Cholinergic top-down influences on the auditory brainstem
-
Thomas Künzel
Thomas Künzel studied biology at the Ruhr University in Bochum. He received his doctoral degree from the RWTH Aachen University with a dissertation on the development of auditory brainstem neurons under the supervision of Prof. Harald Luksch. He then briefly took a position as a research assistant at RWTH Aachen before continuing his postdoctoral training at Erasmus Medical Center in Rotterdam with Prof. Marcel van der Heijden and Prof. Gerard Borst. In 2011, he returned to the RWTH Aachen as an assistant to Prof. Hermann Wagner and has headed an independent group, which is supported by the German science foundation DFG (PP1608 “Ultrafast and temporally precise information processing: Normal and dysfunctional hearing”) since 2012. His research focus is on development and cellular physiology of phase-coding neurons in avian and mammalian auditory brainstem circuits.
 and
Hermann Wagner
and
Hermann Wagner
Hermann Wagner studied biology in Ulm, Erlangen and Tübingen. After his diploma and doctoral thesis with Werner Reichardt at the Max-Planck-Institute of Biological Cybernetics, he was a postdoc with Mark Konishi at the California Institute of Technology, Pasadena, USA. He returned to the Max-Planck Institute to work with Kuno Kirschfeld, received a Heisenberg stipend to work with Geoff Manley at the TU Munich, before he took the chair of Zoology/Animal Physiology at the RWTH Aachen. His interests are in mechanisms of orientation, ranging from ecology via neuro-ethology to system physiology and cognition.

Abstract
Descending connections are present in many sensory systems and support adaptive information processing. This allows the sensory brain to code a wider range of inputs. A well characterized descending system is the olivo-cochlear cholinergic innervation of the inner ear, which mediates a reduction of the sensitivity of the inner ear upon perception of intense sounds. Because this inhibits the response to background noise, the olivo-cochlear system supports detection of transient sound events. Olivo-cochlear neurons also innervate the cochlear nucleus through axon collaterals. Here, acetylcholine increases the excitability of central neurons without reducing their temporal precision. Thus their target neurons in the superior olivary complex can more effectively process binaural temporal cues. We argue that the central effect of the olivo-cochlear system augments the peripheral effect. In addition, olivo-cochlear cholinergic neurons are under top-down control of cortical inputs, providing further adaptability of information processing on the level of the auditory brainstem.
Imagine you return to a cocktail party after a quiet walk outside. Suddenly you are in a room filled with loud music and many talking people and are engulfed in intense noisiness coming from many directions. The sound level is much higher now and relevant information can be overpowered (“masked”) by the various noise sources in the room. Identification and localization of individual speakers is still possible, although more difficult than in a quiet room. What are the mechanisms in the brain that allow you to do this?
Sensory pathways are often mistakenly seen as purely ascending streams of information, where a given neuron feeds information only “upwards” to stations of increasing complexity in a higher hierarchical level. In a well-known example, time-coding neurons in the cochlear nucleus are contacted by giant synaptic terminals of the auditory nerve (Felmy and Künzel, 2014) and pass information about the timing of the perceived sound up along the auditory pathway into binaural nuclei with more complex functions. These neurons are clearly dominated by information flow from hierarchically lower to higher stations, as the ascending synaptic connection from the auditory nerve onto the time-coding neuron is very powerful. Although it is well established that the auditory periphery and the first stages of the auditory pathway indeed show activity dependent dynamics, these adaptive phenomena can only rely on the limited information that is available at the given neuron. In this scheme, however, the background noise from the cocktail party could easily drown the important information, since one can expect it to activate the ascending stream of auditory information as intense as the relevant auditory event. However, recurrent connections within hierarchical levels and descending connections towards neurons in lower hierarchical orders that provide dynamic and adaptive changes to the sensory systems are common in all sensory modalities (Engel et al., 2001; Briggs and Usrey, 2011; Gilbert and Li, 2013; D’Souza and Vijayaraghavan, 2014; Llorca-Torralba et al., 2016), including the auditory brainstem. One especially well characterized top-down connection in a sensory pathway is the olivo-cochlear projection in the lower auditory brainstem, which mediates the so-called olivo-cochlear reflex (Guinan, 2006). This neuronal system is driven by auditory inputs and provides cholinergic innervation back to the inner ear and also to central neurons in the lower auditory pathway, like the time-coding neurons in the cochlear nucleus that we mentioned earlier. So while bottom-up computations definitely play a large role in time-coding neurons, descending projections become more important for optimal encoding of information as it exists, for example, in adaptive processing. In this article, we shall first briefly review the anatomy and physiology of the peripheral olivo-cochlear pathway, discuss its function in hearing and then move on to discuss recent findings about the central effects of the olivo-cochlear innervation on neurons in the cochlear nucleus. We shall use this circuit as a study case for the general problem of the interaction of feedforward and feedback connections. We will also work out that the function of centrally connected collaterals of the olivo-cochlear innervation complement the effects it has on the inner ear. This may in fact provide additional avenues for complex top-down influences on adaptive information processing in the auditory pathway.
Anatomy and physiology of the olivo-cochlear system
Auditory information reaches the brain via the auditory nerve that innervates the three subdivisions of the cochlear nucleus. From there, auditory information is conveyed to the superior olivary complex and other brain nuclei (Fig. 1A). The olivo-cochlear innervation (Fig. 1B) of the inner ear, the so called olivo-cochlear bundle (OCB), is formed by the projecting axons of two major bilateral neuron groups, the medial (MOC) and the lateral (LOC) olivo-cochlear neurons (Brown, 2011). Both neuron groups are situated in close vicinity to the superior olivary complex. The MOC neurons are found close to the medial superior olive, mostly in the ventral nucleus of the trapezoid body, while the LOC neurons are found inside of and surrounding the lateral superior olive (Radtke-Schuller et al., 2015). Axons from both the contralateral and the ipsilateral MOC and LOC innervate the inner ear (Brown, 1993). Axons from MOC neurons are thick and heavily myelinated, indicating rapid action potential conduction. In contrast, axons of LOC neurons are thin and non-myelinated, which indicates a slow function that is not time-critical. MOC axons form cholinergic contacts with the basal parts of outer hair cells (Wersinger and Fuchs, 2011; Elgoyhen and Katz, 2012). In contrast, LOC axons contact auditory nerve endings close to inner hair cells and together with acetylcholine as a neurotransmitter they use a variety of co-transmitters (Guinan, 2006).
The innervation of the LOC neurons is not well understood. It is assumed that LOC neurons are driven by sound and receive inputs from the ipsilateral posteroventral cochlear nucleus (PVCN). Axons of LOC neurons then almost exclusively innervate the ipsilateral cochlea. Thus the majority of LOC fibers are uncrossed. However, it remains unclear what the exact effect of the innervation of auditory nerve endings by the LOC axons is. Generally, LOC activity seems to modulate the excitability of auditory nerve fibers mostly in such a way that increased LOC activity inhibits auditory nerve activity (Groff and Liberman, 2003; Darrow et al., 2006).
In contrast to the LOC system, more is known about MOC anatomy and physiology (Brown et al., 2003). Individual MOC neurons respond to narrow-band sound. They are innervated by neurons in the contralateral PVCN. Most MOC axons cross the midline below the floor of the fourth ventricle and enter the inner ear through the vestibular part of the eighth nerve (Brown, 1993). These axons form the double-crossed pathway for the ipsilateral olivo-cochlear reflex, where sound in the right cochlear nucleus excites MOC neurons in the left brain hemisphere which, in turn, innervate the right inner ear again. However, a certain proportion of MOC axons do not cross the midline. Instead, these axons join the ipsilateral OCB, forming a second pathway, where sound in the right ear excites MOC neurons in the left hemisphere, which then innervate the left inner ear. This contralateral reflex pathway is of special interest, because through the olivo-cochlear innervation sound in one ear may influence the way sounds in the opposing ear are processed. From a mechanistic point of view, it would be interesting to separate ipsi- and contralateral contributions to the olivo-cochlear reflex. This is possible experimentally by decoupling the input to the two ears: When playing broadband noise to one ear, the sensitivity of the other ear is reduced. This is called contralateral suppression and presumably mediated by the olivo-cochlear system (Knudson et al., 2014). The exact proportion of ipsi- vs. contralateral olivo-cochlear innervation is variable and depends on the species and tonotopic position on the cochlea. In general it is assumed, that the ipsilateral component is 2–3 times stronger than the contralateral component (Guinan, 2006). However, in humans the proportion of crossed vs. uncrossed MOC axons seems to be roughly equal. This also holds true for the low frequency parts of the auditory system of animals (Guinan, 2006; Brown, 2011). Both contra- and ipsilateral MOC axons provide feedback to outer hair cells (OHC) with cholinergic synapses. This synaptic contact is well studied (Elgoyhen and Katz, 2012). Acetylcholine, upon release from the MOC axon, binds to pharmacologically unique cholinergic receptors on the postsynaptic site. These receptors contain alpha-9 and alpha-10 nicotinic acetylcholine-receptor subunits, which show a high calcium conductivity (Elgoyhen et al., 2001). The fast effect of the acetylcholine-induced calcium influx is hyperpolarization of the OHC membrane potential mediated by calcium-activated potassium channels (Elgoyhen and Katz, 2012). This hyperpolarization yields effective inhibition of the OHC function by directly influencing the membrane-potential dependent motility of the OHC. Furthermore, on longer time-scales the cholinergic calcium influx also influences the stiffness of the OHC by acting on cytoskeleton and prestin proteins, again impacting the motility of the OHC (Cooper and Guinan, 2003; He et al., 2003). Accordingly, the olivo-cochlear reflex can be physiologically separated into a component having a fast effect and into a component having a slow effect. These two components may be attributed to the two molecular effects described above.

Neuroanatomy of ascending and descending projections in the auditory brainstem. A Simplified depiction of the ascending auditory projections with emphasis on structures involved in sound localization. Idealized coronal section of the brain is shown, dashed line depicts midline to show crossed vs. uncrossed projections. Blue arrows indicate predominantly excitatory connections, red arrows indicate inhibitory connections. Please note that some connections and nuclei were left out for clarity, for greater detail see reviews Pollak et al., 2003; Grothe et al., 2010; Grothe and Pecka, 2014. B Overview of the cholinergic descending projections in the auditory brainstem. Green arrows indicate cholinergic connections (dark green used to distinguish LOC from MOC axons/neurons). Purple arrows indicate descending cortical connections onto cholinergic neurons. Inputs to LOC neurons are not well known (dashed blue arrow with question marks). Adapted from: Guinan, 2006; Schofield et al., 2011. Abbreviations: AVCN anteroventral cochlear nucleus, DCN dorsal cochlear nucleus, LOC lateral olivo-cochlear, LSO lateral superior olive, MNTB medial nucleus of the trapezoid body, MOC medial olivo-cochlear, MSO medial superior olive, OCB olivo-cochlear bundle, PMT Nuclei of the ponto-mesencephalic tegmentum, PVCN posteroventral cochlear nucleus, VCN ventral cochlear nucleus.
Olivo-cochlear activation improves hearing in noise
What is the effect of MOC feedback on hearing? By acting on the OHC motility the MOC effect reduces the gain of the cochlear amplification, which reduces the response of the cochlea to a given sound level (Wersinger and Fuchs, 2011). This manifests as an upward shift of the response curve of auditory nerve fibers to sound levels (Fig. 2A). Thus, activation of the olivo-cochlear reflex per se makes the inner ear less sensitive to sounds (Cooper and Guinan, 2003; Francis and Guinan, 2010). This has two possible functions: 1) the olivo-cochlear reflex may serve to protect the inner ear from loud sounds. 2) the response to sounds in background noise is improved. The first effect is very intuitive. There is indeed evidence that animals with experimentally impaired olivo-cochlear reflex suffer more from noise-induced trauma and show premature auditory aging (Guinan, 2006; Liberman et al., 2014). In other words, the olivo-cochlear reflex augments other otoprotective reflexes like the middle-ear reflex, which actively reduces transmission of sound by the middle-ear through stiffening of the ossicular chain, upon intense sounds. However there is some debate whether this is really the main function of the elaborate olivo-cochlear system (Kirk and Smith, 2003), especially since the olivo-cochlear reflex is already activated by sound intensities way below traumatic levels. Hence it must have at least one or more additional functions.
The second function of the olivo-cochlear reflex is concerned with the response of the inner ear to constant broadband stimulation, which often occurs in real life as background noise (e. g. rushing water or wind in the trees). The changes that occur in the inner ear due to the background noise can be analyzed by measuring the responses to defined stimuli like tones with and without noise. One finds that broadband noise causes reduction or compression of the dynamic response range of auditory nerve fibers to tones. This is due to the increase of the baseline response by background that constantly excites all auditory nerve fibers. At the same time, the maximal response rate to the test stimulus is diminished due to adaptation caused by the constantly high excitation of the hairs cells of the inner ear (Fig. 2B). However, broadband noise of moderate levels already effectively activates the olivo-cochlear reflex. The response to transient sounds, especially tones, that occur among the constant background noise is then improved by the inhibitory function of the olivo-cochlear reflex (Kawase et al., 1993). How is that possible? By inhibiting the constant response to the broadband noise, both the increased baseline response and consecutively also the adaptation is reduced. The olivo-cochlear reflex, thus, allows a stronger response to the test tone hidden in the noise, a phenomenon called “unmasking”. The unmasking of tones is not perfect however: the response to the tone in noise, despite the augmentation by the olivo-cochlear reflex, has lower sensitivity and a smaller dynamic range compared to the response to tone in silence (Fig. 2C). Nevertheless, in this interpretation, the action of olivo-cochlear reflex appears capable to critically improve the response to transient narrow-band sounds in noise, which is a very common and highly relevant listening task. Think about human speech or animal vocalizations, both of which are characterized by brief periods of specific tone-like sounds (“formants”) that must be reliably detected despite strong broadband background noise like wind roaring in the leaves of a tree.
Furthermore, olivo-cochlear activation not only influences detection of tones but also improves the localization of sounds in noise in animals (May et al., 2004) and humans (Andeol et al., 2011; Boothalingam et al., 2016). Sound localization is mostly a computational task of the brain where binaural information must be compared and analyzed. So far it is not fully understood how the peripheral olivo-cochlear reflex could help with this. Below we will show results from our own experiments that may explain how an improvement of tone-detection might be possible on a neuronal level. Furthermore, it was proposed, that the LOC system may balance overall neuronal excitation levels between the two ears, which is needed for sound localization with interaural intensity differences (Darrow et al., 2006; but see: Larsen and Liberman, 2010). Interestingly, in human subjects with transection of the peripheral part of the olivocochlear bundle (Scharf et al., 1994, 1997), hearing was essentially normal. Also, correlations between MOC activation strength, which is usually measured only in the periphery using otoacustic emissions, and success in complex listening tasks are not always strong (Boothalingam et al., 2016). One possible interpretation for these findings that we would like to stress in this article is that the peripheral part of the olivo-cochlear system is less decisive for complex hearing tasks and a central portion of the olivo-cochlear system begins to play a larger role. In the following, we shall therefore review what is known about the innervation of central neurons by the olivo-cochlear axons, especially recent findings on the effects of acetylcholine on time-coding neurons.
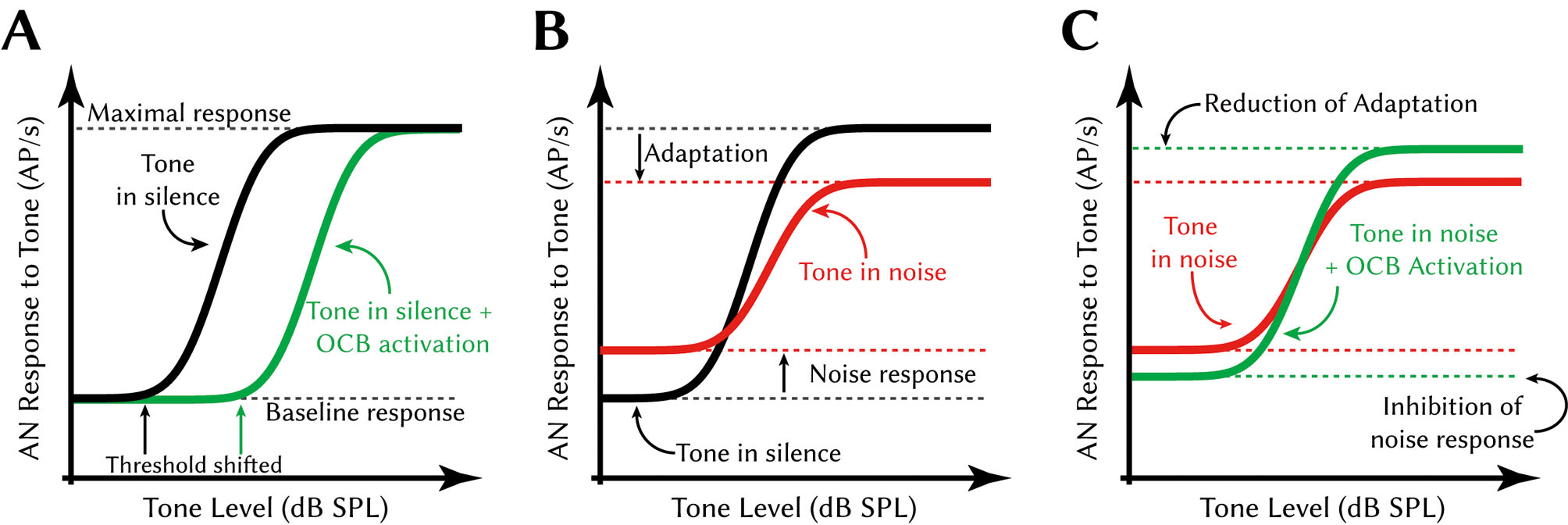
Effect of the olivo-cochlear reflex on auditory nerve response to tones. A Schematic depiction of the threshold shift caused by OCB activation. Black line: rate-level function of auditory nerve responses to ipsilaterally presented tones in silence. Green line: rate-level function of auditory nerve responses to ipsilaterally presented tones with contralaterally presented noise. The contralaterally presented noise activates the olivo-cochlear reflex via MOC axons. B Schematic depiction of the effect of background noise on the tone response. Black line: as in A. Red line: rate-level function of auditory nerve responses to ipsilaterally presented tones hidden in ipsilaterally presented noise. Due to the response to the constant noise stimulus, the baseline response is higher. Due to adaptation effects caused by the constant response to the noise, the maximal response is smaller. C Schematic depiction of the effect of OCB activation on the response to tones in noise. Red line: as in B. Green line: rate-level function of auditory nerve responses to ipsilaterally presented tones hidden in ipsilaterally presented noise with contralaterally presented noise. The olivo-cochlear activation inhibits the constant response to the ipsilaterally presented noise and thus releases the auditory nerve fibers from their adapted state. Adapted from: Kawase et al., 1993; Guinan, 2006. Please note that effects are exaggerated in this figure for clarity. Abbreviations: AN auditory nerve, AP action potential, SPL sound pressure level.
Central effects of olivo-cochlear activation
The efficacy of MOC activation in unmasking the response to a tone in noise has been shown not only on the level of the auditory nerve (Kawase and Liberman, 1993; Kawase et al., 1993) but also in central stations of the auditory pathway, like the cochlear nucleus (Mulders et al., 2008) and the inferior colliculus (Seluakumaran et al., 2008). However, physiological analysis of the olivo-cochlear reflex in central neurons points towards a dual effect of the olivo-cochlear activation, one indirectly via the peripheral projection and a second more direct central effect (Mulders et al., 2002). In accordance with this it was shown in numerous studies for several species that olivo-cochlear axons form collaterals that enter the cochlear nucleus (Brown et al., 1988; Benson and Brown, 1990; Horváth et al., 2000; Brown, 2011; Baashar et al., 2015) and innervate primary neurons in the cochlear nucleus (Benson et al., 1996). There olivo-cochlear activity induced by presenting noise stimuli to the contralateral ear excited ventral cochlear nucleus neurons (VCN) (Mulders et al., 2003, 2009). Other, non olivo-cochlear, commissural connections also exist and cause rapid or onset type contralateral excitation (Bledsoe et al., 2009), but are not discussed further here. We hypothesize that the contralaterally induced olivo-cochlear responses in the VCN discussed above are indicative of a general theme: whenever the inner ear on one side receives olivo-cochlear inhibition (be it ipsi-, contra- or, in most cases, bilaterally induced) axon collaterals of the OCB deliver excitation to the VCN. This is backed by anatomical studies, stating that a large proportion of cholinergic axons entering the VCN comes from MOC/LOC neurons in the superior olivary complex, with about 44 % originating in the ipsilateral SOC (Mellott et al., 2011).
What are the target neurons of this excitatory collateral connection of the OCB in the VCN? In-vivo physiological studies indicated onset neurons (Mulders et al., 2003) or almost all neuron types (Mulders et al., 2008) as potential targets of the olivo-cochlear reflex. Given the complex nature of recurrent connections within the VCN (Campagnola and Manis, 2014), it is hard to decipher how the olivo-cochlear reflex is effecting the function of the VCN in hearing tasks like sound localization, from these results alone. However, in vitro experiments showed that T-stellate neurons in the VCN (Oertel et al., 2011) clearly respond to acetylcholine (Fujino and Oertel, 2001), the neurotransmitter of the OCB. These neurons are excited by acetylcholine through nicotinic and muscarinic cholinergic receptors. T-stellate cells project locally and remotely to diverse targets in the auditory pathway, among others the VCN itself, the dorsal cochlear nucleus, periolivary regions in the superior olivary complex, the nuclei of the lateral lemniscus and ultimately the inferior colliculus (Oertel et al., 2011). Due to the nature of the responses of T-stellate cells to sounds one can derive that they provide information about the level and the envelope modulation of stimulus and thus, as a population, they are able to encode the frequency content of the perceived signal. Interestingly, T-stellate cells provide excitatory inputs to contralateral MOC neurons, forming another positive feedback loop (Oertel et al., 2011). It was hypothesized that the extra excitation provided by the cholinergic collaterals extends the dynamic range of response of the T-stellate cells. However, due to the diffuse nature of the functions and projections of T-stellate cells it is still hard to understand what the exact role of the cholinergic input is. To address this, a recent study analyzed the cholinergic innervation of another type of VCN neurons, the time-coding spherical bushy cells (Goyer et al., 2016).
Cholinergic modulation of time-coding neurons
Spherical bushy cells (and the similar globular bushy cells) are contacted by the auditory nerve with giant somatic terminals, the so called endbulbs of Held (Felmy and Künzel, 2014). Through these powerful synaptic connections and their specialized physiology, these neurons retain the exquisite temporal precision of the action potentials in the auditory nerve or even improve this temporal code with the help of convergent excitatory inputs and inhibition. By exactly locking the times of action potential firing to a certain phase of the perceived sound, the spherical bushy cell encode the relative temporal alignment of the stimulus, which makes sound localization with interaural time differences possible. Action potentials of spherical bushy cells indeed encode the phase of sounds with microsecond precision. The temporal cues are exploited in the medial superior olivary nucleus (Goldberg and Brown, 1969; Grothe et al., 2010), a direct bilateral target of spherical bushy cells. Here, the precision with which the spherical bushy cells can encode the relative timing of the stimulus is translated into how precise the source of a sound can be localized in the horizontal plane.
The computational properties of spherical bushy cells are straightforward. Therefore, these cells are especially well suited to study top-down influences on the auditory brainstem. In a recent study (Goyer et al., 2016) it was thus analyzed how spherical bushy cells respond to acetylcholine. In an in-vitro approach in acute brain slices the authors locally applied carbachol, a cholinergic agonist, to spherical bushy cells while recording in whole-cell configuration (Fig. 3). This experiment also allowed simultaneous stimulation of auditory nerve inputs to the spherical bushy cells. It was found that spherical bushy cells are depolarized through alpha-7 nicotinic acetylcholine receptors on a time-scale that is congruent with the fast component of the olivo-cochlear reflex, i. e. a few hundred milliseconds (Fig. 3A). However, on top of that the study also found that the resting membrane potential of spherical bushy cells is depolarized by carbachol in a time course of minutes (Fig. 3B). The study demonstrated that muscarinic acetylcholine receptors are responsible for this. Since blocking muscarinic receptors slowly hyperpolarized the resting membrane potential (Fig. 3B), it was concluded that a certain constant level of muscarinic activation is involved in setting the actual resting membrane potential of the spherical bushy cell. The authors went on to test the effects of the cholinergic modulation on synaptic transmission and found that under the effect of acetylcholine more action potentials are caused by the same activity of auditory nerve fibers (Fig. 3C). In in-vivo experiments iontophoretic application of carbachol increased the spontaneous firing rate of spherical bushy cells. Remarkably, further in-vitro experiments showed no reduction of the temporal precision in which spherical bushy cells encode the phase of the stimulus, when the excitability is increased by acetylcholine (Fig. 3C). This means that through the increased excitability caused by acetylcholine a higher amount of well-timed action potentials are produced by spherical bushy cells. This likely improves sound localization with temporal cues in the following stages of the auditory pathway. In a computer model of spherical bushy cells, the inner ear and the auditory nerve cholinergic modulation of the spherical bushy cells excitability expanded the dynamic response range of the model neurons and increased the responses to faint sounds, without decreasing the temporal precision of the response (Fig. 3D). This was exactly in line with the hypothesis outlined above: the central effect of the olivo-cochlear reflex augments the peripheral olivo-cochlear effect insofar that it further expands the dynamic response range for tones in noise, which was compressed by the constant background noise (cf. Fig. 2B, C). Taken together, these recent results (Goyer et al., 2016) allow the conclusion that the central part of the olivo-cochlear reflex causes a compensatory increase in excitability in time-coding neurons to counteract the peripheral reduction in sensitivity. It can be concluded that this feedback system stabilizes the quantity of well-timed action-potentials delivered by the spherical bushy cells to higher-order nuclei over a wider range of hearing conditions.

Effect of cholinergic modulation on time-coding central neurons. A Nicotinic transient inward currents recorded from gerbil spherical bushy cell (SBC) in whole-cell voltage clamp (–65mV holding potential) upon brief pressure application (arrow) of carbachol (a stable analogon of acetylcholine) are blocked by the specific alpha-7 nicotinic antagonist methyllycaconitine. Green trace: carbachol induced inward current. Orange trace: current measured upon carbachol application in the presence of methyllycaconitine. B SBC resting membrane potential (RMP) is modulated by muscarinic receptors over minutes. Green data points: changes of RMP measured after repeated pressure application (green bar) of carbachol. Red data points: RMP measured after wash-in (red bar) of the muscarinic antagonist atropine. Orange data points: RMP measured after repeated pressure application of carbachol in the presence of atropine. Error bars depict standard error of the mean for y-axis. C Cholinergic modulation increases spike-probability in vitro but does not decrease temporal precision. Ctrl = control condition, Carb = carbachol application. Blue line denotes input vector strength, a metric for temporal precision of action potentials in relation to the phase of the sound stimulus. Asterisk: p<0.05, paired t-test. D Simulated rate-level functions of SBC, spontaneous rate/baseline rate subtracted. Gray dashed line: simulated auditory nerve input rate. Black line: simulated SBC response, control condition. Green line: simulated SBC response, carbachol condition (RMP depolarized). Red line: simulated SBC response, atropine condition (RMP hyperpolarized). All data in this figure replotted and adapted from Goyer et al., 2016.
Cholinergic top-down influence of the cortex on the auditory brainstem
In addition to what was discussed so far there are further levels of complexity in the anatomy of the cholinergic innervation of the cochlear nucleus: first, the neurons of the olivo-cochlear system in the superior olivary complex not only receive the direct auditory inputs from interneurons in the cochlear nucleus, they are also under top-down control from higher brain areas. It was shown that descending projections from the inferior colliculus (Brown et al., 2013), the auditory cortex (Schofield et al., 2011) and even non-auditory brain areas (Brown et al., 2013) reach all the way down to the olivo-cochlear neurons in the superior olivary complex. Furthermore, there is a whole class of cholinergic neurons in a midbrain area called the ponto-mesencephalic tegmentum that is under cortical control and projects widely into every station of the auditory pathway, including the cochlear nucleus (Behrens et al., 2002; Schofield et al., 2011; Mellott et al., 2011). The latter comprise about 25 % of all cholinergic axons in the cochlear nucleus of rodents. Although strictly speaking many of these circuits still provide a form of sound driven cholinergic feedback, it is fair to assume that it is not as closely stimulus-driven as the more direct local cholinergic feedback generated by the olivo-cochlear loop in the auditory brainstem. Rather, more advanced features of the perceived sound or listening situation, like the novelty of the perceived sound or the “meaning” of the perceived sound for the organism is thought to drive these long cholinergic top-down projections. Again, an increase in cholinergic transmission onto spherical bushy cells would change the excitability of these neurons, likely regardless of the origin of the cholinergic axons. That means that through different effector neurons (either in the tegmentum or the olivo-cochlear system) cortical neurons can strongly influence how neuronal information processing occurs in the inner ear and early stages of the auditory pathway, as can proximal properties of the sound that trigger the olivo-cochlear reflex. Specific behavioral situations like exploration or specific sensory contexts of sound stimuli like in fear conditioning thus potentially can change information processing in the very early stages of the auditory pathway in a quite dramatic way. This is a function classically seen as reserved to occur in the cortex itself.
Conclusion
Up to now we can only speculate on the exact causes and effects of cholinergic top-down control on the auditory brainstem nuclei and why it is useful, from an information processing point of view, to alter the flow of information already so close to the sensory periphery. Nevertheless, this example from the auditory pathway makes it clear that no part of the sensory brain “passively” receives information. Rather, all stages of sensory processing actively influence each other and dynamically modulate the ascending flow of information in the sensory pathway depending on external and internal conditions, under which the sensory perception occurs. It will be interesting to compare the principles of top-down and feedback control in the auditory brainstem with other sensory systems, where functionally similar descending projections have been described, for example in the olfactory and somatosensory systems. What are the common coding principles and computational demands that supported the evolution of these prominent descending pathways? Furthermore, well known feedback connections like the cortico-geniculate projection in the visual system, which at first glance may appear quite dissimilar to the brainstem circuits described here, could also provide worthwhile comparative insights into generalized information processing principles in the sensory brain.
Über die Autoren

Thomas Künzel studied biology at the Ruhr University in Bochum. He received his doctoral degree from the RWTH Aachen University with a dissertation on the development of auditory brainstem neurons under the supervision of Prof. Harald Luksch. He then briefly took a position as a research assistant at RWTH Aachen before continuing his postdoctoral training at Erasmus Medical Center in Rotterdam with Prof. Marcel van der Heijden and Prof. Gerard Borst. In 2011, he returned to the RWTH Aachen as an assistant to Prof. Hermann Wagner and has headed an independent group, which is supported by the German science foundation DFG (PP1608 “Ultrafast and temporally precise information processing: Normal and dysfunctional hearing”) since 2012. His research focus is on development and cellular physiology of phase-coding neurons in avian and mammalian auditory brainstem circuits.

Hermann Wagner studied biology in Ulm, Erlangen and Tübingen. After his diploma and doctoral thesis with Werner Reichardt at the Max-Planck-Institute of Biological Cybernetics, he was a postdoc with Mark Konishi at the California Institute of Technology, Pasadena, USA. He returned to the Max-Planck Institute to work with Kuno Kirschfeld, received a Heisenberg stipend to work with Geoff Manley at the TU Munich, before he took the chair of Zoology/Animal Physiology at the RWTH Aachen. His interests are in mechanisms of orientation, ranging from ecology via neuro-ethology to system physiology and cognition.
Acknowledgments:
We thank Dr. David Goyer and Charlene Gillét for discussion and helpful comments on the manuscript.
Referenes
Andeol, G., Guillaume, A., Micheyl, C., Savel, S., Pellieux, L., Moulin, A. (2011). Auditory Efferents Facilitate Sound Localization in Noise in Humans. J. Neurosci. 31, 6759–6763. 10.1523/JNEUROSCI.0248-11.2011Search in Google Scholar
Baashar, A., Robertson, D., Mulders, W.H.A.M. (2015). A novel method for selectively labelling olivocochlear collaterals in the rat. Hear. Res. 325, 35–41. 10.1016/j.heares.2015.02.011Search in Google Scholar
Behrens, E.G., Schofield, B.R., Thompson, A.M. (2002). Aminergic projections to cochlear nucleus via descending auditory pathways. Brain Res. 955, 34–44. 10.1016/S0006-8993(02)03351-6Search in Google Scholar
Benson, T.E., Berglund, A.M., Brown, M.C. (1996). Synaptic input to cochlear nucleus dendrites that receive medial olivocochlear synapses. J. Comp. Neurol. 365, 27–41. 10.1002/(SICI)1096-9861(19960129)365:1<27::AID-CNE3>3.0.CO;2-LSearch in Google Scholar
Benson, T.E., Brown, M.C. (1990). Synapses formed by olivocochlear axon branches in the mouse cochlear nucleus. J. Comp. Neurol. 295, 52–70. 10.1002/cne.902950106Search in Google Scholar
Bledsoe, S.C., Koehler, S., Tucci, D.L., Zhou, J., Prell, C. Le., Shore, S.E. (2009). Ventral cochlear nucleus responses to contralateral sound are mediated by commissural and olivocochlear pathways. J. Neurophysiol. 102, 886–900. 10.1152/jn.91003.2008Search in Google Scholar
Boothalingam, S., Macpherson, E.A., Allan, C., Allen, P., Purcell, D. (2016). Localization-in-noise and binaural medial olivocochlear functioning in children and young adults. J. Acoust. Soc. Am. 139, 247–262. 10.1121/1.4939708Search in Google Scholar
Briggs, F., Usrey, W.M. (2011). Corticogeniculate feedback and visual processing in the primate. J. Physiol. 589, 33–40. 10.1113/jphysiol.2010.193599Search in Google Scholar
Brown, M.C. (1993). Fiber pathways and branching patterns of biocytin-labeled olivocochlear neurons in the mouse brainstem. J. Comp. Neurol. 337, 600–613. 10.1002/cne.903370406Search in Google Scholar
Brown, M.C. (2011). Anatomy of Olivocochlear Neurons In: D.K. Ryugo, R.R. Fay, A.N. Popper, eds. Auditory and Vestibular Efferents Springer Handbook of Auditory Research. New York, NY: Springer, p. 17–37.10.1007/978-1-4419-7070-1_2Search in Google Scholar
Brown, M.C., Liberman, M.C., Benson, T.E., Ryugo, D.K. (1988). Brainstem branches from olivocochlear axons in cats and rodents. J. Comp. Neurol. 278, 591–603. 10.1002/cne.902780410Search in Google Scholar PubMed
Brown, M.C., Mukerji, S., Drottar, M., Windsor, A.M., Lee, D.J. (2013). Identification of inputs to olivocochlear neurons using transneuronal labeling with pseudorabies virus (PRV). JARO – J. Assoc. Res. Otolaryngol. 14, 703–717. 10.1007/s10162-013-0400-5Search in Google Scholar PubMed PubMed Central
Brown, M.C., Venecia, R.K. De, Guinan, J.J. (2003). Responses of medial olivocochlear neurons: Specifying the central pathways of the medial olivocochlear reflex. Exp. Brain Res. 153, 491–498. 10.1007/s00221-003-1679-ySearch in Google Scholar PubMed
Campagnola, L., Manis, P.B. (2014). A map of functional synaptic connectivity in the mouse anteroventral cochlear nucleus. J. Neurosci. 34, 2214–30. 10.1523/JNEUROSCI.4669-13.2014Search in Google Scholar PubMed PubMed Central
Cooper, N.P., Guinan, J.J. (2003). Separate mechanical processes underlie fast and slow effects of medial olivocochlear efferent activity. J. Physiol. 548, 307–12. 10.1113/jphysiol.2003.039081Search in Google Scholar PubMed PubMed Central
D’Souza, R.D., Vijayaraghavan, S. (2014). Paying attention to smell: cholinergic signaling in the olfactory bulb. Front. Synaptic Neurosci. 6, 21. Search in Google Scholar
Darrow, K.N., Maison, S.F., Liberman, M.C. (2006). Cochlear efferent feedback balances interaural sensitivity. Nat. Neurosci. 9, 1474–1476. 10.1038/nn1807Search in Google Scholar PubMed PubMed Central
Elgoyhen, A.B., Vetter, D.E., Katz, E., Rothlin, C.V., Heinemann, S.F., Boulter, J. (2001). Alpha10: a Determinant of Nicotinic Cholinergic Receptor Function in Mammalian Vestibular and Cochlear Mechanosensory Hair Cells. Proc. Natl. Acad. Sci. U.S.A. 98, 3501–3506. 10.1073/pnas.051622798Search in Google Scholar PubMed PubMed Central
Elgoyhen, A.B., Katz, E. (2012). The efferent medial olivocochlear-hair cell synapse. J. Physiol. Paris 106, 47–56. 10.1016/j.jphysparis.2011.06.001Search in Google Scholar PubMed PubMed Central
Engel, A.K., Fries, P., Singer, W. (2001). Dynamic Predictions: Oscillations and Synchrony in Top–Down Processing. 2, 704–716. 10.1038/35094565Search in Google Scholar PubMed
Felmy, F., Künzel, T. (2014). Giant synapses in the central auditory system. e-Neuroforum 5, 53–59. 10.1007/s13295-014-0060-xSearch in Google Scholar
Francis, N.A., Guinan, J.J. (2010). Acoustic stimulation of human medial olivocochlear efferents reduces stimulus-frequency and click-evoked otoacoustic emission delays: Implications for cochlear filter bandwidths. Hear. Res. 267, 36–45. 10.1016/j.heares.2010.04.009Search in Google Scholar PubMed PubMed Central
Fujino, K., Oertel, D. (2001). Cholinergic modulation of stellate cells in the mammalian ventral cochlear nucleus. J. Neurosci. 21, 7372–7383. 10.1523/JNEUROSCI.21-18-07372.2001Search in Google Scholar
Gilbert, C.D., Li, W. (2013). Top down influences on visual processing. Nat. Rev. Neurosci. 14, 350–363. 10.1038/nrn3476Search in Google Scholar PubMed PubMed Central
Goldberg JM, Brown PB (1969). Response of binaural neurons of dog superior olivary complex to dichotic tonal stimuli: some physiological mechanisms of sound localization. J. Neurophysiol. 32:613–636 10.1152/jn.1969.32.4.613Search in Google Scholar PubMed
Goyer, D., Kurth, S., Gillet, C., Keine, C., Rübsamen, R., Kuenzel, T. (2016). Slow Cholinergic Modulation of Spike Probability in Ultra-Fast Time-Coding Sensory Neurons. eNeuro 3. 10.1523/ENEURO.0186-16.2016Search in Google Scholar PubMed PubMed Central
Groff, J.A., Liberman, M.C. (2003). Modulation of cochlear afferent response by the lateral olivocochlear system: activation via electrical stimulation of the inferior colliculus. J. Neurophysiol. 90, 3178–3200. 10.1152/jn.00537.2003Search in Google Scholar PubMed
Grothe, B., Pecka, M. (2014). The natural history of sound localization in mammals – a story of neuronal inhibition. Front. Neural Circuits 8, 1–19. 10.3389/fncir.2014.00116Search in Google Scholar PubMed PubMed Central
Grothe, B., Pecka, M., McAlpine, D. (2010). Mechanisms of sound localization in mammals. Physiol. Rev. 90, 983–1012. 10.1152/physrev.00026.2009Search in Google Scholar PubMed
Guinan, J.J. (2006). Olivocochlear efferents: anatomy, physiology, function, and the measurement of efferent effects in humans. Ear Hear. 27, 589–607. 10.1097/01.aud.0000240507.83072.e7Search in Google Scholar PubMed
He, D.Z.Z., Jia, S., Dallos, P. (2003). Prestin and the dynamic stiffness of cochlear outer hair cells. J. Neurosci. 23, 9089–96. 10.1523/JNEUROSCI.23-27-09089.2003Search in Google Scholar
Horváth, M., Kraus, K.S., Illing, R.B. (2000). Olivocochlear neurons sending axon collaterals into the ventral cochlear nucleus of the rat. J. Comp. Neurol. 422, 95–105. 10.1002/(SICI)1096-9861(20000619)422:1<95::AID-CNE6>3.0.CO;2-#Search in Google Scholar
Kawase, T., Delgutte, B., Liberman, M.C. (1993). Antimasking effects of the olivocochlear reflex. II. Enhancement of auditory-nerve response to masked tones. J. Neurophysiol. 70, 2533–2549. 10.1152/jn.1993.70.6.2533Search in Google Scholar
Kawase, T., Liberman, M.C. (1993). Antimasking effects of the olivocochlear reflex. I. Enhancement of compound action potentials to masked tones. J Neurophysiol 70, 2519–2532. 10.1152/jn.1993.70.6.2519Search in Google Scholar
Kirk, E.C., Smith, D.W. (2003). Protection from Acoustic Trauma Is Not a Primary Function of the Medial Olivocochlear Efferent System. JARO – J. Assoc. Res. Otolaryngol. 4, 445–465. 10.1007/s10162-002-3013-ySearch in Google Scholar
Knudson, I.M., Shera, C.A., Melcher, J.R. (2014). Increased Contralateral Suppression of Otoacoustic Emissions Indicates a Hyper-responsive Medial Olivocochlear System in Humans with Tinnitus and Hyperacusis. J. Neurophysiol. 112, 3107–3208. 10.1152/jn.00576.2014Search in Google Scholar
Liberman, M.C., Liberman, L.D., Maison, S.F. (2014). Efferent Feedback Slows Cochlear Aging. J. Neurosci. 34, 4599–4607. 10.1523/JNEUROSCI.4923-13.2014Search in Google Scholar
Llorca-Torralba, M., Borges, G., Neto, F., Mico, J.A., Berrocoso, E. (2016). Noradrenergic Locus Coeruleus pathways in pain modulation. Neuroscience 338, 93–113. 10.1016/j.neuroscience.2016.05.057Search in Google Scholar
Mellott, J.G., Motts, S.D., Schofield, B.R. (2011). Multiple origins of cholinergic innervation of the cochlear nucleus. Neuroscience 180, 138–147. 10.1016/j.neuroscience.2011.02.010Search in Google Scholar
Mulders, W.H.A.M., Paolini, A.G., Needham, K., Robertson, D. (2003). Olivocochlear collaterals evoke excitatory effects in onset neurones of the rat cochlear nucleus. Hear. Res. 176, 113–121. 10.1016/S0378-5955(02)00750-5Search in Google Scholar
Mulders, W.H.A.M., Paolini, A.G., Needham, K., Robertson, D. (2009). Synaptic responses in cochlear nucleus neurons evoked by activation of the olivocochlear system. Hear. Res. 256, 85–92. 10.1016/j.heares.2009.07.003Search in Google Scholar
Mulders, W.H.A.M., Seluakumaran, K., Robertson, D. (2008). Effects of centrifugal pathways on responses of cochlear nucleus neurons to signals in noise. Eur. J. Neurosci. 27, 702–714. 10.1111/j.1460-9568.2008.06046.xSearch in Google Scholar
Mulders, W.H.A.M., Winter, I.M., Robertson, D. (2002). Dual action of olivocochlear collaterals in the guinea pig cochlear nucleus. Hear. Res. 174, 264–280. 10.1016/S0378-5955(02)00701-3Search in Google Scholar
Oertel, D., Wright, S., Cao, X.-J., Ferragamo, M., Bal, R. (2011). The multiple functions of T stellate/multipolar/chopper cells in the ventral cochlear nucleus. Hear. Res. 276, 61–69. 10.1016/j.heares.2010.10.018Search in Google Scholar
Pollak, G.D., Burger, R.M., Klug, A. (2003). Dissecting the circuitry of the auditory system. Trends Neurosci. 26, 33–39. 10.1016/S0166-2236(02)00009-7Search in Google Scholar
Radtke-Schuller, S., Seeler, S., Grothe, B. (2015). Restricted loss of olivocochlear but not vestibular efferent neurons in the senescent gerbil (Meriones unguiculatus). Front. Aging Neurosci. 7, 1–12. 10.3389/fnagi.2015.00004Search in Google Scholar
Scharf, B., Magnan, J., Chays, A. (1997). On the role of the olivocochlear bundle in hearing: 16 case studies. Hear. Res. 103, 101–122. 10.1016/S0378-5955(96)00168-2Search in Google Scholar
Scharf, B., Magnan, J., Collet, L., Ulmer, E., Chays, A. (1994). On the role of the olivocochlear bundle in hearing: a case study. Hear. Res. 75, 11–26. 10.1016/0378-5955(94)90051-5Search in Google Scholar
Schofield, B.R., Motts, S.D., Mellott, J.G. (2011). Cholinergic cells of the pontomesencephalic tegmentum: Connections with auditory structures from cochlear nucleus to cortex. Hear. Res. 279, 85–95. 10.1016/j.heares.2010.12.019Search in Google Scholar PubMed PubMed Central
Seluakumaran, K., Mulders, W.H.A.M., Robertson, D. (2008). Unmasking effects of olivocochlear efferent activation on responses of inferior colliculus neurons. Hear. Res. 243, 35–46. 10.1016/j.heares.2008.05.004Search in Google Scholar PubMed
Wersinger, E., Fuchs, P.A. (2011). Modulation of hair cell efferents. Hear. Res. 279, 1–12. 10.1016/j.heares.2010.12.018Search in Google Scholar PubMed PubMed Central
© 2017 by De Gruyter
Articles in the same Issue
- Frontmatter
- Editorial
- Mehr als nur ein neuer Look
- Übersichtsartikel
- Unser hungriges Gehirn: Welche Rolle spielen Gliazellen bei der Energieversorgung?
- Our hungry brain: Which role do glial cells play for the energy supply?
- Untersuchung und Modulation kortikaler Inhibition mittels transkranieller Magnetstimulation
- Assessment and modulation of cortical inhibition using transcranial magnetic stimulation
- Mechanismen des Neuritischen Prunings
- Mechanisms of Neurite Pruning
- Funktionen der GABAergen Übertragung im unreifen Gehirn
- Functions of GABAergic transmission in the immature brain
- Cholinerge Rückkopplungen auf den auditorischen Hirnstamm
- Cholinergic top-down influences on the auditory brainstem
- Forschungsförderung
- DFG Schwerpunktprogramm (SPP) 1926 “Next Generation Optogenetics – Tools and Application”
- Rezension
- Joachim Müller-Jung: Hirnforschung 8: Sprache und Kommunikation (F.A.Z.-Hörbuch)
- Ich glaub, mich trifft der Schlag: Warum das Gehirn tut, was es tun soll, oder manchmal auch nicht
- Nachruf
- Peter H. Seeburg (21.8.1944–22.8.2016)
- Nachrichten
- Nachrichten
Articles in the same Issue
- Frontmatter
- Editorial
- Mehr als nur ein neuer Look
- Übersichtsartikel
- Unser hungriges Gehirn: Welche Rolle spielen Gliazellen bei der Energieversorgung?
- Our hungry brain: Which role do glial cells play for the energy supply?
- Untersuchung und Modulation kortikaler Inhibition mittels transkranieller Magnetstimulation
- Assessment and modulation of cortical inhibition using transcranial magnetic stimulation
- Mechanismen des Neuritischen Prunings
- Mechanisms of Neurite Pruning
- Funktionen der GABAergen Übertragung im unreifen Gehirn
- Functions of GABAergic transmission in the immature brain
- Cholinerge Rückkopplungen auf den auditorischen Hirnstamm
- Cholinergic top-down influences on the auditory brainstem
- Forschungsförderung
- DFG Schwerpunktprogramm (SPP) 1926 “Next Generation Optogenetics – Tools and Application”
- Rezension
- Joachim Müller-Jung: Hirnforschung 8: Sprache und Kommunikation (F.A.Z.-Hörbuch)
- Ich glaub, mich trifft der Schlag: Warum das Gehirn tut, was es tun soll, oder manchmal auch nicht
- Nachruf
- Peter H. Seeburg (21.8.1944–22.8.2016)
- Nachrichten
- Nachrichten