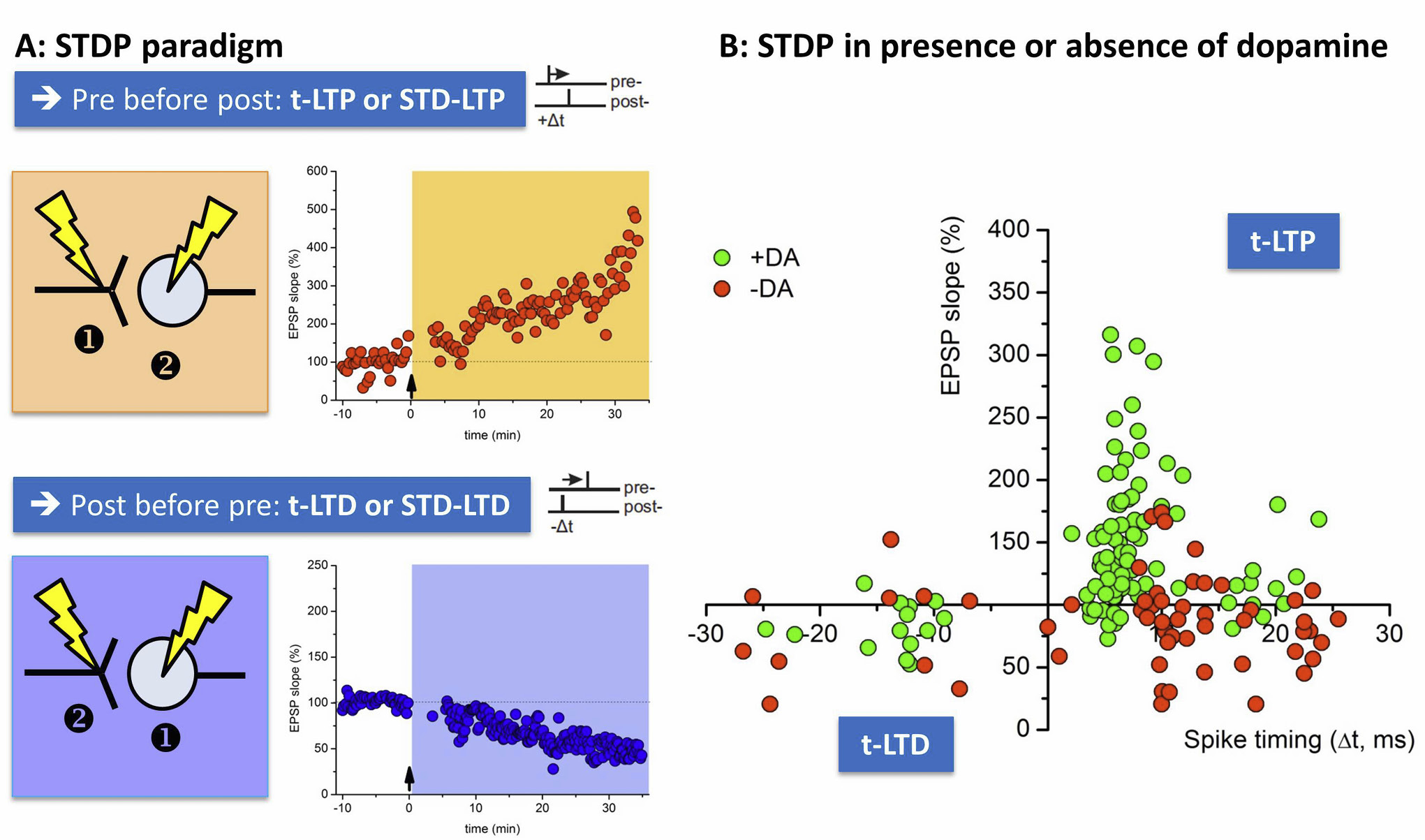
Assessing spike timing-dependent plasticity (STDP) in acute hippocampal slices in the presence or absence of dopamine.
A: STDP can be induced in acute hippocampal slices at Schaffer collateral CA1 synapses with patch clamp techniques. In order to evoke t-LTP (orange) pre-before post pairings are performed by nearly coincident activation of the presynaptic axon through extracellular stimulation of Schaffer collaterals (①) and action potential induction at the soma of the postsynaptic CA1 neuron (②). T-LTD can be elicited by post-pre pairings (②→①, blue boxes). B: The graph shows the outcome of STDP induced by pre-post and post-pre pairings with different spike timing intervals (∆t, ms; spike timing: time delay between pre- and postsynaptic activation). Closed Symbols show t-LTP or t-LTD results for single patch clamp experiments either in the presence (green symbols) or absence of dopamine (red symbols). Note that the typical asymmetric and bidirectional STDP rule can only be observed in the presence of dopamine, where negative spike timings yield t-LTD and positive spike timings induce t-LTP at short time delays. Prolonged time intervals (>20ms) lead to a decline in t-LTP (for positive spike timings) or t-LTD (for negative spike timings) in the presence of dopamine.
Acetylcholine (ACh) releasing neurons originating from the Nucleus basalis Meynert (NBM), as well as dopamine (DA) and noradrenaline (NA) releasing neurons projecting from midbrain structures like the locus coeruleus (LC) and the ventral tegmental area (VTA) to the hippocampus regulate the efficacy of t-LTP and t-LTD in the hippocampus. These neuromodulatory transmitters (neuromodulators) are extracellular messengers that are released in the neighborhood of glutamatergic synapses and render pre- and postsynaptic elements of these excitatory synapses more susceptible to undergo t-LTP or t-LTD. In contrast, neuromediators like brain-derived neurotrophic factor (BDNF), nitric oxide (NO) or endocannabinoids (eCBs) can be synthetized in either pre- or postsynaptic neurons of a glutamatergic synapse where they are released in response to LTP induction paradigms. These neuromediators initiate the pre- and/or postsynaptic processes underlying the long-lasting changes in synaptic efficacy leading to t-LTP and t-LTD. Neuromodulators and neuromediators crucially shape synaptic plasticity at the network level and also at the level of single cells. For STDP it was shown that either a constant tone or a phasic rise of neuromodulatory transmitters in the extracellular space regulate timing-dependent (t-)LTP and t-LTD (e. g., Zhang et al. 2009, Lisman et al., 2010, Edelmann and Lessmann 2011). While some neuromodulators push the system towards LTP, others pull it towards LTD (e. g., Huang et al. 2012). Interestingly, neuromodulators can proactively or retroactively shape LTP or LTD induced by a specific pattern of pre- and postsynaptic activation. If a neuromodulatory transmitter is transiently elevated in the vicinity of a glutamatergic synapse well before STDP induction (i. e. proactive action) t-LTP was facilitated (compare Huang and McNamara, 2012). Likewise, transient activation of a neuromodulatory transmitter system several minutes after STDP induction (i. e., retroactive action) also favors t-LTP (Brzosko et al. 2015). This process is often described as a “higher order of plasticity” or “metaplasticity” (Abraham 2008). Similar proactive or retroactive actions of neuromodulators have also been observed in behavioral experiments testing learning and memory (e. g., Kempadoo et al. 2016, Takeuchi et al. 2016, Moncada 2017) suggesting that synaptic metaplasticity impacts learning capabilities.

Pre- and postsynaptic effects in t-LTP and t-LTD.
A: Example of a typical STDP paradigm yielding timing-dependent LTP (t-LTP). B: The different numbers refer to molecular mechanisms underlying either pre- or postsynaptic changes mediating t-LTP or t-LTD. Presynaptic effects: increased or decreased glutamate release (①). Postsynaptic effects: increased/decreased AMPA receptor function ② or AMPA receptor insertion ③. LTP/LTD induced by activity-dependent release of neuromediators (④⑤⑥). Spine growth ⑦. Central role of postsynaptic Ca2+ increase to induce t-LTP or t-LTP ⑧. Neuromodulators released at rather distant sites (“volume transmission”) can gate t-LTP or t-LTD ⑨. Even subtle changes in repeat number or timing of pre- and postsynaptic APs can dramatically change the locus of t-LTP or t-LTD expression, and the underlying molecular mechanisms.
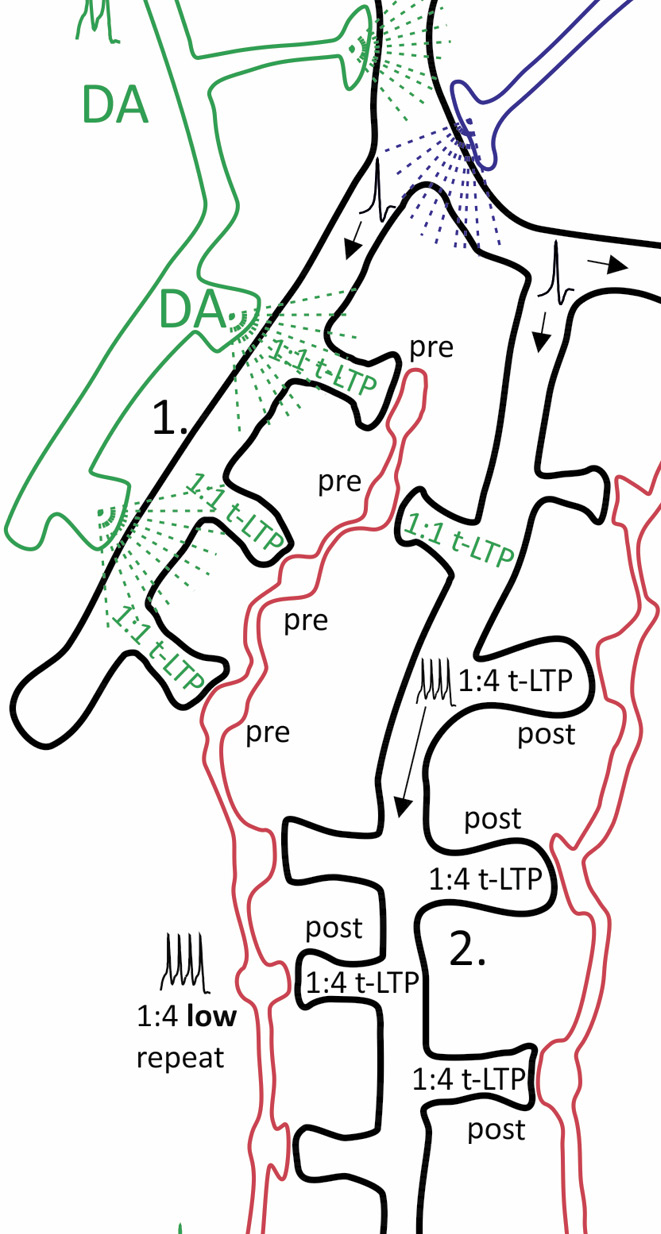
Individual synapses along the dendritic tree of a single postsynaptic CA1 pyramidal neuron can establish distinct forms of t-LTP.
CA1 pyramidal neuron with (branched) axonal projections (Schaffer collaterals, SC) from 2 distinct presynaptic glutamatergic CA3 neurons. Hypothesis: dendritic location of synapses, activity of neuromodulators and stimulation paradigm determine efficacy of t-LTP at this synaptic site. Synaptic transmission between each presynaptic CA3 neuron and the CA1 neuron takes place potentially at 7 (SC2) and 16 (SC1) ultrastructural synapses, each comprising one presynaptic bouton and one postsynaptic spine. STDP paradigms: 1:1 (1 presynaptic: 1 postsynaptic action potential) or 1:4 (1 presynaptic: 4 postsynaptic APs) pairing with either high (>35) or low (<15) number of repeats. Depending on STDP paradigm distinct subsets of ultrastructural synapses undergo pre- or postsynaptically expressed t-LTP (indicated by distinct text colors in the spines; empty spines represent non-potentiated synapses). The different t-LTP types can either be facilitated, enhanced, or inhibited, respectively, by volume transmission of the neuromodulators dopamine (DA), noradrenaline (NA) and acetylcholine (ACh). In addition, local BDNF release mediates t-LTP specifically in response to short bursts of postsynaptic APs. Specific examples shown: ① DA facilitated 1:1 t-LTP (presynaptic expression). ② Non-modulated 1:4 low repeat t-LTP (postsynaptic expression). ③ BDNF-mediated 1:4 high repeat t-LTP (postsynaptic expression). ④ NA-gated 1:1 t-LTP. ⑤ 1:1 t-LTP inhibited by ACh. ⑥ Associative t-LTP in response to 1:4 low repeat paradigm at SC 2 input by locally restricted BDNF spillover from 1:4 high repeat co-stimulated at SC 1 input. Proximal CA1 neuron dendrite: backpropagation of APs is regulated by DA and ACh. Thus, depending on repeat numbers of STDP paradigms (representing high and low activity states of the brain), activated glutamatergic input, and STDP paradigm distinct ultrastructural synapses are fine-tuned in plasticity (Modified from Edelmann et al., Frontiers in synaptic Neuroscience, 2017).
The influence of dopamine on STDP
Dopamine (DA) is known since long to modulate synaptic plasticity in the hippocampus (Frey et al. 1991, Huang and Kandel 1995, reviewed in Edelmann and Lessmann, 2018). Dopamine exerts its actions through activation of different G protein coupled receptors expressed in presynaptic and postsynaptic neurons (e. g., Yao et al. 2008, Tritsch and Sabatini 2012). Among other effects, activation of D1-like receptors induces elevation of cAMP/PKA pathways, whereas D2-like receptors decrease cAMP/PKA signaling (Beaulieu and Gainetdinov, 2012). By activation of different DA receptor signaling cascades, dopamine can facilitate t-LTP induced by different STDP paradigms (e. g., Zhang et al. 2009, Edelmann and Lessmann 2011, 2013, compare Fig. 1). Whether these actions are mediated by exclusive activation of D1-like or D2-like receptors or whether both receptor systems together are involved to successfully gate t-LTP remains to be elucidated. In contrast to this rather well documented effect of DA on synaptic plasticity, the source of DA (e. g. VTA, LC) that is released in the hippocampus and the meandering innervation pattern of dopaminergic fibers in hippocampal subfields (e. g. CA1, Stratum radiatum, Stratum oriens, CA3) along the longitudinal axis of the hippocampus is still controversially discussed (reviewed in e. g., Edelmann and Lessmann, 2018).
STDP and endocannabinoids
Endocannabinoids (eCBs, endogenous lipid mediators) are essential regulators of hippocampal network activity. eCBs were initially discovered to be released from postsynaptic structures of glutamatergic neurons upon Ca2+ elevation initiated by longer lasting postsynaptic depolarization. Thus, eCBs act as retrograde synaptic messengers that – depending on the transmitter phenotype of the opposed presynaptic terminal – inhibit either GABA release from specific sets of GABAergic interneurons or glutamate release at excitatory synapses (Ohno-Shosaku et al., 2002; for recent reviews see e. g., Xu and Cheng, 2015; Lupica et al. 2017). The former mechanisms is called DSI (depolarization induced suppression of inhibition), the latter was named DSE (depolarization induced suppression of excitation). These presynaptic effects of eCB are mediated by G-protein coupled cannabinoid receptors (CB1 and CB2). Up to date, little is known about eCB signaling in STDP in the hippocampus. ECB signaling was described to be involved in expression of t-LTD at hippocampal CA3-CA1 synapses in juvenile mice (Andrade-Talavera et al. 2016). This result is consistent with eCBs being released from postsynaptic neurons even in response to modest numbers of action potentials fired at low frequency, followed by a retrograde action via presynaptic CB1 receptors to reduce glutamate release, overall leading to expression of t-LTD. Interestingly, Cui and colleagues provided evidence for the involvement of eCB release also in low repeat timing-dependent LTP at cortico-striatal synapses (Cui et al. 2015). Surprisingly, this indicates that eCBs can, depending on the protocol used to induce STDP, contribute either to strengthening (t-LTP) or weakening (t-LTD) of synaptic efficacy at glutamatergic synapses. While eCB-dependent t-LTD seems to be mediated by decreased glutamate release, it is likely that eCB mediated t-LTP might rely on different or even postsynaptic mechanisms. In this respect, Dubruc and colleagues (2013) suggest that eCBs can tune CA1 neurons to more reliably fire action potentials during EPSPs. Overall, it remains to be determined, whether these effects rely on the reduced tone of GABAergic inhibition in the investigated networks or rather on eCBs acting as direct mediators of t-LTP.
Modulation of STDP by brain-derived neurotrophic factor and its cognate TrkB receptor
In contrast to endocannabinoids (eCBs), brain-derived neurotrophic factor (BDNF) signaling via its cognate tyrosine kinase receptor TrkB was clearly demonstrated to mediate classical LTP in the hippocampus (see first chapters; for BDNF actions see e. g., Gottmann et al. 2009, Park and Poo 2013, Edelmann et al. 2017). However, only a very limited number of studies addressed BDNF actions in STDP. BDNF-dependent t-LTP was shown at glutamatergic synapses in prefrontal cortex, neocortex and hippocampus at synapses in juvenile rats and mice. Using either chronic depletion of BDNF in ko mice, application of exogenous BDNF, inhibition of BDNF-dependent activation of TrkB receptor signaling, or scavenging of secreted BDNF, all these studies strongly suggested BDNF-dependent expression of t-LTP (Gordon et al. 2006, Lu et al. 2014, Edelmann et al. 2015). Importantly, electrophysiologically recorded synaptic changes leading to t-LTP are converted also into BDNF-dependent structural remodeling of synapses such as growth of postsynaptic spines (Tanaka et al. 2008; Harward et al., 2016). These results clearly suggest that synaptically secreted BDNF is able to serve the function of a mediator of t-LTP and subsequent protein synthesis-dependent structural remodeling of synapses that underlie long-lasting storage of memories in vivo.
However, it is noteworthy that not all types of t-LTP are gated or mediated by DA, eCBs or BDNF and that the recruitment of specific neuromodulators and mediators in t-LTP critically depends on the activity patterns during STDP induction and the location of synapses along the dendritic tree of postsynaptic neurons (reviewed e. g. in, Edelmann et al. 2017).
Summary and conclusion
Spike timing-dependent plasticity (STDP) is a method to investigate memory relevant mechanisms of synaptic plasticity at single cell level. Depending on the employed paradigm, either t-LTP or t-LTD can be studied. By the choice of a specific paradigm (i. e. the number of repeats of coincident synaptic activation, the inclusion of bursts, or the repeat frequency) t-LTP or t-LTD with different signaling and expression mechanisms will be induced. In a given postsynaptic neuron different types of memory traces can either coexist or support each other to strengthen newly incorporated information.
STDP induced plasticity is therefore a versatile instrument to test and challenge synapses for neuronal mechanisms of learning and memory at the level of a single postsynaptic neuron. Moreover, STDP methods will likely provide insights into physiological mechanisms involved in learning and memory processes in health, and might assist the search for strategies aiming to restore memory deficits resulting from pathophysiological conditions such as dementia.
Glossary
- Dementia
deterioration of mental and emotional state with organic or functional origin
- In vitro
in a physiological environment, outside the living body
- In vivo
in the living body of a human or an animal
- Memory engram
means to store memory in the brain (theoretical concept)
- Synaptic plasticity
Strengthening or weakening of synaptic communication between connected neurons
- Neuromediator
intracellular biochemical messenger that is secreted at synaptic junctions during induction of LTP or LTD, that mediates the synaptic changes leading to LTP or LTD.
- Neuromodulator
chemical substance in the extracellular space that modulates electrical responses or biochemical signaling at a potentiated synapse to change the likelihood of inducing LTP or LTD.
- Patch clamp techniques
electrophysiological method to study ionic currents of a single cell (e. g. a neuron)
- Retrograde messenger
a substance which is released from the postsynaptic (2nd) neuron and travels backwards to induce synaptic plasticity in the presynaptic (1st) neuron
- Spike timing-dependent plasticity (STDP)
A physiologically relevant process to induce strengthening or weakening of a synapse. The direction and the magnitude of change rely on the relative timing of electrical activity in pre- and postsynaptic neurons.
- Synaptic transmission
Signal exchange between synaptically connected neurons via a secreted neurotransmitter substance that elicits an electrical response in the postsynaptic neuron upon binding to a neurotransmitter receptor.
About the authors

Elke Edelmann studied biology at the Universities of Hohenheim and Tübingen. After receiving her PhD in Gravitational biology at the University of Hohenheim, she changed her focus from vestibular function in motion sickness to learning and memory related functions of the hippocampus. She was trained as an electrophysiologist at the Institute of Physiology of the University Kiel. Later on, she moved from Kiel to the Otto-von-Guericke University in Magdeburg starting to establish spike timing-dependent plasticity (STDP) to elicit hippocampal synaptic plasticity. In 2016, she became head of the research group “Hippocampal synaptic plasticity” at the Institute of Physiology, where she and her team focus on regulation of synaptic plasticity (e. g. STDP) by dopamine and BDNF. Her work is currently supported by grants from the DFG, the State Saxony-Anhalt and the European Regional Developmental Fund.

Volkmar Leßmann studied biochemistry at the Leibniz University Hannover (until 1990). He completed his PhD thesis at the Max Planck Institute for Psychiatry in Martinsried and at the Ruhr University in Bochum (RUB, 1993). In 2002, he habilitated in Neurobiochemistry at RUB, before becoming an associate professor at the Johannes Gutenberg-University in Mainz, where he further qualified as professor for Physiology. In 2007, he accepted a W3 professorship position at the Otto-von-Guericke-University Magdeburg where he is appointed as head of the Institute of Physiology. His research is focused on learning and memory relevant mechanisms of synaptic plasticity which are investigated at the cellular, network and behavioral level, with a special focus on processes depending on BDNF secretion. His work is currently funded by grants from the DFG, BMBF, and EU Horizon2020.
Reference List
Abraham, W. C. (2008). Metaplasticity: tuning synapses and networks for plasticity. Nat. Rev. Neurosci. 9(5), 387.10.1038/nrn2356Suche in Google Scholar PubMed
Andrade-Talavera, Y., Duque-Feria, P., Paulsen, O. and Rodriguez-Moreno, A. (2016). Presynaptic Spike Timing-Dependent Long-Term Depression in the Mouse Hippocampus. Cereb Cortex 26(8), 3637–3654.10.1093/cercor/bhw172Suche in Google Scholar PubMed PubMed Central
Bi, G. Q. and Poo, M. M. (1998). Synaptic modifications in cultured hippocampal neurons: dependence on spike timing, synaptic strength, and postsynaptic cell type. J. Neurosci 18(24), 10464–10472.10.1523/JNEUROSCI.18-24-10464.1998Suche in Google Scholar PubMed PubMed Central
Bi, G. Q. and Poo, M. M. (2001). Synaptic modification by correlated activity: Hebb’s postulate revisited. Annu. Rev. Neurosci 24, 139–166.10.1146/annurev.neuro.24.1.139Suche in Google Scholar PubMed
Brzosko, Z., W. Schultz and Paulsen, O. (2015). Retroactive modulation of spike timing-dependent plasticity by dopamine. Elife 4.10.7554/eLife.09685Suche in Google Scholar PubMed PubMed Central
Campanac, E. and Debanne, D. (2008). Spike timing-dependent plasticity: a learning rule for dendritic integration in rat CA1 pyramidal neurons. J. Physiol 586(3), 779–793.10.1113/jphysiol.2007.147017Suche in Google Scholar PubMed PubMed Central
Caporale, N. and Dan, Y. (2008). Spike timing-dependent plasticity: a Hebbian learning rule. Annu. Rev. Neurosci 31, 25–46.10.1146/annurev.neuro.31.060407.125639Suche in Google Scholar PubMed
Costa, R. P., Mizusaki, B. E., Sjostrom, P. J. and van Rossum, M. C. (2017). Functional consequences of pre- and postsynaptic expression of synaptic plasticity. Philos Trans R Soc Lond B Biol Sci 372(1715).10.1098/rstb.2016.0153Suche in Google Scholar PubMed PubMed Central
Cui, Y., Paille, V., Xu, H., Genet, S., Delord, B., Fino, E., Berry, H. and Venance, L. (2015). Endocannabinoids mediate bidirectional striatal spike-timing dependent plasticity. J. Physiol.10.1113/JP270324Suche in Google Scholar PubMed PubMed Central
Dubruc, F., Dupret, D., Caillard, O. (2013): Self-tuning of inhibition by endocannabinoids shapes spike-time precision in CA1 pyramidal neurons. J Neurophysiol. 110(8), 1930–44.10.1152/jn.00099.2013Suche in Google Scholar PubMed PubMed Central
Edelmann, E., Cepeda-Prado, E., Franck, M., Lichtenecker, P., Brigadski, T. and Lessmann, V. (2015). Theta Burst Firing Recruits BDNF Release and Signaling in Postsynaptic CA1 Neurons in Spike-Timing-Dependent LTP. Neuron 86(4), 1041–1054.10.1016/j.neuron.2015.04.007Suche in Google Scholar PubMed
Edelmann, E., Cepeda-Prado, E. and Lessmann, V. (2017). Coexistence of Multiple Types of Synaptic Plasticity in Individual Hippocampal CA1 Pyramidal Neurons. Front Synaptic Neurosci 9: 7.10.3389/fnsyn.2017.00007Suche in Google Scholar PubMed PubMed Central
Edelmann, E. and Lessmann, V. (2011). Dopamine Modulates Spike Timing-Dependent Plasticity and Action Potential Properties in CA1 Pyramidal Neurons of Acute Rat Hippocampal Slices. Front Synaptic. Neurosci 3, 6.10.3389/fnsyn.2011.00006Suche in Google Scholar PubMed PubMed Central
Edelmann, E. and Lessmann, V. (2013). Dopamine regulates intrinsic excitability thereby gating successful induction of spike timing-dependent plasticity in CA1 of the hippocampus. Front Neurosci 7, 25.10.3389/fnins.2013.00025Suche in Google Scholar PubMed PubMed Central
Edelmann, E. and Lessmann, V. (2018). Dopaminergic innervation and modulation of hippocampal networks. Cell Tissue Res. doi: 10.1007/s00441-018-2800-710.1007/s00441-018-2800-7Suche in Google Scholar PubMed
Edelmann, E., Lessmann, V. and Brigadski, T. (2014). Pre- and postsynaptic twists in BDNF secretion and action in synaptic plasticity. Neuropharmacology 76 Pt C, 610–627.10.1016/j.neuropharm.2013.05.043Suche in Google Scholar PubMed
Feldman, D. E. (2012). The spike-timing dependence of plasticity. Neuron 75(4), 556–571.10.1016/j.neuron.2012.08.001Suche in Google Scholar PubMed PubMed Central
Frey, U., Matthies, H., Reymann, K. G. and Matthies, H. (1991). The effect of dopaminergic D1 receptor blockade during tetanization on the expression of long-term potentiation in the rat CA1 region in vitro. Neurosci Lett 129(1), 111–114.10.1016/0304-3940(91)90732-9Suche in Google Scholar PubMed
Gordon, U., Polsky, A. and Schiller, J. (2006). Plasticity compartments in basal dendrites of neocortical pyramidal neurons. J. Neurosci 26(49), 12717–12726.10.1523/JNEUROSCI.3502-06.2006Suche in Google Scholar PubMed PubMed Central
Gottmann, K., Mittmann, T. and Lessmann, V. (2009). BDNF signaling in the formation, maturation and plasticity of glutamatergic and GABAergic synapses. Exp. Brain Res 199(3–4): 203–234.10.1007/s00221-009-1994-zSuche in Google Scholar PubMed
Hansen, N. (2017). The Longevity of Hippocampus-Dependent Memory Is Orchestrated by the Locus Coeruleus-Noradrenergic System. Neural Plast 2017: 2727602.10.1155/2017/2727602Suche in Google Scholar PubMed PubMed Central
Hansen, N. and Manahan-Vaughan, D. (2014). Dopamine D1/D5 receptors mediate informational saliency that promotes persistent hippocampal long-term plasticity. Cereb. Cortex 24(4), 845–858.10.1093/cercor/bhs362Suche in Google Scholar PubMed PubMed Central
Huang, S., Trevino, M., He, K., Ardiles, A., Pasquale, R., Guo, Y., Palacios, A., Huganir, R. and Kirkwood, A. (2012). Pull-push neuromodulation of LTP and LTD enables bidirectional experience-induced synaptic scaling in visual cortex. Neuron 73(3), 497–510.10.1016/j.neuron.2011.11.023Suche in Google Scholar PubMed PubMed Central
Huang, Y. Y. and Kandel, E. R. (1995). D1/D5 receptor agonists induce a protein synthesis-dependent late potentiation in the CA1 region of the hippocampus. Proc Natl Acad Sci U S A 92(7), 2446–2450.10.1073/pnas.92.7.2446Suche in Google Scholar PubMed PubMed Central
Huang, Y. Z. and McNamara, J. O. (2012). Neuroprotective effects of reactive oxygen species mediated by BDNF-independent activation of TrkB. J. Neurosci 32(44), 15521–15532.10.1523/JNEUROSCI.0755-12.2012Suche in Google Scholar PubMed PubMed Central
Kempadoo, K. A., Mosharov, E. V., Choi, S. J., Sulzer, D. and Kandel, E. R. (2016). Dopamine release from the locus coeruleus to the dorsal hippocampus promotes spatial learning and memory. Proc Natl Acad Sci U S A 113(51), 14835–14840.10.1073/pnas.1616515114Suche in Google Scholar PubMed PubMed Central
Lisman, J., Grace, A. A. and Duzel, E., (2011). A neoHebbian framework for episodic memory; role of dopamine-dependent late LTP. Trends Neurosci 34(10), 536–547.10.1016/j.tins.2011.07.006Suche in Google Scholar PubMed PubMed Central
Lu, H., Park, H. and Poo, M. M. (2014). Spike-timing-dependent BDNF secretion and synaptic plasticity. Philos. Trans. R. Soc. Lond B Biol. Sci 369(1633), 20130132.10.1098/rstb.2013.0132Suche in Google Scholar PubMed PubMed Central
Lupica, C. R., Hu, Y., Devinsky, O. and Hoffman, A. F. (2017). Cannabinoids as hippocampal network administrators. Neuropharmacology 124, 25–37.10.1016/j.neuropharm.2017.04.003Suche in Google Scholar PubMed
Markram, H., Gerstner, W. and Sjostrom, P. J. (2011). A history of spike-timing-dependent plasticity. Front Synaptic. Neurosci 3, 4.10.3389/fnsyn.2011.00004Suche in Google Scholar PubMed PubMed Central
Markram, H., Lubke, J., Frotscher, M. and Sakmann, B. (1997). Regulation of synaptic efficacy by coincidence of postsynaptic APs and EPSPs. Science 275(5297), 213–215.10.1126/science.275.5297.213Suche in Google Scholar PubMed
Meis, S., Endres, T. and Lessmann, V. (2012). Postsynaptic BDNF signalling regulates long-term potentiation at thalamo-amygdala afferents. J. Physiol 590(Pt 1), 193–208.10.1113/jphysiol.2011.220434Suche in Google Scholar PubMed PubMed Central
Moncada, D. (2017). Evidence of VTA and LC control of protein synthesis required for the behavioral tagging process. Neurobiol Learn Mem 138, 226–237.10.1016/j.nlm.2016.06.003Suche in Google Scholar PubMed
Ohno-Shosaku T., Tsubokawa, H., Mizushima, I., Yoneda, N., Zimmer, A. and Kano, M., (2002). Presynaptic cannabinoid sensitivity is a major determinant of depolarization-induced retrograde suppression at hippocampal synapses. J Neurosci. 15;22(10), 3864–72.10.1523/JNEUROSCI.22-10-03864.2002Suche in Google Scholar PubMed PubMed Central
Otto, T., Eichenbaum, H., Wiener, S. I. and Wible, C. G. (1991). Learning-related patterns of CA1 spike trains parallel stimulation parameters optimal for inducing hippocampal long-term potentiation. Hippocampus 1(2), 181–192.10.1002/hipo.450010206Suche in Google Scholar PubMed
Park, H. and Poo, M. M. (2013). Neurotrophin regulation of neural circuit development and function. Nat. Rev. Neurosci 14(1), 7–23.10.1038/nrn3379Suche in Google Scholar PubMed
Schildt, S., Endres, T., Lessmann, V. and Edelmann, E. (2013). Acute and chronic interference with BDNF/TrkB-signaling impair LTP selectively at mossy fiber synapses in the CA3 region of mouse hippocampus. Neuropharmacology 71, 247–254.10.1016/j.neuropharm.2013.03.041Suche in Google Scholar PubMed
Takeuchi, T., Duszkiewicz, A. J., Sonneborn, A., Spooner, P. A., Yamasaki, M., Watanabe, M., Smith, C. C., Fernandez, G., Deisseroth, K., Greene, R. W. and Morris, R. G. (2016). Locus coeruleus and dopaminergic consolidation of everyday memory. Nature 537(7620), 357–362.10.1038/nature19325Suche in Google Scholar PubMed PubMed Central
Tanaka, J., Horiike, Y., Matsuzaki, M., Miyazaki, T., Ellis-Davies, G. C. and Kasai, H. (2008). Protein synthesis and neurotrophin-dependent structural plasticity of single dendritic spines. Science 319(5870), 1683–1687.10.1126/science.1152864Suche in Google Scholar PubMed PubMed Central
Tritsch, N. X. and Sabatini, B. L. (2012). Dopaminergic modulation of synaptic transmission in cortex and striatum. Neuron 76(1): 33–50.10.1016/j.neuron.2012.09.023Suche in Google Scholar PubMed PubMed Central
Xu, J. Y. and Chen, C. (2015). Endocannabinoids in synaptic plasticity and neuroprotection. Neuroscientist 21(2), 152–168.10.1177/1073858414524632Suche in Google Scholar PubMed PubMed Central
Yao, W. D., Spealman, R. D. and Zhang, J. (2008). Dopaminergic signaling in dendritic spines. Biochem Pharmacol 75(11), 2055–2069.10.1016/j.bcp.2008.01.018Suche in Google Scholar PubMed PubMed Central
Zhang, J. C., Lau, P. M. and Bi, G. Q. (2009). Gain in sensitivity and loss in temporal contrast of STDP by dopaminergic modulation at hippocampal synapses. Proc. Natl. Acad. Sci. U. S. A 106(31), 13028–13033.10.1073/pnas.0900546106Suche in Google Scholar PubMed PubMed Central
Article note
German version available at https://doi.org/10.1515/nf-2017-0064
© 2018 Walter de Gruyter GmbH, Berlin/Boston
Artikel in diesem Heft
- Titelseiten
- Übersichtsartikel
- Hippokampale Langzeitpotenzierung (LTP) – Geschichte, Gegenwart und Zukunft
- Hippocampal long-term potentiation (LTP) – past, present and future
- Langzeitpotenzierung im Hippokampus: Entdeckung, Mechanismen und Funktion
- Long-term potentiation in the hippocampus: discovery, mechanisms and function
- Die Regulation der hippokampalen Informationsenkodierung durch metabotrope Glutamatrezeptoren
- Regulation of hippocampal information encoding by metabotopic glutamate receptors
- Geclusterte Plastizität bei Langzeitpotenzierung: Wie starke Synapsen bestehen bleiben, um Langzeitgedächtnis aufrechtzuerhalten
- Clustered plasticity in Long-Term Potentiation: How strong synapses persist to maintain long-term memory
- Hippocampale synaptische Plastizität bei neurodegenerativen Erkrankungen: Aβ, Tau und darüber hinaus
- Hippocampal synaptic plasticity in neurodegenerative diseases: Aß, tau and beyond
- Die Analyse synaptischer Plastizität auf Einzelzellebene mit Hilfe der STDP
- Analyzing synaptic plasticity at the single cell level with STDP
- Institutsvorstellung
- Die Neurowissenschaftliche Gesellschaft ist Gründungsmitglied des German Brain Council
- DFG-Graduiertenkolleg 1957 „Adipocyte-Brain Crosstalk“
- Nachrichten
- Jugend forscht – Sonderpreis der Neurowissenschaftlichen Gesellschaft 2018
Artikel in diesem Heft
- Titelseiten
- Übersichtsartikel
- Hippokampale Langzeitpotenzierung (LTP) – Geschichte, Gegenwart und Zukunft
- Hippocampal long-term potentiation (LTP) – past, present and future
- Langzeitpotenzierung im Hippokampus: Entdeckung, Mechanismen und Funktion
- Long-term potentiation in the hippocampus: discovery, mechanisms and function
- Die Regulation der hippokampalen Informationsenkodierung durch metabotrope Glutamatrezeptoren
- Regulation of hippocampal information encoding by metabotopic glutamate receptors
- Geclusterte Plastizität bei Langzeitpotenzierung: Wie starke Synapsen bestehen bleiben, um Langzeitgedächtnis aufrechtzuerhalten
- Clustered plasticity in Long-Term Potentiation: How strong synapses persist to maintain long-term memory
- Hippocampale synaptische Plastizität bei neurodegenerativen Erkrankungen: Aβ, Tau und darüber hinaus
- Hippocampal synaptic plasticity in neurodegenerative diseases: Aß, tau and beyond
- Die Analyse synaptischer Plastizität auf Einzelzellebene mit Hilfe der STDP
- Analyzing synaptic plasticity at the single cell level with STDP
- Institutsvorstellung
- Die Neurowissenschaftliche Gesellschaft ist Gründungsmitglied des German Brain Council
- DFG-Graduiertenkolleg 1957 „Adipocyte-Brain Crosstalk“
- Nachrichten
- Jugend forscht – Sonderpreis der Neurowissenschaftlichen Gesellschaft 2018