Annual cycle and ecological interactions of macroalgal epiphytes with Cymodocea nodosa and Ruppia maritima in Epanomi and Vravrona, Greece
-
Vaia Myloneli holds an Integrated Master’s degree in Agricultural Science with a specialization in the marine environment and a Master’s degree in the Inter-Institutional MSc program
Oceanography and Management of the Marine Environment (NKUA & HCMR). Her research focuses on the impact of anthropogenic activities on benthic ecosystems, marine ecology, and seagrass meadows.Georgios D. Dorovinis holds a BSc degree from the Department of Biology of the University of Patras and a MSc degree in Oceanography and Management of the Marine Environment from the National and Kapodistrian University of Athens, in collaboration with the Hellenic Centre for Marine Research. His main research interests include the study of pollution and epiphytic communities in seagrass ecosystems.
Dr. Sarah Faulwetter is a benthic ecologist and taxonomist. She is currently a postdoc in the University of Patras working on the impacts of marine aquaculture on the benthic ecosystem. Her research interests focus on the effect of anthropogenic activities on benthic ecosystems in relation to water quality, community ecology and ecosystem functioning.
 ,
,
Ioannis-Dimosthenis S. Adamakis, an Associate Professor at the National and Kapodistrian University of Athens, specializes in plant biology with an emphasis on cell structure and responses to environmental stressors. His research integrates microscopy, molecular biology, and biochemistry to explore various aspects of plant biology, including the study of seagrasses.
and
Dr. Anastasia Tsirika, currently working as laboratory teaching staff in the School of Agriculture at the Aristotle University of Thessaloniki, is a biologist with a Master’s degree in Hydrobiology & Aquacultures and a PhD in Marine Botany. Her research focuses on water quality and bioindicators, taxonomy, distribution and seasonality of marine macrophytes and mapping of benthic communities.


Abstract
Cymodocea nodosa and Ruppia maritima constitute key seagrasses that provide habitat, stabilize sediment, and support nutrient cycling and carbon sequestration in coastal ecosystems. This study aims to investigate the composition and abundance of epiphytic macroalgae on C. nodosa and R. maritima at the coastal sites of Epanomi and Vravrona, parts of Natura 2000 network in Greece. Seasonal plant material collection was conducted from 2022 to 2023 and epiphytic macroalgae were removed and identified to species or genera level. The coverage and number of epiphytic macroalgal taxa were determined and species were classified into Ecological Status Groups (ESGs). The similarity of macroalgal composition and coverage between seasons, locations and hosts was analyzed using cluster analysis. Vravrona exhibited higher epiphyte coverage compared to Epanomi, while Rhodophyta dominated qualitatively and quantitatively throughout the year. In Epanomi, seasonal shifts in dominance were observed, with different algal groups prevailing in each season. Cluster analysis revealed distinct clusters of epiphyte communities, indicating significant differences between sites and seasons, suggesting that local environmental conditions play a role in determining the composition and structure of epiphytic communities. The findings underscore the need for continuous environmental assessments to protect coastal waters and support conservation strategies for these critical ecosystems.
1 Introduction
Coastal ecosystems, renowned for their high productivity and ecological value, require detailed assessments to ensure effective management and conservation (Costanza et al. 1997). Among the vital components of these ecosystems are seagrass meadows formed by species such as Cymodocea nodosa and Ruppia maritima (Tracheophyta). These seagrasses play crucial roles by providing habitat and food for numerous marine organisms, stabilizing sediment, and contributing to nutrient cycling and carbon sequestration (Duarte 2002; Orth et al. 2006). C. nodosa is a seagrass predominantly found in the Mediterranean Sea and the eastern Atlantic Ocean, where it forms extensive meadows in shallow coastal waters (Montefalcone et al. 2007). These meadows are crucial for maintaining biodiversity, as they offer habitat and nursery grounds for various invertebrates and fish species (Francour et al. 1999). R. maritima, on the other hand, is more cosmopolitan, occurring in a wide range of salinity conditions from brackish to hypersaline environments (Koch and Dawes 1991). Although not considered a seagrass sensu stricto (den Hartog and Kuo 2006), as it is often found in coastal lagoons, estuaries, and salt marshes, it similarly supports diverse biological communities (da Rosa et al. 2023; Sullivan 1977; Verhoeven 1980).
The health and sustainability of seagrass meadows are vital indicators of overall coastal ecosystem health. Seagrasses are sensitive to changes in water quality, such as nutrient levels, light availability, and pollutants, making them effective indicators of the overall health of coastal ecosystems (Orth et al. 2006; Waycott et al. 2009). Monitoring seagrass health is crucial for providing early warnings of ecosystem degradation and informing conservation strategies. By regularly assessing seagrass health, it is possible to detect early signs of environmental stress and degradation, enabling timely management interventions to mitigate negative impacts and preserve ecosystem services (Apostoloumi et al. 2021). Epiphytic algae, which grow on the leaves and stems of seagrasses, are an essential component of these ecosystems. These algae contribute to the productivity and nutrient cycling of seagrass meadows, serving as a primary food source for various herbivores (Borowitzka et al. 2006). However, excessive growth of epiphytic algae, often triggered by nutrient enrichment, can be detrimental. It can block light from reaching seagrass leaves, reduce photosynthetic efficiency, and ultimately lead to seagrass decline (Brush and Nixon 2003).
Various studies have underscored the importance of epiphytic algae as bioindicators of water quality and ecosystem health (Balata et al. 2008). Epiphytes respond rapidly to changes in nutrient levels and other environmental stressors, making them valuable indicators of eutrophication and other anthropogenic impacts (Collado-Vides et al. 2007). In the Mediterranean Sea, epiphytes on Posidonia oceanica (Tracheophyta) have been well-documented for decades (Piazzi et al. 2016; Tsirika et al. 2007; Van der Ben 1971), while those on C. nodosa remain less studied (Tsioli et al. 2021). Epiphytes on R. maritima, though widespread throughout a variety of brackish and marine environments, have also received relatively limited attention (Koch and Dawes 1991). Studies have examined epiphytes at the morphological functional-form group level, as proposed by Littler and Littler (1980) (García-Redondo et al. 2019; Pardi et al. 2006). However, classifications at a functional group level relevant to nutrient and light levels, which are critical aspects of eutrophication, have not been fully developed (Orfanidis et al. 2001, 2011). Recently, research has highlighted the value of analyzing epiphytes at the functional group level to classify coastal waters under the European Water Framework Directive, particularly in the context of anthropogenic stress (Tsioli et al. 2021). Thus, examining the epiphytic communities on C. nodosa and R. maritima can provide critical insights into the ecological status of coastal waters.
The Natura 2000 sites Epanomi Thessaloniki (GR1220012) and Vravrona Attica (GR3000004) are two coastal areas in Greece that are known for their unique biodiversity and ecological importance, as they support numerous species of flora and fauna, some of which are protected under European directives. The coastal water status at Epanomi has been a subject of ongoing environmental monitoring, due to pressures from urbanization and tourism, which have the potential to impact water quality and habitat integrity (Papadimitriou et al. 2012). In addition, the coastal waters of Vravrona have been assessed for their ecological status, which is influenced by both natural and anthropogenic factors, such as agricultural runoff and coastal development. Efforts to maintain and improve the water quality in this region are crucial for the conservation of its biodiversity (Katsoulis et al. 2019). Both sites highlight the importance of continuous environmental assessments to ensure the protection of their coastal waters and the broader ecosystems they support. Taking into consideration the unique features of both sites, combined with the significance of monitoring seagrass communities, this research aims to investigate the composition and abundance of epiphytic macroalgae on C. nodosa and R. maritima, across coastal area of Epanomi and Vravrona Natura 2000 sites. By comparing these two seagrass species, we aim to identify patterns and differences in epiphytic algal communities that could reflect underlying environmental conditions, including seasonality, and anthropogenic impacts. Additionally, through this study, we aim to enhance our understanding of the ecological dynamics between seagrasses and their macroalgal epiphytic communities, providing valuable data for the conservation and management of these critical coastal ecosystems.
2 Materials and methods
2.1 Study area
Sampling of both plant species was carried out at sites of the European Natura 2000 network in the Aegean Sea, Greece: Epanomi Thessaloniki (GR1220012) (Figure 1) in the Thermaikos Gulf, North Aegean Sea, and Vravrona Attica (GR3000004) (Figure 1) in the Petalioi Gulf, South Aegean Sea. The coastal area of Epanomi is located in the relatively enclosed and shallow Thermaikos Gulf, where many economic and productive activities take place (shellfish farming, fisheries, navigation, industrial production, etc.) and rivers in the western coastal zone (Axios, Gallikos, Loudias, Aliakmonas) supply the gulf with nutrients, resulting in its eutrophication. In addition, the city of Thessaloniki, with a population exceeding 1.5 million, is located in the coastal zone of Thermaikos Gulf. The area of Vravrona is part of the Petalioi Gulf, where significant economic, touristic, and productive activities take place. The wetland includes a small, continuous flow system (Erasinos River), which is being supplied with water from seasonal springs and rainwater. Finally, the area of Vravrona is an important archaeological site in Attica with intense tourist activity, especially during the summer months.
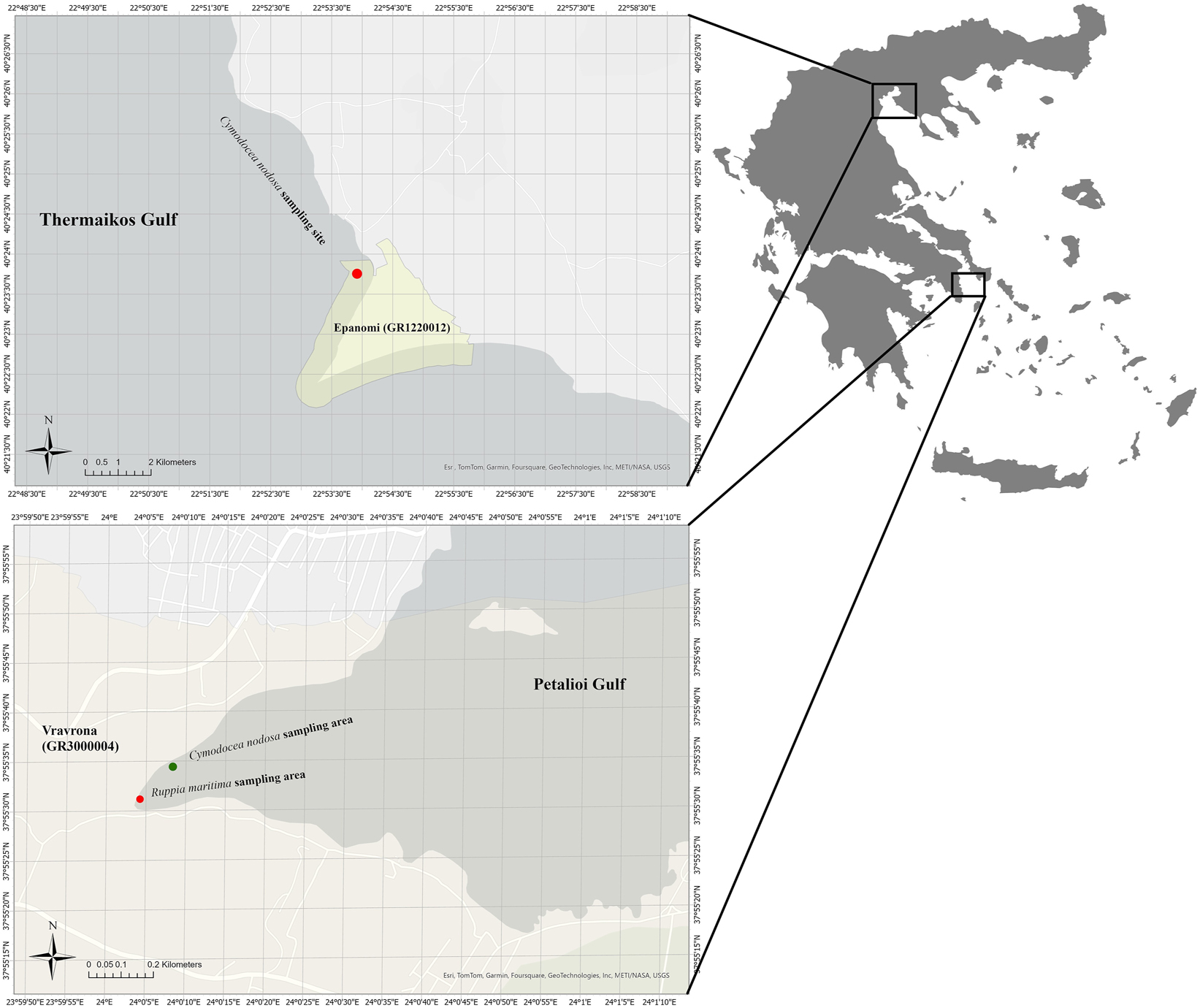
Map of the sampling areas. The Cymodocea nodosa meadows studied are located at the northern side of Epanomi Thessaloniki (GR1220012) in the Thermaikos Gulf, North Aegean Sea and in the protected area of Vravrona Attica (GR3000004) in the Petalioi Gulf, South Aegean Sea. The Ruppia maritima meadow is located in the protected area of Vravrona Attica (GR3000004).
The underwater landscape at both sites consists mostly of soft substrata, either unvegetated or covered by seagrasses. The monospecific meadow of C. nodosa on the northern side of the Epanomi cape (40°23′58.23″ N, 22°54′4.30″ E) covers an area of 61.09 ha (ACOOLAGOONS 2011) and its distribution starts from the first centimeters of the sublittoral zone and reaches up to about 1.5 m depth. Similarly, in the protected area of Vravrona (37° 55′ 34.44″ N, 24°0′ 10.78″ E), the species is found at a distance of 30–70 m from the shore in very shallow waters. The seagrass R. maritima is found only in the protected area of Vravrona, where it forms scattered patches rather than a single meadow, in brackish waters near the river mouth. The Vravrona wetland is permanently flooded, allowing R. maritima to maintain its vegetative form all year round.
2.2 Field work
For the analysis of the macroalgal epiphytic leaf flora of the seagrasses C. nodosa and R. maritima, plant material from both species was collected seasonally during 2022–2023. Samples of C. nodosa were collected from both Epanomi and Vravrona starting in summer 2022, whereas sampling of R. maritima was conducted in Vravrona beginning in autumn 2022.
In this study, a combination of destructive and non-destructive methods was used. First, the density (i.e. shoots m−2) of the C. nodosa meadow was studied in situ by placing, haphazardly, quadrats of 20 × 20 cm along a transect parallel to the shore. The length of the transect was about 100 m, and quadrats were placed about 10 m from each other. The water depth where the field measurements and sample collections were made was approximately 0.5 m. In total, 10 quadrats were measured per site seasonally. A 400 cm2 sampling area is considered the minimum representative sampling area for Mediterranean sublittoral communities according to Dhont and Coppejans (1977) and Boudouresque and Belsher (1979). In addition, basic characteristics of the meadows and the aquatic environment were recorded, such as the lower depth limit or the meadow structure (homogeneity, bottom cover), etc.
Subsequently, plant material was collected from both species. A cylindrical PVC tube of 1 m length and 200 mm diameter (area of 314 cm2) was used to remove 5 haphazard cores of the seabed along the transect, at the respective depths per season and site. The tube was drilled into the seabed (approximately one-third) and then removed, and contained leaves, roots, and rhizomes of each species. The collected plant material was stored in plastic containers of 3 liters, with a solution of 4 % formaldehyde buffered in seawater. The seawater was saline in the containers of C. nodosa and brackish in those of R. maritima.
2.3 Laboratory analyses
In the laboratory, a phenological analysis of the C. nodosa plants was conducted for 20 plants (randomly selected from the 5 cores) for each study site and season. The phenological analysis was conducted in 80 plants of C. nodosa from the Epanomi site (20 per season) and 80 plants from the Vravrona site (20 per season). Each plant was separated into leaves, rhizomes, roots, seeds and flowers. Rhizomes and roots were separated into orthotropic and plagiotropic, while leaves of each shoot were divided into three categories (adult, intermediate and juvenile) according to Giraud (1979). Leaf biometry (length and width of the photosynthetic part of the leaves) was measured using the “Weeder” software (Ramfos et al. 2012), to determine the leaf area index in m2 m−2 (Pergent et al. 1995).
For the study of the macroalgal epiphytic flora, 10 shoots (randomly selected from the 5 cores) for each seagrass, study site and season were used for the estimation of adult leaves’ coverage with epiphytic organisms and 10 shoots (randomly selected from the 5 cores) for each seagrass, study site and season for the identification and quantification of macroalgal epiphytic organisms.
To estimate the % cover of adult leaves with epiphytic organisms, we used the adult leaves of the 10 plants for each seagrass, site and season. Per season we studied 30 shoots (10 C. nodosa shoots from the site of Epanomi, 10 C. nodosa shoots from the site of Vravrona and 10 R. maritima shoots from the site of Vravrona). From each one of the plants, we measured all of the adult leaves. A total of 103 adult leaves were measured for all 4 seasons from C. nodosa meadow from the Epanomi site (Winter: 26, Spring: 26, Summer: 26 and Autumn: 25), 107 adult leaves were measured for all 4 seasons from C. nodosa meadow from the Vravrona site (Winter: 26, Spring: 26, Summer: 27 and Autumn: 28), and 87 leaves were measured for all 4 seasons from R. maritima meadow from the Vravrona site (Winter: 21, Spring: 24, Summer: 21 and Autumn: 21).
For the description of the epiphytic community, we removed the epiphytes from 10 randomly selected adult leaves from each seagrass, site and season. To remove the macroalgal epiphytic taxa from the leaves a blade was used, along the length of the leaf (Dauby and Poulicek 1995). For the observation and identification of macroalgal organisms, a stereoscope and optical microscope as well as relevant specialized taxonomic literature were used. The epiphytic taxa were identified at the level of species or genera.
The parameters that were measured/calculated for the epiphytes are: a) the part of the adult leaves being occupied by epiphytic organisms (including micro- and macroalgae, as well as sedentary fauna), expressed as the percentage of the total leaf surface, b) the total dry biomass of epiphytes on 10 adult leaves, c) the total number of epiphytic macroalgal taxa per species and season, d) the coverage of each macroalgal taxon (the surface covered by each taxon, expressed as percentage coverage of the total surface of leaves) – for crustose species, coverage was estimated as the percentage of the leaf surface occupied by the species (Boudouresque 1971; Reyes et al. 1998) and for erect filamentous or fleshy species coverage was estimated as the percentage of the leaf surface covered by the horizontal projection of the species (Ballesteros et al. 1992; Reyes et al. 1998), e) total epiphyte coverage which was calculated by summing up the coverage of all species per sample (Balata et al. 2007) – as macroalgal vegetation develops on layers (crustose, erect, etc.), the total epiphyte coverage often exceeds 100 % of the leaf’s surface, f) the qualitative and quantitative percentage of taxa belonging to the classes of Chlorophyta, Rhodophyta and Heterokontophyta (Class Phaeophyceae), and g) the percentage of each Ecological Status Group (ESG) according to Ecological Evaluation Index (Orfanidis et al. 2011). The EEI classifies coastal macroalgae in two Ecological Status Groups (ESGs): ESG I (slow-growing, late-successional species) and ESG II (fast-growing, opportunistic species), divided into three and two subclusters respectively.
2.4 Statistical analyses
Differences between locations and seasons were assessed with non-parametric tests (Wilcoxon / Kruskal–Wallis, respectively) as all data showed heteroscedacity and non-normal distributions. The significance level was set to α = 0.05 for all tests.
The similarity of the macroalgal composition and coverage between sampling periods, sites, and host plants was visualized through a cluster analysis (Bray-Curtis dissimilarity coefficient on the untransformed coverage data). To better understand the processes behind seasonal community assembly at the two sites and on the two host plants, temporal beta diversity between seasons was calculated. Beta diversity was then partitioned into its turnover and nestedness components to assess whether species replacement or species loss/recovery was occurring between seasons. All analyses were performed in Microsoft Office Excel 2021 and in R version 4.3.2 (R Core Team 2023), for the analyses of beta diversity the betapart package (Baselga et al. 2023) was used using Sørensen’s dissimilarity index, the dissimilarity and cluster were created using the vegdist (package vegan, Oksanen et al. 2022) and hclust functions.
3 Results
3.1 General description of marine vegetation in the studied sites
The studied sites were characterized by soft substrata, with an alternation of communities of seagrasses and bare substratum.
In Epanomi, a succession of benthic vegetation based on substratum type and depth was observed. In shallow waters, the soft substratum was covered by a monospecific meadow of C. nodosa, whereas the endemic Mediterranean seagrass P. oceanica was present at greater depths. Additionally, isolated areas of both natural and artificial hard substrata covered with macroalgae were present, dominated by the brown algae Gongolaria barbata (formerly Cystoseira barbata) and Padina pavonica (Phaeophyceae) and the green algae Cladophora spp. and Ulva (formerly Enteromorpha) spp. (Chlorophyta).
In Vravrona, the variation of salinity, due to the outflow of fresh water, resulted in the appearance of a mosaic form of the marine vegetation in shallow waters, with an alternation of monospecific meadows of C. nodosa and R. maritima. The distribution of the two seagrass species was determined by salinity, with Ruppia dominating towards the river mouth. The endemic P. oceanica (Tracheophyta) formed an extended meadow at greater depths.
3.2 Meadow density
The shoot density of the C. nodosa meadows in Vravrona and Epanomi varied significantly over the course of the year, presenting similar trends at both sites, with peaks in summer and the lowest density in winter (Figure 2). Specifically, in Vravrona, densities reached an average of 837.5 shoots m−2 in summer, while in autumn and winter a gradual decrease was observed with a small recovery during spring (380 shoots m−2). In Epanomi, the summer density was even higher with an average of 1,185 shoots m−2, while during autumn and winter density decreased to (985 shoots m−2 and 365 shoots m−2). As in Vravrona, an increase was subsequently recorded again in spring (725 shoots m−2). Shoot density differed significantly between the two locations (W = 1,413, p < 0.0001) and in both locations also between seasons (Epanomi: χ2 = 25.826, p < 0.0001, Vravrona: χ2 = 28.019, p < 0.0001).
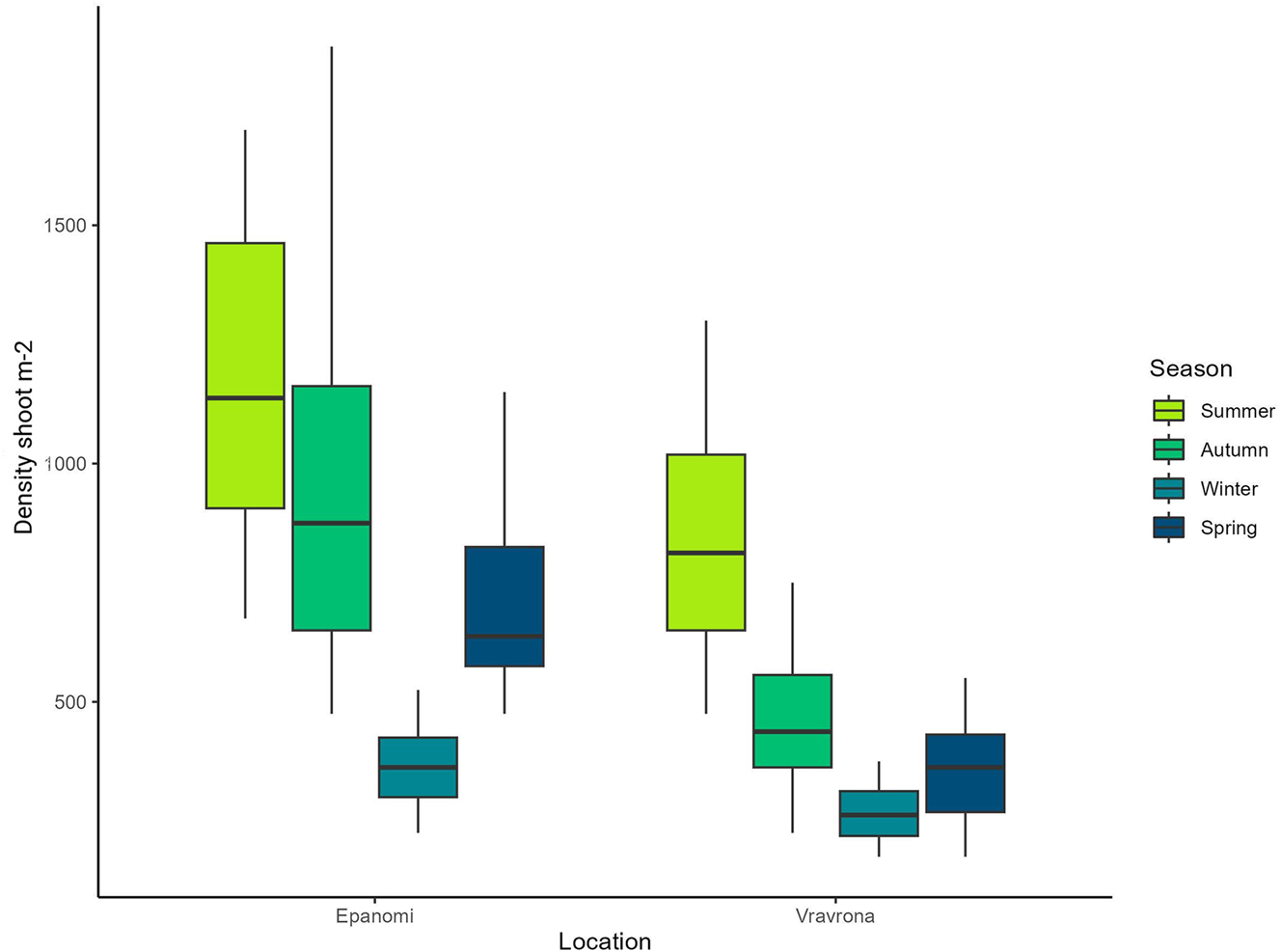
Seasonal variation in shoot density (shoots m−2) of Cymodocea nodosa in the study areas during 2022–2023. Shoot density was measured in situ using a 400 cm2 quadrat, where 10 measurements took place for each season and site (N = 10 for each meadow and season). Whiskers indicate the smallest and largest non-outlier values, while the box encompasses the interquartile range (IQR).
3.3 Leaf area index
The leaf area index (LAI) of adult and intermediate leaves of C. nodosa from Vravrona and Epanomi meadows varied significantly among seasons, and no intermediate leaves were present in Epanomi during Spring and in Vravrona during winter. The lowest values of LAI were recorded during winter in both sites for adult and intermediate leaves and the highest during summer for intermediate leaves in both sites and during summer and autumn for the adult leaves in Epanomi and Vravrona, respectively (Figure 3). In Epanomi, the LAI for adult leaves during spring was comparable to autumn, whereas in the Vravrona site, the LAI for the adult leaves during spring was comparable to winter. The highest values of LAI for both adult (1.244 m2 m−2) and intermediate (0.793 m2 m−2) leaves were recorded in the Epanomi meadow.
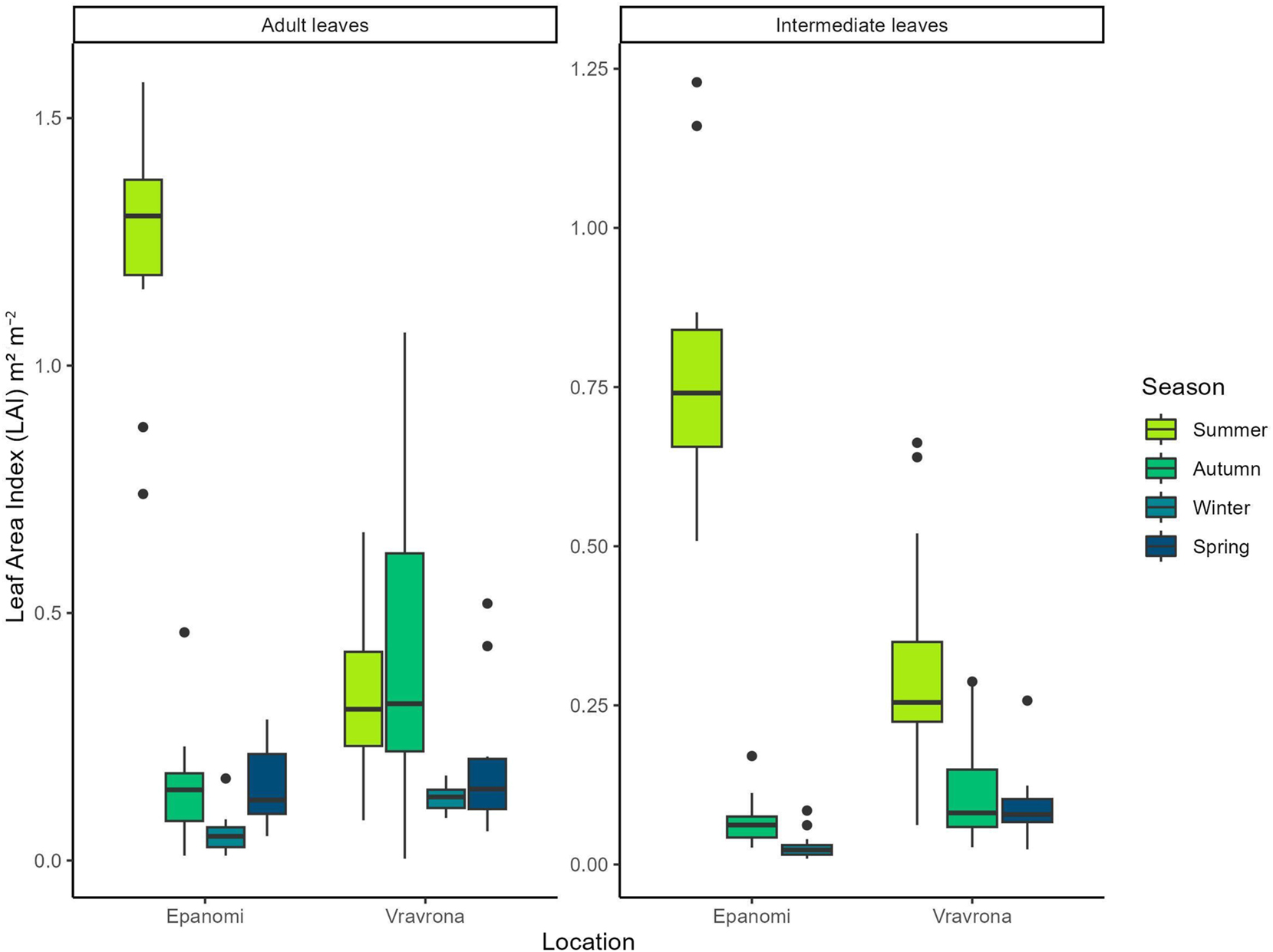
Seasonal variation of leaf area index (LAI) (m2 m−2) of Cymodocea nodosa leaves in the study areas during 2022–2023. The leaf area index of leaves was divided into (a) adult and (b) intermediate leaves. The measurements were conducted in 20 randomly selected plants from each site and season. Whiskers indicate the smallest and largest non-outlier values, while the box encompasses the interquartile range (IQR); black dots denote extreme observed values (outliers).
For both intermediate and adult leaves the LAI did not show any statistically significant differences between the two locations but in both locations significant differences between seasons were found (Epanomi adult leaves: χ2 = 31.812, p < 0.0001, Epanomi intermediate leaves: χ2 = 24.592, p < 0.0001, Vravrona adult leaves: χ2 = 19.294, p = 0.0002, Vravrona intermediate leaves: χ2 = 27.787, p < 0.0001).
3.4 Epiphytes
The epiphytic flora of C. nodosa leaves from both Vravrona and Epanomi sites included macroalgae and periphytic microalgae (mainly diatoms and cyanobacteria). Epiphytic macroalgal diversity was higher in Vravrona compared to Epanomi (Figure 4). A total of 34 taxa at specific and infraspecific rank were identified as epiphytes on C. nodosa leaves at both sites, 27 in Vravrona and 23 in Epanomi. Across the seasons, the highest biodiversity was recorded at both sites during spring and summer, whereas fewer taxa were identified during winter. On R. maritima, a total of 7 macroalgal epiphytic taxa were identified. The above-mentioned data refer to the total number of taxa pooled over the studied adult leaves.
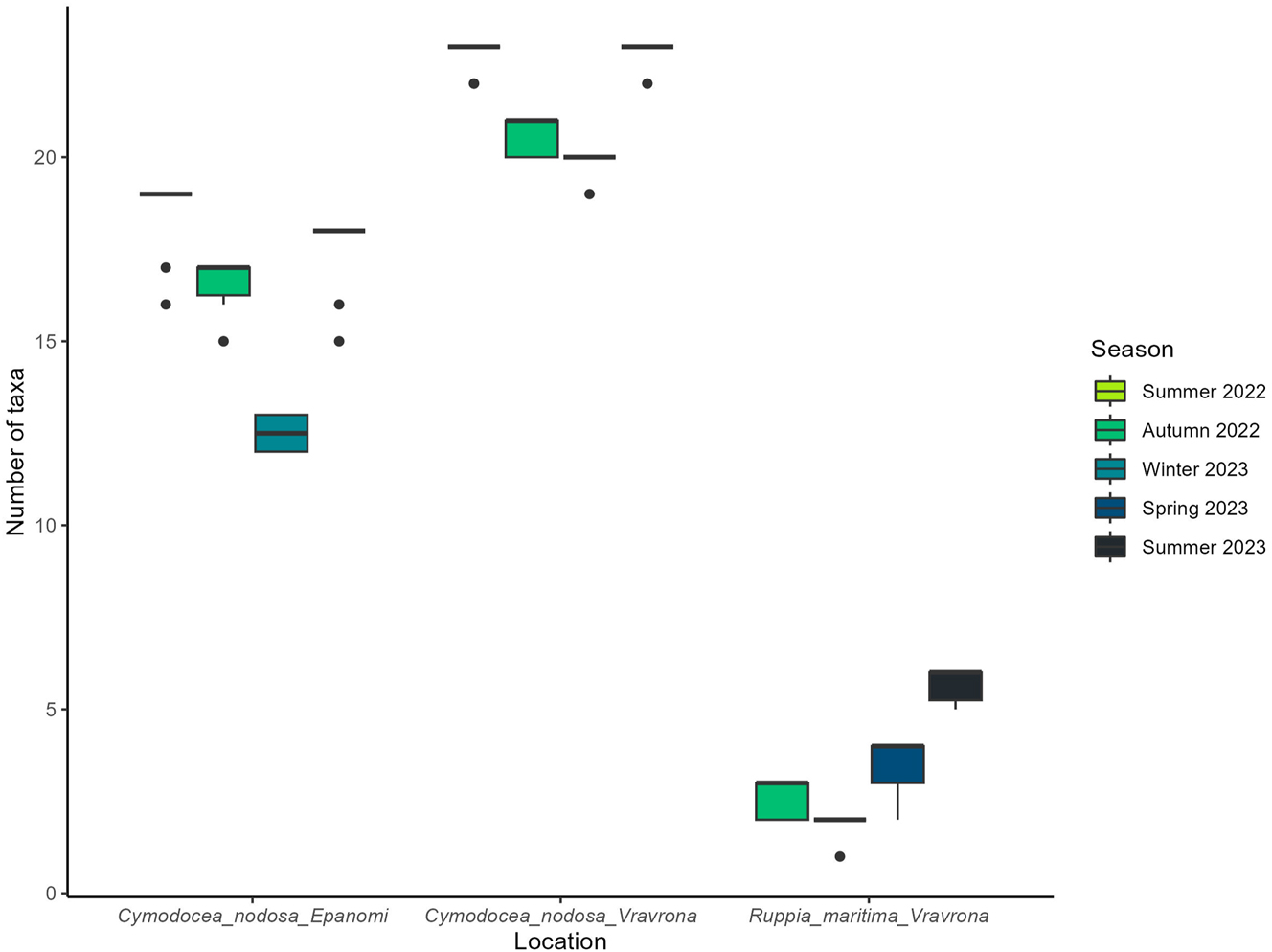
Mean number of identified macroalgal epiphytic taxa of Cymodocea nodosa and Ruppia maritima leaves in the study areas, by season. Mean values represent the number of epiphytic macroalgal taxa identified on the adult leaves studied per site and season (N = 10). Whiskers indicate the smallest and largest non-outlier values, while the box encompasses the interquartile range (IQR); black dots denote extreme observed values (outliers). C. nodosa samples began to be collected in the summer of 2022, while R. maritima sampling began in the autumn of 2022.
As a general observation, the leaf coverage showed a longitudinal gradient, with increasing coverage on the apical (older) part of the leaves.
In Figure 5a the mean percentage coverage of adult leaves’ surface with epiphytic organisms (including microalgae, macroalgae and sedentary fauna) is given, while the mean total coverage of epiphytic macroalgal community (sum of each taxa’s coverage) is presented in Figure 5b. The highest values in leaf coverage with epiphytes for Vravrona were observed during autumn (for both C. nodosa and R. maritima), whereas the highest leaf coverage with epiphytes in Epanomi site were recorded during summer (Figure 5a).
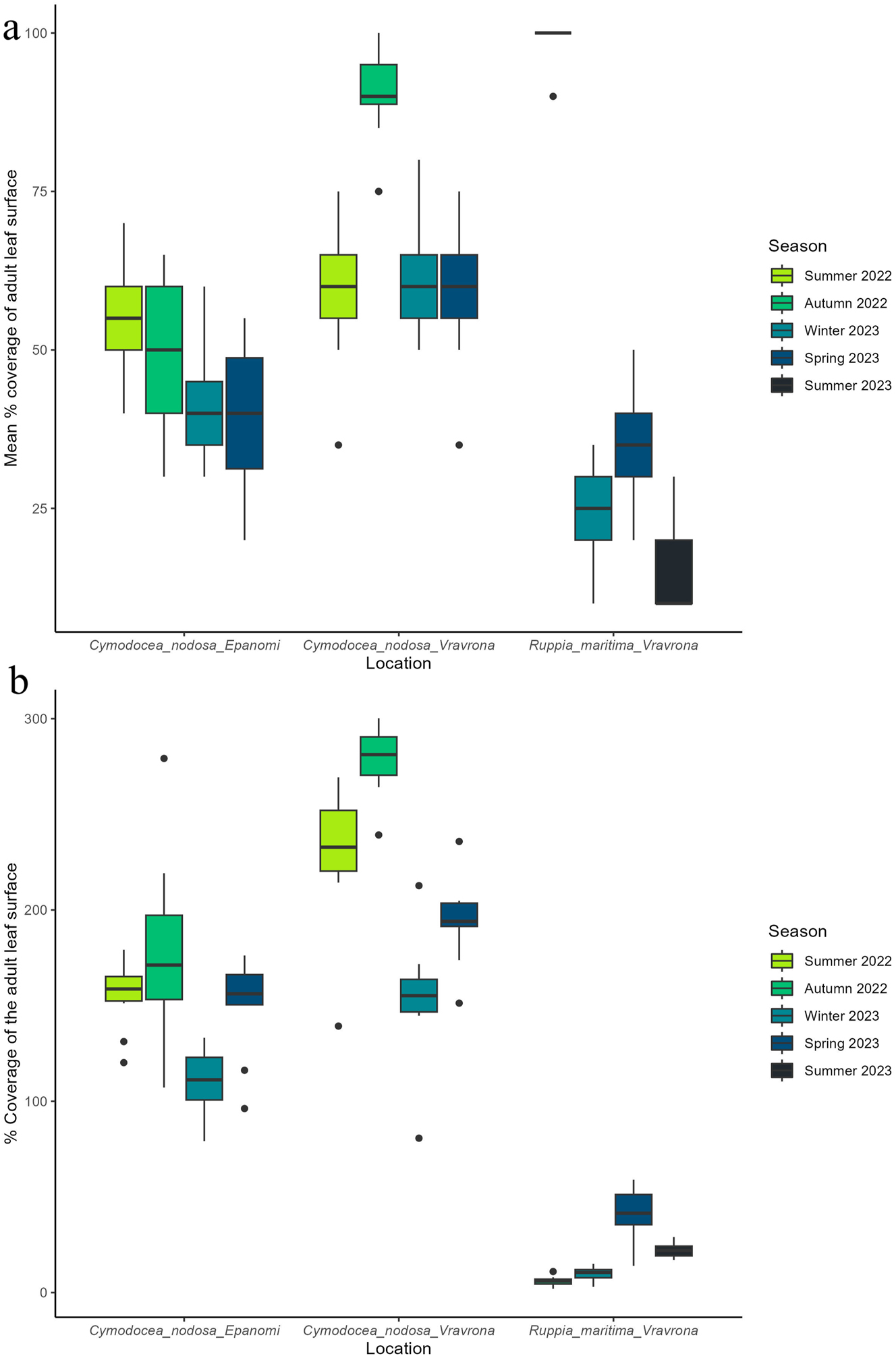
Coverage of adult leaf surface with epiphytic organisms (including microalgae, macroalgae and sedentary fauna) and total macroalgal epiphytic coverage (sum of coverage by macroalgal taxa). (a) Mean % coverage of adult leaf surface of Cymodocea nodosa and Ruppia maritima with epiphytes (including microalgae, macroalgae and sedentary fauna) in the study areas. A total of 103 adult leaves were measured for all 4 seasons from C. nodosa meadow from the Epanomi site (Winter: 26, Spring: 26, Summer: 26 and Autumn: 25), 107 adult leaves were measured for all 4 seasons from C. nodosa meadow from the Vravrona site (Winter: 26, Spring: 26, Summer: 27 and Autumn: 28), and 87 leaves were measured for all 4 seasons from R. maritima meadow from the Vravrona site (Winter: 21, Spring: 24, Summer: 21 and Autumn: 21). (b) Mean total macroalgal epiphytic coverage, expressed as % of adult leaf surface of C. nodosa and R. maritima in the study areas, by season. This figure concerns only macroalgal taxa. The total macroalgal epiphytic coverage was calculated from 10 randomly selected adult leaves from each seagrass, site and season. Whiskers indicate the smallest and largest non-outlier values, while the box encompasses the interquartile range (IQR); black dots denote extreme observed values (outliers). C. nodosa samples began to be collected in the summer of 2022, while R. maritima sampling began in the autumn of 2022.
As total coverage of the epiphytic macroalgal community (Figure 5b) for C. nodosa was calculated as the sum of individual taxon coverage (thus considering its development in layers), it showed much higher numbers (exceeding 100 %) than the percentage of the leaf surface covered with epiphytes (Figure 5a), despite other epiphytic organisms being included in the latter (microalgae, sedentary fauna). This indicates that the macroalgal community is the dominant community on C. nodosa leaves throughout the year. The situation was completely different for the epiphytic community of R. maritima. Here, while the highest coverage of leaves with epiphytes (100 % – Figure 5a) was recorded in autumn, the community of macroalgae had the lowest overall coverage during that period, indicating that the epiphytic macroalgal community was not the dominant one in the R. maritima meadow (Figure 5a and b). Both coverage with all epiphytes and coverage with only macroalgal epiphytes showed significant differences between locations (C. nodosa only, all epiphytes: W = 1,314.5, p < 0.001, macroalgal epiphytes: W = 272.5, p < 0.001) and between seasons in both locations, as well as for R. maritima (All epiphytes: C. nodosa Epanomi: χ2 = 34.988, p < 0.001, C. nodosa Vravrona: χ2 = 61.872, p < 0.001, R. maritima Vravrona: χ2 = 70.702, p < 0.001; macroalgal epiphytes: C. nodosa Epanomi: χ2 = 17.447, p < 0.001, C. nodosa Vravrona: χ2 = 29.483, p < 0.001, R. maritima Vravrona: χ2 = 31.964, p < 0.001).
In Figure 6, the total dry biomass of epiphytes collected from 10 adult leaves of C. nodosa is presented for each region and season. The biomass was measured as a combined value for all 10 leaves per region and season, without recording data separately for each leaf. In Vravrona, the highest value of the epiphytic biomass on adult leaves was recorded during autumn and the lowest during winter. In Epanomi, the highest value of epiphytic biomass was observed during summer and the lowest during spring.
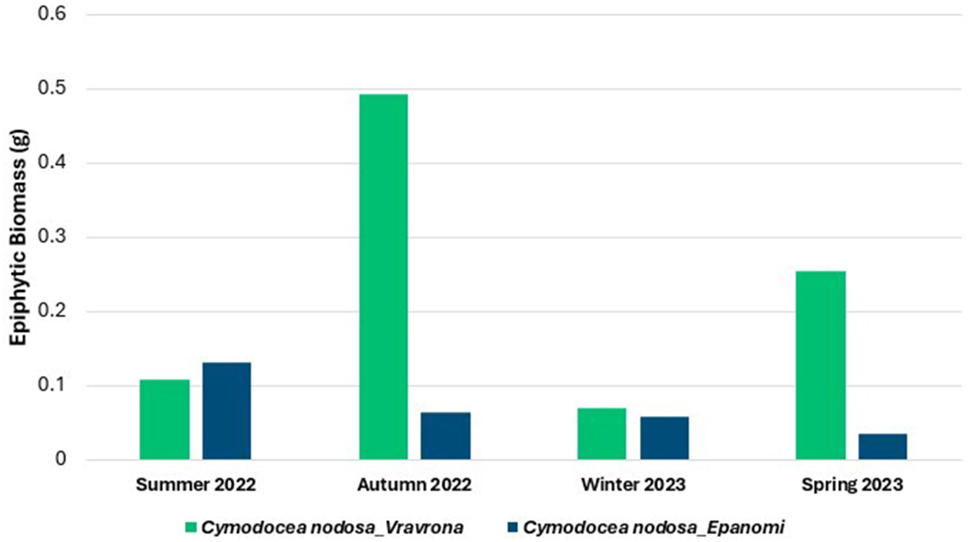
Total dry biomass (g) of epiphytic organisms on 10 adult leaves of Cymodocea nodosa in the meadows of Epanomi and Vravrona, by season, during 2022–2023.
Vravrona was characterized by higher coverage of C. nodosa leaves with epiphytes throughout the year than Epanomi, and especially during autumn (Figure 5a and 6). In all seasons, a qualitative and quantitative dominance of Rhodophyta was recorded in Vravrona (Figure 7a and b). Concerning the identified taxa (Table 1), crustose macroalgae of the genera Hydrolithon and Pneophyllum, as well as erect Rhodophyta of the genera Laurencia, Chondria, Polysiphonia and Ceramium were the most abundant. The genera Cladosiphon, Sphacelaria, Feldmannia and Dictyota dominated among Heterokontophyta (Class Phaeophyceae), whereas of the Chlorophyta, the genus Cladophora was also notable. In addition to macroalgae, periphytic solitary and tube-dwelling colonial diatoms were abundant. The presence of the alien green macroalga Caulerpa cylindracea in the rhizome community was recorded.
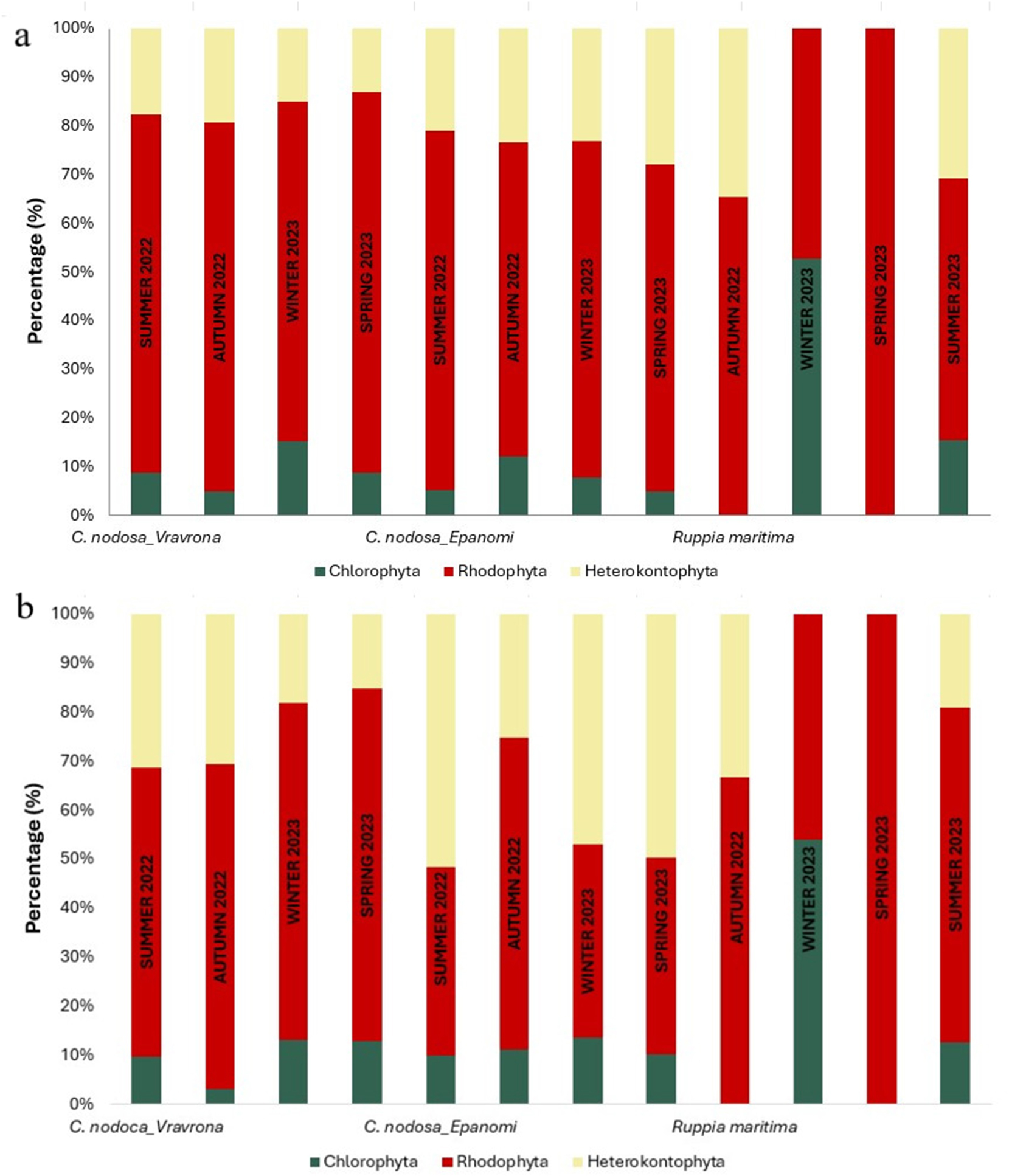
Qualitative and quantitative analysis of the identified macroalgal groups. (a) Relative number of identified taxa in different macroalgal groups observed as epiphytes on leaves of Cymodocea nodosa and Ruppia maritima in the study areas, by season. Values represent the mean number of taxa of each macroalgal group found on the 10 adult leaves studied per seagrass, site and season. (b) Relative total coverage of identified taxa of different algal groups observed on leaves of C. nodosa and R. maritima in the study areas, by season. Values represent the mean total coverage of each macroalgal group found on the 10 adult leaves studied per seagrass, site and season. Cymodocea nodosa samples began to be collected in the summer of 2022, while R. maritima sampling began in the autumn of 2022.
Identified macroalgal taxa growing as epiphytes on Cymodocea nodosa and Ruppia maritima leaves, by season in Epanomi and Vravrona; C. nodosa samples began to be collected in the summer of 2022, while R. maritima sampling began in the autumn of 2022.
| C. nodosa | R. maritima | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Vravrona | Epanomi | Vravrona | ||||||||||
| Summer | Autumn | Winter | Spring | Summer | Autumn | Winter | Spring | Autumn | Winter | Spring | Summer | |
| Chlorophyta | ||||||||||||
|
|
||||||||||||
| Cladophora laetevirens (Dillwyn) Kützing | + | + | + | + | ||||||||
| Cladophora sp. Kützing | + | + | + | + | + | + | + | + | + | + | ||
| Ulva sp. Linnaeus | + | |||||||||||
|
|
||||||||||||
| Rhodophyta | ||||||||||||
|
|
||||||||||||
| Acrochaetium sp. Nägeli | + | + | + | + | + | + | ||||||
| Antithamnion sp. Nägeli | + | |||||||||||
| Carradoriella elongata (Hudson) Savoie et G.W. Saunders | + | |||||||||||
| Ceramium diaphanum (Lightfoot) Roth | + | + | + | + | + | + | + | + | ||||
| Ceramium giacconei Cormaci et G.Furnari | + | |||||||||||
| Ceramium sp. Roth | + | + | + | + | + | + | + | + | + | |||
| Ceramothamnion codii H.Richards | + | + | + | + | + | + | + | + | ||||
| Chondria capillaris (Hudson) M.J.Wynne | + | + | ||||||||||
| Chondria mairei G.Feldmann | + | + | + | + | ||||||||
| Chondria sp. C.Agardh | + | + | ||||||||||
| Chroodactylon ornatum (C.Agardh) Basson | + | + | + | + | + | + | + | + | ||||
| Erythrotrichia carnea (Dillwyn) J.Agardh | + | + | ||||||||||
| Gayliella transversalis (Collins et Hervey) T.O.Cho et Fredericq | + | + | ||||||||||
| Herposiphonia secunda (C.Agardh) Ambronn | + | + | + | + | + | + | ||||||
| Hydrolithon cruciatum (Bressan) Y.M. Chamberlain | + | + | + | + | + | + | + | + | ||||
| Laurencia minuta subsp. scammaccae G.Furnari et Cormaci | + | + | + | + | ||||||||
| Lejolisia mediterranea Bornet | + | + | + | + | + | |||||||
| Lophosiphonia cristata Falkenberg | + | + | + | + | ||||||||
| Melanothamnus sphaerocarpus (Børgesen) Díaz-Tapia et Maggs | + | + | + | + | ||||||||
| Pneophyllum fragile Kützing | + | + | + | + | + | + | + | + | ||||
| Polysiphonia sp. Greville | + | + | + | + | + | + | + | + | + | + | + | |
| Pseudoceramium tenerrimum (G. Martens) Barros-Barreto et Maggs | + | + | + | + | ||||||||
| Stylonema alsidii (Zanardini) K.M. Drew | + | + | + | + | + | + | + | + | ||||
| Womersleyella setacea (Hollenberg) R.E. Norris | + | + | + | + | + | |||||||
|
|
||||||||||||
| Heterokontophyta | ||||||||||||
|
|
||||||||||||
| Cladosiphon irregularis (Sauvageau) Kylin | + | + | + | + | ||||||||
| Dictyota implexa (Desfontaines) J.V. Lamouroux | + | + | + | + | + | + | + | + | ||||
| Feldmannia sp. Hamel | + | + | + | + | + | |||||||
| Giraudia sphacelarioides Derbès et Solier | + | |||||||||||
| Myriactula rivulariae (Suhr ex Areschoug) Feldmann | + | + | + | + | ||||||||
| Myrionema orbiculare J. Agardh | + | + | ||||||||||
| Sphacelaria cirrosa (Roth) C. Agardh | + | + | + | + | + | + | + | + | + | |||
In Epanomi, Rhodophyta dominated qualitatively (number of identified taxa) during all seasons (Figure 7a), while quantitatively (contribution of each group’s coverage on the macroalgal community), i. Heterokontophyta and Chlorophyta dominated in winter, ii. Heterokontophyta in spring, iii. Heterokontophyta and Cyanobacteria in summer and iv. Rhodophyta in autumn (Figure 7b). Among the Rhodophyta, the crustose alga Hydrolithon was the most abundant, followed by erect Rhodophyta of the genera Chondria, Ceramium, Polysiphonia, and Womersleyella. The genera Myriactula, Giraudia, Dictyota and Sphacelaria prevailed among the Heterokontophyta (Phaeophyceae). As for the green algae, the genus Cladophora was abundant. In addition to macroalgae, Cyanobacteria, periphytic diatoms and dinoflagellates, foraminifera and epiphytic hydroids were abundant, while the brown algae Padina and Cystoseira, and the green alga Ulva were present in the rhizomes.
Regarding the community of R. maritima, macroalgae and other autotrophic organisms grew epiphytically on the leaves, with a seasonally varying dominance. A total of 7 macroalgal taxa were identified, most of them belonging to the Rhodophyta (Table 1), whereas the number of identified macroalgal taxa was highest in summer and lowest in winter (Figure 4). The % coverage of leaves’ surface with epiphytes varied significantly during the seasons (Figure 5a), with a peak during autumn, when the coverage of adult leaves with epiphytes was 100 %. On the other hand, the total coverage of the epiphytic macroalgal community was low during this period (Figure 5b), due to the absolute dominance of periphytic erect diatoms. Focusing on macroalgae, in spring the epiphytic Rhodophyta Acrochaetium and Antithamnion dominated, in summer, the epiphytic macroalgae of the genera Cladophora, Feldmannia and Acrochaetium dominated, whereas during autumn and winter the contribution of macroalgae on the epiphytic community was low (Figure 5b).
Regarding the classification of the identified taxa in Ecological Status Groups (ESG I – slow-growing, late-successional species and ESG II – fast-growing, opportunistic species), ESG II dominated, although ESG I group was quantitatively well represented on Cymodocea leaves, due to the significant presence of crustose Rhodophyta. ESG II dominated due to the high development of fast growing, opportunistic species (mainly filamentous). On the leaves of Ruppia, only species of the ESG II group were found (Figure 8).
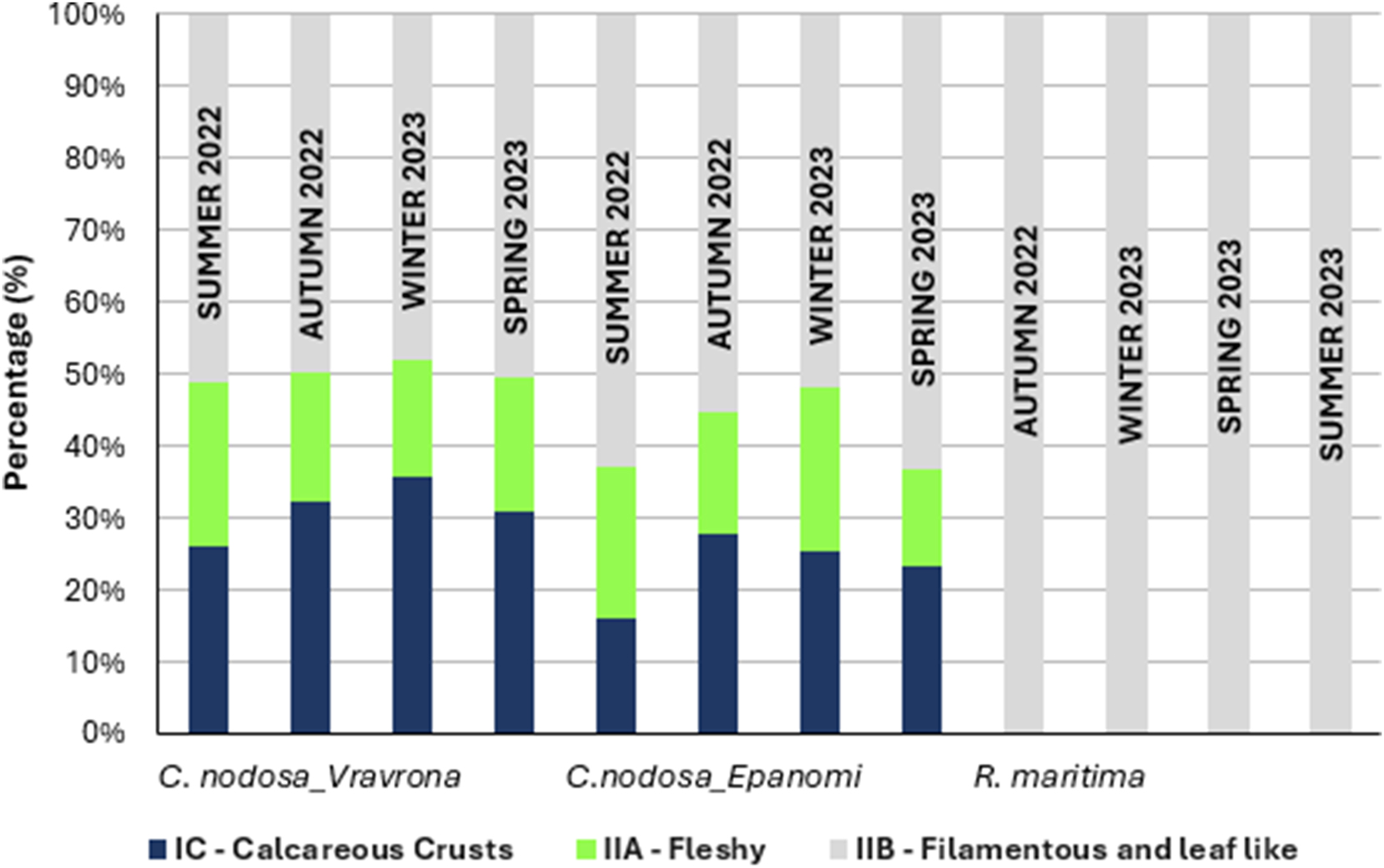
Quantitative dominance of Ecological Status Groups (ESGs) of the epiphytic macroalgae, based on the EEI-c Index (Orfanidis et al. 2011) of Cymodocea nodosa and Ruppia maritima in the study areas, by season. Values represent the mean total coverage of each ESG found on the 10 adult leaves studied per seagrass, site and season; C. nodosa samples began to be collected in the summer of 2022, while R. maritima sampling began in the autumn of 2022.
3.5 Species community analyses
The dendrogram resulting from the hierarchical clustering analysis revealed three distinct clusters of epiphyte communities (Figure 9). The epiphyte communities of R. maritima were almost completely distinct from those of C. nodosa, at a dissimilarity level of 0.94. Within the Cymodocea cluster, samples were separated spatially, with a separation of the two sampling sites at a dissimilarity level of 0.53. Seasonality did not seem to play a major role in separating samples, instead, local processes of community assembly seemed to be more important. However, independent of site or host, the autumn communities always showed the greatest differences compared to the other seasons.
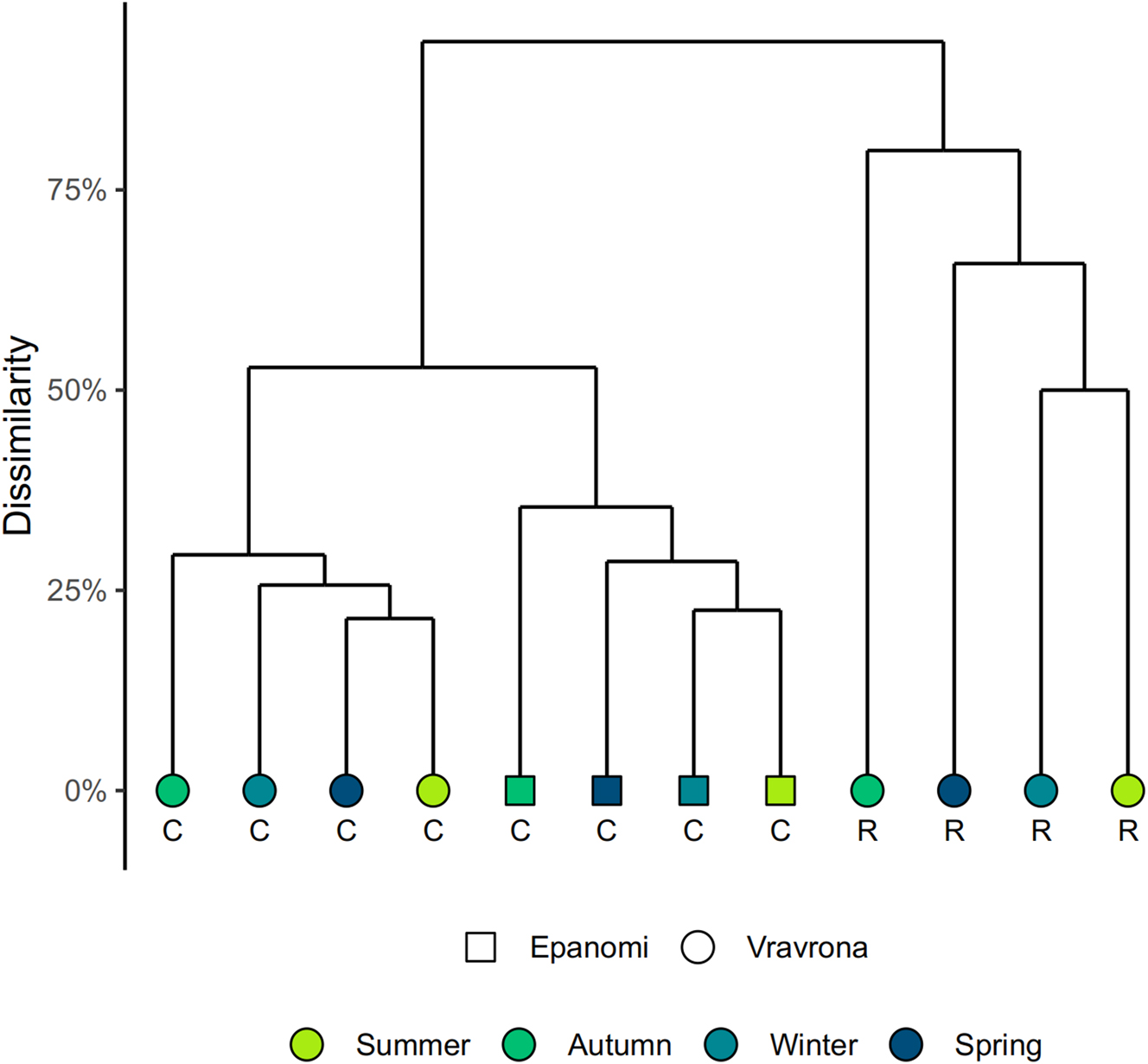
Hierarchical cluster based on Bray-Curtis dissimilarities between samples, by season. C = Cymodocea nodosa, R = Ruppia maritima.
3.6 Beta diversity
Given that the two plant hosts and the two sites showed clearly distinct epiphyte communities, seasonal beta diversity was calculated per site and plant host, in order to gain further insights into the processes behind the epiphyte community assembly. The dissimilarity between seasons was then partitioned into its turnover and nestedness components. Overall, beta diversity was low for Cymodocea at both sites, reaching values up to 0.25, indicating a low variation in species composition over the course of the year. For Ruppia, beta diversity was very high between autumn and winter (no species in common) and substantially lower between spring and summer (Figure 10). Concerning community change between seasons, in Vravrona the turnover component dominated in both Cymodocea and Ruppia epiphyte communities, indicating that the differences in species composition between seasons are dominated by species replacement (i.e. new species added to the community), rather than by species loss or species recovery (i.e. same species returning). This was especially pronounced in the autumn – winter transition of the epiphyte community on Ruppia, where the species community was completely replaced from one season to the next.
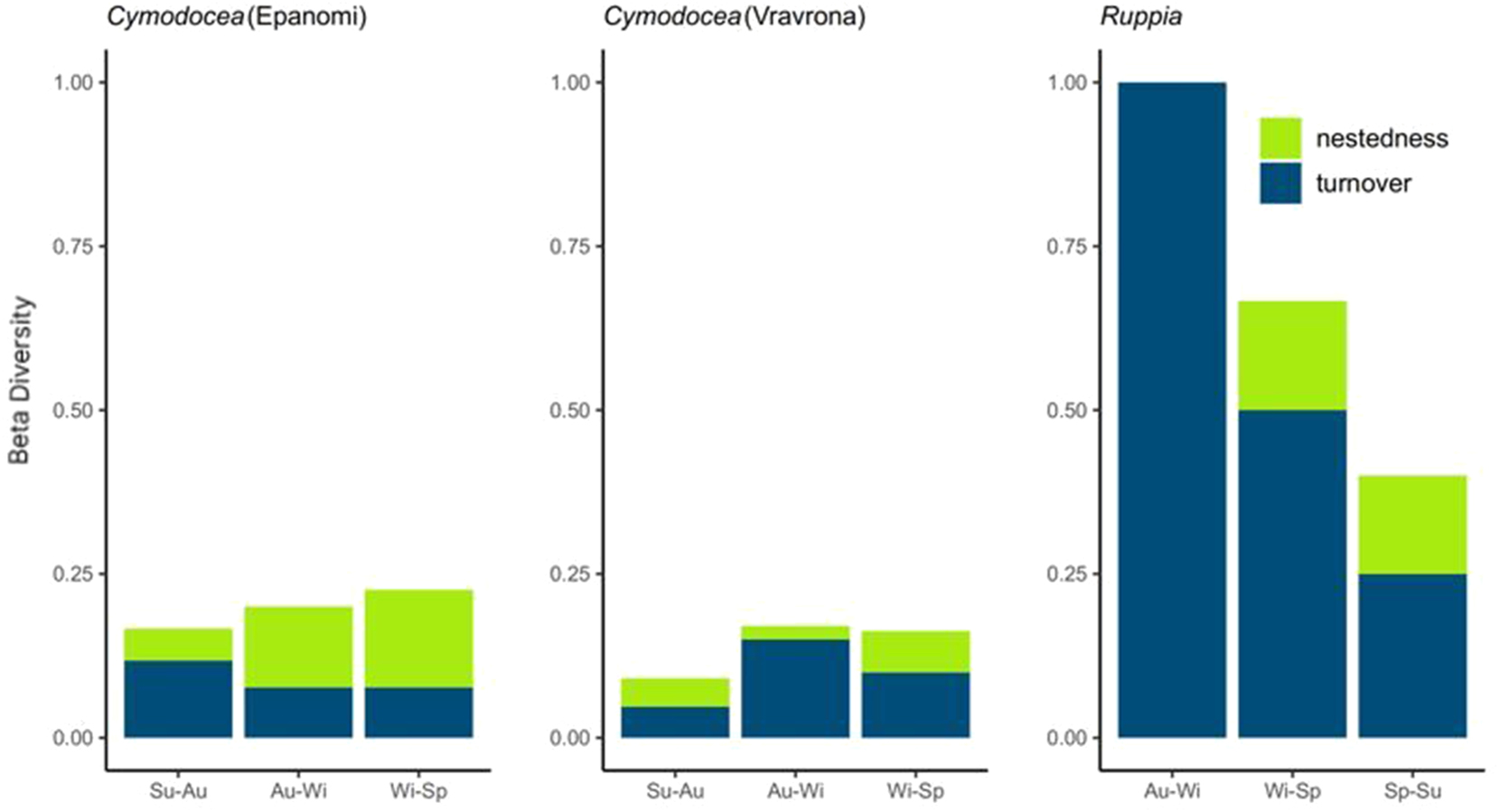
Representation of beta diversity and its components (turnover and nestedness) for each site and host. Sp = spring, Su = summer, Au = autumn, Wi = winter. Cymodocea nodosa samples began to be collected in the summer of 2022, while Ruppia maritima sampling began in the autumn of 2022.
In Epanomi, on the other hand, nestedness was the dominant component from autumn to winter and from winter to spring – only from summer to autumn turnover dominated. This indicates that in autumn different species were added to the community that did not occur in summer, while in winter species were simply lost from the autumn community (and in spring species some species were added back to the community).
4 Discussion
The data collected from seagrass meadows at the Epanomi and Vravrona sites provide a comprehensive overview of their ecological dynamics. The observed differences in density, leaf area index (LAI), epiphytes’ coverage and biomass between the two sites, as well as the differences between the two seagrasses (Ruppia and Cymodocea), suggest that these meadows are subject to varying local and seasonal environmental conditions and potential anthropogenic impacts. Such variations are critical indicators of the health and resilience of these meadows, reflecting how factors such as water quality, nutrient availability, and human activities influence their ecological status (Oliva et al. 2012; Romero et al. 2006). Identifying and understanding specific patterns is crucial for current and future conservation efforts.
4.1 Meadows’ characteristics
Meadow’s density and LAI constitute key indicators of seagrass ecosystems’ health (Papathanasiou et al. 2016), as higher densities and LAIs typically correlate with more robust and productive ecosystems, capable of supporting a diverse array of marine organisms (Schubert et al. 2018). In this study, significant differences between Epanomi and Vravrona sites reflect environmental specificities, both abiotic such as hydrodynamism, nutrient concentrations, water and sediment quality, as well as biotic (competition, grazing). Regarding pressures, both sites are characterized by the development of various activities, resulting in anthropogenic disturbances. As an example, the activities taking place in Epanomi and in Thermaikos Gulf in general, lead to increased levels of turbidity, suspended particulate matter and phytoplankton production – chlorophyll a concentration, as well as the presence of nitrates in the water column and sediment (Poulos et al. 2000). All of the above-mentioned parameters are closely related to meadow characteristics such as depth distribution, density, leaf biometry, etc.
In detail, in the study areas of Epanomi and Vravrona, a significant variation in the density of shoots was observed over the course of the year. The highest density occurred during the summer months, while a gradual decline was observed in autumn and winter, with a recovery in spring, via the formation of new shoots. The decrease in shoot density observed at both sites during autumn and winter is considered a natural response of the plant to maintain a balance between shading and photosynthetic activity, especially when light availability is limited in the temperate regions (Olesen and Sand-Jensen 1993). Previous studies on C. nodosa in the Mediterranean Sea confirm the significant influence of water temperature, solar radiation, nutrients (Mutlu et al. 2022) and salinity on shoot density. Maximum growth rates occur in late spring and summer (Reyes et al. 1995; Sghaier et al. 2017; Terrados and Ros 1992), which constitutes the period with the most favorable conditions for seagrass growth (Guidetti et al. 2002).
The leaf area index is a parameter with numerous and significant seasonal and local variations which are attributed to the annual changes of the formation and growth rates of leaves, determined by the seasonality of abiotic parameters (Enríquez et al. 2004; Máñez-Crespo et al. 2020). C. nodosa exhibits considerable plasticity in leaf size (Pérez et al. 1994) and leaf surface is one of the characteristics most affected by various environmental conditions parameters. In detail, depth (Máñez-Crespo et al. 2020), solar radiation, nutrients, salinity (Sandoval-Gil et al. 2014) and hydrodynamic conditions are strongly related to leaf production and biometry. In the areas of Epanomi and Vravrona, the lowest values of LAI were recorded during winter in both sites and the highest during summer and autumn depending on the leaves’ age and the site. The highest values of LAI for both adult (1.244 m2 m−2) and intermediate (0.793 m2 m−2) leaves were recorded in the Epanomi’s meadow. The differentiation of the two meadows during autumn is noticeable: adult leaves in Vravrona present the highest value of LAI, whereas in Epanomi LAI is low during autumn, in comparison to summer and spring in the same area. Previous studies conducted in the Mediterranean Sea (Cancemi et al. 2002; Reyes et al. 1995; Vidondo et al. 1997), refer to a peak in the development of leaf morphology observed in spring and summer (Cunha and Duarte 2007; Guidetti et al. 2002), and a decrease in autumn and winter (Mutlu et al. 2022). In general, longer and wider leaves seem to reflect the increased photosynthetic rate usually recorded during summer (Malea and Zikidou 2011), following the increased availability of nutrients in the marine environment, whereas smaller leaves characterize plants that use carbon for rhizome growth, in an effort to balance the nutrient deficit during the cold months. Furthermore, it appears that plants with large leaf surfaces occur both in undisturbed (Papathanasiou 2013) and eutrophic areas (Oliva et al. 2012; Orfanidis et al. 2010). The variety of abiotic parameters that affect the appearance and growth of leaves in an area, combined with the strong differentiation (geographical, pressures, etc.) of the project’s sites, does not allow a more accurate interpretation of the differences noticed between the two sites.
4.2 Macroalgal epiphytic community
Seagrass leaves are a temporary, but continuously renewing, substratum for epiphytic organisms of various taxonomic groups such as algae, bacteria, fungi, protozoa, invertebrates, etc. Biodiversity and particularly presence of various photosynthetic groups highlight the ecological significance of seagrass meadows in supporting marine life and maintaining ecosystem services, such as carbon sequestration, nutrient cycling, and habitat provision. These meadows act as nurseries for many marine species, thus enhancing fishery productivity and biodiversity. Among the above-mentioned groups, the epiphytic macroalgae constitute very important primary producers with a significant contribution to food webs. Furthermore, they can account for over 50 % of the standing stock in seagrass meadows and they may act as bioindicators, as they respond to environmental changes more quickly than the seagrasses themselves (Borowitzka et al. 2006).
Epiphyte coverages in Vravrona and Epanomi sites increased along the apical (older) part of the leaves, an observation that is in agreement with other studies. Reyes et al. (1998) mention that the increase in the number of epiphyte species observed along the oldest leaves of C. nodosa from the Canary Islands, from the basal to the apical segments, is in line with the tendency reported along the leaves of different seagrasses by several authors for other seagrasses.
The results of the current study show that the percentage of leaf surface being occupied by epiphytes varied among seasons, sites and seagrass hosts. The highest value of coverage with epiphytes (including flora and fauna) was recorded in Vravrona, specifically in the R. maritima meadow, during autumn (Figure 5a). However, the macroalgal community during this period was characterized by the presence of only three taxa, with low coverage (Figure 5b), whereas the main epiphytes were erect diatoms covering the whole surface.
Regarding the biomass of epiphytes on adult leaves of C. nodosa (Figure 6), significant seasonal variations were observed in Vravrona, with the highest value being recorded during autumn, followed by the spring. In Epanomi, the variations were noticeably smaller. Comparing Figure 5b and 6, no full agreement between the biomass of epiphytes and the coverage of the community of macroalgae was apparent, since many other epiphytic organisms, besides macroalgae, contribute to the biomass as well.
The diversity of epiphytic algae found on seagrass species varies considerably. Mabrouk et al. (2021) described the epiphytic macroalgal assemblages of three seagrasses (P. oceanica, C. nodosa and Halophila stipulacea) from Tunisia. Epiphyte algal assemblages were more abundant and more diversified on the leaves of P. oceanica and C. nodosa, than those on the leaves of H. stipulacea (Tracheophyta), which, according to the authors, is related to species-specific characteristics of each seagrass, such as shape and life span of the leaves, growth rate, etc. In the current study, a total of 34 taxa of epiphytic macroalgae were identified developing on C. nodosa (27 in Vravrona and 23 in Epanomi) and 7 on R. maritima. This significant differentiation between the neighboring meadows of Cymodocea and Ruppia in Vravrona is probably related to the special features of each seagrass as a substratum (leaf length and width, number of leaves, etc), as well as to the specific environmental conditions (Ruppia growing in waters of variable salinity). In addition, as mentioned by Borowitzka et al. (2006), more persistent and structurally complex seagrass species such as P. oceanica tend to have more diverse epiphyte assemblages. Therefore, it is generally accepted that short-lived seagrass species such as R. maritima are likely to be relatively depauperate in epiphyte species richness, compared to persistent seagrass species. The study of epiphytic flora in relation to leaf lifespan could provide valuable insights for evaluating our results. However, the lack of available data, particularly concerning Ruppia species, presents a significant challenge. For example, the lifespan of C. nodosa leaves varies greatly, ranging from nearly 2 to 7 months (Hemminga et al. 1999; Sghaier et al. 2011), and is influenced by factors such as leaf weight, formation rate, and latitude. In contrast, there is a notable absence of data on the lifespan of Ruppia leaves. This gap is partly due to the species’ narrow leaf blades, which make it difficult to apply leaf marking techniques (Koch et al. 2007).
As for the biodiversity of the epiphytic macroalgal assemblages of each seagrass species, our results are in agreement with other studies from the Mediterranean Sea and other areas. Specifically, regarding the C. nodosa community, Mabrouk et al. (2021) identified 36 macroalgal taxa in Tunisia, while Tsioli et al. (2021) found 19 taxa in Northern Greece.
The seasonal variations in epiphytic macroalgal diversity and coverage of C. nodosa underscore the dynamic nature of these ecosystems. The analysis of the epiphytic macroalgal community indicated that species richness is highest in spring and summer months, while the highest coverage is observed in autumn. Notably, Epanomi’s lower epiphyte coverage compared to Vravrona, particularly in autumn, suggests that Vravrona might experience nutrient levels, hydrodynamic conditions, temperature, and/or other factors that favor epiphyte growth (Borowitzka et al. 2006; Cabaço et al. 2008). In general, the seasonal and local variations in the epiphytic community can be attributed to the life cycle of the macroalgae, the growth rhythms of the plant (substratum), light, temperature, salinity, nutrient concentrations, hydrodynamic conditions, and biotic factors such as competition or grazing.
Comparing the studied sites, Vravrona was characterized by a higher coverage of leaves with epiphytes throughout the year, especially during autumn. In the macroalgal epiphytic community the class of Rhodophyceae dominated qualitatively in all samples, at both sites. The encrusting calcareous Rhodophyta belonging to the genera Hydrolithon and Pneophyllum dominated the epiphytic assemblage for C. nodosa. These species are considered among the most ubiquitous in the epiphytic assemblage of marine seagrasses. Although the short lifespan of seagrass leaves contributes to the ephemeral nature of the epiphytic community, preventing long-term succession, crustose corallines play a crucial role as “early colonizers.” They are the pioneering group that dominates after the initial settlement of bacteria and diatoms (Borowitzka et al. 2006; Mabrouk et al. 2015; 2021).
In Vravrona, Rhodophyta dominated quantitatively as well, throughout the year and in addition to macroalgae, the periphytic solitary and tube-dwelling colonial diatoms had a significant presence. In Epanomi, red algae dominated qualitatively (number of identified taxa) during all seasons, while quantitatively different macroalgal as well as microalgal groups dominated in each season. Previous studies have characterized Rhodophyta as the most common group of macroalgae on seagrass leaves (more than 90 %) (Reyes and Sansón 2001), noting that the dominance of other phyla implies unusual environmental conditions, such as nutrient seasonal enrichment resulting in an abundance of green algae or cyanobacteria, changes in energy levels and depth resulting in shifts between diatoms and cyanobacteria, etc. (Borowitzka et al. 2006). Mabrouk et al. (2021), comparing the epiphytic assemblages of three seagrasses, noticed that brown algae were well represented in the epiphytic community, reaching a maximum of 10 species on the leaves of P. oceanica and C. nodosa, while they were poorly represented on the leaves of H. stipulacea (Tracheophyta) (3 species). It has been proposed that the leaves of P. oceanica and C. nodosa are older than those of H. stipulacea and thus they offer more time to erect macroalgal species (as many brown algae are) to settle. In the same article, the low representation of green algae is mentioned, which was noticed in our work as well. Concerning the current survey, high coverage of Rhodophyta on C. nodosa leaves, especially crustose forms like Hydrolithon, indicates relatively stable and potentially nutrient-rich conditions and is often associated with good water quality. In contrast, increased dominance of Heterokontophyta (Phaeophyceae) and Chlorophyta in certain seasons, such as summer in Epanomi, can indicate much higher nutrient levels and warmer water temperatures, as these groups often respond to eutrophic conditions.
The dominant group across both seagrasses and sites consisted of fast-growing, opportunistic species, primarily filamentous, classified within ESG IIB. This pattern aligns with the ephemeral nature of seagrass leaves as substrata and is consistent with previous studies on the epiphytic community of C. nodosa (Tsioli et al. 2021). When examining the classification of identified taxa in Ecological Status Groups (Orfanidis et al. 2011), Hydrolithon cruciatum and Pneophyllum fragile (Rhodophyta) were abundant on C. nodosa leaves but absent on R. maritima leaves. These species are typically characterized as shade-adapted, slow-growing, late-successional red algal calcareous crusts that usually live as epiphytes on seaweed thalli or seagrass leaves (ESG IC). This apparent contradiction, as these species are among the first to settle on seagrass leaves (Borowitzka et al. 2006), may represent an adaptation to the transient nature of leaves as substrata.
As for the epiphytic flora growing on R. maritima, the small number of epiphytic macroalgae and the almost exclusive prevalence of erect diatoms was noteworthy, especially during autumn and winter, which could be attributed to the variability of environmental parameters in ecosystems with salinity fluctuations. In shallow estuarine areas diatoms are the dominant or the most abundant components among the epiphytic algae (McIntyre et al. 1996). Sullivan (1977) notes that epiphytic diatom growth on the leaves of Ruppia under the frame of a study conducted in New Jersey, was so great that it was visible to the naked eye – this was also observed in the Vravrona meadow during autumn. Furthermore, the author supports that the large production of epiphytic diatoms on the leaves of Ruppia may be largely dependent on the uptake of nutrients from the bottom sediments by the roots of Ruppia. As a general remark, the epiphytic diatom communities of seagrasses and seaweeds can vary significantly in space and time, as a result of abiotic environmental factors such as salinity, nutrient availability and temperature, hydrodynamic conditions, etc., biotic factors (especially herbivory action) and intrinsic factors, such as motility (da Rosa and Copertino 2022; Snoeijs 1994; Stanca and Parsons 2021). Concerning the dominance of erect diatoms, previous studies conducted on the epiphytic diatom communities of seaweeds have shown that the growth forms of epiphytic diatoms depend on morphological differences in the thalli of marine seaweeds: those with highly branched or narrow thalli provide a large number of microhabitats, offering several opportunities for colonization of erect forms. Thalli with flat, smooth, or leafy surfaces provide better substratum for adnate forms (da Silva Costa et al. 2016; Totti et al. 2009). Thus, the high abundance of erect diatoms to R. maritima’s leaves could be related to the morphology of the host (narrow leaves).
Concerning the seasonal succession of macroalgal epiphytes, beta diversity analyses suggest that community assembly in the two geographic locations seem to be driven by different underlying processes. In Vravrona, the turnover component largely dominated in both host species, whereas in Epanomi the nestedness component dominated. While it is difficult to disentangle whether these differences are due to random processes such as chance colonization or extinction or due to environmental filtering processes, the fact that both species in Vravrona show a similar pattern could be an indication for deterministic processes underlying these changes (Chase and Myers 2011; Whippo et al. 2018). Beta diversity was much higher on Ruppia than on Cymodocea, however, this is in part also due to the low number of taxa on Ruppia, especially in autumn and winter.
The study of seagrass meadows at Epanomi and Vravrona revealed significant differences in density, leaf area index (LAI), epiphyte coverage, and biomass between sites and species (C. nodosa and R. maritima), reflecting varying environmental conditions and anthropogenic impacts (Oliva et al. 2012; Romero et al. 2006). Higher densities and LAIs indicate more robust ecosystems (Schubert et al. 2018). Seasonal variations in shoot density and LAI, align with changes in light availability and environmental conditions (Enríquez et al. 2004; Máñez-Crespo et al. 2020; Olesen and Sand-Jensen 1993; Reyes et al. 1995; Sghaier et al. 2017; Terrados and Ros 1992). Epiphytic communities also vary by season and site, with more abundant epiphytes on C. nodosa due to species-specific traits (Borowitzka et al. 2006; Mabrouk et al. 2021). Epiphytes affect the photosynthetic abilities of C. nodosa and R. maritima by shading, reducing light availability and photosynthetic efficiency, with peak growth in warmer months (Burnell et al. 2014; Mazzella et al. 1989; Wetzel and Penhale 1979). The significant seasonal variation in epiphytic coverage in Vravrona indicates fluctuating environmental conditions such as temperature, light availability, and nutrient levels throughout the year. For example, warmer months with increased sunlight and higher temperatures can enhance epiphytic growth, while cooler conditions in autumn and winter can reduce their abundance (Hillebrand and Sommer 1997). The dominance of diatoms during autumn and winter suggests these cooler seasons create conditions favorable for their growth. Diatoms thrive in nutrient-rich environments, and cooler temperatures often lead to nutrient dynamics that support their proliferation (Smetacek 1985). During these seasons, nutrient levels such as nitrogen and phosphorus might increase in the upper water layers, further promoting diatom growth (Cloern 1996). In addition, the presence of the alien species C. cylindracea in Vravrona points to significant ecological shifts. This invasive green alga can rapidly spread and outcompete native species, altering habitat structures and impacting nutrient cycles (Piazzi and Ceccherelli 2006). The establishment of C. cylindracea suggests changes in local conditions, possibly due to factors like climate change and nutrient enrichment from human activities (Bulleri and Benedetti-Cecchi 2008).
Sites with higher year-round epiphyte coverage, like Vravrona, offer more stable habitats, supporting diverse epiphytes and fauna, whereas Epanomi’s lower coverage may suggest less stable conditions or different disturbances. These findings highlight the importance of monitoring environmental conditions and anthropogenic impacts to preserve seagrass meadow biodiversity and ecosystem function.
5 Conclusions
The study of seagrass meadows at Epanomi and Vravrona reveals significant differences in density, leaf area index (LAI), epiphyte coverage, and biomass between the sites and between the seagrass species C. nodosa and R. maritima. These differences probably reflect different environmental conditions and anthropogenic influences on the sites seasonall. Epanomi experiences higher disturbances, which lead to increased turbidity and nutrient levels. In contrast, Vravrona exhibits nutrient-rich conditions and distinct hydrodynamic factors that likely promote epiphyte growth.
Seasonal variations in shoot density and LAI, with peaks in summer and autumn and declines in winter, align with changes in light availability and environmental conditions. The macroalgal epiphytic community also varied by season and site, with more abundant epiphytes on C. nodosa due to species-specific traits. Vravrona’s epiphytic macroalgal community, dominated by Rhodophyceae, suggests stable and nutrient-rich conditions, while Epanomi’s lower coverage and biomass indicate less stable conditions or different disturbances.
These findings emphasize the importance of monitoring environmental conditions and anthropogenic impacts to preserve seagrass meadow biodiversity and ecosystem function. Identifying specific environmental drivers and the effects of climate change will aid in developing effective conservation strategies. Future research should focus on the processes driving community assembly and the environmental conditions affecting seagrass health, ensuring the long-term sustainability of these vital marine ecosystems.
Funding source: INTERINSTITUTIONAL POSTGRADUATE PROGRAM NKUA-HCMR, “OCEANOGRAPHY AND MANAGEMENT OF THE MARINE ENVIRONMENTâ€
About the authors

Vaia Myloneli holds an Integrated Master’s degree in Agricultural Science with a specialization in the marine environment and a Master’s degree in the Inter-Institutional MSc program Oceanography and Management of the Marine Environment (NKUA & HCMR). Her research focuses on the impact of anthropogenic activities on benthic ecosystems, marine ecology, and seagrass meadows.

Georgios D. Dorovinis holds a BSc degree from the Department of Biology of the University of Patras and a MSc degree in Oceanography and Management of the Marine Environment from the National and Kapodistrian University of Athens, in collaboration with the Hellenic Centre for Marine Research. His main research interests include the study of pollution and epiphytic communities in seagrass ecosystems.

Dr. Sarah Faulwetter is a benthic ecologist and taxonomist. She is currently a postdoc in the University of Patras working on the impacts of marine aquaculture on the benthic ecosystem. Her research interests focus on the effect of anthropogenic activities on benthic ecosystems in relation to water quality, community ecology and ecosystem functioning.

Ioannis-Dimosthenis S. Adamakis, an Associate Professor at the National and Kapodistrian University of Athens, specializes in plant biology with an emphasis on cell structure and responses to environmental stressors. His research integrates microscopy, molecular biology, and biochemistry to explore various aspects of plant biology, including the study of seagrasses.

Dr. Anastasia Tsirika, currently working as laboratory teaching staff in the School of Agriculture at the Aristotle University of Thessaloniki, is a biologist with a Master’s degree in Hydrobiology & Aquacultures and a PhD in Marine Botany. Her research focuses on water quality and bioindicators, taxonomy, distribution and seasonality of marine macrophytes and mapping of benthic communities.
Acknowledgments
The valuable comments of the anonymous reviewers are acknowledged.
-
Research ethics: The procedures were in accordance with the national laws.
-
Informed consent: Not applicable.
-
Author contributions: V. Myloneli: research; data curation; formal analysis; writing – review and editing. G.D. Dorovinis: research; data curation; formal analysis; writing – review and editing. S. Faulwetter: conceptualization; methodology; review and editing. I.-D.S. Adamakis: conceptualization; methodology; supervision; writing – review and editing. A. Tsirika: conceptualization; research; data curation; formal analysis; writing – original draft preparation; writing – review and editing. All authors have accepted responsibility for the entire content of this manuscript and approved its submission.
-
Use of Large Language Models, AI and Machine Learning Tools: None declared.
-
Conflict of interest: The authors state no conflict of interest.
-
Research funding: This study was undertaken as part of the master’s theses of V. Myloneli and G.D. Dorovinis and received financial support via funds from the Interinstitutional Postgraduate Program NKUA – HCMR, “Oceanography and Management of the Marine Environment”.
-
Data availability: Data are available from the corresponding author upon request.
References
Apostoloumi, C., Malea, P., and Kevrekidis, T. (2021). Principles and concepts about seagrasses: towards a sustainable future for seagrass ecosystems. Mar. Pollut. Bull. 173: 112–936, https://doi.org/10.1016/j.marpolbul.2021.112936.Search in Google Scholar PubMed
Balata, D., Nesti, U., Piazzi, L., and Cinelli, F. (2007). Patterns of spatial variability of seagrass epiphytes in the north-west Mediterranean Sea. Mar. Biol. 151: 2025–2035, https://doi.org/10.1007/s00227-006-0559-y.Search in Google Scholar
Balata, D., Piazzi, L., and Cinelli, F. (2008). Response of a macroalgal community to nutrient enrichment in the Mediterranean Sea: Inferences from a field experiment. Mar. Pollut. Bull. 56: 991–998.Search in Google Scholar
Ballesteros, E., Martin, D., and Uriz, M.J. (1992). Biological activity of extracts from some Mediterranean macrophytes. Bot. Mar. 35: 481–485, https://doi.org/10.1515/botm.1992.35.6.481.Search in Google Scholar
Baselga, A., Orme, D., Villeger, S., De Bortoli, J., Leprieur, F., Logez, M., Martinez-Santalla, S., Martin-Devasa, R., Gomez-Rodriguez, C., and Crujeiras, R. (2023). Betapart: partitioning beta diversity into turnover and nestedness components. R package version 1.6, https://CRAN.R-project.org/package=betapart.Search in Google Scholar
Borowitzka, M.A., Lavery, P.S., and van Keulen, M. (2006) Epiphytes of seagrasses. In: Larkum, A.W.D., Orth, R.J., and Duarte, C.M. (Eds.). Seagrasses: biology, ecology and conservation. Springer, Dordrecht, pp. 441–462.10.1007/1-4020-2983-7_19Search in Google Scholar
Boudouresque, C.F. (1971). Méthodes d’étude qualitative et quantitative du benthos (en particulier du phytobenthos). Tethys 3: 79–104.Search in Google Scholar
Boudouresque, C.F. and Belsher, T. (1979). Une méthode de détermination de l’aire minimale qualitative. Rapp. P.V. Réun. Commiss. Internation. Explor. Sci. Médit. 25: 273–275.Search in Google Scholar
Brush, M.J. and Nixon, S.W. (2003). Biomass layering and metabolic activity of epiphytes on eelgrass (Zostera marina L.) shoots. Mar. Ecol. Progr. Ser. 257: 83–95.Search in Google Scholar
Bulleri, F. and Benedetti-Cecchi, L. (2008). Facilitation of the green alga Caulerpa racemosa by resident algal turfs: evidence from in situ experiments. Mar. Biol. 154: 341–350.Search in Google Scholar
Burnell, O.W., Russell, B.D., Irving, A.D., and Connell, S.D. (2014). Seagrass response to CO2 contingent on epiphytic algae: indirect effects can overwhelm direct effects. Oecologia 176: 871–882, https://doi.org/10.1007/s00442-014-3054-z.Search in Google Scholar PubMed
Cabaço, S., Machás, R., Vieira, V., and Santos, R. (2008). Impacts of urban wastewater discharge on seagrass meadows (Zostera noltii). Estuarine. Coast Shelf Sci. 78: 1–13.10.1016/j.ecss.2007.11.005Search in Google Scholar
Cancemi, G., Buia, M.C., and Mazzella, L. (2002). Structure and growth dynamics of Cymodocea nodosa meadows. Scientia Marina 66: 365–373, https://doi.org/10.3989/scimar.2002.66n4365.Search in Google Scholar
Chase, J.M. and Myers, J.A. (2011). Disentangling the importance of ecological niches from stochastic processes across scales. Philos. Trans. R. Soc. B Biol. Sci. 366: 2351–2363, https://doi.org/10.1098/rstb.2011.0063.Search in Google Scholar PubMed PubMed Central
Cloern, J.E. (1996). Phytoplankton bloom dynamics in coastal ecosystems: a review with some general lessons from sustained investigation of San Francisco Bay, California. Rev. Geophys. 34: 127–168, https://doi.org/10.1029/96rg00986.Search in Google Scholar
Collado-Vides, L., Caballero-Aragón, H., and Blair, S. (2007). Epiphytic macroalgae on Thalassia testudinum in northern Yucatan Peninsula: biomass and composition. Bot. Mar. 50: 191–200.Search in Google Scholar
Costanza, R., d’Arge, R., de Groot, R., Farber, S., Grasso, M., Hannon, B., Limburg, K., Shadid, N., O’Neill, R.V., Paruelo, J., et al.. (1997). The value of the world’s ecosystem services and natural capital. Nature 387: 253–260, https://doi.org/10.1038/387253a0.Search in Google Scholar
Cunha, A.H. and Duarte, C.M. (2007). Biomass and leaf dynamics of Cymodocea nodosa in the Ria Formosa lagoon, South Portugal. Bot. Mar. 50: 1–7, https://doi.org/10.1515/bot.2007.001.Search in Google Scholar
da Rosa, V.C. and Copertino, M. (2022). Diversity and variation of epiphytic diatoms on Ruppia maritima L., related to anthropogenic impact in an estuary in Southern Brazil. Diversity 14: 787, https://doi.org/10.3390/d14100787.Search in Google Scholar
da Rosa, V.C., Martínez-Crego, B., Santos, R.O.P., Odebrecht, C., and Copertino, M.S. (2023). Temporal variation in diatom communities associated to sediments of impacted versus non-impacted seagrass meadows of an estuarine lagoon. Aquat. Bot. 189: 103701.10.1016/j.aquabot.2023.103701Search in Google Scholar
da Silva Costa, M.M., Pereira, S.M.B., da Silva-Cunha, M.da G.G., de Arruda, P.C., and Eskinazi-Leça, E. (2016). Community structure of epiphytic diatoms on seaweeds in Northeastern Brazil. Bot. Mar. 59: 231–240, https://doi.org/10.1515/bot-2015-0014.Search in Google Scholar
Dauby, P. and Poulicek, M. (1995). Methods for removing epiphytes from seagrasses: SEM observations on treated leaves. Aquat. Bot. 52: 217–228, https://doi.org/10.1016/0304-3770(95)00500-5.Search in Google Scholar
den Hartog, C. and Kuo, J. (2006) Taxonomy and biogeography of seagrasses. In: Larkum, A.W.D., Orth, R.J., and Duarte, C.M. (Eds.). Seagrasses: biology, ecology and conservation. Springer, Dordrecht, pp. 1–23.10.1007/1-4020-2983-7_1Search in Google Scholar
Dhont, F. and Coppejans, E. (1977). Résultats d’une étude d’ aire minima des peuplements algaux photophiles sur substrat rocheux à Port-Cros et a Banyuls (France). Rapp. P.V. Reun. CIESM 24: 141–142.Search in Google Scholar
Duarte, C.M. (2002). The future of seagrass meadows. Environ. Conserv. 29: 192–206, https://doi.org/10.1017/s0376892902000127.Search in Google Scholar
Enríquez, S., Marbà, N., Cebriàn, J., and Duarte, C.M. (2004). Annual variation in leaf photosynthesis and leaf nutrient content of four Mediterranean seagrasses. Bot. Mar. 47: 295–306, https://doi.org/10.1515/bot.2004.035.Search in Google Scholar
Francour, P., Ganteaume, A., and Poulain, M. (1999). Effects of boat anchoring in Posidonia oceanica seagrass beds in the Port-Cros national Park (north-western Mediterranean Sea). Aquat. Conserv. Mar. Freshw. Ecosyst. 9: 391–400, https://doi.org/10.1002/(sici)1099-0755(199907/08)9:4<391::aid-aqc356>3.3.co;2-#.10.1002/(SICI)1099-0755(199907/08)9:4<391::AID-AQC356>3.3.CO;2-#Search in Google Scholar
García-Redondo, A., Pérez, M., Romero, J., and Alcoverro, T. (2019). A functional approach to study species invasion processes and impacts in seagrass meadows. Mar. Pollut. Bull. 146: 244–252.Search in Google Scholar
Giraud, G. (1979). Sur une méthode de mesure et de comptage des structures foliaires de Posidonia oceanica (Linnaeus) Delile. Bull. Mus. Hist. Nat. Mars. 39: 33–39.Search in Google Scholar
Guidetti, P., Lorenti, M., Buia, M.C., and Mazzella, L. (2002). Temporal dynamics and biomass partitioning in three Adriatic seagrass apecies: Posidonia oceanica, Cymodocea nodosa, Zostera marina. Mar. Ecol. 23: 51–67, https://doi.org/10.1046/j.1439-0485.2002.02722.x.Search in Google Scholar
Hemminga, M.A., Marbà, N., and Stapel, J. (1999). Leaf nutrient resorption, leaf lifespan and the retention of nutrients in seagrass systems. Aquat. Bot. 65: 141–158.10.1016/S0304-3770(99)00037-6Search in Google Scholar
Hillebrand, H. and Sommer, U. (1997). Response of epilithic microphytobenthos of the western Baltic Sea to in situ experiments with nutrient enrichment. Mar. Ecol. Progr. Ser. 160: 35–46, https://doi.org/10.3354/meps160035.Search in Google Scholar
Katsoulis, B.D., Malakou, M., and Kallimanis, A.S. (2019). Assessing the ecological status of Mediterranean coastal wetlands. J. Environ. Manage. 230: 37–45.Search in Google Scholar
Koch, E.W. and Dawes, C.J. (1991). Ecological studies of Ruppia maritima L. ditch grass in a subtropical estuary. Aquat. Bot. 40: 389–399.10.1016/0304-3770(91)90083-HSearch in Google Scholar
Koch, M.S., Schopmeyer, S.A., Kyhn-Hansen, C., Madden, C.J., and Peters, J.S. (2007). Tropical seagrass species tolerance to hypersalinity stress. Aquat. Bot. 86: 14–24, https://doi.org/10.1016/j.aquabot.2006.08.003.Search in Google Scholar
Littler, M.M. and Littler, D.S. (1980). The evolution of thallus form and survival strategies in benthic marine macroalgae: field and laboratory tests of a functional form model. Am. Nat. 116: 25–44, https://doi.org/10.1086/283610.Search in Google Scholar
Mabrouk, L., Ben Brahim, M., Hamza, A., and Bradai, M.-N. (2015). Temporal and spatial zonation of macroepiphytes on Posidonia oceanica (L.) Delile leaves in a meadow off Tunisia. Mar. Ecol. 6: 77–92, https://doi.org/10.1111/maec.12118.Search in Google Scholar
Mabrouk, L., Ben Brahim, M., Jebara, A., and Jribi, I. (2021). Comparison of epiphyte algal assemblages on the leaves of marine seagrasses Posidonia oceanica (L.) Delile, Cymodocea nodosa (Ucria) Asch, and the lessepsian Halophila stipulacea (Forssk.) Asch in Chebba (East of Tunisia). Mar. Ecol. 42: e12642, https://doi.org/10.1111/maec.12642.Search in Google Scholar
Malea, P. and Zikidou, C. (2011). Temporal variation in biomass partitioning of the seagrass Cymodocea nodosa at the Gulf of Thessaloniki, Greece. J. Biol. Res. Thessalon. 15: 75–90.Search in Google Scholar
Máñez-Crespo, J., Tuya, F., Fernández-Torquemada, Y., Royo, L., del Pilar-Ruso, Y., Espino, F., Manent, P., Antich, L., Castejón-Silvo, I., Curbelo, L., et al.. (2020). Seagrass Cymodocea nodosa across biogeographical regions and times: differences in abundance, meadow structure and sexual reproduction. Mar. Environ. Res. 162: 105159, https://doi.org/10.1016/j.marenvres.2020.105159.Search in Google Scholar PubMed
Mazzella, L., Scipione, M.B., and Buia, M.C. (1989). Spatio-temporal distribution of algal and animal communities in a Posidonia oceanica meadow. Mar. Ecol. 10: 107–129, https://doi.org/10.1111/j.1439-0485.1989.tb00069.x.Search in Google Scholar
McIntyre, H., Geider, R., and Miller, D. (1996). Microphytobenthos: the ecological role of the “secret garden” of unvegetated, shallow-water marine habitats. I. Distribution, abundance and primary production. Estuaries Coasts 19: 186–201, https://doi.org/10.2307/1352224.Search in Google Scholar
Montefalcone, M., Albertelli, G., Bianchi, C.N., and Morri, C. (2007). Urban seagrass: status of Posidonia oceanica facing the Genoa city waterfront (Italy) and implications for management. Mar. Pollut. Bull. 54: 206–213, https://doi.org/10.1016/j.marpolbul.2006.10.005.Search in Google Scholar PubMed
Mutlu, E., Olguner, C., Özvarol, Y., and Gökoğlu, M. (2022). Spatiotemporal biometrics of Cymodocea nodosa in a western Turkish Mediterranean coast. Biologia 77: 649–670, https://doi.org/10.1007/s11756-021-00953-0.Search in Google Scholar
Oksanen, J., Simpson, G., Blanchet, F., Kindt, R., Legendre, P., Minchin, P., O’Hara, R., Solymos, P., Stevens, M., Szoecs, E., et al.. (2022). vegan: community ecology package. R package version 2: 6–4, https://CRAN.R-project.org/package=vegan.Search in Google Scholar
Olesen, B. and Sand-Jensen, K. (1993). Seasonal acclimatization of eelgrass Zostera marina growth to light. Mar. Ecol. Progr. Ser. 94: 91–99, https://doi.org/10.3354/meps094091.Search in Google Scholar
Oliva, S., Mascaró, O., Llagostera, I., Pérez, M., and Romero, J. (2012). Selection of metrics based on the seagrass Cymodocea nodosa and development of a biotic index (CYMOX) for assessing ecological status of coastal and transitional waters. Estuar. Coast Shelf Sci. 114: 7–17, https://doi.org/10.1016/j.ecss.2011.08.022.Search in Google Scholar
Orfanidis, S., Panayotidis, P., and Stamatis, N. (2001). Ecological evaluation of transitional and coastal waters: a marine benthic macrophytes-based model. Mediterr. Mar. Sci. 2: 45–65, https://doi.org/10.12681/mms.266.Search in Google Scholar
Orfanidis, S., Papathanasiou, V., Gounaris, S., and Theodosiou, T.H. (2010). Size distribution approaches for monitoring and conservation of coastal Cymodocea habitats. Aquat. Conserv. Mar. Freshw. Ecosyst. 20: 177–188, https://doi.org/10.1002/aqc.1069.Search in Google Scholar
Orfanidis, S., Panayotidis, P., and Ugland, K. (2011). Ecological evaluation index continuous formula (EEI-c) application: a step forward for functional groups, the formula and reference condition values. Mediterr. Mar. Sci. 12: 199–232, https://doi.org/10.12681/mms.60.Search in Google Scholar
Orth, R.J., Carruthers, T.J., Dennison, W.C., Duarte, C.M., Fourqurean, J.W., Heck, K.L., Hughes, A.R., Kendrick, G.A., Kenworthy, W.J., Olyarnic, S., et al.. (2006). A global crisis for seagrass ecosystems. Bioscience 56: 987–996, https://doi.org/10.1641/0006-3568(2006)56[987:agcfse]2.0.co;2.10.1641/0006-3568(2006)56[987:AGCFSE]2.0.CO;2Search in Google Scholar
Papadimitriou, T., Kagalou, I., and Leonardos, I. (2012). Water quality monitoring in a Mediterranean coastal wetland. Environ. Monit. Assess. 184: 4613–4627.10.1007/s10661-011-2169-5Search in Google Scholar
Papathanasiou, V. (2013). Cymodocea nodosa as a bioindicator of coastal habitat quality: an integrative approach from organism to community scale, Thesis. Plymouth, University of Plymouth.Search in Google Scholar
Papathanasiou, V., Orfanidis, S., and Brown, M.T. (2016). Cymodocea nodosa metrics as bioindicators of anthropogenic stress in N. Aegean, Greek coastal waters. Ecol. Indic. 63: 61–70, https://doi.org/10.1016/j.ecolind.2015.11.059.Search in Google Scholar
Pardi, G., Piazzi, L., and Cinelli, F. (2006). Macroalgal functional groups on Cymodocea nodosa meadows in the north-west Mediterranean Sea: temporal variability and relationships with shoot density. Bot. Mar. 49: 19–26.Search in Google Scholar
Pérez, M., Duarte, C.M., Romero, J., Sand-Jensen, K., and Alcoverro, T. (1994). Growth plasticity in Cymodocea nodosa stands: the importance of nutrient supply. Aquat. Bot. 47: 249–264, https://doi.org/10.1016/0304-3770(94)90056-6.Search in Google Scholar
Pergent, G., Pergent-Martini, C., and Boudouresque, C.F. (1995). Utilization de l’herbier a Posidonia oceanica comme l’indicateur biologique de la qualitè du milieu littoral en Mediterrannée: état des connoissances. Mesogée 54: 3–27.Search in Google Scholar
Piazzi, L. and Ceccherelli, G. (2006). Persistence of biological invasion effects: recovery of macroalgal assemblages after removal of Caulerpa racemosa var. cylindracea. Estuar. Coast Shelf Sci. 68: 455–461, https://doi.org/10.1016/j.ecss.2006.02.011.Search in Google Scholar
Piazzi, L., Balata, D., and Ceccherelli, G. (2016). Epiphyte assemblages of the Mediterranean seagrass Posidonia oceanica: an overview. Mar. Ecol. 37: 3–41, https://doi.org/10.1111/maec.12331.Search in Google Scholar
Poulos, S., Chronis, G., Collins, M.B., and Lykousis, V. (2000). Thermaikos Gulf coastal system, NW Aegean Sea: an overview of water/sediment fluxes in relation to air–land–ocean interactions and human activities. J. Mar. Syst. 25: 47–76, https://doi.org/10.1016/s0924-7963(00)00008-7.Search in Google Scholar
Ramfos, A., Gazis, A., and Katselis, G. (2012). Development and evaluation of an automated digital image analysis software for obtaining seagrass leaf metrics. Bot. Mar. 55: 601–610, https://doi.org/10.1515/bot-2012-0132.Search in Google Scholar
Reyes, J. and Sansón, M. (2001). Biomass and production of the epiphytes on the leaves of Cymodocea nodosa in the Canary Islands. Bot. Mar. 44: 307–313, https://doi.org/10.1515/bot.2001.039.Search in Google Scholar
Reyes, J., Sansón, M., and Afonso-Carrillo, J. (1995). Leaf phenology, growth and production of the seagrass Cymodocea nodosa at El Médano (South of Tenerife, canary Islands). Bot. Mar. 38: 457–465, https://doi.org/10.1515/botm.1995.38.1-6.457.Search in Google Scholar
Reyes, J., Sansón, M., and Afonso-Carrillo, J. (1998). Distribution of the epiphytes along the leaves of Cymodocea nodosa in the canary Islands. Bot. Mar. 41: 543–551, https://doi.org/10.1515/botm.1998.41.1-6.543.Search in Google Scholar
Romero, J., Lee, K.-S., Pérez, M., Mateo, M., and Alcoverro, T. (2006) Nutrient dynamics in seagrass ecosystems. In: Larkum, A.W.D., Orth, R.J., and Duarte, C.M. (Eds.). Seagrasses: biology, ecology and conservation. Springer, Dordrecht, pp. 227–254.10.1007/1-4020-2983-7_9Search in Google Scholar
Sandoval-Gil, J.M., Ruiz, J.M., Marín-Guirao, L., Bernardeau-Esteller, J., and Sánchez-Lizaso, J.L. (2014). Ecophysiological plasticity of shallow and deep populations of the Mediterranean seagrasses Posidonia oceanica and Cymodocea nodosa in response to hypersaline stress. Mar. Environ. Res. 95: 39–61, https://doi.org/10.1016/j.marenvres.2013.12.011.Search in Google Scholar PubMed
Schubert, N., Freitas, C., Silva, A., Costa, M.M., Barrote, I., Horta, R.A., Rodrigues, A.C., Santos, R., and Silva, J. (2018). Photoacclimation strategies in northeastern Atlantic seagrasses: Integrating responses across plant organizational levels. Sci. Rep. 8, https://doi.org/10.1038/s41598-018-33259-4.Search in Google Scholar PubMed PubMed Central
Sghaier, Y.R., Zakhama-Sraieb, R., and Charfi-Cheikhrouha, F. (2011). Primary production and biomass in a Cymodocea nodosa meadow in the Ghar El Melh lagoon, Tunisia. Bot. Mar. 54: 411–418, https://doi.org/10.1515/bot.2011.045.Search in Google Scholar
Sghaier, Y.R., Zakhama-Sraieb, R., and Charfi-Cheikhrouha, F. (2017). Spatio-temporal dynamics and biomass of Cymodocea nodosa in Bekalta (Tunisia, Southern Mediterranean Sea). Mar. Ecol. 38: e12383, https://doi.org/10.1111/maec.12383.Search in Google Scholar
Smetacek, V. (1985). Role of sinking in diatom life-history cycles: ecological, evolutionary and geological significance. Mar. Biol. 84: 239–251, https://doi.org/10.1007/bf00392493.Search in Google Scholar
Snoeijs, P. (1994). Distribution of epiphytic diatom species composition, diversity and biomass on different macroalgal hosts along seasonal and salinity gradients in the Baltic Sea. Diatom Res. 9: 189–211, https://doi.org/10.1080/0269249x.1994.9705296.Search in Google Scholar
Stanca, E. and Parsons, M.L. (2021). Examining the dynamic nature of epiphytic microalgae in the Florida Keys: what factors influence community composition? J. Exp. Mar. Biol. Ecol. 538: 151538, https://doi.org/10.1016/j.jembe.2021.151538.Search in Google Scholar PubMed PubMed Central
Sullivan, M.J. (1977). Edaphic diatom communities associated with Spartina alterniflora and S. patens in New Jersey. Hydrobiologia 52: 207–211, https://doi.org/10.1007/bf00036444.Search in Google Scholar
Terrados, J. and Ros, J.D. (1992). Growth and primary production of Cymodocea nodosa (Ucria) Ascherson in a Mediterranean coastal lagoon: the mar menor (SE Spain). Aquat. Bot. 43: 63–74, https://doi.org/10.1016/0304-3770(92)90014-a.Search in Google Scholar
Totti, C., Poulin, M., Romagnoli, T., Perrone, C., Pennesi, C., and De Stefano, M. (2009). Epiphytic diatom communities on intertidal seaweeds from Iceland. Polar Biol. 32: 1681–1691, https://doi.org/10.1007/s00300-009-0668-4.Search in Google Scholar
Tsioli, S., Papathanasiou, V., Rizouli, A., Kosmidou, M., Katsaros, C., Papastergiadou, E., Küpper, F.C., and Orfanidis, O. (2021). Diversity and composition of algal epiphytes on the Mediterranean seagrass Cymodocea nodosa: a scale-based study. Bot. Mar. 64: 101–118, https://doi.org/10.1515/bot-2020-0057.Search in Google Scholar
Tsirika, A., Skoufas, G., and Haritonidis, S. (2007). Seasonal and bathymetric variations of epiphytic macroflora on Posidonia oceanica (L.) Delile leaves in the national marine Park of Zakynthos (Greece). Mar. Ecol. 28: 146–153, https://doi.org/10.1111/j.1439-0485.2007.00170.x.Search in Google Scholar
Van der Ben, D. (1971). Les épiphytes des feuilles de Posidonia oceanica Delile sur les côtes françaises de la Méditerranée. Inst. R. Sci. Nat. Belg., Mem. 168: 101.Search in Google Scholar
Verhoeven, J.T. (1980). The ecology of Ruppia-dominated communities in western Europe. I. Distribution of Ruppia representatives in relation to their autecology. Aquat. Bot. 6: 197–268, https://doi.org/10.1016/0304-3770(79)90064-0.Search in Google Scholar
Vidondo, B., Duarte, C., Middelboe, A., Stefansen, K., Lützen, T., and Nielsen, S. (1997). Dynamics of a landscape mosaic: size and age distributions, growth and demography of seagrass Cymodocea nodosa patches. Mar. Ecol. Progr. Ser. 158: 131–138, https://doi.org/10.3354/meps158131.Search in Google Scholar
Waycott, M., Duarte, C.M., Carruthers, T.J., Orth, R.J., Dennison, W.C., Olyarnik, S., Calladine, A., Fourqurean, J.W., Heck, K.L.Jr, Hughes, A.R., et al.. (2009). Accelerating loss of seagrasses across the globe threatens coastal ecosystems. Proc. Natl. Acad. Sci. 106: 12377–12381, https://doi.org/10.1073/pnas.0905620106.Search in Google Scholar PubMed PubMed Central
Wetzel, R.G. and Penhale, P.A. (1979). Transport of carbon and excretion of dissolved organic carbon by leaves and roots/rhizomes in seagrasses and their epiphytes. Aquat. Bot. 6: 149–158, https://doi.org/10.1016/0304-3770(79)90058-5.Search in Google Scholar
Whippo, R., Knight, N.S., Prentice, C., Cristiani, J., Siegle, M.R., and O’Connor, M.I. (2018). Epifaunal diversity patterns within and among seagrass meadows suggest landscape-scale biodiversity processes. Ecosphere 9: e02490, https://doi.org/10.1002/ecs2.2490.Search in Google Scholar
© 2025 the author(s), published by De Gruyter, Berlin/Boston
This work is licensed under the Creative Commons Attribution 4.0 International License.
Articles in the same Issue
- Frontmatter
- In this issue
- Physiology and Ecology
- Bacteria-released algal growth and morphogenesis factors regenerate axenic calli derived from the macroalga Ulva (Chlorophyta) and change the fatty acid profile
- Annual cycle and ecological interactions of macroalgal epiphytes with Cymodocea nodosa and Ruppia maritima in Epanomi and Vravrona, Greece
- Past and present fucalean diversity in the island of Marettimo, Egadi Islands Marine Protected Area (Central Mediterranean, Italy)
- Taxonomy/Phylogeny and Biogeography
- Morphological and molecular characterization of Chnoospora minima (Scytosiphonaceae, Ectocarpales) along Mexican coasts, with the description of C. ramosissima sp. nov.
- Phylogeography of the marine benthic alga Gracilaria salicornia (Gracilariales, Rhodophyta) in Southeast Asia
- Unveiling a novel species, Pachymeniopsis shinchonai sp. nov., from Korea, with reference to phylogenetic relationships within the Grateloupiaceae (Halymeniales, Rhodophyta)
- First record of the red alga Ceramium pallidum (Ceramiales, Ceramiaceae) for the Mediterranean Sea
- Chemistry and Applications
- Chemical profiling of seaweeds of the Arabian Gulf by liquid chromatography-mass spectrometry and in-silico screening against MPOX
Articles in the same Issue
- Frontmatter
- In this issue
- Physiology and Ecology
- Bacteria-released algal growth and morphogenesis factors regenerate axenic calli derived from the macroalga Ulva (Chlorophyta) and change the fatty acid profile
- Annual cycle and ecological interactions of macroalgal epiphytes with Cymodocea nodosa and Ruppia maritima in Epanomi and Vravrona, Greece
- Past and present fucalean diversity in the island of Marettimo, Egadi Islands Marine Protected Area (Central Mediterranean, Italy)
- Taxonomy/Phylogeny and Biogeography
- Morphological and molecular characterization of Chnoospora minima (Scytosiphonaceae, Ectocarpales) along Mexican coasts, with the description of C. ramosissima sp. nov.
- Phylogeography of the marine benthic alga Gracilaria salicornia (Gracilariales, Rhodophyta) in Southeast Asia
- Unveiling a novel species, Pachymeniopsis shinchonai sp. nov., from Korea, with reference to phylogenetic relationships within the Grateloupiaceae (Halymeniales, Rhodophyta)
- First record of the red alga Ceramium pallidum (Ceramiales, Ceramiaceae) for the Mediterranean Sea
- Chemistry and Applications
- Chemical profiling of seaweeds of the Arabian Gulf by liquid chromatography-mass spectrometry and in-silico screening against MPOX