Three-dimensional distribution of nutrients and phytoplankton biomass in a semi-enclosed region of the Gulf of California during different ENSO phases
-
Carlos Mauricio Torres-Martínez
Carlos Mauricio Torres-Martínez earned a Master’s degree from the National Autonomous University of Mexico (UNAM). He is currently a Ph.D. student in the graduate program of Marine Sciences and Limnology at UNAM, where his research focuses on the structure of phytoplankton communities and their relationship with hydrodynamic processes.
 ,
María Adela Monreal-Gómez
,
María Adela Monreal-Gómez
Dra. María Adela Monreal-Gómez earned her PhD in oceanology from the University of Liège (Belgium). Her interest in physical oceanography focuses on oceanic processes, such as vortices, fronts, and upwelling events, which emphasize the interactions between physics and biology.
,
Erik Coria-Monter
Erik Coria-Monter has a Ph.D. in biological sciences from the Autonomous Metropolitan University (UAM) of Mexico and is Professor at the National Autonomous University of Mexico (UNAM). His research is focused on plankton ecology, trophic ecology, and physical/biological interactions in marine ecosystems.
 ,
David Alberto Salas-de-León
,
David Alberto Salas-de-León
Dr. David Alberto Salas-de-León obtained his PhD in oceanology at Liege University, Belgium, focusing on ocean models with a strong focus on physics-biology interactions. Currently, he is also studying non-linear systems and chaos.
,
Elizabeth Durán-Campos
Elizabeth Durán-Campos earned a Ph.D. in biological sciences (2015) from the Autonomous Metropolitan University of Mexico. She does research in plankton ecology and physical/biological interactions in both coastal and oceanic environments.
and
Martín Merino-Ibarra

Abstract
This study examined the three-dimensional distribution of nutrients and phytoplankton biomass, as measured by chlorophyll-a (Chl-a) concentration, in the Bay of La Paz, Gulf of California, during different phases of the El Niño Southern Oscillation (ENSO). Two multidisciplinary research cruises were conducted in November 2014 and November 2016, corresponding to El Niño and La Niña conditions, respectively. A CTD Rosette System was used to gather high-resolution hydrographic data and collect seawater samples at different depths for chemical analyses (nutrients and Chl-a). Meteorological data were obtained from a local weather station. The results indicated significant differences between the two study periods. The composition of water masses varied during both events. Notable contrasts were observed in nutrient concentrations, especially at 30 m depth; the maximum concentration of Soluble Reactive Si was 152.46 µM in 2014, compared to only 8.48 µM in 2016. Chl-a levels also showed variations with depth and between the two cruises, being significantly higher in 2014 (7.60 mg m−3) than in 2016 (1.70 mg m−3). This study enhances the understanding of the dynamics of ENSO events, given that the collections were conducted during both an El Niño and a La Niña phase. However, it is worth noting that the La Niña was influenced by an extreme El Niño event occurring in the same year, which likely masked the effects of the subsequent La Niña.
1 Introduction
Phytoplankton is a diverse group of microorganisms that thrive in oceans around the world and are vital to many ecosystem functions. Notably, phytoplankton contributes over 50 % of the primary production on Earth (Falkowski 2012). It plays an essential role in absorbing atmospheric CO2 and releasing oxygen through the process of photosynthesis (Brierley 2017). Additionally, phytoplankton forms the base of pelagic food webs, supporting higher trophic levels, including species that have significant ecological and commercial importance (Richardson 2008).
Chlorophyll-a (Chl-a) is the primary photosynthetic pigment found in all autotrophic species and is widely considered one of the key proxies of phytoplankton biomass (Davies et al. 2018). Its measurement is a standard practice that can be carried out using simple and standardized chemical techniques, such as spectrophotometry, which do not require specialized equipment (Roy et al. 2011). Additionally, Chl-a can be quantified using fluorometric probes, known for their precision and sensitivity (Roesler et al. 2017).
A growing body of scientific evidence shows that both the vertical and horizontal distribution of Chl-a is influenced by complex factors involving numerous physical and biological interactions (McGillicuddy 2016). Consequently, the distribution of Chl-a is highly heterogeneous. This variability affects zooplankton populations because herbivorous and filtering organisms, such as copepods, must adapt to these patterns in order to find food. This adaptation is crucial for the effective functioning of the biological carbon pump (Brierley 2017).
Moreover, the distribution of Chl-a levels is closely linked to physical processes occurring within water columns at various spatial and temporal scales. These processes include internal waves (Villamaña et al. 2017), oceanic fronts (Lévy et al. 2018), eddies (Gaube et al. 2014), and large-scale phenomena at the ocean-atmosphere interface, such as the El Niño Southern Oscillation (ENSO). ENSO has two phases: La Niña, which is characterized by cooling, and El Niño, which is marked by warming of sea surface waters. During the El Niño phase, the advection of oligotrophic water masses can affect phytoplankton biomass, depending on the intensity of the event (McPhaden et al. 2021).
Between 2014 and 2016, several ENSO events of varying magnitudes were observed. The first half of 2014 experienced neutral conditions, followed by an extreme El Niño event that lasted from May 2015 to April 2016. This prolonged and intense phenomenon was dubbed the “Godzilla El Niño” (Schiermeier 2015). Its effects on phytoplankton biomass were significant, leading to extremely low levels of Chl-a in oceanic and coastal environments worldwide (McPhaden et al. 2021).
Coastal environments are well-known for their high biological diversity and productivity, serving as essential areas for refuge, feeding, and spawning for many species (Zhang et al. 2017). Multidisciplinary studies have demonstrated that the species richness and biological production in these coastal areas are closely linked to phytoplankton, which forms the base of the food chain (Dai et al. 2023). Consequently, research on the variability of phytoplankton and its relationship with large-scale processes, such as ENSO events, has become increasingly common in recent years. This research spans different regions around the world, including the Arabian Sea (Shafeeque et al. 2021) and the South China Sea (Xiu et al. 2018).
In the coastal areas of the Southwestern Atlantic, ENSO events, including both El Niño and La Niña, have a significant impact on wind patterns. These changes are closely related to upwellings, which affect the concentration of nutrients and phytoplankton biomass (Machado et al. 2013). In the coastal regions of Brazil, La Niña events, in particular, have been associated with increased levels of precipitation. This increase in rainfall alters the physical, chemical, and biological parameters of the area, leading to reduced salinity and elevated concentrations of nutrients and phytoplankton biomass (Andrade et al. 2016).
Numerous studies have documented the impact of ENSO events on phytoplankton biomass in Mexican waters through satellite and field observations. For instance, research by Sosa-Ávalos et al. (2021) in Manzanillo and Santiago Bays on the Pacific coast found that high temperatures corresponded with El Niño events, while low temperatures were associated with La Niña events. Specifically, surface Chl-a levels were found to be 5.4 times higher during La Niña compared to El Niño.
In the Gulf of Tehuantepec, satellite observations indicated unusually low Chl-a levels (<0.1 mg m−3) during the 2015 El Niño event (Coria-Monter et al. 2019a). Herrera-Cervantes et al. (2010) also examined the influence of ENSO on satellite-derived Chl-a levels along the Gulf of California and found a clear pattern: Chl-a concentrations decreased during El Niño and increased during La Niña. The effects of ENSO were not uniform across the region; the northern area experienced the most significant changes, while the islands exhibited the least variability.
In the Bay of La Paz, located in the southern Gulf of California, Herrera-Cervantes et al. (2020) analyzed interannual variability in satellite-derived Chl-a concentrations. They found that both El Niño and La Niña affected Chl-a levels within the bay, with maximum concentrations occurring during La Niña events and a significant decrease during El Niño due to the advection of warm, oligotrophic waters.
In recent years, significant efforts have been made to understand the relationship between the variability of phytoplankton biomass with ENSO events in Mexican waters. However, a complete characterization is still lacking. This study aims to assess the three-dimensional distribution of nutrients and Chl-a levels during different ENSO phases in the Bay of La Paz, the largest and deepest coastal environment within the Gulf of California, known for its high biological diversity.
Data for this study were collected during two multidisciplinary research cruises on board the R/V El Puma, operated by the National Autonomous University of Mexico. These cruises took place in November during the autumn months of 2014 and 2016. The current study seeks to evaluate how phytoplankton biomass responds to ENSO events, specifically examining the El Niño conditions observed in November 2014 and the subsequent La Niña event following the prolonged “Godzilla” El Niño of 2015/2016. Ultimately, this study aims to enhance scientific understanding of the ecosystems in Mexican environments, particularly in the Bay of La Paz.
2 Materials and methods
2.1 Study area
The Bay of La Paz is the largest and deepest coastal environment within the Gulf of California (Figure 1). It is located approximately 200 km from where the Gulf meets the Pacific Ocean, in the southeastern Baja California peninsula. This area is well-known for its high biological diversity and serves as a refuge and feeding habitat for numerous species, including some that are threatened or endangered (Durán-Campos et al. 2020).

Location of (a) the Gulf of California, and (b) the Bay of La Paz. ᛫ Symbols represent the localities sampled in this study. Bathymetry is shown in m.
The bay connects directly to the Gulf of California through two openings: the San Lorenzo Channel to the southeast and Boca Grande to the northeast. The San Lorenzo Channel is narrow (20 m deep) and shallow (6 km wide). In contrast, Boca Grande is deep (250 m deep) and wide (32 km). Boca Grande is the primary area for the transport and exchange of water masses between the bay and the Gulf of California.
The bathymetry of the bay is complex, featuring a series of bathymetric sills in its northern portion where it connects with the Gulf. The maximum depth of the bay is 420 m, found in the central region known as Cuenca Alfonso. In the southern area, the depth decreases to 20 m and is characterized by slopes and extensive beaches (Molina-Cruz et al. 2002).
The regional climate is classified as desertic (type BwH), characterized by evaporation rates that exceed precipitation (García 1989). Notably, there is no river discharge into the bay. A significant feature of the region is the wind pattern, which experiences considerable seasonal variations. During winter (December to March), strong and persistent dry winds from the northwest, exceeding 12 m s−1, dominate the area. In contrast, during summer (June to September), the wind pattern reverses, bringing weaker and wetter winds from the southeast, with speeds less than 5 m s−1, and frequent calm periods (Monreal-Gomez et al. 2001).
The depth of the thermocline and nutricline, along with the mixing of the water column, is directly linked to these wind patterns. The wind promotes a cyclonic eddy in the bay, which plays a crucial role in supplying nutrients to the euphotic layer, essential for phytoplankton communities.
The thermohaline structure of the bay consists of four main water masses: Tropical Surface Water (TSW, S < 34.90, and T ≥ 18.00 °C), Gulf of California Water (GCW, S > 34.90, and T ≥ 12.00 °C), Subtropical Subsurface Water (StSsW, 34.50 < S < 34.90, and 9.00 ≤ T ≤ 18.00 °C), and Pacific Intermediate Water (PIW, 34.50 ≤ S < 34.80, and 4.00 ≤ T < 9.00 °C). The PIW is specifically found in the Boca Grande region, off the bay (Coria-Monter et al. 2019b; Monreal-Gómez et al. 2001).
Within the bay, both surface and subsurface circulation are dominated by a cyclonic eddy, which significantly affects the distribution of phytoplankton organisms. This cyclonic eddy leads Ekman pumping, which fertilizes the euphotic layer (Coria-Monter et al. 2017) and causes phytoplankton to aggregate differentially from the center towards the periphery (Coria-Monter et al. 2014). Furthermore, additional processes like the propagation of internal waves and hydraulic jumps increase the variability of the water column, thereby enhancing phytoplankton biomass (Coria-Monter et al. 2019c).
2.2 Sampling
The data set used in this study was collected during two oceanographic cruises. The first, known as the Paleaomar-1 cruise, took place in November 2014, while the second cruise, Paleomar-2, was conducted in November 2016. Both cruises were carried out onboard the R/V El Puma, operated by the National Autonomous University of Mexico. The sampling strategy involved an equidistant grid consisting of 44 hydrographic stations that covered the interior of the bay and its connection to the Gulf of California (Figure 1).
At each station, hydrographic data were collected using a CTD probe (SeaBird 19 plus), which has a precision of 0.005 °C for temperature, and 0.0005 S m−1 for conductivity. The CTD was attached to a Rosette System containing 12 Niskin bottles (General Oceanics) with a capacity of 10 l each. The CTD-Rosette system was deployed from the water surface to 5 m above the seafloor, descending at a speed of 1 m s−1, with data being acquired at a frequency of 24 Hz.
Water samples for nutrients and Chl-a analyses were collected at different depths: surface, 10, 30, and 50 m depth. Once the Rosette System was onboard, 100-ml subsamples for nutrient determinations were filtered through two cellulose acetate syringe filters (Merck Millipore, 0.22-μm and 0.45-μm pore sizes). These subsamples were then stored in polypropylene containers that had been previously washed with an acid solution, fixed with chloroform, and kept frozen at −20 °C until laboratory analysis.
For Chl-a determinations, 4-l subsamples were filtered through nitrocellulose membrane filters (Merck Millipore, 0.45-μm pore size) using a stainless-steel vacuum manifold system set at 10 psi. During the filtration process, special precautions were taken to avoid contamination, including rinsing the manifold system with distilled water between each sample and conducting the procedure under low-light conditions to prevent degradation. Finally, the membranes were stored in plastic centrifuge tubes covered with aluminum foil and frozen at −20 °C prior to analysis.
2.3 Laboratory chemical analyses
The water samples collected for nutrient analysis were analyzed in the laboratory immediately after the research cruises. The concentrations of nitrate (precision of 0.10 µM), nitrite (precision of 0.02 µM), soluble reactive phosphorus (SRP, precision of 0.04 µM), and soluble reactive silica (SRSi, precision of 0.10 µM) were determined using a segmented flow analyzer (Skalar San Plus) following the methods outlined by Grasshoff et al. (1983) and Kirkwood (1994).
For the quantification of Chl-a, 90 % acetone was added to the centrifuge tubes containing the membranes. The samples were extracted in the dark for 24 h and then stored at −20 °C. After this extraction period, the tubes were centrifuged at 2,490g (Eppendorf 5,840 centrifuge) for 30 min. Finally, in accordance with the methodology of Strickland and Parsons (1972), the absorbance was measured in triplicate at wavelengths of 750, 664, 647, and 630 nm using a spectrophotometer (Genesys 10S UV-VIS, Thermo Scientific) with quartz cells. This entire process was carried out under extremely low lighting conditions.
2.4 Data analyses
To identify the ENSO phases that occurred during each research cruise, the Ocean Niño Index (ONI) provided by the NOAA National Weather Service was used (https://origin.cpc.ncep.noaa.gov/products/analysis_monitoring/ensostuff/ONI_v5.php). The ONI is a reliable indicator for monitoring the evolution and strength of ENSO events in the El Niño 3.4 oceanic region. According to this index, values higher than +0.5 indicate El Niño conditions, while values lower than −0.5 indicate La Niña conditions.
The CTD data were initially processed with the nominal calibration file, following the routines and subroutines of the manufacturer with SBE Data Processing software V.7.26.7, averaging each dbar. Then, salinity and temperature were derived using standard algorithms. To analyze the presence and proportions of the water masses in both research cruises, temperature-salinity (TS) diagrams were constructed based on the classification outlined by Lavín et al. (2009).
Wind data for November 2014 and November 2016 were obtained from meteorological station # 76404, operated by the National Meteorological Service of Mexico (Servicio Meteorológico Nacional) and located in La Paz city. This dataset included records of wind magnitude and direction recorded every 15 min. To eliminate high-frequency fluctuations, a 24-h running average was applied.
3 Results
3.1 ENSO phases
During the Paleomar-1 cruise in November 2014, the ONI recorded a value of +0.6 indicating the presence of an El Niño event. In contrast, during the Paleomar-2 cruise in November 2016, a value of −0.7 was registered, signifying La Niña conditions. Thus, both events exhibited similar intensity.
3.2 Hydrography
The T-S diagrams generated indicated notable differences in the number and proportion of water masses observed during the two research cruises (Figure 2).

Temperature-salinity diagram of both research cruises considered in this study. Red points, Paleomar-1 cruise, November 2014; blue points, Paleomar-2 cruise, November 2016; StSsW, subtropical subsurface water; GCW, Gulf of California water; TSW, tropical surface water; PIW, Pacific intermediate water.
During the Paleomar-1 cruise, four distinct water masses were identified: (1) Subtropical Subsurface Water, (2) Gulf of California Water, (3) Tropical Surface Water, and (4) Pacific Intermediate Water (Figure 2). The PIW was limited to stations in the region connecting the bay to the Gulf of California.
In contrast, only three water masses were identified during Paleomar-2 cruise, as there was no evidence of TSW. The observed water masses were (1) GCW, (2) StSsW, and (3) PIW, which was also confined to the stations linked to the Gulf.
The horizontal and vertical temperature distributions demonstrated distinct patterns during the two cruises analyzed in this study.
During the Paleomar 1 cruise, it was noted that the temperature at surface (Figure 3a), at 10 m (Figure 3b), and at 30 m depth (Figure 3c) remained largely uniform across the bay, at around 26.50 °C, except for the Boca Grande region, where temperatures dropped to approximately 25.50 °C. In contrast, at a depth of 50 m (Figure 3d), temperature gradients were observed, with colder cores (22–23 °C) located in the northern and southwestern parts of the bay.

Horizontal distribution of temperature (°C) at different depths: (a) surface, (b) 10, (c) 30, and (d) 50 m depth during the Paleomar-1 cruise, (e) surface, (f) 10, (g) 30, and (h) 50 m depth during the Paleomar-2 cruise.
During the Paleomar 2 cruise, temperature distribution showed higher temperatures compared to the Paleomar 1 cruise. In the first three depth levels (surface, 10 and 30 m depth), temperatures exceeded 27.50 °C and were uniformly distributed throughout the bay (Figure 3e–g). However, at 30 m depth, a small area of lower temperature was observed (Figure 3g). At 50 m depth, horizontal temperature gradients began to emerge, revealing a cold core (<22.00 °C) in the Alfonso Basin. In contrast, higher temperatures (>27.00 °C) were recorded in the southern bay, where strong horizontal temperature gradients were also noted (Figure 3h).
3.3 Wind field
The wind dataset collected during each research cruise exhibited notable differences. For example, during the Paleomar-1 cruise held in November 2014, the prevailing winds were northeasterly, with speeds exceeding 4 m s−1. Occasional southwesterly winds were also during the month, but these were brief (Figure 4a). In contrast, during the Paleomar-2 cruise, which took place in November 2016, the northeasterly winds were dominant, with only brief occurrences of weak southwesterly wind, typically with speeds below 1 m s−1 (Figure 4b). Additionally, the wind speeds in 2016 were more consistent compared to those recorded in 2014. It is important to note that during the Paleomar-1 cruise, the northeasterly winds exceeded 4 m s−1 on two lapses (Figure 4a), while during the Paleomar-2 cruise, the wind speeds, were around 3 m s−1 (Figure 4b).
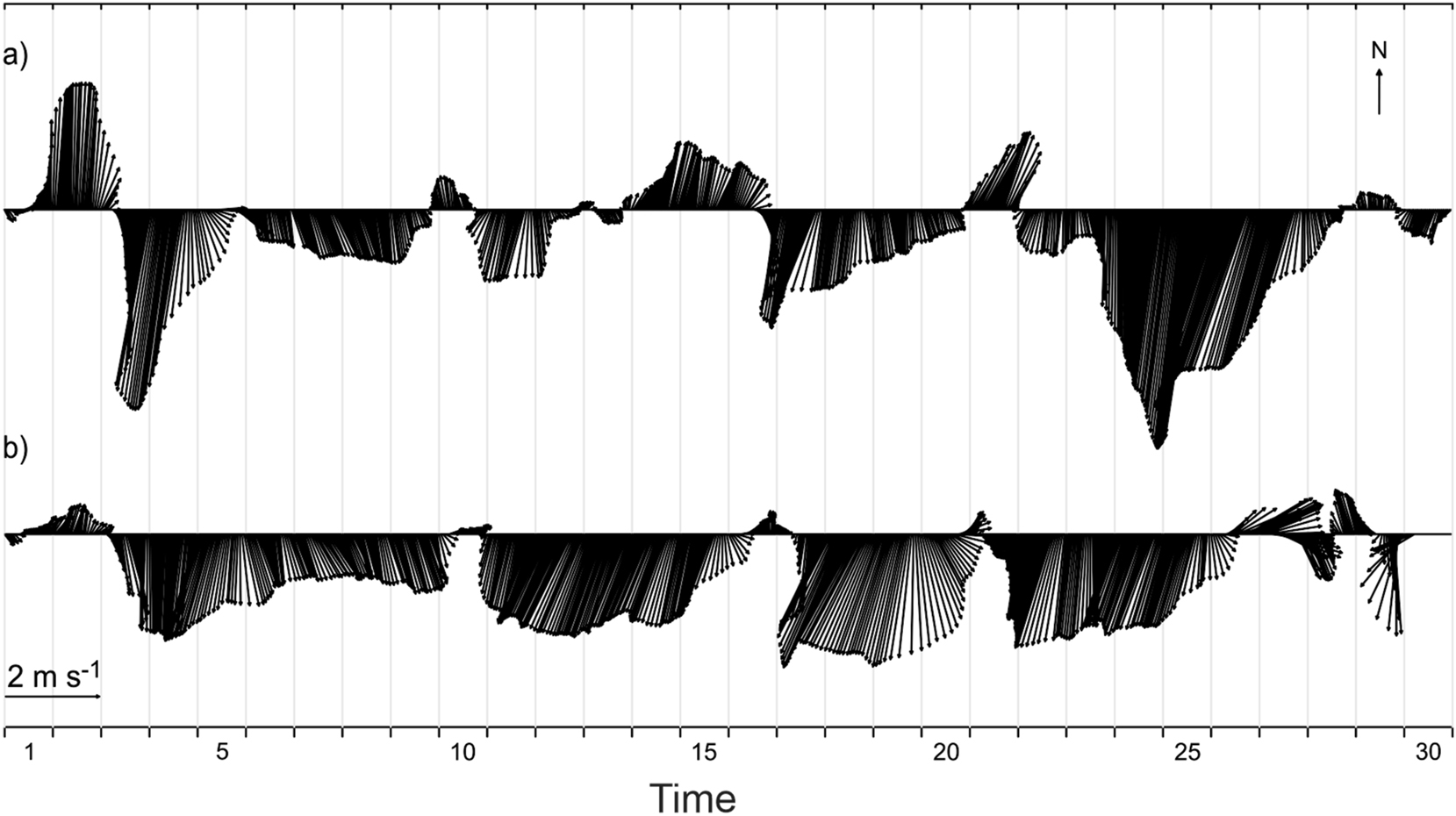
Wind velocity (m s−1) time series at weather station at La Paz, Baja California Sur, during (a) November 2014 (the Paleomar-1 cruise), (b) November 2016 (the Paleomar-2 cruise).
3.4 Nutrients
The nutrient concentrations and their horizontal distribution exhibited variations with depth and between the two research cruises.
During the Paleomar-1 cruise, at the surface, nitrate concentrations ranged from 0.07 to 2.41 µM, with the highest value recorded in a core located in the southwestern region of the bay (Figure 5a). Nitrite concentrations (Figure 6a) varied from 0.04 to 0.46 µM, showing two cores: one at the boundary with the Gulf of California and another in the southwestern region near the San Lorenzo Channel. SRP values ranged from 0.24 to 0.76 µM, exhibiting lower concentrations in the central portion of the bay (Figure 7a). Additionally, SRSi varied from 0.03 to 26.20 µM, with two high-concentration cores observed in the western section of the bay (Figure 8a).

Horizontal distribution of nitrate (µM) at different depths: (a–d) Paleomar-1 cruise; (e–h) Paleomar-2 cruise.

Horizontal distribution of nitrites (µM) at different depths: (a–d) Paleomar-1 cruise; (e–h) Paleomar-2 cruise.

Horizontal distribution of soluble reactive phosphorus (µM) at different depths: (a–d) Paleomar-1 cruise; (e–h) Paleomar-2 cruise.
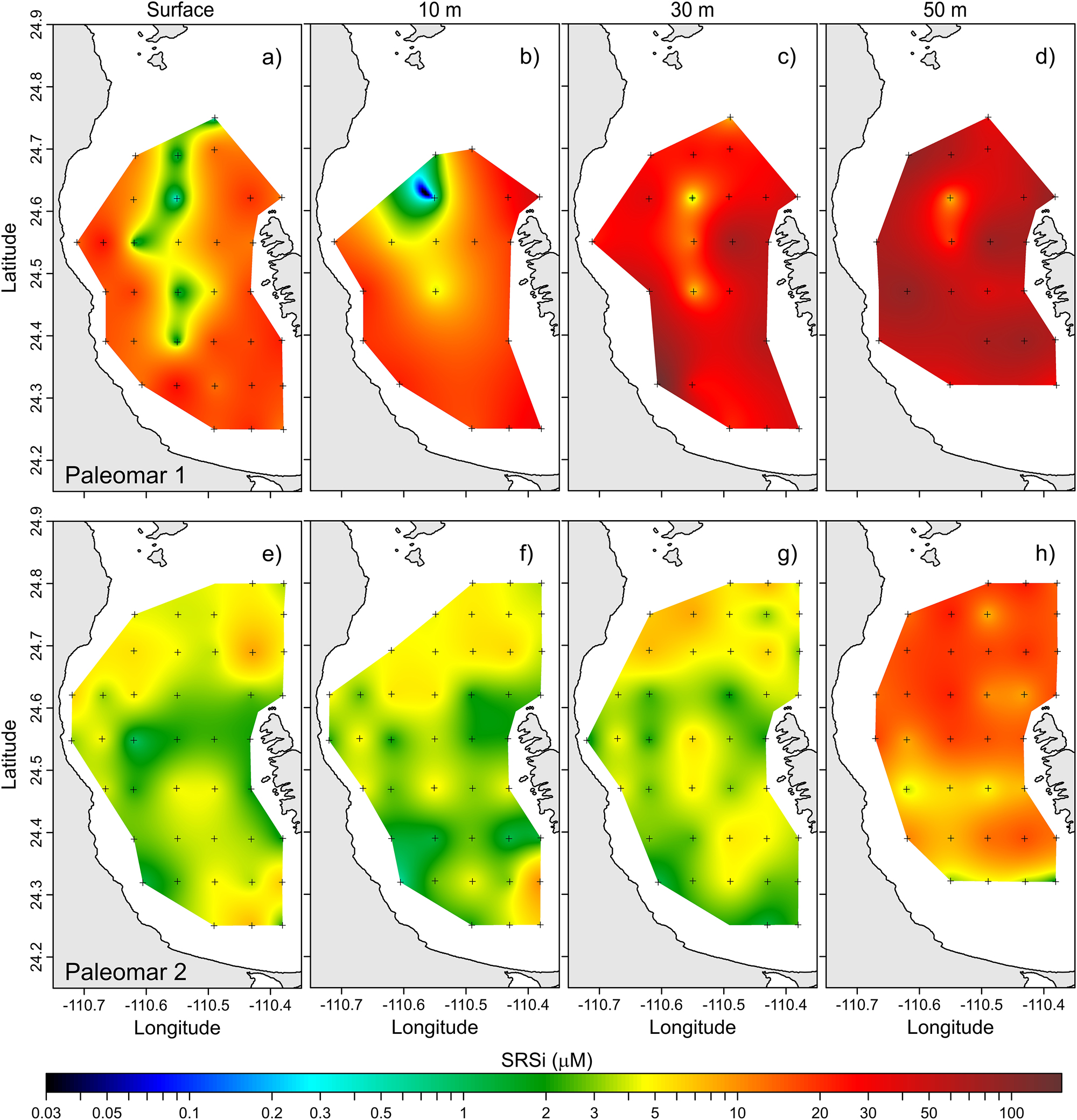
Horizontal distribution of soluble reactive Si (µM) at different depths: (a–d) Paleomar-1 cruise; (e–h) Paleomar-2 cruise.
At 10 m depth, nitrate concentrations ranged from 0.26 to 1.27 µM, with lower levels observed in the central area of the bay (Figure 5b). Nitrite concentrations varied from 0.05 to 0.64 µM, revealing two regions of high levels: one near the Gulf of California and another in the southern region near the San Lorenzo Channel (Figure 6b). SRP values ranged from 0.26 to 0.71 µM, with a core of lower concentrations also noted in the central region of the bay (Figure 7b). Finally, SRSi values ranged from 0.12 to 47.77 µM, showing high concentrations in the southwestern bay, close to the coast (Figure 8b).
At 30 m depth, nutrient concentrations were statistically higher than those observed in surface waters. Nitrate levels increased to a range of 0.56–8.79 µM, with two areas of high concentrations: one in the southwestern region near the coast and another near the San Lorenzo Channel (Figure 5c). Nitrite concentrations ranged from 0.08 to 0.57 µM, with higher values noted close to the San Lorenzo Channel (Figure 6c). SRP values ranged from 0.33 to 1.14 µM, with the highest concentrations found in the western portion of the bay, near San Juan de la Costa (Figure 7c). SRSi ranged from 1.60 to 152.46 µM, with the highest concentration observed in the San Juan de la Costa region (Figure 8c).
At 50 m depth, high concentrations of nutrients were observed. Nitrate levels ranged from 1.21 to 12.57 µM, with the highest concentrations found in the central part of the bay (Figure 5d). Nitrite levels ranged from 0.09 to 0.69 µM, displaying high values in the southern region near the San Lorenzo Channel (Figure 6d). SRP varied from 0.44 to 1.50 µM, with a peak observed in the southern area of the bay (Figure 7d). Furthermore, SRSi ranged from 5.76 to 97.62 µM, with the highest concentrations recorded in the western part of the bay (Figure 8d).
During the Paleomar-2 cruise, surface nitrate concentrations ranged from 0.00 to 0.86 µM, with lower values found in the central portion of the bay (Figure 5e). Nitrite levels varied from 0.02 to 0.37 µM, with higher concentrations observed near Boca Grande, where the bay connects to the Gulf of California (Figure 6e). SRP concentrations ranged from 0.25 to 0.55 µM, showing lower levels in the central part of the bay (Figure 7e). In contrast, SRSi concentrations varied from 0.86 to 8.97 µM, with a peak also recorded at Boca Grande, near the Gulf of California (Figure 8e).
At 10 m depth, nitrate concentrations reached up to 0.82 µM, with the highest levels found in the central portion of the bay (Figure 5f). Nitrite concentrations ranged from 0.02 to 0.32 µM, showing high levels in the central region (Figure 6f). SRP values ranged from 0.25 to 0.58 µM, revealing a core with low concentration near Roca Partida (Figure 7f). SRSi concentrations varied from 0.53 to 10.26 µM, with the highest concentrations located in the northern region of the bay (Figure 8f).
At 30 m depth, the nitrate concentration ranged from 0.02 to 2.73 µM, with the highest values found in the central and northern areas of the bay (Figure 5g). The nitrite concentration ranged from 0.02 to 0.22 µM, peaking in the central region (Figure 6g). SRP levels ranged from 0.24 to 0.67 µM, displaying a uniform distribution pattern throughout the bay (Figure 7g). The concentration of SRSi ranged from 1.10 to 8.48 µM, also showing a uniform distribution pattern (Figure 8g).
Finally, at 50 m depth, nitrate concentrations increased significantly, ranging from 0.21 to 13.22 µM, with high levels observed in the central and northern regions (Figure 5h). Nitrite concentrations varied from 0.04 to 0.50 µM, with a notable peak near the San Lorenzo Channel and a lower value in the central portion (Figure 6h). The SRP levels varied from 0.32 to 2.08 µM, showing a high concentration core in the central region of the bay (Figure 7h). Finally, SRSi concentrations ranged from 1.43 to 23.63 µM, with the highest values also located in the central region of the bay (Figure 8h).
The values of SRSi measured during the Paleomar-1 cruise were significantly higher than those recorded in the Paleomar-2 cruise, especially at 30 m depth. For example, the SRSi concentration reached 152.46 µM during the Paleomar-1 cruise (Figure 8c), whereas the highest concentration observed in the Paleomar-2 cruise was only 8.48 µM (Figure 8g).
Statistical t-tests performed on the dataset indicated significant differences, particularly at the surface, 10, and 30 m depths (Supplementary Table S1).
3.5 Chlorophyll-a
The Chl-a levels at the surface, 10, 30, and 50 m depth, within the euphotic zone exhibited variations with depth between the two cruises, which indicate changes in nutrient concentrations.
During the Paleomar-1 cruise, the surface of Chl-a concentrations ranged from 0.04 to 2.60 mg m−3, with the highest levels detected in a core located in the southern bay (Figure 9a). At 10 m depth, the Chl-a levels remained similar to those observed at the surface, peaking at 2.69 mg m−3 in the northern bay (Figure 9b). At 30 m depth, the concentrations fluctuated from 0 to 2.83 mg m−3, with the highest levels found in the central bay (Figure 9c). Finally, at 50 m depth, high values of up to 7.60 mg m−3 were recorded along the western coast of the bay (Figure 9d).
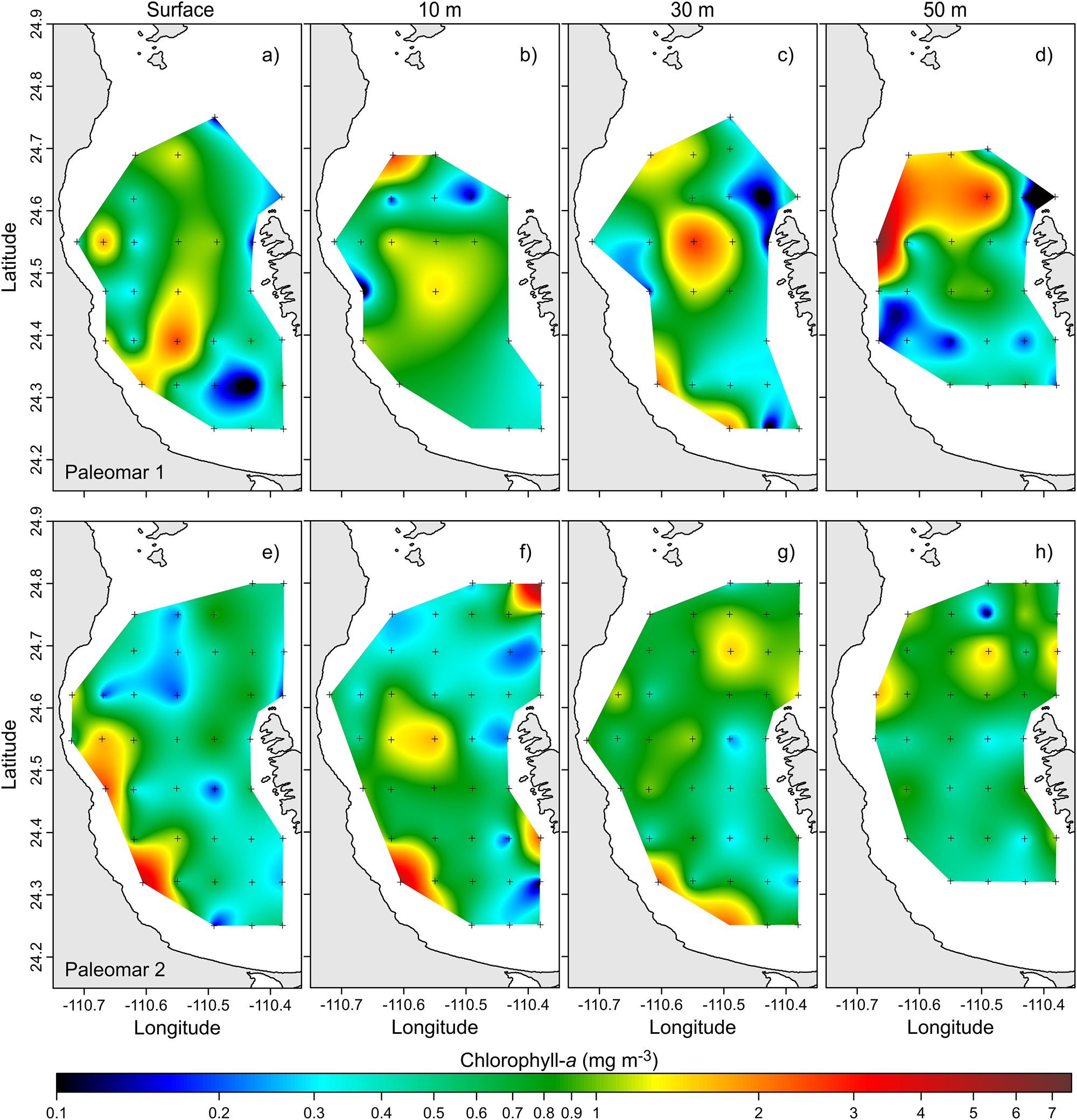
Horizontal distribution of Chl-a (mg m−3) levels at different depths: (a–d) Paleomar-1 cruise; (e–f) Paleomar-2 cruise.
During the Paleomar-2 cruise, the concentrations of Chl-a showed notable differences compared to those recorded in the Paleomar-1 cruise, particularly in the subsurface layers. At the surface, Chl-a values ranged from 0.13 to 4.63 mg m−3, with the highest concentrations found in the southern bay, specifically near San Juan de la Costa (Figure 9e). At 10 m depth, a similar distribution was observed, with peak values reaching 4.69 mg m−3 also in the San Juan de la Costa area (Figure 9f). At 30 m depth, Chl-a values varied from 0.21 to 2.36 mg m−3, again with the highest concentrations found in the southern region of the bay (Figure 9g). At 50 m depth, the maximum Chl-a value recorded was 1.70 mg m−3, displaying a more uniform distribution across the region (Figure 9h).
Comparing the two cruises, the concentrations of Chl-a were higher during the Paleomar-1 cruise, especially in the subsurface layers between 30 and 50 m depth.
4 Discussion
The period from 2014 to 2016 was characterized by significant dynamism, as reflected in the various ENSO phases. Initially, conditions were neutral until mid-2014, after which they transitioned into an extreme El Niño event that lasted until April 2016. Following this prolonged extreme El Niño, the conditions shifted to a La Niña event that continued through the second half of 2016.
The two periods analyzed in this study, El Niño and La Niña, had distinct impacts on the biochemical characteristics of the water column. For example, the hydrographic structure, as examined through the water masses (Figure 2), showed a significant intrusion of TSW during 2014 (Paleomar-1 cruise), while GCW predominated in 2016 (Paleomar-2 cruise). TSW is a water mass that occupies the upper 50 m layer and has a temperature >18.00 °C and a salinity <34.90. Its proportion in the Gulf of California and the Bay of La Paz demonstrates seasonal variability; however, its surface distribution is closely linked to wind patterns (Lavin and Marinone 2003). In this study, it was observed that wind intensity was greater in 2014 than 2016, which may explain the higher proportion of TSW recorded during that period.
The horizontal temperature distribution at different depths during both cruises showed significant differences (Figure 3). For example, during the Paleomar-2 cruise, which occurred under La Niña conditions, temperatures at the first three levels were notably higher than those recorded during the Paleomar-1 cruise, which took place during the El Niño phase. This increase in temperature during La Niña may be linked to the lingering effects of El Niño 2015/2016, which had a prolonged impact on the region. Consequently, the warmer conditions resulting from El Niño persisted until November 2016, despite the onset of the La Niña event. This observation highlights the substantial influence that El Niño 2015/2016 had on the region. The presence of this unusually warm water mass may explain the low concentrations of nutrients, particularly SRSi, and Chl-a observed during this period.
Regarding nutrient concentrations, there were significant differences between the two cruises. Specifically, the concentration of SRSi was markedly higher during the Paleomar-1 cruise (152.46 µM) compared to the Paleomar-2 cruise (8.48 µM). During the Paleomar-1 cruise, SRSi was distributed homogeneously. A similar trend was observed for other nutrients, including nitrates, nitrites, and SRP, all of which had higher concentrations during Paleomar-1 than Paleomar-2, especially at a depth of 30 m.
In this study, in November 2016, during a La Niña event, Chl-a concentrations were significantly lower compared to November 2014, which was under El Niño conditions. Previous research has indicated that Chl-a concentrations often decline during La Niña events compared to those during El Niño events. For example, in the Bali Strait of Indonesia, Chl-a levels typically decrease during La Niña events (Wijaya et al. 2020). Furthermore, Wilkerson et al. (2002) noted that the impacts of ENSO events in the Gulf of Farallones, California, may not be as pronounced as expected. They observed that during El Niño conditions, both nutrient concentrations and Chl-a levels are generally higher.
The results presented here contrast with previous studies conducted in the Bay of La Paz, which reported increased Chl-a values during La Niña events and a significant decrease during El Niño events (Herrera-Cervantes et al. 2020). However, it is important to consider the potential impact of the extreme El Niño event that affected the region from May 2015 to April 2016, as its long duration may help explain the lower concentrations of nutrients and Chl-a observed in November 2016.
El Niño during 2015/2016 was notably extreme. Evidence indicates that this event had a negative impact on phytoplankton biomass in various areas of the Mexican Pacific. For example, in the Gulf of Tehuantepec, a region recognized for its high Chl-a values due to its upwelling system, El Niño 2015/2016 led to unusually low Chl-a levels (<0.10 mg m−3) and an increase in sea surface temperatures of over 5 °C (Coria-Monter et al. 2019a). This situation reflects low nutrient availability. A similar trend was observed in this study, the Bay of La Paz experienced a prolonged warming period and low nutrient concentrations between May 2015 and April 2016 due to El Niño. This negatively impacted phytoplankton biomass. Although conditions transitioned to La Niña afterwards, the intensity of these conditions was not strong enough to cause the increases in Chl-a levels typically associated with La Niña events.
Many aspects of the nature and impact of ENSO events still need to be understood in several environments, both coastal and oceanic. In the case of the Bay of La Paz, there is limited knowledge about how these events affect the organisms at the base of the food chain, in this case the phytoplankton. To enhance our understanding of ENSO events, more comprehensive in situ observations are necessary. These observations should assess different hydrographic parameters and dynamics that could influence phytoplankton communities, which are crucial for supporting many commercially important pelagic fish species.
Funding source: Dirección General de Asuntos del Personal Académico, Universidad Nacional Autónoma de México
Award Identifier / Grant number: IA200120, IG100421 and IA200123
Award Identifier / Grant number: 144, 145 and 627
About the authors

Carlos Mauricio Torres-Martínez earned a Master’s degree from the National Autonomous University of Mexico (UNAM). He is currently a Ph.D. student in the graduate program of Marine Sciences and Limnology at UNAM, where his research focuses on the structure of phytoplankton communities and their relationship with hydrodynamic processes.

Dra. María Adela Monreal-Gómez earned her PhD in oceanology from the University of Liège (Belgium). Her interest in physical oceanography focuses on oceanic processes, such as vortices, fronts, and upwelling events, which emphasize the interactions between physics and biology.

Erik Coria-Monter has a Ph.D. in biological sciences from the Autonomous Metropolitan University (UAM) of Mexico and is Professor at the National Autonomous University of Mexico (UNAM). His research is focused on plankton ecology, trophic ecology, and physical/biological interactions in marine ecosystems.

Dr. David Alberto Salas-de-León obtained his PhD in oceanology at Liege University, Belgium, focusing on ocean models with a strong focus on physics-biology interactions. Currently, he is also studying non-linear systems and chaos.

Elizabeth Durán-Campos earned a Ph.D. in biological sciences (2015) from the Autonomous Metropolitan University of Mexico. She does research in plankton ecology and physical/biological interactions in both coastal and oceanic environments.
Acknowledgments
We would like to thank the captain and crew of R/V El Puma and students who assisted during the fieldwork. Sergio Castillo-Sandoval performed chemical analyses and Jorge Castro improved the quality of the figures. Helpful comments from one anonymous reviewer improved the manuscript.
-
Research ethics: Not applicable.
-
Informed consent: Not applicable.
-
Author contributions: Conceptualization: C.M.T.M., M.A.M.G., E.C.M., D.A.S.L. and E.D.C.; methodology: C.M.T.M., M.A.M.G., E.C.M., D.A.S.L. and E.D.C; software: C.M.T.M., M.A.M.G., E.C.M., D.A.S.L. and E.D.C; formal analysis: C.M.T.M., M.A.M.G., E.C.M., D.A.S.L. and E.D.C; investigation: C.M.T.M., M.A.M.G., E.C.M., D.A.S.L. and E.D.C; data curation: C.M.T.M., M.A.M.G., E.C.M., and E.D.C; writing – original draft preparation: C.M.T.M., M.A.M.G., E.C.M., D.A.S.L. and E.D.C; writing – review and editing: C.M.T.M., M.A.M.G., E.C.M., D.A.S.L. and E.D.C; supervision: M.A.M.G., E.C.M., D.A.S.L. and E.D.C; project administration: M.A.M.G., E.C.M., D.A.S.L. and E.D.C; funding acquisition: M.A.M.G., E.C.M., D.A.S.L. and E.D.C. The authors have accepted responsibility for the entire content of this manuscript and approved its submission. All authors have read and agreed to the published version of the manuscript.
-
Use of Large Language Models, AI and Machine Learning Tools: None declared.
-
Conflict of interest: The authors state no conflict of interest.
-
Research funding: This study was supported by the Institute of Marine Science and Limnology of the National Autonomous University of Mexico (ICML-UNAM, grants # 144, 145 and 627), and partially supported by DGAPA-PAPIIT-UNAM projects # IA200120, IG100421 and IA200123. The ship time for the research cruises on board R/V El Puma was funded by UNAM.
-
Data availability: The datasets generated during this study are available from the corresponding author on request.
References
Andrade, M.P., Magalhães, A., Pereira, L.C.C., Flores-Montes, M.J., Pardal, E.C., Andrade, T.P., and Costa, R.M. (2016). Effects of a La Niña event on hydrological patterns and copepod community structure in a shallow tropical estuary (Taperaçu, Northern Brazil). J. Mar. Syst. 164: 128–143.10.1016/j.jmarsys.2016.07.006Search in Google Scholar
Brierley, A.S. (2017). Plankton. Curr. Biol. 27: R478–R483, https://doi.org/10.1016/j.cub.2017.02.045.Search in Google Scholar PubMed
Coria-Monter, E., Monreal-Gómez, M.A., Salas de León, D.A., and Durán-Campos, E. (2017). Wind driven nutrient and chlorophyll-a enhancement at a cyclonic circulation in Bay of La Paz, Gulf of California, Mexico. Estuar. Coast. Shelf Sci. 196: 290–300.10.1016/j.ecss.2017.07.010Search in Google Scholar
Coria-Monter, E., Monreal-Gómez, M.A., Salas de Leon, D.A., and Durán-Campos, E. (2019b). Water masses and chlorophyll-a distribution in a semi-enclosed bay of the southern Gulf of California, Mexico, after the “Godzilla El Niño”. Arab. J. Geosci. 12: 473.10.1007/s12517-019-4636-1Search in Google Scholar
Coria-Monter, E., Monreal-Gómez, M.A., Salas-de-León, D.A., Aldeco-Ramírez, J., and Merino-Ibarra, M. (2014). Differential distribution of diatoms and dinoflagellates in a cyclonic eddy confined in the Bay of La Paz, Gulf of California. J. Geophys. Res. 119: 6258–6268, https://doi.org/10.1002/2014JC009916.Search in Google Scholar
Coria-Monter, E., Salas de León, D.A., Monreal-Gómez, M.A., and Durán-Campos. (2019a). Satellite observations of the effect of the “Godzilla El Niño” on the Tehuantepec upwelling system in the Mexican Pacific. Helgol. Mar. Res. 73: 3.10.1186/s10152-019-0525-ySearch in Google Scholar
Coria-Monter, E., Salas de Leon, D.A., Monreal-Gómez, M.A., and Durán-Campos, E. (2019c). Internal waves in the Bay of La Paz, southern Gulf of California, Mexico. Vie Milieu 69: 115–122.Search in Google Scholar
Dai, Y., Yang, S., Hu, C., Xu, W., Anderson, D.M., Li, Y., Song, X.-P., Boyce, D.G., Gibson, L., Zheng, C., et al.. (2023). Coastal phytoplankton blooms expand and intensify in the 21st century. Nature 615: 280–284, https://doi.org/10.1038/s41586-023-05760-y.Search in Google Scholar PubMed PubMed Central
Davies, C., Ajani, P., Armbrecht, L., Atkins, N., Baird, M.E., Beard, J., Bonham, P., Burford, M., Clementson, L., Coad, P., et al.. (2018). A database of chlorophyll-a in Australian waters. Sci. Data 5: 180018, https://doi.org/10.1038/sdata.2018.18.Search in Google Scholar PubMed PubMed Central
Durán-Campos, E., Monreal-Gómez, M.A., Salas de León, D.A., and Coria-Monter, E. (2020). Field and satellite observations on the seasonal variability of the surface chlorophyll-a in the Bay of La Paz, Gulf of California, Mexico. IJOO 14: 157–167, https://doi.org/10.37622/ijoo/14.1.2020.157-167.Search in Google Scholar
Falkowski, P. (2012). Ocean science: the power of plankton. Nature 483: 17–20, https://doi.org/10.1038/483s17a.Search in Google Scholar
García, E. (1989). Apuntes de climatología. Universidad Nacional Autónoma de México, Ciudad de México.Search in Google Scholar
Gaube, P., McGillicuddy, D.J., Chelton, D.B., Behrenfeld, M.J., and Strutton, P.G. (2014). Regional variations in the influence of mesoscale eddies on near-surface chlorophyll. J. Geophys. Res. 119: 8195–8220, https://doi.org/10.1002/2014jc010111.Search in Google Scholar
Grasshoff, K., Ehrhardt, M., and Kremling, K. (1983). Methods of seawater analysis, 2nd ed. Verlag Chemie, Weinheim.Search in Google Scholar
Herrera-Cervantes, H., Lluch-Cota, S., Cortés-Ramos, J., Farfan, L., and Morales-Aspeitia, R. (2020). Interannual variability of surface satellite-derived chlorophyll concentration in the bay of La Paz, Mexico, during 2003–2018 period: the ENSO signature. Cont. Shelf Res. 209: 104254.10.1016/j.csr.2020.104254Search in Google Scholar
Herrera-Cervantes, H., Lluch-Cota, S., Lluch-Cota, D., Gutiérrez de Velasco-Sanromán, G., and Lluch-Belda, D. (2010). ENSO influence on satellite-derived chlorophyll trends in the Gulf of California. Atmósfera 23: 253–262.Search in Google Scholar
Kirkwood, D.S. (1994). SanPlus segmented flow analyzer and its applications. Seawater analysis. Skalar, Amsterdam.Search in Google Scholar
Lavín, M.F., Castro, R., Beier, E., Godínez, V.M., Amador, A., and Guest, P. (2009). SST, thermohaline structure, and circulation in the southern Gulf of California in June 2004 during the North American monsoon experiment. J. Geophys. Res. Oceans 114: C02025, https://doi.org/10.1029/2008jc004896.Search in Google Scholar
Lavin, M.F. and Marinone, S.G. (2003). An overview of the physical oceanography of the Gulf of California. In: Velasco Fuentes, O.U., Sheinbaum, J., and Ochoa, J. (Eds.), Nonlinear processes in geophysical fluid dynamics. Springer Dordrecht, Netherlands, pp. 173–204.10.1007/978-94-010-0074-1_11Search in Google Scholar
Lévy, M., Franks, P.J.S., and Smith, K.S. (2018). The role of submesoscale currents in structuring marine ecosystems. Nat. Commun. 9: 4758, https://doi.org/10.1038/s41467-018-07059-3.Search in Google Scholar PubMed PubMed Central
Machado, I., Barreiro, M., and Calliari, D. (2013). Variability of chlorophyll-a in the Southwestern Atlantic from satellite images: seasonal cycle and ENSO influences. Cont. Shelf Res. 53: 102–109, https://doi.org/10.1016/j.csr.2012.11.014.Search in Google Scholar
McGillicuddyJr.D.J. (2016). Mechanisms of physical-biological-biogeochemical interaction at the oceanic mesoscale. Ann. Rev. Mar. Sci. 8: 125–159, https://doi.org/10.1146/annurev-marine-010814-015606.Search in Google Scholar PubMed
McPhaden, M.J., Santoso, S., and Cai, W. (2021). Introduction to El Niño southern oscillation in a changing climate. In: McPhaden, M.J., Santoso, A., and Cai, W. (Eds.). El Niño southern oscillation in a changing climate. American Geophysical Union, Geophysical Monograph Series, USA, pp. 1–19.10.1002/9781119548164.ch1Search in Google Scholar
Molina-Cruz, A., Pérez-Cruz, L., and Monreal-Gómez, M.A. (2002). Laminated sediments in the Bay of La Paz, Gulf of California: a depositional cycle regulated by pluvial flux. Sedimentology 49: 1401–1410, https://doi.org/10.1046/j.1365-3091.2002.00505.x.Search in Google Scholar
Monreal-Gómez, M.A., Molina-Cruz, A., and Salas de León, D.A. (2001). Water masses and cyclonic circulation in Bay of La Paz, Gulf of California, during June 1998. J. Mar. Syst. 30: 305–315, https://doi.org/10.1016/s0924-7963(01)00064-1.Search in Google Scholar
Richardson, A.J. (2008). In hot water: zooplankton and climate change. ICES J. Mar. Sci. 65: 279–295, https://doi.org/10.1093/icesjms/fsn028.Search in Google Scholar
Roesler, C., Uitz, J., Claustre, H., Boss, E., Xing, X., Organelli, E., Briggs, N., Bricaud, A., Schmechtig, C., Poteaaus, A., et al.. (2017). Recommendations for obtaining unbiased chlorophyll estimates from in situ chlorophyll fluorometers: a global analysis of WET Labs ECO sensors. L&O Methods 15: 572–585, https://doi.org/10.1002/lom3.10185.Search in Google Scholar
Roy, S., Llewellyn, C.A., Egeland, E.S., and Johnsen, G. (2011). Phytoplankton pigments: characterization, chemotaxonomy and applications in oceanography. Cambridge University Press, Cambridge.10.1017/CBO9780511732263Search in Google Scholar
Schiermeier, Q. (2015). Hunting the Godzilla El Niño. Nature 526: 490–491, https://doi.org/10.1038/526490a.Search in Google Scholar PubMed
Shafeeque, M., Balchand, A.N., Shah, P., George, G., Smitha, B.R., Varghese, E., Joseph, A.K., Sathyendranath, S., and Platt, T. (2021). Spatio-temporal variability of chlorophyll-a in response to coastal upwelling and mesoscale eddies in the south eastern Arabian Sea. Int. J. Remote Sens. 42: 4840–4867, https://doi.org/10.1080/01431161.2021.1899329.Search in Google Scholar
Sosa-Ávalos, R., Santamaría-del-Ángel, E., Acosta-Chamorro, V., Silva-Iñiguez, L., Pelayo-Martínez, G.C., and Quijano-Scheggia, S.I. (2021). Phytoplankton primary production during the cold and warm seasons in Manzanillo and Santiago Bays, Mexico. Estuar. Coast. Shelf Sci. 261: 107569, https://doi.org/10.1016/j.ecss.2021.107569.Search in Google Scholar
Strickland, J.D.H. and Parsons, T.R. (1972). A practical handbook of seawater analysis. Ottawa Fisheries Research Board of Canada, Bulletin 167, Ottawa, Canada.Search in Google Scholar
Villamaña, M., Mouriño-Carballido, B., Maranon, E., Cermeño, P., Chouciño, P., da Silva, J.C.B., Díaz, P.A., Fernández-Castro, B., Gilcoto, M., Graña, R., et al.. (2017). Role of internal waves on mixing, nutrient supply and phytoplankton community structure during spring and neap tides in the upwelling ecosystem of Ría de Vigo (NW Iberian Peninsula). Limnol. Oceanogr. 62: 1014–1030, https://doi.org/10.1002/lno.10482.Search in Google Scholar
Wijaya, A., Zakiyah, U., Sambah, A.B., and Setyohadi, D. (2020). Spatio-temporal variability of temperature and chlorophyll-a concentration of sea surface in Bali Strait, Indonesia. Biodiv 21: 5283–5290, https://doi.org/10.13057/biodiv/d211132.Search in Google Scholar
Wilkerson, F.P., Dudgale, R.C., Marchi, A., and Collins, C.A. (2002). Hydrography, nutrients and chlorophyll during El Niño and La Niña 1997–1998 winters in the Gulf of Farallones, California. Prog. Oceanogr. 54: 293–310.10.1016/S0079-6611(02)00055-1Search in Google Scholar
Xiu, P., Dai, M., Chai, F., Zhuo, K., Zeng, L., and Du, C. (2018). On contributions by wind-induced mixing and eddy pumping to interannual chlorophyll variability during different ENSO phases in the northern South China Sea. Limnol. Oceanogr. 64: 503–514, https://doi.org/10.1002/lno.11055.Search in Google Scholar
Zhang, H., Qiu, Z., Sun, D., Wang, S., and He, Y. (2017). Seasonal and interannual variability of satellite-derived chlorophyll-a (2000–2012) in the Bohai Sea, China. Remote Sens. 9: 582, https://doi.org/10.3390/rs9060582.Search in Google Scholar
Supplementary Material
This article contains supplementary material (https://doi.org/10.1515/bot-2025-0007).
© 2025 the author(s), published by De Gruyter, Berlin/Boston
This work is licensed under the Creative Commons Attribution 4.0 International License.
Articles in the same Issue
- Frontmatter
- In this issue
- Physiology and Ecology
- Three-dimensional distribution of nutrients and phytoplankton biomass in a semi-enclosed region of the Gulf of California during different ENSO phases
- Proliferation of Undaria pinnatifida along the Atlantic coast of the Iberian Peninsula
- Study on fatty acid methyl ester (FAME) composition of 19 Brazilian thraustochytrid isolates
- Taxonomy/Phylogeny and Biogeography
- Variations in the bacterial and fungal community structure along the hypoxic gradient of the Arabian Sea oxygen-depleted environment based on eDNA metabarcoding analysis
- Studies of North Carolina marine algae XV. DNA sequencing reveals some different Ulva species compared to historical reports and U. carsoniae sp. nov. (Ulvales, Chlorophyta)
- Identification and growth of Prorocentrum cf. cassubicum (Dinoflagellata: Prorocentraceae) from Bahía de La Paz: first record for the Gulf of California
- A new oomycete pathogen Olpidiopsis dasysiphoniae sp. nov. (Oomycota) infecting the red alga Dasysiphonia japonica (Ceramiales, Delesseriaceae)
- Chemistry and Applications
- Exploring the suitable extractive species in an IMTA: inorganic nutrient removal from mariculture effluents by commercially important marine macroalgae
Articles in the same Issue
- Frontmatter
- In this issue
- Physiology and Ecology
- Three-dimensional distribution of nutrients and phytoplankton biomass in a semi-enclosed region of the Gulf of California during different ENSO phases
- Proliferation of Undaria pinnatifida along the Atlantic coast of the Iberian Peninsula
- Study on fatty acid methyl ester (FAME) composition of 19 Brazilian thraustochytrid isolates
- Taxonomy/Phylogeny and Biogeography
- Variations in the bacterial and fungal community structure along the hypoxic gradient of the Arabian Sea oxygen-depleted environment based on eDNA metabarcoding analysis
- Studies of North Carolina marine algae XV. DNA sequencing reveals some different Ulva species compared to historical reports and U. carsoniae sp. nov. (Ulvales, Chlorophyta)
- Identification and growth of Prorocentrum cf. cassubicum (Dinoflagellata: Prorocentraceae) from Bahía de La Paz: first record for the Gulf of California
- A new oomycete pathogen Olpidiopsis dasysiphoniae sp. nov. (Oomycota) infecting the red alga Dasysiphonia japonica (Ceramiales, Delesseriaceae)
- Chemistry and Applications
- Exploring the suitable extractive species in an IMTA: inorganic nutrient removal from mariculture effluents by commercially important marine macroalgae