Proliferation of Undaria pinnatifida along the Atlantic coast of the Iberian Peninsula
-
Mar Humet holds a Bachelor’s degree in Environmental Sciences from the University of Barcelona and is currently pursuing a Master’s degree in Bioinformatics and Biostatistics at the Open University of Catalonia. She works as a research technician with a focus on rocky shore biodiversity and the physiological responses of intertidal invertebrates to thermal stress, and she is also actively engaged in citizen science activities.
 ,
,
Joana Pereira holds a Master’s degree in Biodiversity, Genetics and Evolution from the University of Porto and is currently a PhD student at the BIODIV doctoral program at CIBIO. Her PhD thesis focuses on understanding the mechanistic links between climate change and coastal biodiversity.
João Nunes holds a Master’s degree in Marine Sciences from the University of Porto. His research focuses on intertidal biodiversity monitoring using UAS (drones) and automated image classification. He has contributed to studies on rocky shore communities and the impacts of environmental changes. Additionally, he has been involved in citizen science initiatives and ocean conservation projects.
Rocío Nieto-Vilela holds a PhD in Biological Sciences with a marine focus from the Universidad de la Patagonia San Juan Bosco, Argentina, which she completed in 2020. Following her doctorate, she undertook a postdoctoral research position (2020–2023) focused on the physiological effects caused by environmental contaminants. Since then, she has been working as a researcher at CIBIO/BIOPOLIS, contributing to the ANERIS project through biodiversity data analysis, environmental DNA research, and the coordination of citizen science initiatives.
Ana Sinde-Mano is a marine scientist fascinated by the biodiversity and ecology of rocky coastal communities. She holds a Master’s degree in Marine Biology and Conservation from the Applied Psychology Institute in Lisbon, Portugal. Currently, she is working on identifying and mapping the distribution of intertidal species to make temporal comparisons and understand shifts in these communities due to climate change.


Abstract
The spread of non-indigenous species is a critical concern for marine ecosystems, particularly in regions with high biodiversity and economic reliance on coastal resources. This study investigates the distribution, abundance, and expansion of the invasive kelp Undaria pinnatifida along the Atlantic coast of the Iberian Peninsula. A comprehensive survey was conducted in 2024, covering 59 locations across Spain and Portugal. This dataset was complemented by biodiversity records from citizen science platforms. Historical records were also reviewed to assess changes in its distribution over time. Results indicate that U. pinnatifida has notably increased its presence in Portugal since its first record, now occurring at 16 sites (compared to only 2 in 2007), while remaining well-established in Galicia. However, the species co-occurs with native kelps and, at this stage, does not dominate the communities where it has established itself. Furthermore, it remains absent from southern Iberia and the Bay of Biscay. Hence, environmental conditions and biotic interactions might be shaping their distribution, which warrants further investigation. Given the ongoing environmental changes and human-mediated dispersal, continuous monitoring is essential to track future range shifts. This study also showcases how integrating citizen science data can enhance the spatial and temporal resolution of non-indigenous species monitoring.
1 Introduction
Marine non-indigenous species (NIS) (Richardson et al. 2011) spread beyond their natural range, mainly transported by maritime traffic and marine aquaculture (Hewitt et al. 2007; Schaffelke et al. 2006). NIS are known as “invasive species” when they establish, spread, and proliferate without direct human assistance (according to Mack et al. 2000) or alter the environment (according to Simberloff et al. 2013). Invasive alien species have contributed solely or alongside other drivers to 60 % of recorded global extinctions, and are the only driver in 16 % of the documented global animal and plant extinctions (Roy et al. 2023) by outcompeting native taxa and leading to considerable negative economic impacts (Brondizio et al. 2019; Dudeque Zenni et al. 2021; McKnight et al. 2021).
Intertidal macroalgae play an important role in coastal ecosystems, acting as primary producers and providing habitat and nursery grounds for a wide variety of species, many of which are ecological and economically relevant (Steneck et al. 2002). Their canopies offer a buffer against the harsh physical conditions of the intertidal, helping to maintain ecological balance and support the resilience of coastal ecosystems (Mineur et al. 2015; Thomsen et al. 2012). Nevertheless, some invasive marine macroalgae can disrupt coastal ecosystems, driving ecological change (Çinar et al. 2014). For example, the alteration of competitive relationships within the recipient habitat, often manifested as space monopolization, leads to significant reductions in both the abundance and diversity of native macroalgae and associated fauna. Davidson et al. (2015) showed that, in numerous instances, areas affected by invasive species exhibited a marked decline in the richness and abundance of native macroalgal species compared to non-invaded areas. However, the impacts of invasive species are often species- and region-specific (Epstein and Smale 2017).
Undaria pinnatifida (Harvey) Suringar, commonly known as “Wakame”, is a cold-temperate kelp native to the northwest Pacific Ocean (coastlines of Japan, Korea, Russia, and China) (Epstein and Smale 2017). U. pinnatifida is one of the 100 most invasive species in the world according to the Invasive Species Specialist Group list (Lowe et al. 2000). Nowadays, this kelp is present in nearly all temperate coastal marine ecoregions (Spalding et al. 2007), including the northeast Atlantic, southwest Atlantic, New Zealand, Australia, and the northeast Pacific (Heiser et al. 2014; James et al. 2015; Raffo et al. 2009; Thornber et al. 2004). Due to its photosynthetic adaptability and ability to grow, this species can extend its distribution into deeper subtidal habitats, enhancing its ecological impact across a broader range of marine environments (Desmond et al. 2019; Russell et al. 2008). It has been suggested that U. pinnatifida might act as a passenger rather than a driver of ecological change (sensu) (MacDougall and Turkington 2005), as it colonizes areas that have lost their natural canopy cover (South and Thomsen 2016). Hence, it typically establishes itself first on artificial substrata (Kaplanis et al. 2016) or on disturbed natural substrata with little competition. In a second phase, it often spreads to nearby natural rocky substrata, where the effects on native communities are region-specific, ranging from little impact (e.g., De Leij et al. 2017) to a substantial decrease in local diversity. For example, in Argentina, the presence of Undaria resulted in a decrease in native seaweed diversity and richness (Casas et al. 2004); in Italy, a decrease in the surface area of native seaweeds was observed following Undaria’s establishment (Curiel et al. 2002) and, in New Zealand, there was a reduction in the abundance of epifauna with the spread of Undaria (Suárez-Jiménez et al. 2017).
In the Northeast Atlantic, U. pinnatifida has its southern limit in central Portugal (Pereira et al. 2022) and its northern limit in the island of Sylt, Germany (Schiller et al. 2018). U. pinnatifida was first recorded in Europe in the Thau lagoon on the French Mediterranean coast in 1971, probably as the result of the import of oysters from Japan (Boudouresque et al. 1985; Floc’h et al. 1991). Then it was transferred for farming to northern France in 1983 and, in 1988, reproducing individuals were found growing on mussel lines outside the seaweed farm (Floc’h et al. 1991, 1996).
The species was first recorded in the Iberian Peninsula in Galicia in 1988, probably introduced via oysters imported there (Caamano et al. 1990). In Spain, U. pinnatifida has extended its distribution to various regions, including Asturias and Cantabria, as documented by Peteiro (2008), Pérez Ruzafa et al. (2002) and Salinas et al. (1996). Ugarte et al. (2006) provided a detailed review of its distribution, confirming its expansion in these regions.
In Portugal, U. pinnatifida was first recorded in 2007 by Araújo et al. (2009) at a marina in Póvoa de Varzim (northern Portugal) and in the Ría de Aveiro (central Portugal). Later, Veiga et al. (2014) reported the disappearance of the Aveiro population and documented a new one in Buarcos (central Portugal). More recently, Monteiro et al. (2022) documented the presence of U. pinnatifida at four new sites – two on artificial substrata and two on natural substrata. A study by Carreira-Flores et al. (2023) did not identify any additional locations in mainland Portugal.
The Atlantic coast of the Iberian Peninsula exhibits unique ecological conditions. The nutrient-rich waters and pronounced latitudinal thermal gradient driven by the cooling effect of the Canary upwelling system during summer create a biogeographical transition zone in northern Portugal (Fiuza 1983). This area serves as the southern range limit for various cold-temperate species and as a biogeographical poleward barrier for warmer-water species (Casado-Amezúa et al. 2019; Monteiro et al. 2022). Modelling efforts to predict the fate of the distribution of U. pinnatifida in the region give mixed results, ranging from the identification of favorable conditions for the expansion of U. pinnatifida (Báez et al. 2010), to a low or very low probability of establishment outside the Galician rías (Blanco et al. 2021). Nevertheless, recent distribution shifts in northern Portugal of both native and non-indigenous intertidal and subtidal seaweeds (de Azevedo et al. 2023; Monteiro et al. 2022), highlight the importance of closely tracking the expansion of non-native species and its potential impacts on coastal communities.
This paper describes the expansion of U. pinnatifida along the Atlantic coastlines of the Iberian Peninsula since its initial establishment. It also updates the presence and absence of this species based on recent survey data (summer 2024).
2 Materials and methods
2.1 Data collection
Fieldwork was conducted from February to November of 2024 to monitor the distribution and abundance of the kelp U. pinnatifida in the intertidal zone along the Atlantic coast of the Iberian Peninsula, from the Strait of Gibraltar to the northeast Spanish border with France. Survey sites were selected to ensure broad geographical coverage, focusing on areas where kelp populations were previously known to occur (Figure 1).

Distribution of Undaria pinnatifida before 2024. Red triangle represents the first record within the Atlantic coast of the Iberian Peninsula. Blue points represent presences between 1990 and 2000; green points between 2000 and 2010; yellow points between 2010 and 2020, and purple points between 2020 and 2023.
At each site, a team of 2 people surveyed the rocky shore for 2 h during low tides employing an ad-libitum search focused on the low intertidal zone where U. pinnatifida is typically found. This method enhances the detection of rare species such as an invasive species at its early stages (Monteiro et al. 2022). Furthermore, the SACFOR abundance scale (Burrows et al. 2008; Hiscock 1981) was used to estimate the abundance of the U. pinnatifida populations, providing a standardized assessment for future comparisons. This scale grades abundances as 6: Superabundant (more than 90 % cover), 5: Abundant (60–90 % cover), 4: Common (widespread on the shore, 30–59 %), 3: Frequent (patches apparent, up to 30 % cover), 2: Occasional (3–20 individuals scattered) and 1: Rare (only 1 or 2 individuals). The scale was applied in each site, except for those records obtained from citizen science monitoring platforms. Only the locations where U. pinnatifida was found attached to the substratum were counted as confirmed presence, as specimens floating in the water or lying on the sand could have arrived through other means of transport and were, therefore, not considered established populations.
2.2 Citizen science platforms
We also mined two biodiversity monitoring programs, MINKA (https://minka-sdg.org/) and iNaturalist (https://www.inaturalist.org/) for records of U. pinnatifida in the Atlantic Iberian Peninsula. These platforms allow for the collection of observations from a wide range of contributors, helping to address potential geographical gaps in our dataset. Observations were limited to the geographical distribution within continental Portugal and the Atlantic coast of Spain, excluding any data beyond the borders of the study area (i.e., in the Atlantic coast of France or in the Mediterranean Sea). Both platforms employ a curator system to ensure the accuracy and reliability of records, hence only records with “research grade” were kept. Additionally, data from these platforms were carefully reviewed for accuracy. Only observations documented with pictures of the specimen visibly attached to the substratum were used for the analysis.
2.3 Historical distribution
To compare with the present survey, we conducted a comprehensive review of previous U. pinnatifida records along the intertidal Atlantic coast of the Iberian Peninsula, from the first record in 1988 to January 2025. We performed a systematic literature search using popular scientific search engines, namely Web of Science (WoS) and Google Scholar with the keywords “U. pinnatifida”, “Iberian Peninsula” and “Península Iberica” for Portuguese and Spanish references, as well as species distribution databases such as GBIF and WoRMS. Only studies that provided records of the species’ geographical distribution – including location and year/date of observation – were included, while articles focused on other aspects, e.g. aquaculture, were excluded. We then carefully document the location and date of its first record at each site. In some cases, approximate locations were used due to a lack of precise geographic details in the literature.
2.4 Distribution and abundance – maps
Data analysis was conducted using R Statistical Software (R Development Core Team 2024). We generated distribution maps of the year 2024 and the temporal range from 1988 to 2023 to assess the expansion of U. pinnatifida using ‘ggplot2’ (Wickham 2016), ‘sf’ (Pebesma and Bivand 2023), ‘ggrepel’ (Slowikowski 2024), ‘giscoR’ (Hernangómez 2020) and ‘rnaturalearth’ (Massicotte and South 2023) R packages.
3 Results
3.1 Historical distribution
After compiling all available data on the presence of U. pinnatifida along the Atlantic coast of the Iberian Peninsula, our database showed that the species had been found in 108 locations, 94 in Spain and 14 in Portugal (Supplementary Table S1). To facilitate interpretation, data were divided into four distinct periods: (1) from the late 1980s to 2000, (2) from 2000 to 2010, (3) from 2010 to 2020, and (4) from 2020 to 2023 (Figure 1).
In the first period (1980s–2000), U. pinnatifida was found in 18 locations in Galicia and one site in Asturias, but there were no records in Portugal at that time. During the second period (2000–2010), it was found in 35 locations in Spain, and it was recorded for the first time in Portugal at 2 sites. By the third period (2010–2020), it had been observed in 27 locations in Galicia and 3 locations in central Portugal. Finally, prior to this study (2020–2023), U. pinnatifida was reported in 13 locations in Galicia and 9 locations in Portugal, expanding to the north.
To construct the historical distribution map (Figure 1), we used data from seven published references and seven citizen science observations. Of these references, Araújo et al. (2009) and Veiga et al. (2014) provided records exclusively for Portugal, while Blanco et al. (2021), Caamano et al. (1990), Carreira-Flores et al. (2023), Pérez Ruzafa et al. (2002), Peteiro (2008), Salinas et al. (1996) and Ugarte et al. (2006) contained data points only for Spain. Additionally, Pereira et al. (2022) included distribution records from both countries. Between 2010 and 2020, only one study in the Iberian Peninsula reported Undaria’s distribution.
3.2 Present survey (2024)
The survey carried out in 2024 covered a total of 60 sites, 35 in Spain and 25 in Portugal (Supplementary Table S2 and GBIF – https://doi.org/10.15468/sd2kdv). Of these, U. pinnatifida was found in a total of 31 locations, 15 in Spain and 16 in Portugal (Figure 2). During this survey U. pinnatifida was recorded in four new sites in Spain (Espasante, Punta Arroás, Praia das Cabanas, and A Guarda) and in six new sites in Portugal (Montedor, Viana do Castelo, Praia da Amorosa, Mindelo, Angeiras, and Praia do Homem do Leme). U. pinnatifida was not found in high abundances; it was recorded as rare in five sites (corresponding to an abundance level of 2) and observed as frequent at nine sites (corresponding to an abundance level of 3). All sites visited are urban shores, very close to human settlements. As reported in several other studies, we found Undaria near harbours, marinas and pontoons (3 sites) and often in natural rocky shores near man-made structures (10 sites). An overview of these shores can be found in Figure S1.
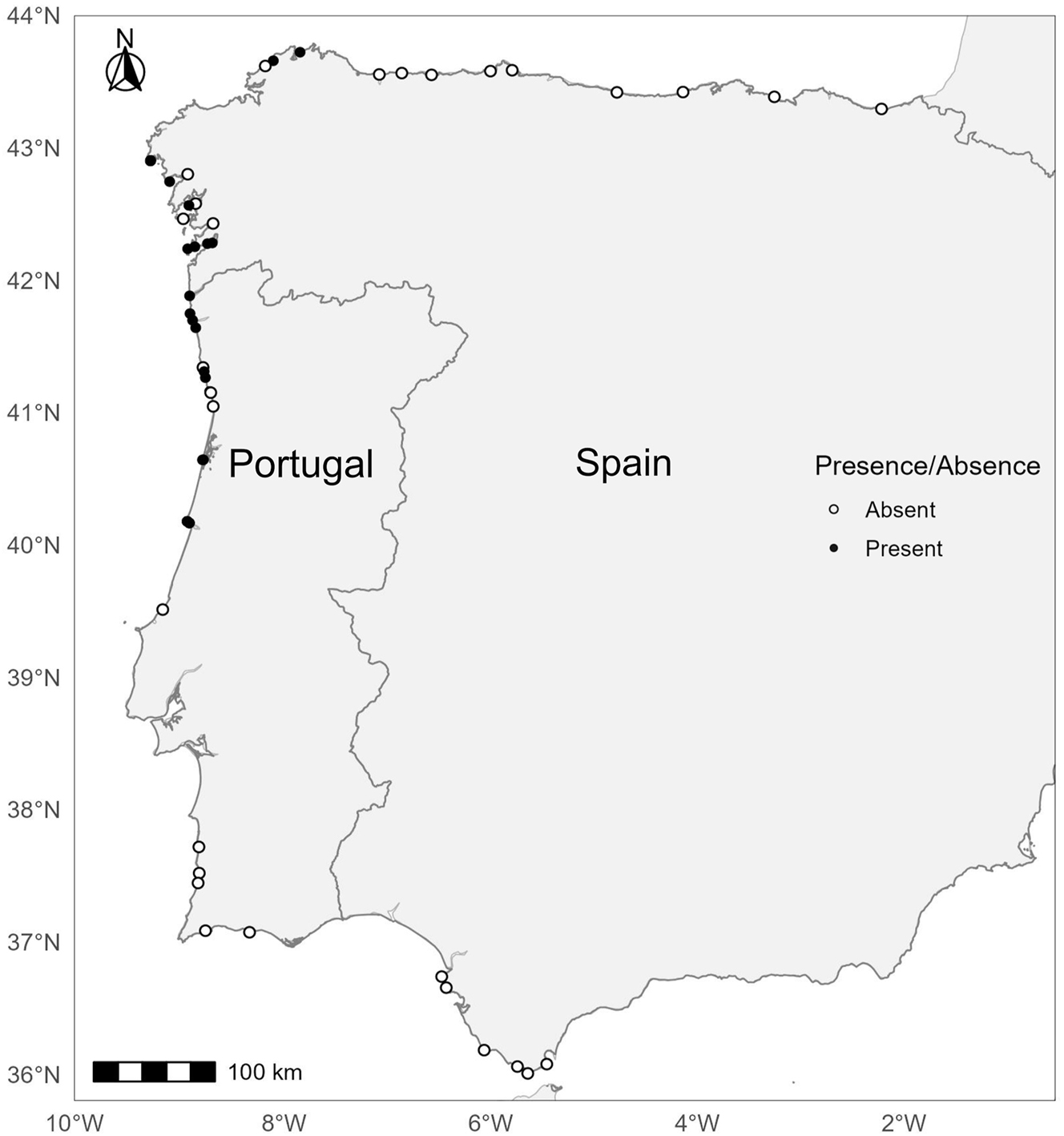
2024 survey of Undaria pinnatifida in the Iberian Península. Black circles indicate presence, while white circles indicate absence.
In total, 24 observations came from citizen science biodiversity platforms, MINKA and iNaturalist, contributed by 15 different observers, spanning from 2016 to 2024. We identified a geographic bias in the data, with most Portuguese observations concentrated in Buarcos and Aveiro, while fewer records were available from northern areas. In Spain, records were mainly localized within rías, with very few observations from the A Coruña area.
The current distribution limits of U. pinnatifida in the Iberian Peninsula are Buarcos (central Portugal, 40° 09′ 58.6ʺ N; 8° 53′ 08.0ʺ W) and Burela (Lugo, Spain, 43° 40′ 17.2ʺ N; 7° 21′ 28.9ʺ W) observed for the first time in 2013 and 2017, respectively. Several sites were surveyed south of Buarcos and east of Lugo, but no specimens were found (Figure 2).
Undaria pinnatifida was observed in the low intertidal zone, commonly alongside other kelp species such as Saccorhiza polyschides and Laminaria spp. (Figure 3). It frequently hosted epibionts such as the gastropods Steromphala spp. and Patella pellucida. Overall, individuals ranged in size from 0.5 m to approximately 1.5 m in length during its maximum size peak in early summer. Sporophytes of U. pinnatifida were recorded throughout the year, with the largest specimens observed in spring, while by the end of summer, blade senescence was common.

Specimens of Undaria pinnatifida found in: (A) Homem do Leme, Portugal (sourced from MINKA, ©juliana09); (B) Praia Norte, Viana do Castelo, Portugal (M. Humet); (C) Muros, Spain (A. Sinde-Mano); (D) Ilha de Arousa, Spain (sourced from iNaturalist, ©javsagg34).
4 Discussion
Our results indicate that U. pinnatifida has (i) proliferated along the Portuguese coast, increasing its distribution from 2 sites in 2007 to 16 sites in 2024, while (ii) persisting in Galicia, where it was first recorded in 1988 and it has been reported in 15 locations in the 2024 survey. The 2024 survey documented Undaria in 10 new sites across Portugal and Spain (out of 31 sites where Undaria was recorded and out of 60 sites surveyed). This pattern aligns with previous predictions that identified the northwest of the Iberian Peninsula as a suitable habitat for the species (Báez et al. 2010). However, its spread and establishment are not uniform throughout the Iberian Peninsula. Populations of U. pinnatifida in the Lower Rías were the first to establish in the Iberian Peninsula, and their abundances are higher there than in the Upper Rías and northern Portugal (this study; Pereira et al. 2022). In Portugal, although northern shores are geographically closer to the introduction site in Galicia (Ria de Arousa), U. pinnatifida was first established in Buarcos (central Portugal), located 255 km from the point of the first introduction in Ria de Arousa. It was only reported in northern Portugal (Póvoa de Varzim) in 2007.
The species was mainly observed co-occurring with native kelps, which supports the hypothesis that it may act as a passenger of ecological change, occupying newly available niches as conditions shift (De Leij et al. 2017). South et al. (2017) highlights that U. pinnatifida’s success as an invasive species is related to its ability to exploit disturbed habitats and to its effective dispersal mechanisms. Additionally, De Leij et al. (2017) emphasize that the establishment and persistence of U. pinnatifida in natural habitats are negatively correlated with the presence of native macroalgal canopies. Dense canopies of native kelp species in the Iberian Peninsula, such as Saccorhiza polyschides and Laminaria spp. (e.g. Boaventura et al. 2002; Casado-Amezúa et al. 2019), can limit the availability of light and space, restricting the colonization and growth of U. pinnatifida (De Leij et al. 2017; South et al. 2017). This apparent lower competitive ability might explain why Undaria does not dominate any of the sites where it has established itself in NW Iberia and that it has spread slowly at its northeastern limit. Apart from competition, other local environmental factors, such as temperature and nutrient availability could be influencing or even determining some of this regional heterogeneity (Gao et al. 2013; Thornber et al. 2004). Although these factors may currently limit its expansion, U. pinnatifida has proven its ability to overcome such constraints (Russell et al. 2008) and today it comprises 70 % of the subtidal seaweed biomass in Otago, New Zealand (Jiménez et al. 2015). This highlights the importance of sustained monitoring efforts in newly invaded areas.
In Asturias, U. pinnatifida was not found in 2024, even though the species was previously reported there between 2002 and 2008. In Portugal, a population in the Ria de Aveiro was first reported in 2007, not observed in 2013, then reported again in 2019 from a citizen science platform, and then again in 2021 (Pereira et al. 2022). These fluctuations indicate that these populations are most likely poorly established and subjected to seasonal or annual fluctuations in abundance. Caution is warranted, though, as these results were not collected in a standardized, consistent way (different research teams and methods) and geographical and temporal gaps exist – ours is the first study to cover the Iberian Peninsula in the same year. For example, in Spain, there is a gap of eight years (2008–2016) where there is no data about the occurrence of Undaria, and there is more information about the distribution in Lower Rías than in Upper Rías.
Given that the species is at its warmer-edge limit in Europe, temperature might be playing a key role in the study area. We show that U. pinnatifida is neither in the south of Portugal nor in the Bay of Biscay (northeast Spain). This is likely due to the specie’s temperature requirements, as it is a temperate kelp with upper thermal limits of around 20 °C for year-round populations and 29 °C for annual (winter) populations (James et al. 2015). The south of Portugal and the Bay of Biscay experience higher summer temperatures than Galicia and northern Portugal (Lima et al. 2007; Ramos et al. 2016). At the same time, the colder, nutrient-rich waters surfaced by the NW Iberian upwelling appear to provide an optimal environment for U. pinnatifida to establish and persist (Gómez-Gesteira et al. 2008). Further studies should explore which life-cycle phases and temperature metrics limit its expansion in the Iberian Peninsula.
Like U. pinnatifida, several non-native species have also recently colonized and spread in northern Portugal and Spain, namely Asparagopsis armata, Codium fragile and various bryozoans (Ramos et al. 2020). Given this, along with the pressure of maritime traffic in the area (James and Shears 2016), U. pinnatifida is expected to continue expanding its distribution. Furthermore, considering the decline in native kelp communities observed at different sites (Arriaga et al. 2024; Pereira et al. 2022; Ramos et al. 2020), large U. pinnatifida populations will likely establish in the future. This calls for intensive spatial and temporal monitoring and early intervention by managing bodies since eradication measures are costly and often have proven unsuccessful elsewhere (e.g. Forrest and Blakemore 2006; Hewitt et al. 2005). Moreover, the impacts of U. pinnatifida populations on biodiversity and ecosystem functioning have not been assessed in this region; therefore, we cannot accurately determine its invasion status on the Iberian Peninsula. Field studies are urgently needed to address this outstanding question.
The integration of observations from citizen science biodiversity monitoring programs provided 24 extra observations, expanding the geographical coverage of our survey. It confirmed the continued presence of U. pinnatifida in previously documented locations not visited during our fieldwork. This highlights the valuable role of public participation in biodiversity and biogeography research, providing complementary data that can enhance monitoring efforts, especially for non-native species (Encarnação et al. 2021). Citizen science initiatives increase temporal and spatial resolution and contribute to early detection and long-term tracking of species distribution (Kelly et al. 2020).
Overall, our findings contribute to better understand the distribution range and temporal dynamics of a non-indigenous species along the Atlantic coast of the Iberian Peninsula. While U. pinnatifida remains restricted to the Galician rias and north Portugal, ongoing environmental changes and human-mediated dispersal may influence its future spread. Further research is needed on the interactions of U. pinnatifida with native macroalgal communities, as well as its physiological response to environmental fluctuations such as potential changes in the upwelling system. Additionally, by integrating field-based studies with citizen science initiatives, we can improve early detection and track long-term trends to help develop effective conservation and mitigation measures for non-indigenous species in the region.
Funding source: FCT through a PhD grant
Award Identifier / Grant number: 2022.12126.BD
Funding source: ANERIS
Award Identifier / Grant number: 101094924
Funding source: LifeWatch ERIC
Award Identifier / Grant number: 101059548
Funding source: CEECInd contracts
Award Identifier / Grant number: 10.54499/CEECIND/03185/2018/CP1546/CP1648/CT0004
Funding source: BIOINTERACT
Award Identifier / Grant number: 10.54499/2022.02887.PTDC
Funding source: OceanLog
Award Identifier / Grant number: PTDC/BIA-BMA/4848/202
About the authors

Mar Humet holds a Bachelor’s degree in Environmental Sciences from the University of Barcelona and is currently pursuing a Master’s degree in Bioinformatics and Biostatistics at the Open University of Catalonia. She works as a research technician with a focus on rocky shore biodiversity and the physiological responses of intertidal invertebrates to thermal stress, and she is also actively engaged in citizen science activities.

Joana Pereira holds a Master’s degree in Biodiversity, Genetics and Evolution from the University of Porto and is currently a PhD student at the BIODIV doctoral program at CIBIO. Her PhD thesis focuses on understanding the mechanistic links between climate change and coastal biodiversity.

João Nunes holds a Master’s degree in Marine Sciences from the University of Porto. His research focuses on intertidal biodiversity monitoring using UAS (drones) and automated image classification. He has contributed to studies on rocky shore communities and the impacts of environmental changes. Additionally, he has been involved in citizen science initiatives and ocean conservation projects.

Rocío Nieto-Vilela holds a PhD in Biological Sciences with a marine focus from the Universidad de la Patagonia San Juan Bosco, Argentina, which she completed in 2020. Following her doctorate, she undertook a postdoctoral research position (2020–2023) focused on the physiological effects caused by environmental contaminants. Since then, she has been working as a researcher at CIBIO/BIOPOLIS, contributing to the ANERIS project through biodiversity data analysis, environmental DNA research, and the coordination of citizen science initiatives.

Ana Sinde-Mano is a marine scientist fascinated by the biodiversity and ecology of rocky coastal communities. She holds a Master’s degree in Marine Biology and Conservation from the Applied Psychology Institute in Lisbon, Portugal. Currently, she is working on identifying and mapping the distribution of intertidal species to make temporal comparisons and understand shifts in these communities due to climate change.
Acknowledgments
We thank the MINKA and iNaturalist community for their valuable contributions which greatly aided this research. Additionally, we extend our gratitude to Marta Cortés for the design of the graphical abstract.
-
Research ethics: Not applicable.
-
Informed consent: Not applicable.
-
Author contributions: MH and JP: fieldwork, data curation, data analysis, writing – original draft, review and editing. JN: fieldwork, data analysis, writing – review and editing. RN: fieldwork, conceptualization, writing – review and editing. AS: fieldwork, writing – review and editing. FL: conceptualization, funding acquisition, writing – review and editing. CM: fieldwork, conceptualization, funding acquisition, writing – review and editing. All authors have accepted responsibility for the entire content of this manuscript and approved its submission.
-
Use of Large Language Models, AI and Machine Learning Tools: None declared.
-
Conflict of interest: The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
-
Research funding: This work was supported by Fundação para Ciência e Tecnologia (FCT) through the projects BIOINTERACT (https://doi.org/10.54499/2022.02887.PTDC) and OceanLog (PTDC/BIA-BMA/4848/202), and by the Horizon 2020 project ANERIS (ref. 101094924). JP received support from FCT through a PhD grant (2022.12126.BD) and FPL through a CEECInd contracts (10.54499/CEECIND/03185/2018/CP1546/CP1648/CT0004). ALM received support from LifeWatch ERIC (101059548).
-
Data availability: The authors confirm that the data supporting the findings of this study are available within the article and its Supplementary material.
References
Araújo, R., Bárbara, I., Tibaldo, M., Berecibar, E., Tapia, P.D., Pereira, R., Santos, R., and Pinto, I.S. (2009). Checklist of benthic marine algae and cyanobacteria of northern Portugal. Bot. Mar. 52: 24–46, https://doi.org/10.1515/bot.2009.026.Search in Google Scholar
Arriaga, O., Wawrzynkowski, P., Muguerza, N., Díez, I., Gorostiaga, J.M., Quintano, E., and Becerro, M.A. (2024). The thermal journey of macroalgae: four decades of temperature-induced changes in the southeastern Bay of Biscay. Mar. Env. Res. 195: 106351, https://doi.org/10.1016/j.marenvres.2024.106351.Search in Google Scholar PubMed
Báez, J.C., Olivero, J., Peteiro, C., Ferri-Yáñez, F., Garcia-Soto, C., and Real, R. (2010). Macro-environmental modelling of the current distribution of Undaria pinnatifida (Laminariales, Ochrophyta) in northern Iberia. Biol. Invasions 12: 2131–2139, https://doi.org/10.1007/s10530-009-9614-1.Search in Google Scholar
Blanco, A., Larrinaga, A.R., Neto, J.M., Troncoso, J., Méndez, G., Domínguez-Lapido, P., Ovejero, A., Pereira, L., Mouga, T.M., Gaspar, R., et al.. (2021). Spotting intruders: species distribution models for managing invasive intertidal macroalgae. J. Environ. Manage. 281: 111861, https://doi.org/10.1016/j.jenvman.2020.111861.Search in Google Scholar PubMed
Boaventura, D., Ré, P., Da Fonseca, L.C., and Hawkins, S.J. (2002). Intertidal rocky shore communities of the continental Portuguese coast: analysis of distribution patterns. Mar. Ecol. 23: 69–90.10.1046/j.1439-0485.2002.02758.xSearch in Google Scholar
Boudouresque, C.F., Gerbal, M., and Knoepffler-Peguy, M. (1985). L’algue japonnaise Undaria pinnatifida (Phaeophyceae, Laminariales) en Méditerranée. Phycologia 24: 364–366, https://doi.org/10.2216/i0031-8884-24-3-364.1.Search in Google Scholar
Brondizio, E.S., Settele, J., Díaz, S., and Ngo, H.T. (2019). Global assessment report on biodiversity and ecosystem services. Intergovernmental Science-Policy Platform on Biodiversity and Ecosystem Services. IPBES, 978-3-947851-13-3.Search in Google Scholar
Burrows, M.T., Harvey, R., and Robb, L. (2008). Wave exposure indices from digital coastlines and the prediction of rocky shore community structure. Mar. Ecol. Prog. Ser. 353: 1–12, https://doi.org/10.3354/meps07284.Search in Google Scholar
Caamano, J.S., Neira, C.D., and Castroviejo, R.A. (1990). Aparición de Undaria pinnatifida en las costas de Galicia (España): un nuevo caso en la problemática de introducción de especies foráneas. Inf. Téc. Cent. Invest. Submar. 3.Search in Google Scholar
Carreira-Flores, D., Rubal, M., Moreira, J., Guerrero-Mesenguer, L., Gomes, P.T., and Veiga, P. (2023). Recent changes on the abundance and distribution of non-indigenous macroalgae along the southwest coast of the Bay of Biscay. Aquat. Bot. 189: 103685, https://doi.org/10.1016/j.aquabot.2023.103685.Search in Google Scholar
Casado-Amezúa, P., Araújo, R., Bárbara, I., Bermejo, R., Borja, A., Díez, I., Fernández, C., Gorostiaga, J.M., Guinda, X., Hernández, I., et al.. (2019). Distributional shifts of canopy-forming seaweeds from the Atlantic coast of Southern Europe. Biodivers. Conserv. 28: 1151–1172, https://doi.org/10.1007/s10531-019-01716-9.Search in Google Scholar
Casas, G., Scrosati, R., and Luz Piriz, M. (2004). The invasive kelp Undaria pinnatifida (Phaeophyceae, Laminariales) reduces native seaweed diversity in Nuevo Gulf (Patagonia, Argentina). Biol. Invasions 6: 411–416, https://doi.org/10.1023/b:binv.0000041555.29305.41.10.1023/B:BINV.0000041555.29305.41Search in Google Scholar
Çinar, M.E., Arianoutsou, M., Zenetos, A., Golani, D., Çinar, M.E., Oztürk, B., Grabowski, M., and Cardoso, A.C. (2014). Impacts of invasive alien marine species on ecosystem services and biodiversity: a pan-European review. Aquat. Invasions 9: 391–423, https://doi.org/10.3391/ai.2014.9.4.01.Search in Google Scholar
Curiel, D., Guidetti, P., Bellemo, G., Scattolin, M., and Marzocchi, M. (2002). The introduced alga Undaria pinnatifida (Laminariales, Alariaceae) in the lagoon of Venice. Hydrobiol. J. 477: 209–219, https://doi.org/10.1023/a:1021094008569.10.1023/A:1021094008569Search in Google Scholar
Davidson, A.D., Campbell, M.L., Hewitt, C.L., and Schaffelke, B. (2015). Assessing the impacts of nonindigenous marine macroalgae: an update of current knowledge. Bot. Mar. 58: 55–79, https://doi.org/10.1515/bot-2014-0079.Search in Google Scholar
de Azevedo, J., Franco, J.N., Vale, C.G., Lemos, M.F.L., and Arenas, F. (2023). Rapid tropicalization evidence of subtidal seaweed assemblages along a coastal transitional zone. Sci. Rep. 13: 11720, https://doi.org/10.1038/s41598-023-38514-x.Search in Google Scholar PubMed PubMed Central
De Leij, R., Epstein, G., Brown, M.P., and Smale, D.A. (2017). The influence of native macroalgal canopies on the distribution and abundance of the non-native kelp Undaria pinnatifida in natural reef habitats. Mar. Biol. 164: 156, https://doi.org/10.1007/s00227-017-3183-0.Search in Google Scholar
Desmond, M.J., Pritchard, D.W., Hurd, C.L., Richards, D.K., Schweikert, K., Wing, S., and Hepburn, C.D. (2019). Superior photosynthetic performance of the invasive kelp Undaria pinnatifida may contribute to continued range expansion in a wave-exposed kelp forest community. Mar. Biol. 166: 1–11, https://doi.org/10.1007/s00227-019-3593-2.Search in Google Scholar
Dudeque Zenni, R., Essl, F., García-Berthou, E., Mcdermott, S.M., and Zenni, R.D. (2021). The economic costs of biological invasions around the world. NeoBiota 67: 1–9, https://doi.org/10.3897/neobiota.67.69971.Search in Google Scholar
Encarnação, J., Teodósio, M.A., and Morais, P. (2021). Citizen science and biological invasions: a review. Front. Environ. Sci. 8: 602980, https://doi.org/10.3389/fenvs.2020.602980.Search in Google Scholar
Epstein, G. and Smale, D.A. (2017). Undaria pinnatifida: a case study to highlight challenges in marine invasion ecology and management. Ecol. Evol. 7: 8624–8642, https://doi.org/10.1002/ece3.3430.Search in Google Scholar PubMed PubMed Central
Fiuza, A.F.G. (1983). Upwelling patterns off Portugal. NATO conference series, (series) 4. Mar. Sci. 10A: 85–98.10.1007/978-1-4615-6651-9_5Search in Google Scholar
Floc’h, J.-Y., Pajot, R., and Mouret, V. (1996). Undaria pinnatifida (Laminariales, Phaeophyta) 12 years after its introduction into the Atlantic Ocean. Hydrobiologia 326: 217–222, https://doi.org/10.1007/978-94-009-1659-3_30.Search in Google Scholar
Floc’h, J.Y., Pajot, R., and Wallentinus, I. (1991). The Japanese brown alga Undaria pinnatifida on the coast of France and its possible establishment in European waters. ICES J. Mar. Sci. 47: 379–390, https://doi.org/10.1093/icesjms/47.3.379.Search in Google Scholar
Forrest, B.M. and Blakemore, K.A. (2006). Evaluation of treatments to reduce the spread of a marine plant pest with aquaculture transfers. Aquaculture 257: 333–345, https://doi.org/10.1016/j.aquaculture.2006.03.021.Search in Google Scholar
Gao, X., Endo, H., Taniguchi, K., and Agatsuma, Y. (2013). Combined effects of seawater temperature and nutrient condition on growth and survival of juvenile sporophytes of the kelp Undaria pinnatifida (Laminariales; Phaeophyta) cultivated in northern Honshu, Japan. J. Appl. Phycol. 25: 269–275.10.1007/s10811-012-9861-xSearch in Google Scholar
Gómez-Gesteira, M., deCastro, M., Alvarez, I., and Gómez-Gesteira, J.L. (2008). Coastal sea surface temperature warming trend along the continental part of the Atlantic Arc (1985–2005). J. Geophys. Res. 113: C04010, https://doi.org/10.1029/2007jc004315.Search in Google Scholar
Heiser, S., Hall-Spencer, J.M., and Hiscock, K. (2014). Assessing the extent of establishment of Undaria pinnatifida in Plymouth Sound Special Area of Conservation, UK. Mar. Biodivers. Rec. 7: e93, https://doi.org/10.1017/s1755267214000608.Search in Google Scholar
Hernangómez, D. (2020). giscoR: download map data from GISCO API – Eurostat (Version 0.6.1). CRAN: Contributed Packages, Available at: https://cran.r-project.org/web/packages/giscoR/index.htmlLowe.10.32614/CRAN.package.giscoRSearch in Google Scholar
Hewitt, C.L., Campbell, M.L., McEnnulty, F., Moore, K.M., Murfet, N.B., Robertson, B., and Schaffelke, B. (2005). Efficacy of physical removal of a marine pest: the introduced kelp Undaria pinnatifida in a Tasmanian Marine Reserve. Biol. Invasions 7: 251–263, https://doi.org/10.1007/s10530-004-0739-y.Search in Google Scholar
Hewitt, C.L., Campbell, M.L., and Schaffelke, B. (2007). Introductions of seaweeds: accidental transfer pathways and mechanisms. Bot. Mar. 50: 326–337, https://doi.org/10.1515/bot.2007.038.Search in Google Scholar
Hiscock, K. (1981). The rocky shore ecology of Sullom Voe. Proc. R. Soc. Edinb. B Biol. Sci. 80: 219–240, https://doi.org/10.1017/s026972700000659x.Search in Google Scholar
James, K. and Shears, N.T. (2016). Proliferation of the invasive kelp Undaria pinnatifida at aquaculture sites promotes spread to coastal reefs. Mar. Biol. 163: 1–12, https://doi.org/10.1007/s00227-015-2811-9.Search in Google Scholar
James, K., Kibele, J., and Shears, N.T. (2015). Using satellite-derived sea surface temperature to predict the potential global range and phenology of the invasive kelp Undaria pinnatifida. Biol. Invasions 17: 3393–3408, https://doi.org/10.1007/s10530-015-0965-5.Search in Google Scholar
Jiménez, R.S., Hepburn, C.D., Hyndes, G.A., McLeod, R.J., and Hurd, C.L. (2015). Contributions of an annual invasive kelp to native algal assemblages: algal resource allocation and seasonal connectivity across ecotones. Phycologia 54: 530–544, https://doi.org/10.2216/15-39.1.Search in Google Scholar
Kaplanis, N.J., Harris, J.L., and Smith, J.E. (2016). Distribution patterns of the non-native seaweeds Sargassum horneri (Turner) C. Agardh and Undaria pinnatifida (Harvey) Suringar on the San Diego and Pacific coast of North America. Aquat. Invasions 11: 111, https://doi.org/10.3391/ai.2016.11.2.01.Search in Google Scholar
Kelly, R., Fleming, A., Pecl, G.T., von Gönner, J., and Bonn, A. (2020). Citizen science and marine conservation: a global review. Philos. Trans. R. Soc. Lond. B Biol. Sci. 375: 20190461, https://doi.org/10.1098/rstb.2019.0461.Search in Google Scholar PubMed PubMed Central
Lima, F.P., Ribeiro, P.A., Queiroz, N., Hawkins, S.J., and Santos, A.M. (2007). Do distributional shifts of northern and southern species of algae match the warming pattern? Glob. Change Biol. 13: 2592–2604, https://doi.org/10.1111/j.1365-2486.2007.01451.x.Search in Google Scholar
Lowe, S., Browne, M., Boudjelas, S., and Poorter, M. De. (2000). 100 of the world’s worst invasive alien species: a selection from the global invasive species database. The Invasive Species Specialist Group (ISSG) a specialist group of the Species Survival Commission (SSC) of the World Conservation Union (IUCN), Auckland, New Zealand.Search in Google Scholar
MacDougall, A.S. and Turkington, R. (2005). Are invasive species the drivers or passengers of change in degraded ecosystems? Ecology 86: 42–55, https://doi.org/10.1890/04-0669.Search in Google Scholar
Mack, R.N., Simberloff, D., Mark Lonsdale, W., Evans, H., Clout, M., and Bazzaz, F.A. (2000). Biotic invasions: causes, epidemiology, global consequences, and control. Ecol. Appl. 10: 689–710.10.1890/1051-0761(2000)010[0689:BICEGC]2.0.CO;2Search in Google Scholar
Massicotte, P. and South, A. (2023). rnaturalearth: world map data from natural earth (version 1.0.1), Available at: https://cran.r-project.org/web/packages/rnaturalearth/index.html.Search in Google Scholar
McKnight, E., Spake, R., Bates, A., Smale, D.A., and Rius, M. (2021). Non-native species outperform natives in coastal marine ecosystems subjected to warming and freshening events. Glob. Ecol. Biogeogr. 30: 1698–1712, https://doi.org/10.1111/geb.13318.Search in Google Scholar
Mineur, F., Arenas, F., Assis, J., Davies, A.J., Engelen, A.H., Fernandes, F., Malta, E.j., Thibaut, T., Van Nguyen, T., Vaz-Pinto, F., et al.. (2015). European seaweeds under pressure: consequences for communities and ecosystem functioning. J. Sea Res. 98: 91–108, https://doi.org/10.1016/j.seares.2014.11.004.Search in Google Scholar
Monteiro, C., Pereira, J., Seabra, R., and Lima, F.P. (2022). Fine-scale survey of intertidal macroalgae reveals recent changes in a cold-water biogeographic stronghold. Front. Mar. Sci. 9: 880074, https://doi.org/10.3389/fmars.2022.880074.Search in Google Scholar
Pebesma, E. and Bivand, R. (2023). Spatial data science. In: Spatial data science: with applications in R, 1st ed. Chapman and Hall/CRC, New York.10.1201/9780429459016Search in Google Scholar
Pereira, J., Monteiro, C., Seabra, R., and Lima, F.P. (2022). Fine-scale abundance of rocky shore macroalgae species with distribution limits in NW Iberia in 2020/2021. Biodivers. Data J. 10: e80798, https://doi.org/10.3897/bdj.10.e80798.Search in Google Scholar PubMed PubMed Central
Pérez Ruzafa, I.M., Menéndez, J.L., and Salinas, J.M. (2002). Mapas de distribución de algas marinas de la Península Ibérica y las Islas Baleares. XV. Undaria pinnatifida (Harvey) Suringar (Laminariales, Fucophyceae). Bot. Complut. 26: 147–152.Search in Google Scholar
Peteiro, C. (2008). A new record of the introduced seaweed Undaria pinnatifida (Laminariales, Phaeophyceae) from the Cantabrian Sea (northern Spain) with comments on its establishment. Aquat. Invasions 3: 413–415, https://doi.org/10.3391/ai.2008.3.4.6.Search in Google Scholar
R Development Core Team (2024). R: a language and environment for statistical computing (version 4.4.2). R Foundation for Statistical Computing, Vienna, Austria.Search in Google Scholar
Raffo, M.P., Eyras, M.C., and Iribarne, O.O. (2009). The invasion of Undaria pinnatifida to a Macrocystis pyrifera kelp in Patagonia (Argentina, south-west Atlantic). J. Mar. Biol. Assoc. U. K. 89: 1571–1580, https://doi.org/10.1017/s002531540900071x.Search in Google Scholar
Ramos, E., Guinda, X., Puente, A., de la Hoz, C.F., and Juanes, J.A. (2020). Changes in the distribution of intertidal macroalgae along a longitudinal gradient in the northern coast of Spain. Mar. Environ. Res. 157: 104930, https://doi.org/10.1016/j.marenvres.2020.104930.Search in Google Scholar PubMed
Ramos, E., Puente, A., and Juanes, J.A. (2016). An ecological classification of rocky shores at a regional scale: a predictive tool for management of conservation values. Mar. Ecol. 37: 311–328, https://doi.org/10.1111/maec.12280.Search in Google Scholar
Richardson, D., Pyšek, P., and Carlton, J.T. (2011). A compendium of essential concepts and terminology in invasion ecology. In: Richarson, D. (Ed.). Fifty years of invasion ecology: the legacy of Charles Elton. Blackwell Publishing Ltd, West Sussex, pp. 409–420.10.1002/9781444329988.ch30Search in Google Scholar
Roy, H.E., Pauchard, A., Stoett, P., Renard Truong, T., Lipinskaya, T., and Vicente, J.R. (2023). Introducing biological invasions and the IPBES thematic assessment of invasive alien species and their control. In: Roy, H.E., Pauchard, A., Stoett, P., and Renard Truong, T. (Eds.). Thematic assessment report on invasive alien species and their control of the intergovernmental science-policy platform on biodiversity and ecosystem services. IPBES Secretariat, Bonn, Germany, chapter 1.Search in Google Scholar
Russell, L.K., Hepburn, C.D., Hurd, C.L., and Stuart, M.D. (2008). The expanding range of Undaria pinnatifida in southern New Zealand: distribution, dispersal mechanisms and the invasion of wave-exposed environments. Biol. Invasions 10: 103–115, https://doi.org/10.1007/s10530-007-9113-1.Search in Google Scholar
Salinas, J.M., Llera, E.M., and Fuertes, C. (1996). Nota sobre la presencia de Undaria pinnatifida (Harvey) Suringar (Laminariales, Phaeophyta) en Asturias (mar Cantábrico). Bol. Inst. Esp. Oceanogr. 12: 77–79.Search in Google Scholar
Schaffelke, B., Smith, J.E., and Hewitt, C.L. (2006). Introduced macroalgae – a growing concern. J. Appl. Phycol. 18: 529–541, https://doi.org/10.1007/s10811-006-9074-2.Search in Google Scholar
Schiller, J., Lackschewitz, D., Buschbaum, C., Reise, K., Pang, S., and Bischof, K. (2018). Heading northward to Scandinavia: Undaria pinnatifida in the northern Wadden Sea. Bot. Mar. 61: 365–371, https://doi.org/10.1515/bot-2017-0128.Search in Google Scholar
Simberloff, D., Martin, J.L., Genovesi, P., Maris, V., Wardle, D.A., Aronson, J., Courchamp, F., Galil, B., García-Berthou, E., Pascal, M., et al.. (2013). Impacts of biological invasions: what’s what and the way forward. Trends Ecol. Evol. 28: 58–66.10.1016/j.tree.2012.07.013Search in Google Scholar PubMed
Slowikowski, K. (2024). ggrepel: automatically position non-overlapping text labels with “ggplot2” (version 0.9.6), Available at: https://cran.r-project.org/web/packages/ggrepel/index.html.Search in Google Scholar
South, P.M. and Thomsen, M.S. (2016). The ecological role of invading Undaria pinnatifida: an experimental test of the driver–passenger models. Mar. Biol. 163: 175, https://doi.org/10.1007/s00227-016-2948-1.Search in Google Scholar
South, P.M., Floerl, O., Forrest, B.M., and Thomsen, M.S. (2017). A review of three decades of research on the invasive kelp Undaria pinnatifida in Australasia: an assessment of its success, impacts and status as one of the world’s worst invaders. Mar. Environ. Res. 131: 243–257, https://doi.org/10.1016/j.marenvres.2017.09.015.Search in Google Scholar PubMed
Spalding, M.D., Fox, H.E., Allen, G.R., Davidson, N., Ferdaña, Z.A., Finlayson, M., Halpern, B.S., Jorge, M.A., Lombana, A., Lourie, S.A., et al.. (2007). Marine ecoregions of the world: a bioregionalization of coastal and shelf areas. BioScience 57: 573–583, https://doi.org/10.1641/b570707.Search in Google Scholar
Steneck, R.S., Graham, M.H., Bourque, B.J., Corbett, D., Erlandson, J.M., Estes, J.A., and Tegner, M.J. (2002). Kelp forest ecosystems: biodiversity, stability, resilience and future. Environ. Conserv. J. 29: 436–459, https://doi.org/10.1017/s0376892902000322.Search in Google Scholar
Suárez-Jiménez, R., Hepburn, C.D., Hyndes, G.A., McLeod, R.J., Taylor, R.B., and Hurd, C.L. (2017). The invasive kelp Undaria pinnatifida hosts an epifaunal assemblage similar to native seaweeds with comparable morphologies. Mar. Ecol. Prog. Ser. 582: 45–55, https://doi.org/10.3354/meps12321.Search in Google Scholar
Thomsen, M.S., Wernberg, T., Engelen, A.H., Tuya, F., Vanderklift, M.A., Holmer, M., McGlathery, K.J., Arenas, F., Kotta, J., and Silliman, B.R. (2012). A meta-analysis of seaweed impacts on seagrasses: generalities and knowledge gaps. PLoS One 7: 28595, https://doi.org/10.1371/journal.pone.0028595.Search in Google Scholar PubMed PubMed Central
Thornber, C.S., Kinlan, B.P., Graham, M.H., and Stachowicz, J.J. (2004). Population ecology of the invasive kelp Undaria pinnatifida in California: environmental and biological controls on demography. Mar. Ecol. Prog. Ser. 268: 69–80, https://doi.org/10.3354/meps268069.Search in Google Scholar
Ugarte, J.C., Gago, Ó.F., and García, C.P. (2006). Biología, distribución e integración del alga alóctona Undaria pinnatifida (Laminariales, Phaeophyta) en las comunidades bentónicas de las costas de Galicia (NW de la Península Ibérica). An. Jard. Bot. Madr. 63: 169–187, https://doi.org/10.3989/ajbm.2006.v63.i2.6.Search in Google Scholar
Veiga, P., Torres, A.C., Rubal, M., Troncoso, J., and Sousa-Pinto, I. (2014). The invasive kelp Undaria pinnatifida (Laminariales, Ochrophyta) along the north coast of Portugal: distribution model versus field observations. Mar. Pollut. Bull. 84: 363–365, https://doi.org/10.1016/j.marpolbul.2014.05.038.Search in Google Scholar PubMed
Wickham, H. (2016). ggplot2: elegant graphics for data analysis. Springer International Publishing, Cham, Switzerland.Search in Google Scholar
Supplementary Material
This article contains supplementary material (https://doi.org/10.1515/bot-2025-0013).
© 2025 the author(s), published by De Gruyter, Berlin/Boston
This work is licensed under the Creative Commons Attribution 4.0 International License.
{kind=link}
Articles in the same Issue
- Frontmatter
- In this issue
- Physiology and Ecology
- Three-dimensional distribution of nutrients and phytoplankton biomass in a semi-enclosed region of the Gulf of California during different ENSO phases
- Proliferation of Undaria pinnatifida along the Atlantic coast of the Iberian Peninsula
- Study on fatty acid methyl ester (FAME) composition of 19 Brazilian thraustochytrid isolates
- Taxonomy/Phylogeny and Biogeography
- Variations in the bacterial and fungal community structure along the hypoxic gradient of the Arabian Sea oxygen-depleted environment based on eDNA metabarcoding analysis
- Studies of North Carolina marine algae XV. DNA sequencing reveals some different Ulva species compared to historical reports and U. carsoniae sp. nov. (Ulvales, Chlorophyta)
- Identification and growth of Prorocentrum cf. cassubicum (Dinoflagellata: Prorocentraceae) from Bahía de La Paz: first record for the Gulf of California
- A new oomycete pathogen Olpidiopsis dasysiphoniae sp. nov. (Oomycota) infecting the red alga Dasysiphonia japonica (Ceramiales, Delesseriaceae)
- Chemistry and Applications
- Exploring the suitable extractive species in an IMTA: inorganic nutrient removal from mariculture effluents by commercially important marine macroalgae
Articles in the same Issue
- Frontmatter
- In this issue
- Physiology and Ecology
- Three-dimensional distribution of nutrients and phytoplankton biomass in a semi-enclosed region of the Gulf of California during different ENSO phases
- Proliferation of Undaria pinnatifida along the Atlantic coast of the Iberian Peninsula
- Study on fatty acid methyl ester (FAME) composition of 19 Brazilian thraustochytrid isolates
- Taxonomy/Phylogeny and Biogeography
- Variations in the bacterial and fungal community structure along the hypoxic gradient of the Arabian Sea oxygen-depleted environment based on eDNA metabarcoding analysis
- Studies of North Carolina marine algae XV. DNA sequencing reveals some different Ulva species compared to historical reports and U. carsoniae sp. nov. (Ulvales, Chlorophyta)
- Identification and growth of Prorocentrum cf. cassubicum (Dinoflagellata: Prorocentraceae) from Bahía de La Paz: first record for the Gulf of California
- A new oomycete pathogen Olpidiopsis dasysiphoniae sp. nov. (Oomycota) infecting the red alga Dasysiphonia japonica (Ceramiales, Delesseriaceae)
- Chemistry and Applications
- Exploring the suitable extractive species in an IMTA: inorganic nutrient removal from mariculture effluents by commercially important marine macroalgae