Chemical profiling of seaweeds of the Arabian Gulf by liquid chromatography-mass spectrometry and in-silico screening against MPOX
-
Hanan Al-Adilah
Hanan Al-Adilah is an associate research scientist at Kuwait Institute for Scientific Research. She has a BSc in plant biology from Kuwait University (2011). She obtained her Master’s degree from Kuwait University with distinction, in the field of marine phytoplankton diversity in 2015. She has a PhD in algal diversity and biochemistry at the University of Aberdeen. Research in her lab includes environmental algal biodiversity, algal taxonomy, biochemistry of seagrasses and algae.
Dr. Gagan Preet has been a Research Fellow at the Marine Biodiscovery Center, University of Aberdeen, UK, since 2019. His expertise lies in the study of marine natural products, with a deep foundation in organic chemistry and computational chemistry applied to drug design and discovery. His research focuses on exploring marine organisms for potential pharmaceutical applications. With years of experience, he has contributed significantly to advancing knowledge in the field of marine medicinal chemistry and drug discovery.
The group of Prof Rainer Ebel at the Marine Biodiscovery Centre, University of Aberdeen focuses on the natural product chemistry of marine and terrestrial microorganisms and marine invertebrates, with special emphasis on endophytes from both marine and terrestrial sources. He is experienced in speciation of small molecules, i.e. structure elucidation of complex natural products using state-of-the-art hyphenated techniques including LC-MS as well as modern NMR experiments.
Puja Kumari is a lecturer in Macroalgal Research at the Scottish Association for Marine Science. She is a former recipient of ARO (Israel), BIDR-Blaustein (Israel) and JSPS (Japan) and Marie Curie (MSCA-IF) postdoctoral fellowships. Her research focus is to understand how different algae interact with its abiotic and biotic environment using biochemical, molecular and applied omics approaches.
 and
Frithjof Christian Küpper
and
Frithjof Christian Küpper
Frithjof Christian Küpper has held the Chair in Marine Biodiversity at the University of Aberdeen since 2011, after 8 years at the Scottish Association for Marine Science, studying the biodiversity and biochemistry of marine plants/algae. He found that iodide serves as an inorganic antioxidant in kelp, the first described from a living system, impacting atmospheric and marine chemistry. A certified scientific diver, Frithjof has worked in the Mediterranean, South Atlantic (Ascension, Falklands), Antarctica, the Arctic and the Gulf.


Abstract
This work conducted metabolic profiling on nine brown, one red, and two green algal species collected from different sampling sites of Kuwait’s coastal waters in the Arabian Gulf. Crude extracts of freeze-dried seaweed samples were prepared using solvents of different polarities by solvent extraction, namely, methanol and dichloromethane, and analysed by liquid chromatography-mass spectrometry for metabolite identification. The Principal Component Analysis scores scatter plot grouped the extracts of Codium papillatum, Chondria sp., Iyengaria stellata, Feldmannia indica, Padina boergesenii, Colpomenia sinuosa, Dictyota dichotoma, Sargassum aquifolium, S. asperifolium and S. ilicifolium together, suggesting their similar chemical profiles. However, S. ilicifolium var. acaraeocarpum and C. vagabunda were separated from the remainder of seaweed taxa by unique metabolite features. As expectable, species-specific differences in chemical profiles were observed in different seaweeds. Dereplication studies were employed to identify differential metabolites and to search for their previously reported bioactivities. The study led to the tentative identification of 22 metabolites using various annotation tools contained within the Global Natural Products Social Molecular Networking platform, most importantly MolNetEnhancer. The most prevalent chemical class annotations were “terpenoids” and “lipids and lipid-like molecules”. The potential modes of action of the metabolites in certain diseases were further studied using computational approaches such as molecular docking and ligand-based pharmacophore generation. This constitutes the first metabolomics study of seaweeds from the Arabian Gulf.
1 Introduction
Marine natural products are a structurally and biosynthetically diverse source of unique bioactive metabolites, including some with unusual mechanisms of action (Costa-Lotufo et al. 2009). In many cases, macroscopic seaweeds are easy to collect and contain a wide variety of compounds with interesting properties and applications in pharmacology and drug development (Gnanavel et al. 2019; Pérez et al. 2016). Seaweeds produce secondary metabolites in response to various abiotic and biotic environmental factors (Pérez et al. 2016).
Among the wide range of bioactive compounds identified from seaweeds, including peptides, phlorotannins, polysaccharides, carotenoids, and sterols, a number exhibit significant anti-hypertensive and anti-obesity properties (Lee et al. 2020; Seca and Pinto 2018; Wan-Loy and Siew-Moi 2016). Phlorotannins, abundant in seaweeds, possess multiple physiological activities, including antioxidant and antidiabetic properties (Agarwal et al. 2023; Gunathilaka et al. 2020; Jacobsen et al. 2019; Zheng et al. 2022). Compounds such as porphyran, carrageenan, halogenated monoterpenes, terpenes, and their derivatives, sourced from seaweeds, have potential against various cancers (Besednova et al. 2019; Khotimchenko et al. 2020; Liu et al. 2019; Rocha et al. 2018). Furthermore, extracts and fractions from red algae containing steroids, flavonoids, and triterpenoids with antidiabetic activity (Nguyen et al. 2019; Sanger et al. 2019).
Numerous studies have been conducted in seaweeds to characterise new bioactive secondary metabolites, including polyphenols, terpenoids, alkaloids, sterols, halogenated compounds and peptides in seaweeds using classical chromatographic separation, combined with liquid chromatography mass spectrometry (LC-MS) and nuclear magnetic resonance (NMR) for structure elucidation (Indergaard 1983). For instance, 54 polyphenolic compounds exhibiting antioxidant properties were tentatively identified in 8 seaweeds belonging to the genera Ulva, Caulerpa, Codium, Dasya, Grateloupia, Centroceros, Sargassum and Ecklonia using LC-ESI-QTOF-MS/MS (Zhong et al. 2020). In a recent review of secondary metabolites isolated from seaweeds, 33 metabolites out of 53 were potentially cytotoxic terpenoids. Furthermore, one-third of them exhibited moderate cytotoxicity (IC50 between 10 and 15 μM) and 20 compounds were significantly cytotoxic (IC50 between 1 and 10 μM). A dimeric sesquiterpene of the cyclolaurane-type and a halogenated monoterpene were extremely cytotoxic (IC50 between 1 and 5 μM) against several cancer lines (Rocha et al. 2018). Additionally, these authors emphasised that the diterpene dictyolactone isolated from Dilophus ligolatus (Bouaicha et al. 1993), the sterol cholest-5-en-3,7-diol isolated from Laurencia tristicha (Sun et al. 2007), and the halogenated monoterpene halomon isolated from Portieria hornemannii (Fuller et al. 1994) displayed cytotoxicity with IC50 values below 1 μM.
An untargeted metabolomics approach is based primarily on the qualitative or semi-quantitative analysis of the most significant metabolites from a diversity of chemical and biological classes contained in a biological specimen (Klassen et al. 2017). The analysis of metabolites based on separation technologies such as liquid chromatography (Yu et al. 2014), mass spectrometry (Won et al. 2014), nuclear magnetic resonance spectroscopy (Hilbert et al. 2015), and gas chromatography-mass spectrometry (GC-MS) can rapidly provide an overall picture of all chemical components, with minimal manipulation of the sample, and clarify the similarities and differences between biological samples (Miliauskas et al. 2006). Untargeted metabolomic studies are common to identify novel unique metabolites, though their annotation can be challenging (Miliauskas et al. 2006). LC-MS analysis generates a large number of spectra and spectral identification of each compound, one by one, using classical dereplication with reference databases (DBs) such as NCBI, Metlin, NIST, or MassBank.
Dereplication is an essential step in natural product discovery, allowing for rapid identification of known and unknown compounds in biological extracts and preventing re-isolation and re-characterisation of known bioactive compounds (Beutler et al. 1990). The isolation and identification of an active compound derived from natural products can be expensive and time-consuming. Therefore, it is necessary to determine early on if a compound was previously reported (Dias et al. 2012). Dereplication methods often involve a combination of bioassay, separation science, spectroscopic techniques and database searching, known as chemical or biological screening processes (Dias et al. 2012). Access to suitable scientific databases is crucial for dereplicating crude extracts in natural product research (Dias et al. 2012; Martins et al. 2014). It has been shown that only 1.8 % of the data can be annotated using existing MS/MS reference libraries in the public domain (da Silva et al. 2015; Wang et al. 2016). The inability to annotate data is a significant bottleneck in metabolomic data analysis that often results in reporting only the known molecules in an untargeted dataset and ignoring the rest.
New approaches have been developed to guide the isolation of new metabolites. In particular, this involves molecular networking which provides a visual overview of all the ions of molecules that were detected and fragmented during an MS experiment and the chemical relationships between them (Watrous et al. 2012). One of the currently most widely used molecular networking platforms is Global Natural Products Social Molecular Networking (GNPS), a molecular networking and data-sharing web-based (Wang et al. 2016), allowing researchers to take advantage of this bioinformatics strategy and perform data-driven, crowd-sourced analysis. GNPS is used to analyse metabolomes of all types of samples from plant extracts, microbial cultures, human cells, tissues, marine, environmental and disease samples, utilising spectral library-based annotation (Quinn et al. 2017). GNPS is based on the principle that structurally similar metabolites display similar MS/MS fragmentation patterns, allowing for the clustering and subsequent instantaneous visual investigation of identical molecules, analogues, or compound families. A vector-based computational algorithm is used to compare the degree of spectral similarity between every MS/MS spectrum in a dataset generating molecular networks (MNs) (Guthals et al. 2012; Watrous et al. 2012). The resulting MNs show diverse clusters composed of several m/z features (defined as “nodes”) characterised by their MS/MS data. Nodes within the same cluster (based on shared fragmentation patterns) are connected with edges (lines) to corresponding nodes, with the degree of similarity in their fragmentation patterns (and by inference, the degree of chemical similarity) being expressed by a so-called cosine score (Wang et al. 2016). Identical fragmentation patterns, e.g. due to similar compounds, will lead to a cosine score of 1. At the same time, it is common to set a threshold cosine score of 0.6–0.7 for clustering chemically related compounds into families in most studies involving secondary metabolites (Beniddir et al. 2021; Wang et al. 2016).
Recently, a combination of molecular networking and a bioassay-guided fractionation approach has been applied to seaweeds to isolate new bioactive compounds. For example, antimicrobial galactolipids and phlorotannins were identified from Baltic Fucus vesiculosus using UPLC-QToF-MS/MS-based bioactive molecular networking on bioactive hexane and n-butanol sub-extracts coupled with in silico dereplication tools (Buedenbender et al. 2020). Similarly, anti-helminthic fatty acids, stearidonic, α-linolenic, eicosapentaenoic, docosahexaenoic and arachidonic acids, and mono-glycerol, MG 20:5) were obtained from the brown seaweeds Saccharina latissima and Laminaria digitata using combined molecular networking and bio-assay guided fractionation (Bonde et al. 2021). Genetic and temporal chemodiversity of extracts of 14 seaweeds of the genera Dictyopteris, Dictyota, Canistrocarpus, and Laurencia collected from Brazil were demonstrated using exploratory molecular networking analysis (Philippus et al. 2018), successfully annotating various bioactive terpenoids and C15-acetogenins previously isolated from the same or related species using GNPS tools. The Carriot group successfully annotated 212 metabolites, including lipids and terpenoids, in the Taonia atomaria metabolome, using integrated LC/MS-based molecular networking and a classical phytochemical approach (Carriot et al. 2021). Recently, discriminant surface metabolites (ulvaline, MGTAs, DGTAs and carboxylic acid derivatives) involved in fouling resistance were identified in Fucus species by employing the GNPS molecular networking approach (Oppong-Danquah et al. 2023). Moreover, the authors also putatively characterised two new lipid classes, including a geranylgeranyl or a farnesyl moiety together with a rare class of acylglycerols bearing a methacrylic acid moiety (fulvenllic acid derivatives) (Carriot et al. 2021). Additionally, several putative primary and secondary metabolites of algal origin, along with bacterial and fungal secondary metabolites, were mapped onto the algal surface (or inner tissues) of F. vesiculosus by desorption electrospray ionisation – imaging mass spectrometry (DESI-IMS) coupled with GNPS tools (Parrot et al. 2019). These studies indicated that GNPS is a powerful and effective tool for analysing both known and unknown metabolites from seaweeds.
The monkeypox virus (MPOX) is a member of the poxviridae family, chordopoxvirinae subfamily, and belongs to the clade Orthopoxviruses, that also includes the Variola virus which is responsible for smallpox, cowpox, vaccinia, and variola (Sapkal and Agrawal 2022; Singh et al. 2023). It is endemic to West and Central Africa (Antunes and Virgolino 2022). The MPOX outbreak has extended its reach to 82 countries outside its usual range. Given the absence of a dedicated vaccine or antiviral (Bisanzio and Reithinger 2022; Sahu et al. 2023), currently identified potential treatments for monkeypox include tecovirimat, brincidofovir, and cidofovir, which are considered possibly effective, as they have also been licensed for the treatment of smallpox (Hoy 2018; Rani et al. 2023); however, their use has not been studied outside of experimental models. The application of other drugs has not been widely explored. Considering the knowledge gap and urgency for obtaining effective MPOX treatments, the present study explored through molecular modelling approaches whether some of the metabolites obtained from seaweeds of the Gulf might have potential in this regard.
This study obtained metabolite profiles of 12 seaweeds from the Arabian Gulf using an integrated LC-MS-based dereplication and GNPS platform. The chemical diversity between different seaweeds was elaborated using multivariate principal component analysis (PCA) and heat maps. In addition, selected compounds were isolated and purified from algal extracts showing distinct metabolite profiles by solid-phase fractionation and HPLC, and their structures were elucidated using NMR. Afterwards, a computational approach comprising molecular docking analysis was performed to understand the binding potentials of selected compounds against three vital MPOX-related poxvirus proteins because these selected compounds have similar scaffolds which have shown bioactivity against number of poxviruses (Preet et al. 2022). Based on the results, a pharmacophore evaluation study was also conducted to generate ligand-based feature pharmacophores for the best-performing selected compound.
This study was conducted following our recent work on halogens (Al-Adilah et al. 2020, 2022), trace elements (Al-Adilah et al. 2021a) and fatty acids in seaweeds (Al-Adilah et al. 2021b) of Kuwait. Significantly, the present work constitutes the first ever metabolomics study of seaweeds in the Gulf region.
2 Materials and methods
2.1 Sampling
A total of 12 different common seaweed species, which were expected to contain promising metabolites and representing three major groups – green, red, and brown seaweeds – were collected from different sampling sites in Kuwait’s coastal waters in the Arabian Gulf between May and June 2018 and February 2021 (Al-Adilah et al. 2020, 2021a, b, for all details of collections). Specifically, these included Cladophora vagabunda (L.) C. Hoek (9.5 kg FW), Codium papillatum C. K. Tseng et W. J. Gilbert (8 kg FW) [both Chlorophyta], Chondria sp. C. Agardh (8 kg FW) [Rhodophyta], Iyengaria stellata (Børgesen) Børgesen (5.8 kg FW), F. indica (Sonder) Womersley et A. Bailey (11.0 kg FW), Padina boergesenii Allender et Kraft (11.0 kg FW), Colpomenia sinuosa (Mertens ex Roth) Derbes et Solier (4.5 kg FW), Dictyota dichotoma (Hudson) J.V.Lamouroux (23.5 kg FW), Sargassum aquifolium (Turner) C.Agardh (8 kg FW), Sargassum asperifolium Hering et G.Martens ex J.Agardh (4.5 kg FW), Sargassum ilicifolium (Turner) C.Agardh (10 kg FW) and S. ilicifolium var. acaraeocarpum Grunow (2 kg FW) [all Ochrophyta]. Seaweeds were identified based on their morphological characteristics and DNA barcoding. The taxonomy of Sargassum species used here is consistent with the latest study of this genus in Kuwait and the Gulf (Hasan et al. 2023). Seaweed samples were rinsed thoroughly with seawater on-site and placed in plastic bags. Date of collection and location were noted. These samples were transferred to the laboratory and washed three times with seawater. Fresh samples were frozen at −20 °C for 24 h, followed by lyophilisation in a freeze dryer (Labconco, USA) at −45 °C for 48 h.
2.2 Seaweed metabolite extraction and LC-MS sample preparation
The freeze-dried seaweed samples were homogenised in methanol (MeOH) for 3 days, followed by dichloromethane for 24 h thrice. The initial crude extract generally consists of a wide variety of chemical compounds differing in polarity. All the initial extracts were combined, evaporated to near dryness under reduced pressure using a rotary evaporator and transferred to a glass vial. Then, the extracts were dried entirely under nitrogen gas, weighed, and stored at −20 °C until analysed. For LC-MS analysis, 1 mg of each crude extract was transferred to an Eppendorf tube, and 1 ml of MeOH was added to dissolve the extract. Subsequently, extracts were diluted to a concentration equivalent to 0.5 mg ml−1 and centrifuged at 16,060g for 10 min before being transferred to LC-MS vials.
2.3 HPLC Q-TOF MS/MS analysis
All analyses were performed using a Bruker MAXIS II Q-ToF mass spectrometer coupled to an Agilent 1290 UHPLC system. Based on single-time injection on HPLC-MS, the data have no replicates since single-run analysis is hard to reproduce with collected algal samples. Separation was achieved using a Phenomenex Kinetex 103 XB-C18 (2.6 mM, 100 × 2.1 mm) column and the following LC gradient profile: 5 % acetonitrile + 0.1 % formic acid to 100 % acetonitrile + 0.1 % formic acid in 15 min even longer to allow for full elution. MS parameters were as follows; mass range m/z 100–2000, capillary voltage 4.5 kV, nebuliser gas 5.0 bar, dry gas 12.0 l min−1, and a dry temperature of 220 °C. MS/MS experiments were conducted under auto MS/MS scan mode with no step collision. The system was then reequilibrated to the starting conditions before the next run. The spectral data were recorded in the range m/z 100–3,000, and data files were converted from Bruker Data Analysis (.d) to mzML .mzXML file format using MS Convert (http://proteowizard.sourceforge.net/index.html) (Chambers et al. 2012) for processing analysis. For background subtraction, blank runs were obtained by injecting solvent only.
After eliminating peaks also present in the blank runs, the individual spectra for the most 7–10 prominent peaks in each run were analysed manually by examining the quasimolecular ions (mainly [M+H]+ or [M+Na]+). Candidate molecular formulae of individual metabolites were predicted using three factors, i.e., low mass error value (±3 ppm), low mSigma value (<20) and the high score percentage above 80 %, representing the software evaluation of the low error and mSigma value. The mass error is a measure of the deviation zn of the experimental from the predicted accurate mass for a given molecular formula, which, according to the standard applied in the field, should be within ± 5 ppm, while the mSigma value represents the match of the measured isotope pattern compared with the theoretical one predicted based on the atoms present in the molecular formula. The score value is a weighted average of these two parameters applied by the instrument manufacturer’s software. The abovementioned thresholds represent empirical values used within the Aberdeen Marine Biodiscovery Centre for the last 10–15 years.
2.3 Data processing and multivariate analysis
Converted mzML mass files were imported to MZmine v2.40.1 (Holman et al. 2014) to compensate for variations in retention times and m/z values between each analysis. Chromatograms were built using a noise level of 1,000, a minimum peak height of 10,000, a minimum time span of 0.2 min, and an m/z tolerance of 0.001 and deconvoluted within a 90 % threshold over a 40-min time range with a minimum relative height of 5 % and a minimum ratio of top/edge of 2, with only the most intense isotope being retained. Peak lists were aligned using the join aligner method with a 5 % relative retention time tolerance and weight for retention time, m/z kept at 20, and gap-filled using a 25 % intensity tolerance and retention time tolerance of 0.5 min. Mass spectral 104 data were exported into a file with a comma-separated values data format (*.csv) as a peak list table, with rows representing the seaweed samples and columns representing the integrated and normalised peak areas (m/z values and retention times). The CSV file (*.csv) was imported to SIMCA version 15 (MKS Data Analytics Solutions, Umeå, Sweden) for principal component analysis (PCA). PCA score plots and loading plots were constructed using positive (ESI+) precursor ions. This model developed a covariance between X data matrices (metabolites spectra) and Y matrices (seaweeds) to produce a scores plot showing the relationship between seaweeds and a loadings plot showing the variations in X matrices that were responsible for the differences between seaweeds. Heat maps were generated to identify discriminant metabolites using ggplot of programming software package R (version_64 2.15.2; R Foundation for Statistical Computing, Bristol, UK) (Wickham 2011) to plot all m/z ratios from the sample set.
The molecular formulae obtained using Bruker Daltonics Data Analysis software were searched against an Excel sheet for known compounds from brown, green and red algae which had been prepared by searching the Antibase 2017 database using ChemFinder (Laatsch 2017). Afterwards, each one of these exact masses for the 5–8 most prominent peaks in each run was manually searched against the data present in the Excel sheet, in addition to calculating the values for [M+H]+ and [M+Na]+, which were expected to appear in the runs for individual species if these compounds were present.
2.5 Annotation of metabolites using molecular networking
Additionally, a molecular network was created using the online workflow of GNPS. The data were filtered by removing all MS/MS peaks within ±17 Da of the precursor m/z. MS/MS spectra were window-filtered by choosing only the top 6 peaks in the ± 50 Da window throughout the spectrum. The data were then clustered with MS-Cluster (Guthals et al. 2012), with a parent mass tolerance of 2.0 Da and an MS/MS fragment ion tolerance of 0.5 Da to create consensus spectra. Also, consensus spectra containing fewer than one spectrum were discarded. Cosine similarity scores, ranging from 0 to 1 (1 being identical spectra), were computationally combined as consensus spectra if more than six ion fragmentation spectra matched. A minimum cosine score of 0.7 was selected to subdue the clustering of different compound classes under the same molecular family when visualising the data as a network of nodes connected by edges (Cline et al. 2007). The spectra in the network were searched against the GNPS spectral libraries. The achieved GNPS molecular networking was further annotated with MolNetEnhancer (Ernst et al. 2019). Network Annotation Propagation, Dereplicator and MS2LDA_MotifDB were incorporated into the network using GNPS MolNetEnhancer. Data were visualised by importing into Cytoscape suite (version 3.0.2) visualisation software, and nodes and edges were displayed (nodes correspond to a specific consensus spectrum; edges represent significant pairwise alignment between nodes).
2.6 Fractionation and isolation of seaweed metabolites
2.6.1 Fractionation of crude extracts
The crude extracts of individual algal samples were fractionated according to the modified Kupchan scheme (Kupchan et al. 1973). The fractionation procedures mainly depend on the polarity of compounds of interest. First, the initial crude extract was partitioned between water and CH2Cl2, after which the CH2Cl2 layer was evaporated under a vacuum, and the resulting residue was partitioned between n-hexane and 10 % aqueous MeOH. The MeOH layer was then phase adjusted to 50 % aqueous MeOH and extracted with CH2CL2. Then, the aqueous layer was partitioned with an equal volume of 2-butanol. These were further purified by RP-HPLC using a 30 min gradient program of 50–100 % MeOH in H2O using instruments of the Agilent HPLC 1100 and 1200 series.
2.6.2 Nuclear magnetic resonance spectroscopy (NMR)
Pure compounds obtained from HPLC-MS were characterised by 1D-NMR Varian 400 MHz (1H and 13C NMR) and 2D-NMR Varian 600 MHz (COSY, HSQC, HMBC and NOESY) in the Marine Biodiscovery Centre, Department of Chemistry, University of Aberdeen. Samples were prepared in either deuterated methanol or deuterated chloroform. Chemical shifts are reported in parts per million (ppm) relative to residual solvent signals.
2.6.3 Molecular docking
Molecular docking analysis was performed using the same computational approach (Preet et al. 2022) Autodock Vina v.1.2.0 (The Scripps Research Institute, La Jolla, CA, USA) docking software (Trott and Olson 2010). The receptor site was predicted using LigandScout (Inte: Ligand) Advanced software. Furthermore, all ligands were set to minimize the preset of 1,000 steps (N = 1000, M = 25 and E t = 0.05 kcal mol−1) where N is the maximum number of minimization steps, M is the number of consecutive minimization steps, and E t is the energy difference between steps that is less than the threshold, before docking experiments were performed in Autodock Vina v.1.2.0.
The X-ray crystal structure of the first protein used for docking was PDB: 2V54 (Caillat et al. 2008), which is the Vaccinia virus with the functional enzyme thymidylate kinase. The second is PDB: 4QWO A42R (Minasov et al. 2022), a profilin-like protein from the monkeypox virus Zaire-96-I-16. The third protein, Vaccinia virus D13, PDB: 6BED (Garriga et al. 2018), was retrieved from the Protein Data Bank and utilized to perform docking simulations. The box center and size coordinates for (PDB: 2V54) were 5.6 × 19.2 × −30.6 and 26.2 × 33.2 × 19.9; for (PDB: 4QWO) they were 1.2 × 16.7 × 16.5 and 24.7 × 21.1 × 28.6; for (PDB: 6BED) they were 106.0 × 100.4 × 14.0 and 27.9 × 38.6 × 96.4 around the active site. All coordinates used Angstrom units. Experimental search parameters set to the number of binding modes were 5, exhaustiveness was 32, and the maximum energy difference was 3 kcal/mol. Chimera 1.16, UCSF_USA, LigPlot+ software (EMBL-EBI, Cambridgeshire, UK) (Laskowski and Swindells 2011) and Samson 2022 (OneAngstrom, French Institute for Research in Computer Science and Automation, Domaine de Voluceau, France), were used for visualization and calculation of protein–ligand interactions.
2.7 Pharmacophore model generation
Pharmacophore model generation was performed using the same approach (Preet et al. 2022). LigandScout by Inte Ligand, Advanced software (Wolber and Langer 2005), Vienna, Austria (Pettersen et al. 2004), was used to generate a pharmacophore model. The Espresso Algorithm was used to generate a ligand-based pharmacophore. Relative Pharmacophore-Fit scoring function, Merged feature pharmacophore type and feature tolerance scale factor were set to 1.0 for Ligand-Based Pharmacophore creation. The best model was selected from the ten models generated.
3 Results and discussion
Twelve of the most common seaweed species belonging to the phyla Chlorophyta, Rhodophyta and Ochrophyta from the Arabian Gulf coast of Kuwait were analysed to obtain their metabolite profiles. At first, the unannotated seaweed metabolome spectra were compared using principal component analysis and heat map to identify any species-specific molecular mass features. Then, the metabolites were integrated and annotated manually using the available databases as mentioned in Section 2. Finally, selected metabolites were investigated computationally for the potential to bind to MPOX, given the critical shortage of treatments against this virus pathogen.
3.1 Comparison of seaweed metabolomes
The metabolite profiles of different seaweeds were compared using PCA and a heat map. PCA displayed an overview of the distribution of various metabolites, explaining 50 % of the variation in the data (PC1 – 27.6 % and PC2 – 22.4 %; Figure 1a). It can be observed that the values of R2 = 0.555 and Q2 = 0.21 indicated that the model had very good fitness and prediction ability as both R2 and Q2 > 0.5, and the difference between R2Xcum and Q2cum was less than 0.2–0.3, highlighting the importance of the models. This result shows that the classification model is feasible and successful regarding variety.
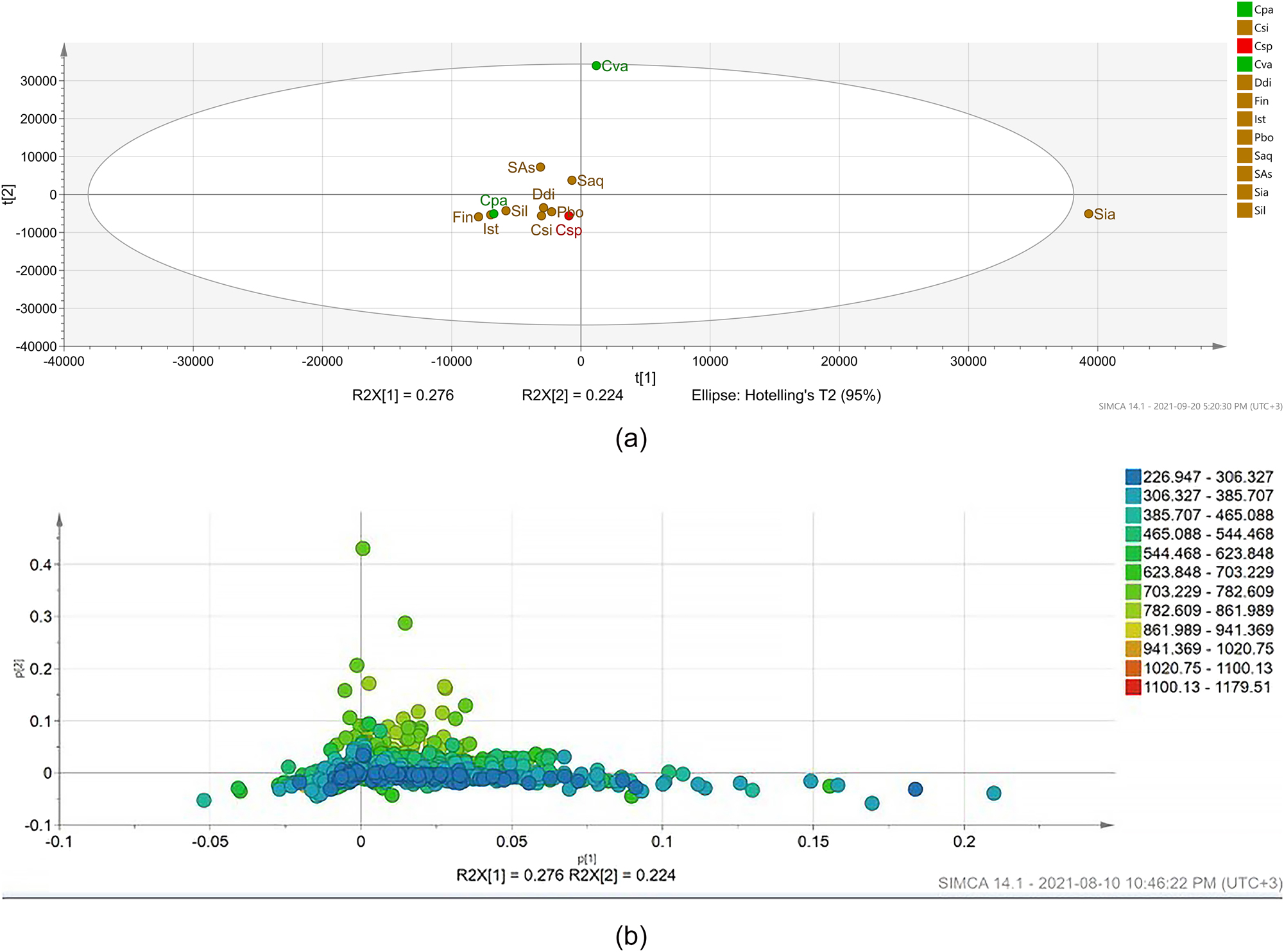
Principal component analysis (PCA) based on LC-MS spectra of 12 seaweed extracts. (a) Scores plot and (b) loading plot sub-figures show metabolites (in molecular weight). Each node represents an m/z value. This figure shows the relationship among seaweeds (scores plot) and molecules in the loading plot that were unique to either one particular seaweed or groups of seaweeds. The loading plot enables identification of metabolites that are most responsible for driving the patterns seen in the score plot of the crude extracts. Abbreviations: Csp, Chondria sp.; Cva, Cladophora vagabunda; Cpa, Codium papillatum; Csi, Colpomenia sinuosa; Ddi, Dictyota dichotoma; Fin, Feldmannia indica; Ist, Iyengaria stellata; Pbo, P. boergesenii; Saq, Sargassum aquifolium; Sas, Sargassum asperifolium; Sia, Sargassum ilicifolium var. acaraeocarpum; Sil, S. ilicifolium. X- and Y-axes show scores for first and second principal components, respectively (X axis represents PC1, and Y axis represents PC2).
From the score plot (Figure 1a), seaweed extracts showed similar metabolite profiles with several compounds shared between most samples and clustered together. However, S. ilicifolium var. acaraeocarpum and C. vagabunda were separated from the rest of the seaweed samples due to unique metabolite features; specifically, these comprised m/z 359.2216 (most likely corresponding to candidate molecular formula C22H30O4) and m/z 301.135 (most likely corresponding to candidate molecular formula C13H21N2O6) for S. ilicifolium var. acaraeocarpum; and m/z 327.223 (most likely corresponding to candidate molecular formula C17H31N2O4) as well as m/z 760.591 (most likely corresponding to candidate molecular formula C46H82NO7) for C. vagabunda.
Furthermore, a heat map analysis of seaweed species and the m/z values of the metabolites identified in them showed the relative levels of different putative metabolites (Figure 2; Table 1). Interestingly, among the 1906 ions encountered in this study, 803 were shared between seaweeds. In general, the metabolic profile of S. ilicifolium var. acaraeocarpum suggested the highest degree of chemical complexity (900 detected unique masses) among all seaweeds analysed here, whereas F. indica contained the lowest number of detected unique masses (159).
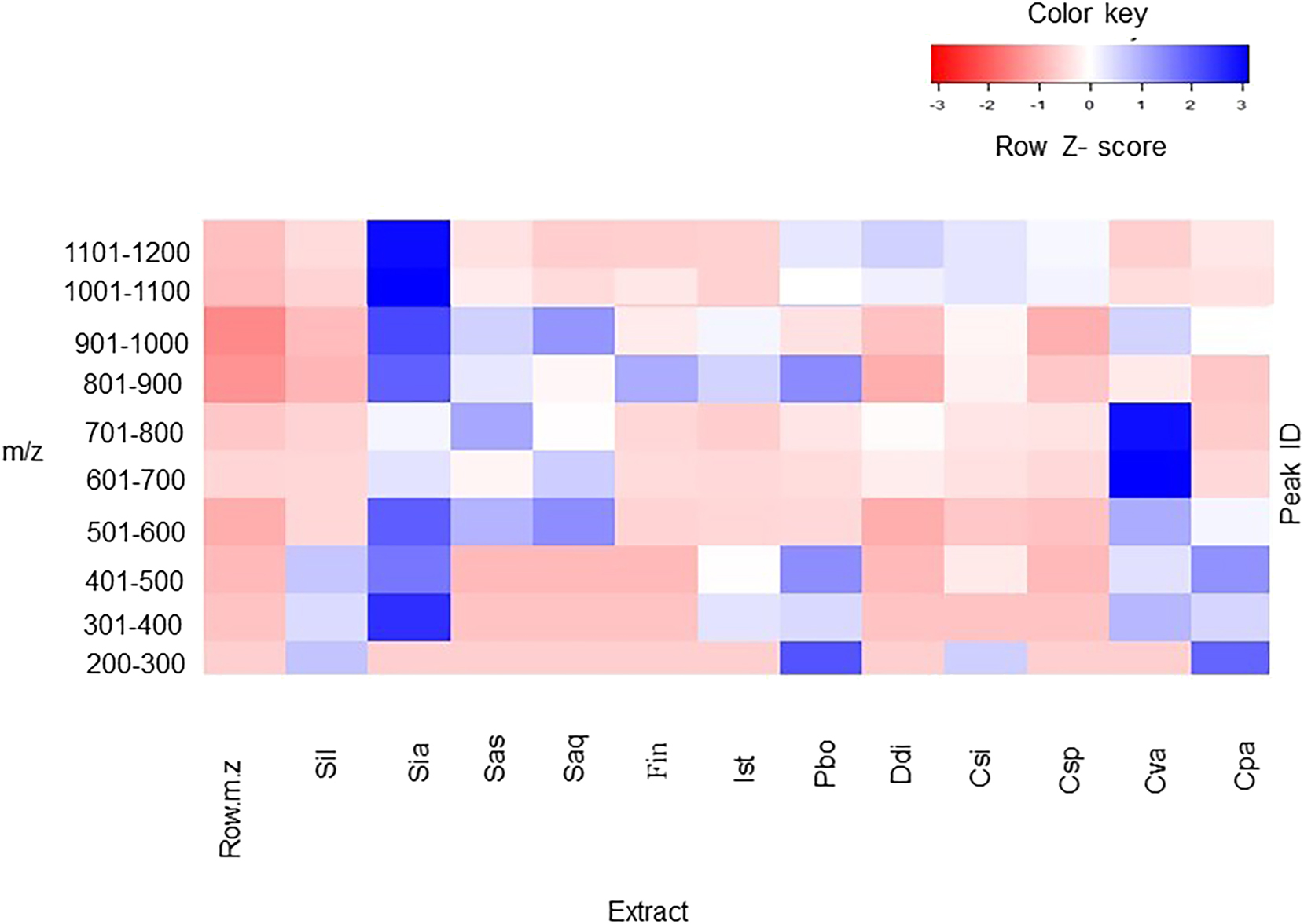
Heatmap showing relative intensities of the 1906 metabolites ranging from m/z 226–1179 in raw spectra across 12 different seaweeds. Abbreviations: Csp, Chondria sp.; Cva, Cladophora vagabunda; Cpa, Codium papillatum; Csi, Colpomenia sinuosa; Ddi, Dictyota dichotoma; Fin, Feldmannia indica; Ist, Iyengaria stellata; Pbo, P. boergesenii; Saq, Sargassum aquifolium; Sas, Sargassum asperifolium; Sia, Sargassum ilicifolium var. acaraeocarpum; Sil, S. ilicifolium. Metabolite features whose levels vary significantly (p < 0.01) across 12 different seaweeds are projected onto the heat map and used for sample clustering. Rows display metabolites and columns represent the samples. The row Z-score of each feature is plotted in red-blue color scale. The blue color of the tile indicates high abundance and red indicates low abundance. The brightness of each color corresponds to the magnitude of the difference when compared with average value. A z-score of zero indicates that a value is on the mean.
Relative intensities of the 1906 metabolites ranging from m/z 226–1179 in raw spectra of different seaweed extracts.
| Species no. | Species | Phylogenetic affinity | Mass range with highest abundances (m/z) | |||||
|---|---|---|---|---|---|---|---|---|
| 301–400 | 401–500 | 501–600 | 601–700 | 701–800 | 801–900 | |||
|
|
||||||||
| Ochrophyta | ||||||||
| 1 | Sargassum ilicifolium | Sargassaceae, Fucales, Phaeophyceae | xx | |||||
| 2 | S. ilicifolium var. acaraeocarpum | Sargassaceae, Fucales, Phaeophyceae | ||||||
| 3 | Sargassum asperifolium | Sargassaceae, Fucales, Phaeophyceae | xx | |||||
| 4 | Sargassum aquifolium | Sargassaceae, Fucales, Phaeophyceae | xx | |||||
| 5 | Feldmannia indica | Acinetosporaceae Ectocarpales, Phaeophyceae | xx | |||||
| 6 | Iyengaria stellata | Scytosiphonaceae, Ectocarpales, Phaeophyceae | x | xx | ||||
| 7 | Padina boergesenii | Dictyotaceae, Dictyotales, Phaeophyceae | xx | |||||
| 8 | Dictyota dichotoma | Phaeophyceae, Dictyotales, Ochrophyta | x | |||||
| 9 | Colpomenia sinuosa | Phaeophyceae, Ectocarpales Ochrophyta | x | x | x | x | x | |
|
|
||||||||
| Rhodophyta | ||||||||
|
|
||||||||
| 10 | Chondria sp. | Rhodomelaceae, Ceramiales, Florideophycea | x | x | x | |||
|
|
||||||||
| Chlorophyta | ||||||||
|
|
||||||||
| 11 | Cladophora vagabunda | Cladophoraceae, Cladophorales, Ulvophyceae | xx | x | ||||
| 12 | Codium papillatum | Codiaceae, Bryopsidales, Ulvophycea | x | x | ||||
-
x, lower abundances; xx, highest abundances.
Most compounds encountered were found between m/z 300–400 in seaweeds in the order S. ilicifolium var. acaraeocarpum > C. sinuosa > S. aquifolium > S. asperifolium > C. vagabunda > P. boergesenii > Chondria sp. > D. dichotoma > I. stellata > C. papillatum > F. indica > S. ilicifolium. No specific pattern was observed for the abundance of molecular masses in other mass ranges among different seaweeds. Further, based on the abundance of masses, strong species-specific differences between abundance of different masses were observed in different seaweeds.
Among Sargassum species, masses in the range of m/z 301–400 showed the highest abundance in S. ilicifolium, S. ilicifolium var. acaraeocarpum, while masses in the range of m/z 701–800 and m/z 501–600 showed highest abundance in S. asperifolium and S. aquifolium respectively. F. indica, I. stellata, and P. boergesenii showed the highest abundance of masses in the range of m/z 601–700, followed by m/z 301–400 in F. indica, and P. boergesenii, and m/z 501–600 in I. stellata. In D. dichotoma, masses in the range m/z 301–400 showed the maximum abundance, followed by m/z 701–799, while all other masses had low abundance. C. sinuosa had mass abundance similar to S. ilicifolium and S. ilicifolium var. acaraeocarpum, with the highest abundance in the range of m/z 301–400, followed by approximately similar intensities for m/z 401–800, marginal abundance for masses less than m/z 300 and very low abundances for more than m/z 800. In the red alga Chondria sp., masses in the range m/z 301–400 showed the highest abundance, similar to most brown algae, followed by masses in the range m/z 701–800, and m/z 401–500 and m/z 601–700. The two green algae, C. vagabunda and C. papillatum, showed very distinct mass abundances. The masses in the range m/z 701–800 showed the highest abundance (due to higher intensities of m/z 760.5915 and 761.5915), followed by m/z 801–900 in C. vagabunda (due to higher Abundance of m/z 760.5914 and 761.5950 m/z) while masses in the range m/z of 301–400 showed the highest abundance in C. papillatum (due to higher intensities of m/z 393.2892 and 301.1346) followed by m/z 500–600 (due to m/z 537.2917).
3.2 Putative identification of compounds based on manual dereplication
A total of 511 metabolites were detected in seaweed samples using manual integration with Data Analysis software v 4.3 (Bruker Maxis). Each metabolite represents a unique combination of the m/z value and chromatogram peak characteristics. However, only 20 metabolites were tentatively identified with AntiBase 2017 (Supplementary Table S1; Supplementary Figures S1, S2). The commercially available Antibase database (version 2017) was employed in this study for a targeted search of individual compounds previously reported in the chemical literature, taking into account the available taxonomic information. Antibase is a comprehensive database which, as of 2017, contained more than 48,000 natural products, in which taxonomic information about the source organism(s) (if provided by the individual studies) is available for most entries relating to the individual compounds. However, it should be noted that this information is taken at the point of publication and is not updated in case later revisions allow for a more precise identification (e.g., a sample of “Codium sp.” being identified as C. papillatum) or even in case a misidentification should be detected. More importantly, the natural product-related literature has a strong, not to say extreme, bias towards the reporting of new compounds, which in many cases means that the occurrence of known compounds can only be reported in high-impact journals if the study in which they are encountered affords at least one new compound. Therefore, it is challenging to obtain an unbiased picture regarding the distribution of natural products over a complex group of organisms such as macroalgae. Based on these constraints, taxonomic information available in Antibase used in this study was restricted to the level of genus, and in case no hits were found, then to the level of the family, then order, and finally phylum. It is clear from visual inspection of the chromatograms (data not shown) that more than half of the peaks were not identified by comparison with the databases, including some of the significant components that showed strong peak intensities and good resolution. The m/z value and the predicted formula of these identified compounds are presented in Supplementary Material (Table S1; Figure S1). Matching the molecular formulae and corresponding taxonomic information was assumed to putatively identify the previously reported compound with sufficient certainty. It is clear that using this approach, only an indicative overview of the chemical diversity of individual algal species could be obtained. In those cases, more than one isomer corresponding to a given molecular formula has been reported in the literature, and no decision relating to the presence of a specific compound could be made. Finally, of course, the presence of any new compounds cannot be established at all. To a given molecular formula reported in the literature, no decision relating to the presence of a specific compound could be made.
Many compounds identified in the studied seaweeds from Antibase (listed and numbered in Table S1, structures in Figure S1) are reported to have cytotoxic properties. Isodecortinol (1) and decortinol (2) are C29-sterols were previously isolated from the ethyl acetate soluble portion of the extract of the green seaweed Codium decorticatum and Cladophora sp. (Ahmad et al. 1993). The sterol, 24-R-stigmasta-4,25-diene-3β,6β-diol (3), was isolated from the green alga Codium divaricatum, a traditional Chinese medicine (He et al. 2010). 29-Hydroxy stigmasta-5,24 (28)-dien-3β-ol (4), another sterol isolated from Chaetomorpha basiretorsa exhibited significant cytotoxic activity (Shi et al. 2008). (20 S)-3β,20-Dihydroxyergosta-5,24(28)-dien-16-one (5) isolated from Cystophora brownie (Bian and van Altena 1998) and 2-(3-methyl-2-butenyl)-2,3-epoxy-1,4-naphthalenedione 4,4-dimethyl ketal (6) isolated from Landsburgia quercifolia were reported to be active against P-388 leukaemia cells (IC50 0.6 μg/ml) and also exhibited anti-fungal properties (Perry et al. 1991). (3β,23E)-stigmasta-5,23-diene-3,28-diol (7) and (3β,28S)-24,28-epoxy-stigmast-5-en-3-ol (8) were isolated from the brown alga Sargassum carpophyllum as active compounds causing morphological abnormality of Pyricularia oryzae mycelia. Both compounds (7) and (8) also exhibited cytotoxic activity against various cancer cell lines (Tang et al. 2002). Compounds (9), a novel diterpene divarinone, and (10), (5R,8S,9S,12R,14S0-9-hydroxydolasta-1,3- dien-13-one have been reported from Dictyota divaricata (Trimurtulu et al. 1992). Dictyol A (information as retrieved from Antibase; 11), a diterpene alcohol isolated from Dictyota volubilis (Wright et al. 1993)was previously described from D. dichotoma (Fattorusso et al. 1976). Most of the putatively identified peaks belong to the class of terpenes. C29H49O2 (429.3719) was found to be present in most of the seaweeds studied here. All compounds that were detected in C. sinuosa were also present in all other brown seaweeds. The ion m/z 429.3724 was only detected in green algae species C. papillatum and C. vagabunda.
Based on PCA and heat map analyses, the metabolite profile of S. ilicifolium var. acaraeocarpum was chosen for further dereplication and annotation of putative compounds. For 32 compounds, candidate molecular formulae were assigned by manual analysis of significant peaks, and these were searched in Reaxys and NP Atlas (Table S2). Interestingly, only 6 of these candidate molecular formulae were represented in Reaxys, and 6 were represented in the MNP Atlas (which include two being present in both databases). Of course, this means that the remaining 22 candidate molecular formulae may indicate the presence of compounds so far not reported in the literature. Thus, this is a promising target for further studies involving their isolation and structural characterisation. It is important to note that the presence of a peak, even a prominent one, in a mass spectrum does not necessarily indicate the presence of a sizeable amount of a compound, due to differences in ionizability. Also, it should be stated that in the current study, the only elements allowed for when identifying candidate molecular formulae were C, H, N and O, thus a priori eliminating the possibility of detecting known (or even potentially new) compounds containing “rarer” elements (as far as natural products are concerned), such as halogens or sulphurs.
3.3 Automated annotation of metabolites using GNPS
In an attempt to extend the annotation limit of seaweed metabolites, the metabolite profiles of different seaweeds were subjected to various annotation tools contained within the GNPS platform, most importantly MolNetEnhancer. Figure 3 and Table 2 display the molecular network coloured by the most prevalent chemical class annotations. The network has 786 nodes comprising 191 clusters and 500 self-looped nodes based on similarities in MS/MS fragmentation patterns. Some nodes represented adducts; thus, not all network nodes correspond to a single molecule. Automated annotation covered 28 chemical classes.
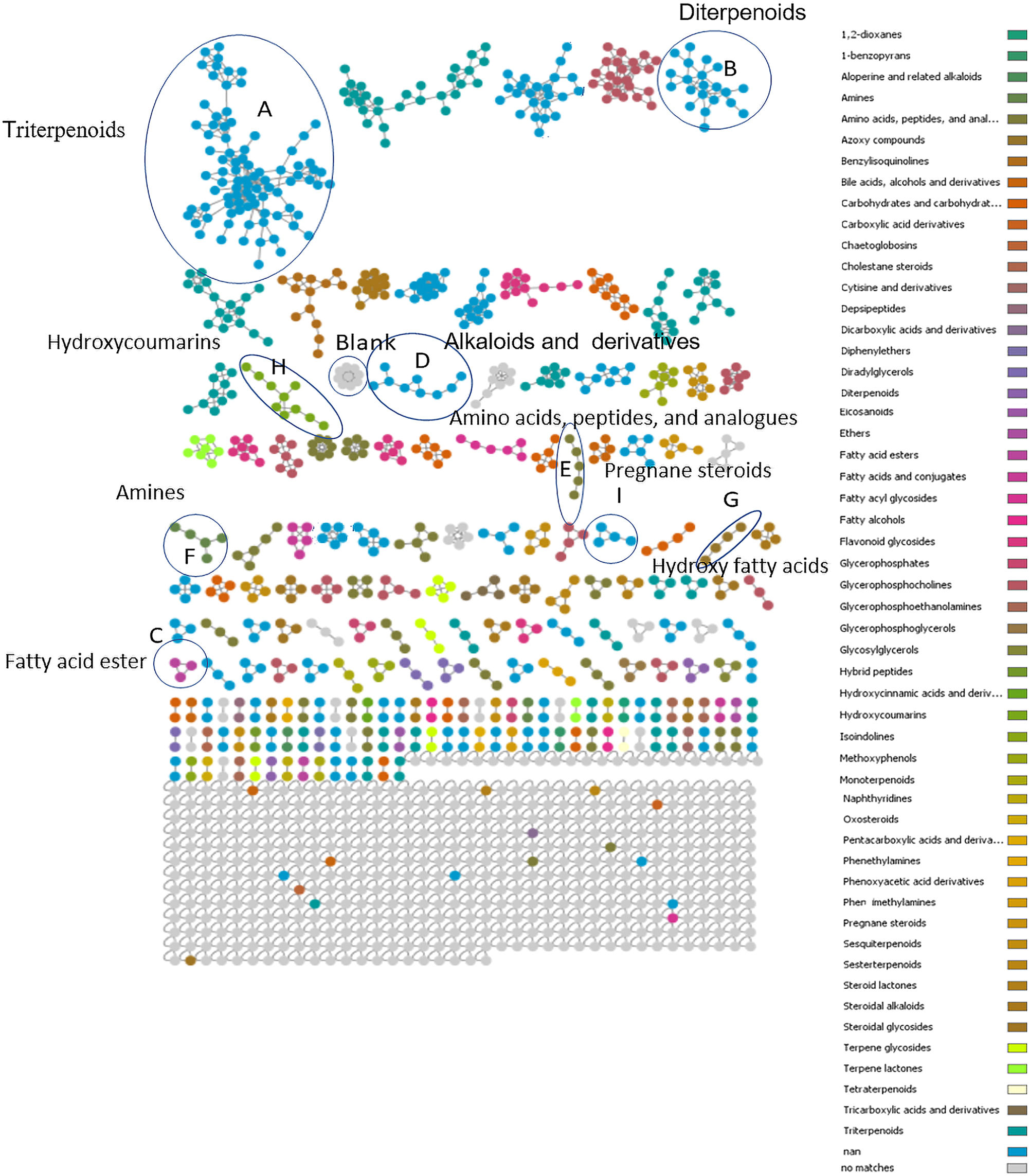
Molecular network for marine algae coloured corresponding to 28 selected chemical class terms as indicated in the legend.
The hits in precursor tolerance against the Global Natural Products Social Molecular Networking (GNPS) library.
| Cluster | Name of the compounds | Molecular mass (m/z) | Molecular formulae | Biological origin | References |
|---|---|---|---|---|---|
| Cluster A (triterpenoids) | Ursolic acid | 455.361 [M+H]+ | C30H48O3 | (Eucalyptus tereticornis) Kingdom: Plantae Phylum: Spermatophyta (Eriobotrya japonica) Kingdom: Plantae Phylum: Spermatophyta 455.361 |
Saraswat et al. (2000)
Tan et al. (2019) |
| Pygenic acid A b – 30eV | 471.356 [M+H]+ | C30H48O4 |
Prunella vulgaris
Kingdom: Plantae Phylum: Spermatophyta Lagerstroemia speciosa Kingdom: Plantae Phylum: Spermatophyta |
Choi et al. (2016), Lim et al. (2020)
Raoof and Mohamed (2018) |
|
| Pygenic acid A b – 40eV | 487.35 [M+H]+ | C30H48O5 |
Prunella vulgaris
Kingdom: Plantae Phylum: Spermatophyta Lagerstroemia speciosa Kingdom: Plantae Phylum: Spermatophyta |
Choi et al. (2016), Lim et al. (2020)
Raoof and Mohamed (2018) |
|
| Pygenic acid A b – 10eV | 471.355 [M+H]+ | C30H48O4 |
Prunella vulgaris
Kingdom: Plantae Phylum: Spermatophyta Lagerstroemia speciosa Kingdom: Plantae Phylum: Spermatophyta |
Choi et al. (2016), Lim et al. (2020)
Raoof and Mohamed (2018) |
|
| Cycloart-23-ene-3, 25 diol | 443.325 [M+H]+ | C30H50O2 |
Trichilia dregeana
Kingdom: Plantae Phylum: Magnoliophyta |
Eldeen et al. (2007) | |
| Cluster B (diterpenoids) | Lagochilin | 379.237 [M+Na]+ | C20H36O5 |
Lagochilus inebrians
Kingdom: Plantae Phylum: Tracheophyta |
Akramov et al. (2019) |
| 5-(1,2,4a,5-tetramethyl-7-oxo-3,4,8,8a-tetrahydro-2H-naphthalen-1-yl)-3-methylpentanoic acid | 220.2279 [M+Na]+ | C20H32O3 |
Clausena dunniana
Kingdom: Plantae Phylum: Tracheophyta Neomirandea angularis Kingdom: Plantae Phylum: Spermatophyta |
He et al. (2003)
Tamayo-Castillo et al. (1989) |
|
| Cluster C (fatty acid ester) | Sorbitane monostearate – polysorbate 60 in-source fragment | 428.364 [M+H]+ | C24H44O6 | Synthetic food additives | Additives et al. (2017) |
| Sorbitane monooleate – polysorbate 20 in-source fragment | 430.38 [M+H]+ | C24H46O6 | Synthetic food additives | Additives et al. (2017) | |
| Sorbitane monopalmitate – polysorbate 40 in-source fragment | 402.349 [M+H]+ | C22H42O6 | Synthetic food additives | Additives et al. (2017) | |
| Cluster D (alkaloids and derivatives) | Strychnine | 335.176 [M+H]+ | C21H22N2O2 |
Strychnos nux-vomica
Kingdom: Plantae Phylum: Spermatophyta |
Miao et al. (1998) |
| Cluster E (amino acids, peptides, and analogues) | Dendroamide C | 428.318 [M+H]+ | C21H24N6O5S3 |
Stigonema dendroideum
Kingdom: Eubacteria Phylum: Cyanobacteria |
Xia and Smith (2001) |
| Decanoyl-l-carnitine | 316.22 [M+H]+ | C17H34NO4.Cl | Synthetic | ||
| Cluster F (amines) | Oleoylethanolamine | 326.335 [M+H]+ | C20H39NO2 | Primary metabolites | |
| Linoleoyl ethanolamide | 324.319 [M+H]+ | C20H37NO2 | |||
| Cluster G (hydroxy fatty acids) | Aleuretic acid | 455.361 [M+H]+ | C16H32O5 | Seedlac | Nagappayya and Gaikar (2010) |
| Cluster H (hydroxycoumarins) | 3-Hydroxy-4-(succin-2-YL)-caryolane delta-lactone | 319.218 [M−H]- | C19H28O4 | ||
| Cluster I (pregnane steroids | Cortisone acetate | 403.237 [M+H]+ | C23H30O6 | Human milk | Al-Salman and Shaker (2017) |
| Cluster J Nan | 1,4a-dimethyl-9-oxo-7-propan-2-yl-3,4,10,10a-tetrahydro-2H-phenanthrene-1-carboxylic acid | 313.207 [M+H]+ | C20H26O3 |
Bathymodiolus
Kingdom: Animalia Phylum: Mollusca |
ChEBI |
| Self nodes | Griseofulvic acid | 361.027 [M+Na]+ | C16H15ClO6 |
Penicillium griseofulvum
Kingdom: Fungi Phylum: Ascomycota |
Petersen et al. (2014) |
| Cocamidopropyl betaine | 343.228 [M+H]+ | C19H37N2O3 | Synthetic | Jacob and Amini (2008) | |
| Chaetoglobosins A | (551.27 [M+Na]+ | C32H36N2O5 |
Penicillium chrysogenum
Kingdom: Fungi Phylum: Ascomycota |
Huang et al. (2016) |
3.4 Isolation and structural characterisation of the selected compound
The presence of loliolide in the extracts of P. boergesenii and F. indica was established by comparison of the NMR data (Suppl. Table S2) obtained for the isolated compound and literature data. Loliolide (12), a C11-monoterpenoid lactone, has previously been reported from plants and seaweeds, including genera Sargassum, Undaria, Padina and Codium (Pan et al. 2009; Percot et al. 2009; Tanaka and Matsunaga 1989). It is considered a photo-oxidative or thermal degradation product of carotenoids (Repeta 1989; Rios et al. 2008). In addition, it has reported antioxidant, anti-fungal, antibacterial and cytotoxic properties (Dias et al. 2020; Li et al. 2020; Silva et al. 2021).
3.5 Determination of antiviral activity using molecular docking approach
Given the current topicality of MPOX and the lack of potential treatments of this virus disease, we conducted a molecular docking approach to predict the antiviral activity of five compounds (6, 9, 10, 11 and 12) indicated in the Results (Sections 3.2 and 3.4). Cidofovir is used as standard which is widely used as antiviral. In the docking study, three proteins were taken to perform the experiments. Poxviruses tend to replicate in the cytoplasm of the host cells as in the case of DNA viruses. The genome replication as well as transcription are processed by them with the help of encoding enzymes, including their own thymidine and thymidylate kinases (Fenner 1990). PDB: 2V54 (Caillat et al. 2008) is the first protein used for docking from the vaccinia virus (Vacc-TMPK) with the functional enzyme thymidylate kinase. PDB: 4QWO (Minasov et al. 2022) A42R profilin-like protein is the second protein used for docking from monkeypox virus Zaire-96-I-16. D13, PDB: 6BED (Garriga et al. 2018) is the third protein used for docking from the Vaccinia virus.
Docking poses were analyzed and compared to the standard Cidofovir. The five compounds (6, 9, 10, 11 and 12) were subjected to docking analysis and the specificities of their interaction with the three targets, as shown in Figures 4 and 5), were investigated. The convergence was done to evaluate all docked results. The best-docked complexes were obtained based on binding energies and interacting residues. Docking poses were compared and analysed with the standard. In all three molecular docking studies (Table 3, Figure 5), compound (11) was the best lead-docked structure.
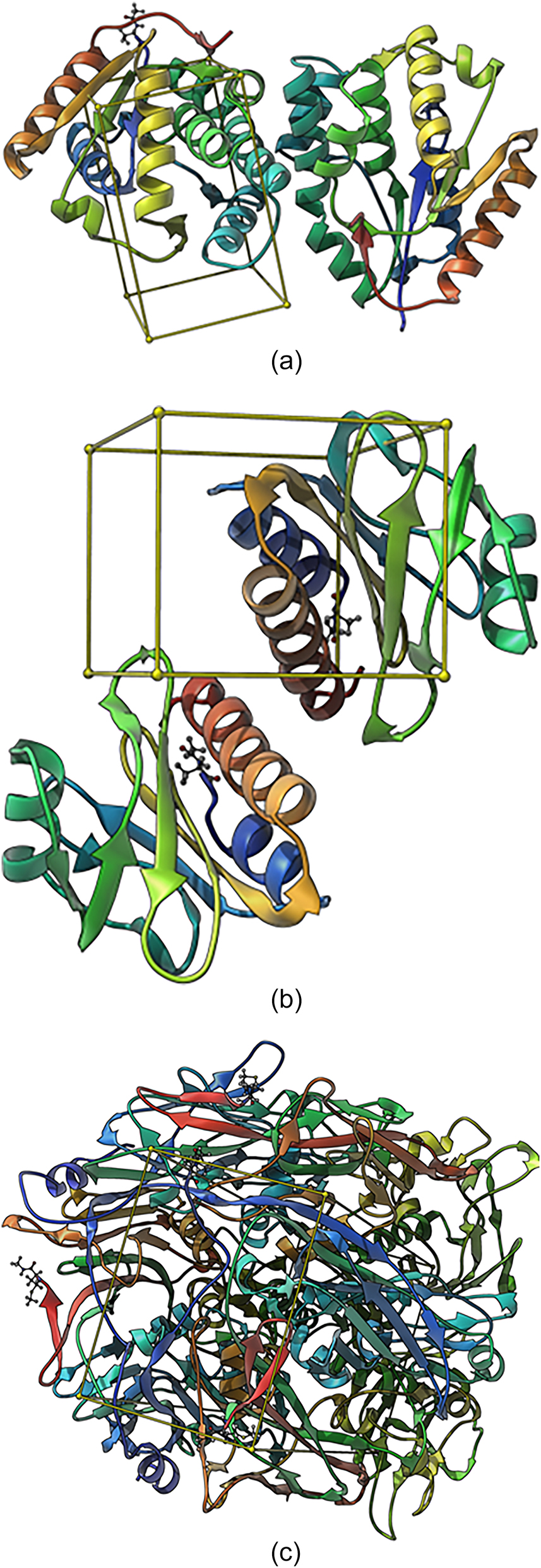
(A) Binding site (Box) of thymidylate kinase of the vaccinia virus; (B) binding site (Box) of A42R profilin-like protein from the monkeypox virus Zaire-96-I-16; (C) binding site (Box) of D13 protein of the vaccinia virus.
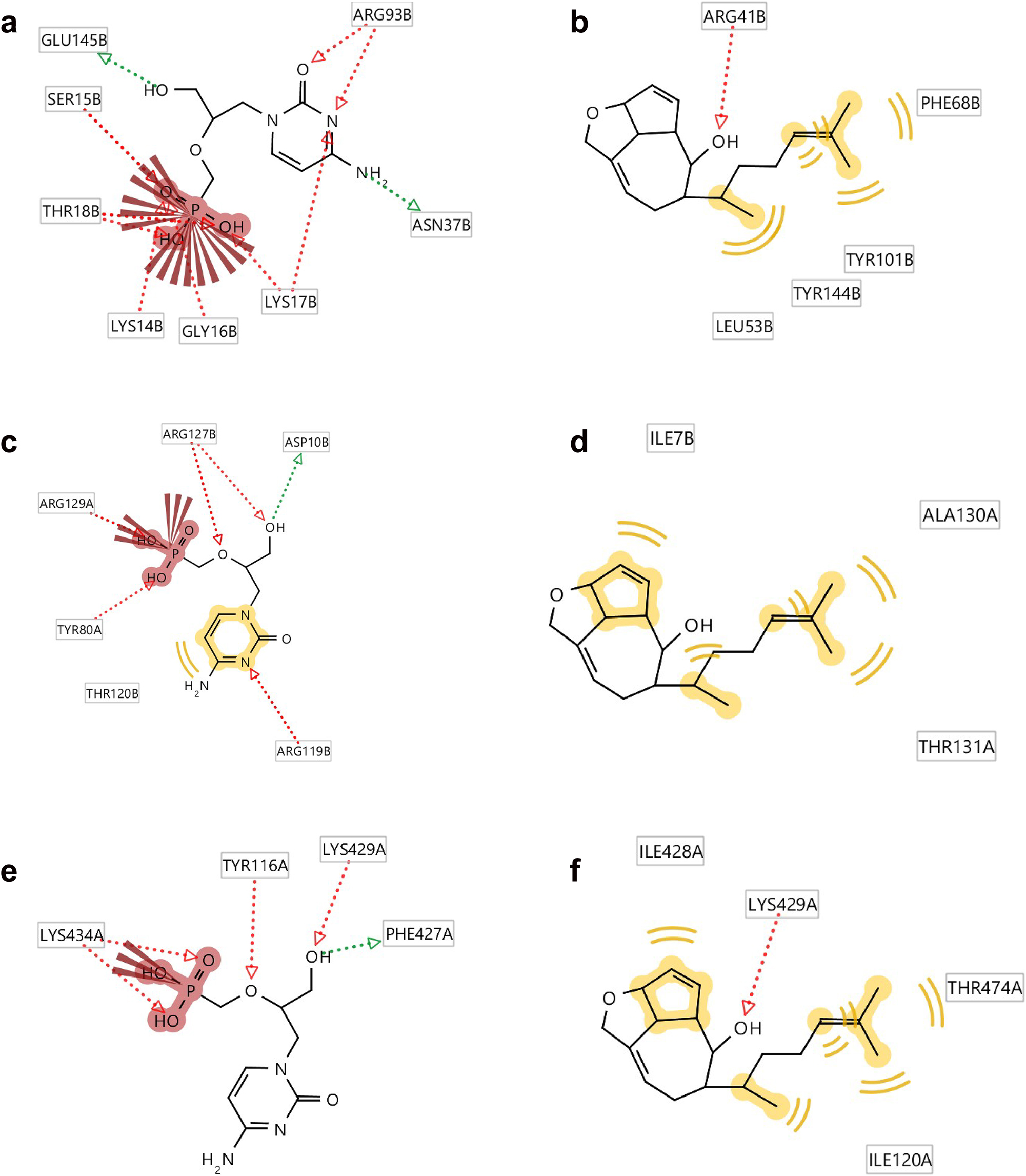
Ligplots showing the interacting amino acid residues of the functional enzyme thymidylate kinase, A42R profilin and D13 protein with (a) standard (b) compound 11, (c) standard (d) compound 11 and (e) standard (f) compound 11. Red arrows and broken lines, hydrogen bond acceptors; green arrows and broken lines, hydrogen bond donors; yellow lines, H (hydrogen bond) interactions, negative ionizable area (NI) interaction. Brick red represents amino acids, e.g. ARG93B as arginine, where 93 is the number given to the amino acid of the protein amino acid sequence and B represents the chain.
Docking analysis of five structures on three different protein receptors with respect to standard (Cidofovir) in kcal mol−1; bold type indicates best lead compound.
| Compounds | Monkeypox virus proteins | ||
|---|---|---|---|
| PDB: 2V54 (docking score) (kcal mol−1) | PDB:4QWO (docking score) (kcal mol−1) | PDB:6BED (docking score) (kcal mol−1) | |
| CIDOFOVIR (Standard) | −7.2 | −6.3 | −6.5 |
| Compound_6 | −6.1 | −6 | −6.6 |
| Compound_9_divarinone | −5.8 | −7.1 | −6.9 |
| Compound_10_5R,8S,9S,12R,14S0-9- | −6 | −6.4 | −6.7 |
| Compound_11_dictyol_A | −8.1 | −6.6 | −7.1 |
| Compound_12_loliolide | −5.3 | −6 | −6.3 |
Ligplots in Figure 5 show that amino acids Glu145B and Asn37B of standard cidofovir are involved as the hydrogen bond donors whereas Lys14B, Ser15B, Gly16B, Thr18B, Lys17B and Arg39B are involved as the hydrogen bond acceptors. Only Lys17B is involved as negative ionisation area interaction with the standard. On the other hand, Phe68B, Tyr101B, Tyr144B as well as Leu53B are involved in hydrogen interactions and Arg41B as hydrogen bond acceptor with the compound (11).
Ligplots in Figure 5 also show that amino acids Tyr116A and Lys434A of standard cidofovir are involved as the hydrogen bond acceptors whereas Phe427A is involved as the hydrogen bond donor. Lys434A is involved as negative ionisation area interaction with the standard. On the other hand, Ile428A, Thr474A, and Ile120A are involved in hydrogen interactions with compound (11). Both the standard and compound (11) are showing hydrogen bond acceptor interaction with the Lys429A amino acid.
3.6 Pharmacophore evaluation
In order to further evaluate the potential of compounds identified here as drug development leads for treatment of MPOX virus infection, a pharmacophore model was generated using the lowest energy conformers of cidofovir as well as compounds (6), (9), (11), (10) and (12). The generated pharmacophore showed three key features: hydrogen bond acceptors (HBAs), hydrogen bond donors (HBDs) and hydrophobic interactions (H). The representative 3D and 2D pharmacophoric features of each compound are shown in Figure 6. Each compound contains individual pharmacophoric features, and from these individual characteristic pharmacophores, a merged pharmacophore with common features was generated (Figure 7). This common feature pharmacophore model with a score of 0.8395 from ten generated models showed certain features: one HBD, three HBAs, and three Hs.
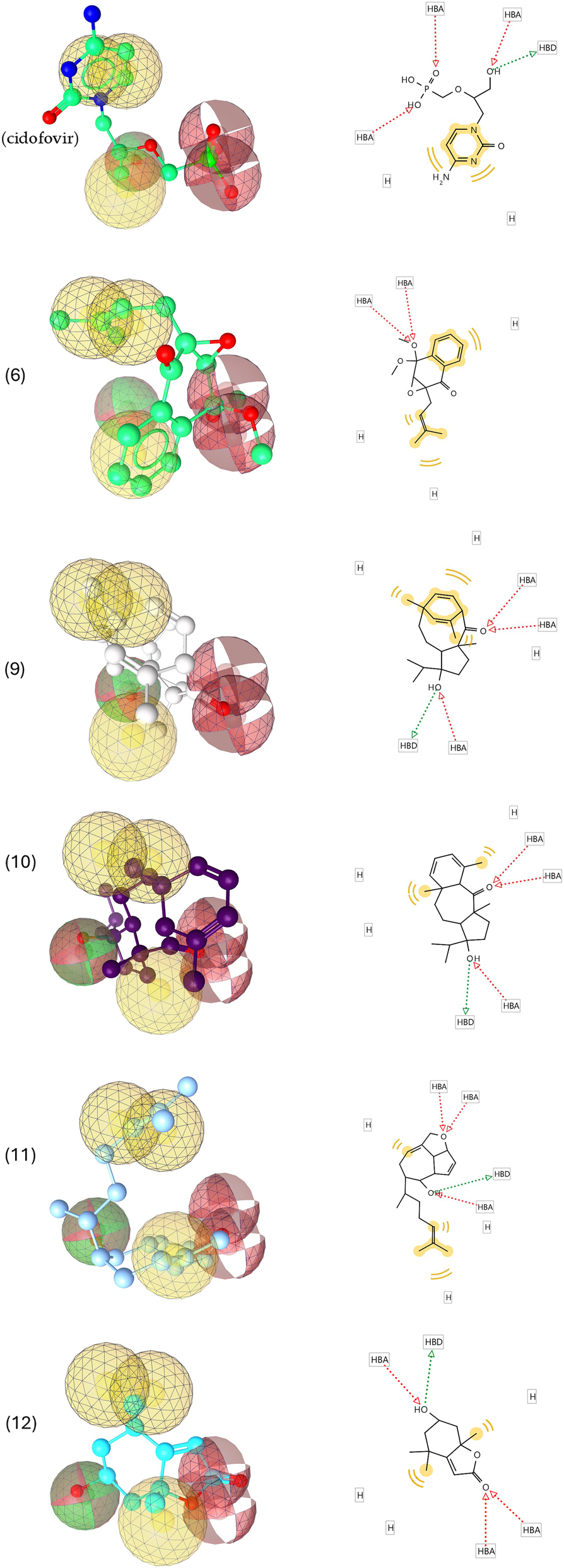
3D and 2D representations of pharmacophoric features of cidofovir, (6), (9), (10), (11) and (12), used in 3D pharmacophore generation. Red arrows and broken lines, hydrogen bond acceptors (HBA); green arrows and broken lines, hydrogen bond donors (HBD); yellow lines, hydrogen (H) bond interactions.
Common feature pharmacophore. Color codes are analogous with Figure 6. Red indicates hydrogen bond acceptors, green, hydrogen bond donators, and yellow, hydrogen bond interactions.
Overall, this study shows that several compounds from seaweeds of the Gulf have potential for developing drug treatments against MPOX, which should be explored further also with regard to other important virus diseases. It should be noted that dolysaccharides extracted from seaweeds, including carrageenan gel from C. crispus, have a demonstrated potential to impede the transmission of the HIV virus and other STD viruses such as gonorrhoea, genital warts, and the Herpes simplex Viruses (HSV) (Carlucci et al. 1997; Mahomoodally et al. 2019; Shi et al. 2017; Witvrouw and De Clercq 1997). Fucoidan, isolated from a different algal species, exhibits in vitro antiviral activity against various viruses, including HIV (Dinesh et al. 2016; Lee et al. 2013; Pagarete et al. 2021; Thuy et al. 2015), HSV (Krylova et al. 2020; Oliyaei et al. 2022; Rabanal et al. 2014), Influenza virus (Besednova et al. 2019; Sun et al. 2018), and murine norovirus (Kim et al. 2020). Furthermore, the green algae Ulva fasciata and Codium decorticatum have remarkable antiviral activity, recording the highest activity (99.9 %) against HSV-1. Triacylglycerols and fatty acids were identified as major components (Soares et al. 2012). The antiviral capacities of Ulvans from diverse Ulva species, such as U. compressa (Lopes et al. 2017), U. clathrata (Aguilar-Briseño et al. 2015), U. intestinalis (Morán-Santibañez et al. 2016), U. armoricana (Hardouin et al. 2016 ) and U. pertusa (Song et al. 2015 ), are widespread. These Ulvans exhibit antiviral activity against a variety of enveloped viruses, including HSV, Newcastle Disease Virus (NDV) (Lopes et al. 2017), Japanese Encephalitis Virus (JEV) (Chi et al. 2020), Dengue Virus (Pujol et al. 2012), Influenza (H1N1) (Jiao et al. 2012), Avian Influenza virus (Song et al. 2015), Vesicular Stomatitis Virus (Chi et al. 2020), and Measles Virus (Morán-Santibañez et al. 2016).
4 Conclusions
Seaweeds contain a variety of natural compounds, and a large variation occurs in the abundance of different compounds indicating the presence of species-specific metabolite patterns. Overall, the present study identified 27 compounds in 12 seaweed species from the territorial waters of Kuwait (with 11 compounds identified using the ChemFinder database, 15 compounds using the GNPs database, and 1 isolated compound). In general, but also in the present study, due to the limitations of mass spectrometry and dedicated seaweed databases for annotation and structure assignment, only a small fraction of the compounds can be identified. Furthermore, the GNPS platform was also not helpful for identifying metabolites in our study, annotating only one node, ursolic acid (as the seaweed metabolite hit) which was considered a “likely” hit based on taxonomic considerations, while a multitude of further automated annotations had to be rejected based on the same criteria. This indicated there is further scope to characterize ‘unannotated and tentatively’ molecular features using appropriate chemical standards. Further, loliolide was isolated and characterized from extracts of P. boergesenii and F. indica using classical separation and NMR spectrometry methods. Furthermore, in the computational study, we report that structure (11) may have the capacity to inhibit the mpox virus. The in silico screening of structures displayed interesting interactions with the binding site of previously reported proteins. These results are supported by a common pharmacophore model that could help guide future studies in identifying, selecting, and designing structures from big compound libraries or synthetically producing those that could work individually or synergistically with other drugs to treat mpox and other poxviruses. Finally, additional studies comprising in silico and in vitro studies related to stability testing could effectively improve future mpox virus inhibitor designs.
Funding source: Marine Alliance for Science and Technology Scotland
Award Identifier / Grant number: HR09011
Funding source: Kuwait Institute for Scientific Research
About the authors

Hanan Al-Adilah is an associate research scientist at Kuwait Institute for Scientific Research. She has a BSc in plant biology from Kuwait University (2011). She obtained her Master’s degree from Kuwait University with distinction, in the field of marine phytoplankton diversity in 2015. She has a PhD in algal diversity and biochemistry at the University of Aberdeen. Research in her lab includes environmental algal biodiversity, algal taxonomy, biochemistry of seagrasses and algae.

Dr. Gagan Preet has been a Research Fellow at the Marine Biodiscovery Center, University of Aberdeen, UK, since 2019. His expertise lies in the study of marine natural products, with a deep foundation in organic chemistry and computational chemistry applied to drug design and discovery. His research focuses on exploring marine organisms for potential pharmaceutical applications. With years of experience, he has contributed significantly to advancing knowledge in the field of marine medicinal chemistry and drug discovery.

The group of Prof Rainer Ebel at the Marine Biodiscovery Centre, University of Aberdeen focuses on the natural product chemistry of marine and terrestrial microorganisms and marine invertebrates, with special emphasis on endophytes from both marine and terrestrial sources. He is experienced in speciation of small molecules, i.e. structure elucidation of complex natural products using state-of-the-art hyphenated techniques including LC-MS as well as modern NMR experiments.

Puja Kumari is a lecturer in Macroalgal Research at the Scottish Association for Marine Science. She is a former recipient of ARO (Israel), BIDR-Blaustein (Israel) and JSPS (Japan) and Marie Curie (MSCA-IF) postdoctoral fellowships. Her research focus is to understand how different algae interact with its abiotic and biotic environment using biochemical, molecular and applied omics approaches.

Frithjof Christian Küpper has held the Chair in Marine Biodiversity at the University of Aberdeen since 2011, after 8 years at the Scottish Association for Marine Science, studying the biodiversity and biochemistry of marine plants/algae. He found that iodide serves as an inorganic antioxidant in kelp, the first described from a living system, impacting atmospheric and marine chemistry. A certified scientific diver, Frithjof has worked in the Mediterranean, South Atlantic (Ascension, Falklands), Antarctica, the Arctic and the Gulf.
-
Research ethics: This work is fully compliant with the ethical standards of the University of Aberdeen and San Diego State University.
-
Informed consent: Not applicable.
-
Author contributions: Conceptualisation, H.A-A and F.C.K.; methodology, H.A-A.; G.P.; P.K. and E.T.O.; software, H.A-A. , G.P., R. V. A.; formal analysis, H.A-A., G.P. and R.V.A; investigation, H.A-A.; resources, F.C.K., R.E., M.J. ; data curation, H.A-A., P.K., G.P.; writing- H.A-A., G.P and P.K.; writing – review and editing, all authors; visualization, H.A-A., G.P and P.K.; supervision, F.C.K., M.J., R.E. and P.K.; project administration, F.C.K.; funding acquisition, H.A-A. and F.C.K. The authors have accepted responsibility for the entire content of this manuscript and approved its submission.
-
Use of Large Language Models, AI and Machine Learning Tools: None declared
-
Conflict of interest: The authors declare no conflict of interest.
-
Research funding: Kuwait Institute for Scientific Research & Marine Alliance for Science and Technology Scotland (HR09011).
-
Data availability: All data are available from the corresponding author (FCK) upon request.
References
Additives, E.P.O.F., Food, N.S.A.T., Mortensen, A., Aguilar, F., Crebelli, R., Di Domenico, A., Dusemund, B., Frutos, M.J., Galtier, P., Gott, D., et al.. (2017). Re-evaluation of sorbitan monostearate (E 491), sorbitan tristearate (E 492), sorbitan monolaurate (E 493), sorbitan monooleate (E 494) and sorbitan monopalmitate (E 495) when used as food additives. EFSA J. 15: e04788, https://doi.org/10.2903/j.efsa.2017.4788.Search in Google Scholar PubMed PubMed Central
Agarwal, S., Singh, V., and Chauhan, K. (2023). Antidiabetic potential of seaweed and their bioactive compounds: a review of developments in last decade. Crit. Rev. Food Sci. Nutr. 63: 5739–5770, https://doi.org/10.1080/10408398.2021.2024130.Search in Google Scholar PubMed
Aguilar-Briseño, J.A., Cruz-Suarez, L.E., Sassi, J.-F., Ricque-Marie, D., Zapata-Benavides, P., Mendoza-Gamboa, E., Rodríguez-Padilla, C., and Trejo-Avila, L.M. (2015). Sulphated polysaccharides from Ulva clathrata and Cladosiphon okamuranus seaweeds both inhibit viral attachment/entry and cell-cell fusion. NDV Infection. Mar. Drugs 13: 697–712.10.3390/md13020697Search in Google Scholar PubMed PubMed Central
Ahmad, V.U., Aliya, R., Perveen, S., and Shameel, M. (1993). Sterols from marine green alga Codium decorticatum. Phytochemistry 33: 1189–1192, https://doi.org/10.1016/0031-9422(93)85047-u.Search in Google Scholar
Akramov, D.K., Bacher, M., Zengin, G., Bohmdorfer, S., Rosenau, T., Azimova, S.S., and Mamadalieva, N. (2019). Chemical composition and anticholinesterase activity of Lagochilus inebrians. Chem. Nat. Compd. 55: 575–577, https://doi.org/10.1007/s10600-019-02748-x.Search in Google Scholar
Al-Adilah, H., Peters, A.F., Al-Bader, D., Raab, A., Akhdhar, A., Feldmann, J., and Küpper, F.C. (2020). Iodine and fluorine concentrations in seaweeds of the Arabian Gulf identified by morphology and DNA barcodes. Bot. Mar. 63: 509–519, https://doi.org/10.1515/bot-2020-0049.Search in Google Scholar
Al-Adilah, H., Al-Bader, D., Elkotb, M., Kosma, I., Kumari, P., and Küpper, F.C. (2021a). Trace element concentrations in seaweeds of the Arabian Gulf identified by morphology and DNA barcodes. Bot. Mar. 64: 327–338, https://doi.org/10.1515/bot-2021-0027.Search in Google Scholar
Al-Adilah, H., Al-Sharrah, T.K., Al-Bader, D., Ebel, R., Küpper, F.C., and Kumari, P. (2021b). Assessment of Arabian Gulf seaweeds from Kuwait as sources of nutritionally important polyunsaturated fatty acids (PUFAs). Foods 10: 2442, https://doi.org/10.3390/foods10102442.Search in Google Scholar PubMed PubMed Central
Al-Adilah, H., Feiters, M.C., Carpenter, L.J., Kumari, P., Carrano, C.J., Al-Bader, D., and Küpper, F.C. (2022). Halogens in seaweeds: biological and environmental significance. Phycology 2: 132–171, https://doi.org/10.3390/phycology2010009.Search in Google Scholar
Al-Salman, H. and Shaker, A. (2017). Estimation of cortisone acetate in pharmaceutical anti-inflammatory drugs by HPLC-UV technique. Int. J. Sci. Res. 6: 2319–7064.Search in Google Scholar
Antunes, F. and Virgolino, A. (2022). Monkeypox mysteries of the new outbreak in non-endemic areas. Int. J. Environ. Res. Public Health 19, https://doi.org/10.3390/ijerph192214881.Search in Google Scholar PubMed PubMed Central
Beniddir, M.A., Kang, K.B., Genta-Jouve, G., Huber, F., Rogers, S., and Van Der Hooft, J.J. (2021). Advances in decomposing complex metabolite mixtures using substructure- and network-based computational metabolomics approaches. Nat. Prod. Rep. 38: 1967–1993, https://doi.org/10.1039/d1np00023c.Search in Google Scholar PubMed PubMed Central
Besednova, N., Zaporozhets, T., Kuznetsova, T., Makarenkova, I., Fedyanina, L., Kryzhanovsky, S., Malyarenko, O., and Ermakova, S. (2019). Metabolites of seaweeds as potential agents for the prevention and therapy of influenza infection. Mar. Drugs 17: 373, https://doi.org/10.3390/md17060373.Search in Google Scholar PubMed PubMed Central
Beutler, J.A., Alvarado, A.B., Schaufelberger, D.E., Andrews, P., and Mccloud, T.G. (1990). Dereplication of phorbol bioactives: Lyngbya majuscula and Croton cuneatus. J. Nat. Prod. 53: 867–874, https://doi.org/10.1021/np50070a014.Search in Google Scholar PubMed
Bian, B. and Van Altena, I.A. (1998). Four new compounds from the Australian brown alga Cystophora brownii. Aust. J. Chem. 51: 1157–1166, https://doi.org/10.1071/c98008.Search in Google Scholar
Bisanzio, D. and Reithinger, R. (2022). Projected burden and duration of the 2022 monkeypox outbreaks in non-endemic countries. Lancet Microbe 3: e643, https://doi.org/10.1016/s2666-5247(22)00183-5.Search in Google Scholar
Bonde, C.S., Bornancin, L., Lu, Y., Simonsen, H.T., Martínez-Valladares, M., Peña-Espinoza, M., Mejer, H., Williams, A.R., and Thamsborg, S.M. (2021). Bio-guided fractionation and molecular networking reveal fatty acids to be principal anti-parasitic compounds in nordic seaweeds. Front. Pharmacol. 12: 674520, https://doi.org/10.3389/fphar.2021.674520.Search in Google Scholar PubMed PubMed Central
Bouaicha, N., Pesando, D., Puel, D., and Tringali, C. (1993). Cytotoxic diterpenoids from the brown alga Dilophus ligulatus. J. Nat. Prod. 56: 1747–1752, https://doi.org/10.1021/np50100a013.Search in Google Scholar PubMed
Buedenbender, L., Astone, F.A., and Tasdemir, D. (2020). Bioactive molecular networking for mapping the antimicrobial constituents of the Baltic brown alga Fucus vesiculosus. Mar. Drugs 18: 311, https://doi.org/10.3390/md18060311.Search in Google Scholar PubMed PubMed Central
Caillat, C., Topalis, D., Agrofoglio, L.A., Pochet, S., Balzarini, J., Deville-Bonne, D., and Meyer, P. (2008). Crystal structure of poxvirus thymidylate kinase: an unexpected dimerization has implications for antiviral therapy. Proc. Natl. Acad. Sci. U.S.A. 105: 16900–16905, https://doi.org/10.1073/pnas.0804525105.Search in Google Scholar PubMed PubMed Central
Carlucci, M.J., Pujol, C.A., Ciancia, M., Noseda, M.D., Matulewicz, M.C., Damonte, E.B., and Cerezo, A.S. (1997). Antiherpetic and anticoagulant properties of carrageenans from the red seaweed Gigartina skottsbergii and their cyclized derivatives: correlation between structure and biological activity. Int. J. Biol. Macromol. 20: 97–105, https://doi.org/10.1016/s0141-8130(96)01145-2.Search in Google Scholar PubMed
Carriot, N., Paix, B., Greff, S., Viguier, B., Briand, J.-F., and Culioli, G. (2021). Integration of LC/MS-based molecular networking and classical phytochemical approach allows in-depth annotation of the metabolome of non-model organisms. The case study of the brown seaweed Taonia atomaria. Talanta 225: 121925, https://doi.org/10.1016/j.talanta.2020.121925.Search in Google Scholar PubMed
Chambers, M.C., Maclean, B., Burke, R., Amodei, D., Ruderman, D.L., Neumann, S., Gatto, L., Fischer, B., Pratt, B., Egertson, J., et al.. (2012). A cross-platform toolkit for mass spectrometry and proteomics. Nat. Biotechnol. 30: 918–920, https://doi.org/10.1038/nbt.2377.Search in Google Scholar PubMed PubMed Central
Chi, Y., Zhang, M., Wang, X., Fu, X., Guan, H., and Wang, P. (2020). Ulvan lyase assisted structural characterization of ulvan from Ulva pertusa and its antiviral activity against vesicular stomatitis virus. Int. J. Biol. Macromol. 157: 75–82, https://doi.org/10.1016/j.ijbiomac.2020.04.187.Search in Google Scholar PubMed
Choi, H.G., Kim, T.H., Kim, S.-H., and Kim, J.A. (2016). Anti-allergic inflammatory triterpenoids isolated from the spikes of Prunella vulgaris. Nat. Prod. Commun. 11, https://doi.org/10.1177/1934578x1601100111.Search in Google Scholar
Cline, M.S., Smoot, M., Cerami, E., Kuchinsky, A., Landys, N., Workman, C., Christmas, R., Avila-Campilo, I., Creech, M., Gross, B., et al.. (2007). Integration of biological networks and gene expression data using Cytoscape. Nat. Protoc. 2: 2366, https://doi.org/10.1038/nprot.2007.324.Search in Google Scholar PubMed PubMed Central
Costa-Lotufo, L.V., Wilke, D.V., Jimenez, P.C., and Epifanio, R.D.A. (2009). Organismos marinhos como fonte de novos fármacos: histórico & perspectivas. Química Nova 32: 703–716.10.1590/S0100-40422009000300014Search in Google Scholar
Da Silva, R.R., Dorrestein, P.C., and Quinn, R.A. (2015). Illuminating the dark matter in metabolomics. Proc. Natl. Acad. Sci. U.S.A. 112: 12549–12550, https://doi.org/10.1073/pnas.1516878112.Search in Google Scholar PubMed PubMed Central
Dias, D.A., Urban, S., and Roessner, U. (2012). A historical overview of natural products in drug discovery. Metabolites 2: 303–336, https://doi.org/10.3390/metabo2020303.Search in Google Scholar PubMed PubMed Central
Dias, M.K.H.M., Madusanka, D.M.D., Han, E.J., Kim, M.J., Jeon, Y.-J., Kim, H.-S., Fernando, I.P.S., and Ahn, G. (2020). (−)-Loliolide isolated from Sargassum horneri protects against fine dust-induced oxidative stress in human keratinocytes. Antioxidants 9: 474, https://doi.org/10.3390/antiox9060474.Search in Google Scholar PubMed PubMed Central
Dinesh, S., Menon, T., Hanna, L.E., Suresh, V., Sathuvan, M., and Manikannan, M. (2016). In vitro anti-HIV-1 activity of fucoidan from Sargassum swartzii. Int. J. Biol. Macromol. 82: 83–88, https://doi.org/10.1016/j.ijbiomac.2015.09.078.Search in Google Scholar PubMed
Eldeen, I., Van Heerden, F., and Van Staden, J. (2007). Biological activities of cycloart-23-ene-3, 25-diol isolated from the leaves of Trichilia dregeana. S. Afr. J. Bot. 73: 366–371, https://doi.org/10.1016/j.sajb.2007.02.192.Search in Google Scholar
Ernst, M., Kang, K.B., Caraballo-Rodríguez, A.M., Nothias, L.-F., Wandy, J., Chen, C., Wang, M., Rogers, S., Medema, M.H., Dorrestein, P.C., et al.. (2019). MolNetEnhancer: enhanced molecular networks by integrating metabolome mining and annotation tools. Metabolites 9: 144, https://doi.org/10.3390/metabo9070144.Search in Google Scholar PubMed PubMed Central
Fenner, F. (1990) Poxviruses. In: Fields, B.N., Chanock, R.M., Hirsch, M.S., Melnick, J., Monath, T.P., and Roizman, B. (Eds.). Virology. Raven Press, New York, USA, pp. 2113–2133.Search in Google Scholar
Fattorusso, E., Magno, S., Mayol, L., Santacroce, C., Sica, D., Amico, V., Oriente, G., Piattelli, M., and Tringali, C. (1976). Dictyol A and B, two novel diterpene alcohols from the brown alga Dictyota dichotoma. J. Chem. Soc. Chem. Commun. 14: 575–576, https://doi.org/10.1039/c39760000575.Search in Google Scholar
Fuller, R.W., Cardellina, J.H., Jurek, J., Scheuer, P.J., Alvarado-Lindner, B., Mcguire, M., Gray, G.N., Steiner, J.R., and Clardy, J. (1994). Isolation and structure/activity features of halomon-related antitumor monoterpenes from the red alga Portieria hornemannii. J. Med. Chem. 37: 4407–4411, https://doi.org/10.1021/jm00051a019.Search in Google Scholar PubMed
Garriga, D., Headey, S., Accurso, C., Gunzburg, M., Scanlon, M., and Coulibaly, F. (2018). Structural basis for the inhibition of poxvirus assembly by the antibiotic rifampicin. Proc. Natl. Acad. Sci. U. S. A. 115: 8424–8429, https://doi.org/10.1073/pnas.1810398115.Search in Google Scholar PubMed PubMed Central
Gnanavel, V., Roopan, S.M., and Rajeshkumar, S. (2019). Aquaculture: an overview of chemical ecology of seaweeds (food species) in natural products. Aquaculture 507: 1–6, https://doi.org/10.1016/j.aquaculture.2019.04.004.Search in Google Scholar
Gunathilaka, T.L., Samarakoon, K., Ranasinghe, P., and Peiris, L.D.C. (2020). Antidiabetic potential of marine brown algae: a mini review. J. Diabetes Res. 1: 1230218, https://doi.org/10.1155/2020/1230218.Search in Google Scholar PubMed PubMed Central
Guthals, A., Watrous, J.D., Dorrestein, P.C., and Bandeira, N. (2012). The spectral networks paradigm in high throughput mass spectrometry. Mol. BioSyst. 8: 2535–2544, https://doi.org/10.1039/c2mb25085c.Search in Google Scholar PubMed PubMed Central
Hardouin, K., Bedoux, G., Burlot, A.-S., Donnay-Moreno, C., Bergé, J.-P., Nyvall-Collén, P., and Bourgougnon, N. (2016). Enzyme-assisted extraction (EAE) for the production of antiviral and antioxidant extracts from the green seaweed Ulva armoricana (Ulvales, Ulvophyceae). Algal Res. 16: 233–239, https://doi.org/10.1016/j.algal.2016.03.013.Search in Google Scholar
Hasan, A.H.H., Al-Bader, D.A., Peters, A.F., and Küpper, F.C. (2023). Revision of the family Sargassaceae (Fucales, Phaeophyceae) from Kuwait (Arabian/Persian Gulf, NW Indian ocean) based on analysis of morphology and ITS2 sequences. Ukr. Bot. J. 80: 3–20, https://doi.org/10.15407/ukrbotj80.01.003.Search in Google Scholar
He, H.P., Shen, Y.M., Zuo, G.Y., Yang, X.S., and Hao, X.J. (2003). Dinorditerpene, diterpenes, alkaloids, and coumarins from Clausena dunniana. Helv. Chim. Acta 86: 3187–3193, https://doi.org/10.1002/hlca.200390259.Search in Google Scholar
He, Z., Zhang, A., Ding, L., Lei, X., Sun, J., and Zhang, L. (2010). Chemical composition of the green alga Codium divaricatum Holmes. Fitoterapia 81: 1125–1128, https://doi.org/10.1016/j.fitote.2010.07.011.Search in Google Scholar PubMed
Hilbert, G., Temsamani, H., Bordenave, L., Pedrot, E., Chaher, N., Cluzet, S., Delaunay, J.-C., Ollat, N., Delrot, S., Mérillon, J.-M., et al.. (2015). Flavonol profiles in berries of wild Vitis accessions using liquid chromatography coupled to mass spectrometry and nuclear magnetic resonance spectrometry. Food Chem. 169: 49–58, https://doi.org/10.1016/j.foodchem.2014.07.079.Search in Google Scholar PubMed
Holman, J.D., Tabb, D.L., and Mallick, P. (2014). Employing ProteoWizard to convert raw mass spectrometry data. Curr. Protoc. Bioinf. 46: 13.24. 1–13.24. 9, https://doi.org/10.1002/0471250953.bi1324s46.Search in Google Scholar PubMed PubMed Central
Hoy, S.M. (2018). Tecovirimat: first global approval. Drugs 78: 1377–1382, https://doi.org/10.1007/s40265-018-0967-6.Search in Google Scholar PubMed
Huang, S., Chen, H., Li, W., Zhu, X., Ding, W., and Li, C. (2016). Bioactive chaetoglobosins from the mangrove endophytic fungus Penicillium chrysogenum. Mar. Drugs 14: 172, https://doi.org/10.3390/md14100172.Search in Google Scholar PubMed PubMed Central
Indergaard, M. (1983) The aquatic resource. In: Côté, W.A. (Ed.). Biomass utilization. NATO advanced science institutes series, Vol. 67. Springer, Boston, MA, pp. 137–168.10.1007/978-1-4757-0833-2_8Search in Google Scholar
Jacob, S.E. and Amini, S. (2008). Cocamidopropyl betaine. DERM 19: 157–160, https://doi.org/10.2310/6620.2008.06043.Search in Google Scholar
Jacobsen, C., Sørensen, A.-D.M., Holdt, S.L., Akoh, C.C., and Hermund, D.B. (2019). Source, extraction, characterization, and applications of novel antioxidants from seaweed. Annu. Rev. Food Sci. Technol. 10: 541–568, https://doi.org/10.1146/annurev-food-032818-121401.Search in Google Scholar PubMed
Jiao, G., Yu, G., Wang, W., Zhao, X., Zhang, J., and Ewart, S.H. (2012). Properties of polysaccharides in several seaweeds from Atlantic Canada and their potential anti-influenza viral activities. J. Ocean Univ. China 11: 205–212, https://doi.org/10.1007/s11802-012-1906-x.Search in Google Scholar
Khotimchenko, M., Tiasto, V., Kalitnik, A., Begun, M., Khotimchenko, R., Leonteva, E., Bryukhovetskiy, I., and Khotimchenko, Y. (2020). Antitumor potential of carrageenans from marine red algae. Carbohydr. Polym. 246: 116568, https://doi.org/10.1016/j.carbpol.2020.116568.Search in Google Scholar PubMed
Kim, H., Lim, C.Y., Lee, D.B., Seok, J.H., Kim, K.H., and Chung, M.S. (2020). Inhibitory effects of Laminaria japonica fucoidans against noroviruses. Viruses 12: 997, https://doi.org/10.3390/v12090997.Search in Google Scholar PubMed PubMed Central
Klassen, A., Faccio, A.T., Canuto, G.A., da Cruz, P.L., Ribeiro, H.C., Tavares, M.F., and Sussulini, A. (2017). Metabolomics: definitions and significance in systems biology. Adv. Exp. Med. Biol. 965: 3–17, https://doi.org/10.1007/978-3-319-47656-8_1.Search in Google Scholar PubMed
Krylova, N.V., Ermakova, S.P., Lavrov, V.F., Leneva, I.A., Kompanets, G.G., Iunikhina, O.V., Nosik, M.N., Ebralidze, L.K., Falynskova, I.N., Silchenko, A.S., et al.. (2020). The comparative analysis of antiviral activity of native and modified fucoidans from brown algae Fucus evanescens in vitro and in vivo. Mar. Drugs 18: 224, https://doi.org/10.3390/md18040224.Search in Google Scholar PubMed PubMed Central
Kupchan, S.M., Britton, R.W., Ziegler, M.F., and Sigel, C.W. (1973). Bruceantin, a new potent antileukemic simaroubolide from Brucea antidysenterica. J. Org. Chem. 38: 178–179, https://doi.org/10.1021/jo00941a049.Search in Google Scholar PubMed
Laatsch, H. (2017). AntiBase: the natural compound identifier. Wiley-Vch, Weinheim, Germany.Search in Google Scholar
Laskowski, R.A. and Swindells, M.B. (2011). LigPlot+: multiple ligand–protein interaction diagrams for drug discovery. J. Chem. Inf. Model. 51: 2778–2786.10.1021/ci200227uSearch in Google Scholar PubMed
Lee, H.-G., Lu, Y.A., Li, X., Hyun, J.-M., Kim, H.-S., Lee, J.J., Kim, T.H., Kim, H.M., Kang, M.-C., and Jeon, Y.-J. (2020). Anti-obesity effects of Grateloupia elliptica, a red seaweed, in mice with high-fat diet-induced obesity via suppression of adipogenic factors in white adipose tissue and increased thermogenic factors in brown adipose tissue. Nutrients 12: 308, https://doi.org/10.3390/nu12020308.Search in Google Scholar PubMed PubMed Central
Lee, J.-C., Hou, M.-F., Huang, H.-W., Chang, F.-R., Yeh, C.-C., Tang, J.-Y., and Chang, H.-W. (2013). Marine algal natural products with anti-oxidative, anti-inflammatory, and anti-cancer properties. Cancer Cell Int. 13: 1–7, https://doi.org/10.1186/1475-2867-13-55.Search in Google Scholar PubMed PubMed Central
Li, L.-L., Zhao, H.-H., and Kong, C.-H. (2020). (–)-Loliolide, the most ubiquitous lactone, is involved in barnyardgrass-induced rice allelopathy. J. Exp. Bot. 71: 1540–1550, https://doi.org/10.1093/jxb/erz497.Search in Google Scholar PubMed
Lim, G.-E., Sung, J.Y., Yu, S., Kim, Y., Shim, J., Kim, H.J., Cho, M.L., Lee, J.-S., and Kim, Y.-N. (2020). Pygenic acid A (PA) sensitizes metastatic breast cancer cells to anoikis and inhibits metastasis in vivo. Int. J. Mol. Sci. 21: 8444, https://doi.org/10.3390/ijms21228444.Search in Google Scholar PubMed PubMed Central
Liu, Z., Gao, T., Yang, Y., Meng, F., Zhan, F., Jiang, Q., and Sun, X. (2019). Anti-cancer activity of porphyran and carrageenan from red seaweeds. Molecules 24: 4286, https://doi.org/10.3390/molecules24234286.Search in Google Scholar PubMed PubMed Central
Lopes, N., Ray, S., Espada, S.F., Bomfim, W.A., Ray, B., Faccin-Galhardi, L.C., Linhares, R.E.C., and Nozawa, C. (2017). Green seaweed Enteromorpha compressa (Chlorophyta, Ulvaceae) derived sulphated polysaccharides inhibit herpes simplex virus. Int. J. Biol. Macromol. 102: 605–612, https://doi.org/10.1016/j.ijbiomac.2017.04.043.Search in Google Scholar PubMed
Mahomoodally, M.F., Lobine, D., Rengasamy, K.R., Gowrishankar, S., Tewari, D., Zengin, G., Kim, D.H., and Sivanesan, I. (2019). Marine algae: a potential resource of anti-HSV molecules. Processes 7: 887, https://doi.org/10.3390/pr7120887.Search in Google Scholar
Martins, A., Vieira, H., Gaspar, H., and Santos, S. (2014). Marketed marine natural products in the pharmaceutical and cosmeceutical industries: tips for success. Mar. Drugs 12: 1066–1101, https://doi.org/10.3390/md12021066.Search in Google Scholar PubMed PubMed Central
Miao, P., Cai, D.-C., Xiang, B.-R., An, D.-K., and Ito, Y. (1998). Separation and purification of strychnine from crude extract of Strychnos nux-vomica L. by high-speed countercurrent chromatography. J. Liq. Chromatogr. Rel. Technol. 21: 163–170, https://doi.org/10.1080/10826079808001945.Search in Google Scholar
Miliauskas, G., Van Beek, T.A., De Waard, P., Venskutonis, R.P., and Sudhölter, E.J. (2006). Comparison of analytical and semi-preparative columns for high-performance liquid chromatography–solid-phase extraction–nuclear magnetic resonance. J. Chromatogr. 1112: 276–284, https://doi.org/10.1016/j.chroma.2005.11.059.Search in Google Scholar PubMed
Minasov, G., Inniss, N.L., Shuvalova, L., Anderson, W.F., and Satchell, K.J. (2022). Structure of the monkeypox virus profilin-like protein A42R reveals potential functional differences from cellular profilins. Acta Crystallogr., Sect. F: Struct. Biol. Commun. 78: 371–377, https://doi.org/10.1107/s2053230x22009128.Search in Google Scholar PubMed PubMed Central
Morán-Santibañez, K., Cruz-Suárez, L.E., Ricque-Marie, D., Robledo, D., Freile-Pelegrín, Y., Peña-Hernández, M.A., Rodríguez-Padilla, C., and Trejo-Avila, L.M. (2016). Synergistic effects of sulfated polysaccharides from Mexican seaweeds against measles virus. BioMed Res. Intl. 1: 8502123.10.1155/2016/8502123Search in Google Scholar PubMed PubMed Central
Nagappayya, S.K. and Gaikar, V.G. (2010). Extraction of aleuritic acid from seedlac and purification by reactive adsorption on functionalized polymers. Ind. Eng. Chem. Res. 49: 6547–6553, https://doi.org/10.1021/ie902061e.Search in Google Scholar
Nguyen, T.H., Nguyen, T.L.P., Tran, T.V.A., Do, A.D., Kim, S.M., and Kim, S. (2019). Antidiabetic and antioxidant activities of red seaweed Laurencia dendroidea. Asian Pac. J. Trop. Biomed. 9: 501–509, https://doi.org/10.4103/2221-1691.271723.Search in Google Scholar
Oliyaei, N., Moosavi-Nasab, M., and Mazloomi, S.M. (2022). Therapeutic activity of fucoidan and carrageenan as marine algal polysaccharides against viruses. 3 Biotech 12: 154, https://doi.org/10.1007/s13205-022-03210-6.Search in Google Scholar PubMed PubMed Central
Oppong-Danquah, E., Bluemel, M., and Tasdemir, D. (2023). Metabolomics and microbiomics insights into the differential surface fouling of three macroalgal species of Fucus (Fucales, Phaeophyceae) that co-exist in the German Baltic sea. Mar. Drugs. 21: 595.10.3390/md21110595Search in Google Scholar PubMed PubMed Central
Pagarete, A., Ramos, A.S., Puntervoll, P., Allen, M.J., and Verdelho, V. (2021). Antiviral potential of algal metabolites: a comprehensive review. Mar. Drugs 19: 94, https://doi.org/10.3390/md19020094.Search in Google Scholar PubMed PubMed Central
Pan, L., Sinden, M.R., Kennedy, A.H., Chai, H., Watson, L.E., Graham, T.L., and Kinghorn, A.D. (2009). Bioactive constituents of Helianthus tuberosus (Jerusalem artichoke). Phytochem. Lett. 2: 15–18, https://doi.org/10.1016/j.phytol.2008.10.003.Search in Google Scholar
Parrot, D., Blümel, M., Utermann, C., Chianese, G., Krause, S., Kovalev, A., Gorb, S.N., and Tasdemir, D. (2019). Mapping the surface microbiome and metabolome of brown seaweed Fucus vesiculosus by amplicon sequencing, integrated metabolomics and imaging techniques. Sci. Rep. 31: 1061, https://doi.org/10.1038/s41598-018-37914-8.Search in Google Scholar PubMed PubMed Central
Percot, A., Yalçın, A., Aysel, V., Erduğan, H., Dural, B., and Güven, K.C. (2009). Loliolide in marine algae. Nat. Prod. Res. 23: 460–465, https://doi.org/10.1080/14786410802076069.Search in Google Scholar PubMed
Pérez, M.J., Falqué, E., and Domínguez, H. (2016). Antimicrobial action of compounds from marine seaweed. Mar. Drugs 14: 52, https://doi.org/10.3390/md14030052.Search in Google Scholar PubMed PubMed Central
Perry, N.B., Blunt, J.W., and Munro, M.H. (1991). A cytotoxic and antifungal 1, 4-naphthoquinone and related compounds from a New Zealand brown alga, Landsburgia quercifolia. J. Nat. Prod. 54: 978–985, https://doi.org/10.1021/np50076a009.Search in Google Scholar PubMed
Petersen, A.B., Rønnest, M.H., Larsen, T.O., and Clausen, M.H. (2014). The chemistry of griseofulvin. Chem. Rev. 114: 12088–12107, https://doi.org/10.1021/cr400368e.Search in Google Scholar PubMed
Pettersen, E.F., Goddard, T.D., Huang, C.C., Couch, G.S., Greenblatt, D.M., Meng, E.C., and Ferrin, T.E. (2004). UCSF chimera: a visualization system for exploratory research and analysis. J. Comput. Chem. 25: 1605–1612, https://doi.org/10.1002/jcc.20084.Search in Google Scholar PubMed
Philippus, A.C., Zatelli, G.A., Wanke, T., Barros, M.G.D.A., Kami, S.A., Lhullier, C., Armstrong, L., Sandjo, L.P., and Falkenberg, M. (2018). Molecular networking prospection and characterization of terpenoids and C 15-acetogenins in Brazilian seaweed extracts. RSC Adv. 8: 29654–29661, https://doi.org/10.1039/c8ra02802h.Search in Google Scholar PubMed PubMed Central
Preet, G., Oluwabusola, E.T., Milne, B.F., Ebel, R., and Jaspars, M. (2022). Computational repurposing of mitoxantrone-related structures against monkeypox virus: a molecular docking and 3D pharmacophore study. Int. J. Mol. Sci. 23: 14287, https://doi.org/10.3390/ijms232214287.Search in Google Scholar PubMed PubMed Central
Pujol, C.A., Ray, S., Ray, B., and Damonte, E.B. (2012). Antiviral activity against dengue virus of diverse classes of algal sulfated polysaccharides. Int. J. Biol. Macromol. 51: 412–416, https://doi.org/10.1016/j.ijbiomac.2012.05.028.Search in Google Scholar PubMed
Quinn, R.A., Nothias, L.F., Vining, O., Meehan, M., Esquenazi, E., and Dorrestein, P.C. (2017). Molecular networking as a drug discovery, drug metabolism, and precision medicine strategy. Trends Pharmacol. Sci. 38: 143–154, https://doi.org/10.1016/j.tips.2016.10.011.Search in Google Scholar PubMed
Rabanal, M., Ponce, N.M., Navarro, D.A., Gómez, R.M., and Stortz, C.A. (2014). The system of fucoidans from the brown seaweed Dictyota dichotoma: chemical analysis and antiviral activity. Carbohydr. Polym. 101: 804–811, https://doi.org/10.1016/j.carbpol.2013.10.019.Search in Google Scholar PubMed
Rani, I., Satapathy, P., Goyal, A., Shamim, M.A., Pal, A., Squitti, R., Goswami, K., Pradhan, K.B., Rustagi, S., Hermis, A.H., et al.. (2023). Viral loads in skin samples of patients with monkeypox virus infection: a systematic review and meta-analysis. Viruses 15: 1386, https://doi.org/10.3390/v15061386.Search in Google Scholar PubMed PubMed Central
Raoof, G.F.A. and Mohamed, K.Y. (2018) Natural products for the management of diabetes. In: Rahman, A. (Ed.). Studies in natural products vhemistry, Vol. 59. Elsevier, Amsterdam, The Netherlands, pp. 323–374.10.1016/B978-0-444-64179-3.00010-4Search in Google Scholar
Repeta, D.J. (1989). Carotenoid diagenesis in recent marine sediments. II. Degradation of fucoxanthin to loliolide. Geochim. Cosmochim. Acta 53: 699–707, https://doi.org/10.1016/0016-7037(89)90012-4.Search in Google Scholar
Rios, J.J., Fernández-García, E., Mínguez-Mosquera, M.I., and Pérez-Gálvez, A. (2008). Description of volatile compounds generated by the degradation of carotenoids in paprika, tomato and marigold oleoresins. Food Chem. 106: 1145–1153.10.1016/j.foodchem.2007.07.045Search in Google Scholar
Rocha, D.H., Seca, A.M., and Pinto, D.C. (2018). Seaweed secondary metabolites in vitro and in vivo anticancer activity. Mar. Drugs 16: 410, https://doi.org/10.3390/md16110410.Search in Google Scholar PubMed PubMed Central
Sahu, A., Gaur, M., Mahanandia, N.C., Subudhi, E., Swain, R.P., and Subudhi, B.B. (2023). Identification of core therapeutic targets for monkeypox virus and repurposing potential of drugs against them: an in silico approach. Comput. Biol. Med. 161: 106971, https://doi.org/10.1016/j.compbiomed.2023.106971.Search in Google Scholar
Sanger, G., Rarung, L., Damongilala, L., Kaseger, B., and Montolalu, L. (2019) Phytochemical constituents and antidiabetic activity of edible marine red seaweed (Halymenia durvilae). In: IOP conference series: earth and environmental science. IOP Publishing, Bristol, p. 012069.10.1088/1755-1315/278/1/012069Search in Google Scholar
Sapkal, A. and Agrawal, S. (2022). Monkeypox: the re-emerging terror. Cureus 14: e28597, https://doi.org/10.7759/cureus.28597.Search in Google Scholar
Saraswat, B., Visen, P.K., and Agarwal, D.P. (2000). Ursolic acid isolated from Eucalyptus tereticornis protects against ethanol toxicity in isolated rat hepatocytes. Phytother. Res. 14: 163–166, https://doi.org/10.1002/(sici)1099-1573(200005)14:3<163::aid-ptr588>3.0.co;2-d.10.1002/(SICI)1099-1573(200005)14:3<163::AID-PTR588>3.0.CO;2-DSearch in Google Scholar
Seca, A.M. and Pinto, D.C. (2018). Overview on the antihypertensive and anti-obesity effects of secondary metabolites from seaweeds. Mar. Drugs 16: 237, https://doi.org/10.3390/md16070237.Search in Google Scholar
Shi, D., Fan, X., Sun, J., Han, L., and Shi, J. (2008). Steroids from green alga Chaetomorpha basiretorsa Setchell. Chin. J. Oceanol. Limnol. 26: 415–418, https://doi.org/10.1007/s00343-008-0415-7.Search in Google Scholar
Shi, Q., Wang, A., Lu, Z., Qin, C., Hu, J., and Yin, J. (2017). Overview on the antiviral activities and mechanisms of marine polysaccharides from seaweeds. Carbohydr. Res. 453: 1–9, https://doi.org/10.1016/j.carres.2017.10.020.Search in Google Scholar
Silva, J., Alves, C., Martins, A., Susano, P., Simões, M., Guedes, M., Rehfeldt, S., Pinteus, S., Gaspar, H., Rodrigues, A., et al.. (2021). Loliolide, a new therapeutic option for neurological diseases? In vitro neuroprotective and anti-inflammatory activities of a monoterpenoid lactone isolated from Codium tomentosum. Int. J. Mol. Sci. 22: 1888, https://doi.org/10.3390/ijms22041888.Search in Google Scholar
Singh, S., Srivastava, V., Godara, P., Banavath, H.N., Tak, H., Nayak, A., Kumari, D., Naik, B., and Prusty, D. (2023). An in-silico-based study identified peptide inhibitors that can block the egression of the monkeypox virus by inhibiting the p37 protein target. Peptide Sci. 115: e24325, https://doi.org/10.1002/pep2.24325.Search in Google Scholar
Soares, A.R., Robaina, M., Mendes, G.S., Silva, T.S., Gestinari, L., Pamplona, O.S., Yoneshigue-Valentin, Y., Kaiser, C.R., and Romanos, M.T.V. (2012). Antiviral activity of extracts from Brazilian seaweeds against herpes simplex virus. Revista Brasileira de Farmacognosia 22: 714–723, https://doi.org/10.1590/s0102-695x2012005000061.Search in Google Scholar
Song, L., Chen, X., Liu, X., Zhang, F., Hu, L., Yue, Y., Li, K., and Li, P. (2015). Characterization and comparison of the structural features, immune-modulatory and anti-avian influenza virus activities conferred by three algal sulfated polysaccharides. Mar. Drugs 14: 4, https://doi.org/10.3390/md14010004.Search in Google Scholar PubMed PubMed Central
Sun, J., Shi, D.Y., Li, S., Wang, S.J., Han, L.J., Fan, X., Yang, Y.C., and Shi, J.G. (2007). Chemical constituents of the red alga Laurencia tristicha. J. Asian Nat. Prod. Res. 9: 725–734, https://doi.org/10.1080/10286020601103338.Search in Google Scholar PubMed
Sun, T., Zhang, X., Miao, Y., Zhou, Y., Shi, J., Yan, M., and Chen, A. (2018). Studies on antiviral and immuno-regulation activity of low molecular weight fucoidan from Laminaria japonica. J. Ocean Univ. China 17: 705–711, https://doi.org/10.1007/s11802-018-3794-1.Search in Google Scholar
Tamayo-Castillo, G., Jakupovic, J., Castro, V., and King, R.M.J.P. (1989). (+)-α-copaen-8-one and other constituents from Neomirandea species. Phytochem. 28: 938–940, https://doi.org/10.1016/0031-9422(89)80149-9.Search in Google Scholar
Tan, H., Zhao, C., Zhu, Q., Katakura, Y., Tanaka, H., Ohnuki, K., and Shimizu, K. (2019). Ursolic acid isolated from the leaves of loquat (Eriobotrya japonica) inhibited osteoclast differentiation through targeting exportin 5. J. Agric. Food Chem. 67: 3333–3340, https://doi.org/10.1021/acs.jafc.8b06954.Search in Google Scholar PubMed
Tanaka, R. and Matsunaga, S. (1989). Loliolide and olean-12-en-3β, 9α, 11α-triol from Euphorbia supina. J. Agric. Food Chem. 28: 1699–1702, https://doi.org/10.1016/s0031-9422(00)97828-2.Search in Google Scholar
Tang, H.F., Yi, Y.H., Yao, X.S., Xu, Q.Z., Zhang, S.Y., and Lin, H.W. (2002). Bioactive steroids from the brown alga Sargassum carpophyllum. J. Asian Nat. Prod. Res. 4: 95–101, https://doi.org/10.1080/10286020290027362.Search in Google Scholar PubMed
Thuy, T.T.T., Ly, B.M., Van, T.T.T., Van Quang, N., Tu, H.C., Zheng, Y., Seguin-Devaux, C., Mi, B., and Ai, U. (2015). Anti-HIV activity of fucoidans from three brown seaweed species. Carbohydr. Polym. 115: 122–128, https://doi.org/10.1016/j.carbpol.2014.08.068.Search in Google Scholar PubMed
Trimurtulu, G., Kushlan, D.M., Faulkner, D.J., and Rao, C.B. (1992). Divarinone, a novel diterpene from the brown alga Dictyota divaricata of the Indian Ocean. Tetrahedron Lett. 33: 729–732, https://doi.org/10.1016/s0040-4039(00)77701-x.Search in Google Scholar
Trott, O. and Olson, A.J. (2010). AutoDock Vina: improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 31: 455–461, https://doi.org/10.1002/jcc.21334.Search in Google Scholar PubMed PubMed Central
Wan-Loy, C. and Siew-Moi, P. (2016). Marine algae as a potential source for anti-obesity agents. Mar. Drugs 14: 222, https://doi.org/10.3390/md14120222.Search in Google Scholar PubMed PubMed Central
Wang, M., Carver, J.J., Phelan, V.V., Sanchez, L.M., Garg, N., Peng, Y., Nguyen, D.D., Watrous, J., Kapono, C.A., Luzzatto-Knaan, T., et al.. (2016). Sharing and community curation of mass spectrometry data with global natural products social molecular networking. Nat. Biotechnol. 34: 828–837, https://doi.org/10.1038/nbt.3597.Search in Google Scholar PubMed PubMed Central
Watrous, J., Roach, P., Alexandrov, T., Heath, B.S., Yang, J.Y., Kersten, R.D., Van Der Voort, M., Pogliano, K., Gross, H., Raaijmakers, J.M., et al.. (2012). Mass spectral molecular networking of living microbial colonies. Proc. Natl. Acad. Sci. 109: E1743–E1752, https://doi.org/10.1073/pnas.1203689109.Search in Google Scholar PubMed PubMed Central
Wickham, H. (2011). ggplot2. WIREs Comput. Stat. 3: 180–185, https://doi.org/10.1002/wics.147.Search in Google Scholar
Witvrouw, M. and De Clercq, E. (1997). Sulfated polysaccharides extracted from sea algae as potential antiviral drugs. Gen. Pharmacol. Vasc. Syst. 29: 497–511, https://doi.org/10.1016/s0306-3623(96)00563-0.Search in Google Scholar PubMed
Wolber, G. and Langer, T. (2005). LigandScout: 3-D pharmacophores derived from protein-bound ligands and their use as virtual screening filters. J. Chem. Inf. Model. 45: 160–169, https://doi.org/10.1021/ci049885e.Search in Google Scholar PubMed
Won, T.H., Liao, L., Kang, S.S., and Shin, J. (2014). Simultaneous analysis of furfural metabolites from Rehmanniae radix preparata by HPLC-DAD–ESI-MS. Food Chem. 142: 107–113, https://doi.org/10.1016/j.foodchem.2013.07.026.Search in Google Scholar PubMed
Wright, A.D., König, G.M., and Sticher, O. (1993). New and highly oxidised hydroazulenoid diterpenes from the tropical marine brown alga Dictyota volubilis. Tetrahedron 49: 571–558.10.1016/S0040-4020(01)86260-8Search in Google Scholar
Xia, Z. and Smith, C.D. (2001). Total synthesis of dendroamide A, a novel cyclic peptide that reverses multiple drug resistance. J. Org. Chem. 66: 3459–3466, https://doi.org/10.1021/jo005783l.Search in Google Scholar PubMed
Yu, C., Wang, C.-Z., Zhou, C.-J., Wang, B., Han, L., Zhang, C.-F., Wu, X.-H., and Yuan, C.-S. (2014). Adulteration and cultivation region identification of American ginseng using HPLC coupled with multivariate analysis. J. Pharm. Biomed. Anal. 99: 8–15, https://doi.org/10.1016/j.jpba.2014.06.031.Search in Google Scholar PubMed PubMed Central
Zheng, H., Zhao, Y., and Guo, L. (2022). A bioactive substance derived from brown seaweeds: phlorotannins. Mar. Drugs 20: 742, https://doi.org/10.3390/md20120742.Search in Google Scholar PubMed PubMed Central
Zhong, B., Robinson, N.A., Warner, R.D., Barrow, C.J., Dunshea, F.R., and Suleria, H.A. (2020). LC-ESI-QTOF-MS/MS characterization of seaweed phenolics and their antioxidant potential. Mar. Drugs 18: 331, https://doi.org/10.3390/md18060331.Search in Google Scholar PubMed PubMed Central
Supplementary Material
This article contains supplementary material (https://doi.org/10.1515/bot-2024-0032).
© 2025 Walter de Gruyter GmbH, Berlin/Boston
Articles in the same Issue
- Frontmatter
- In this issue
- Physiology and Ecology
- Bacteria-released algal growth and morphogenesis factors regenerate axenic calli derived from the macroalga Ulva (Chlorophyta) and change the fatty acid profile
- Annual cycle and ecological interactions of macroalgal epiphytes with Cymodocea nodosa and Ruppia maritima in Epanomi and Vravrona, Greece
- Past and present fucalean diversity in the island of Marettimo, Egadi Islands Marine Protected Area (Central Mediterranean, Italy)
- Taxonomy/Phylogeny and Biogeography
- Morphological and molecular characterization of Chnoospora minima (Scytosiphonaceae, Ectocarpales) along Mexican coasts, with the description of C. ramosissima sp. nov.
- Phylogeography of the marine benthic alga Gracilaria salicornia (Gracilariales, Rhodophyta) in Southeast Asia
- Unveiling a novel species, Pachymeniopsis shinchonai sp. nov., from Korea, with reference to phylogenetic relationships within the Grateloupiaceae (Halymeniales, Rhodophyta)
- First record of the red alga Ceramium pallidum (Ceramiales, Ceramiaceae) for the Mediterranean Sea
- Chemistry and Applications
- Chemical profiling of seaweeds of the Arabian Gulf by liquid chromatography-mass spectrometry and in-silico screening against MPOX
Articles in the same Issue
- Frontmatter
- In this issue
- Physiology and Ecology
- Bacteria-released algal growth and morphogenesis factors regenerate axenic calli derived from the macroalga Ulva (Chlorophyta) and change the fatty acid profile
- Annual cycle and ecological interactions of macroalgal epiphytes with Cymodocea nodosa and Ruppia maritima in Epanomi and Vravrona, Greece
- Past and present fucalean diversity in the island of Marettimo, Egadi Islands Marine Protected Area (Central Mediterranean, Italy)
- Taxonomy/Phylogeny and Biogeography
- Morphological and molecular characterization of Chnoospora minima (Scytosiphonaceae, Ectocarpales) along Mexican coasts, with the description of C. ramosissima sp. nov.
- Phylogeography of the marine benthic alga Gracilaria salicornia (Gracilariales, Rhodophyta) in Southeast Asia
- Unveiling a novel species, Pachymeniopsis shinchonai sp. nov., from Korea, with reference to phylogenetic relationships within the Grateloupiaceae (Halymeniales, Rhodophyta)
- First record of the red alga Ceramium pallidum (Ceramiales, Ceramiaceae) for the Mediterranean Sea
- Chemistry and Applications
- Chemical profiling of seaweeds of the Arabian Gulf by liquid chromatography-mass spectrometry and in-silico screening against MPOX