Direct-acting antiviral therapy may help restore HCV-induced impaired redox balance and liver fibrosis process
-
Tayibe Bal

 ,
Serdar Doğan
,
Oğuzhan Özcan
,
Mehmet Çabalak
and
Berfin Çirkin
,
Serdar Doğan
,
Oğuzhan Özcan
,
Mehmet Çabalak
and
Berfin Çirkin

Abstract
Objectives
The aim of this study was to investigate the changes in thiol/disulfide balance, pro-fibrotic mediators (transforming growth factor-beta [TGF-β] and periostin) and a potential biomarker for the prediction of HCV-induced HCC (3β-hydroxysterol Δ24-reductase [DHCR24]) during direct-acting antiviral (DAA) therapy in chronic hepatitis C (CHC) patients.
Methods
This prospective cohort study included 56 non-cirrhotic, treatment-naive CHC patients who were treated with DAAs between January and June 2020. Laboratory tests, including serum total/native thiol, TGF-β
Results
Of the 56 patients, all achieved a sustained virological response after DAA therapy. There was a significant decrease in serum levels of disulfide and TGF-β, (p=0.020 and p<0.001, respectively) and a significant increase in serum levels of native thiol compared with baseline levels (p=0.010). There was no significant change in levels of total thiol, DHCR24 and periostin levels. Serum TGF-β levels were found to be positively correlated with total bilirubin levels (rs=0.470, p=0.001) and negatively with albumin levels (rs=−0.483, p<0.001). A significant moderate positive correlation was determined between baseline serum DHRC24 and disulfide levels (rs=0.356, p=0.007).
Conclusions
The study results suggest that the DAA therapy may help to restore the impaired thiol/disulfide balance and reduce the pro-fibrotic process in CHC patients by markedly decreasing serum levels of TGF-β, a key player in HCV-induced liver fibrosis.
Introduction
It is estimated that more than 180 million people worldwide are thought to have chronic hepatitis C (CHC) infections. Hepatitis C virus (HCV) is a common pathogen globally and is one of the leading causes of chronic liver disease that can lead to liver cirrhosis and hepatocellular carcinoma (HCC) [1]. Recent evidence has suggested that the progression of liver damage and fibrogenesis in CHC infection is the result of increased inflammatory activity, which might increase virus-induced oxidative stress, rather than direct viral cytotoxicity. Moreover, proteins expressed by HCV, predominantly NS5A proteins, damage to the mitochondrial respiratory chain directly and lead to increased production of reactive oxygen species (ROS) as well as reduced antioxidative capacity, and contribute to oxidative stress [2], [3], [4].
The antioxidative defense system maintains redox homeostasis in the liver by removing ROS, thereby protecting cells from oxidative damage [5]. Dynamic thiol/disulfide homeostasis is one of the major defense mechanisms against oxidative stress. An imbalance in this homeostasis indicates oxidative stress and has been shown to play a role in the pathogenesis of many diseases including chronic hepatitis B. However, there has been very little published information about the role of this imbalance in the pathogenesis of CHC infection [6], [7], [8]. Furthermore, it has been established that chronic oxidative stimuli not only cause hepatocyte death but also increase the production of profibrogenic factors such as transforming growth factor-beta (TGF-β), a key player in the development of liver fibrosis [9]. The serum levels of TGF-β have been reported to be elevated in CHC patients and to be associated with the severity of liver fibrosis which can be significantly decreased with interferon (IFN) and ribavirin (RBV) therapy [10, 11]. However, such data is limited in direct-acting antiviral (DAAs) therapy. Periostin is another critical mediator that contributes to liver fibrosis which is strongly induced by TGF-β [12]. Another important aspect of hepatic oxidative stress is that it is strongly associated with an increased risk of HCC in CHC patients [13]. 3β-hydroxysterol Δ24-reductase (DHCR24) has been one of the most prominent biomarkers recently used to in demonstrate this association [14]. It has been established that the upregulation of DHCR24 expression induced by HCV-mediated oxidative stress suppresses p53 activity (DNA repair), thereby promoting tumorigenicity [4, 13]. There is also a growing body of evidence to suggest that it might be a potential biomarker for the early detection of HCC and for monitoring disease progression in CHC patients [15].
With the use of DAAs, high success rates can be achieved in the sustained suppression of viral replication in CHC patients [16]. What remains unclear, however, is whether or not the DAAs alone are sufficient to control the increased oxidative stress secondary to the HCV and to terminate the progressive tissue damage such as fibrosis and carcinogenesis [17, 18]. If DAAs are not capable of reducing in oxidative stress, CHC patients may still be at an increased risk for the development of cirrhosis and HCC, even if permanent virological suppression has been achieved.
Therefore, the aim of this study was to determine whether DAA therapy alone is sufficient to change the increased oxidative stress and pro-fibrotic mediators induced by HCV in CHC patients.
Materials and methods
Participants and study design
This prospective, single-center, cohort study was conducted on 56 antiviral therapy naïve non-cirrhotic CHC patients, all of whom received DAA therapy between January 2020 and June 2020 at the Infectious Diseases Clinic of Hatay Mustafa Kemal University Hospital, a tertiary level university hospital in Hatay, Turkey. All patients received DAA therapy for 8 or 12 weeks (with GLE/PIB: glecaprevir/pibrentasvir and EXV/VIE: exviera/viekirax, respectively) and all achieved a sustained virological response at 12 weeks after completion of DAA therapy (SVR12). CHC was diagnosed on the basis of anti-HCV positivity and was confirmed by the detection of HCV RNA with a polymerase chain reaction. Patients co-infected with HBV/HIV, other active infections, or chronic diseases (e.g. collagen tissue diseases, malignancy, diabetes mellitus) were excluded.
The aspartate aminotransferase to platelet ratio index (APRI), a noninvasive alternative to was utilized to assess patients’liver fibrosis stages. The upper limit of normal (ULN) of AST was 40 IU/L. APRI scores were calculated as ([AST/ULN AST] × 100)/platelet count (109/L) [19].
Specimen collection
Blood samples were collected at baseline, at 1 month and at the end of therapy (EOT) which is taken within 72 h after the last dose of therapy. Fasting venous blood samples were collected from the HCV patients into biochemistry tubes for serum separation and into lithium-heparin coated tubes for plasma separation. All samples were centrifuged at 1,500 × g for 10 min within 45 min after sampling. Serum and plasma samples were stored at −80 °C until analyses.
Biochemical analysis
Serum levels of albumin, total protein, total bilirubin, direct bilirubin and ALT, AST activities were measured on Advia 1800 autoanalyzer (Siemens, Germany).
Thiol/disulfide parameters
Plasma total and native thiol levels were assayed with a novel, fully automated colorimetric method developed by Erel and Neselioglu [7]. Following the reduction of disulfide bonds with sodium borohydride to form free functional thiol groups, formaldehyde is used to remove the unused sodium borohydride reductant for preventing the reduction of DTNB (5,5′-dithiobis-[2-nitrobenzoic] acid). The levels of reduced and native thiol groups were determined spectrophotometrically at 415 nm after the reaction with DTNB. To calculate the plasma concentration of disulfide the following formula was used: Disulfide levels (µmol/L)=(total thiol – native thiol)/2 [20].
DHCR24 measurement
Serum DHCR-24 levels were measured by the enzyme-linked immunosorbent assay (ELISA) using commercially available ELISA kits (USCN, catalog no: SEF185Hu). The sensitivity of these DHCR24 kits was 0.056 ng/mL and the assay ranges were 0.156–10 ng/mL with intra-and interassay CV% of less than 10%. The results were presented as ng/mL.
Periostin measurement
Serum periostin levels were assayed by the ELISA method using commercially available periostin ELISA Kits (Elabscience catalog no: E-EL-H2452). The sensitivity of these periostin kits was 0.1 ng/mL and the assay ranges were 0.16–10 ng/mL with intra-and interassay CV% of less than 10%. The results were presented as ng/mL.
TGF-β1 measurement
Serum TGF-β1 levels were assayed by the ELISA method using commercially available TGF-β1 ELISA Kits (Elabscience catalog no:E-EL-H0110). The sensitivity of these TGF-β1 kits was 18.75 pg/mL and the assay ranges were 31.25–2000 pg/mL with intra-and interassay CV% of less than 10%. All samples were diluted 10-fold before analysis. The results were presented as pg/mL.
Statistical analysis
All statistical analyses were carried out using SPSS software, version 23 for Mac (SPSS Inc, Chicago IL, USA). The normal distribution of data was evaluated by the Shapiro-Wilk test. Categorical variables were compared with the Chi-square test or Fisher’s Exact test. Comparisons of changes over time of non-normally distributed continuous variables were performed using the Friedman test. To examine the significance of pairwise differences, the Wilcoxon signed-rank test was used with Bonferroni adjustment to account for multiple comparisons. The Spearman rank-order correlation test was applied to determine correlations between different variables. A value of p<0.05 was considered statistically significant.
Results
The total 56 patients comprised 38 (67.9%) males and 18 (32.1%) females with a median age of 55.5 (32.0–70.5) years. The baseline characteristics of the patients are shown in Table 1.
Baseline characteristics of the patients with CHC.
| Characteristics | Total (n=56) |
|---|---|
| Age, years | 55.5 (32.0–70.5) |
| Age
|
20 (35.7) |
| Gender, male | 38 (67.9) |
| Treatment regimen | |
| EXV/VIE | 36 (64.3) |
| GLE/PIB | 20 (35.7) |
| Genotype 1b | 26 (46.4) |
| Log10 serum HCV-RNA, IU/mL | 3.9 (8.3–16.7) |
| ALT, U/L | 46 (28–61) |
| AST, U/L | 38.5 (27.5–47.0) |
| Total bilirubin, mg/dL | 0.73 (0.57–0.97) |
| Direct bilirubin,
|
0.23 (0.16–0.32) |
| Albumin, g/dL | 4.05 (3.80–4.50) |
| Total protein, g/dL | 6.45 (6.20–6.87) |
-
EXV/VIE, exviera/viekirax; GLE/PIB, glecaprevir/pibrentasvir; HCV, hepatitis C virus; RNA, ribonucleic acid; ALT, alanine aminotransferase; AST, aspartate aminotransferase; TGF-β, transforming growth factor beta. Continuous data were expressed as median
The serum levels of the thiol-disulfide hemostasis parameters (native thiol, total thiol and disulfide), TGF-β, periostin, DHCR24 at baseline, 1 month and EOT are shown in Table 2. From baseline to at the EOT, there was a significant decrease in serum levels of disulfide and TGF-β (p=0.020 and p<0.001, respectively) and a significant increase in serum native thiol levels (p=0.010) (Table 2, Figures 1, 2). There were no statistically significant differences in the serum levels of periostin, total thiol, and DHCR24 between those observed at baseline and at the EOT (Table 2, Figures 1, 2).
Changes in serum levels of periostin, TGF-β, DHCR24 and thiol-disulphide hemostasis and the APRI score with direct-acting antiviral therapy on the study population (n=56).
| Baseline | 1 Month | EOT | p-Value | |
|---|---|---|---|---|
| TGF-β, ng/mL | 12,019.25 (9,694.77–19,677.70) | 12,009.35b (8,639.22–19,045.67) | 9,648.50a (2,655.57–17,053.30) | <0.001 |
| Periostin, ng/mL | 4.74 (4.19–5.96) | 4.52 (3.83–5.27) | 4.44 (3.97–5.21) | 0.622 |
| DHCR24,
|
3.71 (3.39–4.33) | 4.05 (3.59–4.64) | 3.96 (3.50–4.40) | 0.561 |
| Total thiol,
|
392.33 (358.14–430.43) | 369.00b (342.97–407.86) | 402.30 (368.00–444.12) | 0.024 |
| Native thiol,
|
341.22 (314.25–363.31) | 342.18b (321.90–388.22) | 371.02a (330.23–396.38) | 0.010 |
| Disulfide,
|
20.54 (13.81–34.26) | 15.33 (10.73–26.34) | 15.00a (10.74–28.03) | 0.020 |
| APRI score | 0.47 (0.33–0.67) | 0.26a (0.19–0.38) | 0.23a (0.19–0.32) | <0.001 |
-
TGF-β, transforming growth factor beta; DHRC24, 3β-hydroxysterol Δ24-reductase; APRI, AST to platelet ratio index. ap<0.016 for comparison with levels observed at baseline, bp<0.016 for comparison with levels observed at the end of therapy. Continuous data were expressed as median

Comparison of the serum (A) native thiol, (B) total thiol and (C) disulphide levels in CHC cases at different time points. (A) *p=0.008 and **p=0.002, (B) *p=0.003, (C) *p=0.004. Data are presented with the Tukey box-and-whisker plot, where the middle line represents the median, and the box represents the IQR (the 25th and 75th percentiles).
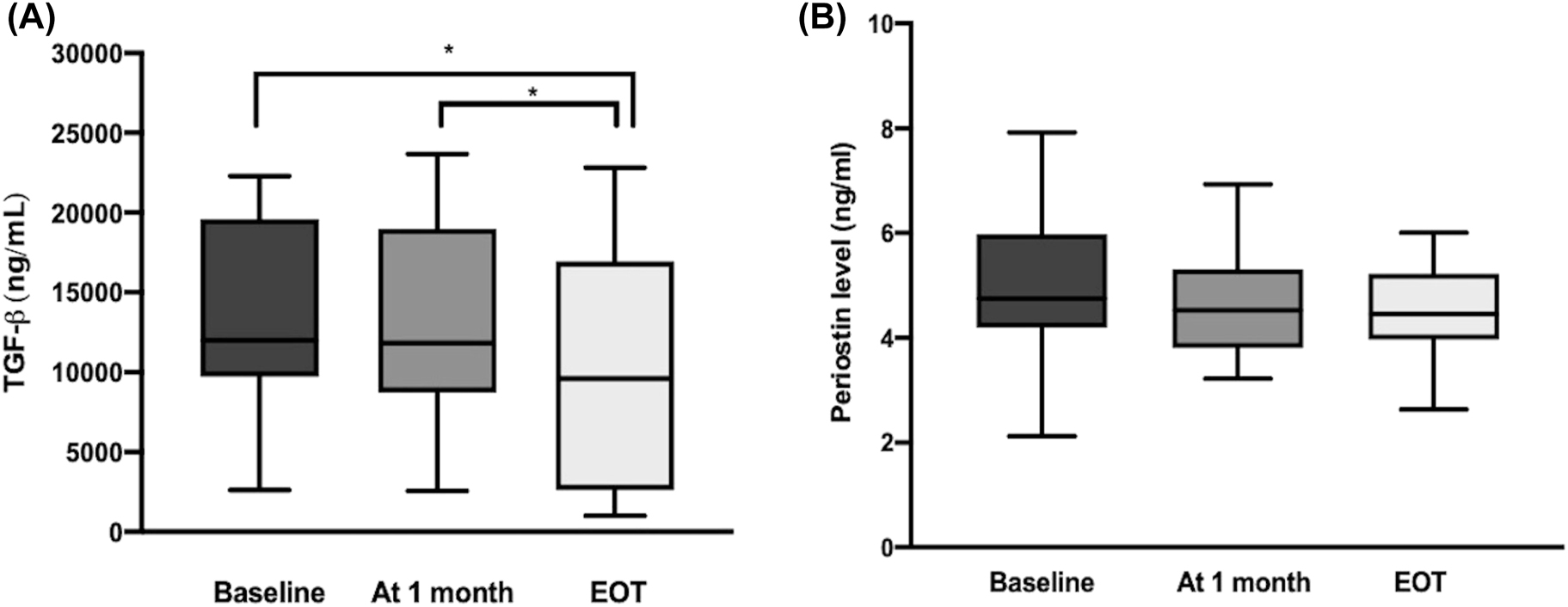
Comparison of (A) the serum TGF-β levels and (B) serum periostin levels in CHC cases at different time points. *p<0.001. Data are presented with the Tukey box-and-whisker plot, where the middle line represents the median, and the box represents the IQR (the 25th and 75th percentiles). TGF-β, transforming growth factor-beta; CHC, chronic hepatitis C.
There was a statistically significant decrease of the APRI scores in CHC patients at the 1 month and EOT of DAA therapy compared to baseline (p<0.001 for both). However, there was no correlation between APRI scores and TGF-beta (Spearman correlation coefficients were 0.126, p=0.384) (Table 2).
There was a significant moderate negative correlation between the levels of baseline serum albumin and TGF-β (rs=−0.483, p<0.001) and a moderate positive correlation between baseline serum total bilirubin levels and TGF-β (rs=0.470, p=0.001). A significant moderate positive correlation was determined between baseline serum DHRC24 and disulfide levels (rs=0.356, p=0.007) (Figure 3).

Correlation coefficients and p values (in parentheses) of the Spearman rank correlations between different variables. (A) TGF-β and albumin, (B) TGF-β and total bilirubin, (C) DHRC24 and disulfide. TGF-β
Figures were plotted with Prism 8.0 (GraphPad Software, La Jolla, CA, USA).
Discussion
It has been clearly established in several studies that there is a strong association between CHC infection and oxidative stress, which may also contribute to HCV-induced liver fibrogenesis and tumorigenesis [4, 5, 13]. However, very little is known about the reversibility of this impaired oxidative balance and associated fibrogenic and tumorigenic processes with new DAA regimens [11, 21]. Thus, in this study it was aimed to address these gaps in the literature. The results of the current study indicated that DAA therapy may help restore impaired oxidative balance and may have the additional benefit of decreasing the levels of TGF-β, the central fibrogenic factor as early as 8 weeks after the initiation of DAA therapy (at the EOT), which suggested that DAA therapy could help reverse the liver fibrosis promoted by TGF-β-induced MSCs.
Thiol-disulfide balance, which shifts towards disulfide bonds rather than thiol group formation has recently been shown to be one of the most prominent alternative indicators of oxidative stress recently. Thiols act as a major antioxidant and neutralize reactive oxygen molecules in cells. They are highly vulnerable to oxidation, which is based on their-SH group [10], [11], [12], [13], [14], [15], [16], [17], [18], [19], [20], [21], [22]. Under oxidative stress, the oxidation of the thiol (C-SH) group results in the formation of disulfide bonds (-S-S-). After the elimination of oxidative stress, those disulfide bonds can be reduced back to free thiol groups (-SH + -SH), and therefore the homeostasis between thiol-disulfide pairs is maintained [6, 23, 24]. In the current study, the significantly decreased disulfide levels might have been due to a decrease in oxidative stress in DAA-treated CHC patients. In addition, the significantly increased native thiol levels may have derived from the regaining of antioxidant defensive capacity after HCV cure. Nevertheless, according to recent evidence, it seems possible that these results are due to the antiviral effect of DAAs, which may reduce viral load, inflammation and possibly virus-induced oxidative stress, rather than a direct antioxidant effect. The parameters of dynamic thiol/disulfide balance have recently been studied as a novel oxidative stress indicator in CHC patients [8]. However, to the best of our knowledge, the current study is the first to have revealed the benefits of DAA therapy in restoring this impaired balance.
Chronic oxidative stress and concomitant increase in ROS have been shown to stimulate the TGF-β signaling pathway, which is a key driver for liver fibrogenesis [9, 25]. TGF-β is a profibrogenic cytokine, that induces hepatic stellate cell (HSC) activation and the production of extracellular matrix (ECM) proteins, such as collagen I and periostin. This consequently results in an enhanced ECM protein deposition, thereby promoting liver fibrosis. Moreover, ROS and TGF-β may also contribute indirectly to the fibrotic process via enhanced inflammation [25, 26]. Serum TGF-β levels have been shown to be increased in HCV-related chronic liver disease compared with control subjects, which correlates with fibrosis score [27, 28]. However, the reducing effect of DAA therapy on TGF-β levels and its relation with the regression on liver fibrosis has not been elucidated. The results of this study showed that DAA therapy has an improving effect on the serum levels of TGF-β.
We also evaluated the APRI score and compared its value before and after DAA therapy. We found that there was a statistically significant decrease in the APRI scores in CHC patients after DAA therapy at month 1, as well as at the EOT compared to baseline. Although there was no correlation between APRI scores and TGF-β, this finding supports the marked decrease in TGF-β levels with DAA therapy. A moderate positive correlation was also determined between baseline serum total bilirubin levels and TGF-β, whereas a moderate negative correlation was found between the levels of serum albumin and TGF-β. Plasma albumin (an indicator of liver synthesis capability) and bilirubin concentrations (an indicator of hepatic detox excretion capability) commonly served as laboratory indicators of the prognosis of liver diseases. Thus, these findings suggest that the lowering of TGF-β levels with DAAs may help reduce the progression of fibrogenesis in these patients [29]. Nevertheless, further studies are required to ascertain these results in the future.
Periostin, which is a matricellular protein involved in the process of liver fibrogenesis, is secreted by activated HSCs after stimulation with TGF-β. In turn, periostin induces the production of TGF-β and enhances its activity, which suggests that there may be a potential reciprocal regulatory interaction between TGF-β and periostin [12]. Interestingly, there was no significant change in serum levels of periostin during DAA therapy compared with the baseline in the current study. It is difficult to explain this result because there are no previous studies in the literature addressing this topic. One possible explanation for this might be that periostin regulation is influenced not only by the expression of TGF-
HCV-induced hepatic oxidative stress leads to increased oxidative DNA damage in hepatocytes, which plays a pivotal role in the initiation and progression of HCC. In this process, the overproduction of ROS can induce 3β-hydroxysterol Δ24-reductase (DHCR24) expression, which plays a part in the protection of hepatocytes from oxidative stress-induced apoptosis [13]. At this point, the DHCR24 suppresses apoptotic responses by impairing p53 activity, which may increase tumorigenicity [31]. Consistent with this evidence, a statistically significant moderate positive correlation was observed in the current study between baseline serum DHRC24 and disulfide levels. The level of serum DHCR24 is expected to fall after DAA therapy, as a result of decreased virus-induced oxidative stress. However, in the present study, no significant change was determined in serum DHCR24 levels with DAA therapy. This result may be explained by the fact that the complete biochemical improvement after DAA therapy might require a longer disease-free period [17]. Thus, future studies with longer follow-up periods are required to investigate the long-term changes in plasma DHCR24 Ab levels after DAA therapy.
Despite these encouraging results, this study had some limitations, primarily that it was a small single-center study. Another limitation was that only non-cirrhotic patients were examined, so these results may not be applicable to patients with cirrhosis. Nevertheless, the fact that three serum samples were obtained at different time points during treatment from each patient for comparison is the main strength of this study.
In conclusion, the results of this study have shown that DAA therapy may help restore impaired thiol/disulfide balance induced by HCV. The second major finding was that DAA therapy can markedly decrease serum levels of TGF-β, which have been found to be associated with a reduction in serum total bilirubin (an indicator of hepatic detox excretion capability) and an increase in plasma albumin concentrations (an indicator of liver synthesis capability), commonly served as laboratory indicators of prognosis of liver diseases. Thus these results suggest that the DAA therapy may help to reduce the pro-fibrotic process in CHC patients by markedly decreasing serum levels of TGF-β, a key player in HCV-induced liver fibrosis. However, there is a need for further studies with larger cohorts and longer follow-up periods in order to clarify this issue.
-
Research funding: This work was supported by the Scientific Research Project Committee of Mustafa Kemal University, Hatay, Turkey (Project number: 19.M.026).
-
Author contributions: All authors have accepted responsibility for the entire content of this manuscript and approved its submission.
-
Competing interests: Authors state no conflict of interest.
-
Informed consent: Informed consent was obtained from all individuals included in this study.
-
Ethical approval: The study protocol was approved by the Ethics Committee of Mustafa Kemal University, Faculty of Medicine (2020/78).
References
1. Gower, E, Estes, C, Blach, S, Razavi-Shearer, K, Razavi, H. Global epidemiology and genotype distribution of the hepatitis C virus infection. J Hepatol 2014;61:45–57. https://doi.org/10.1016/j.jhep.2014.07.027.Search in Google Scholar PubMed
2. Nitshitsuji, H, Funami, K, Shimizu, Y, Ujino, S, Sugiyama, K, Seya, T, et al.. Hepatitis C virus infection induced inflammatory cytokines and chemokines mediated by the cross talk between hepatocytes and stellate cells. J Virol 2013;87:8169–78.10.1128/JVI.00974-13Search in Google Scholar PubMed PubMed Central
3. Anticoli, S, Amatore, D, Matarrese, P, De Angelis, M, Palarama, AT, Nencioni, L, et al.. Counteraction of HCV-induced oxidative stress concurs to establish chronic infection in liver cell cultures. Oxid Med Cell Longev 2019;2019:3712969. https://doi.org/10.1155/2019/6452390.Search in Google Scholar PubMed PubMed Central
4. Rebbani, Tsukiyama-Kohara K. HCV-induced oxidative stress: battlefield-winning strategy. Oxid Med Cell Longev 2016;2016:7425628. https://doi.org/10.1155/2016/7425628.Search in Google Scholar PubMed PubMed Central
5. Li, S, Tan, HY, Wang, N, Zhang, ZJ, Lao, L, Wong, CW, et al.. The role of oxidative stress and antioxidants in liver diseases. Int J Mol Sci 2015;16:26087–124. https://doi.org/10.3390/ijms161125942.Search in Google Scholar PubMed PubMed Central
6. Biswas, S, Chida, AS, Rahman, I. Redox modifications of protein-thiols: emerging roles in cell signaling. Biochem Pharmacol 2006;71:551–64. https://doi.org/10.1016/j.bcp.2005.10.044.Search in Google Scholar PubMed
7. Erel, O, Neselioglu, S. A novel and automated assay for thiol/disulphide homeostasis. Clin Biochem 2014;47:326–32. https://doi.org/10.1016/j.clinbiochem.2014.09.026.Search in Google Scholar PubMed
8. Damar Cakırca, T, Ceylan, MR, Koyuncu, I, Çakırca, G. Thiol-disulfide balance and total oxidant-antioxidant status in patients with chronic hepatitis C. Int J Clin Pract 2021;75:e13988. https://doi.org/10.1111/ijcp.13988.Search in Google Scholar PubMed
9. Dewidar, B, Meyer, C, Dooley, S, Meindl-Beinker, AN. TGF-β in hepatic stellate cell activation and liver fibrogenesis-updated 2019. Cells 2019;8:1419. https://doi.org/10.3390/cells8111419.Search in Google Scholar PubMed PubMed Central
10. Clemente, M, Núñez, O, Lorente, R, Rincon, D, Matilla, A, Salcedo, M, et al.. Increased intrahepatic and circulating levels of endoglin, a TGF-beta1 co-receptor, in patients with chronic hepatitis C virus infection: relationship to histological and serum markers of hepatic fibrosis. J Viral Hepat 2006;13:625–32. https://doi.org/10.1111/j.1365-2893.2006.00733.x.Search in Google Scholar PubMed
11. Kotsiri, I, Hadziyannis, E, Georgiou, A, Papageorgiou, MV, Vlachogiannakos, I, Papatheodoridis, G. Changes in serum transforming growth factor-β1 levels in chronic hepatitis C patients under antiviral therapy. Ann Gastroenterol 2016;29:79–84.Search in Google Scholar
12. Huang, Y, Liu, W, Xiao, H, Maitikabili, A, Lin, Q, Wu, T, et al.. Matricellular protein periostin contributes to hepatic inflammation and fibrosis. Am J Pathol 2015;185:786–97. https://doi.org/10.1016/j.ajpath.2014.11.002.Search in Google Scholar PubMed
13. Tsukiyama-Kohara, K. Role of oxidative stress in hepatocarcinogenesis induced by hepatitis C virus. Int J Mol Sci 2012;13:15271–8. https://doi.org/10.3390/ijms131115271.Search in Google Scholar PubMed PubMed Central
14. Saito, M, Takano, T, Nishimura, T, Kohara, M, Tsukiyama-Kohara, K. 3β-hydroxysterol δ24-reductase on the surface of hepatitis C virus-related hepatocellular carcinoma cells can be a target for molecular targeting therapy. PLoS One 2015;10:e0124197. https://doi.org/10.1371/journal.pone.0124197.Search in Google Scholar PubMed PubMed Central
15. Ezzikouri, S, Kimura, K, Sunagozaka, H, Kaneko, S, Inoue, K, Nishimura, T, et al.. Serum DHCR24 auto-antibody as a new biomarker for progression of hepatitis C. EBioMedicine 2015;2:604–12. https://doi.org/10.1016/j.ebiom.2015.04.007.Search in Google Scholar PubMed PubMed Central
16. Cabalak, M, Bal, T, Onlen, Y, Demir, M. Incidence and predictors of direct-acting antiviral treatment failure in Turkish patients with chronic hepatitis C genotype 1b infection. Trop Doct 2020;50:141–6. https://doi.org/10.1177/0049475519892082.Search in Google Scholar PubMed
17. Hengst, J, Falk, CS, Schlaphoff, V, Deterding, K, Manns, MP, Cornberg, M, et al.. Direct-acting antiviral-induced hepatitis C virus clearance does not completely restore the altered cytokine and chemokine milieu in patients with chronic hepatitis C. J Infect Dis 2016;214:1965–74. https://doi.org/10.1093/infdis/jiw457.Search in Google Scholar PubMed
18. Li, H, Huang, MH, Jiang, JD, Peng, ZG. Hepatitis, C: from inflammatory pathogenesis to anti-inflammatory/hepatoprotective therapy. World J Gastroenterol 2018;24:5297–311. https://doi.org/10.3748/wjg.v24.i47.5297.Search in Google Scholar PubMed PubMed Central
19. Wai, CT, Greenson, JK, Fontana, RJ, Kalbfleisch, JD, Marrero, JA, Conjeevaram, HS, et al.. A simple noninvasive index can predict both significant fibrosis and cirrhosis in patients with chronic hepatitis C. Hepatology 2003;38:518–26. https://doi.org/10.1053/jhep.2003.50346.Search in Google Scholar PubMed
20. Uyanıkoğlu, A, Sabuncu, T, Yıldız, R, Cindioğlu, Ç, Kirmit, A, Erel, Ö. Impaired thiol/disulfide homeostasis in patients with mild acute pancreatitis. Turk J Gastroenterol 2019;30:899–902. https://doi.org/10.5152/tjg.2019.18775.Search in Google Scholar PubMed PubMed Central
21. Serejo, F, Emerit, I, Filipe, PM, Fernandes, AC, Costa, MA, Freitas, JP, et al.. Oxidative stress in chronic hepatitis C: the effect of interferon therapy and correlation with pathological features. Can J Gastroenterol 2003;17:644–50. https://doi.org/10.1155/2003/710693.Search in Google Scholar PubMed
22. Erel, Ö, Erdoğan, S. Thiol-disulfide homeostasis: an integrated approach with biochemical and clinical aspects. Turk J Med Sci 2020;50:1728–38. https://doi.org/10.3906/sag-2003-64.Search in Google Scholar PubMed PubMed Central
23. Cremers, CM, Jakob, U. Oxidant sensing by reversible disulfide bond formation. J Biol Chem 2013;288:26489–96. https://doi.org/10.1074/jbc.r113.462929.Search in Google Scholar PubMed PubMed Central
24. Vázquez-Torres, A. Redox active thiol sensors of oxidative and nitrosative stress. Antioxidants Redox Signal 2012;17:1201–14.10.1089/ars.2012.4522Search in Google Scholar PubMed PubMed Central
25. Fabregat, I, Moreno-Càceres, J, Sánchez, A, Dooley, S, Dewidar, B, Giannelli, G, et al.. IT-LIVER Consortium. TGF-β signalling and liver disease. FEBS J 2016;283:2219–32. https://doi.org/10.1111/febs.13665.Search in Google Scholar PubMed
26. Richter, K, Kietzmann, T. Reactive oxygen species and fibrosis: further evidence of a significant liaison. Cell Tissue Res 2016;365:591–605. https://doi.org/10.1007/s00441-016-2445-3.Search in Google Scholar PubMed PubMed Central
27. Nagaraja, T, Chen, L, Balasubramanian, A, Groopman, JE, Ghoshal, K, Jacob, ST, et al.. Activation of the connective tissue growth factor (CTGF)-transforming growth factor β 1 (TGF-β 1) axis in hepatitis C virus-expressing hepatocytes. PLoS One 2012;7:e46526. https://doi.org/10.1371/journal.pone.0046526.Search in Google Scholar PubMed PubMed Central
28. Marcellin, P, Asselah, T, Boyer, N. Fibrosis and disease progression in hepatitis C. Hepatology 2002;36:S47–56. https://doi.org/10.1053/jhep.2002.36993.Search in Google Scholar PubMed
29. Lucey, MR, Brown, KA, Everson, GT, Fung, JJ, Gish, R, Keefe, EB, et al.. Minimal criteria for placement of adults on the liver transplant waiting list: a report of a national conference organized by the American Society of Transplant Physicians and the American Association for the Study of Liver Diseases. Liver Transplant Surg 1997;3:628–37. https://doi.org/10.1002/lt.500030613.Search in Google Scholar PubMed
30. Kudo, A, Kii, I. Periostin function in communication with extracellular matrices. J Cell Commun Signal 2018;12:301–8. https://doi.org/10.1007/s12079-017-0422-6.Search in Google Scholar PubMed PubMed Central
31. Nishimura, T, Kohara, M, Izumi, K, Kasama, Y, Hirata, Y, Huang, Y, et al.. Hepatitis C virus impairs p53 via persistent overexpression of 3beta-hydroxysterol Delta24-reductase. J Biol Chem 2009;284:36442–52. https://doi.org/10.1074/jbc.m109.043232.Search in Google Scholar PubMed PubMed Central
© 2022 the author(s), published by De Gruyter, Berlin/Boston
This work is licensed under the Creative Commons Attribution 4.0 International License.
Articles in the same Issue
- Frontmatter
- Opinion Paper
- Ischemia – modified albumin by albumin cobalt binding test: a false myth or reality
- Research Articles
- Machine learning models can predict the presence of variants in hemoglobin: artificial neural network-based recognition of human hemoglobin variants by HPLC
- Investigation of the preanalytical process practices in primary care in Istanbul regarding the newborn screening tests
- Cell counting chamber vs. Sysmex XN-1000 for determining white blood cell count and differentiation for body fluids
- Synthesis of PEITC-loaded gold nanoparticles and evaluation of the hepatoprotective effect on CCl4-induced damage through Nrf2 pathway
- Concentrations of B cell-activating factor, aquaporin-4 antibody and brain-derived neurotrophic factor in neuromyelitis optica spectrum disorder
- Macula, choroid and retinal nerve fiber layer optical coherence tomography biomarkers in liver dysfunction
- Direct-acting antiviral therapy may help restore HCV-induced impaired redox balance and liver fibrosis process
- Are VEGF and SCUBE1 gene expressions increased in diabetic retinopathy?
- Biochemical analysis of microbiotas obtained from healthy, prediabetic, type 2 diabetes, and obese individuals
- Inflammation parameters, monocyte subgroups and toll-like receptor expression before and after dialysis in patients with chronic kidney disease
- Matrix metalloproteinase 9 gene-MMP9-DNA methylation status in Turkish schizophrenia patients
- HLA DRB1 alleles, IFN-γ and TGF-β Gene Variants in childhood ALL patients
- Epithelial-mesenchymal transition as a potential route for DAPT resistance in breast cancer cells
- Comparison with molecular effects of ukrain, tamoxifen, and docetaxel on human breast cancer cell lines
- In vitro evaluation of 2-pyrazoline derivatives as DPP-4 inhibitors
- Effects of polyphenolic-rich extracts from Citrus hystrix on proliferation and oxidative stress in breast and colorectal cancer
- The effects of sodium benzoate exposure on learning and neurobehavior during the prepubertal period in rats
- The association between βeta 2-microglobulin and bronchopulmonary dysplasia
Articles in the same Issue
- Frontmatter
- Opinion Paper
- Ischemia – modified albumin by albumin cobalt binding test: a false myth or reality
- Research Articles
- Machine learning models can predict the presence of variants in hemoglobin: artificial neural network-based recognition of human hemoglobin variants by HPLC
- Investigation of the preanalytical process practices in primary care in Istanbul regarding the newborn screening tests
- Cell counting chamber vs. Sysmex XN-1000 for determining white blood cell count and differentiation for body fluids
- Synthesis of PEITC-loaded gold nanoparticles and evaluation of the hepatoprotective effect on CCl4-induced damage through Nrf2 pathway
- Concentrations of B cell-activating factor, aquaporin-4 antibody and brain-derived neurotrophic factor in neuromyelitis optica spectrum disorder
- Macula, choroid and retinal nerve fiber layer optical coherence tomography biomarkers in liver dysfunction
- Direct-acting antiviral therapy may help restore HCV-induced impaired redox balance and liver fibrosis process
- Are VEGF and SCUBE1 gene expressions increased in diabetic retinopathy?
- Biochemical analysis of microbiotas obtained from healthy, prediabetic, type 2 diabetes, and obese individuals
- Inflammation parameters, monocyte subgroups and toll-like receptor expression before and after dialysis in patients with chronic kidney disease
- Matrix metalloproteinase 9 gene-MMP9-DNA methylation status in Turkish schizophrenia patients
- HLA DRB1 alleles, IFN-γ and TGF-β Gene Variants in childhood ALL patients
- Epithelial-mesenchymal transition as a potential route for DAPT resistance in breast cancer cells
- Comparison with molecular effects of ukrain, tamoxifen, and docetaxel on human breast cancer cell lines
- In vitro evaluation of 2-pyrazoline derivatives as DPP-4 inhibitors
- Effects of polyphenolic-rich extracts from Citrus hystrix on proliferation and oxidative stress in breast and colorectal cancer
- The effects of sodium benzoate exposure on learning and neurobehavior during the prepubertal period in rats
- The association between βeta 2-microglobulin and bronchopulmonary dysplasia