Molekulare Dynamik der neuronalen Informationsübertragung
-
Martin Heine
Martin Heine studierte Biologie an der Friedrich-Schiller-Universität Jena. In seiner Doktorarbeit an der Georg-August-Universität Göttingen untersuchte er im Labor von Prof. D.W. Richter die Verteilung von Serotonin-Rezeptoren und deren Einfluss auf die dendritische Signalverarbeitung in CA1 – Neuronen des Hippocampus. Nach dem Abschluss der Doktorarbeit 2002 wechselte er in die Gruppe von Prof. D. Choquet an der Universität Bordeaux (Frankreich). Hier untersuchte Martin Heine die funktionellen Konsequenzen der Oberflächendynamik von AMPA-Rezeptoren in der neuronalen Membran. Ende 2007 begann Martin Heine in der Abteilung für Neurochemie von Prof. E. Gundelfinger am Leibniz-Institut für Neurobiologie in Magdeburg zu arbeiten. Hier ist er seit 2009 Leiter der Forschungsgruppe „Molekulare Physiologie“. Das Hauptinteresse der Arbeitsgruppe liegt in der Erforschung dynamischer Interaktionen von Adhäsionsproteinen und Ionenkanälen in der neuronalen Membran. Dabei wird vordergründig die funktionelle Bedeutung der lateralen Mobilität dieser Moleküle in Synapsen und neuronalen Netzwerken untersucht.
 and
Arthur Bikbaev
and
Arthur Bikbaev
Arthur Bikbaev studierte Biologie an der Bashkir State Universität in Ufa (Rußland). In seiner Doktorarbeit an der Bashkir State Universität Ufa und der Lomonosov-Universität in Moskau untersuchte er die pathogenen Mechanismen von Schläfenlappen- und Absenzenepilepsie. Nach der Verteidigung seiner Doktorarbeit, wechselte er in die Gruppe von Prof. D. Manahan-Vaughan an die Ruhr-Universität Bochum. Hier untersuchte er die Wirkung hippocampaler synaptischer Plastizität auf Netzwerk-Oszillationen
in vivo . Arthur begann 2009 in der Gruppe von Dr. M. Heine, die Entwicklung und Konnektivität in neuronalen Netzwerkenin vitro zu untersuchen. Hier arbeitet er vorrangig an der Analyse der Informationsverarbeitung in neuronalen Netzwerken.

Zusammenfassung
Die detaillierte Analyse von Synapsen als verbindende Elemente zwischen Nervenzellen ist von zentraler Bedeutung, um sowohl die kognitiven Leistungen des Gehirns als auch deren Einschränkungen zu verstehen. Modernste optische Verfahren erlauben es heute, einzelne Moleküle in lebenden Zellen zu detektieren. Insbesondere kann so die Dynamik der molekularen Komposition in kleinsten Kompartimenten wie etwa den prä- und postynaptischen Membranen erforscht werden. Die Beobachtung der Verteilung von Rezeptoren, Ionenkanälen und Adhäsionsmolekülen über die Zeit lässt erkennen, dass diese einer ständigen stochastischen Bewegung unterliegen. Dies ist überraschend, sind doch Synapsen gerade für solche Proteinmoleküle als Akkumulationsorte mit vielen molekularen Anknüpfungspunkten beschrieben. Durch gezielte Beeinflussung der lateralen Bewegung von Glutamatrezeptoren in Verbindung mit klassischer Elektrophysiologie konnte gezeigt werden, dass die molekulare Dynamik entscheidend für die Induktion synaptischer Plastizität ist und ihrerseits durch synaptische Aktivität beeinflusst werden kann. Sie sollte daher auch mit Blick auf übergeordnete Hirnfunktionen und neurologische Erkrankungen näher untersucht werden.
Einleitung
Die Fähigkeit eines Gehirns, aus multiplen und sich ständig ändernden sensorischen Reizen biologisch sinnvolle Informationen für das einzelne Lebewesen zu generieren, ist für uns besonders faszinierend. Einerseits können sich Erfahrungen und erlernte Fähigkeiten über lange Zeit entwickeln und erhalten, andererseits sind oftmals schnelle impulsive Reaktionen auf Umgebungsreize nötig. Dieses breite Spektrum an Reaktionsfähigkeit wird der Funktion neuronaler Netzwerke zugeschrieben, in denen die Erregbarkeit und die synaptischen Verknüpfungen der Nervenzellen von zentraler Bedeutung sind. Nervenzellen sind, unterstützt durch Gliazellen, die zellulären Bausteine neuronaler Netzwerke. Typischerweise empfangen sie dabei Signale von zahlreichen anderen Nervenzellen über synaptische Eingänge, die auf den Zellkörpern, vor allem aber auf verzweigten Fortsätzen, den sogenannten Dendriten, lokalisert sind. So kann beispielsweise eine einzelne Pyramidenzelle in der CA1-Region des Hippocampus einer Ratte ca. 30.000 glutamaterge (erregende) und zusätzlich etwa 1.700 GABAerge (hemmende) synaptische Eingänge entlang dendritischer Verzweigungen mit einer Gesamtlänge von ca. 12.000 µm aufweisen (Megias et al., 2001). Die sinnvolle Integration der eintreffenden Signale stellt somit eine enorme Herausforderung dar. Ein wesentlicher Parameter ist dabei die Stärke einer jeden Synapse sowie ihre Lokalisation auf der Zelle. Hemmende Synapsen sind vorzugsweise an proximalen Dendriten, den Zellkörper und den Axonhügel anzutreffen, während erregende Synapsen sich sowohl am proximalen als auch distalen Dendriten befinden (Gulyas et al., 1999; Megias et al., 2001).
Dendritische Strukturen können die Integration synaptischer Information über verschiedene Mechanismen beeinflussen. Die elektrischen Eigenschaften dendritischer Membranen und die unterschiedliche Expression von spannungsabhängigen Ionenkanälen entlang von Dendriten können den Beitrag der einzelnen Synapse verstärken oder schwächen (Tran-Van-Minh et al., 2015). Entscheidend für den Informationsaustausch in neuronalen Netzwerken ist jedoch die Stärke und Aktivierungsfrequenz der Synapsen. Die kurzzeitige, wiederholte Aktivierung von Synapsen kann zur Bahnung (engl.: facilitation) oder Depression führen und abhängig von der Stimulationsdauer längerfristig die Effizienz der synaptischen Übertragung verändern. Diese plastischen Prozesse können in wenigen Millisekunden bis Sekunden ablaufen und dabei die Synapsen für Minuten bis Stunden verändern. Die Mechanismen, die die Transmissionseigenschaften einer Synapse kurzfristig verändern, sind oft präsynaptischen Ursprungs und können durch die Akkumulation von präsynaptischem Kalzium, die Änderung der Anzahl freisetzungsbereiter Vesikel oder durch retrograde Signale ausgelöst werden. Jedoch sind auch postsynaptische Mechanismen wie die Desensitisierung oder Sättigung von Rezeptoren beteiligt (Zucker und Regehr, 2002). Diese schnelle Plastizität von Synapsen ist die entscheidende Variable, um Aktivitätsänderungen über die Zeit umzusetzen und kurzzeitig (innerhalb von Sekunden-Minuten) zu erinnern (Abbott und Regehr, 2004). Länger anhaltende Veränderungen der synaptischen Aktivität sind hingegen häufig durch postsynaptische Mechanismen dominiert, können aber auch durch eine anhaltende Änderung der präsynaptischen Transmitterfreisetzung getragen werden. Oft gehen solche langanhaltenden Änderungen mit strukturellen Veränderungen von Synapsen einher und werden weithin als zelluläres Korrelat für Lernen und Gedächtnisbildung betrachtet. Die Übergänge zwischen kurzzeitigen Änderungen und anhaltender Verstellung der synaptischen Aktivität sind indes fließend.
Früh wurde beobachtet, dass die Richtung der kurzzeitigen Plastizität, ob Bahnung oder Depression, von der postsynaptischen Zelle mitbestimmt ist. Das heißt, die zeitliche Integration der synaptischen Aktivität scheint vorgegeben. Dies wurde erstmals an Motorneuronen von Krebsen gezeigt, bei denen die Synapsen derselben Nervenfaser eine unterschiedliche Kurzzeitplastizität an verschiedenen Muskelfasern zeigen (Atwood und Bittner, 1971). Ähnliche Beobachtungen wurden später auch in primären neuronalen Zellkulturen und im Gehirn von Säugern gemacht (Bao et al., 2010; Branco et al., 2008; Markram et al., 1998; Pouille und Scanziani, 2004), wo Endigungen derselben Pyramidenzelle verschiedene Populationen von Neuronen kontaktieren und abhängig von der postsynaptischen Zelle eine spezifische Kurzzeitplastizität zeigen. Die Lokalisation und molekulare Zusammensetzung der Synapse sind entscheidend für ihre Funktion. Lange wurden diese beiden Parameter als sehr stabile Elemente neuronaler Netzwerke wahrgenommen. Allein die kinetischen Eigenschaften von Ionenkanälen und ionotropen Rezeptoren, die Spezifität von Adhäsionsmolekülen sowie die Modulation der Synapse durch metabotrope Rezeptoren ergeben eine Fülle von Möglichkeiten, die Plastizität von Synapsen zu beschreiben. Die Beobachtung, dass Synapsen ständig auf und abgebaut werden (Attardo et al., 2015; Engert und Bonhoeffer, 1999) und vom Austausch (Recycling) membranständiger Rezeptoren abhängig sind (Kerchner und Nicoll, 2008; Kittler und Moss, 2003), waren entscheidende Impulse, zusätzlich die Dynamik molekularer Mechanismen für die synaptische Plastizität und Integrität neuronaler Netzwerke zu erkennen. Vor allem die Manipulation dynamischer Prozesse in der postsynaptischen Membran glutamaterger Synapsen hat gezeigt, dass die lokale Endo-/Exozytose von Glutamatrezeptoren des AMPA-Typs sowie ihre laterale Verteilung und Diffusion in der Membran die Plastizität der Synapse wesentlich bestimmen (Carroll et al., 1999; Heine et al., 2008; Luscher et al., 1999; Penn et al., 2017). Im Folgenden wird die Oberflächendiffusion von synaptischen Signalmolekülen, deren Schlüsselfunktion für die Plastizität der Synapse unlängst gezeigt wurde, näher erläutert.
Methoden zur Messung molekularer Mobilität in der Membran
Die Aktivität und Verteilung einzelner Moleküle kann mittels elektrophysiologischer und optischer Methoden dargestellt werden. Die Voraussetzung hierfür ist die Kenntnis kinetischer Parameter, im Falle der Nutzung der Elektrophysiologie, oder die spezifische Markierung des Moleküls mit optisch detektierbaren Partikeln. Während die elektrophysiologischen Methoden die kinetischen Eigenschaften von Ionenkanälen und ionotropen Rezeptoren nutzen und somit auf elektrisch aktive Proteine beschränkt sind, können optische Methoden auf nahezu alle Moleküle angewendet werden.
Elektrophysiologisch sind mobile Rezeptorpopulationen mithilfe spezifischer Pharmaka darstellbar. Diese als funktionelles Markieren (functional tagging) beschriebene Methode wurde erstmalig zur Analyse des lokalen Austausches von synaptischen NMDA-Rezeptoren (NMDAR) in glutamatergen Synapsen genutzt (Tovar und Westbrook, 1999). Durch evozierte oder spontane Aktivität werden synaptische NMDAR irreversibel mit dem Antagonisten MK-801 inaktiviert. Die lokale synaptische Freisetzung von Glutamat führt somit zur Inaktivierung synaptischer NMDAR. Schnelles Auswaschen von ungebundenem MK-801 verhindert die weitere Inaktivierung von neu in die Synapse einwandernden NMDAR. In der Folge kann die Erholung NMDAR-vermittelter synaptischer Ströme als Austausch von inaktiven gegen aktive Rezeptoren interpretiert werden. Solche Experimente ergaben, dass schon innerhalb weniger Minuten wieder eine NMDAR-vermittelte synaptische Antwort messbar ist. Die kurzfristige Erholung lässt schließen, dass der laterale Austausch membranständiger NMDAR hier der vorherrschende Mechanismus ist, da ein Austausch synaptischer NMDAR über die Exozytose intrazellulärer Rezeptoren wesentlich langsamer erfolgt (>1h). Dennoch können auch neu in die Membran inserierte Rezeptoren teilweise beteiligt sein. Ähnliche Strategien wurden für spezifische Populationen von AMPA-Rezeptoren (AMPAR) und GABAA-Rezeptoren entwickelt (Thomas und Smart, 2006).
Optische Methoden hingegen ermöglichen es, sowohl Populationen als auch einzelne Moleküle zu beobachten. Voraussetzung ist, dass die Moleküle über ein optisch sichtbares Teilchen markiert werden können (optical tagging). Oft sind dies fluoreszierende Farbstoffe, die durch die Absorption von Licht und die Emission einer spezifischen Fluoreszenz nachweisbar sind. Es gibt jedoch auch die Möglichkeit, z. B. mithilfe von kleinen Goldpartikeln, andere optische Methoden zu nutzen. Hier wird die Änderung der Absorption des Lichts oder die Änderung des lokalen Brechungsindex als Messgröße verwendet (Celebrano et al., 2011; Lasne et al., 2006; Ritchie und Kusumi, 2003).
Der erste Nachweis der Mobilität synaptischer Rezeptoren wurde mit FRAP-Experimenten (fluorescence recovery after photobleach, FRAP, Fluoreszenzrückkehr nach Photoausbleichung) erbracht. Hier wird die Rückkehr der Fluoreszenz nach dem lokalen Ausbleichen der Fluoreszenz-markierten Rezeptoren über die Zeit beobachtet. Der Zeitverlauf der Erholung der Fluoreszenzintensität in der ausgeblichenen Region gibt eine direkte Abschätzung der mobilen Fraktion von Molekülen und kann zur Analyse der Diffusionseigenschaften genutzt werden. Mithilfe von Fluoreszenz-markiertem α-Bungarotoxin war es z. B. möglich, die lokale Diffusion von Acetylcholin-Rezeptoren in der postsynaptischen Membran neuromuskulärer Synapsen der Kaulquappe des Krallenfroschs spezifisch zu bestimmen (Young und Poo, 1983). Basierend auf der Entwicklung von genetisch kodierbaren pH-sensitiven Fluorophoren (z. B. pHluorin) können Oberflächenmoleküle in der Membran direkt dargestellt werden. Die pH-Abhängigkeit der Fluorophore erlaubt, membranständige von intrazellulär, in Vesikeln lokalisierten Rezeptoren, zu unterscheiden, was die spezifische Analyse der Rezeptormobilität in der Membran wesentlich vereinfacht. Generell birgt FRAP in Kombination mit Transgen-kodierten, Fluoreszenzprotein-markierten Rezeptoren aber die Gefahr, dass letztere in Konkurrenz zur endogenen Rezeptorpopulation treten. In Folge dessen übersteigt die anhand der markierten Moleküle ausgelesene Dynamik oftmals die tatsächlich anzunehmende Mobilität der endogenen Rezeptorpopulation ohne zusätzliche Expression.
Die direkte Beobachtung einzelner Moleküle in der neuronalen Membran ist demgegenüber sehr viel informativer. Individuelle Bewegungen membranständiger Moleküle geben uns Einblick in deren Verweildauer und Dichteverteilung. Besonders interessant werden solche Experimente, wenn die Lokalisation individueller Moleküle wichtig für ihre Funktion ist. Dies lässt sich z. B. an der Freisetzung eines Transmittervesikels illustrieren. Hier ist die Aktivierung weniger Kalziumkanäle, im Extremfall eines einzigen Kalziumkanals ausreichend, um ein Transmittervesikel in der näheren Umgebung (20–50 nm) desselben freizusetzen (Eggermann et al., 2012). Bezogen auf die räumliche Dimension der meisten Synapsen, die mit einem Durchmesser der aktiven Zone bzw. postsynaptischen Dichte von ca. 300–500 nm angegeben wird, könnten diffuse molekulare Bewegungen durchaus relevant sein, um die Übertragungseigenschaften einer Synapse zu beeinflussen. Für typische Diffusionskonstanten membranständiger Proteine (0.005–0.5 µm²/s) bedeutet das, dass eine Verschiebung von ca. 10–100 nm in 10 ms möglich ist. Die Zahl spannungsabhängiger Kalziumkanäle, Adhäsionsmoleküle und Rezeptoren ist für kortikale Synapsen (z. B. hippocampale CA3-CA1 Synapse) mit 10–150 Molekülen angegeben. Die Frage ist nun, inwieweit die Dichte der Moleküle deren Bewegung beeinträchtigt.
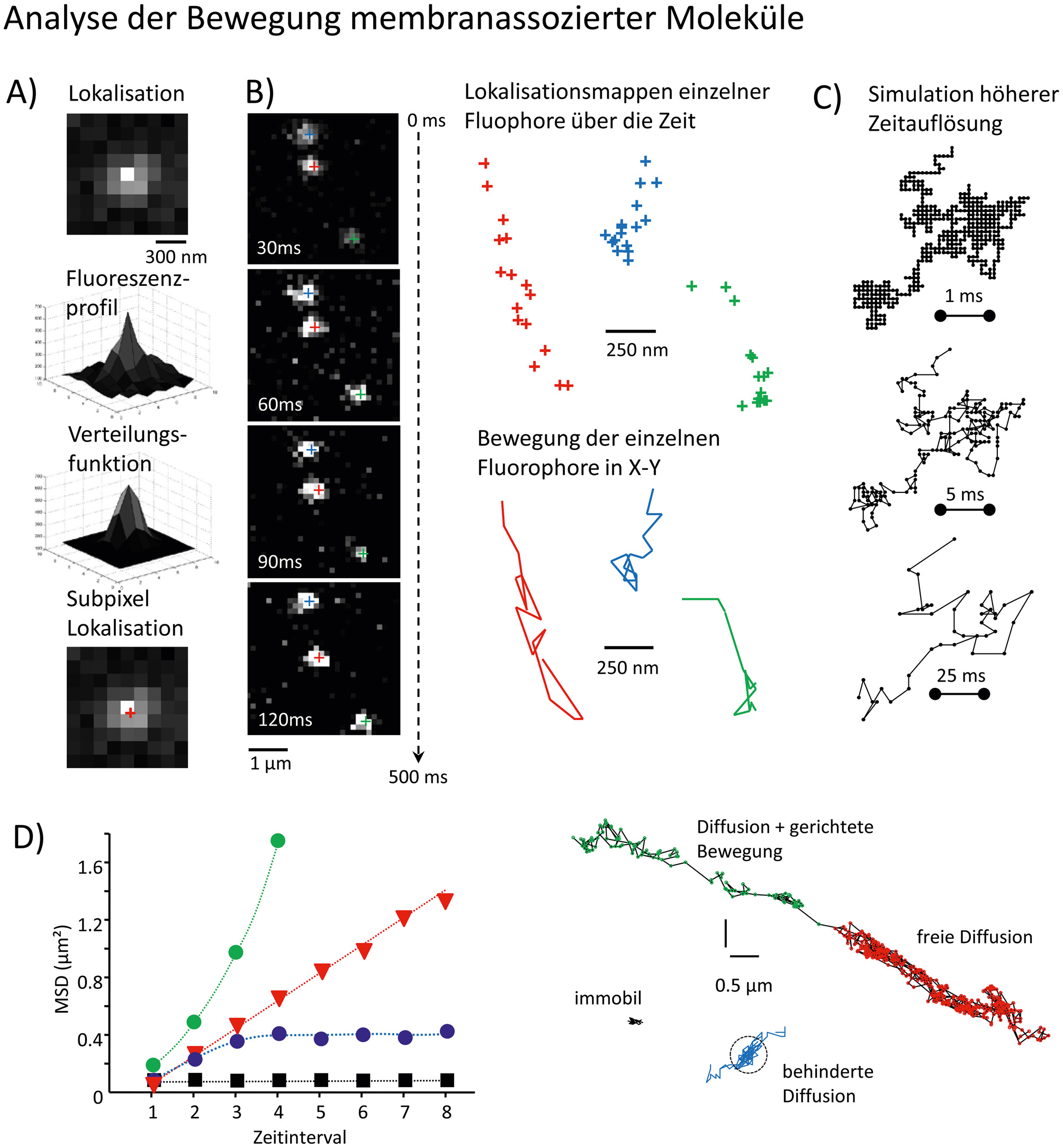
Analyse der Mobilität von Molekülen in der zellulären Membran mittels Single Particle Tracking (SPT). A) Dargestellt sind die einzelnen Schritte für die Lokalisation eines Fluorophores im Bild der Kamera. Die Aufteilung der detektierten Lichtintensitäten auf mehrere Pixel der Kamera erlaubt die Lokalisation des Fluorophores innerhalb eines Pixels. B) Die zeitliche Abfolge der Lokalisation identifizierbarer Fluorophore erlaubt die Erstellung von Lokalisationskarten und die Verbindung der einzelnen Positionen über die Zeit. Diese Trajektorien können dann für die Analyse der Diffusion und explorierten Oberfläche verwendet werden. C) Simulation einer höheren Aufnahmerate für die Diffusion eines Partikels in der Membran. Deutlich ist die wesentlich bessere Repräsentation der explorierten Oberfläche bei Aufnahmeraten ≥1 kHz. Die Parameter für die Simulation waren Beobachtungsdauer 1s, Diffusionskoeffizient 0.2 µm²/s bei Bildraten von 1.000, 200 und 25 Bildern pro Sekunde. D) Quantifizierung verschiedener Bewegungsarten mittels der mittleren quadratischen Verschiebung (MSD) über die Zeit. Die Abweichung von freier Brown´scher Diffusionsbewegung (gerade Linie) können mittels dieser Darstellung eindeutig identifiziert werden. Dargestellt sind freie Diffusion (schwarz), begrenzte Diffusion (rot), Diffusion mit gerichteter Bewegung (grün) und immobile Teilchen (schwarz). Die Fläche unter der Kurve stellt die explorierte Oberfläche zum gegebenen Zeitpunkt dar. Die Abweichung von y=0 für immobile Teilchen entspricht der Lokalisationsgenauigkeit des Experiments. Beispiele für Trajektorien unterschiedlicher Diffusionsbewegung sind in der entsprechenden Farbe dargestellt.
Diese lokalen Verteilungen und Bewegungen in Membrankompartimenten wie der Synapse darzustellen verlangt die direkte Detektion einzelner Moleküle über die Zeit. Die Emissionseigenschaften der Fluorophore, die Größe, Affinität und Bindungsvalenz der Liganden (z. B. primäre Antikörper, Nanobodies etc.) sowie die Markierungsdichte sind hier entscheidende Parameter. Neben der Fluoreszenzkorrelationsspektroskopie ist die kamerabasierte Detektion einzelner Moleküle über die Zeit (single particle tracking, SPT) eine inzwischen häufig verwendete Methode. Das lokale Fluoreszenzmaximum (Centroid) erlaubt es, die Lokalisation eines Fluorophors innerhalb von 10–40 nm zu bestimmen (Abbildung 1A, B). Nach der Bestimmung individueller Positionen pro Bild werden sie über die Zeit (Bildrate) miteinander verbunden, um die Bewegungsbahnen (Trajektorien) jedes einzelnen Moleküls zu generieren (Abbildung 1B). Die Bildrate zur Darstellung der Diffusionsbewegungen ist abhängig von der Menge der emitierten Photonen pro Zeiteinheit, die die Lokalisationsgenauigkeit des einzelnen Fluorophores ermitteln. Beim Arbeiten mit lebenden Zellen muss man hier einen Kompromiss zwischen Beleuchtungsstärke, Phototoxizität für die Zelle und der Messgenauigkeit finden. Daher sind die explorierten Oberflächen einzelner Trajektorien oft nur eine grobe Abschätzung der Wirklichkeit (Abbildung 1C). Die Analyse dieser Bewegungsbahnen mittels Berechnung der mittleren quadratischen Verschiebung (mean square displacment, MSD) erlaubt es, die verschiedenen Bewegungsarten (freie Diffusion, beschränkte Diffusion, gerichtete Bewegung etc.), die explorierte Oberfläche und den Diffusionskoeffizienten zu bestimmen (Abbildung 1D). Abhängig vom genutzten Fluorophor können Moleküle für Sekunden (einzelne Fluoreszenzfarbstoffe) bis hin zu mehreren Minuten (Quantum Dots) verfolgt werden. Damit können Änderungen des Diffusionsverhaltens in Korrelation zur Position in der Membran direkt verfolgt werden (Abbildung 2). Individuelle molekulare Interaktionen sind oft sehr kurzlebig und durch die limitierte zeitliche Auflösung nicht direkt zu beobachten. Die Verwendung von statistischen Analysemethoden erlaubt es, sich auf die Verschiebung der Moleküle zwischen zwei Zeitpunkten zu beziehen und so unabhängig von der Länge der individuellen Beobachtungszeit die mittlere Verweildauer und Packungsdichte der Moleküle innerhalb kleiner Membranoberflächen zu bestimmen (Hoze et al., 2012; Renner et al., 2017).
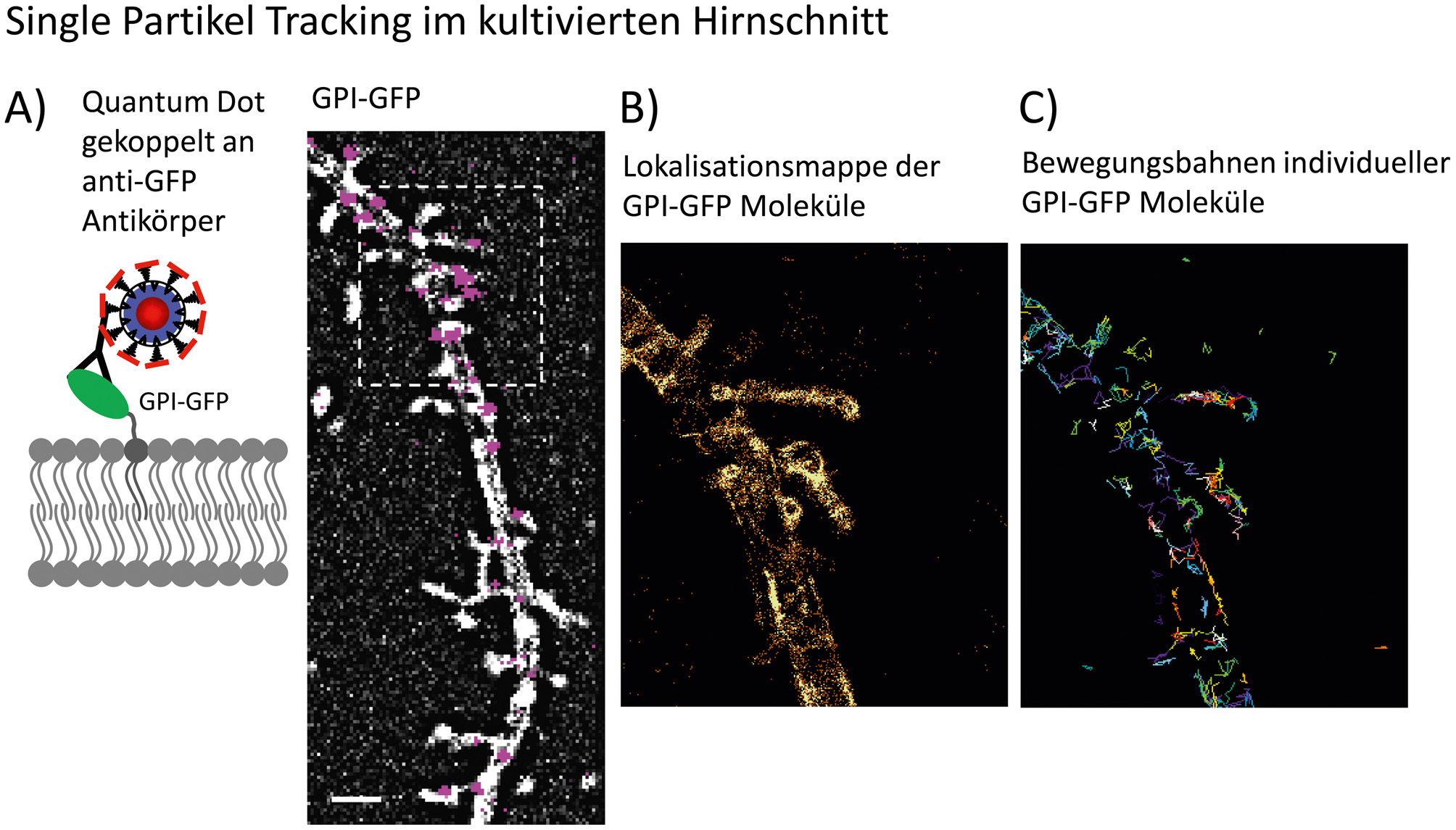
Single Particle Tracking in kultivierten Hirnschnitten. A) Schematische Darstellung eines lipidverankerten GFPs, welches über einen monoclonalen anti-GFP – Antikörper mit einem Quantum Dot (QD) markiert ist. Quantum Dots sind Einzelphotonen – Emitter, die durch ihre lang anhaltende Fluoreszenz und dem stochastischen Blinken den Vorteil bieten, sie als einzelne Fluorophore zu erkennen. Des Weiteren haben QDs ein sehr breites Anregungsspektrum. Diese Eigenschaften ermöglichen es, einzelne QDs zweifelsfrei zu detektieren. B) Im Bild sind die Positionen einzelner Qantum Dots auf den mit GPI-GFP transfizierten Dendritensegment eine hippocampalen Nervenzelle gezeigt. Die Verfolgung individueller QDs über die Zeit ermöglicht die Darstellung der äußeren Membran, basierend auf der Lokalisation der QDs über die Zeit (hier 1.000 Bilder über 30 s). C) Die Rekonstruktion der Bewegungsbahn ist oft durch das Blinken der QDs oder die Überschneidung der Lokalisation benachbarter Partikel unterbrochen, wobei ein Teil der Lokalisationen nicht für die Rekonstruktion der Trajektorien verwendet werden kann. Für weitere Informationen siehe auch (Biermann et al., 2014).
Die Zellmembran als dynamisches Kompartiment
Die ladungstrennenden Eigenschaften der Zellmembran zusammen mit der spezifischen Verteilung von Ionenkanälen sind entscheidend für die Aktivität von Nervenzellen. Andererseits ist die in der Membran gespeicherte thermische Energie immer Ursache für die Brown‘sche Molekularbewegung von Lipiden und membranenständigen Proteinen. Die Aufrechterhaltung der lokalen Dichte von Ionenkanälen, z. B. innerhalb der synaptischen Membran oder im Axonhügel, ist abhängig von der Verankerung oder bestehenden Diffusionsbarrieren innerhalb der Membran. Dem entsprechend ist die Diffusionsbewegung individueller Moleküle in der Zellmembran sehr heterogen und durch direkte und indirekte Interaktionen mit benachbarten Molekülen beeinflusst. Hier sind laterale Interaktionen innerhalb der Membran sowie vertikale Interaktionen mit assoziierten Strukturen an den Grenzflächen der Membran dominant. Die laterale Bewegungsfreiheit innerhalb der Zellmembran wird durch die Dichte integraler Proteinkomplexe und die lokale Komposition der membranbildenden Lipide bestimmt. Ein beeindruckendes Beispiel für die inhomogene Zusammensetzung der Membran ist der Axonhügel oder das axonale Initialsegment. Dieses Membransegment ist mit seiner hohen Dichte an Ionenkanälen Initiationsstelle des Aktionspotenzials und Diffusionsbarriere zwischen somato-dendritischer und axonaler Membran (Nakada et al., 2003; Winckler et al., 1999). Das in allen tubulären Fortsätzen von Neuronen vorhandene reguläre Zytoskelett (Xu et al., 2013), aufgebaut aus Aktinringen, die über das Linkerprotein Adducin mit längs verlaufenden Spektrinfilamenten verbunden sind, ist im Initialsegment Ankerstruktur für Gerüstproteine (AnkyrinG) und damit Aggregationspunkt für Ionenkanäle. Die tubuläre Geometrie der Membran von Neuriten hat neben der Komplexität der Membranzusammensetzung auch Einfluss auf die Diffusionsbewegung der Moleküle (Domanov et al., 2011). Je dünner die Zellfortsätze werden, umso restriktiver ist die Diffusionsbewegung, was vor allem für dünne postsynaptische Dornenfortsätze (Spines) von Bedeutung ist. Weitere Mechanismen für die unterschiedliche Verteilung von Membranproteinen sind die lokale Endo/Exozytose, lokale transmembranöse Proteincluster, Interaktionen mit der extrazellulären Matrix und elektrostatische Interaktionen (Trimble und Grinstein, 2015). Die Bewegung von transmembranösen Proteinen kann also durch die Struktur der Membran selbst und die an sie angelagerten Strukturen stark beeinflusst werden.
Dynamik postsynaptischer Signalproteine
Für die Effizienz der Informationsübertragung ist die dynamische Verteilung postsynaptischer Rezeptoren in hemmenden und erregenden Synapsen gleichermaßen als wichtig beschrieben worden. Die Bewegung der Rezeptoren in glutamatergen Synapsen ist hierbei am weitesten erforscht, weshalb sie nun näher behandelt werden soll.
Beginnend mit der Freisetzung von Glutamat in den synaptischen Spalt, ist die Diffusion von Glutamat im extrazellulären Raum entscheidend für die Signalwirkung des Transmitters. Die Freisetzung eines Glutamatvesikels verliert innerhalb von ca. 100 µs seine Signalfunktion. Ursache ist die geringe Affinität von AMPAR zu Glutamat. Abhängig von der Zusammensetzung der AMPAR liegt der EC50-Wert zwischen 0,5–2 mM. Das heißt, dass nur innerhalb eines Radius von ca. 100 nm AMPAR während der Freisetzung eines Glutamatvesikels (2.000–5.000 Glutamatmoleküle) aktiviert werden können (Diamond und Jahr, 1997). Die Anordnung, Dichte und Zusammensetzung der postsynaptischen AMPAR bestimmt die Amplitude der postsynaptischen Stromantwort. In Bezug auf die hier im Mittelpunkt stehende laterale Bewegung von Molekülen in der Membran stellen sich folgende Fragen: Wie viele Rezeptoren werden durch die Transmitterfreisetzung eines Vesikels aktiviert? Wie stabil sind Rezeptoren in der Membran verankert? Wie kann die lokale Dichte von Rezeptoren in der Synapse verändert werden?
Elektronenmikroskopische Bilder von synaptischen Strukturen vermitteln den Eindruck, dass der stochastischen Bewegung von postsynaptischen Signalproteinen massiv Einhalt geboten wird. Dafür spricht auch ihre durch zahlreiche Studien belegte Vernetzung mit membranassoziierten Gerüstproteinen und dem Zytoskelet. Die intensive Erforschung des Transports und der lateralen Dynamik von AMPAR hat aber die Fluktuation von Rezeptoren und assoziierter Signalmoleküle als zentrale Mechanismen synaptischer Plastizität aufgedeckt. Die Analyse des lateralen Diffusionsverhaltens von Rezeptoren und Lipiden zeigt, dass innerhalb der Synapse membranständige Moleküle zwischen freien und behinderten Diffusionsverhalten wechseln. Im Falle der Rezeptoren reflektiert dieses „Stop and Go“ potenziell eine wechselnde Bindung an zum Beispiel intrazelluläre Gerüstmoleküle. AMPAR werden hauptsächlich über das Gerüstprotein PSD-95 in der Synapse akkumuliert, während an inhibitorischen GABAergen und glycinergen Synapsen Gephyrin die Rolle des zentralen Gerüstproteins für die betreffenden Rezeptoren erfüllt. Ursprünglich waren C-terminale Bindungsmotive einzelner AMPAR – Untereinheiten als Basis für eine direkte Interaktion mit PSD-95 diskutiert worden (Shi et al., 2001). Jedoch sind diese Interaktionen der Rezeptor – Untereinheiten mit PSD-95 zu schwach, um somit die synaptische Stabilisierung von AMPAR während der Induktion von Langzeitplastizität zu erklären (Kim et al., 2005). Vielmehr zeigte sich, dass die synaptische Verankerung der Rezeptoren auch über weitere Bindungspartner realisiert werden kann. Insbesondere mit der Entdeckung von Rezeptor-assoziierten Transmembranproteinen wie den TARPs (transmembrane AMPA receptor regulatory proteins) oder CKAMP44 (cysteine-Knoten AMPAR modulierendes Protein 44), Shisa und Cornichon konnten hier wesentliche Einsichten gewonnen werden. Diese Moleküle sind für den intrazellulären Transport der AMPAR zur Plasmamembran notwendig und vermitteln zudem eine weitere Bindung der Rezeptoren an Gerüstproteine. Geschuldet der Halbwertszeit eines jeden Rezeptors oder der mit ihm assoziierten Moleküle ist die Stabilisierung in der Synapse nur von kurzer Dauer. Somit ist die transiente Interaktion von Rezeptoren und assoziierten Proteinen wesentlicher Bestandteil der Funktion und Plastizität von Synapsen. Die Beobachtung einzelner Rezeptoren in der aktiven Synapse ermöglicht es, die zeitliche Kodierung der Plastizität als eine Abfolge molekularer Interaktionen näher zu verstehen. Am Beispiel verschiedener Studien zur Funktion der TARPs sollen diese flexiblen Interaktionen für verschiedene Zeiteinheiten illustriert werden.
Stargazin (TARPγ-2), das zunächst als akzessorische γ-Untereinheit von spannungsabhängigen Kalziumkanälen beschrieben wurde, ist essenziell für den Transport und die synaptische Expression von AMPAR in glutamatergen Synapsen des Kleinhirns (Stargazer, dt. „Sterngucker“, ist der Name der entsprechenden spontanen Mausmutante). TARPγ-2 ist nicht nur essenziell für die Oberflächenexpression und die kinetischen Eigenschaften von AMPAR, sondern es vermittelt auch die Assoziation von AMPAR mit dem Gerüstprotein PSD-95 (Chen et al., 2000). Mit der Beobachtung individueller AMPAR und TARPγ-2 konnte nachgewiesen werden, dass die Assoziation von TARPγ-2 zu PSD-95 für die Stabilität und Akkumulation von AMPAR wesentlich ist. Diese Experimente zeigten auch, dass sich die Mehrheit der Rezeptoren sowohl innerhalb als auch außerhalb der Synapse in Assoziation mit TARPγ-2 befindet (Bats et al., 2007). Eine Aktivitätsänderung der Synapse kann zur C-terminalen Phosphorylierung von TARPγ-2 führen und somit die Bindungseigenschaften zu PSD-95 verändern (Opazo et al., 2010). Demnach kann in einem Zeitfenster von Sekunden bis wenigen Minuten die synaptische Akkumulation der AMPAR verändert werden, da ein Verlust (Mobilisierung) oder Gewinn (Immobilisierung) von Bindungen an intrazelluläre Gerüstproteine stets in Konkurrenz zur vorhandenen thermischen Energie der Membran steht.
Ein besonders interessanter Modus, über den die Assoziation bzw. Dissoziation von AMPAR und TARPγ-2 gleichzeitig an die Dynamik von AMPAR und deren funktionellen Zustand gekoppelt wird, wurde kürzlich vorgeschlagen. Demnach ist die Assoziation von AMPAR mit TARPγ-2 entscheidend für die postsynaptische Sensitivität für Glutamat. Die Aktivierung eines AMPAR durch Bindung von Glutamat führt stets zu einer strukturellen Änderung der extrazellulären Domänen des Rezeptors, welche die Öffnung des Ionenkanals induziert. Diese Glutamat-abhängige Konformationsänderung ist jedoch nur von kurzer Dauer und wird von einer weiteren Konformationsänderung gefolgt, welche zur Desensitisierung des Rezeptors führt. In diesem Zustand dissoziieren AMPAR und TARPγ-2 und die Rezeptoren verlieren die TARPγ-2-vermittelte Verankerung an das Gerüstprotein PSD-95. Erst wenn sich Glutamat vom Rezeptor wieder löst, kann dieser in den geschlossenen, aber aktivierbaren Zustand zurückkehren und erneut mit TARPγ-2 assoziieren. Eine wichtige Implikation dieses Zyklus ist, dass der kurzzeitige Verlust der Bindung an PSD-95 den Austausch von desensitisierten gegen aktivierbare AMPAR in der Synapse potenziell begünstigt. Allerdings kann die Packungsdichte von benachbarten integralen Proteinen in der Membran, auch als „molecular crowding“ bekannt, dem Wegdiffundieren desensitisierter Rezeptoren aus der Synapse entgegenwirken. Somit initiiert also die präsynaptische Freisetzung von Glutamat eine postsynaptische Rezeptorverschiebung innerhalb weniger Nanometer (Constals et al., 2015).
Diese zwei Beispiele zeigen, dass über indirekte Interaktionen des Rezeptors mit Gerüstproteinen eine wesentlich feinere Steuerung der Rezeptorverteilung möglich ist. Durch proteomische Analyse von AMPAR – Komplexen konnten weitere Bindungspartner identifiziert werden, die in unterschiedlichen Kombinationen mit AMPAR assoziiert sind (Schwenk et al., 2012; Schwenk et al., 2009). Die differenzielle Verteilung von AMPAR – Untereinheiten und assoziierter Proteine in verschiedenen Hirnregionen vermittelt ein sehr viel komplexeres Bild von der molekularen Konfiguration glutamaterger Synapsen. Funktionell ist die laterale Mobilität von AMPAR sowohl für kurzzeitige als auch für längerfristige Veränderungen der Synapse relevant.
Oberflächendiffusion von AMPAR als Variable synaptischer Plastizität
Die beispielhaft dargestellten Mechanismen der dynamischen Verteilung von AMPAR führen zu der Frage, inwieweit stochastische Veränderungen der Rezeptorpopulation die Plastizität der Synapse beeinflussen können. Postsynaptische Variablen der Kurzzeitplastizität sind die kinetischen Eigenschaften der AMPA-Rezeptoren, die durch die Assoziation mit akzessorischen Proteinen beeinflusst werden. In Synapsen mit hoher Freisetzungswahrscheinlichkeit sind AMPAR teilweise durch vorherige Stimulationen desensitisiert und tragen somit zur Verringerung der postsynaptischen Antwort bei. Eine artifizielle Immobilisierung der AMPAR während der wiederholten Benutzung der Synapse führt sogar zu einer noch stärkeren Depression der Synapse (Heine et al., 2008). Dies bedeutet, dass der laterale Austausch von Rezeptoren in der postsynaptischen Membran einen Teil der Depression kompensiert, indem aktivierbare gegen desensitisierte Rezeptoren ausgetauscht werden (Abbildung 3A, B). Die beschriebene Dissoziation zwischen Rezeptor und TARPγ-2 könnte hier einen molekularen Mechanismus darstellen, wie über die schnelle Änderung von intrazellulären Bindungsaffinitäten der laterale Austausch der Rezeptoren induziert werden kann (siehe oben). Andere mit AMPAR assoziierte Proteine haben unterschiedlichen Einfluss auf die kinetischen Eigenschaften der Rezeptoren (von Engelhardt et al., 2010). Die Dynamik innerhalb der postsynaptischen Membran muss daher immer in Reflektion mit der molekularen Zusammensetzung betrachtet werden, welche sehr variabel in verschiedenen Hirnregionen ist (Schwenk et al., 2014).
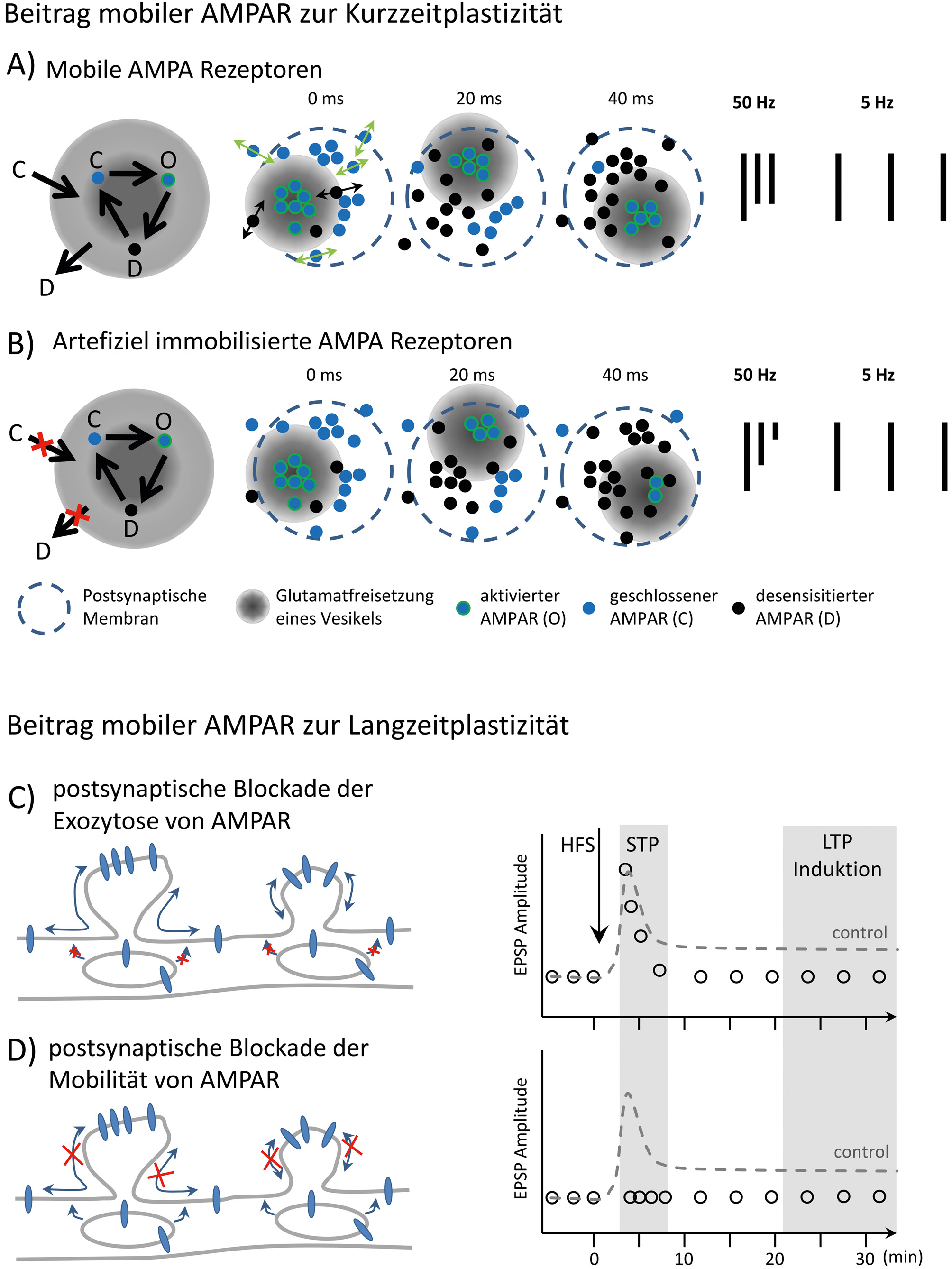
Schematische Darstellung des Einflusses mobiler AMPAR auf die synaptische Plastizität. A) Dargestellt sind die kinetischen Zustände von AMPAR, wobei der desensitisierte Zustand der zeitlimitierende ist. Die Verweildauer im desensitisierten Zustand kann je nach Rezeptorzusammensetzung variieren und die Aktivierbarkeit der postsynaptischen AMPAR – Population beeinflussen. Die einzelnen Zustände sind: der geschlossene Rezeptor (C), der geöffnete Rezeptor nach Bindung von Glutamat (O) und der desensitisierte Rezeptor (D). Die lokale Dynamik der Rezeptoren erlaubt eine teilweise Kompensation der Akkumulation desensitisierter Rezeptoren während hochfrequenter Stimulation der Synapse. B) Die artifizielle Immobilisierung der AMPAR führt zu einer stärkeren Akkumulation von desensitisierten Rezeptoren in der postsynaptischen Membran, welche bei hochfrequenten Stimulationen der Synapse zur Verstärkung der Depression beiträgt, hier schematisch gezeigt für 50 Hz. Stimulationen im Frequenzbereich <5 Hz sind nicht von der Immobilisierung der AMPAR abhängig (modifiziert nach Heine et al. 2008). C) Schematische Darstellung des Beitrags der Mobilität von AMPAR zur Langzeitpotenzierung in CA3-CA1 – Synapsen des Hippocampus. Durch die postsynaptische Blockade der Exocytose von AMPAR mittels Tetanus Toxin wird die Induktion der LTP verhindert, jedoch nicht die kurzzeitige posttetanische Potenzierung. D) Die globale Immobilisierung der AMPAR verhindert sowohl die posttetanische Potenzierung als auch die Induktion von LTP (modifiziert nach Penn et al. 2017).
Die Langzeitpotenzierung der synaptischen Transmission (LTP), postuliert als Prozess der Gedächtnisbildung, kann initial entweder durch eine Änderung der Freisetzungswahrscheinlichkeit von Transmitter-Vesikeln und/oder eine postsynaptische Änderung der Anzahl, Leitfähigkeit und kinetischen Eigenschaften der Rezeptoren hervorgerufen werden. Die Veränderung der Anzahl von AMPAR ist vor allem über die Rekrutierung von Rezeptoren in die Synapse beschrieben worden. Bislang wurde hauptsächlich die lokale Exozytose intrazellulärer AMPAR in die postsynaptische Membran als molekularer Mechanismus aufgezeigt. Vorstellbar ist aber auch eine Veränderung der lokalen Anzahl von Rezeptoren durch die Rekrutierung von perisynaptischen AMPAR, die bereits in der Plasmamembran vorhanden sind. Extrasynaptische AMPAR sind mittels iontophoretischer oder photolytischer Applikation von Glutamat funktionell detektierbar (Liu et al., 1999). Mit der Markierung einzelner AMPAR konnte die extrasynaptische Population als eine sehr mobile Population von Rezeptoren nachgewiesen werden (Heine et al., 2008). Durch die direkte Manipulation des Rezeptorrecyclings (Endo-/ Exozytose) einerseits und der Verhinderung der lateralen Mobilität der AMPAR über Biotin-Avidin-vermittelte Vernetzung der Oberflächenpopulation andererseits, ist es gelungen, die zeitliche Abfolge der Rekrutierung von Rezeptoren in die Synapse darzustellen. Eine anhaltende Immobilisierung von AMPAR in der Zellmembran führt hier zur vollständigen Blockade der Induktion von LTP. Die Verhinderung der Exozytose von AMPAR durch die Perfusion der postsynaptischen Zelle mit Tetanus-Toxin hat zwar keine Wirkung auf die transiente posttetanische Potenzierung, verhindert aber die Induktion von LTP (Abbildung 3C, D). Weitere Variationen dieser zwei Experimente in verschiedenen experimentellen Systemen (kultivierter Hirnschnitt, akuter Hirnschnitt, in vivo) bestätigten, dass die laterale Mobilität der AMPAR essenziell für die Induktion von LTP ist. Tatsächlich konnte zudem durch die Verhinderung der lateralen Mobilität von AMPAR in vivo eine Blockade des Hippocampus-abhängigen Angstlernens in der initialen Phase erzielt werden (Penn et al., 2017). Somit ist die schnelle Umverteilung von Rezeptoren in der Zellmembran offensichtlich ein notwendiger Prozess, um auf akute Änderungen synaptischer Aktivität zu reagieren und ein „mobiles Gedächtnis“ der Aktivität zu bewahren. Die Konfiguration der Rezeptorkomplexe aus verschiedenen Interaktionspartnern kann dieses „mobile Gedächtnis“ entscheidend beeinflussen.
Oberflächendiffusion präsynaptischer Signalproteine
Anwendungen von lokalisationsmikroskopischen Methoden haben gezeigt, dass sich auch präsynaptische Signalmoleküle, wie spannungsabhängige Kalziumkanäle, SNARE-Proteine und Adhäsionsmoleküle diffus in der Membran bewegen (Bademosi et al., 2017; Fu und Huang, 2010; Neupert et al., 2015; Schneider et al., 2015). Welche Funktion dieser Dynamik für die Freisetzung von synaptischen Vesikeln zukommt, ist noch nicht gut erforscht. Es lässt sich aber spekulieren, dass die geringe Dichte von z. B. Kalziumkanälen in präsynaptischen Membranen eher für eine dynamische Lokalisation dieser Kanäle spricht. Ausschlaggebend für diese Interpretation ist die vielfach demonstrierte Notwendigkeit, dass Vesikel und Kalziumkanal in sehr enger Nachbarschaft zueinander stehen müssen, damit es zur Freisetzung fusionsbereiter Transmittervesikel kommt (<100 nm (Eggermann et al., 2012)). Die eher geringe Affinität vesikulärer Kalziumsensoren für freies intrazelluläres Kalzium, die durch das Aktionspotenzial definierte kurze Öffnungszeit des Kalziumkanals und die Kontrolle des intrazellulären Kalziums durch kalziumbindende Proteine definieren die räumlichen Dimensionen (Eggermann et al., 2012). Mobile Kanäle erlauben es, die räumliche Nähe von Kanal und Vesikel vorübergehend zu erreichen. Der zu Beginn erwähnte Einfluss der Postsynapse auf die Freisetzungswahrscheinlichkeit der Präsynapse ist Hinweis auf unterschiedliche Komplexe von Adhäsionsmolekülen, die an der lokalen Organisation präsynaptischer Kalziumkanäle (Missler et al., 2003; Sylwestrak und Ghosh, 2012) teilhaben. Eine weitere Stellschraube, um die Wahrscheinlichkeit der „transienten Kopplung“ zwischen Kanal und Vesikel zu erhöhen, ist die direkte oder indirekte Interaktion der Kalziumkanäle mit Gerüstproteinen der aktiven Zone. Direkte Protein-Protein-Interaktionen des C-Terminus von Kalziumkanälen mit RIM und RIM Binding Protein (RBP) haben massiven Einfluss auf die Akkumulation und Positionierung von Kalziumkanälen (Acuna et al., 2016; Kaeser et al., 2011). Interaktionen dieser Proteine mit anderen Gerüstproteinen in der Zytomatrix der Präsynapse (Bassoon, Munc13, ELKS) sind ebenfalls beteiligt an der Positionierung und Rekrutierung der Kanäle (Biederer et al., 2017).
Die Beschreibung der lokalen Dichte von prä- (RIM) und postsynaptischen (PSD-95) Gerüstproteinen mit lokalisationsmikroskopischen Methoden (STORM/PALM) schlägt die Existenz von sogenannten Nanosäulen vor, die das Zusammenspiel von präsynaptischer Vesikelfreisetzung und postsynaptischer Rezeptordichte optimieren (Tang et al., 2016). Diese Nanosäulen können als transmembranale Hindernisse und intrazelluläre Dockstation für diffundierende Kalziumkanäle dienen und so deren Aktionsradius definieren. Andererseits erlaubt die Beweglichkeit der Kanäle (Schneider et al. 2015), dass es individuelle Zusammentreffen von Kanal und fusionsbereitem Vesikel gibt. Die Zeitspanne dieser Zusammentreffen sollte direkten Einfluss auf die Anzahl von freizusetzenden Vesikeln haben. Die direkte Beobachtung von Vesikelfusionen innerhalb einer Präsynapse über die Zeit zeigte, dass die Lokalisation der Freisetzungsorte zwischen zwei Zeitpunkten sich immer ändert und so die Idee einer transienten Kopplung von Kanal und Vesikel unterstützt (Maschi und Klyachko, 2017; Tang et al., 2016).
Ein zusätzlicher Effekt einer transienten Kopplung von Kanal und Vesikel ist die geringe Anzahl von Kalziumkanälen in der präsynaptischen Membran. Induziert durch das Ruhemembranpotenzial sind ca. 50% der Kalziumkanäle inaktiviert, also nicht verfügbar. Eine stete Umordnung der Kanäle in der präsynaptischen Membran kann diesen Effekt teilweise kompensieren.
Ausblick
In den letzten Jahrzehnten wurde eine Vielfalt von synaptischen Proteinen beschrieben, die die große strukturelle Komplexität der Synapse bestimmen. Mit der Entwicklung neuer Mikroskopietechniken und molekularer Hilfsmittel wird immer deutlicher, dass die Zusammensetzung von Synapsen sehr heterogen ist. Die stochastische Dynamik von membranständigen Proteinen ist in der Synapse von besonderer Bedeutung, da hier viele verschiedene Proteine miteinander interagieren und aktivitätsbedingt variable Bindungsaffinitäten zueinander haben. Damit ist auf kleinsten Raum eine hohe Flexibilität möglich. Diese veränderliche Organisation der Synapse über die Zeit ist in der Etablierung und Reifung von Synapsen und während der Induktion von Plastizität wichtig. Bezüglich der neuesten Arbeiten über die Funktion mobiler AMPAR kann man davon ausgehen, dass Änderungen der Oberflächendynamik von Rezeptoren als „molekulares Gedächtnis“ vorausgegangener synaptischer Aktivität angesehen werden können. Obwohl es nur für kurze Momente gespeichert wird, ist dies eine zeitliche Repräsentation vorausgegangener Aktivitätsmuster. Die Möglichkeit, in jedem Moment innerhalb einer sehr heterogenen Umgebung molekulare Konfigurationen und damit Synapsen zu ändern, kann den initialen Schritt für die Etablierung von Plastizität darstellen. Die weitere Erforschung der molekularen Dynamik von Synapsen wird helfen, die Plastizität in neuronalen Netzwerken besser zu verstehen. Ein weiterer Aspekt dieser Forschung liegt darin, die spezifische Expression von Molekülen in Hirnregionen zu nutzen, um lokale neurologische Störungen zu behandeln. Dies wurde kürzlich für das akzessorische AMPAR Protein TARPγ-8 vorgeschlagen (Maher et al., 2017).
Abkürzungen
AMPAR α-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid (AMPA) receptors
Cornichon CNIHs sind kleine, drei transmembranale Domainen besitzende Proteine und binden direkt an AMPAR
CKAMP44 Cysteine-Knoten AMPAR modulierendes Protein 44, wird der Shisa-Proteinfamilie zugerechnet
ELKS Protein reich an den Aminosäuren E, L, K and S
GPI-GFP GGlycosylphosphatidylinositol – gekoppeltes grün fluoreszierendes Protein, genutzt als Reporter der Beweglichkeit von Lipiden in der äußeren Lipidschicht von Membranen
LTP Langzeitpotenzierung
Munc13 Primingfaktor für die Fusion von synaptische Vesikeln
NMDAR N-methyl-D-aspartate receptor
PDZ PSD-95/discs large/zonula occludens-1
PSD-95 postsynaptic density protein 95, which has three PDZ-domains and a MAGUK-domain (membrane-associated guanylate kinase)
RIM RAB interacting molecule
RIM binding protein RAB interacting molecule binding protein
SNARE soluble N-ethylmaleimide-sensitive-factor attachment receptor
Shisa single-transmembrane proteins is characterized by an N-terminal cysteine-rich domain and a proline-rich C-terminal region
TARP transmembrane AMPA receptor regulatory proteins, zu Typ I TARPs gehören die Untereinheiten γ2 (Stargazin), γ3, γ4, and γ8, zu den Typ II TARPs die Untereinheiten γ5 and γ7
About the authors

Martin Heine studierte Biologie an der Friedrich-Schiller-Universität Jena. In seiner Doktorarbeit an der Georg-August-Universität Göttingen untersuchte er im Labor von Prof. D.W. Richter die Verteilung von Serotonin-Rezeptoren und deren Einfluss auf die dendritische Signalverarbeitung in CA1 – Neuronen des Hippocampus. Nach dem Abschluss der Doktorarbeit 2002 wechselte er in die Gruppe von Prof. D. Choquet an der Universität Bordeaux (Frankreich). Hier untersuchte Martin Heine die funktionellen Konsequenzen der Oberflächendynamik von AMPA-Rezeptoren in der neuronalen Membran. Ende 2007 begann Martin Heine in der Abteilung für Neurochemie von Prof. E. Gundelfinger am Leibniz-Institut für Neurobiologie in Magdeburg zu arbeiten. Hier ist er seit 2009 Leiter der Forschungsgruppe „Molekulare Physiologie“. Das Hauptinteresse der Arbeitsgruppe liegt in der Erforschung dynamischer Interaktionen von Adhäsionsproteinen und Ionenkanälen in der neuronalen Membran. Dabei wird vordergründig die funktionelle Bedeutung der lateralen Mobilität dieser Moleküle in Synapsen und neuronalen Netzwerken untersucht.

Arthur Bikbaev studierte Biologie an der Bashkir State Universität in Ufa (Rußland). In seiner Doktorarbeit an der Bashkir State Universität Ufa und der Lomonosov-Universität in Moskau untersuchte er die pathogenen Mechanismen von Schläfenlappen- und Absenzenepilepsie. Nach der Verteidigung seiner Doktorarbeit, wechselte er in die Gruppe von Prof. D. Manahan-Vaughan an die Ruhr-Universität Bochum. Hier untersuchte er die Wirkung hippocampaler synaptischer Plastizität auf Netzwerk-Oszillationen in vivo. Arthur begann 2009 in der Gruppe von Dr. M. Heine, die Entwicklung und Konnektivität in neuronalen Netzwerken in vitro zu untersuchen. Hier arbeitet er vorrangig an der Analyse der Informationsverarbeitung in neuronalen Netzwerken.
Danksagung
Wir möchten uns für die Korrektur und Kritik am Manuskript bei Dr. Ulrich Thomas, Jennifer Heck und Oliver Klatt bedanken. Des Weiteren bedanken wir uns bei Dr. A. Voigt für die Monte-Carlo-Simulationen des Diffusionsverhaltens einzelner Moleküle in der Membran. Beim Land Sachsen – Anhalt (Förderung durch EFRE-Mittel), dem CBBS, dem Leibniz-Institut für Neurobiologie (LIN) Magdeburg sowie der Schram-Stiftung danken wir für die finanzielle Unterstützung unserer Arbeit.
Literatur
Abbott, L. F. and Regehr, W. G. (2004). Synaptic computation. Nature 431, 796–803.10.1038/nature03010Search in Google Scholar PubMed
Acuna, C., Liu, X. and Sudhof, T. C. (2016). How to Make an Active Zone: Unexpected Universal Functional Redundancy between RIMs and RIM-BPs. Neuron 91, 792–807.10.1016/j.neuron.2016.07.042Search in Google Scholar PubMed
Attardo, A., Fitzgerald, J. E. and Schnitzer, M. J. (2015). Impermanence of dendritic spines in live adult CA1 hippocampus. Nature 523, 592–596.10.1038/nature14467Search in Google Scholar PubMed PubMed Central
Atwood, H. L. and Bittner, G. D. (1971). Matching of excitatory and inhibitory inputs to crustacean muscle fibers. J. Neurophysiol. 34, 157–170.10.1152/jn.1971.34.1.157Search in Google Scholar PubMed
Bademosi, A. T., Lauwers, E., Padmanabhan, P., Odierna, L., Chai, Y. J., Papadopulos, A., Goodhill, G. J., Verstreken, P., van Swinderen, B. and Meunier, F. A. (2017). In vivo single-molecule imaging of syntaxin1A reveals polyphosphoinositide- and activity-dependent trapping in presynaptic nanoclusters. Nat. Commun. 8, 13660.10.1038/ncomms13660Search in Google Scholar PubMed PubMed Central
Bao, J., Reim, K. and Sakaba, T. (2010). Target-dependent feedforward inhibition mediated by short-term synaptic plasticity in the cerebellum. J. Neurosci. 30, 8171–8179.10.1523/JNEUROSCI.0276-10.2010Search in Google Scholar PubMed PubMed Central
Bats, C., Groc, L. and Choquet, D. (2007). The interaction between Stargazin and PSD-95 regulates AMPA receptor surface trafficking. Neuron 53, 719–734.10.1016/j.neuron.2007.01.030Search in Google Scholar PubMed
Biederer, T., Kaeser, P. S. and Blanpied, T. A. (2017). Transcellular Nanoalignment of Synaptic Function. Neuron 96, 680–696.10.1016/j.neuron.2017.10.006Search in Google Scholar PubMed PubMed Central
Biermann, B., Sokoll, S., Klueva, J., Missler, M., Wiegert, J. S., Sibarita, J. B. and Heine, M. (2014). Imaging of molecular surface dynamics in brain slices using single-particle tracking. Nat. Commun. 5, 3024.10.1038/ncomms4024Search in Google Scholar PubMed PubMed Central
Branco, T., Staras, K., Darcy, K. J. and Goda, Y. (2008). Local dendritic activity sets release probability at hippocampal synapses. Neuron 59, 475–485.10.1016/j.neuron.2008.07.006Search in Google Scholar PubMed PubMed Central
Carroll, R. C., Beattie, E. C., Xia, H., Luscher, C., Altschuler, Y., Nicoll, R. A., Malenka, R. C. and von Zastrow, M. (1999). Dynamin-dependent endocytosis of ionotropic glutamate receptors. Proc. Natl. Acad. Sci. U.S.A. 96, 14112–14117.10.1073/pnas.96.24.14112Search in Google Scholar PubMed PubMed Central
Celebrano, M., Kukura, P., Renn, A. and Sandoghdar, V. (2011). Single-molecule imaging by optical absorption. Nat. Photonics 5, 95.10.1038/nphoton.2010.290Search in Google Scholar
Chen, L., Chetkovich, D. M., Petralia, R. S., Sweeney, N. T., Kawasaki, Y., Wenthold, R. J., Bredt, D. S. and Nicoll, R. A. (2000). Stargazin regulates synaptic targeting of AMPA receptors by two distinct mechanisms. Nature 408, 936–943.10.1038/35050030Search in Google Scholar PubMed
Constals, A., Penn, A. C., Compans, B., Toulme, E., Phillipat, A., Marais, S., Retailleau, N., Hafner, A. S., Coussen, F., Hosy, E. et al. (2015). Glutamate-Induced AMPA Receptor Desensitization Increases Their Mobility and Modulates Short-Term Plasticity through Unbinding from Stargazin. Neuron 85, 787–803.10.1016/j.neuron.2015.01.012Search in Google Scholar PubMed
Diamond, J. S. and Jahr, C. E. (1997). Transporters buffer synaptically released glutamate on a submillisecond time scale. J. Neurosci. 17, 4672–4687.10.1523/JNEUROSCI.17-12-04672.1997Search in Google Scholar PubMed
Domanov, Y. A., Aimon, S., Toombes, G. E., Renner, M., Quemeneur, F., Triller, A., Turner, M. S. and Bassereau, P. (2011). Mobility in geometrically confined membranes. Proc. Natl. Acad. Sci. U.S.A. 108, 12605–12610.10.1073/pnas.1102646108Search in Google Scholar PubMed PubMed Central
Eggermann, E., Bucurenciu, I., Goswami, S. P. and Jonas, P. (2012). Nanodomain coupling between Ca(2)(+) channels and sensors of exocytosis at fast mammalian synapses. Nat. Rev. Neurosci. 13, 7–21.10.1038/nrn3125Search in Google Scholar PubMed PubMed Central
Engert, F. and Bonhoeffer, T. (1999). Dendritic spine changes associated with hippocampal long-term synaptic plasticity. Nature 399, 66–70.10.1038/19978Search in Google Scholar PubMed
Fu, Y. and Huang, Z. J. (2010). Differential dynamics and activity-dependent regulation of alpha- and beta-neurexins at developing GABAergic synapses. Proc. Natl. Acad. Sci. U.S.A. 107, 22699–22704.10.1073/pnas.1011233108Search in Google Scholar PubMed PubMed Central
Gulyas, A. I., Megias, M., Emri, Z. and Freund, T. F. (1999). Total number and ratio of excitatory and inhibitory synapses converging onto single interneurons of different types in the CA1 area of the rat hippocampus. J. Neurosci. 19, 10082–10097.10.1523/JNEUROSCI.19-22-10082.1999Search in Google Scholar PubMed
Heine, M., Groc, L., Frischknecht, R., Beique, J. C., Lounis, B., Rumbaugh, G., Huganir, R. L., Cognet, L. and Choquet, D. (2008). Surface mobility of postsynaptic AMPARs tunes synaptic transmission. Science 320, 201–205.10.1126/science.1152089Search in Google Scholar PubMed
Hoze, N., Nair, D., Hosy, E., Sieben, C., Manley, S., Herrmann, A., Sibarita, J. B., Choquet, D. and Holcman, D. (2012). Heterogeneity of AMPA receptor trafficking and molecular interactions revealed by superresolution analysis of live cell imaging. Proc. Natl. Acad. Sci. U.S.A. 109, 17052–17057.10.1073/pnas.1204589109Search in Google Scholar PubMed
Kaeser, P. S., Deng, L., Wang, Y., Dulubova, I., Liu, X., Rizo, J. and Sudhof, T. C. (2011). RIM proteins tether Ca2+ channels to presynaptic active zones via a direct PDZ-domain interaction. Cell 144, 282–295.10.1016/j.cell.2010.12.029Search in Google Scholar
Kerchner, G. A. and Nicoll, R. A. (2008). Silent synapses and the emergence of a postsynaptic mechanism for LTP. Nat. Rev. Neurosci. 9, 813–825.10.1038/nrn2501Search in Google Scholar PubMed
Kim, C. H., Takamiya, K., Petralia, R. S., Sattler, R., Yu, S., Zhou, W., Kalb, R., Wenthold, R., Huganir, R. and M-334* (2005). Persistent hippocampal CA1 LTP in mice lacking the C-terminal PDZ ligand of GluR1. Nat. Neurosci. 8, 985–987.10.1038/nn1432Search in Google Scholar PubMed
Kittler, J. T. and Moss, S. J. (2003). Modulation of GABAA receptor activity by phosphorylation and receptor trafficking: implications for the efficacy of synaptic inhibition. Curr. Opin. Neurobiol. 13, 341–347.10.1016/S0959-4388(03)00064-3Search in Google Scholar PubMed
Lasne, D., Blab, G. A., Berciaud, S., Heine, M., Groc, L., Choquet, D., Cognet, L. and Lounis, B. (2006). Single nanoparticle photothermal tracking (SNaPT) of 5-nm gold beads in live cells. Biophys. J. 91, 4598–4604.10.1529/biophysj.106.089771Search in Google Scholar PubMed
Liu, G., Choi, S. and Tsien, R. W. (1999). Variability of neurotransmitter concentration and nonsaturation of postsynaptic AMPA receptors at synapses in hippocampal cultures and slices. Neuron 22, 395–409.10.1016/S0896-6273(00)81099-5Search in Google Scholar PubMed
Luscher, C., Xia, H., Beattie, E. C., Carroll, R. C., von Zastrow, M., Malenka, R. C. and Nicoll, R. A. (1999). Role of AMPA receptor cycling in synaptic transmission and plasticity. Neuron 24, 649–658.10.1016/S0896-6273(00)81119-8Search in Google Scholar PubMed
Maher, M. P., Matta, J. A., Gu, S., Seierstad, M. and Bredt, D. S. (2017). Getting a Handle on Neuropharmacology by Targeting Receptor-Associated Proteins. Neuron 96, 989–1001.10.1016/j.neuron.2017.10.001Search in Google Scholar PubMed
Markram, H., Wang, Y. and Tsodyks, M. (1998). Differential signaling via the same axon of neocortical pyramidal neurons. Proc. Natl. Acad. Sci. U.S.A. 95, 5323–5328.10.1073/pnas.95.9.5323Search in Google Scholar PubMed
Maschi, D. and Klyachko, V. A. (2017). Spatiotemporal Regulation of Synaptic Vesicle Fusion Sites in Central Synapses. Neuron 94, 65–73 e63.10.1016/j.neuron.2017.03.006Search in Google Scholar
Megias, M., Emri, Z., Freund, T. F. and Gulyas, A. I. (2001). Total number and distribution of inhibitory and excitatory synapses on hippocampal CA1 pyramidal cells. Neuroscience 102, 527–540.10.1016/S0306-4522(00)00496-6Search in Google Scholar PubMed
Missler, M., Zhang, W., Rohlmann, A., Kattenstroth, G., Hammer, R. E., Gottmann, K. and Sudhof, T. C. (2003). Alpha-neurexins couple Ca2+ channels to synaptic vesicle exocytosis. Nature 423, 939–948.10.1038/nature01755Search in Google Scholar PubMed
Nakada, C., Ritchie, K., Oba, Y., Nakamura, M., Hotta, Y., Iino, R., Kasai, R. S., Yamaguchi, K., Fujiwara, T. and Kusumi, A. (2003). Accumulation of anchored proteins forms membrane diffusion barriers during neuronal polarization. Nat. Cell Biol. 5, 626–632.10.1038/ncb1009Search in Google Scholar PubMed
Neupert, C., Schneider, R., Klatt, O., Reissner, C., Repetto, D., Biermann, B., Niesmann, K., Missler, M. and Heine, M. (2015). Regulated Dynamic Trafficking of Neurexins Inside and Outside of Synaptic Terminals. J. Neurosci. 35, 13629–13647.10.1523/JNEUROSCI.4041-14.2015Search in Google Scholar PubMed PubMed Central
Opazo, P., Labrecque, S., Tigaret, C. M., Frouin, A., Wiseman, P. W., De Koninck, P. and Choquet, D. (2010). CaMKII triggers the diffusional trapping of surface AMPARs through phosphorylation of stargazin. Neuron 67, 239–252.10.1016/j.neuron.2010.06.007Search in Google Scholar PubMed
Penn, A. C., Zhang, C. L., Georges, F., Royer, L., Breillat, C., Hosy, E., Petersen, J. D., Humeau, Y. and Choquet, D. (2017). Hippocampal LTP and contextual learning require surface diffusion of AMPA receptors. Nature 549, 384–388.10.1038/nature23658Search in Google Scholar PubMed PubMed Central
Pouille, F. and Scanziani, M. (2004). Routing of spike series by dynamic circuits in the hippocampus. Nature 429, 717–723.10.1038/nature02615Search in Google Scholar PubMed
Renner, M., Wang, L., Levi, S., Hennekinne, L. and Triller, A. (2017). A Simple and Powerful Analysis of Lateral Subdiffusion Using Single Particle Tracking. Biophys. J. 113, 2452–2463.10.1016/j.bpj.2017.09.017Search in Google Scholar PubMed PubMed Central
Ritchie, K. and Kusumi, A. (2003). Single-particle tracking image microscopy. Methods Enzymol. 360, 618–634.10.1016/S0076-6879(03)60131-XSearch in Google Scholar PubMed
Schneider, R., Hosy, E., Kohl, J., Klueva, J., Choquet, D., Thomas, U., Voigt, A. and Heine, M. (2015). Mobility of calcium channels in the presynaptic membrane. Neuron 86, 672–679.10.1016/j.neuron.2015.03.050Search in Google Scholar PubMed
Schwenk, J., Baehrens, D., Haupt, A., Bildl, W., Boudkkazi, S., Roeper, J., Fakler, B. and Schulte, U. (2014). Regional diversity and developmental dynamics of the AMPA-receptor proteome in the mammalian brain. Neuron 84, 41–54.10.1016/j.neuron.2014.08.044Search in Google Scholar PubMed
Schwenk, J., Harmel, N., Brechet, A., Zolles, G., Berkefeld, H., Muller, C. S., Bildl, W., Baehrens, D., Huber, B., Kulik, A. et al. (2012). High-resolution proteomics unravel architecture and molecular diversity of native AMPA receptor complexes. Neuron 74, 621–633.10.1016/j.neuron.2012.03.034Search in Google Scholar PubMed
Schwenk, J., Harmel, N., Zolles, G., Bildl, W., Kulik, A., Heimrich, B., Chisaka, O., Jonas, P., Schulte, U., Fakler, B. et al. (2009). Functional proteomics identify cornichon proteins as auxiliary subunits of AMPA receptors. Science 323, 1313–1319.10.1126/science.1167852Search in Google Scholar PubMed
Shi, S., Hayashi, Y., Esteban, J. A. and Malinow, R. (2001). Subunit-specific rules governing AMPA receptor trafficking to synapses in hippocampal pyramidal neurons. Cell 105, 331–343.10.1016/S0092-8674(01)00321-XSearch in Google Scholar PubMed
Sylwestrak, E. L. and Ghosh, A. (2012). Elfn1 regulates target-specific release probability at CA1-interneuron synapses. Science 338, 536–540.10.1126/science.1222482Search in Google Scholar PubMed PubMed Central
Tang, A. H., Chen, H., Li, T. P., Metzbower, S. R., MacGillavry, H. D. and Blanpied, T. A. (2016). A trans-synaptic nanocolumn aligns neurotransmitter release to receptors. Nature 536, 210–214.10.1038/nature19058Search in Google Scholar PubMed PubMed Central
Thomas, P. and Smart, T. G. (2006). Receptor Dynamics at the Cell Surface Studied Using Functional Tagging. In The Dynamic Synapse: Molecular Methods in Ionotropic Receptor Biology, Kittler, J. T. and Moss, S. J., eds. (Boca Raton (FL)).Search in Google Scholar
Tovar, K. R. and Westbrook, G. L. (1999). The incorporation of NMDA receptors with a distinct subunit composition at nascent hippocampal synapses in vitro. J. Neurosci. 19, 4180–4188.10.1523/JNEUROSCI.19-10-04180.1999Search in Google Scholar PubMed
Tran-Van-Minh, A., Caze, R. D., Abrahamsson, T., Cathala, L., Gutkin, B. S. and DiGregorio, D. A. (2015). Contribution of sublinear and supralinear dendritic integration to neuronal computations. Front. Cell. Neurosci. 9, 67.10.3389/fncel.2015.00067Search in Google Scholar PubMed PubMed Central
Trimble, W. S. and Grinstein, S. (2015). Barriers to the free diffusion of proteins and lipids in the plasma membrane. J. Cell Biol. 208, 259–271.10.1083/jcb.201410071Search in Google Scholar PubMed PubMed Central
von Engelhardt, J., Mack, V., Sprengel, R., Kavenstock, N., Li, K. W., Stern-Bach, Y., Smit, A. B., Seeburg, P. H. and Monyer, H. (2010). CKAMP44: a brain-specific protein attenuating short-term synaptic plasticity in the dentate gyrus. Science 327, 1518–1522.10.1126/science.1184178Search in Google Scholar PubMed
Winckler, B., Forscher, P., Mellman, I. and M-253* (1999). A diffusion barrier maintains distribution of membrane proteins in polarized neurons. Nature 397, 698–701.10.1038/17806Search in Google Scholar PubMed
Xu, K., Zhong, G. and Zhuang, X. (2013). Actin, spectrin, and associated proteins form a periodic cytoskeletal structure in axons. Science 339, 452–456.10.1126/science.1232251Search in Google Scholar PubMed PubMed Central
Young, S. H. and Poo, M. M. (1983). Rapid lateral diffusion of extrajunctional acetylcholine receptors in the developing muscle membrane of Xenopus tadpole. J. Neurosci. 3, 225–231.10.1523/JNEUROSCI.03-01-00225.1983Search in Google Scholar PubMed
Zucker, R. S. and Regehr, W. G. (2002). Short-term synaptic plasticity. Annu. Rev. Physiol. 64, 355–405.10.1146/annurev.physiol.64.092501.114547Search in Google Scholar PubMed
Anmerkung
Englische Version des Artikels online verfügbar unter https://doi.org/10.1515/nf-2017-A042
© 2018 by De Gruyter
Articles in the same Issue
- Frontmatter
- Übersichtsartikel
- Gedächtnismechanismen in Drosophila
- Making Memories. On the fly.
- Bewegungssehen: Zellen, Schaltkreise und Algorithmen
- Motion detection: cells, circuits and algorithms
- Molekulare Dynamik der neuronalen Informationsübertragung
- Molecular dynamics of neuronal information transfer
- Die Rolle von Histonmethylierungen in der Entwicklung des zentralen Nervensystems und bei Neuralrohrdefekten
- Histone methylations in the developing central nervous system and in neural tube defects
- „Mind from Matter?“ – Über Verhalten und Gehirn
- Mind from Matter? – Via Brain and Behavior
- Forschungsförderung
- Sonderforschungsbereich (SFB 1280) „Extinktionslernen“
- Kollaboratives Forschungszentrum (Sonderforschungsbereich, SFB) 1286 „Quantitative Synaptologie“
- Nachruf
- Nachruf auf Prof. Dr. Dr. h. c. mult. Franz Huber
- Nachruf auf Robert F. Schmidt
- Rezension
- Norman Doidge: Neustart im Kopf – Wie sich unser Gehirn selbst repariert
- Nachrichten
- Einladung zur Mitgliederversammlung während des FENS Forum 2018 in Berlin (7.–11. Juli 2018)
Articles in the same Issue
- Frontmatter
- Übersichtsartikel
- Gedächtnismechanismen in Drosophila
- Making Memories. On the fly.
- Bewegungssehen: Zellen, Schaltkreise und Algorithmen
- Motion detection: cells, circuits and algorithms
- Molekulare Dynamik der neuronalen Informationsübertragung
- Molecular dynamics of neuronal information transfer
- Die Rolle von Histonmethylierungen in der Entwicklung des zentralen Nervensystems und bei Neuralrohrdefekten
- Histone methylations in the developing central nervous system and in neural tube defects
- „Mind from Matter?“ – Über Verhalten und Gehirn
- Mind from Matter? – Via Brain and Behavior
- Forschungsförderung
- Sonderforschungsbereich (SFB 1280) „Extinktionslernen“
- Kollaboratives Forschungszentrum (Sonderforschungsbereich, SFB) 1286 „Quantitative Synaptologie“
- Nachruf
- Nachruf auf Prof. Dr. Dr. h. c. mult. Franz Huber
- Nachruf auf Robert F. Schmidt
- Rezension
- Norman Doidge: Neustart im Kopf – Wie sich unser Gehirn selbst repariert
- Nachrichten
- Einladung zur Mitgliederversammlung während des FENS Forum 2018 in Berlin (7.–11. Juli 2018)