Untersuchung und Modulation kortikaler Inhibition mittels transkranieller Magnetstimulation
-
Andreas Vlachos
Prof. Dr. Andreas Vlachos studierte Medizin an der Goethe-Universität in Frankfurt/M. (2001–2008) und promovierte nach einem Forschungsaufenthalt am Weizmann Institute of Science (Rehovot, Israel) im Jahre 2010 in Frankfurt/M. über die Rolle des aktin-bindenden Proteins Synaptopodin bei synaptischer Plastizität. 2011 erhielt er den Nachwuchspreis der Anatomischen Gesellschaft und habilitierte 2014 für das Gesamtfach Anatomie. Im März 2016 folgte er einem Ruf auf eine W2-Professur für Anatomie an die Heinrich-Heine-Universität in Düsseldorf. Der wissenschaftliche Schwerpunkt seiner Arbeitsgruppe liegt in der Untersuchung und Modulation homöostatischer Struktur-Funktions-Beziehungen im Zentralnervensystem.
 ,
Klaus Funke
,
Klaus Funke
Prof. Dr. Klaus Funke erwarb 1983 sein Diplom in Biologie mit Studien am spinalen somatosensorischen System der Taube. Die elektrophysiologische und histologische Charakterisierung der spezifischen und multimodalen somatosensorischen Repräsentationsareale im Vorderhirn der Taube war dann Gegenstand seiner Promotion im Jahr 1988. Als wissenschaftlicher Mitarbeiter und später akademischer Rat in der Abteilung für Neurophysiologie der Medizinischen Fakultät der Ruhr-Universität Bochum widmete er sich der zustandsabhängigen (EEG, Neuromodulatoren) Verarbeitung visueller Signale im Thalamus und Kortex und habilitierte sich 1996 in Physiologie. Seit nunmehr 15 Jahren untersucht er die zellulären Mechanismen der transkraniellen Magnetstimulation mittels elektrophysiologischer und histologisch/biochemischer Ansätze und ist derzeit Mitglied des German Center for Brain Stimulation (GCBS) als Teil des BMBF Forschungsnetzes zu psychiatrischen Erkrankungen.
Prof. Dr. Ulf Ziemann ist Klinischer Direktor der Abteilung Neurologie mit Schwerpunkt neurovaskuläre Erkrankungen, und Direktor am Hertie-Institut für Klinische Hirnforschung, Universität Tübingen. Seine wissenschaftlichen Schwerpunkte liegen im Bereich der systemischen Neurowissenschaften, Motorkortex Erregbarkeit und Plastizität, motorisches Lernen, nicht-invasive Hirnstimulation inklusive hirnzustandsabhängiger Stimulation und Neuropharmakologie. Er ist Chef-Editor von Clinical Neurophysiology. Er erhielt mehrere Preise, darunter den Richard-Jung-Preis der Dt. Gesellschaft für klinische Neurophysiologie und funktionelle Bildgebung, den NIH Merit Award und den NIH Fellowship Award for Research Excellence.

Zusammenfassung
Die Transkranielle Magnetstimulation (TMS) ist eine nicht-invasive Hirnstimulationstechnik, die zu diagnostischen, therapeutischen und wissenschaftlichen Zwecken in der Neurologie und Psychiatrie eingesetzt wird. Sie beruht auf dem physikalischen Grundprinzip der elektromagnetischen Induktion und ermöglicht die lokale Aktivierung kortikaler Areale durch den intakten Schädel des wachen Probanden oder Patienten. Eine sogenannte repetitive TMS (rTMS) kann zu anhaltenden Veränderungen der kortikalen Erregbarkeit führen. Die TMS stellt somit einen vielversprechenden Ansatz zur nicht-invasiven Untersuchung neuronaler Netzwerke, sowie deren lang anhaltender Beeinflussung dar. Trotz ihres mittlerweile breiten klinischen Einsatzes, sind die zellulären und molekularen Wirkmechanismen der TMS noch nicht genau geklärt. Etablierte Therapiekonzepte gehen von einer Wiederherstellung einer krankheitsbedingt veränderten kortikalen Erregbarkeit aus, die auf einer „Langzeit-Potenzierung“ oder „Langzeit-Depression“ erregender Synapsen beruhen könnte. Tatsächlich zeigen tierexperimentelle Studien, dass die rTMS in der Lage ist, anhaltende Veränderungen erregender Neurotransmission zu bewirken. Es bleibt allerdings unklar, auf welche Weise synaptische Veränderungen, die durch eine elektromagnetische Aktivierung der Hirnrinde bewirkt werden und demnach nicht kontext- oder verhaltensspezifisch sind, einen positiven Einfluss auf Hirnfunktionen haben. Neuere Befunde deuten nun darauf hin, dass die rTMS in der Lage ist, neben erregenden auch hemmende neuronale Netzwerke anhaltend zu beeinflussen. So gelang es nachzuweisen, dass 10 Hz repetitive Magnetstimulation zu einer kalziumabhängigen „Langzeit-Depression“ inhibitorischer GABAerger Synapsen führt. Da der Verminderung hemmender Neurotransmission (=Disinhibition) eine wichtige Rolle bei der Bahnung assoziativer Plastizität erregender Synapsen beigemessen wird, ist davon auszugehen, dass rTMS-induzierte Disinhibition die Ausbildung kontext- und verhaltensspezifischer synaptischer Veränderungen begünstigt. Das Modell derrTMS-induzierten lokalen Disinhibition bietet somit einen attraktiven Erklärungsansatz für die Beobachtung, dass eine scheinbar unspezifische exogene Magnetstimulation spezifische endogene strukturelle, funktionelle und molekulare Veränderungen kortikaler Synapsen fördern könnte. Untersuchungen zu Wirkungen rTMS-induzierter Disinhibition auf synaptische Plastizität in geeigneten Tiermodellen (in vitro und in vivo) sind Gegenstand aktueller Forschung. Neben ihrem diagnostischen und therapeutischen Potenzial stellt die TMS somit auch einen vielversprechenden methodischen Ansatz zur Durchführung klinisch orientierter translationaler Plastizitätsstudien dar.
Einführung
Die Fähigkeit des Gehirns, auf einen spezifischen Stimulus mit strukturellen, funktionellen und molekularen Veränderungen zu reagieren wird im Allgemeinen als Plastizität bezeichnet (Konorski 1948). Aktivitätsabhängige Veränderungen an den Kontaktstellen zwischen Nervenzellen, sogenannte synaptische Plastizität, spielt hierbei eine außerordentlich wichtige Rolle bei physiologischen Hirnfunktionen, wie beispielsweise Lern- und Gedächtnisprozessen oder Orientierung in Raum und Zeit. In den letzten Jahrzehnten wurden in diesem Kontext zelluläre und molekulare Wirkmechanismen der sogenannten assoziativen Plastizität (Hebb 1949), die als zelluläres Korrelat kontext- und verhaltensspezifischer Anpassungsreaktionen im Gehirn gilt, intensiv beforscht (siehe z. B. Bliss und Collingridge 1993; Nicoll und Roche 2013).
Hinweise für plastische Veränderungen im Gehirn des Menschen gehen unter anderem auf TMS-Studien zurück (Ziemann et al. 2008). Bei der TMS handelt es sich um ein nicht-invasives Hirnstimulationsverfahren, das auf dem physikalischen Prinzip der elektromagnetischen Induktion beruht. Hierbei wird ein sehr kurzes (~500µs) und starkes Magnetfeld (>1Tesla) mittels einer geeigneten Spule über dem Schädel des Probanden/Patienten aufgebaut (Abb. 1a). Dieses Feld führt innerhalb des Schädels zur Induktion elektrischer Felder im Gehirn, die eine Aktivierung von Nervenzellen bewirken. Da die Magnetfelder die Haut und den intakten Schädelknochen durchdringen (= transkranial), gilt dieses Stimulationsverfahren als nicht-invasiv. Die TMS gilt unter Beachtung etablierter Sicherheitsrichtlinien (Rossi et al. 2009) als ungefährlich und wird im Allgemeinen gut toleriert.
Die Effekte einzelner TMS-Pulse über dem primären Motorkortex werden typischerweise durch die Ableitung sogenannter Motorisch Evozierter Potenziale (MEP) im Zielmuskel erfasst und quantifiziert (Abb. 1b). Kombinierte Stimulationen des Motorkortex und Rückenmarks dienen hierbei der diagnostischen Bestimmung der zentralmotorischen Leitungszeit, die z. B. bei demyelinisierenden Erkrankungen wie Multipler Sklerose verlängert ist. Die Applikation von Reizzügen mit mehreren hundert TMS-Pulsen (repetitive TMS, rTMS) über dem Motorkortex kann zu einer anhaltenden Veränderung der kortikalen Erregbarkeit führen: Hochfrequente rTMS (≥5Hz) bewirkt typischerweise eine Erhöhung der kortiko-spinalen Erregbarkeit, gemessen als Zunahme der MEP-Amplitude, wohingegen niederfrequente rTMS (1Hz) selbige herabsetzt. Basierend auf pharmakologischen Studien und Analogien zu klassischen Plastizitätsexperimenten der Grundlagenforschung wurde die Hypothese aufgestellt, dass rTMS-induzierte Veränderungen der kortikalen Erregbarkeit eine Langzeitpotenzierung bzw. Langzeitdepression erregender Kontakte zwischen Nervenzellen darstellen könnte (Ziemann et al. 2008). Tatsächlich gelang es in organotypischen Hirnschnittkulturen nachzuweisen, dass repetitive Magnetstimulation strukturelle, funktionelle und molekulare Veränderungen an erregenden Synapsen induziert, die einer Langzeitpotenzierung und somit assoziativer synaptischer Plastizität entsprechen (Vlachos et al. 2012). Scheinbar konsistent mit diesen Befunden zeigte sich, dass rTMS das Lernverhalten im Tierexperiment verbessert (Mix et al. 2010) und Bewegungslernen nach einem Schlaganfall beim Menschen fördern kann (Brodie et al. 2014; Volz et al., 2016). Es bleibt allerdings unklar, auf welche Weise aktivitätsabhängige Modifikationen von Nervenverbindungen, die durch mehrere hundert exogene Magnetpulse bewirkt werden, komplexe Hirnfunktionen beim Gesunden und Kranken verbessern könnten.

Transkranielle Magnetstimulation (TMS). (a) Darstellungen transkranieller Magnetstimulation (b) des Motorkortex und Rückenmarks. Die Effekte der TMS werden mittels Ableitung von Motorisch Evozierten Potentialen (MEP) quantifiziert.
Rolle inhibitorischer Netzwerke bei assoziativer synaptischer Plastizität
Die Bedeutung hemmender neuronaler Netzwerke für physiologische Hirnfunktionen wurde in den letzten Jahren intensiv beforscht (Tremblay et al. 2016). Davon ausgehend, dass inhibitorische Interneurone die Aktivität und Erregbarkeit neuronaler Netzwerke kontrollieren, wurde die Hypothese aufgestellt, dass eine Reduktion hemmender Neurotransmission, sogenannte Disinhibition, Lern- und Gedächtnisprozesse und somit auch assoziative synaptische Plastizität erregender Synapsen bahnen könnte. Tatsächlich gelang es in Tierexperimenten aufzuzeigen, dass eine Reduktion GABAerger Inhibition Lern- und Gedächtnisprozesse begünstigt, wohingegen erhöhte Inhibition Plastizität hemmt (Tremblay et al. 2016). Mittlerweile gilt als gesichert, dass lokale disinhibitorische Netzwerke eine zentrale regulatorische Rolle bei Lern-/Gedächtnisprozessen spielen (Letzkus et al. 2015).
Neben der Plastizität GABAerger Synapsen (Froemke 2015) kann eine lokale Disinhibition durch eine Reihe unterschiedlicher Netzwerkmotive erzielt werden: z. B. durch (1) herabgesetzte glutamaterge Aktivierung lokaler hemmender Interneurone, (2) erhöhte Aktivität inhibitorischer Projektionsneurone, die lokale inhibitorische Interneurone anderer kortikaler Areale hemmen, oder (3) durch neuromodulatorische Projektionsbahnen, die aktivierend auf spezifische lokale Interneurone wirken, die wiederum die Hemmung anderer lokaler Interneurone vermitteln (Abb. 2; Letzkus et al. 2015).
Die klinische Relevanz dieser Mechanismen spiegelt sich auch in der Tatsache wider, dass Störungen der sogenannten Erregungs-/Hemmungs-Balance für das Auftreten neurologischer und psychiatrischer Symptome verantwortlich gemacht werden (z. B. Nelson und Valakh 2015). Aus klinischer Sicht erscheint somit die Untersuchung und Modulation (dis)inhibitorischer Netzwerke äußerst attraktiv. Geeignete diagnostische und/oder interventionelle Maßnahmen könnten diesbezüglich die Fähigkeit des menschlichen Gehirns zur Expression endogener Plastizität erfassen und darüber hinaus beeinflussen. Die TMS stellt in diesem Kontext ein überaus interessantes klinisches Werkzeug dar.
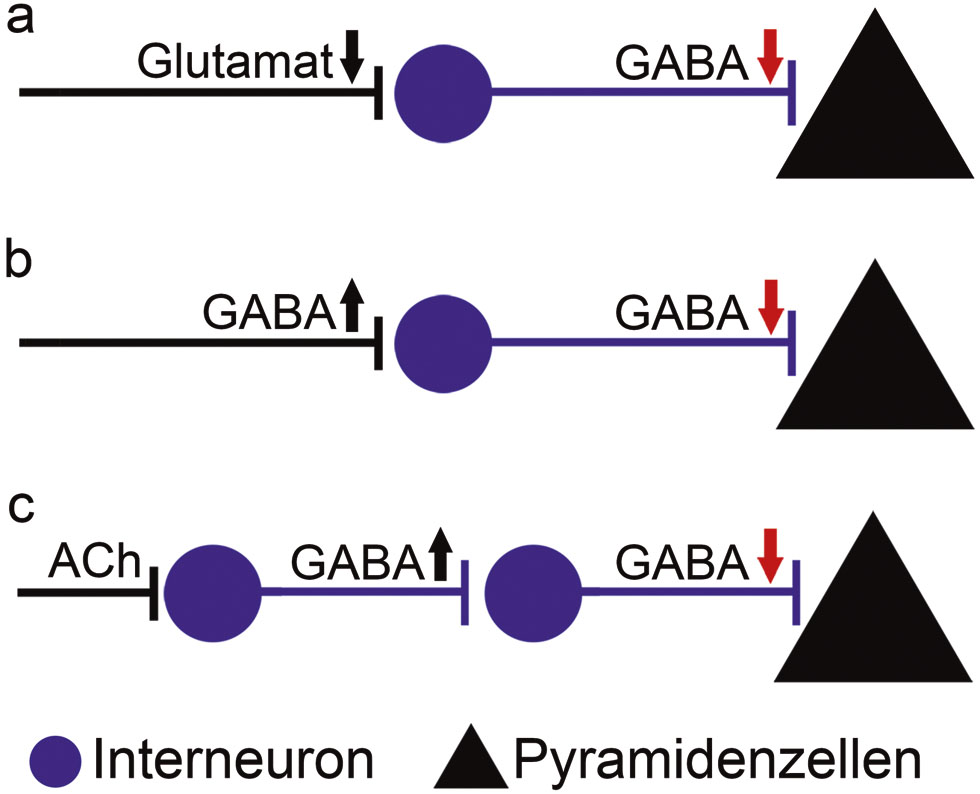
Netzwerkmotive lokaler Disinhibition. Neben einer „Langzeit-Depression“ inhibitorischer Synapsen auf Pyramidenzellen kann eine lokale Disinhibition durch eine Reihe unterschiedlicher Netzwerkmotive erzielt werden: z. B. durch (a) herabgesetzte glutamaterge Aktivierung lokaler hemmender Interneurone, (b) erhöhte Aktivität inhibitorischer Projektionsneurone, die lokale Interneurone innervieren, oder (c) durch neuromodulatorische Projektionsbahnen (z. B. cholinerge Projektionen; Acetylcholin, ACh), die aktivierend auf spezifische lokale Interneurone wirken, die wiederum die Hemmung anderer lokaler Interneurone vermitteln.
TMS-basierte Untersuchung inhibitorischer kortikaler Netzwerke
Funktionelle Untersuchungen intra- und interkortikaler Inhibition im Motorkortex des Menschen gehen überwiegend auf TMS-Studien zurück (Di Lazzaro und Ziemann 2013). In diesem Bereich wurde in den letzten Jahren eine Reihe spezifischer Doppelpulsprotokolle etabliert: So beruht die Untersuchung der sogenannten „short-interval intracortical inhibition“ (SICI) auf der Applikation eines unterschwelligen konditionierenden TMS-Pulses, der selbst kein MEP induziert, gefolgt von einem überschwelligen Test-Puls innerhalb eines kurzen Intervalls (1–5ms), wohingegen bei der Untersuchung sogenannter „long-interval intracortical inhibition“ (LICI) zwei überschwellige TMS-Pulse mit längerem Intervall (50–200ms) gepaart werden. In beiden Fällen kommt es zu einer intervallabhängigen Inhibition des durch den Test-Puls ausgelösten MEPs. Während die zugrundeliegenden Netzwerkmechanismen noch ungeklärt sind (Di Lazzaro und Ziemann 2013), geht man davon aus, dass der erste Puls eine charakteristische (direkte oder indirekte) Aktivierung inhibitorischer Interneurone bewirkt, die eine quantifizierbare Abschwächung des Test-MEPs zur Folge hat. Pharmakologische Studien deuten darauf hin, dass SICI den Einfluss ionotroper GABAA-Rezeptoren abbildet, während LICI eher auf metabotrope GABAB-Rezeptor zurückzuführen ist (Ziemann et al. 2015). Demnach ermöglichen charakteristische TMS-Protokolle prinzipiell die nicht-invasive, funktionelle Untersuchung lokaler kortikaler Inhibition im Motorkortex des Menschen.
Eine bedeutende Limitierung stellt in diesem Kontext allerdings die Tatsache dar, dass zur Quantifizierung der TMS-Effekte die Ableitung von MEPs erforderlich ist. Abgesehen von der Tatsache, dass es sich hierbei um die Messung von Potenzialen handelt, die nicht unmittelbar am Ort der Stimulation abgeleitet werden, sondern nachgeschaltet mindestens zwei Synapsen vom stimulierten Motorkortex entfernt liegen (Abb. 1b), lassen sich diese Protokolle auch nicht auf andere (krankheitsrelevante) Hirnareale übertragen. Ein direkter Bezug zwischen MEP-Amplitude und kortikaler Erregbarkeit bleibt durch den ungeklärten Einfluss der „Filterfunktionen“ kortiko-spinaler Synapsen sowie der motorischen Endplatte kontrovers. In diesem Zusammenhang zeichnen sich intensive Bemühungen ab, mittels TMS-evozierter elektroenzephalografischer (EEG) Potenziale kortikale Inhibition unabhängig vom stimulierten Kortexareal direkter zu erfassen (Rogasch und Fitzgerald 2013; Premoli et al. 2014). Unterstützt werden diese Ansätze durch weitere Verfahren, wie z. B. GABA-Magnetresonanzspektroskopie (Stagg et al. 2011). Inwiefern plastizitäts-induzierende rTMS-Protokolle allerdings in der Lage sind, TMS-evozierte EEG-Signale lang anhaltend zu modulieren und welche Mechanismen diesen Änderungen zugrunde liegen, wurde bislang nicht systematisch untersucht.
rTMS-induzierte Modulation inhibitorischer Netzwerke
Während der Einfluss der rTMS auf erregende Synapsen tierexperimentell als gesichert gilt (Tang et al. 2015), bleiben rTMS-induzierte Veränderungen hemmender Synapsen wenig erforscht. Es liegen allerdings robuste Grundlagendaten vor, die aufzeigen, dass spezifische rTMS-Protokolle kalziumbindende Moleküle beeinflussen, die als Aktivitäts- und Plastizitätsmarker inhibitorischer Interneurone gewertet werden können (Caroni 2015). So bewirkt beispielsweise die sogenannte intermittierende theta-burst Stimulation (iTBS) eine Reduktion der Parvalbumin (PV)-Expression in hemmenden Neuronen des Rattenkortex, wohingegen eine kontinuierliche theta-burst Stimulation (cTBS) und eine niederfrequente rTMS (1 Hz) eher die Calbindin-Expression in einer anderen Klasse von inhibitorischen Neuronen reduzieren (Benali et al. 2011). Da PV-positive Interneurone die somatische Inhibition von Projektionsneuronen vermitteln, während Calbindin-positive Interneurone auf Dendriten wirken, deuten diese Arbeiten daraufhin, dass die rTMS je nach Stimulationsprotokoll spezifische Aspekte der Netzwerk-Inhibition (z. B. Kontrolle der Ausgangs- vs. Eingangsaktivität an Pyramidenzellen) beeinflussen könnte. Interessanterweise zeigte sich in den bisherigen Studien vornehmlich eine Abnahme der entsprechenden Markermoleküle im Sinne einer rTMS-induzierten Disinhibition.
Bisher nicht publizierte Daten deuten darauf hin, dass Interneurone mit reduzierter PV-Expression weniger kortikale glutamaterge Eingänge haben, als solche mit einer hohen PV-Expression (siehe hierzu auch Abb. 2). Ähnliche Zusammenhänge zwischen PV-Expression und der Balance erregender zu hemmenden synaptischen Eingängen wurden auch im Hippokampus von Mäusen beschrieben (Donato et al. 2013). Eine elektrophysiologische Studie im primären somatosensorischen Kortex der Ratte zeigte dann auch, dass eine durch iTBS-rTMS-induzierte Abnahme der PV-Expression von einer Steigerung sensorischer Antworten begleitet ist (Thimm und Funke 2015), die ursächlich für die zuvor beschriebene Steigerung der Lernleistung sein könnte (Mix et al. 2010), sodass diese Studien das Model der rTMS-induzierten Disinhibition stützen. Allerdings lassen Untersuchungen inhibitorischer Aktivitäts- und Plastizitätsmarker nur indirekte Aussagen über rTMS-induzierte funktionelle Veränderungen inhibitorischer Synapsen zu.
rTMS-induzierte LangzeitDepression inhibitorischer Synapsen
In einer aktuellen Studie untersuchten wir die Effekte der repetitiven Magnetstimulation (rMS) auf strukturelle, funktionelle und molekulare Eigenschaften inhibitorischer Synapsen (Lenz et al. 2016). In organotypischen Hirnschnittkulturen der Maus gelang es mittels Einzelzellableitungen zu zeigen, dass rMS tatsächlich in der Lage ist, eine Abschwächung inhibitorischer Neurotransmission zu bewirken. Diese funktionellen Veränderungen gehen mit strukturellen und molekularen Änderungen inhibitorischer Postsynapsen einher: einer Verkleinerung und Destabilisierung von Gephyrin-Aggregaten, den wichtigsten postsynaptischen Ankermolekülen der GABAA-Rezeptoren (Tyagarajan und Fritschy 2014; Kneussel und Hausrat 2016).
Die rMS-induzierte Abschwächung inhibitorischer Neurotransmission erfordert die Aktivierung von spannungsabhängigen Natriumkanälen, NMDA-Rezeptoren und L-Typ spannungsabhängigen Kalziumkanälen während der Stimulation. Außerdem sind die Effekte in Anwesenheit eines Inhibitors der Calcineurin-abhängigen Phosphatase nicht nachweisbar. Wir schließen aus diesen Befunden, dass rMS kalziumabhängige Phosphorylierungs-/Dephosphorylierungsreaktionen induziert, die zu einer Destabilisierung von Gephyrin-Aggregaten führen können und dadurch eine lang anhaltende Verminderung der GABAergen Neurotransmission bewirken. Die durch extrasynaptische GABA-Rezeptoren vermittelte tonische Inhibition war in unseren Experimenten nach rMS nicht verändert (Lenz et al., 2016).
Die pharmakologische Hemmung Calpain-abhängiger Proteasen konnte die Effekte der rMS nicht verhindern. Da Calpain unter anderem für die kalziumabhängige Degradation von Gephyrin unter Bedingungen einer Übererregung verantwortlich gemacht werden kann (Tyagarajan und Fritschy 2014), legen diese Daten nahe, dass das eingesetzte 10Hz rMS-Protokoll nicht zu einer übermäßigen und potenziell toxischen Erregung neuronaler Netzwerke führt, die sonst mit einem proteolytischen Abbau essenzieller synaptischer Bestandteile einhergeht.
Die rTMS-induzierten Veränderungen von Gephyrin ließen sich auch in vivo in anästhesierten Mäusen nachweisen, sodass unsere Arbeiten insgesamt aufzeigen, dass die rTMS ein vielversprechendes Interventionsverfahren zur Beeinflussung Gephyrin-abhängiger inhibitorischer Neurotransmission darstellt. Dieser Schluss ist von klinischer Bedeutung, da Veränderungen des humanen Genlocus für Gephyrin mit einem erhöhten Risiko zur Ausbildung von Autismus, Schizophrenie und Epilepsie in Verbindung gebracht wird (Lionel et al. 2013). Gemeinsam mit den oben beschriebenen Arbeiten zu den Effekten der rTMS auf Markermoleküle hemmender Interneurone zeigen diese Arbeiten, dass die rTMS in der Lage ist, anhaltende Veränderungen inhibitorischer Neurotransmission im Sinne einer Disinhibition zu bewirken.
Wechselwirkung zwischen rTMS-induzierten Veränderungen an hemmenden und erregenden Synapsen?
Offensichtlich ist es mittels rTMS möglich, sowohl hemmende als auch erregende Synapsen zu modulieren. Interessanterweise zeigen unsere Experimente, dass beide Prozesse kalziumabhängig sind und die Aktivierung von NMDA-Rezeptoren und L-Typ spannungsabhängigen Kalziumkanälen erfordern (Vlachos et al. 2012; Lenz et al. 2015, 2016). Hieraus ergibt sich die wichtige Frage nach der zeitlichen, örtlichen und molekularen Wechselwirkung zwischen rTMS-induzierten inhibitorischen und exzitatorischen synaptischen Veränderungen.
Basierend auf dem vorgestellten Modell der rTMS-induzierten Disinhibition ergibt sich die interessante Hypothese, dass die rTMS vornehmlich auf hemmende Synapsen wirken könnte, wohingegen die Veränderungen erregender Synapsen ein Ergebnis der „Disinhibitions-bedingten Bahnung assoziativer synaptischer Plastizität“ darstellen könnte. Tatsächlich zeigen unsere in vitro Experimente, dass es nach 10 Hz rMS nicht zu einer unmittelbaren post-tetanischen Verstärkung erregender Synapsen kommt, wie sie bei lokaler elektrischer Stimulation in klassischen LTP-Experimenten beobachtet wird. Vielmehr kommt es zu einer langsamen Potenzierung erregender Synapsen innerhalb der ersten 1–2 h nach Stimulation, die 2–4 h nach Stimulation ein Plateau erreicht (Vlachos et al. 2012). Dagegen lässt sich eine Veränderung der Expression inhibitorischer Markermoleküle im Rattenkortex bereits ca. 30 Minuten nach rTMS nachweisen (Hoppenrath und Funke 2013). Diese Beobachtungen unterstützen die Hypothese, dass die Plastizität erregender Synapsen nicht unmittelbarer Effekt der rTMS ist. Vor diesem Hintergrund erscheint die Untersuchung der genauen zeitlichen Abfolge rTMS-induzierter Langzeit-Depression inhibitorischer Synapsen (Lenz et al. 2016) und rTMS-induzierter Veränderungen erregender Synapsen (Vlachos et al. 2012) überaus wichtig. Experimente, in denen die Netzwerkaktivität vor, während und nach rTMS systematisch moduliert wird (z. B. mittels pharmakologischer oder optogenetischer Techniken) könnten in diesem Zusammenhang wichtige Erkenntnisse zur Frage beitragen, inwiefern die Effekte der rTMS vom Aktivitätszustand des Netzwerkes abhängig sind. Wir sind zuversichtlich, dass diese Studien den Einfluss rTMS-induzierter Disinhibition auf Metaplastizität und homöostatische Plastizität beleuchten werden und darüber hinaus wichtige Beiträge zu der jüngst geführten Diskussion bezüglich einer erheblichen inter- und intra-individuellen Variabilität der rTMS-Effekte beim Menschen beitragen können (Muller-Dahlhaus et al. 2008; Hamada et al. 2013; Lopez-Alonso et al. 2014). Es ist interessant in diesem Kontext zu vermuten, dass die zellulären und molekularen Prozesse, die durch ein spezifisches rTMS-Protokoll per se induziert werden, wenig variabel sind, die Langzeiteffekte aber sehr wohl von inter- und intra-individuellen Faktoren abhängig sein könnten (z. B. Alter, Aufmerksamkeitszustand, Tageszeit der Stimulation etc. – Ridding und Ziemann 2010).
Synapsenspezifische Effekte der rTMS
Das Modell der rTMS-induzierten Disinhibition bietet einen interessanten Erklärungsansatz für die Beobachtung, dass eine scheinbar unspezifische exogene Stimulation kortikaler Netzwerke zu kontext- und verhaltensspezifischen Veränderungen führen kann. So wäre beispielsweise denkbar, dass vor einer rehabilitativen Trainingsmaßnahme nach einem Schlaganfall 10Hz rTMS oder iTBS eingesetzt wird, um den Trainingseffekt durch „Disinhibitions-bedingte Bahnung assoziativer Plastizität“ an spezifischen erregenden Synapsen zu unterstützen (Volz et al. 2016). Der Einfluss rTMS-induzierter Disinhibition auf physiologische und pathologische Hirnfunktionen ist Gegenstand aktueller Forschung.
Tatsächlich gelang es bereits zu zeigen, dass die rMS in der Lage ist, Plastizität an spezifischen Synapsen einer Zelle zu induzieren (Abb. 3). So konnten nachgewiesen werden, dass 10 Hz rMS vornehmlich auf dendritische aber nicht somatische Inhibition wirkt (Lenz et al. 2016). Dasselbe Stimulationsprotokoll führt wiederum zu einer Potenzierung somanaher erregender Synapsen auf kleinen dendritischen Dornen (Vlachos et al. 2012; Lenz et al. 2015). Die genauen Mechanismen, die zu diesen spezifischen Effekten führen, sind noch nicht geklärt. Nichtsdestotrotz ist es aus anatomischer Sicht ein interessantes Konzept, dass spezifische rTMS-Parameter zu einer selektiven Verstärkung somanaher erregender Synapsen auf Projektionsneuronen führen könnten, z. B. thalamo-kortikaler Projektionen in Schicht IV des Kortex, während dendritische Disinhibition die Fähigkeit zur Ausbildung assoziativer Plastizität an Dendriten der äußeren Assoziationsschichten bahnen könnte. Inwiefern diese synapsenspezifischen Effekte der rMS, die auf CA1 – Pyramidenzellen in organtypischen Schnittkulturen des Hippokampus nachweisbar sind, auch auf andere Neurone und Netzwerke des Kortex übertragen werden können, muss geklärt werden.

Synapsenspezifische Effekte der repetitiven Magnetstimulation (rMS). Trotz des verhältnismäßig großen induzierten elektromagnetischen Feldes kann die rMS differenzielle Effekte an spezifischen Synapsen einer Pyramidenzelle induzieren. In vitro Experimente zeigen, dass 10 Hz rMS zu einer Abschwächung dendritischer Inhibition sowie Verstärkung somanaher erregender Synapsen auf kleinen dendritischen Dornen führt. Die genauen Mechanismen, die zu diesen spezifischen Effekten führen sind noch nicht geklärt.
Wie wirkt TMS während der Stimulation?
Eine der wichtigen noch ungeklärten Fragen ist die nach der Wirkung der TMS während der Stimulation. Die meisten Annahmen in diesem Bereich beruhen auf computerbasierten Modellen (Esser et al. 2005; Rusu et al. 2014).In diesem Zusammenhang besteht die große technische Herausforderung in den starken elektromagnetischen Feldern, die während der Stimulation aufgebaut werden. Diese Felder erschweren elektrophysiologische Untersuchungen, d. h. Ableitungen neuronaler Aktivität auf Einzelzellebene, auch wenn es einige wenige Arbeiten gibt, die bei niedriger Feldstärke und geeigneter Ausrichtung der Ableitelektroden durchgeführt werden konnten (Moliadze et al. 2005; Mueller et al. 2014; Pashut et al. 2014). Der Einsatz „kontaktfreier“ funktioneller Mikroskopieverfahren, z. B. mittels kalzium- oder spannungssensitiver Farbstoffe (Kozyrev et al. 2014; Murphy et al. 2016), deutet darauf hin, dass bei geringer Stimulationsintensität vornehmlich inhibitorische Interneurone aktiviert werden, während Projektionsneurone erst bei höheren Intensitäten rekrutiert werden (siehe auch Pashut et al. 2014). Somit bieten diese Studien einen plausiblen Erklärungsansatz für das bereits beschriebene SICI-Doppelpulsprotokoll, bei dem der unterschwellige konditionierende TMS-Puls zu einer GABAA-abhängigen Inhibition der durch den TMS Test-Puls induzierten MEP-Antwort führt.
Um die Effekte der rTMS genauer zu verstehen, ist es dringend erforderlich, das Aktivitätsmuster ganzer Populationen von Neuronen simultan mit hoher zeitlicher und örtlicher Auflösung abzuleiten. Diese Studien könnten auch dazu beitragen, die Rolle sogenannter rückwärts propagierender Aktionspotenziale zu beleuchten, die eine wichtige Rolle bei der rTMS-induzierten synaptischen Plastizität spielen könnten (Lenz et al. 2015). Fest steht, dass die gegenwärtig vorliegenden Informationen nicht ausreichen, um mittels computerbasierter Simulationen zuverlässige Voraussagen über die Wirkung der rTMS zu machen. Unsere pharmakologischen Studien zeigen bislang auf, dass die Aktivierung von spannungsabhängigen Natriumkanälen, NMDA-Rezeptoren und L-Typ spannungsabhängigen Kalziumkanälen während der Stimulation erforderlich ist, um rTMS-induzierte Veränderungen erregender und hemmender Synapsen in vitro zu induzieren. Der potenzielle Einfluss der TMS auf Gliazellen (Astrozyten, Oligodendrozyten und Mikrogliazellen) und auf das neurovaskuläre System müssen in diesem Kontext ebenfalls berücksichtigt werden.
Offene Fragen und einige Ausblicke
30 Jahre nach der Entwicklung des ersten TMS-Gerätes durch Anthony Barker und Kollegen (Barker et al. 1985) zeichnen sich wichtige Hinweise auf die zellulären und molekularen Wirkmechanismen der TMS ab. Gegenwärtige TMS-basierte Therapien führen allerdings nicht zu einer ausgeprägten und nachhaltigen Verbesserung neurologischer und psychiatrischer Symptome (Lefaucheur et al. 2014). Ein substanzieller Fortschritt in diesem Bereich wird durch das immer noch eingeschränkte Wissen über die Wirkmechanismen der TMS erschwert. Des Weiteren steht unser limitiertes Wissen bezüglich der Rolle synaptischer Plastizität unter pathologischen Bedingungen einem effizienten Einsatz der rTMS in der Klinik entgegen.
Arbeiten aus den letzten Jahren konnten zeigen, dass eine Störung assoziativer synaptischer Plastizität nicht zwingend als Merkmal neurologischer oder psychiatrischer Erkrankungen gewertet werden kann (Maggio und Vlachos 2014). Tatsächlich könnte eine Verminderung der Fähigkeit von Nervenzellen, synaptische Plastizität zu exprimieren auch protektiven Charakter haben, indem sogenannte „maladaptive Veränderungen“ verhindert werden. Gleichzeitig müssen mögliche negative Auswirkungen auf den Erfolg von Rehabilitationsmaßnahmen in diesem Kontext diskutiert werden. Unser gegenwärtiges Verständnis ist, dass sich der Schwellenwert zur Expression von Plastizität im Laufe einer Erkrankung des Gehirns ändern kann. Änderungen der Erregungs-/Hemmungs-Balance scheinen hierbei eine wichtige Rolle zu spielen, da wie in diesem Artikel diskutiert, Veränderungen inhibitorischer Synapsen die Plastizität an erregenden Synapsen beeinflussen. Fest steht, dass ein übergreifendes Verständnis zur Rolle unterschiedlicher Formen synaptischer Plastizität (assoziative, homöostatische Plastizität und Metaplastizität) unter pathologischen Bedingungen dringend erforderlich ist.
Zukünftige diagnostische und therapeutische Interventionen könnten darauf abzielen, zu unterschiedlichen Zeitpunkten im Laufe einer Erkrankung (und ggf. davor), in einzelnen kortikalen Arealen die Fähigkeit von Nervenzellen zur Expression von Plastizität zu erfassen und zu modulieren. Die TMS stellt in diesem Zusammenhang offensichtlich ein attraktives klinisches Werkzeug dar. Tatsächlich könnte es mittels der TMS auch gelingen, Nachteile der klassischen pharmakologischen Behandlung, der die erforderliche örtliche und zeitliche Spezifität fehlt, zu überwinden bzw. zu komplementieren. Während das diagnostische Potenzial der rTMS in Bezug auf (dis)inhibitorische Netzwerke weiter ausgearbeitet werden muss und weitere zelluläre und molekulare Effekte der rTMS berücksichtigt werden sollten, z. B. Einfluss auf Genexpression, mRNA-Transport/Stabilität, lokale Proteinbiosynthese, Interaktionen mit Mitochondrien, Rolle glialer und vaskulärer Faktoren, liefert das vorgestellte Konzept der rTMS-induzierten Disinhibition eine vielversprechende Arbeitshypothese für zukünftige Studien.
In diesem Kontext wird es nun auch erforderlich sein, den Einfluss der rTMS auf die beschriebenen Netzwerkmotive lokaler (Dis)Inhibition zu erfassen (Abb. 2); also den Einfluss der rTMS auf (1) glutamaterge Synapsen auf inhibitorischen Interneuronen, (2) auf interkortikale inhibitorische Projektionen und (3) neuromodulatorische Projektionsbahnen. Des Weiteren müssen die Effekte unterschiedlicher Stimulationsparameter systematisch untersucht werden, um beispielsweise zu überprüfen, ob mittels rTMS auch die Verstärkung inhibitorischer Synapsen oder die Modulation somatischer Inhibition möglich ist (Abb. 3). Ferner wird derzeit daran gearbeitet, Aktivitätszustände neuronaler Netzwerke des Gehirns (“brainstates”) mittels EEG in Echtzeit auszulesen und TMS-Pulse spezifisch repetitiv immer nur zu Zeitpunkten eines bestimmten Zustandes, z. B. verminderter instantaner Inhibition, zu applizieren (Zrenner et al. 2016). Dieser Ansatz basiert auf der Theorie, dass die Erregbarkeit neuronaler Netzwerke durch gepulste Inhibition reguliert wird (Jensen und Mazaheri 2010). Wir erwarten, dass hirnzustandsabhängige TMS gegenüber konventioneller “open-loop” Stimulation die rTMS-Effektstärke in Richtung lang anhaltender Plastizität erheblich steigern kann und somit die Zukunft therapeutischer rTMS-Protokolle in “closed-loop” Ansätzen liegt, die spontane rhythmische Oszillationen inhibitorischer Aktivität konsequent nutzen.
Perspektivisch könnten computerbasierte Simulationen in diesem Kontext äußerst hilfreich sein. Allerdings fehlen gegenwärtig die hierzu erforderlichen Grundlageninformationen zu den genauen Wirkmechanismen der TMS während und nach der Stimulation. Insgesamt ist zu erwarten, dass ein besseres Verständnis rTMS-induzierter Plastizität zu einer Optimierung bestehender Therapiekonzepte führen wird, die auf eine Beeinflussung der Erregungs-/Hemmungs-Balance abzielen (z. B. im Rahmen der Behandlung nach einem Schlaganfall oder bei der Epilepsietherapie). Wir sind überzeugt, dass das neue Konzept der rTMS-induzierten lokalen Disinhibition an Beachtung gewinnen wird. Neben ihrem diagnostischen und therapeutischen Potenzial, stellt die TMS somit auch einen vielversprechenden methodischen Ansatz zur Durchführung klinisch-orientierter translationaler Plastizitätsstudien dar.
Über die Autoren

Prof. Dr. Andreas Vlachos studierte Medizin an der Goethe-Universität in Frankfurt/M. (2001–2008) und promovierte nach einem Forschungsaufenthalt am Weizmann Institute of Science (Rehovot, Israel) im Jahre 2010 in Frankfurt/M. über die Rolle des aktin-bindenden Proteins Synaptopodin bei synaptischer Plastizität. 2011 erhielt er den Nachwuchspreis der Anatomischen Gesellschaft und habilitierte 2014 für das Gesamtfach Anatomie. Im März 2016 folgte er einem Ruf auf eine W2-Professur für Anatomie an die Heinrich-Heine-Universität in Düsseldorf. Der wissenschaftliche Schwerpunkt seiner Arbeitsgruppe liegt in der Untersuchung und Modulation homöostatischer Struktur-Funktions-Beziehungen im Zentralnervensystem.

Prof. Dr. Klaus Funke erwarb 1983 sein Diplom in Biologie mit Studien am spinalen somatosensorischen System der Taube. Die elektrophysiologische und histologische Charakterisierung der spezifischen und multimodalen somatosensorischen Repräsentationsareale im Vorderhirn der Taube war dann Gegenstand seiner Promotion im Jahr 1988. Als wissenschaftlicher Mitarbeiter und später akademischer Rat in der Abteilung für Neurophysiologie der Medizinischen Fakultät der Ruhr-Universität Bochum widmete er sich der zustandsabhängigen (EEG, Neuromodulatoren) Verarbeitung visueller Signale im Thalamus und Kortex und habilitierte sich 1996 in Physiologie. Seit nunmehr 15 Jahren untersucht er die zellulären Mechanismen der transkraniellen Magnetstimulation mittels elektrophysiologischer und histologisch/biochemischer Ansätze und ist derzeit Mitglied des German Center for Brain Stimulation (GCBS) als Teil des BMBF Forschungsnetzes zu psychiatrischen Erkrankungen.

Prof. Dr. Ulf Ziemann ist Klinischer Direktor der Abteilung Neurologie mit Schwerpunkt neurovaskuläre Erkrankungen, und Direktor am Hertie-Institut für Klinische Hirnforschung, Universität Tübingen. Seine wissenschaftlichen Schwerpunkte liegen im Bereich der systemischen Neurowissenschaften, Motorkortex Erregbarkeit und Plastizität, motorisches Lernen, nicht-invasive Hirnstimulation inklusive hirnzustandsabhängiger Stimulation und Neuropharmakologie. Er ist Chef-Editor von Clinical Neurophysiology. Er erhielt mehrere Preise, darunter den Richard-Jung-Preis der Dt. Gesellschaft für klinische Neurophysiologie und funktionelle Bildgebung, den NIH Merit Award und den NIH Fellowship Award for Research Excellence.
Danksagung:
Unsere Arbeiten wurden von der Deutschen Forschungsgemeinschaft und dem Bundesministerium für Bildung und Forschung gefördert.
Literatur
Barker, A.T., Jalinous, R., Freeston, I.L. (1985). Non-invasive magnetic stimulation of human motor cortex. Lancet 1, 1106–1107.10.1016/S0140-6736(85)92413-4Search in Google Scholar
Benali, A., Trippe, J., Weiler, E., Mix, A., Petrasch-Parwez, E., Girzalsky, W., Eysel, U.T., Erdmann, R., Funke, K. (2011). Theta-burst transcranial magnetic stimulation alters cortical inhibition. J Neurosci 31, 1193–1203.10.1523/JNEUROSCI.1379-10.2011Search in Google Scholar PubMed PubMed Central
Bliss, T.V., Collingridge, G.L. (1993). A synaptic model of memory: long-term potentiation in the hippocampus. Nature 361, 31–39.10.1038/361031a0Search in Google Scholar PubMed
Brodie, S.M., Meehan, S., Borich, M.R., Boyd, L.A. (2014). 5 Hz repetitive transcranial magnetic stimulation over the ipsilesional sensory cortex enhances motor learning after stroke. Front Hum Neurosci 8, 143.10.3389/fnhum.2014.00143Search in Google Scholar PubMed PubMed Central
Caroni, P. (2015). Regulation of Parvalbumin Basket cell plasticity in rule learning. Biochem Biophys Res Commun 460, 100–103.10.1016/j.bbrc.2015.02.023Search in Google Scholar PubMed
Di Lazzaro, V., Ziemann, U. (2013). The contribution of transcranial magnetic stimulation in the functional evaluation of microcircuits in human motor cortex. Front Neural Circuits 7, 18.10.3389/fncir.2013.00018Search in Google Scholar PubMed PubMed Central
Donato, F., Rompani, S.B., Caroni, P. (2013). Parvalbumin-expressing basket-cell network plasticity induced by experience regulates adult learning. Nature 504, 272–276.10.1038/nature12866Search in Google Scholar PubMed
Esser, S.K., Hill, S.L., Tononi, G. (2005). Modeling the effects of transcranial magnetic stimulation on cortical circuits. J Neurophysiol 94, 622–639.10.1152/jn.01230.2004Search in Google Scholar PubMed
Froemke, R.C. (2015). Plasticity of cortical excitatory-inhibitory balance. Annu Rev Neurosci 38, 195–219.10.1146/annurev-neuro-071714-034002Search in Google Scholar PubMed PubMed Central
Hamada, M., Murase, N., Hasan, A., Balaratnam, M., Rothwell, J.C. (2013). The role of interneuron networks in driving human motor cortical plasticity. Cereb Cortex 23, 1593–1605.10.1093/cercor/bhs147Search in Google Scholar PubMed
Hebb, D.O. (1949). The organization of behavior; a neuropsychological theory. New York,: Wiley.Search in Google Scholar
Hoppenrath, K., Funke, K. (2013). Time-course of changes in neuronal activity markers following iTBS-TMS of the rat neocortex. Neurosci Lett 536, 19–23.10.1016/j.neulet.2013.01.003Search in Google Scholar PubMed
Jensen, O., Mazaheri, A. (2010). Shaping functional architecture by oscillatory alpha activity: gating by inhibition. Front Hum Neurosci 4, 186.10.3389/fnhum.2010.00186Search in Google Scholar PubMed PubMed Central
Kneussel, M., Hausrat, T.J. (2016). Postsynaptic neurotransmitter receptor reserve pools for synaptic potentiation. Trends Neurosci 39, 170–182.10.1016/j.tins.2016.01.002Search in Google Scholar PubMed
Konorski, J. (1948). Conditioned reflexes and neuron organization. Cambridge Eng.: University Press.Search in Google Scholar
Kozyrev, V., Eysel, U.T., Jancke, D. (2014). Voltage-sensitive dye imaging of transcranial magnetic stimulation-induced intracortical dynamics. Proc Natl Acad Sci USA 111, 13553–13558.10.1073/pnas.1405508111Search in Google Scholar PubMed PubMed Central
Lefaucheur, J.P., Andre-Obadia, N., Antal, A., Ayache, S.S., Baeken, C., Benninger, D.H., Cantello, R.M., Cincotta, M., De Carvalho, M., De Ridder, D., Devanne, H., Di Lazzaro, V., Filipovic, S.R., Hummel, F.C., Jaaskelainen, S.K., Kimiskidis, V.K., Koch, G., Langguth, B., Nyffeler, T., Oliviero, A., Padberg, F., Poulet, E., Rossi, S., Rossini, P.M., Rothwell, J.C., Schonfeldt-Lecuona, C., Siebner, H.R., Slotema, C.W., Stagg, C.J., Valls-Sole, J., Ziemann, U., Paulus, W., Garcia-Larrea, L. (2014). Evidence-based guidelines on the therapeutic use of repetitive transcranial magnetic stimulation (rTMS). Clin Neurophysiol 125, 2150–2206.10.1016/j.clinph.2014.05.021Search in Google Scholar PubMed
Lenz, M., Galanis, C., Muller-Dahlhaus, F., Opitz, A., Wierenga, C.J., Szabo, G., Ziemann, U., Deller, T., Funke, K., Vlachos, A. (2016). Repetitive magnetic stimulation induces plasticity of inhibitory synapses. Nat Commun 7, 10020.10.1038/ncomms10020Search in Google Scholar PubMed PubMed Central
Lenz, M., Platschek, S., Priesemann, V., Becker, D., Willems, L.M., Ziemann, U., Deller, T., Muller-Dahlhaus, F., Jedlicka, P., Vlachos, A. (2015). Repetitive magnetic stimulation induces plasticity of excitatory postsynapses on proximal dendrites of cultured mouse CA1 pyramidal neurons. Brain Struct Funct 220, 3323–3337.10.1007/s00429-014-0859-9Search in Google Scholar PubMed
Letzkus, J.J., Wolff, S.B., Luthi, A. (2015). Disinhibition, a Circuit Mechanism for Associative Learning and Memory. Neuron 88, 264–276.10.1016/j.neuron.2015.09.024Search in Google Scholar PubMed
Lionel, A.C., Vaags, A.K., Sato, D., Gazzellone, M.J., Mitchell, E.B., Chen, H.Y., Costain, G., Walker, S., Egger, G., Thiruvahindrapuram, B., Merico, D., Prasad, A., Anagnostou, E., Fombonne, E., Zwaigenbaum, L., Roberts, W., Szatmari, P., Fernandez, B.A., Georgieva, L., Brzustowicz, L.M., Roetzer, K., Kaschnitz, W., Vincent, J.B., Windpassinger, C., Marshall, C.R., Trifiletti, R.R., Kirmani, S., Kirov, G., Petek, E., Hodge, J.C., Bassett, A.S., Scherer, S.W. (2013). Rare exonic deletions implicate the synaptic organizer Gephyrin (GPHN) in risk for autism, schizophrenia and seizures. Hum Mol Genet 22, 2055–2066.10.1093/hmg/ddt056Search in Google Scholar PubMed
Lopez-Alonso, V., Cheeran, B., Rio-Rodriguez, D., Fernandez-Del-Olmo, M. (2014). Inter-individual variability in response to non-invasive brain stimulation paradigms. Brain Stimul 7, 372–380.10.1016/j.brs.2014.02.004Search in Google Scholar PubMed
Maggio, N., Vlachos, A. (2014). Synaptic plasticity at the interface of health and disease: New insights on the role of endoplasmic reticulum intracellular calcium stores. Neuroscience 281, 135–146.10.1016/j.neuroscience.2014.09.041Search in Google Scholar PubMed
Mix, A., Benali, A., Eysel, U.T., Funke, K. (2010). Continuous and intermittent transcranial magnetic theta burst stimulation modify tactile learning performance and cortical protein expression in the rat differently. Eur J Neurosci 32, 1575–1586.10.1111/j.1460-9568.2010.07425.xSearch in Google Scholar PubMed
Moliadze, V., Giannikopoulos, D., Eysel, U.T., Funke, K. (2005). Paired-pulse transcranial magnetic stimulation protocol applied to visual cortex of anaesthetized cat: effects on visually evoked single-unit activity. J Physiol 566, 955–965.10.1113/jphysiol.2005.086090Search in Google Scholar PubMed PubMed Central
Mueller, J.K., Grigsby, E.M., Prevosto, V., Petraglia, F.W., 3rd, Rao, H., Deng, Z.D., Peterchev, A.V., Sommer, M.A., Egner, T., Platt, M.L., Grill, W.M. (2014). Simultaneous transcranial magnetic stimulation and single-neuron recording in alert non-human primates. Nat Neurosci 17, 1130–1136.10.1038/nn.3751Search in Google Scholar PubMed PubMed Central
Muller-Dahlhaus, J.F., Orekhov, Y., Liu, Y., Ziemann, U. (2008). Interindividual variability and age-dependency of motor cortical plasticity induced by paired associative stimulation. Exp Brain Res 187, 467–475.10.1007/s00221-008-1319-7Search in Google Scholar PubMed
Murphy, S.C., Palmer, L.M., Nyffeler, T., Müri, R.M., Larkum, M.E. (2016). Transcranial magnetic stimulation (TMS) inhibits cortical dendrites. Elife pii:e13598.10.7554/eLife.13598Search in Google Scholar PubMed PubMed Central
Nelson, S.B., Valakh, V. (2015). Excitatory/Inhibitory Balance and Circuit Homeostasis in Autism Spectrum Disorders. Neuron 87, 684–698.10.1016/j.neuron.2015.07.033Search in Google Scholar PubMed PubMed Central
Nicoll, R.A., Roche, K.W. (2013). Long-term potentiation: peeling the onion. Neuropharmacology 74, 18–22.10.1016/j.neuropharm.2013.02.010Search in Google Scholar PubMed PubMed Central
Pashut, T., Magidov, D., Ben-Porat, H., Wolfus, S., Friedman, A., Perel, E., Lavidor, M., Bar-Gad, I., Yeshurun, Y., Korngreen, A. (2014). Patch-clamp recordings of rat neurons from acute brain slices of the somatosensory cortex during magnetic stimulation. Front Cell Neurosci 8, 145.10.3389/fncel.2014.00145Search in Google Scholar PubMed PubMed Central
Premoli, I., Castellanos, N., Rivolta, D., Belardinelli, P., Bajo, R., Zipser, C., Espenhahn, S., Heidegger, T., Muller-Dahlhaus, F., Ziemann, U. (2014). TMS-EEG signatures of GABAergic neurotransmission in the human cortex. J Neurosci 34, 5603–5612.10.1523/JNEUROSCI.5089-13.2014Search in Google Scholar PubMed PubMed Central
Ridding, M.C., Ziemann, U. (2010). Determinants of the induction of cortical plasticity by non-invasive brain stimulation in healthy subjects. J Physiol 588, 2291–2304.10.1113/jphysiol.2010.190314Search in Google Scholar PubMed PubMed Central
Rogasch, N.C., Fitzgerald, P.B. (2013). Assessing cortical network properties using TMS-EEG. Hum Brain Mapp 34, 1652–1669.10.1002/hbm.22016Search in Google Scholar PubMed PubMed Central
Rossi, S., Hallett, M., Rossini, P.M., Pascual-Leone, A., Safety of TMS Consensus Group. (2009). Safety, ethical considerations, and application guidelines for the use of transcranial magnetic stimulation in clinical practice and research. Clin Neurophysiol 120, 2008–2039.10.1016/j.clinph.2009.08.016Search in Google Scholar PubMed PubMed Central
Rusu, C.V., Murakami, M., Ziemann, U., Triesch, J. (2014). A model of TMS-induced I-waves in motor cortex. Brain Stimul 7, 401–414.10.1016/j.brs.2014.02.009Search in Google Scholar PubMed
Stagg, C.J., Bestmann, S., Constantinescu, A.O., Moreno, L.M., Allman, C., Mekle, R., Woolrich, M., Near, J., Johansen-Berg, H., Rothwell, J.C. (2011). Relationship between physiological measures of excitability and levels of glutamate and GABA in the human motor cortex. J Physiol 589, 5845–5855.10.1113/jphysiol.2011.216978Search in Google Scholar PubMed PubMed Central
Tang, A., Thickbroom, G., Rodger, J. (2015). Repetitive Transcranial Magnetic Stimulation of the Brain: Mechanisms from Animal and Experimental Models. Neuroscientist.Search in Google Scholar
Thimm, A., Funke, K. (2015). Multiple blocks of intermittent and continuous theta-burst stimulation applied via transcranial magnetic stimulation differently affect sensory responses in rat barrel cortex. J Physiol 593, 967–985.10.1113/jphysiol.2014.282467Search in Google Scholar PubMed PubMed Central
Tremblay, R., Lee, S., Rudy, B. (2016). GABAergic Interneurons in the Neocortex: From Cellular Properties to Circuits. Neuron 91, 260–292.10.1016/j.neuron.2016.06.033Search in Google Scholar PubMed PubMed Central
Tyagarajan, S.K., Fritschy, J.M. (2014). Gephyrin: a master regulator of neuronal function? Nat Rev Neurosci 15, 141–156.10.1038/nrn3670Search in Google Scholar PubMed
Vlachos, A., Muller-Dahlhaus, F., Rosskopp, J., Lenz, M., Ziemann, U., Deller, T. (2012). Repetitive magnetic stimulation induces functional and structural plasticity of excitatory postsynapses in mouse organotypic hippocampal slice cultures. J Neurosci 32, 17514–17523.10.1523/JNEUROSCI.0409-12.2012Search in Google Scholar PubMed PubMed Central
Volz, L.J., Rehme, A.K., Michely, J., Nettekoven, C., Eickhoff, S.B., Fink, G.R., Grefkes, C. (2016). Shaping early reorganization of neural networks promotes motor function after Stroke. Cereb Cortex 26, 2882–2894.10.1093/cercor/bhw034Search in Google Scholar PubMed PubMed Central
Ziemann, U., Paulus, W., Nitsche, M.A., Pascual-Leone, A., Byblow, W.D., Berardelli, A., Siebner, H.R., Classen, J., Cohen, L.G., Rothwell, J.C. (2008). Consensus: Motor cortex plasticity protocols. Brain Stimul 1, 164–182.10.1016/j.brs.2008.06.006Search in Google Scholar PubMed
Ziemann, U., Reis, J., Schwenkreis, P., Rosanova, M., Strafella, A., Badawy, R., Muller-Dahlhaus, F. (2015). TMS and drugs revisited 2014. Clin Neurophysiol 126, 1847–1868.10.1016/j.clinph.2014.08.028Search in Google Scholar PubMed
Zrenner, C., Belardinelli, P., Muller-Dahlhaus, F., Ziemann, U. (2016). Closed-Loop Neuroscience and Non-Invasive Brain Stimulation: A Tale of Two Loops. Front Cell Neurosci 10, 92.10.3389/fncel.2016.00092Search in Google Scholar PubMed PubMed Central
© 2017 by De Gruyter
Articles in the same Issue
- Frontmatter
- Editorial
- Mehr als nur ein neuer Look
- Übersichtsartikel
- Unser hungriges Gehirn: Welche Rolle spielen Gliazellen bei der Energieversorgung?
- Our hungry brain: Which role do glial cells play for the energy supply?
- Untersuchung und Modulation kortikaler Inhibition mittels transkranieller Magnetstimulation
- Assessment and modulation of cortical inhibition using transcranial magnetic stimulation
- Mechanismen des Neuritischen Prunings
- Mechanisms of Neurite Pruning
- Funktionen der GABAergen Übertragung im unreifen Gehirn
- Functions of GABAergic transmission in the immature brain
- Cholinerge Rückkopplungen auf den auditorischen Hirnstamm
- Cholinergic top-down influences on the auditory brainstem
- Forschungsförderung
- DFG Schwerpunktprogramm (SPP) 1926 “Next Generation Optogenetics – Tools and Application”
- Rezension
- Joachim Müller-Jung: Hirnforschung 8: Sprache und Kommunikation (F.A.Z.-Hörbuch)
- Ich glaub, mich trifft der Schlag: Warum das Gehirn tut, was es tun soll, oder manchmal auch nicht
- Nachruf
- Peter H. Seeburg (21.8.1944–22.8.2016)
- Nachrichten
- Nachrichten
Articles in the same Issue
- Frontmatter
- Editorial
- Mehr als nur ein neuer Look
- Übersichtsartikel
- Unser hungriges Gehirn: Welche Rolle spielen Gliazellen bei der Energieversorgung?
- Our hungry brain: Which role do glial cells play for the energy supply?
- Untersuchung und Modulation kortikaler Inhibition mittels transkranieller Magnetstimulation
- Assessment and modulation of cortical inhibition using transcranial magnetic stimulation
- Mechanismen des Neuritischen Prunings
- Mechanisms of Neurite Pruning
- Funktionen der GABAergen Übertragung im unreifen Gehirn
- Functions of GABAergic transmission in the immature brain
- Cholinerge Rückkopplungen auf den auditorischen Hirnstamm
- Cholinergic top-down influences on the auditory brainstem
- Forschungsförderung
- DFG Schwerpunktprogramm (SPP) 1926 “Next Generation Optogenetics – Tools and Application”
- Rezension
- Joachim Müller-Jung: Hirnforschung 8: Sprache und Kommunikation (F.A.Z.-Hörbuch)
- Ich glaub, mich trifft der Schlag: Warum das Gehirn tut, was es tun soll, oder manchmal auch nicht
- Nachruf
- Peter H. Seeburg (21.8.1944–22.8.2016)
- Nachrichten
- Nachrichten