Mechanismen des Neuritischen Prunings
-
Sebastian Rumpf
Dr. Sebastian Rumpf (*1975) Biologiestudium in Heidelberg,
Dr. rer. nat. am Max-Planck-Institut für Biochemie (2006). Postdoc an der University of California San Francisco im Labor von Prof. Yuh Nung Jan, Juniorgruppenleiter an der WWU Münster seit 2013. Dr. Rumpf untersucht Pruning-Mechanismen inDrosophila . ,
Sandra Rode
,
Sandra Rode
Sandra Rode (*1987) Biologiestudium an der WWU Münster, Masterarbeit im Labor von C. Klämbt über die Funktion von Gliazellen. Seit 2013 Promotionsstudentin an der WWU zum Thema
Pruning von C4da-Neuronen .Svende Herzmann (*1988) Biologiestudium an der WWU Münster, Masterarbeit im Labor von C. Klämbt über die Entwicklung von Gliazellen. Seit 2013 Promotionsstudentin an der WWU zum Thema
Pruning von C4da-Neuronen .Rafael Krumkamp (*1987) Biologiestudium an der WWU Münster, Masterarbeit im Labor von S. Rumpf über
Pruning von C4da-Neuronen . Seit 2015 Promotionsstudent an der WWU zum selben Thema.

Zusammenfassung
Die Entwicklung des Nervensystems beinhaltet sowohl progressive Phasen, in denen Neuriten und Synapsen geformt werden, als auch regressive Phasen, in denen überzählige Fortsätze zurückgezogen und abgebaut werden. Solche regressiven Mechanismen werden auch alsPruning-Prozesse bezeichnet. Pruning wird zur Spezifizierung neuronaler Verbindungen benötigt, aber auch, um temporär benötigte Strukturen wieder zu entfernen. Dabei können sowohl einzelne Synapsen als auch lange Neuriten entfernt werden. Hier stellen wir vier Beispiele für entwicklungsabhängiges Pruning aus Säugern und Drosophila mit ihrem biologischen Zweck und zellbiologischem Mechanismus vor. Dabei wollen wir mechanistische Gemeinsamkeiten der verschiedenen Modelle hervorheben.
Einleitung
Nervenzellen besitzen charakteristische lange und verzweigte Fortsätze (Neuriten), die Dendriten und Axone, die über Synapsen miteinander verbunden sind. Sie dienen der Aufnahme bzw. der Weiterleitung von Information in Form von elektrischen Reizen. Während der Entwicklung wachsen Neuriten mit hoher Zielgenauigkeit und über lange Strecken, bis sie ihre jeweilige Zielregion erreicht haben. Aber nicht nur Neuritenwachstum und Synapsenbildung sind für die Vernetzung im Nervensystem wichtig, sondern auch die regulierte Entfernung von Neuriten und Synapsen. Solche regressiven Mechanismen werden oft unter dem Begriff Pruning zusammengefasst (englisch to prune: auslauben, stutzen, beschneiden) (Schuldiner und Yaron, 2015). So wächst die Synapsendichte im menschlichen Gehirn während des ersten Lebensjahrs rapide, fällt dann jedoch bis nach der Pubertät wieder stetig (Huttenlocher, 1990). Durch Pruning können nicht nur kleine Synapsen, sondern auch lange Abschnitte von Axonen oder Dendriten entfernt werden. Hier stellen wir vier Beispiele für entwicklungsabhängiges Pruning und die dazugehörigen zellbiologischen Mechanismen vor: Synapseneliminierung an der Neuromuskulären Endplatte in Säugern, Pruning bei der Ausbildung von augenspezifischen Verbindungen im visuellen System, Spezifizierung von efferenten Axonen von Neuronen in der kortikalen Schicht V, und das Pruning von sensorischen class IV dendritic arborization (c4da)-Neuronen in der Fruchtfliege Drosophila. Anhand dieser Beispiele diskutieren wir dann allgemeine zellbiologische Mechanismen des Pruning.
Pruning: Spezifizierung neuronaler Schaltkreise und mehr
Warum werden während der Entwicklung Neuriten und Synapsen gebildet, nur um kurz darauf wieder abgebaut zu werden? Zum einen müssen die Milliarden Nervenzellen im Säugergehirn eine Vielzahl hochspezifischer Synapsen bilden (manche Neuronen haben über tausend Synapsen). Diese hohe Anzahl erfordert Mechanismen, die die Zielgenauigkeit von Axonen und Dendriten und die Spezifität der Synapsenbildung regulieren. Solche Mechanismen beruhen meist auf spezifischen Paaren von Transmembranrezeptoren, die jeweils passende Prä- und Postsynapsen markieren. Die Diversität solcher Rezeptorpaare im Säugergenom ist tatsächlich relativ hoch und wird durch alternatives Splicing noch erhöht (einige axon guidance-Rezeptoren haben potenziell Hunderte Isoformen). Wahrscheinlich sind die kombinatorischen Möglichkeiten von einigen tausend Rezeptoren aber nicht genug, um Billionen von Synapsen im Genom eindeutig zu spezifizieren. Das hätte zwangsweise Verknüpfungsfehler bei der Gehirnentwicklung zur Folge. Erst nachfolgende Korrekturmechanismen wie Pruning könnten dann die hohe Spezifität der Gehirnverschaltungen ermöglichen. Möglicherweise erfolgt die initiale Verschaltung des Gehirns auch nach dem trial-and-error-Prinzip. Danach würden die geeignetsten Synapsen durch funktionelle Tests – wie häufiger Gebrauch – identifiziert und ungeeignete durch Pruning wieder entfernt. In jedem Fall dient Pruning der Spezifität von synaptischen Verbindungen.
Pruning-Mechanismen werden auch in Fällen benötigt, in denen das Nervensystem an unterschiedliche Entwicklungsphasen adaptiert ist. Dies ist bei holometabolen Insekten der Fall, deren Lebenszyklus in die morphologisch sehr verschiedenen Larven- und adulte Phasen unterteilt sind. Die Nervensysteme von Larve und Fliege sind an die jeweilige Lebensweise angepasst: Kriechen und Fressen bzw. Fliegen und Paarung. Große Teile des larvalen Nervensystems sind daher in der Fliege nutzlos. Solche Entwicklungsintermediate werden in der Metamorphose u. a. durch Pruning entfernt.
Allgemein wird zwischen klein- und großskaligem Pruning unterschieden. Beim kleinskaligen Pruning werden meist nur einzelne Synapsen oder kleine Teile von Axonen oder Dendriten eliminiert, während beim großskaligen Pruning lange Abschnitte von primären Axonen oder Dendriten abgebaut werden. Beim kleinskaligen Pruning können Synapsen einfach durch Zurückziehen entfernt werden. Beim großskaligen Pruning dagegen fragmentieren und degenerieren abzubauende Neuriten. Diese Art von Pruning erfolgt meist nach stereotypischen Mustern: Die Identität des abzubauenden Neuriten sowie der Zeitpunkt sind genau vorhersagbar. Kleinskaliges Pruning ist oft das Ergebnis aktivitätsabhängiger Synapsenkompetition, sodass die Identität der abzubauenden Synapse nicht von vornherein feststeht. Großskaliges Pruning spielt oft eine Rolle beim Entfernen von Entwicklungsintermediaten, etwa bei der Metamorphose von Insekten.
Beispiel 1: Synapseneliminierung an der Neuromuskulären Endplatte
Bei der Geburt sind die meisten neuromuskulären Endplatten in Säugern von mehreren Motoneuron-Axonen innerviert. Während der ersten beiden Wochen nach der Geburt werden dann systematisch Axone eliminiert, bis jede Endplatte nur noch von einem Axon innerviert wird (Abb. 1A). Schwächere Axone werden zunächst dünner, um sich schließlich von der Endplatte bis zum nächsten Verästelungspunkt zurückzuziehen, während die stärkeren Axone die gesamte Endplatte ausfüllen. Beim Rückzug bilden Axone am Ende eine charakteristische runde Struktur, die retraction bulb genannt wird, und hinterlassen Axosomen, kleine Membranpartikel (Bishop et al., 2004).
Motoneuron-Axone werden durch Kompetition eliminiert. An jeder Endplatte wird dasjenige Axon stabilisiert, das mehr Neurotransmitter ausschüttet und den Muskel besser stimuliert, während das schwächere Axon destabilisiert wird. Wenn man experimentell einzelne Axone aktiviert oder inhibiert, erhöht bzw. verringert dies seine Chancen, die Endplatte zu übernehmen. Stabilisierung und Destabilisierung beinhalten retrogrades Feedback vom Muskel an die individuellen Präsynapsen. Dieses Feedback wird von verschiedenen Isoformen des Brain-derived Neurotrophic Factor (BDNF) durch Stimulation verschiedener präsynaptischer Rezeptoren vermittelt (Je 2012). Schwächere Axone werden systematisch destabilisiert: Sie enthalten weniger stabilisierende Zytoskelett-Elemente, und retraction bulbs enthalten Lysosomen (Membranabbau-Organellen). Synapseneliminierung an der Endplatte ist ein Beispiel für kleinskaliges Pruning, das der Spezifizierung dient.

Beispiele für Pruning während der neuronalen Entwicklung in Säugern. (A) Synapseneliminierung an der Neuromuskulären Endplatte. Anfänglich sind Endplatten von mehreren Motoneuronen (MN) innerviert (links), dann ziehen sich die schwächeren Axone zurück (rechts). (B) Augenspezifische Segregation der Axone von Retinalen Ganglienzellen (RGC) im Corpus geniculatum laterale (dLGN). Kurz nach der Geburt überlappen die Zielareale der ipsi- und kontralateralen RGC-Axone (links), die nicht-spezifischen Inputs werden dann eliminiert, sodass die Zielareale segregiert sind (rechts). C Pruning von Axonen von kortikalen L5-Neuronen. L5-Neurone aus visuellem und Motorkortex projizieren anfänglich sowohl ins Rückenmark als auch in visuelle Zentren wie den Superior Colliculus (SC) (links), dann degenerieren nichtspezifische Axone (rechts).
Beispiel 2: Augenspezifische Segregation der Axone von Retinalen Ganglienzellen
Retinale Ganglienzellen (engl., retinal ganglion cells, RGCs) leiten visuelle Information aus der Netzhaut ins zentrale Nervensystem (ZNS). Einer der ersten Verschaltungspunkte ist der Corpus geniculatum laterale (engl., dorsal Lateral Geniculate Nucleus, dLGN) im Thalamus. RGCs aus dem ipsi- und dem kontralateralen Auge senden Axone in beide Hemisphären des dLGN (Abb. 1B). Durch Injektion von verschiedenen Farbstoffen in beide Augen konnte gezeigt werden, dass die ipsi- und kontralateralen RGCs in streng voneinander getrennte fleckförmige Bereiche des dLGN projizieren. Diese räumliche Trennung ermöglicht den höheren visuellen Zentren die Kalkulation von Stereobildern. Die Segregation der ipsi- und kontralateralen Axone im dLGN während der Entwicklung entsteht durch Pruning von fehlgeleiteten Axonen. Wenn neuronale Aktivität in einem Auge experimentell vermindert wird, werden RGC-Axone aus diesem Auge präferenziell durch Pruning entfernt, was zu Segregationsdefekten führt (Schafer et al., 2012) und auf einen aktivitätsabhängigen (Kompetitions-?)Mechanismus hindeutet. Abzubauende RGC-Synapsen werden durch Komplementproteine markiert. Diese sind eigentlich Komponenten des angeborenen Immunsystems, die im Blut an Bakterien binden und diese für die Phagozytose markieren. Während der Entwicklung können sie aber auch im ZNS nachgewiesen werden. Komplement-markierte Bakterien werden durch Phagozyten eingeschlossen und verdaut. In einer ähnlichen Weise werden Komplement-markierte RGC-Synapsen von hirnständigen Immunzellen, den Mikroglia, phagozytiert. Diese exprimieren ebenfalls während der Entwicklung entsprechende Komplement-Rezeptoren. Außer von Mikroglia können RGC-Synapsen auch von Astrozyten, einer anderen Art von Gliazellen, abgebaut werden. Diese benutzen dazu den MEGF10-Rezeptor, der normalerweise apoptotische (absterbende) Zellen erkennt (Chung, 2013). Mausmutanten, die einen dieser Rezeptoren nicht besitzen, zeigen Segregations- und neurophysiologische Defekte im dLGN. Die Segregation von Inputs im dLGN dient der Spezifizierung der Kontakte im visuellen System und beruht auf kleinskaligem Pruning.
Beispiel 3: Pruning von kortikospinalen Axonen von L5-Neuronen
Der Neokortex von Säugern ist in fünf Schichten aufgebaut. Pyramidalzellen der Schicht V (engl. Layer V, L5) senden Axone in verschiedene ausführende ZNS-Regionen. L5-Neurone des visuellen Kortex projizieren Axone in die Sehhügel (engl., superior colliculus, SC) im Mittelhirn, während L5-Neurone des Motorkortex Axone ins Rückenmark senden. Die Axone von L5-Neuronen fangen kurz nach der Geburt an zu wachsen. Alle L5-Axone verlassen den Kortex zunächst über einen gemeinsamen Pfad und wachsen gemeinsam ins Rückenmark. Dann bilden alle L5-Axone Verästelungen, die in die anderen Zielregionen wie den SC wachsen (Abb. 1C). Eine Spezifizierung wird erst dann durch Pruning der falschen Projektionen erreicht: Im Rückenmark werden die L5-Axone aus dem visuellen Kortex entfernt, und im SC die L5-Axone aus dem Motorkortex. Diese Spezifizierung erfolgt in der zweiten postnatalen Woche, wenn die verschiedenen Kortexareale beginnen, sich zu spezialisieren. Dabei fragmentieren Axonäste in den falschen Zielregionen bis zur Verzweigung und lösen sich auf. Pruning von L5-Axonen erfolgt also durch einen degenerativen Mechanismus. Dabei werden die falschen Axonäste wahrscheinlich von den jeweiligen Zielarealen abgestoßen. Visuelle L5-Neurone, aber nicht jene aus dem Motorkortex, exprimieren die repulsiven axon guidance-Rezeptoren Neuropilin-2 und PlexinA3/A4, deren repulsiver Ligand Semaphorin 3F (Sema 3F) im Rückenmark exprimiert wird. Durch Aktivierung des Rezeptors werden die visuellen L5-Axonäste im Rückenmark abgestoßen. In PlexinA3/A4-Mutanten persistieren diese Äste im Rückenmark (Low et al., 2008). Pruning von L5-Axonen ist großskalig und stereotypisch, da die Identität der zu entfernenden Axonäste und der Zeitverlauf genau vorhergesagt werden können.
Beispiel 4: Dendritisches Pruning von Drosophila c4da-Neuronen
C4da-Neurone sind nozizeptive sensorische Nervenzellen in der Haut von Larven von Drosophila-Fruchtfliegen. Diese Neurone haben lange und verzweigte sensorische Dendriten, die die gesamte Haut der Larve abdecken. Die Dendriten sind eng mit den Epidermiszellen verbunden. Während der Puppenphase werden die larvalen Epidermiszellen durch neue adult-spezifische Zellen ersetzt. Dadurch verlieren die c4da-Dendriten ihr Substrat, sie müssen durch Pruning abgebaut werden (Abb. 2A). Sowohl der Zellkörper als auch das Axon bleiben intakt. Da c4da-Neurone leicht genetisch manipuliert und sehr gut mikroskopisch untersucht werden können, ist dies eines der am besten verstandenen Modelle für Pruning. Dendriten-Pruning von c4da-Neuronen wird durch dasVerpuppungshormon Ecdyson ausgelöst. Ecdyson aktiviert den Ecdysonrezeptor EcR-B1, einen Transkriptionsfaktor, der die Transkription einer kleinen Zahl von Pruning-Genen aktiviert (Abb. 2B). Dazu gehört Sox14, ein Transkriptionsfaktor, der das MICAL-Gen aktiviert (Kirilly, 2009). MICAL kodiert für ein Enzym, das Aktinfilamente – stabilisierende Zytoskelettkomponenten – auflöst. In den Dendriten können die ersten Anzeichen von Pruning etwa fünf Stunden nach Beginn der Puppenphase (engl. hours after puparium formation, h APF) detektiert werden. Dabei wird im proximalen Dendritenabschnitt das Zytoskelett (insbesondere die Mikrotubuli) abgebaut. Für den Abbau der Mikrotubuli wird das Enzym Katanin p60-like 1 benötigt, das Mikrotubuli zerbrechen kann (Lee, 2009). Außerdem werden in proximalen Dendriten vermehrt endozytische Vesikel gebildet, die Membranmaterial von der Plasmamembran abziehen (Kanamori et al., 2015). Durch die Kombination von Zytoskelettabbau und Membranabzug wird der Dendrit an der Basis ausgedünnt und destabilisiert (Abb. 2B). Die Ausdünnung der proximalen Dendriten isoliert die Dendriten vom Soma. In ihnen kann man nun häufig räumlich und zeitlich begrenzte Kalziumströme beobachten, sogenannte transients. Das Auftreten dieser transients ist ein Indikator, dass der betreffende Dendrit demnächst im proximalen Bereich abbrechen wird (Kanamori et al., 2015), was ab etwa 6 h APF passiert. Zur selben Zeit kann man in den abgeschnürten Dendriten aktive Caspasen detektieren – Proteasen, die normalerweise den regulierten Zelltod auslösen (Williams, 2006). Die Caspasen bewirken, dass abgebrochene Dendriten in kleine Fragmente zerfallen, die dann von den umgebenden Epidermiszellen phagozytisch aufgenommen werden (Han, 2014) (Abb. 2B). Dabei benutzen sie den Rezeptor Draper, der dem glialen Synapsenrezeptor MEGF10 ähnlich ist. Nach etwa 16 h APF sind die Dendriten von c4da-Neuronen komplett abgebaut. Nun werden die umgebenden Epidermis- und Muskelzellen erneuert. Nach etwa zwei Tagen wachsen den c4da-Neuronen neue Dendriten, die an die neue Umgebung angepasst sind. Larvale Dendriten von c4da-Neuronen sind Entwicklungsintermediate, und das dendritische Pruning ist ein Beispiel für stereotypisches großskaliges Pruning.
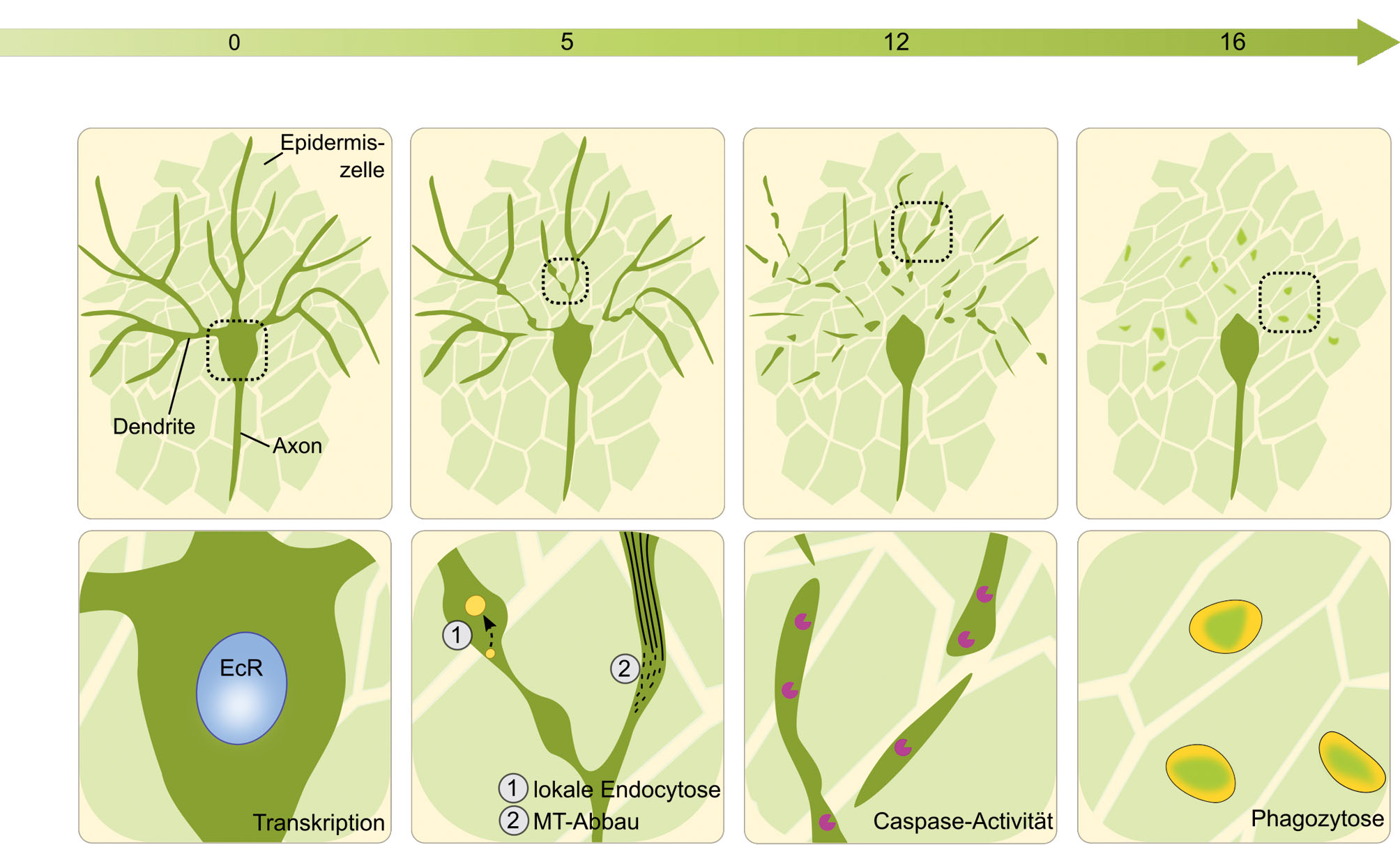
Zeitverlauf und zellbiologische Mechanismen des dendritischen Pruning von Drosophila c4da-Neuronen. (A) Zeitverlauf. Larvale c4da-Neurone haben zu Beginn der Puppenphase (0 hours after puparium formation, h APF) lange und verzweigte sensorische Dendriten. Nach etwa 5 h APF dünnen die Dendriten proximal aus und brechen. Bis 12 h APF fragmentieren die abgebrochenen Dendriten und werden anschließend phagozytisch entfernt. Nach 16 Stunden ist der Prozess beendet, die Axone bleiben bestehen. (B) Zellbiologische Mechanismen. In der Reihenfolge des zeitlichen Auftretens: Ecdyson-abhängige Transkription von Pruning-Genen im Zellkern (blau) ab 0 h APF; Dendritendestabilisierung um 5 h APF: Mikrotubuli (schwarze Linien) werden lokal abgebaut, und Plasmamembran wird lokal durch Endozytose entfernt (gelbe Vesikel); Fragmentierung von Dendriten durch Caspasen (lila) zwischen 5–12 h APF; anschließend Phagozytose durch Epidermiszellen (gelbe Vesikel).
Wie wird Pruning initiiert?
Es scheint mindestens zwei prinzipielle Mechanismen zu geben, die Pruning auslösen. An der neuromuskulären Endplatte und bei der Spezifizierung der axonalen Kontakte von RGCs wird Pruning wahrscheinlich durch synaptische Kompetition ausgelöst. Diejenigen Synapsen, die weniger synaptische Aktivität (als ihre Nachbarsynapsen) haben, werden abgebaut. Bei Motoneuron-Synapsen führt starke Aktivität zu einem positiven Feedback von der postsynaptischen Muskelzelle (trophic support), das wahrscheinlich aus Wachstumsfaktoren besteht. Schwache Aktivität hat ein negatives Feedback zur Folge, entweder Entzug von trophic support, oder sogar ein punishment- (d. h., Straf-)Signal. Wie solche Signale lokal auf engem Raum vermittelt werden, ist unbekannt. Bei der dLGN-Entwicklung findet man die Komplement-Untereinheit C1q als Phagozytosesignal auf präsynaptischen Strukturen, die nicht in unmittelbarer Nachbarschaft einer Postsynapse sind. In beiden Beispielen wird durch einen funktionellen Test entschieden, ob die Synapse entfernt wird oder nicht.
In Drosophila c4da-Neuronen wird Pruning durch das Verpuppungshormon Ecdyson ausgelöst, dessen Konzentration zu Beginn der Puppenphase stark ansteigt. Ecdyson induziert die Expression von Pruning-Genen, die den Dendritenabbau intrinsisch, also ohne Feedback von anderen Zellen bewirken. Pruning von L5-corticospinalen Axonen beruht auf Abstoßung durch die repulsiven axon guidance-Rezeptoren PlexinA3/A4. Die Spezifität dieser Abstoßung wird dadurch erreicht, dass nur L5-Neuronen aus dem visuellen Kortex, aber nicht jene aus dem Motorkortex, diese Rezeptoren exprimieren. Daher sind die Axone von L5-Neuronen aus dem Motorkortex insensitiv gegen das Sema 3F-Signal und persistieren, während die der L5-Neurone aus dem visuellen Kortex im Rückenmark entfernt werden. Der entscheidende Faktor ist hier ebenfalls ein (gewebespezifischer) Unterschied in der Genexpression – ob Unterschiede in der neuronalen Aktivität hier auch eine Rolle spielen, ist noch unbekannt. Stereotypisches großskaliges Pruning wird also oft durch Änderungen der Genexpression kontrolliert.
Lokale intrinsische Effekte auf Zytoskelett und Plasmamembran
Sowohl beim Pruning durch Retraktion als auch beim Pruning durch Degeneration spielen lokale Änderungen in Zytoskelett und Plasmamembran eine wichtige Rolle. Im Rückzug befindliche Motoneuron-Axone an der neuromuskulären Endplatte zeigen im Vergleich zu stabilen Axonen weniger Mikrotubuli und desorganisierte Neurofilamente (Bishop et al., 2004).Wie diese Aktivitäten durch negatives Feedback vom Muskel ausgelöst werden, ist unklar.
In c4da-Neuronen werden Mikrotubuli lokal in proximalen Dendriten zerstört, wo der Dendrit schließlich abbrechen wird. Die Plasmamembran von c4da-Neuronen ist zu Beginn der Puppenphase ebenfalls starken Änderungen unterworfen. So wird durch lokale Endozytose in proximalen Dendriten Membran abgezogen (Kanamori et al., 2015). Durch welche Signalwege Ecdyson diese lokalen Effekte bewirkt, ist noch unbekannt.
Membran- und Zytoskelettregulation spielen höchstwahrscheinlich auch eine wichtige Rolle beim Pruning von L5-Axonen. Ob RGC-Synapsen vor der Phagozytose intrinsisch destabilisiert werden müssen, ist unklar.
Fragmentierung durch Caspasen und Phagozytose
Beim stereotypischen großskaligen Pruning zerfallen Neuriten meist noch vor ihrer Phagozytose in kleine Fragmente. Interessanterweise fragmentieren Neuriten auch, wenn Nervenzellen durch Apoptose absterben, was auf einen ähnlichen Mechanismus hinweist. Tatsächlich wird die Fragmentierung in c4da-Neuronen durch Caspasen vermittelt, und Caspasen sind mittlerweile in mehreren Pruning-Modellen impliziert worden (Schuldiner und Yaron, 2015). Ob die Fragmentierung von L5-Axonen oder die Entstehung von Axosomen beim Pruning von Motoneuron-Axonen ebenfalls auf Caspasen beruht, ist noch nicht untersucht worden.
Ein sehr wichtiger Mechanismus beim Pruning ist Phagozytose. In c4da-Neuronen werden die fragmentierten Dendriten durch Epidermiszellen phagozytisch entfernt (Han, 2014). Phagozytose ist auch für das Pruning von RGC-Synapsen wichtig, diese können also nicht allein durch Retraktion entfernt werden. Wenn die Phagozytose von RGC-Synapsen im dLGN inhibiert ist, bleiben die überzähligen Synapsen funktional, offensichtlich haben hier die Phagozyten auch eine Funktion bei der Inaktivierung der Synapsen. Bei der Motoneuron-Synapseneliminierung werden Axosomen von myelinierten Schwannzellen aufgenommen, eine potenzielle Rolle der Phagozytose beim Pruning von L5-Axonen ist wahrscheinlich, wurde aber noch nicht untersucht.
Ausblick
Viele Mechanismen des Pruning sind in den Grundrissen schon gut verstanden. Eine der wichtigsten offenen Fragen ist die der räumlichen Regulation – wie wird bestimmt, bis wohin ein Axon oder ein Dendrit abstirbt? Ebenfalls noch nicht sehr gut verstanden ist der Mechanismus der Synapsenkompetition. Die Erforschung der genauen molekularen Signalwege, durch die die Pruning-Auslöser wirken, wird sicher bald Antworten auf diese Fragen geben.
Über die Autoren

Dr. Sebastian Rumpf (*1975) Biologiestudium in Heidelberg, Dr. rer. nat. am Max-Planck-Institut für Biochemie (2006). Postdoc an der University of California San Francisco im Labor von Prof. Yuh Nung Jan, Juniorgruppenleiter an der WWU Münster seit 2013. Dr. Rumpf untersucht Pruning-Mechanismen in Drosophila.

Sandra Rode (*1987) Biologiestudium an der WWU Münster, Masterarbeit im Labor von C. Klämbt über die Funktion von Gliazellen. Seit 2013 Promotionsstudentin an der WWU zum Thema Pruning von C4da-Neuronen.

Svende Herzmann (*1988) Biologiestudium an der WWU Münster, Masterarbeit im Labor von C. Klämbt über die Entwicklung von Gliazellen. Seit 2013 Promotionsstudentin an der WWU zum Thema Pruning von C4da-Neuronen.

Rafael Krumkamp (*1987) Biologiestudium an der WWU Münster, Masterarbeit im Labor von S. Rumpf über Pruning von C4da-Neuronen. Seit 2015 Promotionsstudent an der WWU zum selben Thema.
Literatur
Bishop, D.L., Misgeld, T., Walsh, M.K., Gan, W.B., Lichtman, J.W. (2004). Axon branch removal at developing synapses by axosome shedding. Neuron 44, 651–661.10.1016/j.neuron.2004.10.026Search in Google Scholar PubMed
Chung, W.S., Clarke, L.E., Wang, G.X., Stafford, B.K., Sher, A., Chakraborty, C., Joung, J., Foo, L.C., Thompson, A., Chen, C., Smith, S.J., Barres, B.A. (2013). Astrocytes mediate synapse elimination through MEGF10 and MERTK pathways. Nature 504, 394–400.10.1038/nature12776Search in Google Scholar PubMed PubMed Central
Han, C., Song, Y., Xiao, H., Wang, D., Franc, N.C., Jan, L.Y., Jan, Y.N. (2014). Epidermal cells are the primary phagocytes in the fragmentation and clearance of degenerating dendrites in Drosophila. Neuron 81, 544–560.10.1016/j.neuron.2013.11.021Search in Google Scholar PubMed PubMed Central
Huttenlocher, P.R. (1990). Morphometric study of human cerebral cortex development. Neuropsychologia 28, 517–527.10.1002/9780470753507.ch8Search in Google Scholar
Je, H.S., Yang, F., Ji, Y., Nagappan, G., Hempstead, B.L., Lu, B. (2012). Role of pro-brain-derived neurotrophic factor (proBDNF) to mature BDNF conversion in activity-dependent competition at developing neuromuscular synapses.Proc. Natl. Acad. Sci. USA 109, 15924–15929.10.1073/pnas.1207767109Search in Google Scholar PubMed PubMed Central
Kanamori, T., Yoshino, J., Yasunaga, K., Dairyo, Y., Emoto, K. (2015). Local endocytosis triggers dendritic thinning and pruning in Drosophila sensory neurons. Nat. Comm. 12, 6515.10.1038/ncomms7515Search in Google Scholar PubMed
Kirilly, D., Gu, Y., Huang, Y., Wu, Z., Bashirullah, A., Low, B.C., Kolodkin, A.L., Wang, H., Yu, F. (2009). A genetic pathway composed of Sox14 and Mical governs severing of dendrites during pruning. Nat. Neurosci. 12, 1497–1505.10.1038/nn.2415Search in Google Scholar PubMed PubMed Central
Lee, H.H., Jan, L.Y., Jan, Y.N. (2009). Drosophila IKK-related kinase Ik2 and Katanin p60-like 1 regulate dendrite pruning of sensory neuron during metamorphosis. Proc. Natl. Acad. Sci. USA. 106, 6363–6368.10.1073/pnas.0902051106Search in Google Scholar PubMed PubMed Central
Low, L.K., Liu, X.B., Faulkner, R.L., Coble, J., Cheng, H.J. (2008). Plexin signaling selectively regulates the stereotyped pruning of corticospinal axons from visual cortex. Proc. Natl. Acad. Sci. USA 105, 8136–8141.10.1073/pnas.0803849105Search in Google Scholar PubMed PubMed Central
Schafer, D.P., Lehrman, E.K., Kautzman, A.G., Koyama, R., Mardinly, A.R., Yamasaki, R., Ransohoff, R.M., Greenberg, M.E., Barres, B.A., Stevens, B. (2012). Microglia sculpt postnatal neural circuits in an activity- and complement-dependent manner. Neuron 24, 691–705.10.1016/j.neuron.2012.03.026Search in Google Scholar PubMed PubMed Central
Schuldiner, O., Yaron, A. (2015). Mechanisms of developmental neurite pruning. Cell Mol Life Sci 72, 101–119.10.1007/s00018-014-1729-6Search in Google Scholar PubMed PubMed Central
Williams, D.W., Kondo, S., Krzyzaowska, A., Hiromi, Y., Truman, J.W. (2006). Local caspase activity directs engulfment of dendrites during pruning. Nat. Neurosci. 9, 1234–1236.10.1038/nn1774Search in Google Scholar PubMed
© 2017 by De Gruyter
Articles in the same Issue
- Frontmatter
- Editorial
- Mehr als nur ein neuer Look
- Übersichtsartikel
- Unser hungriges Gehirn: Welche Rolle spielen Gliazellen bei der Energieversorgung?
- Our hungry brain: Which role do glial cells play for the energy supply?
- Untersuchung und Modulation kortikaler Inhibition mittels transkranieller Magnetstimulation
- Assessment and modulation of cortical inhibition using transcranial magnetic stimulation
- Mechanismen des Neuritischen Prunings
- Mechanisms of Neurite Pruning
- Funktionen der GABAergen Übertragung im unreifen Gehirn
- Functions of GABAergic transmission in the immature brain
- Cholinerge Rückkopplungen auf den auditorischen Hirnstamm
- Cholinergic top-down influences on the auditory brainstem
- Forschungsförderung
- DFG Schwerpunktprogramm (SPP) 1926 “Next Generation Optogenetics – Tools and Application”
- Rezension
- Joachim Müller-Jung: Hirnforschung 8: Sprache und Kommunikation (F.A.Z.-Hörbuch)
- Ich glaub, mich trifft der Schlag: Warum das Gehirn tut, was es tun soll, oder manchmal auch nicht
- Nachruf
- Peter H. Seeburg (21.8.1944–22.8.2016)
- Nachrichten
- Nachrichten
Articles in the same Issue
- Frontmatter
- Editorial
- Mehr als nur ein neuer Look
- Übersichtsartikel
- Unser hungriges Gehirn: Welche Rolle spielen Gliazellen bei der Energieversorgung?
- Our hungry brain: Which role do glial cells play for the energy supply?
- Untersuchung und Modulation kortikaler Inhibition mittels transkranieller Magnetstimulation
- Assessment and modulation of cortical inhibition using transcranial magnetic stimulation
- Mechanismen des Neuritischen Prunings
- Mechanisms of Neurite Pruning
- Funktionen der GABAergen Übertragung im unreifen Gehirn
- Functions of GABAergic transmission in the immature brain
- Cholinerge Rückkopplungen auf den auditorischen Hirnstamm
- Cholinergic top-down influences on the auditory brainstem
- Forschungsförderung
- DFG Schwerpunktprogramm (SPP) 1926 “Next Generation Optogenetics – Tools and Application”
- Rezension
- Joachim Müller-Jung: Hirnforschung 8: Sprache und Kommunikation (F.A.Z.-Hörbuch)
- Ich glaub, mich trifft der Schlag: Warum das Gehirn tut, was es tun soll, oder manchmal auch nicht
- Nachruf
- Peter H. Seeburg (21.8.1944–22.8.2016)
- Nachrichten
- Nachrichten