A new species of marine algae from the Mexican Atlantic based on morphology and molecular data: Gelidium rodrigueziae sp. nov. (Gelidiaceae, Rhodophyta)
-
Nataly Quiroz-González
Nataly Quiroz-González is a professor at the Facultad de Ciencias, Universidad Nacional Autónoma de México. Her lines of research are systematics, taxonomy and ecology of marine algae. She currently focuses on molecular systematics of red marine algae and algal ecological interactions such as epibiosis.
Maria Edith Ponce-Márquez completed her Bachelor studies in coral reefs, worked with marine pharmacology, detection of antibacterial activity. Master of Science (Biology) in the Faculty of Sciences, UNAM, in cytogenetics of marine algae; PhD of Science in Marine Biology at the Institute of Marine Sciences and Limnology, in molecular biology with marine macroalgae. Her current line of research is molecular biology, DNA extraction, PCR amplification, among others, with various groups of marine organisms, especially macroalgae.
 ,
Luis Gabriel Aguilar-Estrada
,
Luis Gabriel Aguilar-Estrada
Luis Gabriel Aguilar-Estrada is a professor in Facultad de Ciencias of Universidad Nacional Autónoma de México (UNAM). His current research interests are systematics, ecology and biogeography of marine mollusks, as well as the ecological interactions of invertebrates and aquatic vegetation such as macroalgae, seagrasses and mangroves.
Ivette Ruiz-Boijseauneau completed all her studies at the National Autonomous University of Mexico, and has worked with invertebrates for 27 years, both pelagic and benthic. In the last 10 years she has worked with macroalgae and particularly with amphipods associated with macroalgae. She has worked in the rocky intertidal of the Mexican tropical Pacific coast, with topics of taxonomy, ecology and histology.

Abstract
The algae of the genus Gelidium can be found as frequent and abundant inhabitants in the rocky intertidal zone of the Mexican Atlantic. Six species of this genus have been reported in this region, but only Gelidium americanum has been confirmed with morphological and molecular tools, despite how important both tools have proven to be for the group. This research aimed to evaluate the Mexican specimens previously identified as Gelidium through morphological and molecular approaches, using rbcL and COI-5P markers. This study examined 20 specimens found living attached to the shell of a gastropod Stramonita rustica in the mid-high intertidal zone of the Mexican Atlantic and ten specimens found living on rocks. Phylogenetic analyses resolved these specimens to be an independent species closely related to G. gonzalezii, G. yangmeikengense, and G. indonesianum. Genetic distances and morphological information confirmed this. Gelidium rodrigueziae sp. nov. is described as a new species. It is distinguished from its related species by having a simpler branching pattern, a smaller number and size of medullary cells, and the smaller quantity and distribution of restricted internal rhizoid filaments around the medulla.
1 Introduction
The genus Gelidium J.V. Lamouroux is a complex group containing 144 species in the Gelidiaceae of Rhodophyta (Guiry and Guiry 2023) and is characterized by plants with pinnate or simple branches, brush-like haptera, rhizoidal filaments in the cortex and/or medulla, bilocular cystocarps with an ostiole on each side, and apical tetrasporangial sori (Boo et al. 2022b; Rodríguez et al. 2008). The genus comprises perennial red seaweeds with ecological and economic value that occur on rocks in lower intertidal areas around the world (Rodríguez et al. 2008; Santelices 1990).
Gelidium is a genus with a cosmopolitan distribution that, together with its high morphological plasticity, has presented a significant challenge for taxonomic determination using only morphological characters (Boo et al. 2022b; Quiroz-González et al. 2021a). Molecular tools used by other disciplines were adopted and used in phycological studies (Freshwater and Rueness 1994). These analyses, in conjunction with morphological studies, increased the ability to differentiate Gelidium species and resulted in the description of new taxa, as well as the extension of their distributions or important nomenclatural changes (Boo et al. 2013, 2016a,b,c, 2022b; Freshwater and Rueness 1994; Kim et al. 2012a,b; Quiroz-González et al. 2020a).
The algae of the genus Gelidium are an essential element of the flora of the coasts of Mexico because they are abundant and contribute to ecological interactions. They have been recorded in different intertidal and subtidal environments, being an important part of the floristic inventories in the country (García-García 2020; Ortega et al. 2001; Rodríguez et al. 2008), and they are found in rocky intertidal areas, in coastal lagoons, in coral reefs, and attached to rocks or other organisms, such as invertebrates or other algae (Quiroz-González et al. 2020b). The genus is a structuring element of Mexican coast communities, and the taxonomy of its species must be adequately delimited.
Knowledge of these species has been developed mainly on the Mexican Pacific coasts, where 20 species have been recorded to date (Quiroz-González et al. 2020a, 2021a; Rodríguez et al. 2008), while in the Mexican Atlantic, only six species have been recorded: Gelidium americanum, G. corneum, G. crinale, G. floridanum, G. pusillum and G. spinosum (Garcia-Garcia et al. 2020; González-González et al. 1996; Ortega et al. 2001). In the states of Veracruz and Tabasco, in particular, five of the six species recorded for the Mexican Atlantic have been found, the exception being G. spinosum (Garcia-Garcia et al. 2020).
The vast majority of the Gelidium species in the Mexican Atlantic have not yet been evaluated with morphological and molecular tools, and only G. americanum has been characterized (Quiroz-González et al. 2021b). In Mexico, this type of study has been developed in the Pacific region, with the proposal of two new species (Gelidium gonzalezii and G. nayaritense) and the taxonomic certification of three more (G. sanyaense, G. jhonstonii and G. sclerophyllum) (Boo et al. 2022b; Quiroz-González et al. 2020a, 2021a; Wang et al. 2017).
Given the value that the genus Gelidium represents and the need to evaluate its taxonomic status with molecular and morphological tools, the present study aimed to contribute to the knowledge of the genus through the characterization of specimens found as epibionts of the mollusk Stramonita rustica Lamarck 1822 in an Atlantic locality of Southern Mexico, using molecular markers and morphological data, which gives rise to the proposal of a new species G. rodrigueziae sp. nov.
2 Materials and methods
A total of 20 specimens were found growing on the shell of the mollusk Stramonita rustica that were collected in Playa Hermosa, Veracruz (18°39′ 51″ N, 95°7′ 48″ W) in May 2018. More recently, seven specimens were found growing on a rock in the Faro Verde locality, Veracruz (19°11′40″N, 96°7 ′24″W), and three more were found on this same substratum in the Andrés Sánchez Magallanes Breakwater, Tabasco (18°18′ 2″N, 93°51′ 1″ W) in May 2022 (Figure 1). Specimens were fixed with 4 % formaldehyde solution in seawater for morphological study, and a part of each thallus was also placed in silica gel for molecular analysis. Images of their general morphological features were obtained with a Canon Eos Rebel T3 camera (Canon Inc., Tokyo, Japan). Transverse and longitudinal thallus sections were cut manually using a razor blade under a Nikon C-LEDS stereomicroscope (Nikon, Inc., Konan, Minato-ku, Tokyo, Japan). Anatomical observations were made with an Olympus CX23 optical microscope (Olympus America Inc., Center Valley, Pennsylvania, USA).

Sampling sites for Gelidium spp. in the Mexican Atlantic.
Total DNA was extracted from 8 specimens using the CTAB (Doyle and Doyle 1990) and Master Pure (Epicentre Illumina Inc., San Diego, California, USA) protocol. PCR reactions were set up with the Thermoscientific Phire Plant Direct Kit with the following reagent proportions: 10 μl of 2× buffer, 1.0 μl of each forward and reverse primer at 10 pmol, 0.4 μl of Taq Polymerase, 1.0 μl of DNA and 11.6 μl of injectable water, for a final volume of 25 μl. The primers used for amplifying and sequencing were F-rbcL, starting with R-753 and F-577 with R-1381 for rbcL (Freshwater and Rueness 1994). COI-5P was amplified and sequenced using the GHaIF with GazR primers (Saunders 2005). The reactions were run in a Techne Flexi-gene thermal cycler (Bibby scientific, Maryland, USA), following the cycling protocol described by Freshwater and Rueness (1994). The PCR products were sequenced by Macrogen Inc. (South Korea), edited with the Bioedit (Hall 1999) or Sequencher (Gene Codes Corporation, Ann Arbor, MI, USA) programs, and aligned with Clustal W (Thompson et al. 1994).
The rbcL and COI-5P alignments comprised 13 new and 41 GenBank sequences, and 10 new and 37 GenBank sequences, respectively (Supplementary Table S1). Ptilophora scalaramosa, Gelidiophycus freshwateri, and Capreolia implexa were used as the outgroups. MEGA 7.0 (Kumar et al. 2016) was used to calculate genetic distances. The best evolutionary model for phylogenetic analyses selected with the JModelTest 2.1.10 program (Dariba et al. 2012) was GTR + G + I for both data sets. Phylogenies of COI-5P and rbcL datasets were inferred using maximum likelihood (ML) and Bayesian inference (BI). The ML analyses were conducted using RAxML v8.0.X (Stamatakis 2014) set as follows: a rapid bootstrap analysis and search for the best scoring ML tree in one single program run with 1000 bootstrap replicates under the GTR + G + I substitution model. BI was performed for individual datasets with MrBayes v3.2.1 (Ronquist et al. 2012) using the Metropolis coupled Markov chain Monte Carlo under the GTR + G + I model. For each matrix, two million generations of two independent runs were performed with four chains and sampling trees every 100 generations. The burn-in period was identified graphically by tracking the likelihood at each generation to determine whether they reached a plateau. Twenty-five percent of the saved trees were removed, and the remaining trees were used to calculate Bayesian posterior probabilities (BPPs).
3 Results
3.1 Molecular phylogeny
Maximum Likelihood and Bayesian Inference analyses produced trees with similar topologies for both markers. For both markers, the specimens of this work formed a clade fully supported by high posterior probability and bootstrap values (1 and 100, respectively) (Figures 2 and 3). The clade formed by these Mexican specimens for both markers was the sister species of G. gonzalezii and closely related to G. indonesianum Kim, Gerung et Boo and G. yangmeikengense Wang et Wang. For rbcL, genetic distances among the Mexican specimens were 0.0–0.1 % at the intraspecific level, and 1.2–1.8 % for G. gonzalezii and 1.6–2.2 % for G. yangmeikengense and G. indonesianum. The intraspecific genetic distances among the specimens for the COI-5P marker were 0.0 %, while the interspecific distances between them and G. gonzalezii were 3.2–3.9 % and between them and G. yangmeikengense and G. indonesianum were 3.4–6.7 %.

Maximum-likelihood (ML) tree inferred from rbcL sequences of Gelidium. ML bootstrap and Bayesian posterior probability values are shown at branches. Bold letters indicate newly generated sequences in this study. * indicates the holotype.

Maximum-likelihood (ML) tree inferred from COI-5P sequences of Gelidium. ML bootstrap and Bayesian posterior probability values are shown at branches. Bold letters indicate newly generated sequences in this study. * indicates the holotype.
3.2 Morphology
3.2.1 Gelidium rodrigueziae Quiroz-González et Ponce-Márquez
Holotype: Playa Hermosa, Veracruz, México (18°39′ 51″y 95°7′ 48″ O). Thalli was growing on the shell of gastropod Stramonita rustica in the intertidal zone: GM 898, vegetative, deposited in the Herbarium of Facultad de Ciencias, Universidad Nacional Autónoma de México (FCME); collected by L. G. Aguilar-Estrada on 18 May 2018. GenBank accession number OQ850753 (rbcL) and OR133600 (COI-5P). Isotype: GM 899. GenBank accession number OQ850754 (rbcL) and OR133601 (COI-5P). Paratypes: Faro Verde, Veracruz, México GM900, GM 901 deposited in the Herbarium of Facultad de Ciencias, Universidad Nacional Autónoma de México (FCME); collected by N. Quiroz-González on 13 May 2022. GenBank accession number OQ850755, OQ850756 (rbcL) and OR133602, OR133603 (COI-5P).
Etymology: Dedicated to Dra. Dení Rodriguez a Mexican phycologist, for her invaluable contribution to our knowledge of the Gelidiales from Mexico and the world.
Diagnosis: Thallus light to dark red, up to 0.7 cm high, forming a turf on Stramonita rustica (Figure 4A and B), with terete prostrate branches attached by brush-like holdfasts (Figure 4C and D). Erect axes cylindrical at the base and flattened above 150–350 µm in width. Branches are rare, and when branches are occasionally present, they are irregularly arranged with one order of ramification (Figure 4B–D).
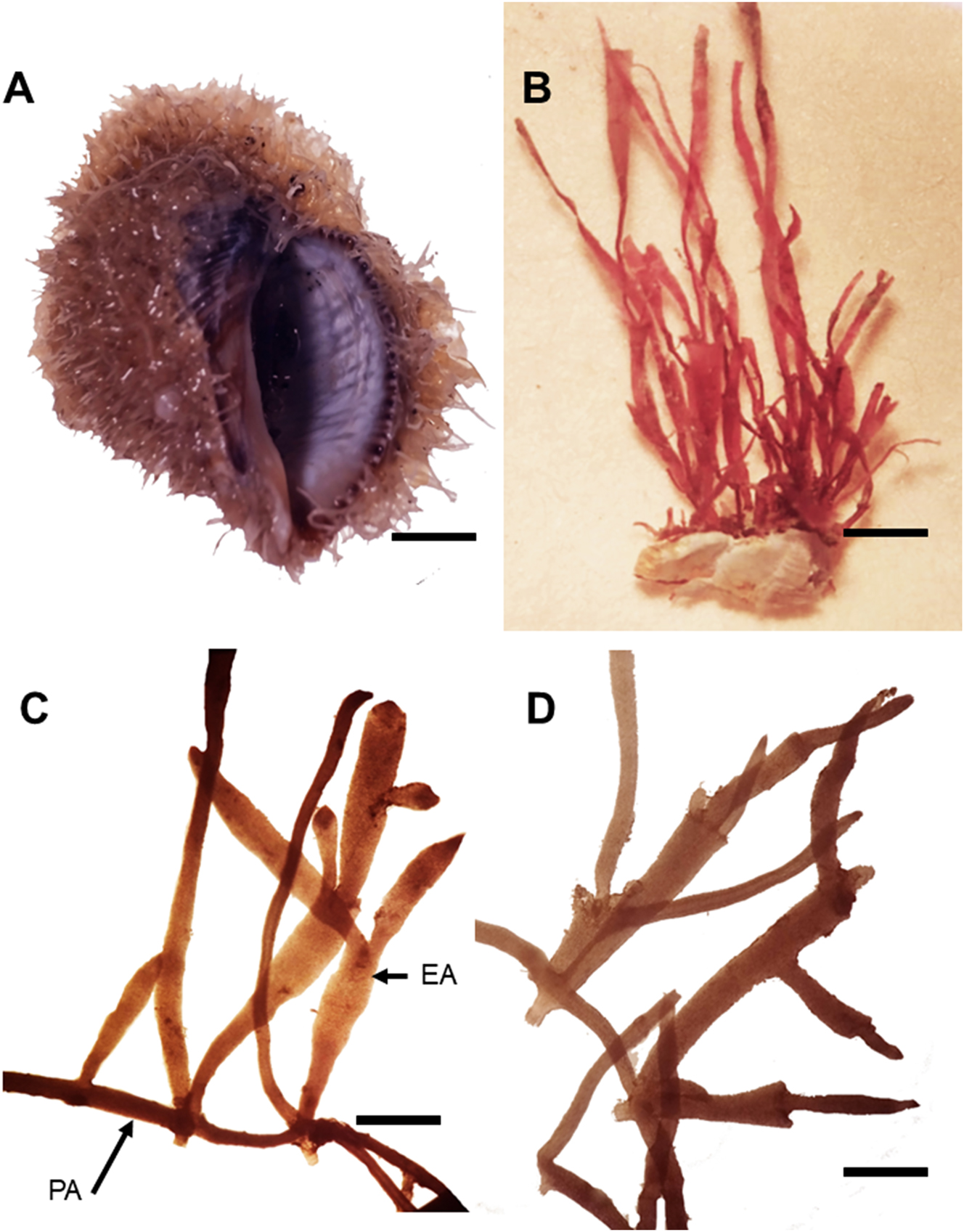
Vegetative features of Gelidium rodrigueziae sp. nov. (A) Thalli on Stramonita rustica (GM 898). Scale = 0.5 cm. (B) Vegetative holotype thalli (GM 898). Scale = 600 μm. (C and D) Vegetative paratype thalli (GM 900) with prostrate axes (PA) and erect axes (EA). Scale = 400 µm. (D) Stolon. Scale = 400 µm.
Description: Plants a turf of purple-red thalli, with cylindrical prostrate axes attached to substratum with brush-like haptera (Figure 5A) and erect linear to clavate axes, with irregular to smooth margins, cylindrical at base, 0.3–0.7 cm high and 150–350 µm in width (Figure 4B–D). Axes in transverse section are ovoid, and elliptical. The shape of vegetative apices is acute (Figure 5B). Cortex of 1–2 layers of rounded cells, 3.75–4.5 µm in diameter (Figure 5C and D), outer cortical cells irregularly arranged in surface view. Medulla of 2–4 rows of circular or subcircular cells in transverse section, 10–12.5 μm in diameter, and with very large intercellular spaces. Internal rhizoidal filaments in the medulla (Figure 5C and D).

Vegetative features of Gelidium rodrigueziae sp. nov. (A) Haptera (HAP). Scale = 100 µm. (B) Apical cell (AC). Scale = 50 µm. (C and D) Transverse section, with internal rhizoidal filaments (arrows). Scale = 25 µm.
Additional material examined: Playa Hermosa, Veracruz, México GM 902-GM905 and GM911-GM922 vegetative, deposited in the Herbarium of Facultad de Ciencias, Universidad Nacional Autónoma de México (FCME); collected by L. G. Aguilar-Estrada on 18 May 2018. Faro Verde, Veracruz, México GM 906-GM 908 and GM 923-GM926 vegetative, deposited in the Herbarium of Facultad de Ciencias, Universidad Nacional Autónoma de México (FCME); collected by N. Quiroz-González on 13 May 2022. Andrés Sánchez Magallanes Breakwater, Tabasco, México GM909-GM910, GM 927 vegetative, deposited in the Herbarium of Facultad de Ciencias, Universidad Nacional Autónoma de México (FCME); collected by L. G. Aguilar-Estrada on 16 May 2022.
4 Discussion
Our analyses of plastid rbcL and mitochondrial COI-5P sequences, with morphological observations, clearly revealed the occurrence of a new species, Gelidium rodrigueziae, from Mexican Atlantic. Gelidium rodrigueziae is characterized by a combination of the following vegetative features: small size (0.3–0.7 cm), axes erect, terete at the base, flattening as they approach the apex, with simple or scarce branching, rhizines moderately abundant to abundant in medulla. Table 1 provides a comparison of G. rodrigueziae with the other species recorded in the Mexican Atlantic and phylogenetically related species. Gelidium rodrigueziae is notably different from the species to which it is closest phylogenetically. It has a much smaller and simpler thallus, with fewer layers of medullary and cortical cells than G. gonzalezii, G. indonesianum and G. yangmeikengense (Quiroz-González et al. 2020a; Wang et al. 2017), is clearly distinguished from G. gonzalezii by its simpler branching and fewer orders, as well as a smaller number of rows of cortical and medullary cells. There is no morphological similarity with the rest of the species registered for the Mexican Atlantic, since they have much larger and more branched thalli.
Morphological comparison of Gelidium rodrigueziae sp. nov with closely related species, and those of similar morphology along the Mexican Atlantic coast.
| Characters | G. americanum (W.R. Taylor) Santelices (Perrone et al. 2019; Quiroz-González et al. 2021b; Taylor 1945) | G. corneum (Hudson) Lamouroux (Taylor 1960) | G. crinale (Hare ex Turner) Gaillon (Taylor 1960) | G. floridanum W.R. Taylor (Taylor 1960) | G. gonzalezii Rodríguez, Quiroz-González et Ponce-Márquez (Quiroz-González et al. 2020a) | Gelidium indonesianum K.M. Kim, G.S. Gerung et S.M. Boo (Kim et al. 2011) | G. pusillum (Stackhouse) Le Jolis (Taylor 1960) | G. rodrigueziae sp. nov. Quiroz-González et Ponce-Márquez (this study) | G. yangmeikengense G.C. Wang et X.L. Wang (Wang et al. 2017) |
|---|---|---|---|---|---|---|---|---|---|
| Thallus size | 0.5–6 cm | 3–50 cm | 0.4–1.20 | 3–6.5 cm | 0.7–2.5 cm | Up to 15 cm | 0.1–1.70 | 0.3–0.7 cm | 1–2 cm |
| Branching pattern | Irregular below, alternating to subpinnate | Pinnate | Simple to multiple irregular | Pinnate, alternating or irregular branching | Multiple irregular | Simple | Irregular | Scarce or simple | Simple or pinnate |
| Branching orders | 2–4 | 1–3 | 1–3 | 2–3 | 1–3 | – | 1–3 | 1 | 1–2 |
| Apex | Acute | Rounded | Acute | Obtuse/bilobed | Obtuse/bilobed | Lanceolate, blunt, or truncate apices | Acute/obtuse | Acute | – |
| Stolon | Cylindrical | Cylindrical | Cylindrical | Cylindrical | Cylindrical | Cylindrical | Cylindrical | Cylindrical | Cylindrical |
| Internal rhizoidal filaments | Abundant to scarce in the medulla | Abundant in the subcortex | Abundant in the subcortex and outlying medulla | Abundant in medulla | Abundant to scarce in the subcortex, abundant in the inner medulla | Abundant around the medulla | Abundant in the medulla, scarce to abundant in the subcortex | Abundant to scarce in the medulla | Abundant in the medulla |
| Medullary cells | 3 rows, circular, 14–16.5 μm diameter | – | 4-8 rows, circular or subcircular, 10–20 μm diameter | – | 3-8 rows, 8–27 μm diameter | – | 3-8 rows, 6–21 μm diameter | 2-4 rows, 10–12.5 μm diameter | – |
| Cortical cells | 1 row | – | 2–3 rows | – | 2–4 rows, elliptical to rounded | Rounded to ovate, 4–7 µm diameter | 1–3 rows | 1–2 rows, rounded | 3 rows; rounded, 2.8–4.6 μm diameter |
-
(−): no information.
Phylogenetic analysis of both rbcL and COI-5P sequences consistently demonstrated a marked difference between Gelidium rodrigueziae and other species in the genus. The nucleotide divergence values established in the genus to distinguish species with rbcL and COI-5P have variable ranges. There are proposals for new species based on very small ranges, with values of 0.3–1 % for rbcL and 2.6–4.6 % for COI-5P, for example, between Gelidium palmatum and G. millarianum, between G. sanyaense and G. sentosaense G.H. Boo, as well as the divergence between G. linoides and G. tenuifolium. Thus, the genetic distance values between G. gonzalezii and G. rodrigueziae resemble the small values reported for other Gelidium species (Boo et al. 2014, 2022a; Boo and Kim 2020; Brunelli et al. 2019; Jamas et al. 2017; Quiroz-González et al. 2020a, 2021a; Wang et al. 2017).
Gelidium rodrigueziae is a sister species of Gelidium gonzalezii recently described for the Mexican Pacific (Quiroz-González et al. 2020a), although they belong to very different ecoregions and biogeographic provinces according to the classification proposed by Spalding et al. (2007) (Figures 2 and 3). A similar close relationship between disjunct species has been recorded in other species of Gelidium, such as Gelidium brasiliense and G. sentosaense, which are phylogenetically close but are found in Brazil and China, respectively (Brunelli et al. 2019) or G. sclerophyllum and G. floridanum that are close but have been recorded in the Pacific from Mexico to Ecuador and Brazil. Their divergence has been attributed to the formation of the Isthmus of Panama (Boo et al. 2022b). Thus, the genetic distance values and their geographical distribution support the proposal of a new species of Gelidium for the Mexican Atlantic.
Gelidium rodrigueziae has been found growing in most abundance on the mollusk Stramonita rustica, which is frequent in the intertidal zone of the Gulf of Mexico. Shells provide substrata for the settlement of epibionts, provide shelter from predation, and from physical or physiological stresses (Gutiérrez et al. 2003; Levenets et al. 2010; Quiroz-González et al. 2020b). Recently, epibiont relationships have been of special interest, as they have served to indicate numerous new species among very different marine organisms, such as ciliates on nematodes, entoprocts on bryozoans and even flatworms on crustaceans (Borisanova and Krylova 2014; Chatterjee and Fernandez-Leborans 2013; Soares et al. 2021).
As Boo and Kim (2020) point out, small species of Gelidium, such as G. rodrigueziae are difficult to define using a few diagnostic characteristics, so the use of molecular tools is essential for the knowledge of this genus whose species richness is increasing substantially thanks to their use. Therefore, evaluating the rest of the algae of this genus on the coasts of Mexico is important, especially on substrata as promising as animal shells.
About the authors

Nataly Quiroz-González is a professor at the Facultad de Ciencias, Universidad Nacional Autónoma de México. Her lines of research are systematics, taxonomy and ecology of marine algae. She currently focuses on molecular systematics of red marine algae and algal ecological interactions such as epibiosis.

Maria Edith Ponce-Márquez completed her Bachelor studies in coral reefs, worked with marine pharmacology, detection of antibacterial activity. Master of Science (Biology) in the Faculty of Sciences, UNAM, in cytogenetics of marine algae; PhD of Science in Marine Biology at the Institute of Marine Sciences and Limnology, in molecular biology with marine macroalgae. Her current line of research is molecular biology, DNA extraction, PCR amplification, among others, with various groups of marine organisms, especially macroalgae.

Luis Gabriel Aguilar-Estrada is a professor in Facultad de Ciencias of Universidad Nacional Autónoma de México (UNAM). His current research interests are systematics, ecology and biogeography of marine mollusks, as well as the ecological interactions of invertebrates and aquatic vegetation such as macroalgae, seagrasses and mangroves.

Ivette Ruiz-Boijseauneau completed all her studies at the National Autonomous University of Mexico, and has worked with invertebrates for 27 years, both pelagic and benthic. In the last 10 years she has worked with macroalgae and particularly with amphipods associated with macroalgae. She has worked in the rocky intertidal of the Mexican tropical Pacific coast, with topics of taxonomy, ecology and histology.
Acknowledgments
We thank the Laboratory of Phycology (Marine Biodiversity), Facultad de Ciencias, Universidad Nacional Autónoma de México for the loan of the equipment and facilities for the taxonomic determination of the material. Thanks also to E. Julieta Sarmiento Ponce for reviewing the English language.
-
Research ethics: Registro Nacional de Pesca y Acuacultura, Folio DF00000208.
-
Author contributions: All authors conceived the project idea and developed the taxonomic work, interpreted the data, and contributed to the draft and critical review of the final manuscript. All authors discussed the results and contributed to the final manuscript.
-
Competing interests: The authors declare that they have no conflict of interest. All data was obtained and collected in accordance with local research protocols.
-
Research funding: This work was not funded.
-
Data availability: The DNA sequences are available for consultation in GenBank.
References
Boo, G.H. and Kim, K.M. (2020). A new species of marine algae from Korea based on morphology and molecular data: Gelidium palmatum sp. nov. (Gelidiales, Rhodophyta). Algae 35: 33–43, https://doi.org/10.4490/algae.2020.35.3.6.Suche in Google Scholar
Boo, G.H., Park, J.K., and Boo, S.M. (2013). Gelidiophycus (Rhodophyta: Gelidiales): a new genus of marine algae from East Asia. Taxon 62: 1105–1116, https://doi.org/10.12705/626.7.Suche in Google Scholar
Boo, G.H., Kim, K.M., Nelson, W.A., Riosmena-Rodríguez, R., Yoon, K.J., and Boo, S.M. (2014). Taxonomy and distribution of selected species of the agarophyte genus Gelidium (Gelidiales, Rhodophyta). J. Appl. Phycol. 26: 1243–1251, https://doi.org/10.1007/s10811-013-0111-7.Suche in Google Scholar
Boo, G.H., Cai, Y., and Boo, S.M. (2016a). Molecular identification of gelidioid algae (Gelidiales, Rhodophyta) from Singapore with a description of Gelidium sentosaense sp. nov. Phycologia 55: 247–256, https://doi.org/10.2216/15-133.1.Suche in Google Scholar
Boo, G.H., Le Gall, L., Miller, K.A., Freshwater, D.W., Wernberg, T., Terada, R., Yoon, K.J., and Boo, S.M. (2016b). A novel phylogeny of the Gelidiales (Rhodophyta) based on five genes including the nuclear CesA, with descriptions of Orthogonacladia gen. nov. and Orthogonacladiaceae fam. nov. Mol. Phylogenet. Evol. 101: 359–372, https://doi.org/10.1016/j.ympev.2016.05.018.Suche in Google Scholar PubMed
Boo, G.H., Hughey, J., Miller, K.A., and Boo, S.M. (2016c). Mitogenomes from type specimens, a genotyping tool for morphologically simple species: ten genomes of agar-producing red algae. Sci. Rep. 6: 35337, https://doi.org/10.1038/srep35337.Suche in Google Scholar PubMed PubMed Central
Boo, G.H., Park, J.K., Han, K.S., and Yoon, H.S. (2022a). Gelidium rosulatum (Gelidiales, Rhodophyta), a new species of subtidal marine algae from Korea. Phycologia 61: 332–340, https://doi.org/10.1080/00318884.2022.2046933.Suche in Google Scholar
Boo, G.H., Le Gall, L., Hwang, I.K., Rousseau, F., and Yoon, H.S. (2022b). Species Diversity of Gelidium from Southern Madagascar evaluated by an integrative taxonomic approach. Diversity 14: 826, https://doi.org/10.3390/d14100826.Suche in Google Scholar
Borisanova, A.O. and Krylova, E.M. (2014). A new species of loxosomatidae (Entoprocta, Solitaria) from the white sea: Loxosomella unicornis sp. nov. Zootaxa 3861: 290–296, https://doi.org/10.11646/zootaxa.3861.3.6.Suche in Google Scholar PubMed
Brunelli, B., Jamas, M., Milstein, D., Boo, S.M., and Fujii, M.T. (2019). Gelidium brasiliense sp. nov. (Gelidiales, Rhodophyta): a diminutive agarophyte from Brazil. J. Appl. Phycol. 31: 951–958, https://doi.org/10.1007/s10811-018-1678-9.Suche in Google Scholar
Chatterjee, T. and Fernandez-Leborans, G. (2013). Ciliate epibionts on Melita petronio Senna et al., 2012 (Crustacea: Amphipoda) from Brazil. Cah. Biol. 54: 393–404.Suche in Google Scholar
Dariba, D., Taboada, G.L., Doallo, R., and Posada, D. (2012). jModelTest 2: more models, new heuristics and parallel computing. Nat. Methods 9: 772, https://doi.org/10.1038/nmeth.2109.Suche in Google Scholar PubMed PubMed Central
Doyle, J.J. and Doyle, J.L. (1990). Isolation of plant DNA from fresh tissue. Focus 12: 13–15.10.2307/2419362Suche in Google Scholar
Freshwater, D.W. and Rueness, J. (1994). Phylogenetic relationships of some European Gelidium (Gelidiales, Rhodophyta) species, based on rbcL nucleotide sequence analysis. Phycologia 33: 187–194, https://doi.org/10.2216/i0031-8884-33-3-187.1.Suche in Google Scholar
García-García, A.M.E., Cabrera-Becerril, E., Núñez-Resendiz, M.L., Dreckmann, K.M., and Sentíes, A. (2020). Actualización taxonómica de las algas rojas (Rhodophyta) marinas bentónicas del Atlántico mexicano. Acta Bot. Mex. 127: e1677, https://doi.org/10.21829/abm127.2020.1677.Suche in Google Scholar
González-González, J., Gold-Morgan, M., León-Tejeda, H., Candelaria, C., León-Álvarez, D., Serviere-Zaragoza, E., and Fragoso, D. (1996). Catálogo onomástico (nomenclator) y bibliografía indexada de las algas bentónicas marinas de México. Cuadernos No. 34. Instituto de Biología, Universidad Nacional Autónoma de México, México.Suche in Google Scholar
Guiry, M.D. and Guiry, W.D. (2023). AlgaeBase version 4.2. Worldwide electronic publication, National University of Ireland, Galway, Available at: http://www.algaebase.org (Accessed 15 April 2023).Suche in Google Scholar
Gutiérrez, J.L., Jones, C.G., Strayer, D.L., and Iribarne, O.O. (2003). Mollusks as ecosystem engineers: the role of shell production in aquatic habitats. Oikos 101: 79–90, https://doi.org/10.1034/j.1600-0706.2003.12322.x.Suche in Google Scholar
Hall, T.A. (1999). BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 41: 95–98.Suche in Google Scholar
Jamas, M., Iha, C., Oliveira, M.C., Guimarães, S.M.P.B., and Fujii, M.T. (2017). Morphological and molecular studies on Gelidiaceae and Gelidiellaceae (Gelidiales, Rhodophyta) from Brazil with description of the new species Gelidium calidum. Phytotaxa 314: 195–218, https://doi.org/10.11646/phytotaxa.314.2.2.Suche in Google Scholar
Kim, K.M., Hoarau, G., and Boo, S.M. (2012a). Genetic structure and distribution of Gelidium elegans (Gelidiales, Rhodophyta) in Korea based on mitochondrial cox1 sequence data. Aquat. Bot. 98: 27–33, https://doi.org/10.1016/j.aquabot.2011.12.005.Suche in Google Scholar
Kim, K.M., Hwang, I.K., Park, J.K, and Boo, S.M. (2011). A new agarophyte species, Gelidium eucorneum sp. nov. (Gelidiales, Rhodophyta), based on molecular and morphological data. J. Phycol. 47: 904–910, https://doi.org/10.1111/j.1529-8817.2011.01005.Suche in Google Scholar
Kim, K.M., Hwang, I.K., Yoon, H.S., Boo, S.M., coreanum, G., jejuensis, G., minimum, G., and prostratum, G. (2012b). Four novel Gelidium species (Gelidiales, Rhodophyta) discovered in Korea: G. coreanum, G. jejuensis, G. minimum and G. prostratum. Phycologia 51: 461–474, https://doi.org/10.2216/11-117.1.Suche in Google Scholar
Kumar, S., Stecher, G., and Tamura, K. (2016). MEGA7: molecular evolutionary genetics analysis version 7.0 for bigger datasets. Mol. Biol. Evol. 33: 1870–1874, https://doi.org/10.1093/molbev/msw054.Suche in Google Scholar PubMed PubMed Central
Levenets, I.R., Ovsyannikova, I.I., and Lebedev, E.B. (2010). Epibiotic macroalgae on the scallop Mizuhopecten yessoensis in Peter the Great Bay, Sea of Japan. Russ. J. Mar. Biol. 36: 340–349, https://doi.org/10.1134/s1063074010050032.Suche in Google Scholar
Ortega, M.M., Godínez, J.L., and Garduño-Solórzano, G. (2001). Catálogo de algas bénticas de las costas mexicanas del Golfo de México y Mar Caribe. Instituto de Biología. Universidad Nacional Autónoma de México, México.Suche in Google Scholar
Quiroz-González, N., Ponce-Márquez, M.E., Fernández-García, C., and Rodríguez, D. (2020a). Gelidium gonzalezii sp. nov. (Gelidiales, Rhodophyta) from the Mexican tropical Pacific based on molecular and morphological evidence. Phytotaxa 459: 124–138, https://doi.org/10.11646/phytotaxa.459.2.4.Suche in Google Scholar
Perrone, C., Bottalico, A., Boo, G.H., Boo, S.M., Miller, K.A, and Freshwater, D.W. (2019). Gelidium adriaticum sp. nov. and Gelidium carolinianum sp. nov. (Gelidiales, Rhodophyta) from the Mediterranean Sea. Phycologia 58: 359–373, https://doi.org/10.1080/00318884.2019.1580102.Suche in Google Scholar
Quiroz-González, N., Aguilar-Estrada, L.G., Ruiz-Boijseauneau, I., and Rodríguez, D. (2020b). Biodiversidad de algas epizoicas en el Pacífico tropical mexicano. Acta Bot. Mex. 127: e1645, https://doi.org/10.21829/abm127.2020.1645.Suche in Google Scholar
Quiroz-GonzálezPonce-Márquez, N.M.E., López-Gómez, N., Rodríguez, D., and Rodríguez, D. (2021a). Morphological and molecular characterization of two species of Gelidium (Gelidiales, Rhodophyta) from Mexico: G. nayaritense sp. nov. and the new record of G. sanyaense. Phytotaxa 527: 161–176, https://doi.org/10.11646/phytotaxa.527.3.1.Suche in Google Scholar
Quiroz-González, N., Rivas Acuña, M.G., and Ponce Márquez, M.E. (2021b). Confirmación taxonómica de Gelidium americanum (Gelidiaceae, Rhodophyta) en Tabasco, México, usando un enfoque morfológico y molecular. Acta Bot. Mex. 128: e1966, https://doi.org/10.21829/abm128.2021.1966.Suche in Google Scholar
Rodríguez, D., López, N., and González-González, J. (2008) Gelidiales (Rhodophyta) en las costas del Pacifico mexicano con énfasis en las especies tropicales. In: Sentíes, A. and Dreckmann, K. (Eds.), Monografías ficológicas, Vol. 3. Universidad Autónoma Metropolitana, Mexico, pp. 27–74.Suche in Google Scholar
Ronquist, F., Teslenko, M., van der Mark, P., Ayres, D.L., Darling, A., Höhna, S., Larget, B., Liu, L., Suchard, M.A., and Huelsenbeck, J.P. (2012). MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 61: 539–542, https://doi.org/10.1093/sysbio/sys029.Suche in Google Scholar PubMed PubMed Central
Santelices, B. (1990). New and old problems in the taxonomy of the Gelidiales (Rhodophyta). Hydrobiologia 204-205: 125–135, https://doi.org/10.1007/978-94-009-2049-1_18.Suche in Google Scholar
Saunders, G.W. (2005). Applying DNA barcoding to red macroalgae: a preliminary appraisal holds promise for future applications. Philos. Trans. R. Soc. Lond. B, Biol. Sci. 360: 1879–1888, https://doi.org/10.1098/rstb.2005.1719.Suche in Google Scholar PubMed PubMed Central
Soares, G.B., Domingues, M.V., and Adriano, E.A. (2021). Morphological and molecular characterization of Udonella brasiliensis n. sp. (Monogenoidea), an epibiont on Caligus sp. parasite of Ariidae from the southeastern coast of Brazil. Parasitol. Int. 83: 1–8, https://doi.org/10.1016/j.parint.2021.102371.Suche in Google Scholar PubMed
Spalding, M.D., Fox, H.E., Allen, G.R., Davidson, N., Ferdaña, Z.A., Finlayson, M., Harlpen, B.S., Jorge, M.A., Lombana, A., Lourie, S.A., et al.. (2007). Marine ecoregions of the world: a bioregionalization of coastal and shelf areas. BioScience 57: 573–583, https://doi.org/10.1641/b570707.Suche in Google Scholar
Stamatakis, A. (2014). RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 22: 2688–2690, https://doi.org/10.1093/bioinformatics/btl446.Suche in Google Scholar PubMed
Taylor, W.R. (1945). Pacific marine algae of the Allan Hancock Expeditions to the Galapagos Islands. Allan Hancock Pac. Exped. 12: 1–528.Suche in Google Scholar
Taylor, W.R. (1960). Marine algae of the eastern tropical and subtropical coasts of the Americas. The University of Michigan Press, Ann Arbor.Suche in Google Scholar
Thompson, J.D., Higgins, D.G., and Gibson, T.J. (1994). CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic Acids Res. 22: 4673–4680, https://doi.org/10.1093/nar/22.22.4673.Suche in Google Scholar PubMed PubMed Central
Wang, X.L., Xia, B.M., Bottalico, A., and Wang, G.C. (2017). Taxonomic revision of Gelidium tsengii and Gelidium honghaiwanense sp. nov. (Gelidiales, Rhodophyta) from China based upon molecular and morphological data analyses. Chinese J. Oceanol. Limnol. 35: 1442–1453, https://doi.org/10.1007/s00343-017-5340-1.Suche in Google Scholar
Supplementary Material
This article contains supplementary material (https://doi.org/10.1515/bot-2023-0046).
© 2024 the author(s), published by De Gruyter, Berlin/Boston
This work is licensed under the Creative Commons Attribution 4.0 International License.
Artikel in diesem Heft
- Frontmatter
- In this issue
- Obituary
- A life for science and seaweeds: Klaus-Otto Lüning (1941–2023)
- Review
- Chemical ecology of plankton parasitism in algae
- Physiology and Ecology
- Saccharina sporophytes settling on kelp cultivation rafts come from wild intertidal populations and not from the farmed cultivars
- Feasibility of ex-situ recruitment and outplanting of Gongolaria nodicaulis (Fucales, Phaeophyceae) for restoration of warm temperate marine forests in Atlantic Morocco
- Taxonomy/Phylogeny and Biogeography
- Phymatolithopsis acervata comb. nov., and P. roseola sp. nov. (Hapalidiales, Rhodophyta) from South Africa with a comment on infraordinal classification
- A new species of marine algae from the Mexican Atlantic based on morphology and molecular data: Gelidium rodrigueziae sp. nov. (Gelidiaceae, Rhodophyta)
- Occurrence of Dictyosphaeria (Cladophorales, Chlorophyta) species on the Brazilian coast and oceanic islands, based on morphological and molecular data
- New morphological data for two rare species of sand-dwelling marine dinoflagellates, Amphidiniella sedentaria and Pachena cf. leibnizii
Artikel in diesem Heft
- Frontmatter
- In this issue
- Obituary
- A life for science and seaweeds: Klaus-Otto Lüning (1941–2023)
- Review
- Chemical ecology of plankton parasitism in algae
- Physiology and Ecology
- Saccharina sporophytes settling on kelp cultivation rafts come from wild intertidal populations and not from the farmed cultivars
- Feasibility of ex-situ recruitment and outplanting of Gongolaria nodicaulis (Fucales, Phaeophyceae) for restoration of warm temperate marine forests in Atlantic Morocco
- Taxonomy/Phylogeny and Biogeography
- Phymatolithopsis acervata comb. nov., and P. roseola sp. nov. (Hapalidiales, Rhodophyta) from South Africa with a comment on infraordinal classification
- A new species of marine algae from the Mexican Atlantic based on morphology and molecular data: Gelidium rodrigueziae sp. nov. (Gelidiaceae, Rhodophyta)
- Occurrence of Dictyosphaeria (Cladophorales, Chlorophyta) species on the Brazilian coast and oceanic islands, based on morphological and molecular data
- New morphological data for two rare species of sand-dwelling marine dinoflagellates, Amphidiniella sedentaria and Pachena cf. leibnizii