Bacteria-released algal growth and morphogenesis factors regenerate axenic calli derived from the macroalga Ulva (Chlorophyta) and change the fatty acid profile
-
Hermann Holbl
Hermann Holbl is a PhD candidate in chemistry and a member of the Chemical Ecology of Cross Kingdom Interactions research group at the Friedrich Schiller University of Jena in Germany. He holds a bachelor’s degree in chemistry and a master’s degree in “Chemistry – Energy – Environment”. He studies the symbiotic relationships between
Ulva and its associated bacteria,Roseovarius sp. andMaribacter sp., concentrating on the algal growth- and morphogenesis-promoting factor thallusin synthesised byMaribacter sp.Nico Dunger is presently a trainee educator in biology and chemistry. While pursuing his education, he worked as a research assistant in the Chemical Ecology of Cross Kingdom Interactions research group at Friedrich Schiller University of Jena, Germany. He mainly examined the fatty acid composition of several morphotypes of
Ulva prior to commencing his high school teaching career.Thomas Wichard is a research group leader and lecturer in Analytical Chemistry at the Institute for Inorganic and Analytical Chemistry of the Friedrich Schiller University Jena (Germany). After being awarded a PhD in Biochemistry (Max Planck Institute for Chemical Ecology), he investigated the metal recruitment of nitrogen fixers at the Princeton Environmental Institute (USA). His team applies various analytical chemistry, chemical ecology, and molecular biology methodologies to understand the basis of eco-physiological processes in bacteria-macroalgae interactions (cross-kingdom interactions).



Abstract
Ulva, a green marine macroalga, lives in a symbiotic relationship with bacteria like Roseovarius sp. and Maribacter sp., which produce algal growth- and morphogenesis-promoting factors (AGMPFs) crucial for algal development. In the absence of all AGMPFs including thallusin Ulva exhibit a callus-like morphotype characterised by a lack of cellular differentiation. This study introduces calli as a novel model to investigate the AGMPF action. The metabolic impacts of AGMPFs were investigated by comparing the axenic calli of Ulva compressa (cultivar Ulva mutabilis) with those exposed to AGMPFs. Adding AGMPFs regenerated the callus and promoted germling-like structures, along with changes in the metabolome. The bouquet of AGMPFs, containing (−)-thallusin, induced significant variations in fatty acid metabolism, notably an increase of palmitic acid and several polyunsaturated fatty acids, along with a change of the ω6:ω3 ratio (<0.7), which may directly affect growth as well as membrane and cell wall formation. The results highlight the significant implications of bacterial-derived AGMPFs instead of live bacteria and underscore the necessity of optimising AGMPF distribution in cultivation to improve the nutritional quality of Ulva for food and feed.
The marine green seaweed genus Ulva (Chlorophyta) has drawn the attention of numerous research groups worldwide, having become a model system for studying the morphogenesis, chemical ecology and phylogeny of the Chlorophyta lineage (Blomme et al. 2023; Steinhagen et al. 2019; Wichard 2015, 2023). Ulva appears ubiquitously in coastal benthic regions; it copes well with stressors like changes in salinity, temperature, irradiance and polluted water (Ghaderiardakani et al. 2020). Together with its high nutrient uptake rate, these properties contribute to the appearance of Ulva blooms, also known as ‘green tides’ (Zhang et al. 2019). Therefore, many concepts for applying Ulva spp. were established over the past few decades for the bioremediation of wastewater polluted with heavy metals, nutrients or micropollutants (Hardegen et al. 2023; Rahhou et al. 2023; Shpigel et al. 1993). Furthermore, Ulva spp. can be cultivated as an alternative to land-based crops for food and animal feed (Khan and Mamat 2024). The biorefinery of, for example, the polysaccharide ulvan, protein sources, lipids and alcohols of low molecular weight open up further applications for Ulva in aquaculture (Bikker et al. 2016; Calheiros et al. 2021; Simon et al. 2022). Our study aimed to understand how bacteria-released algal growth and morphogenesis factors (AGMPF) promote callus regeneration and influence the metabolome and fatty acid profile of axenic calli derived from the model organism Ulva mutabilis Føyn, which was grown under standardised conditions in an artificial Ulva culture medium (UCM, Califano and Wichard 2018) (Figure 1).
![Figure 1:
Experimental workflow. Gametes of Ulva mutabilis were gathered and separated from bacteria (Califano and Wichard 2018). These axenic gametes were cultivated for six weeks to a callus stage. Afterwards, the stock cultures of calli were unified, mixed and distributed in six culture flasks under sterile conditions. One biological triplicate served as control (in 100 % UCM), while the cultures for the AGMPF-treated biological triplicate [in sterile-filtered Roseovarius UCM-based supernatant (75 %) and UCM (25 %); v:v] were inoculated subsequently with (−)-thallusin (2 × 10−8 mol l−1 final concentration). The calli were incubated for two weeks, separated from the culture medium for solvent extraction, and flash-frozen. After further sample preparation, the ground calli were extracted using a commercial kit based on the Folch lipid extraction method (Folch et al. 1957). In the ensuing sample processing, one aliquot of each sample extract was subjected to a UHPLC-MS Orbitrap system to conduct endo-metabolomics. The data were analysed and evaluated using Compound Discoverer (vers. 3.3) and Metaboanalyst (vers. 6.0). Fatty acids were derivatised to obtain methyl esters for quantification by GC-EI-MS. The analysed fatty acids were quantified based on their response factor through a comparison with an external standard mix. For detailed information, see Supplementary Material.](/document/doi/10.1515/bot-2024-0101/asset/graphic/j_bot-2024-0101_fig_001.jpg)
Experimental workflow. Gametes of Ulva mutabilis were gathered and separated from bacteria (Califano and Wichard 2018). These axenic gametes were cultivated for six weeks to a callus stage. Afterwards, the stock cultures of calli were unified, mixed and distributed in six culture flasks under sterile conditions. One biological triplicate served as control (in 100 % UCM), while the cultures for the AGMPF-treated biological triplicate [in sterile-filtered Roseovarius UCM-based supernatant (75 %) and UCM (25 %); v:v] were inoculated subsequently with (−)-thallusin (2 × 10−8 mol l−1 final concentration). The calli were incubated for two weeks, separated from the culture medium for solvent extraction, and flash-frozen. After further sample preparation, the ground calli were extracted using a commercial kit based on the Folch lipid extraction method (Folch et al. 1957). In the ensuing sample processing, one aliquot of each sample extract was subjected to a UHPLC-MS Orbitrap system to conduct endo-metabolomics. The data were analysed and evaluated using Compound Discoverer (vers. 3.3) and Metaboanalyst (vers. 6.0). Fatty acids were derivatised to obtain methyl esters for quantification by GC-EI-MS. The analysed fatty acids were quantified based on their response factor through a comparison with an external standard mix. For detailed information, see Supplementary Material.
Ulva mutabilis Føyn – described as conspecific with Ulva compressa by Steinhagen et al. (2019) – maintains a tripartite symbiotic relationship with Roseovarius sp. strain MS2 and Maribacter sp. strain MS6 in developing into a regular differentiated macroalga (Spoerner et al. 2012; Steinhagen et al. 2019; Wichard 2023). The enhanced growth effect induced by Roseovarius sp. and cell differentiation mediated by Maribacter sp. are triggered by AGMPFs, which are released into the culture medium, including the morphogen thallusin (Alsufyani et al. 2020; Ghaderiardakani et al. 2019; Matsuo et al. 2003). Therefore, filtered bacterial supernatant can be used as a replacement for the bacterial effect (Spoerner et al. 2012).
It is not just U. mutabilis that benefits from the interaction with these specific bacteria; the growth and development of other Ulva species are also promoted within the same phycosphere (Ghaderiardakani et al. 2017). Therefore, the studied effects from bacterial interactions with Ulva are likely generalizable to the genus. Without the symbiotic bacteria, certain substitute species, or the corresponding AGMPFs within the chemosphere, the alga develops into a callus of unorganised cell masses (Wichard 2023). Undifferentiated callus-like morphotypes have also been investigated extensively in flowering plants, such as soybeans, and are interesting ‘seeds’ for aquaculture (Ram Kumar et al. 2021).
Given that AGMPFs shape Ulva significantly, a change in metabolism occurs accordingly (Califano 2022). A recent study on the impact of individual bacterial partners on the axenic microalgae Isochrysis galbana reinforced these observations (Wu et al. 2023). Co-cultivation had effects on several parameters connected to primary metabolism (i.e., the growth, chlorophyll content, soluble protein content, or fatty acid profile of the microalgae) as well as in flowering plants (He et al. 2009; Ram Kumar et al. 2021).
To investigate the hypothesis regarding changes in the metabolome of Ulva influenced by the presence or absence of AGMPFs, (i) axenic U. mutabilis calli (strain FSU-UM5-1) were inoculated with AGMPFs and compared to (ii) an untreated control group of axenic calli, as well as (iii) an adult stationary Ulva culture containing its natural microbiome (Figure 1).
The complete development and morphogenesis occur only in the presence of all AGMPFs released by Roseovarius sp. and Maribacter sp. When only the Roseovarius factor or the Maribacter factor (i.e., thallusin) is present, morphogenesis remains incomplete, and the resulting developmental differences influence the algal metabolome. Therefore, we administered both AGMPFs together to avoid secondary effects due to different phenological stages.
A six-week-old axenic callus culture covered a colonisation area of 15 ± 2.5 cm2 on the 75 cm2 bottom surface of the tissue culture flask in 50 ml UCM. To induce growth and morphogenesis of the calli, the sterile-filtered AGMPF cocktail was supplemented with the Roseovarius factor (supernatant from a Roseovarius sp. culture grown on glycerol in UCM) and the Maribacter factor, (−)-thallusin, at a concentration of 2 × 10−8 mol l−1. This concentration was 4,000 times higher than the half-effective concentration of thallusin (EC50 = 5 × 10−12 mol l−1; Dhiman et al. 2022; Ulrich et al. 2022) and remained effective throughout the two-week cultivation period. The cultures underwent microscopic assessment before harvesting and extracting the endo-metabolome on day 14 after AGMPF inoculation (Figure 2 and Supplementary Material for details).
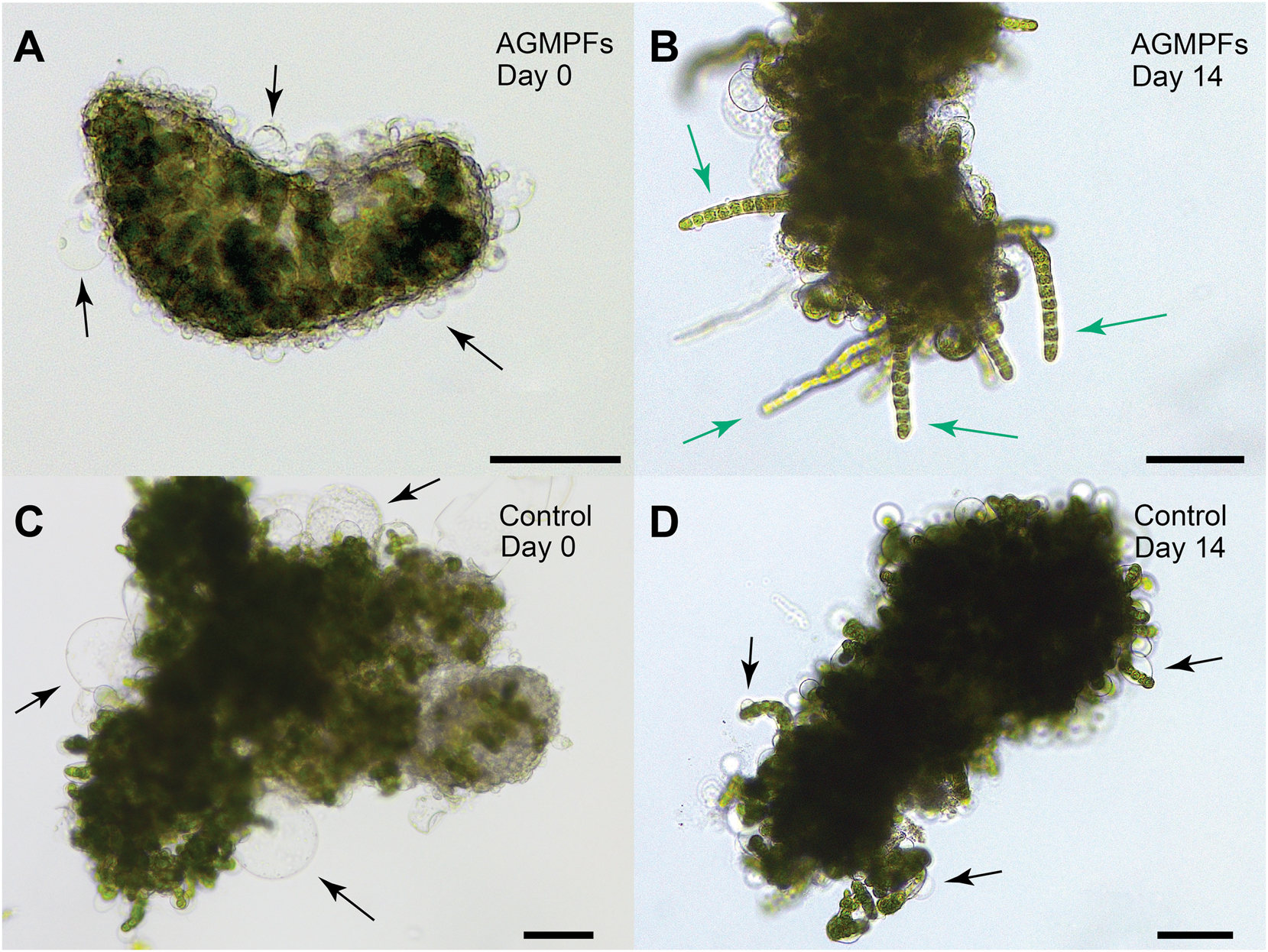
Bioassays with 6-week-old axenic calli of Ulva mutabilis and application of algal growth and morphogenesis-promoting factors (AGMPFs). Exemplary microscopic pictures of the examined treatments show (Α) a callus with cell wall protrusions (arrows) on day 0 in the group with added AGMPFs and displaying (B) a callus after 14 days with AGMPF-induced germling-like structures (green arrows) and no cell wall protrusions. The control group without AGMPFs shows (C, D) the typical cell wall protrusions (arrows) on day 0 and day 14. Scale bars = 100 µm.
Throughout the studied period, the calli treated with AGMPFs exhibited partial recovery. They developed new tissue characterised by the absence of cell wall protrusions and a general decline in the size and number of cell protrusions (compare Figure 2A and B). Overall, AGMPFs containing (−)-thallusin in excess induced callus transformation into a growing tissue (Figure 2B). This contrasted with the non-treated control group, where large protrusions persisted (Figure 2C and D). The well-developed branches (germling-like structures) can be separated from the callus clump and cultured individually for proper development in the presence of the AGMPFs.
To monitor changes in the metabolome upon application of the AGMPF cocktail, the calli were extracted using a mixture of chloroform, methanol and water (Folch et al. 1957). After extraction, the samples were analysed using ultra-high-pressure liquid chromatography coupled with a high-resolution electrospray ionisation mass spectrometer (UHPLC-HR-ESI-MS). The data was processed using the Compound Discoverer (vers. 3.3) and visualised using Metaboanalyst (vers. 6.0) (see Figure 1 and Supplementary Material for details).
Principal component analysis (PCA) showed the similarity between the two sample groups when PC1 was plotted against PC2 (Figure 3A). However, PC3, which accounted for 15 % of the variance, was indicated in a divergence between the control and the AGMPF-treated groups. The volcano plot (Figure 3B, Table S1) demonstrated the significant alterations in metabolite composition resulting from the addition of AGMPFs. Significantly, 88 upregulated and 42 down-regulated m/z features (i.e., potential compounds) were determined in the group treated with AGMPFs, while 3,103 m/z features did not vary between the groups. The up-regulation of palmitic acid (C16:0, indicated by the green circle in Figure 3B) piqued our interest because it is the most abundant fatty acid in Ulva spp. (García-Poza et al. 2022; Kostetsky et al. 2018) and implied further alterations in lipid metabolism, which might affect the membrane composition and development of Ulva due to the application of AGMPFs. It is noteworthy that plant growth regulators can influence plant lipid composition and lipid metabolism in terms of fatty acid induction and alteration of composition (Aly et al. 2008). In contrast to these plants, Ulva relies on an external supply of specifically associated and necessary bacteria for enhanced algal growth and development (Wichard 2023).
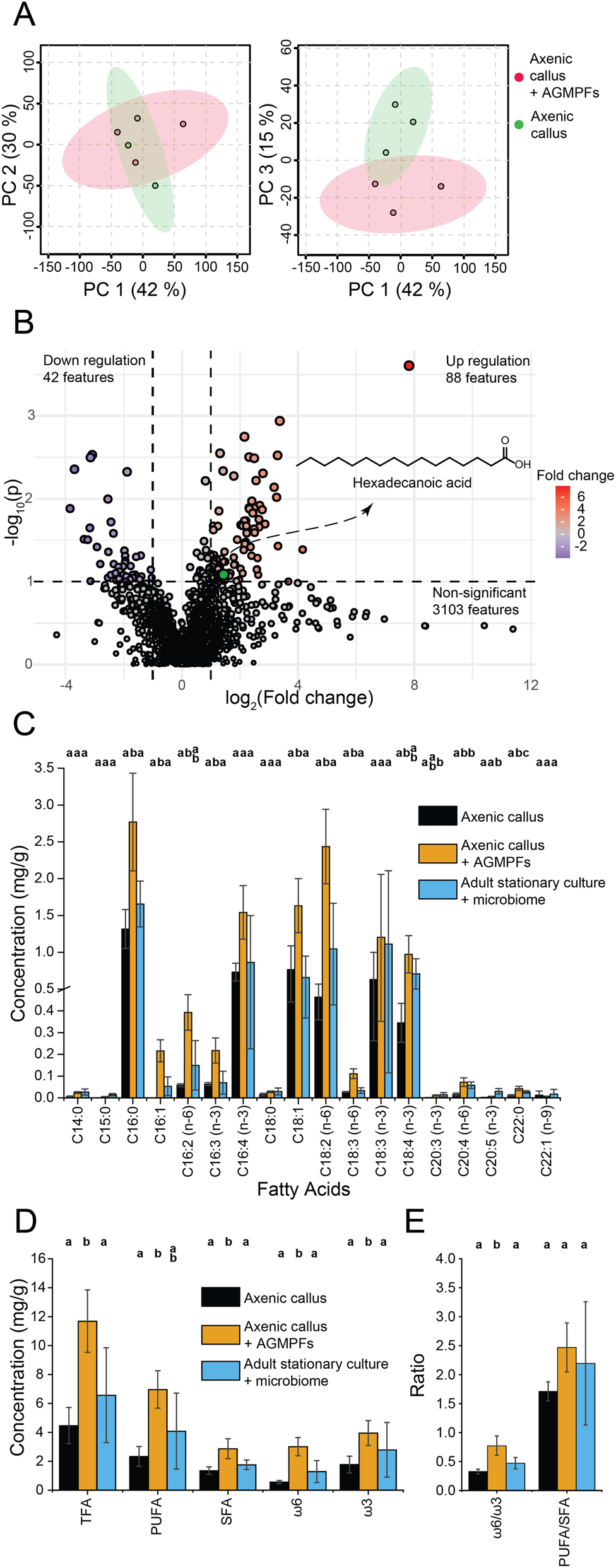
Metabolomics and fatty acid analysis. (A) Principal component analysis (PCA) scores plot from the endo-metabolomic LC-MS data comparison between the axenic calli of Ulva mutabilis treated with the AGMPF group (red) and the axenic calli control group (green). The left plot displays PC 2 versus PC 1, while the right plot shows PC 3 versus PC 1. (B) Volcano plot describing the 88 up- and 42 down-regulated m/z features in the AGMPF-treated samples versus the control samples (p-value threshold = 0.1, log2 fold change threshold = 1). The hexadecanoic acid (C16:0) is highlighted in green as the upregulated compound in samples treated with AGMPFs. (C) Bar chart illustrating the concentrations of individual fatty acids in mg per g algal dry weight and comparing the axenic calli control group (black bars) with axenic calli + AGMPFs (orange bars) and the adult algal samples (blue bars) in a stationary phase with the usual microbiome and normal tubular morphotype (‘slender’). (D) Bar chart of concentrations (mg per g algal dry weight) of different groups of fatty acids in the treatment and control groups. Total fatty acids (TFA), polyunsaturated fatty acids (PUFA), saturated fatty acids (SFA), ω6-FA(n-6), and ω3-FA(n-3) were compared. (E) Bar chart of ω6/ω3 and PUFA/SFA ratios corresponding to each treatment and control group. Error bars in (C), (D) and (E) correspond to the mean ± standard deviation (n = 3). Means denoted by a different letter indicate significant differences between treatments (one-way ANOVA, Fisher or Games-Howell post hoc test, depending on homoscedasticity, p < 0.05) (See also Tables S1–S3 in the Supplementary Material).
For targeted total fatty analyses, the free and bound fatty acids contained in the extract were converted into fatty acid methyl esters (FAME) using methanolysis through trans-esterification (Figure 1) (Alsufyani et al. 2014). In addition to the axenic calli, stationary adult cultures of U. mutabilis, complete with the entire native microbiome, were surveyed for the fatty acid profile. The areas of FAME peaks analysed using gas chromatography (Figure S1) coupled with electron impact ionisation mass spectrometry (GC-EI-MS) were compared to signals from external standards to estimate the concentration per sample dry weight (Figure 3C).
In stationary adult U. mutabilis cultures with their associated microbiomes, high levels of C18-PUFAs were detected including linoleic acid [C18:2(n-6), 15 %], α-linolenic acid [C18:3(n-3), 15 %], stearidonic acid [C18:4(n-3), 12 %] along with trace amounts of γ-linolenic acid, collectively accounting for 42 % of the total fatty acids (Table S2). C20-PUFAs such as arachidonic acid [C20:4(n-6)] and eicosapentaenoic acid [C20:5(n-3)] contributed with around 1 %, only a minor amount. Notably, the fatty acid profile of U. mutabilis is characterised by unsaturated C16 and C18 fatty acids (Figure S1), a typical feature of this genus, as previously reported in several studies (Alsufyani et al. 2014; McCauley et al. 2016; Monteiro et al. 2022; Pereira et al. 2012). Overall, our data aligned well with the range of concentrations previously measured in Ulva species; for example, those that originated from Morraceira Island (Portugal) or Brittany (France) and were cultivated under different conditions (García-Poza et al. 2022; Monteiro et al. 2022).
The fatty acid profile of the axenic callus of U. mutabilis was also predominantly rich in ω3 and C18 fatty acids besides palmitic acid (C16:0), which has already been determined using the endo-metabolomics approach (Figure 3B, Table S1). Our analysis, excluding bacterial fatty acid contamination, found all of the typical fatty acids – such as C16:0, C16:4, C18:1, C18:2, αC18:3, C18:4 and a few C20 fatty acids – in the axenic U. mutabilis cultures, both with and without AGMPFs (Table S1 and S2).
However, the application of AGMPFs containing thallusin enhanced fatty acid synthesis and changed their abundance. The targeted analysis revealed that 10 out of the 18 fatty acids, including 7 PUFAs, increased significantly in cultures supplemented with AGMPFs compared to the untreated control calli. Interestingly, additional AGMPFs even increased the production of fatty acids compared to the adult stationary Ulva culture and its associated microbiome. The excess of AGMPFs corresponded with elevated levels of seven fatty acids – C16:0, C16:1, C16:3(n-3), C18-1, C18-2(n-6), γC18:3(n-6) and C22:0 – whereas the concentrations of other fatty acids remained unchanged (Figure 3C, Table S3).
Distinct characteristics could be noted following the application of AGMPFs. For instance, the ratio of 18:1 to 18:2(n-6) was inverted in favour of linoleic acid, which also affected the ω6:ω3 ratio (Table S2). The alterations in the concentrations of polyunsaturated fatty acids (PUFA) and saturated fatty acids (SFA) (Figure 3D) influenced the ratios of ω6:ω3 and PUFA/SFA, nearly doubling the ω6:ω3 ratio (p < 0.05) in AGMPF-treated calli (Figure 3E). With ω6:ω3 = 0.77, the ratio under AGMPFs-treated conditions was higher than recently published values in Ulva (which, for example, ranged from 0.15 to 0.37; Monteiro et al. 2022), but was still an excellent ratio (ω6:ω3 < 1) for human nutrition (Simopoulos 2000).
In our study, the concentration of specific fatty acids [16:4(n-3)] normalised per mg of dry mass was over ten times higher than in some previous studies (e.g., García-Poza et al. 2022) (Figure 3C, Table S3). These variations in fatty acid concentrations can result from multiple factors. Species-specific metabolic traits play a crucial role as they inherently produce and regulate fatty acids in distinct ways. Abiotic conditions such as temperature and salinity, along with variations in nutrient composition, can significantly influence lipid metabolism (Gnayem et al. 2025). Our study underscores another critical factor: the role of AGMPFs released by the associated microbiome. These bacterial-derived compounds may actively regulate fatty acid biosynthesis in Ulva, modulating lipid composition, and this suggests that future studies should further explore how specific AGMPFs contribute to metabolic regulation.
In any case, comparing the fatty acid composition of differentiated tissue to that of callus tissue in flowering plants (e.g., Cucurbitaceae; Halder and Gadgil 1983) yielded comparable results to those in U. mutabilis. The total lipid content of callus cells significantly diminished from 20–30 % to 2–4 %, and the PUFA/SFA ratio fell in undifferentiated callus cells, indicating an even more pronounced alteration than in Ulva calli (Figure 3D and E). Comparable to our research approach, other investigations have revealed alterations in fatty acid content by adding several growth regulators to callus cells (Pandey and Gadgil 1984).
Our results demonstrated that bacteria-derived AGMPFs triggered molecular processes that served as switches for lipid or fatty acid biosynthesis. Thus, AGMPFs function as stimulants, enhancing fatty acid production in Ulva, a finding that has also been suggested for bacteria-microalgae interactions (Zhang et al. 2025). Changes in fatty acid profile are often correlated with stress parameters. Certainly, the lack of, or a change in, the microbiome can harm algal morphogenesis (Marshall et al. 2006; Spoerner et al. 2012). Therefore, it is tempting to assume that the different quantities and ratios of individual fatty acids might partially explain the callus morphology and its susceptibility to stress in U. mutabilis, considering that those fatty acids have diverse cell functions. These functions are vital membrane components for bilayer establishment, energy storage in the form of triglycerides, and cell signalling (De Carvalho and Caramujo 2018). Membrane fluidity is, for example, higher if more unsaturated fatty acids are employed than saturated fatty acids. Many organisms alter membrane properties based on fatty acid composition, including sphingo- and phospholipids, to cope with their environment (e.g., when confronted with temperature stress). Additional chemical analysis of Ulva calli is necessary to evaluate the appearance of malformed cell walls and protrusion formation in relation to changes in the fatty acid profile. Using Raman spectroscopy imaging, Schultz et al. (2024) have already demonstrated the change in chemical composition due to malformations and cell wall protrusions in axenic cultures.
Furthermore, the β-oxidation of fatty acids from triacylglycerols into fatty acyl CoA is a critical process for supporting the algae during growth. Lipids serve as storage matrices for fat-soluble vitamins, including A, D, and E. Therefore, cell functions can be impaired if essential vitamins are not available within the cell. Also, free PUFA and their derivatives, like oxylipins, often have allelopathic functions (De Carvalho and Caramujo 2018).
To sum up, an impeded lipid or fatty acid metabolism can harm the organism. Based on the change in fatty acid composition and the up- and down-regulation of many (unknown) compounds according to metabolomics, we consider that the AGMPF effect is certainly not limited to lipid metabolism only. Our results underline the importance of an optimal distribution of AGMPF during cultivation for the establishment of a healthy population of Ulva and the provision of high levels of unsaturated fatty acids in Ulva used for food and feed.
Overall, lipid and fatty acid quantities and profiles change with season, sampling location, and abiotic conditions (Gnayem et al. 2024, 2025), as well as with the availability of AGMPFs (Figure 3), which are directly related to the prevalence and functional traits of the macroalgal microbiome. Environmental changes may thus drive shifts in the microbiome or its functional traits, ultimately affecting, for example, the bioavailability of AGMPFs in the algal chemosphere and, subsequently, the fatty acid profile of Ulva. Therefore, measuring the quantities of AGMPFs is essential for effective monitoring of algal aquaculture. Thallusin, the AGMPF produced by Maribacter sp., is being monitored (Ulrich et al. 2022), whereas the AGMPF released by Roseovarius sp. has not yet been identified. Further research is needed to elucidate the regulatory mechanisms of these compounds and to assess their applicability to other systems, including terrestrial plants.
In summary, our study focused on the metabolic effects of AGMPFs, mainly through fatty acid profiles. Our key findings include:
Experimental design: The advantage of using bacterial-derived AGMPFs instead of live bacteria lies in their precise control and reduced risk of unintended bacteria-algae interactions, making them a powerful tool for targeted studies and applications without bacterial contaminants.
Morphological transformation: AGMPF treatment on axenic Ulva calli prompts regeneration, suggesting a reversal from a callus into a differentiated form. This transformation highlights the significance of bacterial interactions in the normal development of algal morphology and demonstrates that axenic calli can be reactivated.
Chemical analysis: Axenic calli facilitate the identification and quantification of metabolites without background noise from microbial contaminants.
Metabolomic shifts: Exposure to AGMPFs is associated with notable changes in fatty acid metabolism. Specifically, there is an increase in hexadecanoic acid and polyunsaturated fatty acids. These compounds play a crucial role in maintaining the structure and fluidity of the cell membrane, which are essential for cellular functionality and integrity in aquatic environments.
Implications for food and feed applications: Given that AGMPFs improve the morphological and metabolic profile of U. mutabilis, optimising AGMPF cultivation conditions could enhance the nutritional value of Ulva as a source of beneficial fatty acids.
About the authors

Hermann Holbl is a PhD candidate in chemistry and a member of the Chemical Ecology of Cross Kingdom Interactions research group at the Friedrich Schiller University of Jena in Germany. He holds a bachelor’s degree in chemistry and a master’s degree in “Chemistry – Energy – Environment”. He studies the symbiotic relationships between Ulva and its associated bacteria, Roseovarius sp. and Maribacter sp., concentrating on the algal growth- and morphogenesis-promoting factor thallusin synthesised by Maribacter sp.

Nico Dunger is presently a trainee educator in biology and chemistry. While pursuing his education, he worked as a research assistant in the Chemical Ecology of Cross Kingdom Interactions research group at Friedrich Schiller University of Jena, Germany. He mainly examined the fatty acid composition of several morphotypes of Ulva prior to commencing his high school teaching career.

Thomas Wichard is a research group leader and lecturer in Analytical Chemistry at the Institute for Inorganic and Analytical Chemistry of the Friedrich Schiller University Jena (Germany). After being awarded a PhD in Biochemistry (Max Planck Institute for Chemical Ecology), he investigated the metal recruitment of nitrogen fixers at the Princeton Environmental Institute (USA). His team applies various analytical chemistry, chemical ecology, and molecular biology methodologies to understand the basis of eco-physiological processes in bacteria-macroalgae interactions (cross-kingdom interactions).
Acknowledgments
The article is also based upon work from COST Action CA20106 “Tomorrow’s wheat of the sea’: Ulva, a model for an innovative mariculture”, supported by COST (European Cooperation in Science and Technology, www.cost.eu). We thank Prof. Dr. Hans-Dieter Arndt (Friedrich Schiller University Jena) and his coworkers, Dr. Seema Dhiman and Dr. Paul Wienecke, for synthesising and providing (−)-thallusin and their great support. We sincerely thank Johann F. Ulrich for his assistance in preparing the Roseovarius raw extract and Prof. Dr. Georg Pohnert (Friedrich Schiller University Jena) for his invaluable support throughout this study.
-
Research ethics: Not applicable.
-
Informed consent: Not applicable.
-
Author contributions: All authors have accepted responsibility for the entire content of this manuscript and approved its submission.
-
Use of Large Language Models, AI and Machine Learning Tools: Not applicable.
-
Conflict of interest: The authors declare that they have no conflict of interest regarding this article.
-
Research funding: The Deutsche Forschungsgemeinschaft (DFG, German Research Foundation) supported the study within the CRC 1127 ChemBioSys (project no. 239748522, HH, and TW).
-
Data availability: The endo-metabolomics data are available in the public database Zenodo: https://zenodo.org/records/14938653 (DOI 10.5281/zenodo.14938653).
References
Alsufyani, T., Califano, G., Deicke, M., Grueneberg, J., Weiss, A., Engelen, A.H., Kwantes, M., Mohr, J.F., Ulrich, J.F., and Wichard, T. (2020). Macroalgal–bacterial interactions: identification and role of thallusin in morphogenesis of the seaweed Ulva (Chlorophyta). J. Exp. Bot 71: 3340–3349, https://doi.org/10.1093/jxb/eraa066.Suche in Google Scholar PubMed PubMed Central
Alsufyani, T., Engelen, A.H., Diekmann, O.E., Kuegler, S., and Wichard, T. (2014). Prevalence and mechanism of polyunsaturated aldehydes production in the green tide forming macroalgal genus Ulva (Ulvales, Chlorophyta). Chem. Phys. Lipids 183: 100–109, https://doi.org/10.1016/j.chemphyslip.2014.05.008.Suche in Google Scholar PubMed
Aly, M.A.M., Amer, E.A., Al-Zayadneh, W.A., and Eldin, A.E.N. (2008). Growth regulators influence the fatty acid profiles of in vitro induced jojoba somatic embryos. Plant Cell Tiss. Org. Cult. 93: 107–114, https://doi.org/10.1007/s11240-008-9348-2.Suche in Google Scholar
Bikker, P., van Krimpen, M.M., van Wikselaar, P., Houweling-Tan, B., Scaccia, N., van Hal, J.W., Huijgen, W.J., Cone, J.W., and Lopez-Contreras, A.M. (2016). Biorefinery of the green seaweed Ulva lactuca to produce animal feed, chemicals and biofuels. J. Appl. Phycol. 28: 3511–3525, https://doi.org/10.1007/s10811-016-0842-3.Suche in Google Scholar PubMed PubMed Central
Blomme, J., Wichard, T., Jacobs, T.B., and De Clerck, O. (2023). Ulva: an emerging green seaweed model for systems biology. J. Phycol. 59: 433–440, https://doi.org/10.1111/jpy.13341.Suche in Google Scholar PubMed
Calheiros, A.C., Sales, L.P.M., Pereira Netto, A.D., Cavalcanti, D.N., Castelar, B., and Reis, R.P. (2021). Commercial raw materials from algaculture and natural stocks of Ulva spp. J. Appl. Phycol. 33: 1805–1818, https://doi.org/10.1007/s10811-021-02413-3.Suche in Google Scholar
Califano, G. (2022). Inter-kingdom interactions shapes macroalga Ulva holobiont: studies on algal growth and morphogenesis promoting factors (phytohormones and thallusin) and algal associated microbial community, PhD dissertation. Friedrich-Schiller-Universität Jena, Jena.Suche in Google Scholar
Califano, G. and Wichard, T. (2018). Preparation of axenic cultures in Ulva (Chlorophyta). In: Charrier, B., Wichard, T., and Reddy, C.R.K. (Eds.). Protocols for macroalgae research. C.R.K. CRC Press, Boca Raton, pp. 159–171.10.1201/b21460-9Suche in Google Scholar
De Carvalho, C. and Caramujo, M.J. (2018). The various roles of fatty acids. Molecules 23: 2583, https://doi.org/10.3390/molecules23102583.Suche in Google Scholar PubMed PubMed Central
Dhiman, S., Ulrich, J.F., Wienecke, P., Wichard, T., and Arndt, H.-D. (2022). Stereoselective total synthesis of (−)-thallusin for bioactivity profiling. Angew. Chem., Int. Ed. 61: e202206746, https://doi.org/10.1002/anie.202206746.Suche in Google Scholar PubMed PubMed Central
Folch, J., Lees, M., and Stanley, G.H.S. (1957). A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 226: 497–509, https://doi.org/10.1016/s0021-9258(18)64849-5.Suche in Google Scholar
García-Poza, S., Morais, T., Leandro, A., Cotas, J., Marques, J.C., Pereira, L., and Gonçalves, A.M.M. (2022). A Comparative study of the fatty acids and monosaccharides of wild and cultivated Ulva sp. J. Mar. Sci. Eng. 10: 233, https://doi.org/10.3390/jmse10020233.Suche in Google Scholar
Ghaderiardakani, F., Coates, J.C., and Wichard, T. (2017). Bacteria-induced morphogenesis of Ulva intestinalis and Ulva mutabilis (Chlorophyta): a contribution to the lottery theory. FEMS Microbiol. Ecol. 93: fix094, https://doi.org/10.1093/femsec/fix094.Suche in Google Scholar PubMed PubMed Central
Ghaderiardakani, F., Califano, G., Mohr, J.F., Abreu, M.H., Coates, J.C., and Wichard, T. (2019). Analysis of algal growth- and morphogenesis-promoting factors in an integrated multi-trophic aquaculture system for farming Ulva spp. Aquacult. Environ. Interact. 11: 375–391, https://doi.org/10.3354/aei00319.Suche in Google Scholar
Ghaderiardakani, F., Quartino, M.L., and Wichard, T. (2020). Microbiome-dependent adaptation of seaweeds under environmental stresses: a perspective. Front. Mar. Sci. 7: 575228, https://doi.org/10.3389/fmars.2020.575228.Suche in Google Scholar
Gnayem, N., Unis, R., Gnaim, R., Chemodanov, A., Israel, Á., Gnaim, J., and Golberg, A. (2024). Seasonal and culture period variations in the lipid and fatty acid content of Ulva lactuca cultivated in Mikhmoret onshore (Israel). Bot. Mar. 67: 101–114, https://doi.org/10.1515/bot-2023-0027.Suche in Google Scholar
Gnayem, N., Unis, R., Gnaim, R., Chemodanov, A., Israel, Á., Gnaim, J., and Golberg, A. (2025). Fatty Acid content and profile in Ulva lactuca in response to exposure to variable growth conditions in indoor photobioreactors. Life 15: 57, https://doi.org/10.3390/life15010057.Suche in Google Scholar PubMed PubMed Central
Halder, T. and Gadgil, V.N. (1983). Fatty acids of callus tissues of six species of cucurbitaceae. Phytochemistry 22: 1965–1967, https://doi.org/10.1016/0031-9422(83)80024-7.Suche in Google Scholar
Hardegen, J., Amend, G., and Wichard, T. (2023). Lifecycle-dependent toxicity and removal of micropollutants in algal cultures of the green seaweed Ulva (Chlorophyta). J. Appl. Phycol. 5: 2031–2048, https://doi.org/10.1007/s10811-023-02936-x.Suche in Google Scholar
He, Y., Guo, X., Lu, R., Niu, B., Pasapula, V., Hou, P., Cai, F., Xu, Y., and Chen, F. (2009). Changes in morphology and biochemical indices in browning callus derived from Jatropha curcas hypocotyls. Plant Cell 98: 11–17, https://doi.org/10.1007/s11240-009-9533-y.Suche in Google Scholar
Khan, N. and Mamat, R. (2024). Eco-friendly nutrient from ocean: exploring Ulva seaweed potential as a sustainable food source. J. Agric. Food Res. 17: 101239.10.1016/j.jafr.2024.101239Suche in Google Scholar
Kostetsky, E., Chopenko, N., Barkina, M., Velansky, P., and Sanina, N. (2018). Fatty acid composition and thermotropic behavior of glycolipids and other membrane lipids of Ulva lactuca (Chlorophyta) inhabiting different climatic zones. Mar. Drugs 16: 494, https://doi.org/10.3390/md16120494.Suche in Google Scholar PubMed PubMed Central
Marshall, K., Joint, I., Callow, M.E., and Callow, J.A. (2006). Effect of marine bacterial isolates on the growth and morphology of axenic plantlets of the green alga Ulva linza. Micro. Ecol. 52: 302–310, https://doi.org/10.1007/s00248-006-9060-x.Suche in Google Scholar PubMed
Matsuo, Y., Suzuki, M., Kasai, H., Shizuri, Y., and Harayama, S. (2003). Isolation and phylogenetic characterization of bacteria capable of inducing differentiation in the green alga Monostroma oxyspermum. Environ. Microbiol. 5: 25–35, https://doi.org/10.1046/j.1462-2920.2003.00382.x.Suche in Google Scholar PubMed
McCauley, J.I., Meyer, B.J., Winberg, P.C., and Skropeta, D. (2016). Parameters affecting the analytical profile of fatty acids in the macroalgal genus Ulva. Food Chem. 209: 332–340, https://doi.org/10.1016/j.foodchem.2016.04.039.Suche in Google Scholar PubMed
Monteiro, J.P., Costa, E., Melo, T., Domingues, P., Fort, A., Domingues, M.R., and Sulpice, R. (2022). Lipidome in-depth characterization highlights the nutritional value and species-specific idiosyncrasies of different Ulva species. Algal Res. 64: 102694, https://doi.org/10.1016/j.algal.2022.102694.Suche in Google Scholar
Pandey, B. and Gadgil, V.N. (1984). Fatty acids in callus cultures: influence of growth factors on fatty acid composition of total lipids in callus cells. Phytochemistry 23: 51–53, https://doi.org/10.1016/0031-9422(84)83076-9.Suche in Google Scholar
Pereira, H., Barreira, L., Figueiredo, F., Custódio, L., Vizetto-Duarte, C., Polo, C., Rešek, E., Engelen, A., and Varela, J. (2012). Polyunsaturated fatty acids of marine macroalgae: potential for nutritional and pharmaceutical applications. Mar. Drugs 10: 1920–1935, https://doi.org/10.3390/md10091920.Suche in Google Scholar PubMed PubMed Central
Rahhou, A., Layachi, M., Akodad, M., El Ouamari, N., Rezzoum, N.E., Skalli, A., Oudra, B., El Bakali, M., Kolar, M., Imperl, J., et al.. (2023). The bioremediation potential of Ulva lactuca (Chlorophyta) causing green tide in Marchica Lagoon (NE Morocco, Mediterranean Sea): biomass, heavy metals, and health risk assessment. Water 15: 1310, https://doi.org/10.3390/w15071310.Suche in Google Scholar
Ram Kumar, M., Abhishek, K., Baburajendra Prasad, V., and Kathiresan, S. (2021). IAA combine with kinetin elevates the α-linolenic acid in callus tissues of soybean by stimulating the expression of FAD3 gene. Plant Gene. 28: 100336, https://doi.org/10.1016/j.plgene.2021.100336.Suche in Google Scholar
Schultz, C., Zopf, D., Holzinger, A., Silge, A., Meyer-Zedler, T., Schmitt, M., Wichard, T., and Popp, J. (2024). Raman spectral analysis in the CHx-stretching region as a guiding beacon for non-targeted, disruption-free monitoring of germination and biofilm formation in the green seaweed Ulva. ChemPhysChem 25: e202400173, https://doi.org/10.1002/cphc.202400173.Suche in Google Scholar PubMed
Shpigel, M., Neori, A., Popper, D.M., and Gordin, H. (1993). A proposed model for “environmentally clean” land-based culture of fish, bivalves and seaweeds. Aquaculture 117: 115–128, https://doi.org/10.1016/0044-8486(93)90128-l.Suche in Google Scholar
Simon, C., McHale, M., and Sulpice, R. (2022). Applications of Ulva biomass and strategies to improve its yield and composition: a perspective for Ulva aquaculture. Biology 11: 1593, https://doi.org/10.3390/biology11111593.Suche in Google Scholar PubMed PubMed Central
Simopoulos, A.P. (2000). Human requirement for n-3 polyunsaturated fatty acids. Poult. Sci. 79: 961–970, https://doi.org/10.1093/ps/79.7.961.Suche in Google Scholar PubMed
Spoerner, M., Wichard, T., Bachhuber, T., Stratmann, J., and Oertel, W. (2012). Growth and thallus morphogenesis of Ulva mutabilis (Chlorophyta) depends on a combination of two bacterial species excreting regulatory factors. J. Phycol. 48: 1433–1447, https://doi.org/10.1111/j.1529-8817.2012.01231.x.Suche in Google Scholar PubMed
Steinhagen, S., Barco, A., Wichard, T., and Weinberger, F. (2019). Conspecificity of the model organism Ulva mutabilis and Ulva compressa (Ulvophyceae, Chlorophyta). J. Phycol. 55: 25–36, https://doi.org/10.1111/jpy.12804.Suche in Google Scholar PubMed
Ulrich, J.F., Gräfe, M.S., Dhiman, S., Wienecke, P., Arndt, H.-D., and Wichard, T. (2022). Thallusin quantification in marine bacteria and algae cultures. Mar. Drugs 20: 690, https://doi.org/10.3390/md20110690.Suche in Google Scholar PubMed PubMed Central
Wichard, T. (2015). Exploring bacteria-induced growth and morphogenesis in the green macroalga order Ulvales (Chlorophyta). Front. Plant Sci. 6: 86, https://doi.org/10.3389/fpls.2015.00086.Suche in Google Scholar PubMed PubMed Central
Wichard, T. (2023). From model organism to application: bacteria-induced growth and development of the green seaweed Ulva and the potential of microbe leveraging in algal aquaculture. Semin. Cell Biol. 134: 69–78, https://doi.org/10.1016/j.semcdb.2022.04.007.Suche in Google Scholar PubMed
Wu, M., Cao, J., Xu, Y., Xu, S., Zhou, Z., Shao, M., Wang, Z., Zhang, L., Chen, D., Wu, Y., et al.. (2023). Impact of three phycospheric bacterial strains on the growth and fatty acid composition of Isochrysis galbana. Algal Res. 74: 103183, https://doi.org/10.1016/j.algal.2023.103183.Suche in Google Scholar
Zhang, M., Zhao, X., and Ren, X. (2025). Research progress on the mechanisms of algal-microorganism symbiosis in enhancing large-scale lipid production. J. Agric. Food. Chem 73: 6345–6360, https://doi.org/10.1021/acs.jafc.4c11580.Suche in Google Scholar PubMed
Zhang, Y., He, P., Li, H., Li, G., Liu, J., Jiao, F., Zhang, J., Huo, Y., Shi, X., Su, R., et al.. (2019). Ulva prolifera green-tide outbreaks and their environmental impact in the Yellow Sea, China. Natl. Sci. Rev. 6: 825–838, https://doi.org/10.1093/nsr/nwz026.Suche in Google Scholar PubMed PubMed Central
Supplementary Material
This article contains supplementary material (https://doi.org/10.1515/bot-2024-0101).
© 2025 the author(s), published by De Gruyter, Berlin/Boston
This work is licensed under the Creative Commons Attribution 4.0 International License.
Artikel in diesem Heft
- Frontmatter
- In this issue
- Physiology and Ecology
- Bacteria-released algal growth and morphogenesis factors regenerate axenic calli derived from the macroalga Ulva (Chlorophyta) and change the fatty acid profile
- Annual cycle and ecological interactions of macroalgal epiphytes with Cymodocea nodosa and Ruppia maritima in Epanomi and Vravrona, Greece
- Past and present fucalean diversity in the island of Marettimo, Egadi Islands Marine Protected Area (Central Mediterranean, Italy)
- Taxonomy/Phylogeny and Biogeography
- Morphological and molecular characterization of Chnoospora minima (Scytosiphonaceae, Ectocarpales) along Mexican coasts, with the description of C. ramosissima sp. nov.
- Phylogeography of the marine benthic alga Gracilaria salicornia (Gracilariales, Rhodophyta) in Southeast Asia
- Unveiling a novel species, Pachymeniopsis shinchonai sp. nov., from Korea, with reference to phylogenetic relationships within the Grateloupiaceae (Halymeniales, Rhodophyta)
- First record of the red alga Ceramium pallidum (Ceramiales, Ceramiaceae) for the Mediterranean Sea
- Chemistry and Applications
- Chemical profiling of seaweeds of the Arabian Gulf by liquid chromatography-mass spectrometry and in-silico screening against MPOX
Artikel in diesem Heft
- Frontmatter
- In this issue
- Physiology and Ecology
- Bacteria-released algal growth and morphogenesis factors regenerate axenic calli derived from the macroalga Ulva (Chlorophyta) and change the fatty acid profile
- Annual cycle and ecological interactions of macroalgal epiphytes with Cymodocea nodosa and Ruppia maritima in Epanomi and Vravrona, Greece
- Past and present fucalean diversity in the island of Marettimo, Egadi Islands Marine Protected Area (Central Mediterranean, Italy)
- Taxonomy/Phylogeny and Biogeography
- Morphological and molecular characterization of Chnoospora minima (Scytosiphonaceae, Ectocarpales) along Mexican coasts, with the description of C. ramosissima sp. nov.
- Phylogeography of the marine benthic alga Gracilaria salicornia (Gracilariales, Rhodophyta) in Southeast Asia
- Unveiling a novel species, Pachymeniopsis shinchonai sp. nov., from Korea, with reference to phylogenetic relationships within the Grateloupiaceae (Halymeniales, Rhodophyta)
- First record of the red alga Ceramium pallidum (Ceramiales, Ceramiaceae) for the Mediterranean Sea
- Chemistry and Applications
- Chemical profiling of seaweeds of the Arabian Gulf by liquid chromatography-mass spectrometry and in-silico screening against MPOX