Gedächtnismechanismen in Drosophila
-
Johannes Felsenberg
Johannes Felsenberg ist Post-Doc in Scott Waddells Labor am Centre for Neural Circuits and Behaviour an der Universität Oxford. Sein Diplom in Biologie und seine Promotion hat er an der Freien Universität Berlin erlangt. In den Laboren von Randolf Menzel und Dorothea Eisenhardt hat er die Rolle des Ubiquitin Proteasome Systems beim Lernen und der Gedächtnisbildung von Honigbienen erforscht. In Oxford arbeitet Johannes an den Mechanismen der Gedächtnis-Rekonsolidierung und der extinktionsinduzierten synaptischen Plastizität im Pilzkörper der Taufliege
Drosophila . Sein Projekt im Waddell-Labor ist unterstützt durch ein DFG-Stipendium. and
David Owald
and
David Owald
David Owald leitet die Emmy-Noether-Nachwuchsgruppe
Behavioral and Physiological Neurogenetics am Institut für Neurophysiologie der Charité in Berlin. Er promovierte an der Universität Göttingen über die molekulare Anatomie und entwicklungsbedingte Plastizität von präsynaptischen aktiven Zonen inDrosophila . Danach war er als Junior Research Fellow für Physiologie am Wolfson College Oxford und als EMBO Long-term und als Sir Henry Wellcome Postdoctoral Fellow am Centre for Neural Circuits and Behaviour an der Universität Oxford tätig. Dort beschäftigte er sich hauptsächlich mit den Mechanismen lernrelevanter Plastizität imDrosophila -Pilzkörper. David erhielt 2017 denSchilling-Forschungspreis der Neurowissenschaftlichen Gesellschaft .

Zusammenfassung
Zielgerichtetes Verhalten wird durch neuronale Schaltkreise im Gehirn gesteuert. Erfahrungen können die Dynamiken dieser neuronalen Schaltkreise verändern, indem spezifische synaptische Verbindungen modifiziert werden. Die Lokalisierung einer solchen lerninduzierten synaptischen Plastizität hat sich jedoch bis heute als anspruchsvoll erwiesen. Technische Fortschritte in der Kontrolle und Messung von neuronaler Aktivität haben es nun ermöglicht, die dem Lernen und der Gedächtnisbildung zugrunde liegenden Strukturen im Gehirn des Modelsystems Drosophila in vivo zu charakterisieren. Die Taufliege (Drosophila melanogaster) hat ein numerisch einfacheres Gehirn, was die Erforschung identifizierter Netzwerkkomponenten realisierbar macht. In diesem Artikel erörtern wir die Mechanismen, auf denen assoziatives Lernen, Gedächtnisabruf sowie die Reevaluierung von existierenden Gedächtnissen in Drosophila beruhen.
Sensorische Reize werden durch neuronale Operationen als Aktivitätsmuster spezifischer Neurone in definierten Schaltkreisen im Gehirn abgebildet. Assoziative Gedächtnisse bilden sich beispielsweise aus, wenn einem zuvor neutralen Reiz eine Wertigkeit zugeschrieben wird. Die hierbei involvierten Netzwerke lassen sich in Hirnarealen lokalisieren, die der primären Reizverarbeitung nachgeschaltet sind. Dort können Informationen sowohl miteinander verknüpft als auch entsprechende nachfolgende motorische Programme angesteuert werden. Im Säugetiergehirn sind solche Regionen beispielsweise der Hippocampus oder die Amygdala (Tovote et al., 2015). Im Insektengehirn erfüllt der Pilzkörper eine solche Funktion (Stevens, 2015; Menzel, 2014).
Neurone sind über Synapsen miteinander verknüpft und bilden so neuronale Schaltkreise. Es wird allgemein angenommen, dass Gedächtnisbildung durch die anhaltende Stärkung oder Abschwächung synaptischer Verknüpfungen realisiert wird, wobei angemerkt werden muss, dass direkte Evidenzen für solch ein Model im sich verhaltenden Tier rar sind. Eine solche lerninduzierte Plastizität setzt voraus, dass die Aktivität von sekundären Botenstoffen sowie komplexer Proteinmaschinerien an Synapsen exakt reguliert werden kann. Tatsächlich können Mutationen von Genen, die involvierte Proteine kodieren, zu anormaler neuronaler Aktivität führen (Südhof, 2012).
Die Grundlage für die Erkenntnis des engen Zusammenhangs zwischen genetischer Information, neuronalen Netzwerken und Verhaltensmerkmalen wurde in den frühen 1970er-Jahren im Labor von Seymour Benzer gelegt. Die angewandten Methoden waren einfach, aber brillant. Taufliegen, Drosophila melanogaster, wurden genetisch verändert und spezifischen Verhaltenstests unterzogen. Dieser Ansatz führte zur Identifikation zahlreicher genetischer Programme, die auf evolutionärer Ebene hochkonserviert sind. Ein in diesem Zusammenhang entdeckter Genlokus ist etwa der zirkadiane Faktor Period (Konopka und Benzer, 1971). Zu den im Benzer-Labor untersuchten Verhaltensparadigmen gehörten auch die ersten Analysen von assoziativen Gedächtnissen (Quinn und Dudai, 1976), die als Grundstein der hier diskutierten Erkenntnisse zu betrachten sind.
Fortschritte im Verständnis der neuronalen Prinzipien, die dem Lernen und der Gedächtnisspeicherung zugrunde liegen, werden durch die numerische Komplexität des Vertebraten-Nervensystems erschwert. Der Modellorganismus Drosophila erlaubt genetische und verhaltensorientierte Ansätze und bietet darüber hinaus die Möglichkeit, physiologische Fragestellungen in einem relativ simplen Gehirn zu bearbeiten. So ist es unter anderem möglich, mithilfe genetischer Strategien einzelne Neurone reproduzierbar zu manipulieren, d. h. zu aktivieren oder zu blockieren und/oder die Aktivität dieser Neurone in vivo zu messen (siehe Exkursion 1). Die relative (numerische) Einfachheit und die experimentelle Zugänglichkeit des Drosophila-Nervensystems haben dazu geführt, dass ein grundlegendes Verständnis entwickelt werden konnte, welche synaptischen Verbindungen im Drosophila-Gehirn lerninduziert verändert werden, wie diese Veränderungen induziert werden und wie diese Plastizität wiederum mit dem erlernten Verhalten eines Tieres in Verbindung steht.
Exkursion 1: Drosophila können problemlos in großen Mengen im Labor gehalten werden und genetische Modifizierungen bestimmter Chromosome ermöglichen es, Mutationen über Generationen hinweg stabil zu halten. Darüber hinaus besitzen Drosophila nur drei Chromosomen (sowie ein sehr kleines Viertes), sodass die genetische Komplexität gewissermaßen überschaubar bleibt.
Die Anwendung der Transgenese (Rubin and Spradling, 1982) und der Einsatz binärer Expressionssysteme ermöglichten wichtige Durchbrüche bezogen auf die Beschreibung verhaltensrelevanter neuronaler Netzwerke. So wurden Transgene hergestellt, die es ermöglichen, Neurotransmission in definierten Zeitfenstern zu blockieren oder Neurone durch licht- beziehungsweise temperaturabhängige Ionenkanäle zeitlich präzise zu aktivieren (Optogenetik bzw. Thermogenetik). Diese Techniken sind von enormen Wert für die Untersuchung der aktiven Beteiligung von Neuronen an Verhaltensprogrammen. Transgenese ermöglicht außerdem den Gebrauch binärer Expressionssysteme (Brand and Perrimon, 1993), in denen Transkriptionsfaktoren und die zugehörigen DNS-Elemente von anderen Organismen entliehen und unter die Kontrolle von Drosophila-Enhancer-Sequenzen gesetzt werden. Solche Systeme erlauben die Kontrolle darüber, wo und wann spezifische Gene exprimiert werden. Mithilfe verschiedener Verfeinerungen dieses Systems lassen sich Transgene, wie zum Beispiel solche, die optogenetische Effektoren kodieren, spezifisch in identifizierten Populationen von Neuronen – in vielen Fällen sogar in einzelnen Neuronen – exprimieren (Owald et al., 2015b). Diese genetischen Prinzipien ermöglichen nicht nur die Aktivitätsmanipulation von Neuronen, sondern erlauben auch, Aktivitätsmuster definierter Neurone zu messen. Der kombinierte Gebrauch von Effektoren und Aktivitätsreportern, beispielsweise genetisch kodierter Kalzium-Indikatoren, hat sich als essentiell für die neuronale Schaltkreiskartierung erwiesen.
Vom Stimulus zur Bedeutung
Die Welt stellt Tieren die ständige Herausforderung, unzählige sensorische Reize wahrzunehmen und über deren Relevanz zu entscheiden. Um grundlegende Aufgaben des Lebens wie Fortpflanzung und Überleben sicherzustellen, müssen Tiere lernen, mithilfe dieser sensorischen Reize Futterquellen sowie Fortpflanzungsmöglichkeiten zu finden und potenzielle Gefahren zu vermeiden. Es ist somit nicht überraschend, dass Taufliegen olfaktorische und visuelle Reize mit Nahrungsquellen (Belohnung) oder unangenehmen und gefährlichen Stimuli (Bestrafung) assoziieren können. Forschungen der letzten Jahrzehnte haben den Pilzkörper, ein Neuropil dritter Ordnung, als das Zentrum für assoziatives Lernen im Insektengehirn identifiziert (Heisenberg, 2003; Menzel, 2014). Das für die Gedächtnisspeicherung essenzielle Pilzkörpernetzwerk der Taufliege besteht aus weniger als 3000 Neuronen pro Gehirnhälfte (Hemisphäre).
Die Aktivität von intrinsischen Pilzkörperzellen, den Kenyon-Zellen (Kenyon cells, KCs, ca. 2000 Zellen pro Hemisphäre), erstellt eine Repräsentation der sensorischen, besonders der olfaktorischen, Umwelt. Gerüche werden von sensorischen Rezeptorneuronen detektiert und der entstandene Reiz wird im Antennallobus weiterverarbeitet und schließlich über Projektionsneurone an die KCs übermittelt. Jeder eingehende Duft aktiviert eine kleine spezifische Gruppe an KCs. Es ist dieses Aktivitätsprofil, das die Geruchsidentität innerhalb des Pilzkörpernetzwerks kodiert (Stevens, 2015). Die überwiegend cholinergen KCs (Barnstedt et al., 2016) entsenden parallele Axonenbündel, welche die Pilzkörperloben formen, und bilden erregende En-passant-Synapsen mit einer geringen Anzahl an nachgeschalteten Partnern, den pilzkörperextrinsischen Neuronen (Mushroom body output neurons, MBONs; siehe Abbildung 1a).
Aufgrund ihrer Anatomie werden die 35 MBONs pro Gehirnhälfte in 21 Typen unterteilt. Jeder MBON-Typ ist definiert über sein spezifisches, abgegrenztes, dendritisches Innervierungsmuster des Pilzkörpers. Dieses eindeutige Muster erlaubt, die entsprechenden MBON-Typen in jedem Fliegengehirn verlässlich zu identifizieren. Im Generellen überlappen diese Bereiche nicht mit denen anderer MBON-Typen. So „kacheln“ die Dendriten der MBON-Typen die Pilzkörperloben und bilden dezidierte Kompartimente (Tanaka et al., 2008; Aso et al., 2014b)
Interessanterweise genügt die Manipulation der Aktivität einzelner MBON-Typen, um die Verhaltensreaktion einer Taufliege auf einen Duft zu verändern (Aso et al., 2014a; Owald et al., 2015a; Perisse et al., 2016). So fördern bestimmte Typen von MBONs eine geruchsinduzierte Hinwendungsreaktion (positive MBONs), während andere Typen geruchsgesteuertes Vermeidungsverhalten vermitteln (aversive MBONs, Abbildung 1b). Naive Taufliegen, denen die Wahl zwischen einem reinen Luftstrom und einem abstoßenden Geruch gegeben wird, vermeiden normalerweise diesen Duft. Wenn aber experimentell aversive MBONs akut blockiert werden, verändern die Taufliegen ihr Verhalten und sie wenden sich nun dem initial abstoßenden Geruch zu. Werden dagegen die positiven MBONs während der Entscheidungssituation ausgeschaltet, verstärkt sich das Vermeidungsverhalten (Owald et al., 2015a; Perisse et al., 2016). Diese Klassifizierung der extrinsischen Pilzkörperneurone in positive und aversive MBONs wird weiter durch optogenetische Experimente gestützt (Aso et al., 2014a; Owald et al., 2015a): Naive Fliegen, die lichtaktivierbare Kationenkanäle (zum Beispiel CsChRimson) in einzelnen MBONs exprimieren, werden vor die Wahl gestellt, diese MBONs entweder zu aktivieren oder deren Aktivierung zu vermeiden. Tatsächlich vermeiden die Taufliegen, bei denen CsChRimson in aversiven MBONs exprimiert wird, überwiegend deren lichtinduzierte Aktivierung. Sie nähern sich hingegen der Lichtquelle, wenn diese Kationenkanäle in positiven MBONs exprimiert werden. Zusammen genommen legen diese Experimente ein Netzwerkmodel nahe, in dem der Identität eines Geruchs an der Synapse zwischen KCs und MBONs (Heisenberg, 2003) eine Bedeutung, eine Valenz, zugeordnet wird.
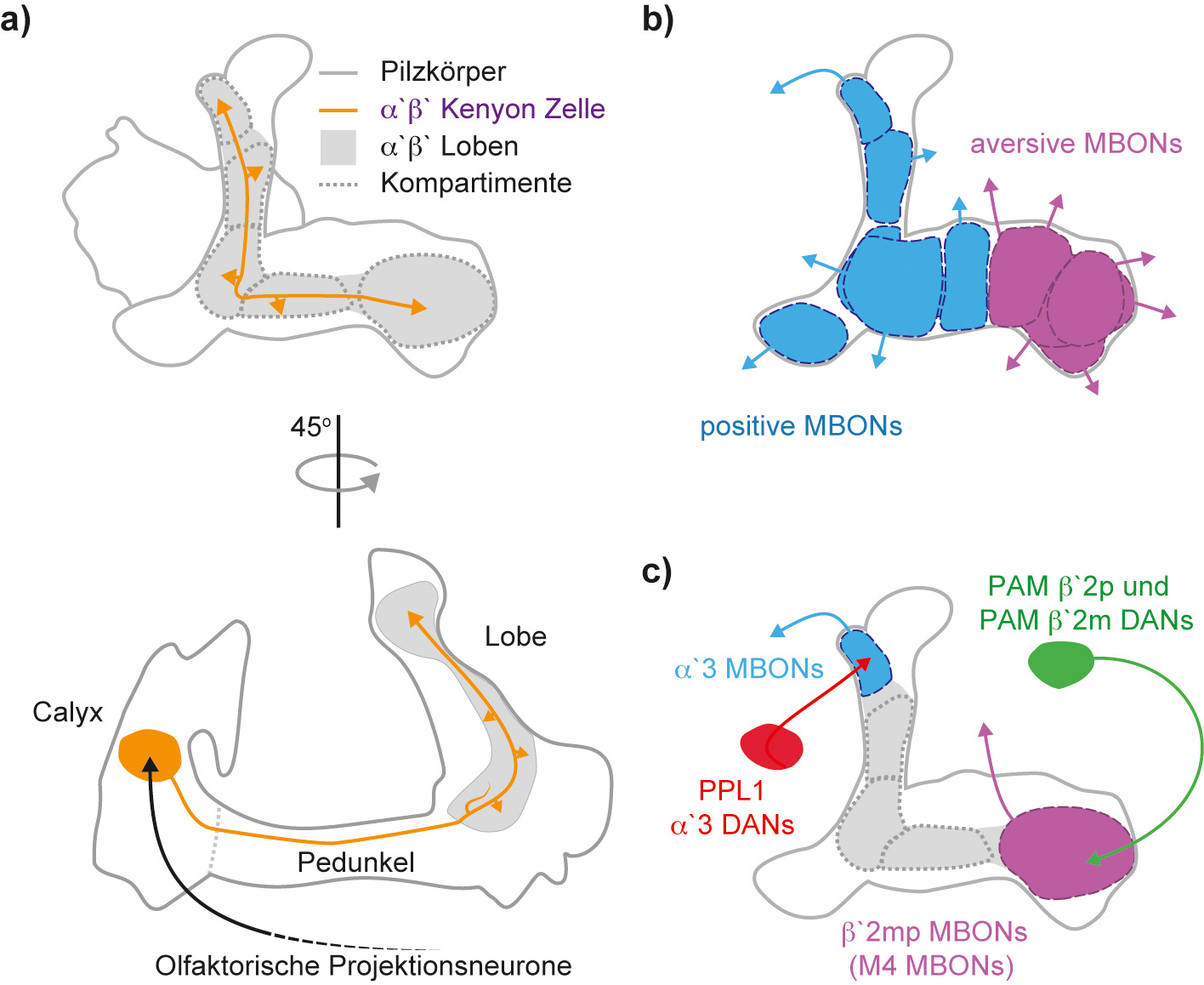
Komponenten des Pilzkörpers a) Kenyon-Zellen (Kenyon cells; KCs) formen die Pilzkörper (mushroom bodies; MBs; graue durchgezogene Linie). Es gibt drei Haupttypen von KCs: die αβ, die α′β′ (in violett, eine Beispiel-Kenyon-Zelle) und die γ KCs. KCs erhalten sensorischen Input von Projektionsneuronen (schwarz) im Calyx des Pilzkörpers. KC-Neuriten projizieren entlang des Pedunkels zu den Loben (grau). In den Loben bilden sie en passant erregende Synapsen (Pfeilspitzen) mit nachgeschalteten Neuronen, den pilzkörperextrinsischen Neuronen (mushroom body output neurons; MBONs). b) Bestimmte Typen von MBONs vermitteln Vermeidungsreaktionen bzw. Hinwendungsverhalten (positive MBONs in blau und aversive MBONs in violett). c) Die dendritischen Innervationsmuster jedes MBON-Typs (zwei Beispiele, blau und violett) decken sich mit der Innervation einer entsprechenden Dopamin-Neuronen-Gruppe (DANs, rot und grün). Zusammen definieren sie Kompartimente des Pilzkörpers (gestrichelte Linien). Nicht alle MBON-Typen vermitteln aversives Verhalten oder Hinwendungsverhalten. Kürzlich wurde gezeigt, dass das α′3 MBON die Neuartigkeit eines Reizes kodiert (Haitori et al 2017). Es bleibt festzuhalten, dass die genauere Untersuchung einiger MBONs noch aussteht.
Werte zuordnen
Wie bei Säugetieren ist das assoziative Lernen der Fliege eng mit dem dopaminergen System verknüpft. Die Pilzkörperloben werden von Dopamin-Neuronen aus zwei verschiedenen Clustern innerviert, deren Aktivität während des assoziativen Lernens notwendig ist. Aversives Lernen, die Assoziation eines Dufts mit einer Bestrafung, bedarf der Aktivität dopaminerger Neurone des kleineren PPL1-Clusters. Unterschiedliche Bestrafungen, so wie Elektroschocks, Hitze oder bittere Substanzen, scheinen hierbei alle über die gleichen bestrafungsvermittelnden PPL1-Dopamin-Neurone vermittelt zu werden [zur Übersicht siehe (Waddell, 2013)].
Das Belohnungslernen dagegen benötigt die Aktivität unterschiedlicher belohnungsspezifischer Dopamin-Neurone aus dem sogenannten PAM-Cluster. So wird das Signal für eine Wasserbelohnung über bestimmte Dopamin-Neurone vermittelt, die sich von den dopaminergen Zellen, die Zuckerbelohnung vermitteln, unterscheiden (Burke et al., 2012; Liu et al., 2012; Lin et al., 2014). Die zuckerbasierte Belohnung lässt sich auf neuronaler Ebene sogar noch weiter differenzieren. Die für das Erlernen eines belohnenden süßen Geschmacks verantwortlichen Dopamin-Neurone unterscheiden sich von solchen, die das Lernen über den Nährwertgehalt vermitteln (Huetteroth et al., 2015; Yamagata et al., 2015). Auffällig ist hierbei, dass die Axone dieser spezifischen Gruppen von Dopamin-Neuronen separate, nicht-überlappende Kompartimente in den Pilzkörperloben innervieren. Diese diskreten Innervierungsmuster decken sich exakt mit der Kompartimentalisierung des Pilzkörpers durch die Dendriten der unterschiedlichen MBON-Typen (Aso et al., 2014a; Owald et al., 2015a).
Dadurch ergibt sich, dass die einzelnen Gruppen von Dopamin-Neuronen bestimmte Synapsen zwischen KCs und einzelnen MBON-Typen in jeweils einem spezifischen Kompartiment beeinflussen. Diese anatomische Separation lässt vermuten, dass Erinnerungen an Bestrafungen und verschiedene Belohnungen in spezifischen KC-MBON-Synapsen des jeweiligen Kompartiments gespeichert werden. Somit bildet diese Anordnung das Grundgerüst für ein differenzierbares Gedächtnisauslese und -schreibsystem.
Gedächtnisse schreiben und lesen: Von der Synapse zum Netzwerk
Bestrafungsvermittelnde Dopamin-Neurone innervieren die Kompartimente, in denen sich die Dendriten der positiven MBON-Typen befinden, wohingegen belohnungsvermittelnde Dopamin-Neurone Kompartimente mit den Dendriten der aversiven MBON-Typen innervieren. Diese Struktur impliziert, dass während des olfaktorischen Lernens die kompartimentspezifische Dopaminausschüttung eine Depression (eine Abschwächung der synaptischen Stärke) der synaptischen Verbindungen zwischen KCs und den nachgeschalteten MBONs induziert.
Die ersten Hinweise für lerninduzierte Plastizität pilzkörperextrinsischer Neurone (MBONs) lieferten extrazelluläre Ableitungen der elektrischen Aktivität von MBONs in der Honigbiene (Menzel und Manz, 2005). In Drosophila werden vorrangig Kalzium-Indikatoren benutzt, um neuronale Aktivität in genetisch-identifizierten Populationen von Neuronen zu messen [zur Übersicht siehe (Owald und Waddell, 2015)]. So konnte gezeigt werden, dass sich die duftinduzierte Aktivität bestimmter MBON-Typen für einen gelernten Duft verändert: Wenn ein Geruch mit einer Belohnung assoziiert wird, verringert sich die Erregung der durch diesen Duft aktivierten aversiven MBON-Typen nachhaltig (Owald et al., 2015a). Wird ein Duft jedoch mit einer Bestrafung, zum Beispiel einem Elektroschock, assoziiert, so verringert sich die Reizantwort von positiven MBON-Typen für den erlernten Geruch (Séjourné et al., 2011; Hige et al., 2015; Perisse et al., 2016). Diese Befunde decken sich mit den oben beschriebenen Änderungen des duftinduzierten Verhaltens, wenn aversive oder positive MBON-Typen geblockt werden: Ein initial aversiver Duft induziert eine Hinwendungsreaktion, wenn aversive MBON-Typen geblockt werden, oder Düfte werden noch aversiver, wenn positive MBON-Typen geblockt werden. Zusammen genommen zeigen diese Ergebnisse, dass die synaptische Plastizität zwischen den die Duftidentität repräsentierenden KCs und den valenzkodierenden MBONs die Grundlage des erlernten Verhaltens bildet und somit einen wichtigen Teil des Korrelats zwischen Belohnungs- und Bestrafungsgedächtnis darstellt.
Die Depression von synaptischen Verbindungen scheint das vorherrschende Motiv der lerninduzierten synaptischen Plastizität im Pilzkörper zu sein. Es gibt allerdings Ausnahmen. Manche MBON-Typen (z. B. V2-MBONs, γ2α’1-MBONs) zeigen eine verstärkte Reizantwort auf erlernte Düfte (Bouzaiane et al., 2015; Owald et al., 2015a; Felsenberg et al., 2017) und wieder andere (beispielsweise die sogenannten M4/6 MBONs) zeigen bidirektionale Plastizität: Eine Depression nach belohnendem Lernen sowie eine Potenzierung der Duftantwort auf einen aversiv erlernten Geruch. Interessanterweise scheint die beobachtete Verstärkung im Fall der bidirektionalen Plastizität jedoch nicht allein auf ein lokales synaptisches Phänomen zurückführbar zu sein; vielmehr entsteht die gesteigerte Aktivität nach aversiver Konditionierung aus einem Netzwerkeffekt (Owald et al., 2015a; Perisse et al., 2016). Es konnte gezeigt werden, dass das GABAerge MBON MVP2 eine reduzierte Duftantwort auf einen mit Bestrafung assoziierten Duft zeigt. Dieses Neuron hat nicht nur die Funktion eines positiven MBONs, es hemmt auch die Aktivität von anderen MBON-Typen. Die vorwärtsgerichtete Hemmung ist hierbei nach aversivem Lernen reduziert und führt somit zu einer Enthemmung der aversiven M4/6 MBONs. Es bleibt festzuhalten, dass beide MBON-Typen, aversive M4/6 und positive MVP2 MBONs, Input von den geruchskodierenden KCs erhalten, sodass die Geruchsidentität in diesem Netzwerk-Motiv erhalten bleibt.

Duftgedächtnisse werden als Ungleichgewicht zwischen aversiven und positiven MBON-Typen gespeichert. Die Exposition der Fliege zu einem Duft führt zu einer Aktivierung spezifischer KCs (nicht abgebildet). KCs verschalten mit erregenden Synapsen auf MBONs. Hier werden exemplarisch das positive GABAerge MVP2 MBON (blau, auch bekannt als MBON γ1pedc>α/β) und die aversiven M4/6 MBONs (violett, auch bekannt als MBON β′2mp, MBON β2β′2a, MBON γ5β′2a) dargestellt. a) Dem aktuellen Modell folgend führt die Präsentation eines neutralen Dufts zu einer ausgeglichenen Erregung positiver und aversiver MBONs. Daher ist der Einfluss des Pilzkörpers auf geruchsgesteuertes Verhalten in diesem Fall nicht wirksam. b) Während des Belohnungslernens wird die Präsentation eines Duftes mit einer Belohnung gepaart. Belohnungsvermittelnde dopaminerge Neurone (DANs) innervieren Kompartimente, in denen die KCs auf aversive MBONs verschalten, d. h. während des Belohnungslernens kommt es zu einer gleichzeitigen Erregung von duftaktivierten KCs und der kompartimentspezifischen Ausschüttung von Dopamin. Dieses Zusammentreffen der Signale induziert Plastizität an den Synapsen zwischen aktiven KCs und aversiven MBONs. c) Während des aversiven Lernens wird ein Duft mit einer Bestrafung zeitlich gepaart präsentiert. Die Bestrafung aktiviert dopaminerge Neurone, die Plastizität an aktiven KCs zu positiven MBON-Synapsen induzieren. d–e) Nach dem Training ist die Stärke der Synapsen zwischen den vom erlernten Geruch aktivierten KCs und den entsprechenden MBONs herabgesetzt. Diese Veränderung im Ausgangssignal des MBON-Netzwerks, entweder in Richtung positiver MBONs nach Belohnungslernen oder in Richtung aversiver MBONs nach Bestrafungslernen, führt zur Ausprägung des erlernten Verhaltens.
Insgesamt lassen die diskutierten Resultate auf ein integratives Modell schließen, in dem sich olfaktorische Gedächtnisse als ein Ungleichgewicht zwischen positiven und aversiven MBON-Typen des Pilzkörper-Output-Netzwerks manifestieren [Abbildung 2, (Owald und Waddell, 2015)]. In einem solchen Modell würde ein Geruch ohne erlernte Bedeutung die positiven und aversiven MBON-Typen mit gleicher Intensität anregen. Während des assoziativen Lernens erregt ein Geruch ein bestimmtes KC-Aktivitätsmuster, und der zeitgleich gegebene bedeutungsvolle Reiz (Belohnung oder Bestrafung) aktiviert die entsprechenden Dopamin-Neurone. Die Koinzidenz von Dopamin und der spezifischen KC-Aktivität in einem bestimmten Kompartiment führt dann zu einer Veränderung, meist einer Depression, der Effektivität der lokalen KC-MBON-Synapse. Diese Veränderung der Reizantwort einer bestimmten Gruppe von MBONs verschiebt die Netzwerkaktivität dann entweder nach Belohnungstraining in Richtung der positiven MBONs oder nach dem Bestrafungslernen in Richtung der aversiven MBONs. Das resultierende Ungleichgewicht im MBON-Netzwerk steuert schließlich das duftabhängige Verhalten der Fliege hin zu einer Hinwendungs- oder einer Vermeidungsreaktion.
Interessanterweise suggeriert die Anatomie der MBONs, dass sie nicht nur vom Pilzkörper zu prämotorischen Arealen projizieren, um Verhalten zu induzieren, sondern auch eine Verknüpfung zu den oben diskutierten Dopamin-Neuronen besitzen, die den Pilzkörper innervieren (Aso et al., 2014b; Lewis et al., 2015; Owald et al., 2015a; Eichler et al., 2017). Da assoziatives Lernen das Gleichgewicht des MBON-Netzwerkes umformt, wird also nicht nur das Verhalten, sondern auch die Rückkopplung zum Dopamin-System (Riemensperger et al., 2005) beeinflusst. Diese Feedback-Loops zwischen MBONs und Dopamin-Neuronen scheinen maßgeblich für die Neubewertung erlernter Informationen zu sein (Felsenberg et al., 2017).
Überdenken, was wahr ist
In einer sich ständig verändernden Welt müssen erlernte Informationen kontinuierlich neu bewertet werden, um adäquates Verhalten sicherzustellen. Daher ist einer der entscheidendsten Aspekte von Gedächtnissen, dass sie modifizierbar sind. Wenn ein Tier auf eine Situation trifft, in der die gespeicherte Information nicht mit der aktuellen Ereignislage übereinstimmt (ein Umstand, der als sogenannter Vorhersagefehler formalisiert werden kann), müssen die erlernten Informationen und die entsprechenden Gedächtnisse angepasst werden. Im Allgemeinen lässt sich dies auf zwei Wegen erreichen: Entweder durch die Aktualisierung der gespeicherten Gedächtnisse oder durch die Ausbildung einer neuen opponierenden Gedächtnisspur, einem Extinktionsgedächtnis. Es scheint vom Vorhersagefehler abzuhängen, welcher der beiden Prozesse in Gang gesetzt wird. In Drosophila kann ein Extinktionsgedächtnis ausgebildet werden, indem Fliegen einem zuvor belohnten Geruch erneut ausgesetzt werden, die erwartete Belohnung aber ausbleibt. Solch ein Widerspruch verändert das erlernte Verhalten der Taufliegen; es hebt das erlernte Hinwendungsverhalten auf (Tempel et al., 1983; Felsenberg et al., 2017). Dieses neue Lernen, das Extinktionslernen, bedarf der Aktivität von bestrafungsvermittelnden Dopamin-Neuronen. Die Aktivität dieser Neurone scheint zusätzliche Plastizität zu induzieren, die das entstandene Ungleichgewicht im MBON-Netzwerk nivelliert. Das Ausbleiben von erwarteter Belohnung wird demnach als Bestrafung kodiert und führt zur Entstehung eines parallelen Extinktionsgedächtnisses, das dem initialen Belohnungsgedächtnis opponierend entgegensteht [Abbildung 3; (Felsenberg et al., 2017)].

Die Extinktion eines Belohnungsgedächtnisses wird über das dopaminerge Bestrafungssystem vermittelt. a) MBONs bilden rekurrierende Verschaltungen (Feedback-Loops) mit dopaminergen Neuronen (DAN), die Plastizität während des assoziativen Lernens induzieren: positive MBONs verschalten auf bestrafungsvermittelnde Dopamin-Neurone und aversive MBONs sind mit belohnungsvermittelnden Dopamin-Neuronen verknüpft. b) Nach dem Lernen wird das Ausgangssignal des MBON-Netzwerks für den belohnungsassoziierten Geruch in Richtung der positiven MBONs verschoben, d. h. positive MBONs sind stärker aktiviert als aversive MBONs. Da dieses Ungleichgewicht sich auf die nachgeschalteten Neurone überträgt, kommt es zu einer relativ stärkeren Erregung der bestrafungsvermittelnden Dopamin-Neurone, wenn die Fliege den erlernten Duft ohne die erwartete Belohnung wahrnimmt (Extinktionslernen). c) Nach der Extinktion befindet sich das Verhältnis der Ausgangssignale von positiven und aversiven MBONs wieder im Gleichgewicht. Jedoch beruht dieses Gleichgewicht, im Unterschied zum „naiven“ Zustand, auf der Ausbildung eines parallelen aversiven Gedächtnisses, das die Veränderung nach dem initialen Lernen kompensiert.
Erwartung und Realität können jedoch auch weitestgehend übereinstimmen. Wenn Taufliegen an ein Belohnungsgedächtnis erinnert werden, ohne dass dieser Abruf der grundlegenden Information des Erlernten widerspricht, durchläuft das Gedächtnis einen Zyklus von De- und Restabilisierung. Dieser Rekonsolidierung genannte Vorgang ist ein über das gesamte Tierreich hinweg konservierter Gedächtnisaktualisierungsmechanismus, der geringfügige Anpassungen der erlernten Informationen erlaubt (Nader, 2015).
Verfestigte, langanhaltende Erinnerungen sind gegenüber Störungen unempfindlich; so sind stabile Belohnungsgedächtnisse in der Taufliege resistent gegenüber drastischen Störungen, wie z. B. einer durch Kälte induzierten Betäubung. Wird das Gedächtnis jedoch abgerufen und damit destabilisiert, ist es während eines kritischen Zeitfensters nach dem Abruf anfällig. Das heißt, wird die Drosophila in diesem Zeitraum betäubt und der Zyklus von De- und Restabilisierung damit unterbrochen, so kann der Gedächtnisinhalt dauerhaft gelöscht werden. Circa eine Stunde nach dem Abruf ist das Gedächtnis jedoch restabilisiert, und eine Betäubung der Taufliegen hat keine Auswirkungen mehr auf das erlernte Verhalten. Zur Restabilisierung des Gedächtnisses bedarf es der abgestimmten Aktivität eines spezifischen MBON-Typs sowie der zeitlich koordinierten Aktivität bestimmter dopaminerger Neuronencluster (Felsenberg et al., 2017).
Obwohl die Neubewertung von Gedächtnisinhalten Möglichkeiten bietet, problematische Erinnerungen bei Menschen, wie zum Beispiel extreme Angstgedächtnisse oder mit Drogen assoziierte, den Entzug erschwerende Gedächtnisse, abzuschwächen, sind die zugrunde liegenden Mechanismen noch wenig verstanden. Die wesentlichen Verhaltensphänomene der Extinktion und der Rekonsolidierung sind jedoch über Tiergruppen hinweg, von Wirbellosen zu Wirbeltieren, in hohem Maße ähnlich (Eisenhardt, 2014; Nader, 2015). Daher erscheint es gerechtfertigt anzunehmen, dass die einzigartigen experimentellen Möglichkeiten in Drosophila zu weiteren Erkenntnissen über diese Prozesse führen werden, die generelle Kodierungsprinzipien beschreiben und welche die Forschung zu diesem Thema vorantreiben werden.
Ausblick
Die Erkenntnisse, wie und wo Lernen und Gedächtnisspeicherung im Drosophila-Gehirn realisiert werden, bieten in Kombination mit der genetischen Zugänglichkeit einzelner Neuronenpopulationen eine einmalige Möglichkeit, mechanistische Einblicke in Lernprozesse in vivo zu gewinnen. Die experimentellen Arbeiten der letzten Jahre haben spezifische Synapsen als physische Manifestation des Belohnungs- und Bestrafungsgedächtnisses identifiziert und ermöglichen somit die Untersuchung der molekularen und genetischen Prozesse, die lerninduzierter synaptischer Plastizität zugrunde liegen. Die präzise Kenntnis des Netzwerks sollte erlauben, zu einem Verständnis von Kodierungsprinzipien des Gedächtnisabrufs, der Konsolidierung von Gedächtnissen sowie der Neubewertung von Erinnerungen zu gelangen.
About the authors

Johannes Felsenberg ist Post-Doc in Scott Waddells Labor am Centre for Neural Circuits and Behaviour an der Universität Oxford. Sein Diplom in Biologie und seine Promotion hat er an der Freien Universität Berlin erlangt. In den Laboren von Randolf Menzel und Dorothea Eisenhardt hat er die Rolle des Ubiquitin Proteasome Systems beim Lernen und der Gedächtnisbildung von Honigbienen erforscht. In Oxford arbeitet Johannes an den Mechanismen der Gedächtnis-Rekonsolidierung und der extinktionsinduzierten synaptischen Plastizität im Pilzkörper der Taufliege Drosophila. Sein Projekt im Waddell-Labor ist unterstützt durch ein DFG-Stipendium.

David Owald leitet die Emmy-Noether-Nachwuchsgruppe Behavioral and Physiological Neurogenetics am Institut für Neurophysiologie der Charité in Berlin. Er promovierte an der Universität Göttingen über die molekulare Anatomie und entwicklungsbedingte Plastizität von präsynaptischen aktiven Zonen in Drosophila. Danach war er als Junior Research Fellow für Physiologie am Wolfson College Oxford und als EMBO Long-term und als Sir Henry Wellcome Postdoctoral Fellow am Centre for Neural Circuits and Behaviour an der Universität Oxford tätig. Dort beschäftigte er sich hauptsächlich mit den Mechanismen lernrelevanter Plastizität im Drosophila-Pilzkörper. David erhielt 2017 den Schilling-Forschungspreis der Neurowissenschaftlichen Gesellschaft.
Literatur
Aso, Y. et al. (2014a). Mushroom body output neurons encode valence and guide memory-based action selection in Drosophila. eLife 3, e04580.10.7554/eLife.04580.039Search in Google Scholar
Aso, Y., Hattori, D., Yu, Y., Johnston, R. M., Iyer, N. A., Ngo, T.-T., Dionne, H., Abbott, L. F., Axel, R., Tanimoto, H., Rubin, G. M. and Griffith, L. C. (2014b). The neuronal architecture of the mushroom body provides a logic for associative learning. eLife 3, e04577.10.7554/eLife.04577Search in Google Scholar PubMed PubMed Central
Barnstedt, O., Owald, D., Felsenberg, J., Brain, R., Moszynski, J.-P., Talbot, C. B., Perrat, P. N. and Waddell, S. (2016). Memory-Relevant Mushroom Body Output Synapses Are Cholinergic. Neuron 89, 1237–1247.10.1016/j.neuron.2016.02.015Search in Google Scholar PubMed PubMed Central
Bouzaiane, E., Trannoy, S., Scheunemann, L., Plaçais, P.-Y. and Preat, T. (2015). Two independent mushroom body output circuits retrieve the six discrete components of Drosophila aversive memory. Cell Rep. 11, 1280–1292.10.1016/j.celrep.2015.04.044Search in Google Scholar PubMed
Brand, A. H. and Perrimon, N. (1993). Targeted gene expression as a means of altering cell fates and generating dominant phenotypes. Development 118, 401–415.10.1242/dev.118.2.401Search in Google Scholar PubMed
Burke, C. J., Huetteroth, W., Owald, D., Perisse, E., Krashes, M. J., Das, G., Gohl, D., Silies, M., Certel, S. and Waddell, S. (2012). Layered reward signalling through octopamine and dopamine in Drosophila. Nature 492, 433–437.10.1038/nature11614Search in Google Scholar PubMed PubMed Central
Eichler, K., Li, F., Litwin-Kumar, A., Park, Y., Andrade, I., Schneider-Mizell, C. M., Saumweber, T., Huser, A., Eschbach, C., Gerber, B., Fetter, R. D., Truman, J. W., Priebe, C. E., Abbott, L. F., Thum, A. S., Zlatic, M. and Cardona, A. (2017). The complete connectome of a learning and memory centre in an insect brain. Nature 548, 175–182.10.1038/nature23455Search in Google Scholar PubMed PubMed Central
Eisenhardt, D. (2014). Molecular mechanisms underlying formation of long-term reward memories and extinction memories in the honeybee ( Apis mellifera). Learn Mem. 21, 534–542.10.1101/lm.033118.113Search in Google Scholar PubMed PubMed Central
Felsenberg, J., Barnstedt, O., Cognigni, P., Lin, S. and Waddell, S. (2017). Re-evaluation of learned information in Drosophila. Nature 544, 240–244.10.1038/nature21716Search in Google Scholar PubMed PubMed Central
Heisenberg, M. (2003). Mushroom body memoir: from maps to models. Nat. Rev. Neurosci. 4, 266–275.10.1038/nrn1074Search in Google Scholar PubMed
Hige, T., Aso, Y., Modi, M. N., Rubin, G. M., Turner, G. C. (2015). Heterosynaptic Plasticity Underlies Aversive Olfactory Learning in Drosophila. Neuron 88, 985–998.10.1016/j.neuron.2015.11.003Search in Google Scholar PubMed PubMed Central
Huetteroth, W., Perisse, E., Lin, S., Klappenbach, M., Burke, C. and Waddell, S. (2015). Sweet taste and nutrient value subdivide rewarding dopaminergic neurons in Drosophila. Curr. Biol. 25, 751–758.10.1016/j.cub.2015.01.036Search in Google Scholar PubMed PubMed Central
Konopka, R. J. and Benzer, S. (1971). Clock mutants of Drosophila melanogaster. PNAS 68, 2112–2116.10.1073/pnas.68.9.2112Search in Google Scholar PubMed PubMed Central
Lewis, L. P. C., Siju, K. P., Aso, Y., Friedrich, A. B., Bulteel, A. J. B., Rubin, G. M. and Grunwald Kadow, I. C. (2015). A Higher Brain Circuit for Immediate Integration of Conflicting Sensory Information in Drosophila. Curr. Biol. 25, 2203–2214.10.1016/j.cub.2015.07.015Search in Google Scholar PubMed
Lin, S., Owald, D., Chandra, V., Talbot, C., Huetteroth, W. and Waddell, S. (2014). Neural correlates of water reward in thirsty Drosophila. Nat. Neurosci. 17, 1536–1542.10.1038/nn.3827Search in Google Scholar PubMed PubMed Central
Liu, C., Plaçais, P.-Y., Yamagata, N., Pfeiffer, B. D., Aso, Y., Friedrich, A. B., Siwanowicz, I., Rubin, G. M., Preat, T. and Tanimoto, H. (2012). A subset of dopamine neurons signals reward for odour memory in Drosophila. Nature 488, 512–516.10.1038/nature11304Search in Google Scholar PubMed
Menzel, R. (2014). The insect mushroom body, an experience-dependent recoding device. J. Physiol. (Paris) 108, 84–95.10.1016/j.jphysparis.2014.07.004Search in Google Scholar PubMed
Menzel, R. and Manz, G. (2005). Neural plasticity of mushroom body-extrinsic neurons in the honeybee brain. J. Exp. Biol. 208, 4317–4332.10.1242/jeb.01908Search in Google Scholar PubMed
Nader, K. (2015). Reconsolidation and the Dynamic Nature of Memory. Cold Spring Harbor Perspect. Biol. 7, a021782.10.1101/cshperspect.a021782Search in Google Scholar PubMed PubMed Central
Owald, D., Felsenberg, J., Talbot, C. B., Das, G., Perisse, E., Huetteroth, W. and Waddell, S. (2015a). Activity of defined mushroom body output neurons underlies learned olfactory behavior in Drosophila. Neuron 86, 417–427.10.1016/j.neuron.2015.03.025Search in Google Scholar PubMed PubMed Central
Owald, D., Lin, S. and Waddell, S. (2015b). Light, heat, action: neural control of fruit fly behaviour. Philos. Trans. R. Soc., B 370, 20140211.10.1098/rstb.2014.0211Search in Google Scholar PubMed PubMed Central
Owald, D. and Waddell, S. (2015). Olfactory learning skews mushroom body output pathways to steer behavioral choice in Drosophila. Curr. Opin. Neurobiol. 35, 178–184.10.1016/j.conb.2015.10.002Search in Google Scholar PubMed PubMed Central
Perisse, E., Owald, D., Barnstedt, O., Talbot, C. B., Huetteroth, W. and Waddell, S. (2016). Aversive Learning and Appetitive Motivation Toggle Feed-Forward Inhibition in the Drosophila Mushroom Body. Neuron 90, 1086–1099.10.1016/j.neuron.2016.04.034Search in Google Scholar PubMed PubMed Central
Quinn, W. G. and Dudai, Y. (1976). Memory phases in Drosophila. Nature 262, 576–577.10.1038/262576a0Search in Google Scholar PubMed
Riemensperger, T., Völler, T., Stock, P., Buchner, E. and Fiala, A. (2005). Punishment prediction by dopaminergic neurons in Drosophila. Curr. Biol. 15, 1953–1960.10.1016/j.cub.2005.09.042Search in Google Scholar PubMed
Rubin, G. M. and Spradling, A. C. (1982). Genetic transformation of Drosophila with transposable element vectors. Science 218, 348–353.10.1126/science.6289436Search in Google Scholar PubMed
Séjourné, J., Plaçais, P.-Y., Aso, Y., Siwanowicz, I., Trannoy, S., Thoma, V., Tedjakumala, S. R., Rubin, G. M., Tchénio, P., Ito, K., Isabel, G., Tanimoto, H. and Preat, T. (2011). Mushroom body efferent neurons responsible for aversive olfactory memory retrieval in Drosophila. Nat. Neurosci. 14, 903–910.10.1038/nn.2846Search in Google Scholar PubMed
Stevens, C. F. (2015). What the fly“s nose tells the fly”s brain. Proc. Natl. Acad. Sci. USA. 112, 9460–9465.10.1073/pnas.1510103112Search in Google Scholar PubMed PubMed Central
Südhof, T. C. (2012). The presynaptic active zone. Neuron 75, 11–25.10.1016/j.neuron.2012.06.012Search in Google Scholar PubMed PubMed Central
Tanaka, N. K., Tanimoto, H. and Ito, K. (2008). Neuronal assemblies of the Drosophila mushroom body. J. Comp. Neurol. 508, 711–755.10.1002/cne.21692Search in Google Scholar PubMed
Tempel, B. L., Bonini, N., Dawson, D. R. and Quinn, W. G. (1983). Reward learning in normal and mutant Drosophila. Proc. Natl. Acad. Sci. USA. 80, 1482–1486.10.1073/pnas.80.5.1482Search in Google Scholar PubMed PubMed Central
Tovote, P., Fadok, J. P. and Lüthi, A. (2015). Neuronal circuits for fear and anxiety. Nat. Rev. Neurosci. 16, 317–331.10.1038/nrn3945Search in Google Scholar PubMed
Waddell, S. (2013). Reinforcement signalling in Drosophila; dopamine does it all after all. Curr. Opin. Neurobiol. 23, 324–329.10.1016/j.conb.2013.01.005Search in Google Scholar PubMed PubMed Central
Yamagata, N., Ichinose, T., Aso, Y., Plaçais, P.-Y., Friedrich, A. B., Sima, R. J., Preat, T., Rubin, G. M. and Tanimoto, H. (2015). Distinct dopamine neurons mediate reward signals for short- and long-term memories. Proc. Natl. Acad. Sci. USA. 112, 578–583.10.1073/pnas.1421930112Search in Google Scholar PubMed PubMed Central
Danksagungen
Wir danken Kerstin Germer für die Übersetzung. JF (FE 1563/1-1) und DO (OW 84/2-1) werden von der DFG gefördert.
Anmerkung
Englische Version des Artikels online verfügbar unter https://doi.org/10.1515/nf-2017-A048
© 2018 by De Gruyter
Articles in the same Issue
- Frontmatter
- Übersichtsartikel
- Gedächtnismechanismen in Drosophila
- Making Memories. On the fly.
- Bewegungssehen: Zellen, Schaltkreise und Algorithmen
- Motion detection: cells, circuits and algorithms
- Molekulare Dynamik der neuronalen Informationsübertragung
- Molecular dynamics of neuronal information transfer
- Die Rolle von Histonmethylierungen in der Entwicklung des zentralen Nervensystems und bei Neuralrohrdefekten
- Histone methylations in the developing central nervous system and in neural tube defects
- „Mind from Matter?“ – Über Verhalten und Gehirn
- Mind from Matter? – Via Brain and Behavior
- Forschungsförderung
- Sonderforschungsbereich (SFB 1280) „Extinktionslernen“
- Kollaboratives Forschungszentrum (Sonderforschungsbereich, SFB) 1286 „Quantitative Synaptologie“
- Nachruf
- Nachruf auf Prof. Dr. Dr. h. c. mult. Franz Huber
- Nachruf auf Robert F. Schmidt
- Rezension
- Norman Doidge: Neustart im Kopf – Wie sich unser Gehirn selbst repariert
- Nachrichten
- Einladung zur Mitgliederversammlung während des FENS Forum 2018 in Berlin (7.–11. Juli 2018)
Articles in the same Issue
- Frontmatter
- Übersichtsartikel
- Gedächtnismechanismen in Drosophila
- Making Memories. On the fly.
- Bewegungssehen: Zellen, Schaltkreise und Algorithmen
- Motion detection: cells, circuits and algorithms
- Molekulare Dynamik der neuronalen Informationsübertragung
- Molecular dynamics of neuronal information transfer
- Die Rolle von Histonmethylierungen in der Entwicklung des zentralen Nervensystems und bei Neuralrohrdefekten
- Histone methylations in the developing central nervous system and in neural tube defects
- „Mind from Matter?“ – Über Verhalten und Gehirn
- Mind from Matter? – Via Brain and Behavior
- Forschungsförderung
- Sonderforschungsbereich (SFB 1280) „Extinktionslernen“
- Kollaboratives Forschungszentrum (Sonderforschungsbereich, SFB) 1286 „Quantitative Synaptologie“
- Nachruf
- Nachruf auf Prof. Dr. Dr. h. c. mult. Franz Huber
- Nachruf auf Robert F. Schmidt
- Rezension
- Norman Doidge: Neustart im Kopf – Wie sich unser Gehirn selbst repariert
- Nachrichten
- Einladung zur Mitgliederversammlung während des FENS Forum 2018 in Berlin (7.–11. Juli 2018)