Regulation of hippocampal information encoding by metabotopic glutamate receptors
-
Denise Manahan-Vaughan
Denise Manahan-Vaughan is a Neurophysiologist, Neuroscientist and Head of the Department of Neurophysiology within the Medical Faculty of the Ruhr University Bochum (www.rub.de/neurophys). She is also director and Dean of Studies of the International Graduate School of Neuroscience (www.rub.de/igsn) and speaker (primary coordinator) of the DFG-funded Collaborative Research Centre on Integration and Representation of Sensory Information Processes at the same university (www.rub.de/sfb874). After completion of her primary degree in Natural Sciences at Trinity College Dublin (TCD), Ireland, she obtained a PhD in Neuropharmacology at TCD and then moved to Germany to the Leibniz Institute for Neurobiology in Magdeburg. Having obtained an Habilitation degree in Physiology from the Otto von Guericke University in Magdeburg, she moved to the Johannes Müller Institute for Physiology at the Charité, Berlin, before moving to Bochum in 2003. Her research focusses on understanding the relationship between hippocampal and cortical synaptic plasticity and long-term associative and spatial memory.


Abstract
The hippocampus supports the acquisition of both spatial representations and long-term spatial memory. This is enabled by a triumvirate of physiological processes comprising information organisation and transfer by means of neuronal oscillations, creation of context-dependent spatial maps by means of place cells, and long-term storage of spatial experience by means of synaptic plasticity. All three processes are enabled by the glutamatergic system. Glutamate binding to ionotropic glutamate receptors enables both fast excitatory synaptic transmission (via AMPA receptors) and the initiation of long-term synaptic storage (via NMDA receptors). But glutamate also binds to metabotropic glutamate (mGlu) receptors. These receptors not only contribute to the stability of hippocampal encoding and the longevity of synaptic plasticity, they can also support synaptic information storage independent of NMDA receptor activation and are important for the acquisition and retention of long-term memory.
Introduction
The hippocampus serves as a central hub for the creation of memories for events (in non-human animals (Eichenbaum, 2017), or episodes (in humans)(Horner & Doeller, 2017), and its prominent role in enabling spatial cognition is likely to support these processes. The hippocampus integrates sensory experience into spatial representations and long-term spatial memory (Manahan-Vaughan, 2017). It does so by enabling long-term alterations of synaptic efficacy in the form of synaptic plasticity (Bliss and Collingridge, 1993; Martin and Buno 2005; Kemp and Manahan-Vaughan 2007; Manahan-Vaughan, 2017), network oscillatory activity (Buzsaki and Draguhn 2004; Hasselmo 2005), and place field formation (O’Keefe and Dostrovsky 1971; Knierim et al. 1995). The neurotransmitter glutamate is of paramount importance for these processes. Glutamate binds to two categories of neurotransmitter receptors, comprising ionotropic and metabotropic glutamate receptors. Ionotropic glutamate receptors are ligand-gated ion channels that include the α-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid (AMPA), kainate and N-methyl-D-aspartate (NMDA) receptors. Whereas AMPA receptors enable fast excitatory synaptic transmission, NMDA receptors are key elements in the induction of synaptic plasticity processes, such as long-term potentiation (LTP) (Bliss et al., this Neuroforum issue). Metabotropic glutamate (mGlu) receptors are G-protein coupled receptors that act via second messenger systems and regulate intracellular levels of adenylyl cyclase or phospholipase C (Table 1). These receptors play a very important role in aspects of hippocampal information processing that relate to the long-term retention of spatial experience (Mukherjee and Manahan-Vaughan, 2012).
Classification of mGlu receptors and synaptic distribution
mGlu Receptor | Subtypes | Hippocampal expression | Coupling |
Group I | mGlu1, mGlu5 | Mainly postsynaptic | Phospholipase C |
Group II | mGlu2 mGlu3 | Mainly presynaptic | Adenylyl cyclase |
Group III | mGlu4,mGlu6*, mGlu7,mGlu8 | Mainly presynaptic | Adenylyl cyclase |
Receptors are classified on the basis of their signal transduction mechanisms and pharmacological properties (see: Mukherjee and Manahan-Vaughan, 2013 for overview). Group I mGlu receptors are positively coupled to phospholipase C and are mainly postsynaptically localised in the hippocampus. Groups II and III receptors are negatively coupled to adenylyl cyclase and presynaptically localised (Ohishi et al., 1993; Mukherjee and Manahan-Vaughan, 2012; Goddyn et al., 2015;Tanabe et al., 1993; Okamoto et al., 1994; Corti et al., 1998). *N. B.: mGlu6 is expressed exclusively in the retina (Nomura et al., 1994).
Place cells, synaptic plasticity, neuronal oscillations
Place cells are pyramidal neurons that are mainly found in the CA1 and CA3 regions of the hippocampus of rodents (Grieves & Jeffery, 2017) and humans (Ekstrom et al., 2003), that exhibit high-frequency discharges when animals traverse a specific location of an environment. The location where the place cell fires, is called the ‘place field’ (O’Keefe and Dostrovsky in 1971) (Fig. 1). The firing behaviour of place cells is specifically related to the context in which the animal finds itself, whereby it can use sensory cues from different modalities as a substrate to create a place map (Zhang and Manahan-Vaughan, 2015). Whereas place fields develop and stabilise while an animal navigates through and becomes familiar with a spatial environment, the long-term retention of spatial experience in the form of long-term memory is enabled by hippocampal synaptic plasticity: it has been shown in rodents, that synaptic plasticity, in the forms of long-term potentiation (LTP) and long-term depression (LTD), that persist for more than 24h, is strongly associated with the acquisition and retention of spatial memory (Kemp and Manahan-Vaughan, 2007, 2008; Manahan-Vaughan, 2017). In fact, in mice, it has been shown that synaptic plasticity can be directly triggered by spatial experience (Goh and Manahan-Vaughan, 2013a).
Neuronal oscillations are an intrinsic component of functional neuronal networks, that in turn support real-time and long-term information encoding related to spatial experience. Modification of hippocampal synaptic strength occurs rapidly during animal behaviour, whereby neuronal oscillations at theta and gamma frequencies play a crucial role in this process (see for review: Buzsáki, 2005; Buzsáki and Draguhn, 2004). Hence, hippocampal theta and gamma oscillations are functionally associated and derive from intrinsic oscillatory properties of principal cells and interneurons, the rhythmic activation of which is driven by intra- and extrahippocampal connections (Bartos et al., 2007). Synaptic plasticity and theta-gamma neuronal oscillations are interdependent: changes in theta-gamma frequency coupling during the induction of LTP predict whether the induction attempt will be successful (Bikbaev and Manahan-Vaughan, 2007, 2008) and applying stimuli on the peak or the trough of hippocampal theta results in the induction of LTP, or LTD, respectively (Hölscher at al, 1997).
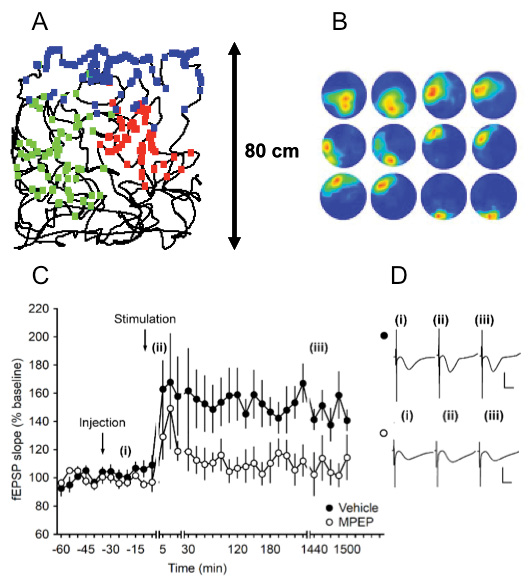
Regulation of hippocampal place fields and synaptic plasticity by mGlu5 receptors
A: The thin black line shows the path taken by a rat as it explored a 80 × 80cm square environment during tetrode recordings from the hippocampal CA1 region.
The green, blue and red dots show the respective regions of the envronment where three different place cells fired, the region covered by one colour corresponds to the place field of that cell.
B: Animals that first showed stable place fields during exploration of a square environment were treated with an mGlu5 receptor antagonist and then allowed to explore a novel (round) environment. Exposure to the same environment one day later was associated with place field remapping, indicating that the stabilisation and consolidation of the place fields was compromised by mGlu5 antagonism. The examples represent (left to right: two exposures to the novel environment on day 1 followed by two exposures to the same environment on day 2. Top to bottom: place fields of place cells recorded from three different animals. (From Zhang and Manahan-Vaughan, 2014).
C: Antagonism of mGlu5 prevents hippocampal LTP that would normally last for longer than 24h in rats.
High-frequency stimulation (HFS) (4 trains of 100 pulses at 100 Hz) of MF-CA3 synapses, elicits LTP (>24h) in vehicle-injected freely behaving animals. Intracerebral application of the mGlu5 antagonist, MPEP (1.8 µg), prior to HFS significantly blocks LTP.
D: Analog responses evoked during LTP experiment
Analogs represent fEPSPs evoked (i) pre-HFS, (ii) 5 min post-HFS and (iii) 24 h post-HFS from) MF-CA3 synapses of vehicle-treated (closed circles i. e. top row) or MPEP-treated animals (open circles i. e. bottom row). (From Hagena and Manahan-Vaughan, 2015).
Involvement of mGlu receptors in place fields, synaptic plasticity and neuronal oscillations
Group I mGlu receptors
Although the involvement of NMDA receptors in the establishment of place fields has been shown (Kentros et al., 1998), few studies have addressed to what extent mGlu receptors contribute to place field generation or stability. In rats, pharmacological antagonism of the mGlu5 receptor prevents long-term stability of place fields and reduces informational content and place cell firing rates in a novel environment (Zhang and Manahan-Vaughan 2014, Fig. 1). This finding creates an intriguing link between spatial mapping by means of place cells and the long-term encoding of spatial experience by means of synaptic plasticity: the mGlu5 receptor is of pivotal importance for both persistent forms of synaptic plasticity and for long-term memory (Hagena and Manahan-Vaughan, 2017). Furthermore, mGlu5 receptors are required for the abovementioned tetanisation-induced changes of theta and gamma oscillations that in turn predict the successful expression of LTP (Bikbaev and Manahan-Vaughan, 2017), and are also required for cell-specific plasticity induced by neuronal oscillations (Zarndadze et al., 2016). Of all of the mGlu receptors, the mGlu5 receptor may be the most important for hippocampal encoding processes: activation of this receptor is required for reference memory (Naie and Manahan-Vaughan, 2004; Manahan-Vaughan and Braunewell, 2005), recognition memory (Marszalek-Grabska et al., 2018) and extinction learning (André et al, 2015). MGlu5 receptors also mediate improvements of LTP that occur following environmental enrichment (Buschler and Manahan-Vaughan, 2017). Activation of the receptor is essential for the expression of forms of LTP and LTD that are facilitated by spatial learning (Popkirov and Manahan-Vaughan, 2011; Goh and Manahan-Vaughan, 2013b; Hagena and Manahan-Vaughan, 2015) and can induce protein-synthesis forms of synaptic plasticity in the absence of NMDA receptor activation (Huber et al., 2001; Naie and Manahan-Vaughan, 2006).
Its counterpart the mGlu1 receptor, is also intrinsically involved in hippocampal synaptic plasticity and memory processes (Naie and Manahan-Vaughan, 2005), including in hippocampus-dependent associative learning forms that are not related to spatial learning (Gil-Sanz et al., 2008). Whereas mGlu5 supports the (late) protein-synthesis phase of LTP and LTD (Balschun and Wetzel, 2002; Naie and Manahan-Vaughan, 2004; Popkirov and Manahan-Vaughan, 2011), mGlu1 receptors support the induction of LTP (Neymann and Manahan-Vaughan 2008; Naie and Manahan-Vaughan, 2005), The mechanism is likely to involve increases in intracellular Ca2+ concentrations, neuronal depolarization, elevations in the frequency of spontaneous inhibitory post-synaptic potentials (Mannaioni et al., 2001), as well as regulation of NMDA receptor currents (Skeberdis et al. 2001) and cycling (Lan et al., 2001).
Group II mGlu receptors
Group I mGlu receptors regulate both hippocampal LTP and LTD, but group II mGlu receptors may only be directly involved in LTD. Antagonism of these receptors prevents the expression of persistent forms of LTD (Manahan-Vaughan, 1997; Kulla et al., 1999) and interestingly also prevents long-term reference memory (Altinbilek and Manahan-Vaughan, 2009). Agonist activation of these receptors can prevent LTP, however, even when ligand doses are used that have no effect on basal synaptic transmission (Kulla et al., 1999). This may relate to regulation of hippocampal excitability: group II receptors are mostly located presynaptically in the hippocampus (Shigemoto et al., 1997) and predominantly subserve an autoreceptor function (Mukherjee and Manahan-Vaughan, 2013). Although their postsynaptic expression is weaker (Petralia et al., 1996), a role for postsynaptic group II mGlu receptors in the regulation of CA3 network activity has been described (Ster et al., 2011) that may be crucial for information processing related to theta activity.
MGlu2 receptor involvement in recognition memory has been proposed (Marszalek-Grabska et al., 2018) and a role for this receptor in spatial working memory has been demonstrated in transgenic mice (de Filippis et al., 2015). Transgenic mice that lack both mGlu2 and mGlu3 receptors are also impaired in spatial working memory tasks (Lyon et al., 2011). These findings in turn provide valuable insights as to the possible relationship between LTP, LTD and components of spatial memory: novel object-place learning, related to recognition memory, triggers LTD in the mouse hippocampus, whereas it has been suggested that the magnitude of LTD can serve as an index of spatial working memory ability (Nakao et al., 2002).
Group III mGlu receptors
Similar to reports with regard to group II mGlu receptors, antagonism of group III mGlu receptors impair the expression of persistent (>24 h) LTD, but not LTP, in both the CA1 region and dentate gyrus (Klausnitzer et al., 2004; Altinbilek and Manahan-Vaughan, 2007). Transgenic mice that lack mGlu7, also exhibit deficits in short-term potentiation (Bushell et al., 2002). Receptor antagonism impairs spatial reference memory in a radial arm maze task (Altinbilek and Manahan-Vaughan, 2007) and both group II and III mGlu receptors have been implicated in the retrieval of context-dependent fear memory (Szapiro et al, 2001) Transgenic mice that lack mGlu7 receptors are impaired in reference memory acquisition in a water maze task, whereas mGlu4 and mGlu8 receptor knockout mice did not exhibit deficits of this kind (Goddyn et al., 2015). This suggests that mGlu7 receptors may be of particular importance for hippocampal information processing. In line with this it was shown that positive allosteric modulation of the mGlu7 receptor restores LTP in a mouse model of Rett syndrome and improves both contextual fear learning and novel object recognition (Gogliotti et al., 2017).
Conclusions
MGlu receptors not only are intrinsically important for hippocampal information encoding, but the different subtypes of these receptors also assume different functional roles. This is reflected by the role of group I receptors in bidirectional forms for hippocampal synaptic plasticity, the bias towards group II and II receptor involvement in LTD and the distinctions in terms of memory forms that are regulated by mGlu receptor groups. It is clear that a tight interplay between mGlu receptors is a key determinant of hippocampal information processing: on the one hand, it has been shown that group I receptors regulate the expression of group I and II receptors (Marszalek-Grabska et al., 2008; Bikbaev et al., 2008). On the other hand, specific role of these receptors in enabling synaptic plasticity within synaptic subcompartments has been demonstrated. For example, the mGlu7 receptor is important for bidirectional plasticity at the mossy fibre synapses, whereby the direction of change of synaptic weight is determined by the relative activation and expression state of the receptor (Pelkey et al., 2005). By contrast, the mGlu5 receptor determines the direction of change of synaptic strength at mossy fibre versus commissural association (AC)-CA3 synapses: pharmacological antagonism of mGlu5 receptors impairs mossy fibre LTP, but not LTD (Fig. 1), whereas mGlu5 receptor antagonism impairs AC-CA3 LTD, but not LTP (Hagena and Manahan-Vaughan, 2015). Taken together, the importance of these receptors for hippocampal information encoding and the long-term storage of spatial experience should not be underestimated.
Acknowledgements: We thank Heide Brusis and Olga Neumann for assistance. Research by the author on this topic was supported by the German Research Foundation (DFG, MA 1843, SFB 874/B1,B3,B10).
About the author

Denise Manahan-Vaughan is a Neurophysiologist, Neuroscientist and Head of the Department of Neurophysiology within the Medical Faculty of the Ruhr University Bochum (www.rub.de/neurophys). She is also director and Dean of Studies of the International Graduate School of Neuroscience (www.rub.de/igsn) and speaker (primary coordinator) of the DFG-funded Collaborative Research Centre on Integration and Representation of Sensory Information Processes at the same university (www.rub.de/sfb874). After completion of her primary degree in Natural Sciences at Trinity College Dublin (TCD), Ireland, she obtained a PhD in Neuropharmacology at TCD and then moved to Germany to the Leibniz Institute for Neurobiology in Magdeburg. Having obtained an Habilitation degree in Physiology from the Otto von Guericke University in Magdeburg, she moved to the Johannes Müller Institute for Physiology at the Charité, Berlin, before moving to Bochum in 2003. Her research focusses on understanding the relationship between hippocampal and cortical synaptic plasticity and long-term associative and spatial memory.
References
André, M. E., Güntürkün, O., Manahan-Vaughan, D (2015) The metabotropic glutamate receptor, mGlu5 is required for extinction learning that occurs in the absence of a context change. Hippocampus, 25, 149–158.10.1002/hipo.22359Search in Google Scholar
Altinbilek, B., Manahan-Vaughan, D (2007) Antagonism of group III metabotropic glutamate receptors results in impairment of LTD but not LTP in the hippocampal CA1 region, and prevents long-term spatial memory. Eur J Neurosci. 26, 1166–1172.10.1111/j.1460-9568.2007.05742.xSearch in Google Scholar
Altinbilek, B., Manahan-Vaughan, D (2009) A specific role for group II metabotropic glutamate receptors in hippocampal long-term depression and spatial memory.Neuroscience 158,149–158.10.1016/j.neuroscience.2008.07.045Search in Google Scholar
Balschun, D., Wetzel, W (2002) Inhibition of mGluR5 blocks hippocampal LTP in vivo and spatial learning in rats. Pharmacol. Biochem. Behav 73, 375–380.10.1016/S0091-3057(02)00847-XSearch in Google Scholar
Bartos, M., Vida, I., and Jonas, P (2007) Synaptic mechanisms of synchronized gamma oscillations in inhibitory interneuron networks. Nat. Rev. Neurosci. 8, 45–56.10.1038/nrn2044Search in Google Scholar PubMed
Bikbaev, A., Manahan-Vaughan, D. (2007). Hippocampal network activity is transiently altered by induction of long-term potentiation in the dentate gyrus of freely behaving rats. 1, 1–7.10.3389/neuro.08.007.2007Search in Google Scholar PubMed PubMed Central
Bikbaev, A., Manahan-Vaughan, D. (2008). Relationship of hippocampal theta and gamma oscillations to potentiation of synaptic transmission. Front Neurosci. 2, 56–63.10.3389/neuro.01.010.2008Search in Google Scholar PubMed PubMed Central
Bikbaev, A., Neyman, S., Ngomba, R., Nicoletti, F., Conn, P.J, Manahan-Vaughan, D (2008) The metabotropic glutamate receptor, mGluR5, mediates the functional interaction between late-LTP, hippocampal network activity, and learning by a mechanism involving regulation of mGluR1 expression. PloS One. 13, e2155.10.1371/annotation/58313075-ff2e-4268-9272-e942aed8d2f6Search in Google Scholar
Bikbaev, A., Manahan-Vaughan, D (2017) Metabotropic glutamate receptor, mGlu5, regulates hippocampal synaptic plasticity and is required for tetanisation-triggered changes in theta and gamma oscillations. Neuropharmacology. 115,20–29.10.1016/j.neuropharm.2016.06.004Search in Google Scholar PubMed
Bliss, T.V., Collingridge, G.L (1993) A synaptic model of memory: long-term potentiation in the hippocampus. Nature 361,31–39.10.1038/361031a0Search in Google Scholar PubMed
Buschler, A., Manahan-Vaughan, D (2017) Metabotropic glutamate receptor, mGlu5, mediates enhancements of hippocampal long-term potentiation after environmental enrichment. Neuropharmacology. 115,42–50.10.1016/j.neuropharm.2016.06.003Search in Google Scholar
Bushell, T.J., Sansig, G., Collett, V.J., van der Putten, H., Collingridge, G.L (2002) Altered short-term synaptic plasticity in mice lacking the metabotropic glutamate receptor mGlu7. ScientificWorldJournal. 2,730-7.10.1100/tsw.2002.146Search in Google Scholar
Buzsáki, G (2005) Theta rhythm of navigation:link between path integration and landmark navigation, episodic and semantic memory. Hippocampus 15, 827–840.10.1002/hipo.20113Search in Google Scholar
Buzsaki, G., Draguhn, A (2004) Neuronal oscillations in cortical networks. Science. 304,1926–1929.10.1126/science.1099745Search in Google Scholar
Corti, C., Aldegheri, L., Somogyi, P., Ferraguti, F (2002) Distribution and synaptic localisation of the metabotropic glutamate receptor 4 (mGluR4) in the rodent CNS. Neuroscience 110, 403e420.10.1016/S0306-4522(01)00591-7Search in Google Scholar
De Filippis B, Lyon L, Taylor A, Lane T, Burnet PW, Harrison PJ, Bannerman DM (2015) The role of group II metabotropic glutamate receptors in cognition and anxiety: comparative studies in GRM2(-/-), GRM3(-/-) and GRM2/3(-/-) knockout mice. Neuropharmacology. 89, 19–32.10.1016/j.neuropharm.2014.08.010Search in Google Scholar PubMed PubMed Central
Eichenbaum, H. (2017) On the Integration of Space, Time, and Memory. Neuron. 95,1007–1018.10.1016/j.neuron.2017.06.036Search in Google Scholar PubMed PubMed Central
Ekstrom, A. D., Kahana, M.J., Caplan, J.B., Fields, T.A., Isham, E. A., Newman, E.L., Fried, I. (2003) Cellular networks underlying human spatial navigation. Nature. 425,184-810.1038/nature01964Search in Google Scholar PubMed
Gil-Sanz, C., Delgado-García, J.M., Fairén, A., Gruart, A (2008) Involvement of the mGluR1 receptor in hippocampal synaptic plasticity and associative learning in behaving mice. Cereb Cortex18, 1653–63.10.1093/cercor/bhm193Search in Google Scholar PubMed
Gogliotti RG, Senter RK, Fisher NM, Adams J, Zamorano R, Walker AG, Blobaum AL, Engers DW, Hopkins CR, Daniels JS, Jones CK, Lindsley CW, Xiang Z, Conn PJ, Niswender CM. (2017) mGlu7 potentiation rescues cognitive, social, and respiratory phenotypes in a mouse model of Rett syndrome. Sci Transl Med. 9, eaai7459.Search in Google Scholar
Goh, J., Manahan-Vaughan, D. (2013a) Endogenous hippocampal LTD that is enabled by spatial object recognition requires activation of NMDA receptors and the metabotropic glutamate receptor, mGlu5. Hippocampus. 23,129–138.10.1002/hipo.22072Search in Google Scholar PubMed
Goh, J., Manahan-Vaughan, D (2013b) Endogenous hippocampal LTD that is enabled by spatial object recognition requires activation of NMDA receptors and the metabotropic glutamate receptor, mGlu5. Hippocampus. 23,129–138.10.1002/hipo.22072Search in Google Scholar
Grieves, R.M., Jeffery, K.J. (2017) The representation of space in the brain. Behav Processes. 135,113–131.10.1016/j.beproc.2016.12.012Search in Google Scholar
Hagena, H., Manahan-Vaughan, D (2015) mGlu5 acts as a switch for opposing forms of synaptic plasticity at mossy fiber-CA3 and commissural associational-CA3 synapses JNeurosci 35, 4999–5006.10.1523/JNEUROSCI.3417-14.2015Search in Google Scholar
Hagena, H., Manahan-Vaughan, D (2017) mGlu5: a metabotropic glutamate receptor at the hub of hippocampal information processing, persistent synaptic plasticity and long-term memory. In: mGlu Receptors. Eds.: Ngomba, R.T., DiGiovanni, G., Battaglia, G., Nicoletti, F. Humana Press, pp 79–102. doi:10.1007/978-3-319-56170-7_5.10.1007/978-3-319-56170-7_5Search in Google Scholar
Hasselmo, M.E (2005) What is the function of hippocampal theta rhythm?—linking behavioral data to phasic properties of field potential and unit recording data. Hippocampus. 15, 936–949.10.1002/hipo.20116Search in Google Scholar
Hölscher, C., Anwyl, R., and Rowan, M. J. (1997). Block of theta-burst-induced long-term potentiation by (1S,3S)-1-aminocyclopentane-1,3-dicarboxylic acid: further evidence against long-term potentiation as a model for learning. Neuroscience 81, 17–22.10.1016/S0306-4522(97)00183-8Search in Google Scholar
Horner, A.J., Doeller, C.F. (2017) Plasticity of hippocampal memories in humans. Curr Opin Neurobiol. 43,102–109.10.1016/j.conb.2017.02.004Search in Google Scholar PubMed PubMed Central
Huber, K.M., Roder, J.C., Bear, M.F (2001) Chemical induction of mGluR5-and protein synthesis--dependent long-term depression in hippocampal area CA1. J Neurophysiol 86, 321–325.10.1152/jn.2001.86.1.321Search in Google Scholar PubMed
Kemp, A., Manahan-Vaughan, D (2007) Hippocampal long-term depression: master or minion in declarative memory processes? Trends Neurosci. 30,111–118.10.1016/j.tins.2007.01.002Search in Google Scholar PubMed
Kemp, A., Manahan-Vaughan, D (2008) The hippocampal CA1 region and dentate gyrus differentiate between environmental and spatial feature encoding through long-term depression. Cerebral Cortex. 18,968–977.10.1093/cercor/bhm136Search in Google Scholar PubMed
Kentros, C., Hargreaves, E., Hawkins, R.D., Kandel, E.R., Shapiro, M., Muller, R.V (1998) Abolition of long-term stability of new hippocampa lplace cell maps by NMDA receptor blockade. Science 280, 2121–2126.10.1126/science.280.5372.2121Search in Google Scholar PubMed
Klausnitzer, J., Kulla, A., Manahan-Vaughan, D. (2004) Role of the group III metabotropic glutamate receptor in LTP, depotentiation and LTD in dentate gyrus of freely moving rats. Neuropharmacology. 46, 160–70.10.1016/j.neuropharm.2003.09.019Search in Google Scholar PubMed
Knierim, J.J., Kudrimoti, H.S., McNaughton, B.L (1995) Place cells, head direction cells, and the learning of landmark stability. J Neurosci. 15,1648–1659.10.1523/JNEUROSCI.15-03-01648.1995Search in Google Scholar
Kulla, A., Reymann K.G., Manahan-Vaughan, D (1999) Time-dependent induction of depotentiation in the dentate gyrus of freely moving rats: involvement of group 2 metabotropic glutamate receptors. Eur. J. Neurosci. 11, 3864–72.10.1046/j.1460-9568.1999.00807.xSearch in Google Scholar PubMed
Lan, J.Y., Skeberdis, V. A., Jover, T., Zheng, X., Bennett, M.V., Zukin, R.S (2001) Activation of metabotropic glutamate receptor 1 accelerates NMDA receptor trafficking. J Neurosci. 21, 6058–68.10.1523/JNEUROSCI.21-16-06058.2001Search in Google Scholar
Lyon, L., Burnet, P.W., Kew, J.N., Corti, C., Rawlins, J.N., Lane, T., De Filippis, B., Harrison, P.J., Bannerman, D.M (2011) Fractionation of spatial memory in GRM2/3 (mGlu2/mGlu3) double knockout mice reveals a role for group II metabotropic glutamate receptors at the interface between arousal and cognition. Neuropsychopharmacology. 36, 2616–28.10.1038/npp.2011.145Search in Google Scholar PubMed PubMed Central
Manahan-Vaughan, D (1997) Group 1 and 2 metabotropic glutamate receptors play differential roles in hippocampal long-term depression and long-term potentiation in freely moving rats. J Neurosci 17,3303–3311.10.1523/JNEUROSCI.17-09-03303.1997Search in Google Scholar
Manahan-Vaughan, D (2017) Learning-Related Hippocampal Long-Term Potentiation and Long-Term Depression. In: Learning and Memory: A Comprehensive Reference (2nd Edition). Reference Module in Neuroscience and Biobehavioral Psychology, Elsevier, pp 585–609. http://dx.doi.org/10.1016/B978-0-12-809324-5.21104-810.1016/B978-0-12-809324-5.21104-8Search in Google Scholar
Manahan-Vaughan, D., Braunewell, K.H. (2005) The metabotropic glutamate receptor, mGluR5, is a key determinant of good and bad spatial learning performance and hippocampal synaptic plasticity.Cereb Cortex. 15,1703-13.10.1093/cercor/bhi047Search in Google Scholar PubMed
Mannaioni, G., Marino, M.J., Valenti, O., Traynelis, S.F., Conn, P.J (2001) Metabotropic glutamate receptors 1 and 5 differentially regulate CA1 pyramidal cell function. J. Neurosci. 21, 5925e5934.10.1523/JNEUROSCI.21-16-05925.2001Search in Google Scholar
Marszalek-Grabska M, Gibula-Bruzda E, Bodzon-Kulakowska A, Suder P, Gawel K, Filarowska J, Listos J, Danysz W, Kotlinska JH (2018) Effects of the Positive Allosteric Modulator of Metabotropic Glutamate Receptor 5, VU-29, on Impairment of Novel Object Recognition Induced by Acute Ethanol and Ethanol Withdrawal in Rats.Neurotox Res. doi: 10.1007/s12640-017-9857-z.10.1007/s12640-017-9857-zSearch in Google Scholar
Martin, E.D., Buno, W (2005) Stabilizing effects of extracellular ATP on synaptic efficacy and plasticity in hippocampal pyramidal neurons. Eur J Neurosci. 21, 936–944.10.1111/j.1460-9568.2005.03925.xSearch in Google Scholar
Mukherjee. S,. Manahan-Vaughan, D (2013) Role of metabotropic glutamate receptors in persistent forms of hippocampal plasticity and learning. Neuropharmacology 66, 65–81.10.1016/j.neuropharm.2012.06.005Search in Google Scholar
Naie, K., Manahan-Vaughan, D (2004) Regulation by metabotropic glutamate receptor 5 of LTP in the dentate gyrus of freely moving rats: relevance for learning and memory formation. Cereb. Cortex 14, 189–198.10.1093/cercor/bhg118Search in Google Scholar
Naie, K., Manahan-Vaughan, D (2005) Pharmacological antagonism of metabotropic glutamate receptor 1 (mGluR1) regulates LTP and spatial reference memory in the dentate gyrus of freely moving rats via NMDA and mGluR-dependent mechanisms. Eur. J. Neurosci. 21, 411–421.10.1111/j.1460-9568.2005.03864.xSearch in Google Scholar
Naie, K., Manahan-Vaughan, D (2006) Investigations of the protein synthesis dependency of mGluR-induced long-term depression in the dentate gyrus of freely moving rats. Neuropharmacology 49 (Suppl. 1), 35–44.10.1016/j.neuropharm.2005.06.001Search in Google Scholar
Nakao K, Ikegaya Y, Yamada MK, Nishiyama N, Matsuki N (2002) Hippocampal long-term depression as an index of spatial working memory. Eur J Neurosci. 16, 970-4.10.1046/j.1460-9568.2002.02159.xSearch in Google Scholar
Neyman, S., Manahan-Vaughan, D (2008) Metabotropic glutamate receptor 1 (mGluR1) and 5 (mGluR5) regulate late phases of LTP and LTD in the hippocampal CA1 region in vitro Eur J Neurosci. 27,1345-52.10.1111/j.1460-9568.2008.06109.xSearch in Google Scholar
Nomura, A., Shigemoto, R., Nakamura, Y., Okamoto, N., Mizuno, N., Nakanishi, S (1994) Developmentally regulated postsynaptic localization of a metabotropic glutamate receptor in rat rod bipolar cells. Cell 77, 361–369.10.1016/0092-8674(94)90151-1Search in Google Scholar
Ohishi, H., Shigemoto, R., Nakanishi, S., Mizuno, N (1993) Distribution of the messenger RNA for a metabotropic glutamate receptor, mGluR2, in the central nervous system of the rat. Neuroscience 53, 1009–1018.10.1016/0306-4522(93)90485-XSearch in Google Scholar
Okamoto, N., Hori, S., Akazawa, C., Hayashi, Y., Shigemoto, R., et al., (1994) Molecular characterization of a new metabotropic glutamate receptor mGluR7 coupled to inhibitory cyclic AMP signal transduction. J. Biol. Chem. 269, 1231–123610.1016/S0021-9258(17)42247-2Search in Google Scholar
O’Keefe, J., Dostrovsky, J (1971) The hippocampus as a spatial map. Preliminary evidence from unit activity in the freely-moving rat. Brain Res. 34,171–175.10.1016/0006-8993(71)90358-1Search in Google Scholar
Petralia, R.S., Wang, Y.X., Niedzielski, A.S., Wenthold, R.J. (1996) The metabotropic glutamate receptors, mGluR2 and mGluR3, show unique postsynaptic, presynaptic and glial localizations. Neuroscience. 71, 949–976.10.1016/0306-4522(95)00533-1Search in Google Scholar
Popkirov, S.G., Manahan-Vaughan, D (2011) Involvement of metabotropic glutamate receptor mGluR5 in learning facilitated-plasticity at CA1 synapses. Cerebral Cortex. 21, 501–519.10.1093/cercor/bhq093Search in Google Scholar
Shigemoto, R., Kinoshita, A., Wada, E., Nomura, S., Ohishi, H., Takada, M., Flor, P.J., Neki, A., Abe, T., Nakanishi, S., Mizuno, N. (1997) Differential presynaptic localization of metabotropic glutamate receptor subtypes in the rat hippocampus. J Neurosci. 17, 7503-22.10.1523/JNEUROSCI.17-19-07503.1997Search in Google Scholar
Skeberdis, V. A., Lan, J., Opitz, T., Zheng, X., Bennett, M.V., Zukin, R.S (2001) mGluR1-mediated potentiation of NMDA receptors involves a rise in intracellular calcium and activation of protein kinase C. Neuropharmacology 40, 856–865.10.1016/S0028-3908(01)00005-3Search in Google Scholar
Ster, J., Mateos, J.M., Grewe, B.F., Coiret, G., Corti, C., Corsi, M., Helmchen, F., Gerber, U. (2011) Enhancement of CA3 hippocampal network activity by activation of group II metabotropic glutamate receptors.Proc Natl Acad Sci USA. 108, 9993-7.10.1073/pnas.1100548108Search in Google Scholar PubMed PubMed Central
Szapiro, G., Barros, D.M., Ardenghi, P., Vianna, M.R., Choi, H., Silva, T., Medina, J.H., Izquierdo, I. (2001) Facilitation and inhibition of retrieval in two aversive tasks in rats by intrahippocampal infusion of agonists of specific glutamate metabotropic receptor subtypes. Psychopharmacology (Berl). 156:397-40110.1007/s002130100755Search in Google Scholar PubMed
Tanabe, Y., Nomura, A., Masu, M., Shigemoto, R., Mizuno, N., Nakanishi, S., 1993. Signal transduction, pharmacological properties, and expression patterns of two rat metabotropic glutamate receptors, mGluR3 and mGluR4. J. Neurosci. 13,1372-378.10.1523/JNEUROSCI.13-04-01372.1993Search in Google Scholar
Zarnadze, S., Bäuerle, P., Santos-Torres, J., Böhm, C., Schmitz, D., Geiger, J.R., Dugladze, T., Gloveli, T. (2016) Cell-specific synaptic plasticity induced by network oscillations. Elife. 24;5.10.7554/eLife.14912Search in Google Scholar PubMed PubMed Central
Zhang, S., Manahan-Vaughan, D (2014) Place field stability requires the metabotropic glutamate receptor, mGlu5. Hippocampus, 24,1330-40.10.1002/hipo.22314Search in Google Scholar PubMed PubMed Central
Zhang, S., Manahan-Vaughan, D (2015) Spatial olfactory learning facilitates place field formation in the hippocampus. Cerebral Cortex 25, 423–432.10.1093/cercor/bht239Search in Google Scholar PubMed PubMed Central
Zhou, R., Chen, F., Feng, X., Zhou, L., Li, Y., Chen, L. (2015) Perinatal exposure to low-dose of bisphenol A causes anxiety-like alteration in adrenal axis regulation and behaviors of rat offspring: a potential role for metabotropic glutamate 2/3 receptors. J Psychiatr Res. 64,121-9.10.1016/j.jpsychires.2015.02.018Search in Google Scholar PubMed
Article note
German version available at https://doi.org/10.1515/nf-2018-0007
© 2018 Walter de Gruyter GmbH, Berlin/Boston
Articles in the same Issue
- Titelseiten
- Übersichtsartikel
- Hippokampale Langzeitpotenzierung (LTP) – Geschichte, Gegenwart und Zukunft
- Hippocampal long-term potentiation (LTP) – past, present and future
- Langzeitpotenzierung im Hippokampus: Entdeckung, Mechanismen und Funktion
- Long-term potentiation in the hippocampus: discovery, mechanisms and function
- Die Regulation der hippokampalen Informationsenkodierung durch metabotrope Glutamatrezeptoren
- Regulation of hippocampal information encoding by metabotopic glutamate receptors
- Geclusterte Plastizität bei Langzeitpotenzierung: Wie starke Synapsen bestehen bleiben, um Langzeitgedächtnis aufrechtzuerhalten
- Clustered plasticity in Long-Term Potentiation: How strong synapses persist to maintain long-term memory
- Hippocampale synaptische Plastizität bei neurodegenerativen Erkrankungen: Aβ, Tau und darüber hinaus
- Hippocampal synaptic plasticity in neurodegenerative diseases: Aß, tau and beyond
- Die Analyse synaptischer Plastizität auf Einzelzellebene mit Hilfe der STDP
- Analyzing synaptic plasticity at the single cell level with STDP
- Institutsvorstellung
- Die Neurowissenschaftliche Gesellschaft ist Gründungsmitglied des German Brain Council
- DFG-Graduiertenkolleg 1957 „Adipocyte-Brain Crosstalk“
- Nachrichten
- Jugend forscht – Sonderpreis der Neurowissenschaftlichen Gesellschaft 2018
Articles in the same Issue
- Titelseiten
- Übersichtsartikel
- Hippokampale Langzeitpotenzierung (LTP) – Geschichte, Gegenwart und Zukunft
- Hippocampal long-term potentiation (LTP) – past, present and future
- Langzeitpotenzierung im Hippokampus: Entdeckung, Mechanismen und Funktion
- Long-term potentiation in the hippocampus: discovery, mechanisms and function
- Die Regulation der hippokampalen Informationsenkodierung durch metabotrope Glutamatrezeptoren
- Regulation of hippocampal information encoding by metabotopic glutamate receptors
- Geclusterte Plastizität bei Langzeitpotenzierung: Wie starke Synapsen bestehen bleiben, um Langzeitgedächtnis aufrechtzuerhalten
- Clustered plasticity in Long-Term Potentiation: How strong synapses persist to maintain long-term memory
- Hippocampale synaptische Plastizität bei neurodegenerativen Erkrankungen: Aβ, Tau und darüber hinaus
- Hippocampal synaptic plasticity in neurodegenerative diseases: Aß, tau and beyond
- Die Analyse synaptischer Plastizität auf Einzelzellebene mit Hilfe der STDP
- Analyzing synaptic plasticity at the single cell level with STDP
- Institutsvorstellung
- Die Neurowissenschaftliche Gesellschaft ist Gründungsmitglied des German Brain Council
- DFG-Graduiertenkolleg 1957 „Adipocyte-Brain Crosstalk“
- Nachrichten
- Jugend forscht – Sonderpreis der Neurowissenschaftlichen Gesellschaft 2018