Studies of North Carolina marine algae XV. DNA sequencing reveals some different Ulva species compared to historical reports and U. carsoniae sp. nov. (Ulvales, Chlorophyta)
-
Skye Dibner is a research analyst at the University of North Carolina at Wilmington. She earned her undergraduate degree at Eckerd College, where she completed a thesis on the molecular systematics of holopelagic
Sargassum . She conducted her master’s research at UNCW, focusing onUlva species diversity in North Carolina. Her current work at UNCW’s Shellfish Research Hatchery centers on the molecular genetics ofCrassostrea virginica (Eastern Oyster).Paul W. Gabrielson is an algal systematist. He obtained his PhD at the University of North Carolina, Chapel Hill with Max Hommersand and was a post-doc with Gerald T. Kraft and the late Robert F. Scagel. He retooled from morpho-anatomy to DNA sequencing while teaching at a small, liberal arts college for nine years, before returning to UNC, Chapel Hill. He collaborates with phycologists worldwide sequencing contemporary field-collected specimens, as well as type specimens of seaweeds from the 18th, 19th and early 20th centuries.
 und
und
D. Wilson Freshwater is researcher at the Center for Marine Science, University of North Carolina, Wilmington. A recent graduate of genomics kindergarten, he has research interests in the molecular phylogeny and taxonomy of marine algae, marine floristics, and the ecology of marine hard bottom communities.


Abstract
Ulva is a genus of green algae common to intertidal and shallow subtidal marine, estuarine, and freshwater habitats worldwide, including North Carolina. To accurately determine the Ulva species diversity in North Carolina, both DNA sequencing and morphological/anatomical assessments were performed. Specimens were collected from sites across the coastal region of the state, and three DNA loci, nuclear-encoded ITS, plastid-encoded rbcL and plastid-encoded tufA, were sequenced for each specimen. Species names were determined where possible by homology of sequence data with that from type specimens. Through DNA sequence cluster and phylogenetic analyses, nine species of Ulva were identified in the collections. Seven, U. gigantea, U. lacinulata, U. lactuca, U. ohnoi, U. prolifera, U. ‘torta’, and U. tepida were confirmed by comparing to available type or topotype sequences. The remaining two are either undescribed species or species for which there is no sequence available from the type specimen. One of these is described as a new species, U. carsoniae. Morpho-anatomical characters were not useful in distinguishing these nine species or determining their identifications. Only the DNA sequenced loci provided an accurate diversity assessment of Ulva species in North Carolina.
1 Introduction
There are 100 species and 51 sub-specific taxa currently accepted in the green algal genus Ulva L. (Guiry and Guiry 2025). Ulva is characterized as a monostromatic tubular or distromatic blade-forming macroalga with cells containing one plastid with one or more pyrenoids. Ulva is increasingly being considered as a potential source of food or biofuel for humans and may be used as an environmental indicator (Charrier et al. 2017; Dominguez and Loret 2019; Rybak 2018). Ulva may also play a role as a fertilizer or natural stimulant for plant growth (Dmytryk and Chojnacka 2018). Across the globe, Ulva species have formed “green tides”, large blooms that can occur locally in coves and embayments or cover long stretches of coastline (Cai et al. 2021; Chávez Sanchez et al. 2018; Hughey et al. 2018; Lee et al. 2019; Liu et al. 2010; Rinehart et al. 2014; Runca et al. 1996). These blooms often occur in spring and summer months and can impact tourism and local revenue where they occur. Major blooms result in the fouling and degradation of beaches and may impede boat travel. Correct identification of Ulva species is crucial to assess species diversity worldwide and to understand which taxa are causing “green tides”.
Ulva was one of the earliest algal genera named by Linnaeus (1753), and it has undergone multiple taxonomic revisions. Link (1820) divided Ulva into two genera based on morphology, with monostromatic tubular species placed in a new genus, Enteromorpha Link, whereas distromatic blade-forming species were kept in Ulva. This distinction, however, was difficult when classifying species such as Ulva ‘linza’ L., which has a distromatic blade with hollow, tubular blade margins and tubular stipes at its base (e.g., Marshall et al. 2006; Schneider and Searles 1991). [Note that throughout species whose type specimens have not been sequenced are indicated by single quotes]. Enteromorpha was considered a distinct genus until Tan et al. (1999) and Hayden et al. (2003) showed that phylogenetic analyses of ITS and rbcL sequences from bladed and tubular species were not placed in separate evolutionary clades. Some recent publications do not follow this synonymy (e.g., Peng et al. 2008; Wang et al. 2010; Wassie et al. 2021).
The correct identification of Ulva species using morphological and/or anatomical characters is difficult due to intraspecific variation of these characters as well as the interspecific overlap of morphological characters. It has been suggested that environmental factors, such as salinity and temperature, may influence the morphology of Ulva species (Blomster et al. 1998; Gao et al. 2016; Rybak 2018), but morphological plasticity of species cultured under the same conditions is known (Føyn 1958; Steinhagen et al. 2019a). Additionally, Ulva species grow symbiotically with bacterial microbiomes, which have been shown to play a role in morphological plasticity and growth (Ghaderiardakani et al. 2017; Marshall et al. 2006; Spoerner et al. 2012; Wichard et al. 2015). Additionally, the correct application of names is hampered by the many Ulva type specimens that have not been sequenced (Hofmann et al. 2010).
In most cases, Ulva diversity reports based on specimens identified by morphology or anatomy do not accurately reflect what species are present in an ecosystem (Chávez-Sánchez et al. 2017, 2019). Also there are numerous misnamed DNA sequences in public databases when type specimens of those species have not been sequenced. For example, prior to its lectotype being sequenced in 2021, no DNA sequences in any public database had been correctly attributed to U. rigida C. Agardh (Hughey et al. 2021a). Even U. lactuca, the generitype, has until recently been misidentified both through incorrect sequence naming in public databases and confusion over the locality and morphology of the type specimen (Hughey et al. 2019). Data mining in concert with DNA sequencing of type specimens has been useful in confirming Ulva species misidentifications (Fort et al. 2022). Sequences of type specimens are available for only 42 of the 400+ potential Ulva species names (Table 1; Guiry and Guiry 2025), which is essential for the correct application of species names for taxonomic (e.g., Hughey et al. 2018, 2019, 2021a, 2021b, 2024) and floristic studies (e.g. Kuba et al. 2022, Gabrielson et al. 2024).
Sequences of rbcL, tufA, and ITS currently available for Ulva species type specimens.
| Species | rbcL accession # | tufA accession # | ITS accession # | Citation | Associated accession # |
|---|---|---|---|---|---|
| Ulva australis Areschoug | LC331300 | – | LC331301 | Hanyuda and Kawai (2018) | |
| Ulva arbuscula Lagourgue et Payri | MZ902936 | – | MZ870607 | Lagourgue et al. (2022) | |
| Ulva batuffolosaa Lagourgue et Payri | MZ902977 | MZ870679 | MZ870641 | Lagourgue et al. (2022) | |
| Ulva californica Wille | OR723827 | OR723828 | OR722833 | Hughey et al. (2024) | |
| Ulva capillata S.Steinhagena | OP265117 | OL421407 | Steinhagen et al. (2022) | ||
| Ulva chaulugei Kavale et Kazi | KP710833 | – | KT429218 | Kazi and Kavale (2015) | |
| Ulva conglobataa Kjellman | MT815850 | – | – | Hughey et al. (2021b) | |
| Ulva clathratioidesa L.G.Kraft, Kraft, et R.F.Waller | EU933951 | – | EU933978 | Kraft et al. (2010) | |
| Ulva curvataa (Kützing) De Toni | OR635441 | – | – | Maggs et al. (2024) | |
| Ulva dactylifera Setchell et N.L.Gardner | OR003918 | OR003918 | OR730543 | Hughey et al. (2024) | |
| Ulva expansa (Setchell) Setchell et N.L.Gardner | MH731009 | MH731007 | MH730160 | Hughey et al. (2021b) | |
| Ulva fasciata Delile | MK456397 | – | AY260561 | Hughey et al. (2019) | |
| Ulva fenestrata Postels et Ruprecht | MK456393 | MK456404 | – | Hughey et al. (2021b) | MT894725 (ITS) |
| Ulva finissima Lagourgue et Payri | MZ902989 | MZ870687 | MZ870652 | Lagourgue et al. (2022) | |
| Ulva giganteaa (Kützing) Bliding | OR427286 | OR427287 | OR428167 | Maggs et al. (2024) | |
| Ulva iliohaha Spalding et Sherwood | KT932995 | KT932976 | KT881223 (ITS1) | Spalding et al. (2016) | |
| Ulva kanagawae Carniero, Martins et Cassano | OQ687080 | OQ653885 | – | Carneiro et al. (2023) | |
| Ulva lacinulata (Kützing) Wittrock | MW543061 | MW543061 | MW544060 | Hughey et al. (2021a) | |
| Ulva lactuca Linnaeus | MK456395 | – | – | Hughey et al. (2021b) | KT882614 (chloroplast genome) |
| Ulva lobata (Kützing) Harvey | MH730972 | MH730972 | – | Hughey et al. (2019) | |
| Ulva nematoidea Bory | OL542489 | – | – | Hughey and Gabrielson (2022) | |
| Ulva ohiohilulu Spalding et Sherwood | – | KT932977 | KT881224 (ITS1), KT881226 (ITS2) | Spalding et al. (2016) | |
| Ulva ohnoi Hiraoka et Shimada | AP018696 | AP018696 | – | Hughey et al. (2021b) | |
| Ulva pennata Lagourgue et Payri | MZ936429 | MZ921402 | MZ921397 | Lagourgue et al. (2022) | |
| Ulva piritokaa Ngāti Kuri, Heesch et W.A. Nelson | MW389665 | – | – | Heesch et al. (2021) | |
| Ulva planiramosa Lagourgue et Payri | MZ902965 | MZ870671 | MZ870630 | Lagourgue et al. (2022) | |
| Ulva pluriramosa Lagourgue et Payri | MZ902984 | – | MZ870647 | Lagourgue et al. (2022) | |
| Ulva pouliotiia Golden et S.C. Lindstrom | – | MW921464 | MW939397 | Lindstrom et al. (2021) | |
| Ulva prolifera Müller | – | – | AJ012276 | Cui et al. (2018) | |
| Ulva pseudocurvataa Koeman et C.Hoek | OR635440 | OR539204 | OR536260 (ITS1) | Maggs et al. (2024) | |
| Ulva pseudo-ohnoia H.W.Lee, J.C.Kang et M.S.Kim | MT624844 | MT625015 | Lee et al. (2019) | ||
| Ulva rigida C. Agardh | MW543060 | MW543060 | MW544059 | Hughey et al. (2021a) | |
| Ulva rotundata Bliding | MW570779 | – | – | Hughey et al. (2019) | |
| Ulva scolopendra Lagourgue et Payri | MZ936426 | MZ870675 | MZ870636 | Lagourgue et al. (2022) | |
| Ulva siganiphyllia Lagourgue et Payri | MZ902959 | – | MZ870627 | Lagourgue et al. (2022) | |
| Ulva spumosa Lagourgue et Payri | MZ902952 | MZ870664 | MZ870621 | Lagourgue et al. (2022) | |
| Ulva stenophylla Setchell et N.L.Gardner | OR723831 or OR723832 | OR723834 | OR722834 | Hughey et al. (2024) | OR723833 (topotype) |
| Ulva taeniata (Setchell) Setchell et N.L.Gardner | OQ349516 | OQ349516 | OR730542 | Hughey et al. (2024) | |
| Ulva tanneri Hayden et Waaland | OR732837 | OR732838 | OR722835 | Hughey et al. (2024) | |
| Ulva tentaculosa Lagourgue et Payri | MZ902954 | MZ870666 | – | Lagourgue et al. (2022) | |
| Ulva tepida Masakiyo et Shimada | – | – | AB904766 | Masakiyo and Shimada (2014) | |
| Ulva vietnamensis L.-A.T.Tran, Leliaert et De Clerck | MZ582335 | MZ582185 | MZ582100 | Tran et al. (2023) |
-
Associated accession # are indicated when additional sequences have been listed as representatives of the type; - indicates no type sequence is available for that locus. aNot included in phylogenetic trees.
Ulva species diversity has recently been assessed from multiple areas along the east coast of the USA using a combination of DNA sequence data, morphological data, and historical reports. Bloom-forming species in Rhode Island were examined using nuclear-encoded ITS1-based and plastid-encoded rbcL-based operational taxonomic units (OTUs) (Guidone et al. 2013). Nine OTUs were identified, some of which were assigned species names, some were assigned to species complexes, and some could not be identified. One of the OTU’s was previously unreported in the area, increasing local diversity. The ITS region has also been used in the northeastern USA to identify four blade-forming species of Ulva in the Great Bay Estuarine System, three more than had previously been reported (Hofmann et al. 2010). A molecular diversity analysis of the northern Gulf of Mexico included specimens with a wide geographic distribution, and of the 16 species clades defined (13 named and 3 unnamed putative species), eight of them included specimens collected from the southeastern USA coast, including the present region of study (Melton and Lopez-Bautista 2021). Species names were placed on specimens in this study based on tufA, rbcL, and ITS sequences that matched GenBank sequences. Only six of the 16 species that Melton and Lopez-Bautista (2021) identified were based on sequence homology to type specimens, and only one of those, U. lactuca, had been historically reported in North Carolina.
The classification of Ulva species in North Carolina has a long and complicated history (Figure 1). All names used in the literature were based on morphological characters, and the most recent floristic treatments predate the Hayden et al. (2003) synonymy of Enteromorpha under Ulva. The first report of Ulva species in North Carolina included two species of Enteromorpha and three of Ulva (Curtis 1867). Subsequent research and floristic publications recorded eight additional species prior to 1984 (Aziz and Humm 1962; Blomquist and Humm 1946; Hoyt 1920; Rhyne 1973; Williams 1948). The most recent flora published for the area is Schneider and Searles (1991), which mirrored the Enteromorpha and Ulva treatment by Kapraun (1984). After updating the classification and names of the taxa in these two publications, there are currently 11 Ulva and two Blidingia Kylin species reported in North Carolina. However, as previously discussed it is difficult to know whether historical accounts of Ulva species reported from North Carolina are correct or incorrect. These species were identified using a variety of different morphological characters (e.g., Kapraun 1984; Schneider and Searles 1991), but they have never been characterized by DNA sequences. The objectives of this study were: 1) to determine the diversity of North Carolina Ulva species based on DNA sequences; 2) to determine if there is utility in morphological and anatomical characters for the identification of North Carolina species, 3) to determine, if possible, the correct application of specific epithets for North Carolina Ulva taxa, and 4) to provide a baseline, based on DNA sequenced specimens, of Ulva species in North Carolina.
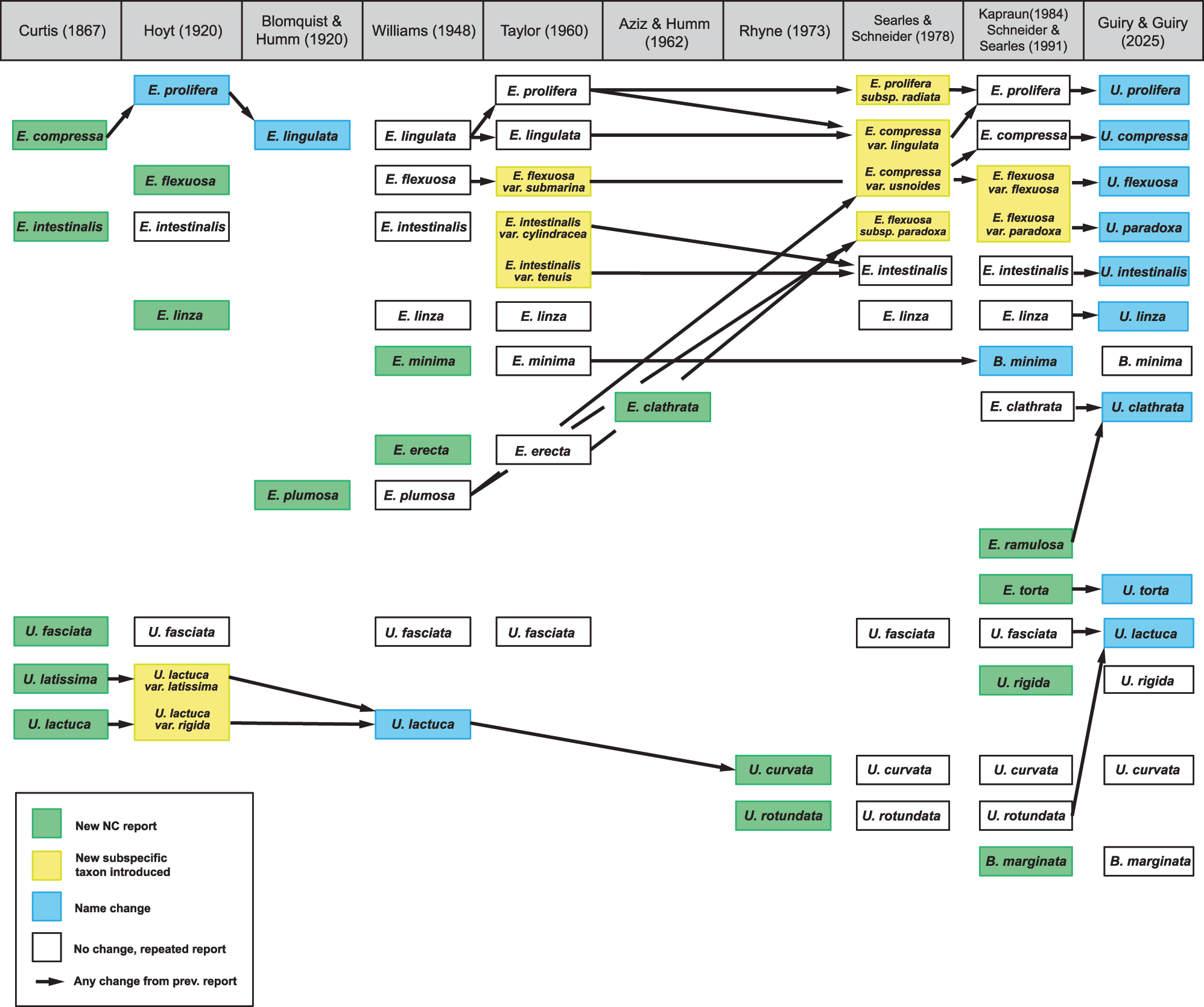
Taxonomic history of North Carolina Ulvaceae species.
2 Materials and methods
2.1 Collection and preservation
Samples were collected during 2022 and 2023 from various locations along the coast of North Carolina (Figure 2). Collected specimens were quickly dried in silica gel desiccant (Chase and Hills 1991) and pressed on herbarium paper in the field when they could not be brought back to the lab for processing. When processed in the lab, a 2 mm2 fragment was placed in 70 µl of sterile water and frozen for later DNA extractions. The remaining material was used for morphological analyses and pressed on herbarium paper with vouchers deposited in WNC (herbarium acronyms according to Index Herbariorum online, Thiers 2025). Sample metadata was submitted to the “Marine Algae of North Carolina” Barcode of Life Database project (BOLDsystems; Ratnasingham and Hebert 2007) and is publicly available at dx.doi.org/10.5883/DS-NCULVA.

Location of sites where Ulva species were collected along the North Carolina coast.
2.2 Morphological assessment
Twenty-five morpho-anatomical characters and their character states (Table 2) were identified for analyses using previous descriptions and keys of North Carolina Ulva species (Kapraun 1984; Schneider and Searles 1991). These included characters generally used in morphological treatments of Ulva [including Enteromorpha] (e.g. Brodie et al. 2007; Norris 2010). Samples were examined using an Olympus SZH (Olympus, Tokyo, Japan) dissecting microscope, and both whole-mount and cross-section slides were made and examined using a Nikon Labophot-2 compound microscope (Nikon, Tokyo, Japan). Character states for the morpho-anatomical characters listed in Table 2 were compiled for every specimen and the consistency of states determined within species as defined by DNA sequences. All continuous characters were plotted as single-axis scatter plots to visually determine where character states could be designated by gaps in the data. All specimens from which morpho-anatomical characters were reported, were also sequenced.
Twenty-five characters and their character states used for morpho-anatomical analyses of North Carolina Ulva species.
| Characters and character states |
|---|
|
2.3 Phylogenetic analyses of DNA sequences
DNA was extracted from each sample using a modified protocol of the MyTaq Extract-PCR Kit (meridian Bioscience, Memphis, TN, USA) following the protocol of Kuba et al. (2022) with small modifications as follows. Samples were incubated in 100 µl of extraction solution at 75 °C, and ground with a microcentrifuge tube pellet pestle every 5–10 min until the tissue was broken up, usually about 20–30 min. The extraction buffer enzymes were denatured at 95 °C for 10 min before centrifugation to pellet cell debris, and the DNA solution diluted 1:5 with water before PCR. Loci to be analyzed were amplified in 12.5 µl PCR reactions containing 1–5 ng of template DNA, 0.4 µM of each amplification primer and MyTaq HS Red DNA polymerase and buffer per manufactures suggestions (meridian Bioscience) and using the thermocycling protocol of Kuba et al. (2022). PCR was performed to amplify the internal transcribed spacer ITS1-5.8S-ITS2 region (ITS) and the rbcL and tufA markers using the primers of Shimada et al. (2003) for ITS and rbcL and Fama et al. (2002) for tufA). Amplification products were cleaned using ExoSap (Thermo Fisher Scientific, Grand Island, NY, USA) and sequenced in both directions using BDX64 enhancing buffer (Molecular Cloning Lab [MCLAB], San Francisco, CA, USA) and BigDye Terminator v.3.1 (Thermo Fisher Scientific) following the MACLAB BDX64 protocol; cleaned using the Zymo ZR DNA Sequencing Clean-up Kit (Zymo Research, Irvine, CA), and run on an ABI3500 Genetic Analyzer (Thermo Fisher Scientific). Sequences were edited and contigs assembled using the Sequencher 5.2.4 software package (GeneCodes Corp., Ann Arbor, MI, USA) or the De Novo Assembly program in Geneious Prime (Biomatters, Auckland, New Zealand).
North Carolina Ulva sequences for each locus were aligned using MUSCLE (Edgar 2004) as implemented in Geneious Prime and adjusted by eye. UPGMA cluster diagrams were created for each locus to identify sequence-based species, and Basic Local Alignment Search Tool (BLAST) searches of GenBank were conducted to identify other Ulva specimens with closely homologous sequences and to determine if any of the North Carolina species could be named based on similarity to sequences from type specimens.
Each haplotype present in North Carolina specimens was included in the phylogenetic analyses. Sequences were aligned as described above using sequences from species clades in the Fort et al. (2022) phylogenetic trees. Additional sequences from the southeastern USA coast (Melton and Lopez-Bautista 2021), most all currently available type sequences (Table 1), and three Umbraulva E.H.Bae et I.K.Lee species as outgroups were also included in these analyses. Accession numbers and detailed sample information, where available, are provided in Supplementary Table S1. ITS was not used for phylogenetic analyses because of uncertain homology among insertion/deletion associated sites. Maximum likelihood (ML) and Bayesian inference (BI) analyses were carried out using the randomized axelerated maximum likelihood (RAxML) (Stamatakis et al. 2005) and MrBayes (Huelsenbeck and Ronquist 2001) Geneious plugins, respectively. RAxML was run using a rapid hill-climbing algorithm for 10 random starting trees with the GTR GAMMA I model, and data partitioned by codon position. Support for nodes was determined through 1000 replications of bootstrapping (BS) using the rapid bootstrapping algorithm and same model and data partitioning as above. Bayesian inference analyses were performed with MrBayes using the GTR + gamma + invariable sites model and two simultaneous runs with 4 (3 heated and 1 cold) Monte-Carlo Markov chains for 4,000,000 generations, sampling every 3000 generations, and with a burn-in value of 1,000,000 generations.
3 Results
A total of 114 Ulva specimens were collected from 52 locations along the coast of North Carolina (Figure 2). Previously unpublished sequences of an additional six Ulva specimens were included in DNA sequence analyses, but these specimens were not analyzed morphologically. DNA was extracted from all specimens, and 97 tufA sequences, 119 rbcL sequences, and 106 ITS sequences were generated and aligned. UPGMA cluster analyses were used to initially delimit nine sequence-based species (Supplementary Figures S1-S3). Phylogenetic analyses including each unique North Carolina rbcL and tufA sequence also resolved nine species among the collected Ulva specimens (Figures 3 and 4, Supplementary Figures S4 and S5).

Maximum likelihood tufA tree with bootstrap and Bayesian posterior probability values shown for branches when greater than 50 % and 0.75, respectively. North Carolina samples are indicated by green lettering. Collapsed clades are indicated with a black triangle and are labeled with information for a representative specimen and the number of additional specimens in the clade. Species epithets applied to sequences that have not been verified by comparisons with type specimen sequences are shown in single quotes.

Maximum likelihood rbcL tree with bootstrap and Bayesian posterior probability values shown for branches when greater than 50 % and 0.75, respectively. North Carolina samples are indicated by green lettering. Collapsed clades are indicated with a black triangle and are labeled with information for a representative specimen and the number of additional specimens in the clade. Species epithets applied to sequences that have not been verified by comparisons with type specimen sequences are shown in single quotes.
Phylogenetic analyses of Ulva rbcL and tufA sequences resolved the North Carolina specimens within moderately to fully supported species clades (Figures 3 and 4, Supplementary Figures S4 and S5). Seven of the nine North Carolina Ulva species have names. The most common ITS, rbcL and tufA sequences from Ulva sp. NC 1 specimens were identical to those of the holotype specimen of Ulva gigantea (Kützing) Bliding (GenBank accessions OR428167, OR427886, OR427287). One of the ITS sequences of Ulva sp. NC 2 was an exact match to that of the U. prolifera O.F.Müller epitype (GenBank accession AJ012276 but see discussion). Ulva sp. NC 5 was confirmed to be Ulva tepida Masakiyo et S.Shimada based on one of the two ITS2 sequences matching that of the holotype specimen (GenBank accession AB904766) and the other being only 1 bp different (0.39 %). Ulva sp. NC 6 was confirmed to be U. lactuca. One of the North Carolina rbcL haplotypes was identical to the sequence of the Florida, USA specimen (GenBank accession KT882614) designated by Hughey et al. (2019) as being representative of rbcL from the U. lactuca lectotype, which has large unsequenced segments throughout it. The rbcL sequence of Ulva sp. NC 7 was 1 bp (0.07 %) different from that of the U. ohnoi Hiraoka et S.Shimada holotype, and their tufA sequences were identical (GenBank accession AP018696 [plastid genome]). Both the rbcL and tufA sequences of Ulva sp. NC 8 were identical to those from the lectotype of U. lacinulata (Kützing) Wittrock (GenBank accession MW543061 [plastid genome]). Using tufA analysis, Ulva sp. NC 4 is tentatively named U. ‘torta’ Trevisan as one of the haplotypes present in this species was 1 bp (0.07 %) different from sequences of specimens considered to be topotypes by Steinhagen et al. (2019b).
None of the 25 analyzed morphological characters had consistent character states for any of the nine sequence-based species (Table 3). Even when examined within the individual species, the number of characters with consistent character states was only two to thirteen, albeit the higher numbers of consistent characters were only for species with two or three collected samples. Morphological identifications of the studied specimens using the Kapraun (1984) key for North Carolina Ulva revealed that the sequence-based species included specimens that fit the descriptions of multiple species. The number of morphology-based species identifications within a sequence-based species ranged from seven named species in Ulva gigantea (sp. NC 1) (n = 46 specimens) and Ulva prolifera (sp. NC 2) (n = 21) to two named species in Ulva tepida (sp. NC 5) (n = 3 specimens) and U. lacinulata (sp. NC 8) (n = 2 specimens).
States of analyzed morphological characters for sampled North Carolina Ulva specimens.
| Specimen ID | Ch 1 | Ch 2 | Ch 3 | Ch 4 | Ch 5 | Ch 6 | Ch 7 | Ch 8 | Ch 9 | Ch 10 | Ch 11 | Ch 12 | Ch 13 | Ch 14 | Ch 15 | Ch 16 | Ch 17 | Ch 18 | Ch 19 | Ch 20 | Ch 21 | Ch 22 | Ch 23 | Ch 24 | Ch 25 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Ulva sp. NC1 - Ulva gigantea | |||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||
| NCweed-2052 | 0 | 1 | N/A | 1 | 0 | 0 | 0 | 1 | 1 | N/A | N/A | N/A | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 1 | 3 | N/A | N/A | 2 | 1 |
| NCweed-2053 | 0 | 1 | N/A | 1 | 0 | 0 | 0 | 1 | 1 | N/A | N/A | N/A | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 0 | 3 | N/A | N/A | 2 | 1 |
| NCweed-2057 | 1 | 1 | 1 | N/A | N/A | N/A | N/A | N/A | 0 | 2 | 0 | 1 | 1 | 1 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | N/A | N/A | 0 | 4 |
| NCweed-2062 | 0 | 1 | N/A | 2 | 0 | 0 | 0 | 1 | 1 | N/A | N/A | N/A | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0/1 | N/A | N/A | 2 | 1 |
| NCweed-2067 | 0 | 0 | N/A | 1 | 1 | 0 | 0 | 1 | 1 | N/A | N/A | N/A | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 4 | 0 | N/A | N/A | 2 | 1 |
| NCweed-2068 | 0 | 0 | N/A | 1 | 0 | 0 | 0 | 1 | 1 | N/A | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 4 | 3 | N/A | N/A | 2 | 4 |
| NCweed-2071 | 0 | 0 | N/A | 1 | 0 | 0 | 0 | 1 | 1 | N/A | N/A | N/A | 0 | 1 | 0 | 0 | 0/2 | 1 | 1 | 2 | 3 | N/A | N/A | 2 | 4 |
| NCweed-2073 | 0 | 1 | N/A | 1 | 0 | 0 | 0 | 1 | 1 | N/A | N/A | N/A | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 0 | 2 | N/A | N/A | 2 | 4 |
| NCweed-2076 | 1 | 0 | 1 | N/A | N/A | N/A | N/A | N/A | N/A | N/A | N/A | N/A | N/A | 0 | 0 | 0 | 0 | 1 | 1 | 4 | 3 | N/A | N/A | 1 | 4 |
| NCweed-2077 | 0 | 2 | N/A | 2 | 0 | 0 | 0 | 1 | 1 | N/A | N/A | N/A | 0 | 0 | 1 | 0 | 0 | 1 | 1 | 4 | 0 | N/A | N/A | 2 | 4 |
| NCweed-2078 | 0 | 2 | N/A | 2 | 0 | 0 | 0 | 1 | 1 | N/A | N/A | N/A | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 4 | 2 | N/A | N/A | 2 | 4 |
| NCweed-2079 | 0 | 2 | N/A | 2 | 0 | 0 | 0 | 1 | 1 | N/A | N/A | N/A | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 4 | 1 | N/A | N/A | 2 | 4 |
| NCweed-2080 | 2 | 1 | N/A | 0 | 0 | 1 | 0 | 0 | 1 | N/A | N/A | N/A | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 4 | 3 | 2 | N/A | 1 | 0 |
| NCweed-2081 | 2 | 1 | N/A | 0 | 0 | 1 | 0 | 0 | 1 | N/A | N/A | N/A | 0 | 1 | 1 | 1 | 0/2 | 2 | 2 | 4 | 2 | N/A | 0 | 1 | 0 |
| NCweed-2083 | 1 | 0 | 1 | N/A | N/A | N/A | N/A | N/A | 1 | N/A | N/A | N/A | 0 | 1 | 0 | 0 | 0 | 2 | 2 | 4 | 2 | N/A | N/A | 1 | 0 |
| NCweed-2090 | 0 | 2 | N/A | 2 | 0 | 0 | 0 | 1 | 1 | N/A | N/A | N/A | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 4 | 2 | N/A | N/A | 2 | 4 |
| NCweed-2091 | 0 | 0 | N/A | 1 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 0/1 | 1 | 1 | 4 | 2 | N/A | N/A | 2 | 4 |
| NCweed-2094 | 0 | 0 | N/A | 1 | 0 | 0 | 0 | 1 | 1 | 1 | N/A | N/A | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | N/A | N/A | 2 | 2 |
| NCweed-2095 | 0 | 1 | N/A | 1 | 0 | 0 | 0 | 1 | 1 | N/A | N/A | N/A | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 2 | 0 | N/A | N/A | 2 | 2 |
| NCweed-2096 | 0 | 0 | N/A | 1 | 0 | 0 | 0 | 1 | 1 | N/A | N/A | N/A | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 2 | 0 | N/A | N/A | 2 | 1 |
| NCweed-2097 | 1 | 1 | 1 | N/A | N/A | N/A | N/A | N/A | 1 | N/A | N/A | N/A | 0 | 1 | 0 | 0 | 0 | 1 | 2 | 4 | 2 | N/A | N/A | 1 | 0 |
| NCweed-2101 | 0 | 1 | N/A | 2 | 0 | 0 | 0 | 1 | 1 | 1 | N/A | N/A | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 3 | 1 | N/A | 2 | 4 |
| NCweed-2103 | 0 | 0 | N/A | 1 | 0 | 0 | 0 | 1 | 1 | 1 | N/A | N/A | 0 | 0 | 0 | 1 | 0/1 | 2 | 1 | 1 | 1 | N/A | N/A | 2 | 0 |
| NCweed-2113 | 1 | 1 | 1 | N/A | N/A | N/A | N/A | N/A | 1 | N/A | N/A | N/A | 0 | 2 | 0 | 1 | 0/1 | 1 | 1 | 5 | 2 | N/A | 1 | 1 | 0 |
| NCweed-2118 | 0 | 0 | N/A | 1 | 0 | 0 | 0 | 1 | 1 | N/A | N/A | N/A | 0 | 1 | 0 | 0 | 0/1 | 1 | 1 | 4 | 0 | N/A | N/A | 2 | 1 |
| NCweed-2121 | 0 | 1 | N/A | 1 | 0 | 0 | 0 | 1 | 1 | N/A | N/A | N/A | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 2 | 2 | N/A | 2 | 2 |
| NCweed-2125 | 0 | 1 | N/A | 1 | 0 | 0 | 0 | 1 | 1 | N/A | N/A | N/A | 0 | 0 | 0 | 0 | 0/1 | 1 | 1 | 0 | 0 | N/A | N/A | 2 | 2 |
| NCweed-2127 | 0 | 1 | N/A | 1 | 0 | 0 | 1 | 1 | 1 | N/A | N/A | N/A | N/A | 1 | 0 | 1 | 0 | 1 | 1 | 4 | 2 | N/A | N/A | 2 | 4 |
| NCweed-2154 | 0 | 0 | N/A | 0 | 0 | 0 | 0 | 0 | 1 | N/A | N/A | N/A | 0 | 0 | 0 | 0 | 0/1 | 1 | 1 | 2 | 0 | N/A | N/A | 1 | 2 |
| NCweed-2155 | 0 | 0 | N/A | 0 | 0 | 0 | 0 | 1 | 1 | N/A | N/A | N/A | 0 | 1 | 0 | 1 | 0 | 1 | 2 | 4 | 2 | N/A | N/A | 2 | 4 |
| NCweed-2184 | 0 | 0 | N/A | 0 | 0 | 0 | 0 | 0 | 1 | 1 | N/A | N/A | N/A | 1 | 0 | 0 | 0 | 1 | 1 | 1 | 0/2 | N/A | N/A | 1 | 0 |
| NCweed-2190 | 0 | 2 | N/A | 2 | 0 | 0 | 0 | 2 | 1 | N/A | N/A | N/A | 0 | 0 | 2 | 1 | 0 | 1 | 1 | 1 | 0 | N/A | N/A | 2 | 1 |
| NCweed-2192 | 0 | 2 | N/A | 1 | 0 | 0 | 0 | 0 | 1 | N/A | N/A | N/A | 0 | 1 | 0 | 1 | 0 | 1 | 1 | 4 | 2 | N/A | N/A | 1 | 2 |
| NCweed-2194 | 0 | 1 | N/A | 0 | 0 | 0 | 0 | 1 | 1 | N/A | N/A | N/A | 0 | 1 | 0 | 1 | 0 | 1 | 1 | 1 | 0 | N/A | N/A | 2 | 4 |
| NCweed-2203 | 0 | 0 | N/A | 0 | 0 | 0 | 0 | 1 | 1 | N/A | N/A | N/A | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 0 | N/A | N/A | 2 | 4 |
| NCweed-2205 | 0 | 0 | N/A | 1 | 0 | 0 | 0 | 1 | 1 | N/A | N/A | N/A | 0 | N/A | 0 | 0 | 0 | 0 | 1 | 1 | 0 | N/A | N/A | 2 | 1 |
| NCweed-2206 | 0 | 1 | N/A | 1 | 0 | 0 | 0 | 1 | 1 | N/A | N/A | N/A | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 1 | 0 | N/A | N/A | 2 | 4 |
| NCweed-2209 | 0 | 2 | N/A | 2 | 0 | 0 | 0 | 1 | 1 | N/A | N/A | N/A | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 0 | N/A | N/A | 2 | 4 |
| NCweed-2217 | 0 | 0 | N/A | 1 | 0 | 0 | 0 | 1 | 1 | N/A | N/A | N/A | 0 | 1 | 0 | 0 | 0/2 | 1 | 1 | 2 | 3 | 2 | N/A | 2 | 2 |
| NCweed-2220 | 0 | 1 | N/A | 1 | 0 | 0 | 1 | 2 | 1 | N/A | N/A | N/A | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | N/A | N/A | 2 | 2 |
| NCweed-2222 | 0 | 1 | N/A | 2 | 0 | 0 | 0 | 0 | 1 | N/A | N/A | N/A | 0 | 0 | 0 | 0 | 0 | 2 | 1 | 4 | 0 | N/A | N/A | 2 | 2 |
| NCweed-2224 | 0 | 2 | N/A | 1 | 0 | 0 | 0 | 1 | 1 | N/A | N/A | N/A | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 2 | 2 | N/A | N/A | 2 | 4 |
| NCweed-2226 | 0 | 0 | N/A | 1 | 0 | 0 | 0 | 1 | 1 | N/A | N/A | N/A | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 3 | 3 | 2 | N/A | 2 | 4 |
| NCweed-2231 | 0 | 1 | N/A | 1 | 0 | 0 | 0 | 1 | 1 | N/A | N/A | N/A | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | N/A | N/A | 2 | 4 |
| NCweed-2232 | 0 | 0 | N/A | 1 | 0 | 0 | 1 | 2 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 0 | 0/2 | 1 | 1 | 4 | 0 | N/A | N/A | 2 | 0/3 |
| NCweed-2236 | 0 | 2 | N/A | 1 | 0 | 0 | 0 | 1 | 1 | N/A | N/A | N/A | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 2 | 1 | N/A | N/A | 2 | 4 |
|
|
|||||||||||||||||||||||||
| Ulva sp. NC2 - Ulva prolifera | |||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||
| NCweed-2054 | 2 | 0 | N/A | 0 | 0 | 1 | 0 | 0 | 1 | N/A | N/A | N/A | 0 | 1 | 0 | 1 | 0 | 0 | 1 | 4 | 3 | 2 | N/A | N/A | 2 |
| NCweed-2069 | 1 | 0 | 1 | N/A | 0 | 0 | 0 | N/A | 0 | 2 | 0 | 0 | 1 | 2 | 0 | 0 | 0 | 1 | 1 | 4 | 0 | N/A | N/A | 1 | 4 |
| NCweed-2070 | 0 | 0 | N/A | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 2 | 3 | N/A | N/A | 1 | 2 |
| NCweed-2072 | 1 | 1 | 1 | N/A | N/A | N/A | N/A | N/A | 0 | 0 | 0/1 | 0 | 2 | 1 | 0 | 0 | 0 | 1 | 1 | 2 | 2 | N/A | 1 | 1 | 0 |
| NCweed-2074 | 2 | 1 | N/A | 0 | 0 | 1 | 0 | 0 | 1 | N/A | N/A | N/A | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 4 | 3 | N/A | N/A | 2 | 2 |
| NCweed-2075 | 1 | 1 | 1 | N/A | N/A | N/A | N/A | N/A | 2 | 2 | N/A | 0 | N/A | 1 | 0 | 0 | 0/1 | 1 | 1 | 2 | 3 | 1 | N/A | 0 | 0 |
| NCweed-2084 | 0 | 0 | N/A | 1 | 0 | 0 | 0 | 1 | 1 | N/A | 0 | 1 | 0 | 1 | 0 | 0 | 1/3 | 1 | 1 | 2 | 2 | N/A | N/A | 2 | 2 |
| NCweed-2093 | 1 | 0 | 1 | N/A | N/A | N/A | N/A | N/A | 0 | 1 | N/A | 1 | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 2 | 3 | 1 | N/A | 1 | 4 |
| NCweed-2098 | 2 | 0 | N/A | 0 | 0 | 1 | 1 | 0 | 1 | N/A | 0 | 0 | 0 | 1 | 0 | 1 | 0/1 | 1 | 1 | 4 | 0 | N/A | N/A | 0 | 2 |
| NCweed-2102 | 1 | 0 | 1 | N/A | N/A | N/A | N/A | N/A | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0/1 | 1 | 1 | 4 | 0 | N/A | N/A | 1 | 2 |
| NCweed-2117 | 1 | 0 | 0 | N/A | N/A | N/A | N/A | N/A | 1 | N/A | N/A | N/A | 0 | 1 | 0 | 0 | 1 | 1 | 1 | 5 | 2 | 2 | 1 | 1 | 4 |
| NCweed-2202 | 1 | 1 | 0 | N/A | N/A | N/A | N/A | N/A | 0 | 1 | 1 | 0 | 2 | 0 | 0 | 0 | 0 | 1 | 1 | 5 | 0 | N/A | 1 | 0 | 0 |
| NCweed-2207 | 1 | 1 | 1 | N/A | N/A | N/A | N/A | N/A | 1 | N/A | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 2 | 0 | N/A | N/A | 0 | 3 |
| NCweed-2208 | 1 | 0 | 0 | N/A | N/A | N/A | N/A | N/A | 0 | 2 | N/A | 1 | 1 | 1 | 0 | 0 | 0/1 | 1 | 1 | 3 | 0 | N/A | 1 | 1 | 0 |
| NCweed-2210 | 2 | 2 | N/A | 0 | 0 | 1 | 1 | 2 | 1 | N/A | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 1 | 1 | 2 | 0 | N/A | N/A | 1 | 3 |
| NCweed-2214 | 1 | 2 | 2 | N/A | N/A | N/A | N/A | N/A | 0 | 0 | 1 | 0 | 2 | 0 | 0 | 0 | 0 | 1 | 1 | 2 | 0 | N/A | 2 | 1 | 0 |
| NCweed-2216 | 2 | 1 | N/A | 0 | 0 | N/A | 2 | 0 | 1 | N/A | N/A | N/A | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 4 | 0 | N/A | N/A | 0 | 0 |
| NCweed-2225 | 1 | 2 | 2 | N/A | N/A | N/A | N/A | N/A | 0 | 2 | N/A | 1 | 1 | 1 | 1 | 0 | 0/1 | 1 | 1 | 1 | 0 | N/A | N/A | 1 | 0 |
| NCweed-2227 | 1 | 1 | 2 | N/A | N/A | N/A | N/A | N/A | 0 | 2 | N/A | 0 | 1 | 2 | 0 | 1 | 0 | 1 | 1 | 2 | 2 | N/A | N/A | 0 | 0 |
| NCweed-2228 | 2 | 0 | N/A | 0 | N/A | N/A | N/A | N/A | 1 | N/A | 0 | 1 | 0 | 2 | 0 | 0 | 0 | 1 | 1 | 1 | 0 | N/A | N/A | 0 | 3 |
| NCweed-2229 | 2 | 0 | N/A | 0 | 0 | 0 | 0 | 0 | 1 | N/A | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 1 | 1 | 2 | 0 | N/A | N/A | 2 | 3 |
|
|
|||||||||||||||||||||||||
| Ulva sp. NC3 | |||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||
| NCweed-2050 | 2 | 2 | N/A | 0 | 0 | 1 | 0 | 0 | 1 | N/A | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 2 | 1 | 4 | 3 | 2 | N/A | 1 | 2 |
| NCweed-2051 | 1 | 1 | 0 | N/A | N/A | N/A | N/A | N/A | 0 | 0 | N/A | N/A | 2 | N/A | 1 | 1 | 0/1 | 1 | 2 | 0 | 3 | N/A | 1 | 1 | 4 |
| NCweed-2060 | 1 | 2 | 2 | N/A | N/A | N/A | N/A | N/A | 0 | 1 | 1 | 1 | 1 | 1 | 0 | 0 | 0/3 | 1 | 0 | 3 | 0 | N/A | 0 | 1 | 4 |
| NCweed-2061 | 1 | 1 | 1 | N/A | N/A | N/A | N/A | N/A | 0 | 1 | 1 | 0 | 1 | 0 | 2 | 1 | 3 | 1 | 1 | 0 | 3 | N/A | N/A | 1 | 4 |
| NCweed-2065 | 1 | 1 | 1 | N/A | N/A | N/A | N/A | N/A | 0 | 2 | 0 | 1 | 1 | 1 | 0 | 0 | 0 | 2 | 1 | 4 | 0 | N/A | N/A | 1 | 4 |
| NCweed-2066 | 1 | 2 | 0 | N/A | N/A | N/A | N/A | N/A | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 1 | 1 | 1 | 0 | 3 | 1 | N/A | 1 | 4 |
| NCweed-2092 | 1 | 0 | 1 | N/A | N/A | N/A | N/A | N/A | 1 | N/A | N/A | N/A | 0 | 0 | 0 | 1 | 0/1 | 1 | 1 | 3 | 0 | N/A | 0/1 | 1 | 0 |
| NCweed-2126 | 1 | 1 | 1 | N/A | N/A | N/A | N/A | N/A | 0 | 2 | N/A | N/A | 1 | 1 | 1 | 1 | 0/1 | 1 | 1 | 0 | 3 | N/A | N/A | 1 | 0 |
| NCweed-2182 | 1 | 1 | 0 | N/A | N/A | N/A | N/A | N/A | 1 | N/A | N/A | N/A | N/A | 2 | 0 | 1 | 0 | 1 | 1 | 1 | 2 | N/A | N/A | 1 | 0 |
| NCweed-2183 | 1 | 1 | 1 | N/A | N/A | N/A | N/A | N/A | 1 | 1 | N/A | N/A | N/A | 1 | 0 | 1 | 0/2 | 1 | 1 | 1 | 2 | N/A | N/A | 0 | 0 |
| NCweed-2187 | 1 | 2 | 0 | N/A | N/A | N/A | N/A | N/A | 0 | 1 | 1 | 0 | 2 | 0 | 1 | 2 | 0/1 | 1 | 2 | 4 | 2 | N/A | 1 | 1 | 0 |
| NCweed-2188 | 1 | 1 | 1 | N/A | N/A | N/A | N/A | N/A | 0 | 2 | N/A | N/A | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 1 | 2 | 2 | N/A | 1 | 0 |
| NCweed-2191 | 1 | 2 | 2 | N/A | N/A | N/A | N/A | N/A | 0 | 1 | 1 | 0 | 2 | 1 | 1 | 1 | 0 | 1 | 1 | 4 | 0 | N/A | 0/1 | 1 | 0 |
| NCweed-2193 | 1 | 2 | 0 | N/A | N/A | N/A | N/A | N/A | 0 | 1 | 1 | 0 | 2 | 1 | 0 | 0 | 0 | 1 | 1 | 1 | 2 | N/A | 0/1 | 1 | 0 |
| NCweed-2195 | 1 | 1 | 1 | N/A | N/A | N/A | N/A | N/A | 0 | 1 | 1 | 1 | 2 | 2 | 1 | 1 | 0 | 1 | 1 | 1 | 2 | N/A | 1 | 0 | 0 |
| NCweed-2198 | 1 | 0 | 2 | N/A | N/A | N/A | N/A | N/A | 0 | 0 | 1 | 0 | 3 | 1 | 0 | 1 | 0 | 1 | 1 | 1 | 0 | N/A | N/A | 0 | 0 |
| NCweed-2199 | 1 | 1 | 1 | N/A | N/A | N/A | N/A | N/A | 0 | 1 | 0 | 0 | 1 | 1 | 0 | 1 | 0 | 1 | 1 | 0 | 0 | N/A | N/A | 0 | 0 |
| NCweed-2201 | 1 | 1 | 1 | N/A | N/A | N/A | N/A | N/A | 0 | 1 | N/A | 0 | 2 | 1 | 1 | 1 | 0/1 | 1 | 1 | 2 | 0 | N/A | N/A | 0 | 0 |
| NCweed-2204 | 1 | 1 | 0 | N/A | N/A | N/A | N/A | N/A | 0 | 0 | 1 | 0 | 3 | 0 | 1 | 1 | 0/1 | 0 | 1 | 0 | 3 | 2 | N/A | 0 | 0 |
| NCweed-2218 | 1 | 2 | 0 | N/A | N/A | N/A | N/A | N/A | 0 | 2 | N/A | 0 | 1 | 2 | 0 | 0 | 0 | 1 | 1 | 0 | 3 | 2 | N/A | 0 | 0 |
| NCweed-2221 | 2 | 0 | N/A | 0 | 0 | 0 | 1 | 2 | 1 | N/A | N/A | N/A | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | N/A | N/A | 1 | 3 |
| NCweed-2223 | 1 | 2 | 0 | N/A | N/A | N/A | N/A | N/A | 0 | 0 | 1 | 0 | 2 | 2 | 1 | 1 | 0 | 0 | 1 | 2 | 1 | N/A | 0/1 | 1 | 0 |
|
|
|||||||||||||||||||||||||
| Ulva sp. NC4 - Ulva ‘torta’ | |||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||
| NCweed-2063 | 1 | 1 | 2 | N/A | N/A | N/A | N/A | N/A | 0 | 1 | N/A | N/A | 1 | 1 | 1 | 1 | 0/2 | 1 | 1 | 2 | 0 | N/A | N/A | 1 | 4 |
| NCweed-2099 | 1 | 1 | 0 | N/A | N/A | N/A | N/A | N/A | 0 | 1 | 1 | 0 | 2 | 1 | 1 | 1 | 1 | 1 | 1 | 5 | 3 | 2 | N/A | 1 | 0 |
| NCweed-2100 | 1 | 1 | 0 | N/A | N/A | N/A | N/A | N/A | 0 | 1 | 1 | 0 | 2 | 2 | 1 | 2 | 0/1 | 1 | 2 | 0 | 3 | 2 | 1 | 1 | 0 |
| NCweed-2120 | 1 | 1 | 1 | N/A | N/A | N/A | N/A | N/A | 0 | 2 | N/A | N/A | 1 | N/A | 1 | 1 | 0/1 | 1 | 2 | 0 | 3 | N/A | N/A | 1 | 0 |
| NCweed-2215 | 1 | 0 | 1 | N/A | N/A | N/A | N/A | N/A | 0 | 2 | N/A | N/A | 1 | 1 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | N/A | N/A | 1 | 0 |
| NCweed-2230 | 1 | 0 | 1 | N/A | N/A | N/A | N/A | N/A | 0 | 1 | 1 | 1 | 1 | 2 | 0 | 1 | 0 | 1 | 2 | 0 | 2 | N/A | 2 | 1 | 0 |
| NCweed-2234 | 1 | 0 | 0 | N/A | N/A | N/A | N/A | N/A | 0 | 0 | 1 | 0 | 2 | 2 | 0 | 1 | 0/1 | 0 | 1 | 0 | 1 | N/A | 1 | 0 | 0 |
|
|
|||||||||||||||||||||||||
| Ulva sp. NC5 - Ulva tepida | |||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||
| NCweed-2112 | 1 | 0 | 1 | N/A | N/A | N/A | N/A | N/A | 0 | N/A | N/A | N/A | 1 | 2 | 1 | 1 | 0/1 | 0 | 1 | 2 | 2 | N/A | 2 | 0 | 0 |
| NCweed-2159 | 1 | 0 | 0 | N/A | N/A | N/A | N/A | N/A | 0 | 1 | 1 | 1 | 2 | 1 | 0 | 1 | 0 | 0 | 1 | 0 | 3 | N/A | N/A | 1 | 0 |
| NCweed-2160 | 1 | 0 | 0 | N/A | N/A | N/A | N/A | N/A | 2 | 2 | 0 | 0 | 1 | N/A | 1 | 1 | 1 | 0 | 2 | 6 | 3 | N/A | N/A | 0 | 0 |
|
|
|||||||||||||||||||||||||
| Ulva sp. NC6 - Ulva lactuca | |||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||
| NCweed-2055 | 0 | 1 | N/A | 0 | 1 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 1 | 2 | 0 | 0 | 0 | 1 | 1 | 4 | 0 | N/A | N/A | 2 | 2 |
| NCweed-2082 | 0 | 1 | N/A | 1 | 0 | 0 | 1 | 2 | 0 | 1 | 0 | 1 | 0 | 1 | 1 | 1 | 0/2 | 1 | 1 | 1 | 3 | 1 | N/A | 2 | 4 |
| NCweed-2109 | 0 | 1 | N/A | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 2 | 1 | 0 | 1 | 0/2 | 1 | 2 | 1 | 1 | N/A | N/A | 1 | 2 |
| NCweed-2111 | 0 | 1 | N/A | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 2 | 1 | 0 | 1 | 0 | 2 | 2 | 4 | 2 | N/A | N/A | 1 | 2 |
| NCweed-2114 | 0 | 0 | N/A | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 2 | 0 | 2 | 0 | 0 | 1 | 1 | 3 | N/A | N/A | 2 | 2 |
| NCweed-2149 | 0 | 1 | N/A | 2 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 2 | 1 | 1 | 0/2 | 2 | 2 | 4 | 2 | N/A | N/A | 2 | 1 |
| NCweed-2150 | 0 | 0 | N/A | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 2 | 0 | 1 | 0 | 2 | 2 | 4 | 1 | N/A | N/A | 2 | 4 |
| NCweed-2152 | 0 | 0 | N/A | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 0 | 1 | 2 | 4 | 1 | N/A | N/A | 1 | 2 |
| NCweed-2212 | 0 | 2 | N/A | 0 | 0 | 0 | 1 | 2 | 0 | 1 | 0 | 1 | 0 | 1 | 1 | 2 | 0/2 | 1 | 2 | 2 | 1/2 | 2 | N/A | 2 | 4 |
| NCweed-2233 | 0 | 1 | N/A | 1 | 0 | 0 | 0 | 2 | 0 | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 0 | 1 | 2 | 0 | 2 | N/A | N/A | 2 | 2 |
|
|
|||||||||||||||||||||||||
| Ulva sp. NC7 - Ulva ohnoi | |||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||
| NCweed-2064 | 0 | 0 | N/A | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 1 | 0 | 1 | 1 | 1 | 0 | 1 | 2 | 4 | 3 | 1 | N/A | 2 | 2 |
| NCweed-2177 | 0 | 0 | N/A | 0 | 1 | 0 | 0 | 1 | 0 | 1 | 0 | N/A | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 0 | 3 | N/A | N/A | 2 | 1 |
| NCweed-2186 | 0 | 1 | N/A | 0 | 0 | 0 | 0 | 2 | 0 | 1 | 0 | 1 | 1 | 1 | 1 | 1 | 0 | 1 | 1 | 2 | 0 | N/A | N/A | 2 | 2 |
|
|
|||||||||||||||||||||||||
| Ulva sp. NC8 - Ulva lacinulata | |||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||
| NCweed-2200 | 0 | 2 | N/A | 2 | 0 | 0 | 0 | 1 | 1 | N/A | N/A | N/A | 0 | 1 | 1 | 1 | 0 | 1 | 1 | 0 | 0 | N/A | N/A | 2 | 4 |
| NCweed-2235 | 0 | 2 | N/A | 1 | 0 | 0 | 0 | 1 | 1 | N/A | N/A | N/A | 0 | 0 | 0 | 1 | 0 | 1 | 1 | 2 | 0 | N/A | N/A | 2 | 4 |
|
|
|||||||||||||||||||||||||
| Ulva sp. NC9 - Ulva carsoniae sp. nov. | |||||||||||||||||||||||||
|
|
|||||||||||||||||||||||||
| NCweed-2189 | 0 | 0 | N/A | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 1 | 1 | 0 | 0 | 2 | 0 | 1 | N/A | N/A | 1 | 2 |
-
Characters and character state codes are as in Table 2. N/A, not applicable/undetermined.
3.1 Ulva sp. NC 1 – Ulva gigantea
Ulva gigantea (sp. NC 1) was resolved in a clade that had strong to full support in both the tufA and rbcL analyses (tufA: BS 98; BI posterior probability (PP) 1.0, and rbcL: BS 99; PP 1.0). It was the most frequently collected species in this study (n = 46) and was sampled from one of the southernmost sites and the northernmost site, in brackish to oceanic environments (Figure 5). Morphologically U. gigantea was especially plastic, having bladed, tubular, and ‘linza-type’ morphologies represented (Table 3, Figure 6A–D). Only two characters, tube width (0.1–1.0 cm) and branching pattern (basal) were consistent within this species, but these characters were not applicable to all specimens. These 46 samples also exhibited large variation in maximal length, from less than 10 cm to greater than 20 cm.

Distribution of eight North Carolina Ulva species among the 52 collection sites in this study.

Variation in morphology of North Carolina Ulva specimens. (A–D) U. gigantea: (A) NCweed-2077 sample voucher (WNC-34210), (B) NCweed-2094 sample voucher (WNC-34222), (C) NCweed-2083 sample voucher (WNC-34215), (D) NCweed-2151 sample voucher (WNC-34361). (E–G) U. prolifera: (E) NCweed-2229 sample voucher (WNC-34394), (F) NCweed-2210 sample voucher (WNC-34310), (G) NCweed-2202 sample voucher (WNC-34389). (H–J) Ulva sp. NC3: (H) NCweed-2126 sample voucher (WNC-34254), (I) NCweed-2187 sample voucher (WNC-34317), (J) NCweed-2223 sample voucher (WNC-34407). (K–L) U. ‘torta’: (K) NCweed-2120 sample voucher (WNC-34246), (L) NCweed-2063 sample voucher (WNC-34195), (M) NCweed-2234 sample voucher (WNC-34411).
3.2 Ulva sp. NC 2 - Ulva prolifera
Ulva prolifera (sp. NC 2) was resolved in a clade that had moderate to full (BS 83; PP 1.0) and good to full (BS 90; PP 1.0) support in the tufA and rbcL analyses, respectively. Ulva prolifera was geographically dispersed across the North Carolina coast and found in a variety of habitats with varying salinities (Figure 5). Morpho-anatomically, North Carolina U. prolifera specimens exhibited all three general morphology types: bladed, tubular, and ‘linza-morphotype’, and only morpho-anatomical characters 5, absence of dentate blade margins, and 19, cell length:width ratios, were consistent within the species (Table 3, Figure 6E–G).
3.3 Ulva sp. NC 3
The species clade including Ulva sp. NC 3 was unsupported in the tufA tree, but this clade had good to full (BS 91; PP 1.0) support in the rbcL tree. This species was sister to U. californica Wille. It was found in a variety of habitats and locations along the coast (Figure 5), and morphologically was almost entirely comprised of tubular specimens, save two that had ‘linza-morphotype’ hollow blade margins (Table 3, Figure 6H–J). These two specimens were collected at different mesohaline sound locations with similar salinities.
3.4 Ulva sp. NC 4 - Ulva ‘torta’
Ulva ‘torta’ (sp. NC 4) was resolved in a fully supported tufA tree clade (BS 100; PP 1.0) and was resolved in a poor to fully supported clade in the rbcL tree (BS 71; PP 1.0). Both the Ulva sp. NC 3 and Ulva ‘torta’ species clades were part of a large mostly unsupported clade that also includes U. californica, U. kanagawae V.Carneiro, M.T.Martins et Cassano, U. tanneri H.S.Hayden et Waaland, and other unidentified species. This species was found at southern collection sites in Brunswick County, North Carolina, and as far north as Ocracoke, North Carolina (Figure 5). Most of the samples were collected in protected, brackish environments, save the Outer Banks sample that was found in a euhaline environment. Morphologically, all North Carolina specimens were tubular, four had maximal lengths between 10 and 20 cm, and three had maximal lengths less than 10 cm (Table 3). These latter specimens fit the common description of U. ‘torta’ as small, densely intertwined tubes (Steinhagen et al. 2019b). All were branched, but the maximal width of the tubes and sizes of cells (both in surface view and transection) varied greatly in the North Carolina specimens (Table 3, Figure 6K and L) and exceeded the ranges cited in morpho-anatomical descriptions of U. ‘torta’ in North Carolina (Kapraun 1984; Schneider and Searles 1991).
3.5 Ulva sp. NC 5 - Ulva tepida
Ulva tepida (sp. NC 5) was resolved in a clade that had poor to no (BS 72; PP 0.76) and moderate to full (BS 86; PP 1.0) support in the tufA and rbcL analyses, respectively. This clade was sister to U. chaugulei M.G.Kavale et Kazi and part of a larger clade with little to no support that also included Ulva iliohaha H.L.Spalding et A.R.Sherwood and multiple species recently described from New Caledonia (Lagourgue et al. 2022). The North Carolina U. tepida samples were all from New Hanover County, besides one from Carteret County (Figure 5). This species was found in euhaline and mesohaline environments. Morpho-anatomically, North Carolina samples were tubular, all had maximal lengths less than 10 cm, their cells had length:width ratios of less than 1 in transection, and all had a proliferous base (Table 3, Figure 7A and B).

Variation in morphology of North Carolina Ulva specimens. (A–B) U. tepida: (A) NCweed-2042 sample voucher (WNC-22798), (B) NCweed-2112 sample voucher (WNC-34239). (C–E) U. lactuca: (C) NCweed-2233 sample voucher (WNC-34415), (D) NCweed-2111 sample voucher (WNC-34238), (E) NCweed-2150 sample voucher (WNC-34362). (F–G) U. ohnoi: (F) NCweed-2064 sample voucher (WNC-34196), (G) NCweed-2186 sample voucher (WNC-34316). (H–I) U. lacinulata: (H) NCweed-2200 sample voucher (WNC-34414). (I) NCweed-2111 sample voucher (WNC-34413).
3.6 Ulva sp. NC 6 - Ulva lactuca
Ulva lactuca (sp. NC 6) was resolved in a clade that received strong to full (BS 99; PP 1.0) support in the tufA analysis, but in the rbcL analysis, received no BS support but good PP (0.93) support. This clade included the type sequences for currently recognized species U. lacinulata (Kützing) Wittrock, U. ohnoi Hiraoka et S.Shimada, U. lactuca, U. dactylifera Setchell et N.L.Gardner, U. taeniata Setchell et N.L.Gardner, and U. conglobata Kjellman. Also in this clade were type sequences for the following names, all of which had been synonymized under U. lactuca: U. nematoidea Bory, U. lobata (Kützing) Harvey, and U. rotundata Bliding.
Ulva lactuca was collected from the southern sites in New Hanover and Brunswick Counties (Figure 5). It was found in oceanic and high to low salinity habitats along the Atlantic Intracoastal Waterway. Morphologically, all members of this species were distromatic blades (Table 3, Figure 7C–E), but the shape of blades was highly mixed, with some having a more linear/lanceolate shape, others being more lobed, and some being linear/lanceolate with lobing present along the margins. All branched U. lactuca samples exhibited basal branching, and they ranged from less than 10 cm to greater than 20 cm in length and from less than 5 cm to greater than 15 cm in width. North Carolina specimens also exhibited a wide range of pyrenoid number per cell.
3.7 Ulva sp. NC 7 - Ulva ohnoi
Ulva ohnoi (sp. NC 7) was part of the same clade that included U. lactuca and U. lacinulata. It was positioned in a subclade with U. dactylifera, U. lactuca and U. taeniata, but the relationships within this clade were not supported.
Ulva ohnoi was collected at three sites from the mid-to southern coastal areas of North Carolina (Figure 5). The three collected samples of U. ohnoi shared character states for ten of the 25 observed morpho-anatomical characters (Table 3). They were all narrow blades with obtuse apices, but without hollow or undulating margins (Figure 7F and G). Their cells were polygonal in surface view with length-width ratios of 1–2 in transection and had 1–3 pyrenoids. The three samples were each distinct however in blade shape, presence of dentated margins and basal branching.
3.8 Ulva sp. NC 8 - Ulva lacinulata
Ulva lacinulata was also in the same species clade as U. lactuca and U. ohnoi in rbcL and tufA trees. Relationships among the species within this clade were unsupported, but U. lacinulata as well as U. conglobata were consistently resolved as basal lineages.
Two specimens of U. lacinulata were collected, one from the mid-coast and one from the southern coast (Figure 5). Both were >20 cm long, unbranched blades and only differed in character states for blade width and shape, pyrenoid count, cell diameter, and shape (Table 3, Figure 7H and I).
3.9 Ulva sp. NC 9
This species was represented in this study by a single cluster of blades that was collected at Carrot Island, near Beaufort, North Carolina (Figure 8). Ulva sp. NC 9 was resolved in a fully supported tufA species clade (BS 100; PP 1.0) and was resolved in a strongly to fully supported rbcL species clade (BS 99; PP 1.0). All the resolved species clades that included North Carolina specimens showed a wide global distribution and have samples from outside the United States, apart from that including Ulva sp. NC 9 for which there was only one sequence confirmed specimen from outside of the USA east coast. Ulva sp. NC 9 is here described as a new species.

Collection location and holotype of Ulva carsoniae sp. nov. (Ulva sp. NC9). (A) Site where holotype was collected; (B) holotype (WNC-34319).
3.10 Ulva carsoniae Dibner, P.W.Gabrielson et Freshwater (Figure 8B)
Holotype: WNC- 34319, Collection NCweed-2189, attached to small shell in intertidal of Taylor Creek, eastern end of Carrot Island, Rachel Carson National Estuarine Research Reserve, Carteret Co., North Carolina, 34°42.443′ N 076°37.645′ W, 26 January 2023, leg. S. Dibner and M. A. Lytle.
Description: Thallus a membranous, distromatic blade, light green in color, up to 7.0 cm high and 1.5 cm wide. Blades clavate to lanceolate with smooth, slightly undulating margins. Cells in surface view polygonal, irregularly arranged, 10–35 µm in diameter. In transverse section, cells rectangular with length:width ratios varying from <1 to 2, and maximum diameters up to 25 µm. One pyrenoid per cell. Holotype DNA sequences: ITS GenBank PQ099646; tufA GenBank PQ099755; rbcL GenBank PQ099533.
Etymology: The name honors Rachel Carson, renowned marine biologist, environmentalist, and author, and also reflects the holotype collection location, which was inside the Rachel Carson National Estuarine Research Reserve.
4 Discussion
Nine different species of Ulva were found in North Carolina based on cluster analyses of nuclear encoded ITS and plastid encoded rbcL and tufA sequences as well as phylogenetic analyses of the latter two genes. Morpho-anatomical characters were determined to not be useful for distinguishing or identifying these species. Definitive names could be applied to six of these species by comparisons to type specimen sequences – U. gigantea, U. lacinulata, U. lactuca, U. ohnoi, U. prolifera, and U. tepida, and U. ‘torta’ is tentatively assigned to one of the North Carolina species based on comparisons with topotype specimen sequences.
To place specimens into species based upon the level of DNA sequence divergence, the difference between the minimum interspecific and maximum intraspecific sequence divergences or ‘barcode gap’ of closely related species must be established (Freshwater et al. 2010; Meier et al. 2008; Saunders and Kucera 2010). The most critical way to assess this is through comparisons of sequences from specimens used in crossing experiments, and Ulva is one of the few macroalgal genera for which such data exist (Cui et al. 2018; Hiraoka et al. 2003, 2011, 2017; Steinhagen et al. 2019a). Unfortunately, partial compatibility among tested cultures and contrasting sequence divergence versus compatibility results have currently made the application of a barcode gap value, or even a narrow barcode gap range, questionable. However, the indistinct barcode gap in Ulva should not be unexpected considering the incompatibility observed between some specimens within the same clade and partial compatibility and hybridization between some specimens from different clades (Cui et al. 2018; Hiraoka et al. 2011, 2017). In lieu of a definitive barcode gap, North Carolina Ulva species were distinguished in this study through cluster analyses of generated ITS, rbcL, and tufA DNA sequences from collected specimens (Supplementary Figures S1–S3) and their resolution as part of monophyletic clades within rbcL and tufA phylogenies (Figures 3 and 4). Some past DNA sequencing studies have only used the ITS1 region (e.g., Guidone 2013) and others have only used the ITS2 region (e.g., Cui et al. 2018). In the present study, the entire ITS, ITS1-5.8S rRNA-ITS2, was sequenced when possible to capture all species-rank variation across the region. However, insertion/deletion mutations within the ITS1 and ITS2 sections make it difficult to use these sequences for phylogenetic analyses because of uncertain homology among indel associated sites (e.g. Saunders and Kucera 2010). Therefore, only rbcL and tufA sequences were used for phylogenetic analyses.
4.1 Ulva sp. NC 1 - Ulva gigantea
The most common sequences of ITS, rbcL and tufA found in the collected Ulva sp. NC 1 specimens matched those from the holotype specimen of U. gigantea published in Maggs et al. (2024). Many other sequences in GenBank are identical or closely match those of the U. gigantea holotype (e.g., 30 rbcL and 50+ ITS with >99 % similarity in BLAST searches, January 2025) and most of the specimens from which they were generated have been identified as U. ‘compressa’. A drawing in Dillenius (1742) was designated as the lectotype of U. compressa by Blomster et al. (1998). A specimen in the Dillenius herbarium that closely matched the lectotype drawing was also selected as a typotype, a designation not included in the International Code of Nomenclature for algae, fungi, and plants (Turland et al. 2018). This specimen was functionally used like an epitype to identify, based on morpho-anatomical similarity, ITS-sequenced contemporary specimens as conspecific with U. ‘compressa’ (Blomster et al. 1998). The ITS sequences of some of these U. ‘compressa’ specimens are nearly identical to that of the U. gigantea holotype (1 bp substitution and 2 ambiguous sites between GenBank AF035345 and OR428167). The synonymy of U. gigantea under U. compressa has been proposed for these reasons (Maggs et al. 2024). However, as has been demonstrated in recent taxonomic studies of Ulva (Hughey and Gabrielson 2022, Hughey et al. 2019, 2021a, 2021b) the application of species names without verification with sequences of type specimens is untenable. As discussed above, the type specimen of Ulva compressa has not been sequenced. However, a sequence-based concept of U.‘compressa’ predicated upon the morpho-anatomical similarity of sequenced contemporary specimens with an historical specimen of no designated status per the International Code of Nomenclature for algae, fungi, and plants (Turland et al. 2018) is often currently used. While this may change once sequence data becomes available from a designated epitype of Ulva compressa, Ulva gigantea is currently the oldest type-sequence-verified name for specimens within the species clade including Ulva sp. NC 1.
Phylogenetic analyses and BLAST searches revealed that Ulva gigantea is distributed around the North Atlantic, the Western Pacific, and South Africa. It is distributed from Texas to New Hampshire, along the Gulf of Mexico and Atlantic coasts of the USA (Guidone et al. 2013; Hofmann et al. 2010; Melton and Lopez-Bautista 2021). Specimens were collected all along the North Carolina coast in both this study and that of Melton and Lopez-Bautista (2021) in which they were identified as Ulva ‘compressa’. Ulva gigantea is sister to a clade containing U. expansa Setchell et N.L.Gardner, U. fenestrata Postels et Ruprecht, U. rigida, and a number of other species of uncertain identification (Figures 3 and 4).
Intraspecific plasticity has been previously noted for this species in other studies where it was identified as Ulva ‘compressa’ or Ulva ‘mutabilis’ Föyn (Guidone et al. 2013; Hofmann et al. 2010; Krupnik et al. 2018; Steinhagen et al. 2019a, 2019c). When the morphological characters from specimens of U. gigantea are compared to the identification guide in Kapraun (1984), the following species names could be applied: U. curvata (Kützing) De Toni, U. fasciata Delile, U. ‘flexuosa subsp. flexuosa’ Wulfen, U. ‘flexuosa subsp. paradoxa’ (C.Agardh) M.J.Wynne, U. linza, U. rigida, and U. rotundata. Surprisingly, U. ‘compressa’ sensu Kapraun (1984) is a name that does not apply based on the morphological characters, despite the phylogenetic analyses indicating its close relationship to other specimens that were morpho-anatomically identified as U. ‘compressa’.
All tubular and ‘linza-morphotype’ specimens of U. gigantea were collected from intertidal oceanic sites, including one tubular specimen that was collected from the northernmost sampling area in the state, Nags Head. Hofmann et al. (2010) hypothesized that there may be a correlation between low-salinity environments and bladed morphology, however, in the current study bladed specimens were found in lower salinity inner estuaries and sounds and also in oceanic environments with higher salinities, which does not support this hypothesis. Other studies have also found no relationship between the switch from bladed to tubular morphologies and salinity (e.g. Tan et al. 1999; Wichard et al. 2015).
4.2 Ulva sp. NC 2 - Ulva prolifera
This species was confirmed as Ulva prolifera by one of the North Carolina ITS2 haplotypes being an exact match to the epitype designated by Cui et al. (2018). While the ITS2 sequence of the epitype specimen was not made publicly available, Cui et al. (2018) reported that it was identical to the ITS2 region of GenBank accession AJ012276, a sequence of the entire ITS region. The other three ITS2 haplotypes present in North Carolina specimens of U. prolifera were only 2–4 nucleotides different from that of the epitype. BLAST results for North Carolina specimens in this species include various specimens in the Ulva linza-procera-prolifera (LPP) complex clade (e.g., Cui et al. 2018; Kang et al. 2019). This species has a wide global distribution, including specimens from the Yellow Sea, Baltic Sea, Mediterranean, and East Pacific. Regionally, this species has been identified by DNA sequencing in the Northeast Pacific (Kuba et al. 2022) and Northwest Atlantic (Melton and Lopez-Bautista 2021).
Ulva ‘linza’ is the oldest of the three names applied to the specimens in the LPP clade, but no sequence data is available from its type specimen, and specimens morphologically identified as U. ‘linza’ are parts of multiple clades resolved by phylogenetic analyses including those in this study (Figures 3 and 4). It is possible that when sequence data is generated from the type of U. linza it will show that U. prolifera is a heterotypic synonym of U. linza, which will then be the correct name to apply to this clade.
The bladed specimens of this species are of interest because U. prolifera traditionally has been described as having a tubular branched morphology (Burrows 1991; Kapraun 1969, 1984; Schneider and Searles 1991), but our results mirror that of Kuba et al. (2022), who found that specimens of U. prolifera from the San Juan Islands, Washington, USA had a ‘linza-morphotype’ of blades that tapered and became tubes near and through the stipe. When attempts to identify the North Carolina specimens of this species strictly based on morphology are made using Kapraun (1984), the following names can be applied: U. ‘linza’, U. ‘flexuosa subsp. flexuosa’, U. ‘flexuosa subsp. paradoxa’, U. prolifera, U. ‘compressa’, Ulva fasciata, and Ulva rotundata.
4.3 Ulva sp. NC 3
This species remains unnamed as its sequences did not match any available from type specimens. Ulva sp. NC 3 was well dispersed across the North Carolina coast, with samples at one of the southernmost North Carolina collection sites and one of the northernmost sites. It also spanned a wide range of salinities and habitats. Globally, this species is distributed in the Indian Ocean, Northeast Atlantic, and East Pacific, as well as the Mediterranean Sea and West Pacific. Phylogenetic analyses of both rbcL and tufA place this species as a sister taxon to U. californica and in a clade with U. tanneri and U. kanagawae (Figures 3 and 4).
Three species names can be applied to the specimens of Ulva sp. NC 3 based on the morphological descriptions of Kapraun (1984): U. ‘linza’, U. ‘flexuosa subsp. flexuosa’, and Ulva ‘flexuosa subsp. paradoxa’. Ulva ‘flexuosa’ is a name that has been applied to other specimens resolved in the tufA species clade that includes Ulva sp. NC 3 (Figure 3), but there is currently no sequence from the type of U. ‘flexuosa’ to confirm these identifications. Ulva ‘aragoënsis’ (Bliding) Maggs is another name that has been applied to specimens within this species clade, including some from North Carolina (Melton and Lopez-Bautista 2021), but similarly there are no type sequences to verify this identification.
4.4 Ulva sp. NC 4 - Ulva ‘torta’
One of the tufA haplotypes for this species was 1 bp (0.067 %) different from the sequence of an Ulva ‘torta’ topotype specimen. It is not a sequence from the type specimen upon which the name is based (Turland et al. 2018). The specimens considered as topotypes for Ulva ‘torta’ match its general morphology but are from the Belt Sea area of the Baltic Sea and not the North Sea coast of Germany from where the species was originally described (Steinhagen et al. 2019b). Thus, we tentatively identified this species as Ulva ‘torta’ until a type specimen sequence becomes available to confirm its identity.
One tufA haplotype from North Carolina specimens was identical to those of specimens from Rhode Island, Texas, and New Jersey, and previously collected North Carolina specimens (Guidone et al. 2013; Melton and Lopez-Bautista 2021). Specimens of this species were also >99.5 % similar to samples from Australia, China, and Tunisia, and >98.5 % similar to specimens from the Northeast Pacific (British Columbia, Canada and Washington. The only other name applied to rbcL and ITS sequences of this species in public databases is Ulva clathratioides L.G.Kraft, Kraft et R.F.Waller, a name that has now been synonymized under Ulva ’torta’. Due to size discrepancies between North Carolina U. ‘torta’ specimens and the species description in Kapraun (1984), this species was identified morphologically as either U. ‘ramulosa’ Smith or U. ‘clathrata’ (Roth) C.Agardh. Ulva ‘ramulosa’ has since been synonymized under U. ‘clathrata’.
4.5 Ulva sp. NC 5 - Ulva tepida
Using only ITS sequences, this sample was confirmed as U. tepida. The ITS sequence of the U. tepida holotype was an exact match to one of the ITS sequences found in North Carolina specimens and was only 1 bp (0.004 %) different from the other North Carolina ITS sequence for this species. Phylogenetically, this species groups with samples from the Mediterranean, East Pacific and Indian Ocean, as well as from the Northwest Atlantic (Melton and Lopez-Bautista 2021).
Krupnik et al. (2018) found U. tepida samples that were all tubular, narrow, and hair-like. The original description of the species states that thalli are up to 11.5 cm in length and up to 8 mm in diameter, with cells containing 1–5 pyrenoids (Masakiyo and Shimada 2014). The North Carolina samples fall within this size description, and their pyrenoid counts fell into the categories of 1–3 or greater than 3 per cell, which matches the original description. However, this description is very broad and overlaps that of many other Ulva species.
Ulva tepida was described as a species subsequent to Kapraun (1984), but when specimens of this species are identified using that dichotomous key, they identify as U. ‘flexuosa subsp. paradoxa’ or U. ‘torta’. Our sequence data indicates that the name U. ‘torta’ is more applicable to Ulva sp. NC 4, but the description of U. ‘torta’ in North Carolina is also applicable to U. tepida.
4.6 Ulva sp. NC 6 - Ulva lactuca
All North Carolina specimens of this species shared an rbcL sequence that was an exact match to that of the U. lactuca specimen from Florida identified by Hughey et al. (2019) as representative for the entire sequence of the lectotype specimen. North Carolina U. lactuca specimens also shared the same tufA sequence as that of the Florida specimen and one of the two ITS sequences was identical to the Florida specimen as well. The other North Carolina U. lactuca ITS sequence was only 3 bp (0.42 %) different. Similar to other studies, U. lactuca is resolved as part of a clade including U. conglobata, U. dactylifera, U. taeniata, U. ohnoi, and U. lacinulata, the latter two species also found in North Carolina. Relationships among the species in this clade were poorly supported in single locus trees generated in this and other studies (Gabrielson et al. 2024; Hughey et al. 2021b, 2024). However, analyses of Ulva spp. plastid and mitochondrial genomes resolved U. lactuca and U. ohnoi as sister species with full support, and also fully supported a basal position for U. lacinulata within this clade (Hughey et al. 2024).
Globally, U. lactuca has been confirmed by sequencing to be distributed in the North Atlantic, Mediterranean Sea, Indian and Pacific Oceans. The Linnaeus’ (1753) original description of U. lactuca mirrors the gross superficial morphology of what was later described as U. fasciata (Gray 1821), an alga with strap-like blades all stemming from a centralized stipe at the base that has been synonymized under U. lactuca (Hughey et al. 2019). All the collected North Carolina U. lactuca specimens could be interpreted as having the fasciculately branched morphology of U. lactuca, but the states of other characters did not always match the descriptions of U. lactuca (as U. fasciata) in North Carolina provided by Kapraun (1984) and Schneider and Searles (1991). For example, one of the defining characters of U. fasciata sensu Kapraun (now U. lactuca) is the presence of very distinct pyrenoids, ranging from 1-3 per cell, a character state that is present in the majority, but not all North Carolina samples of this species.
4.7 Ulva sp. NC 7 - Ulva ohnoi
Three North Carolina specimens were identified as U. ohnoi by their rbcL and tufA sequences being exact matches to those of the holotype specimen. Attempts to sequence ITS from these specimens failed. As noted above, U. ohnoi and U. lactuca have been resolved as sister species in single locus, and plastid and mitochondrial genome analyses, but this relationship is only supported in the genome-level analyses (Hughey et al. 2024).
Ulva ohnoi has been confirmed by DNA sequences to be distributed in the Mediterranean Sea, and Pacific, Indian and Atlantic Oceans. It had previously been reported from the Gulf of Mexico and Atlantic coasts of Florida, USA (Melton et al. 2016), and its Western Atlantic distribution is herein expanded northward to the North Carolina coast. Morpho-anatomically the North Carolina specimens varied at characters considered significant for identification and keyed to three different species (U. curvata, U. fasciata, and U. rigida) using Kapraun (1984).
4.8 Ulva sp. NC 8 - Ulva lacinulata
Ulva lacinulata in North Carolina was positively identified by the tufA and rbcL sequences of the collected specimens being exact matches to those of the U. lacinulata lectotype. The plastid and mitochondrial genome analyses of Hughey et al. (2024) supported a basal position for U. lacinulata within the Ulva spp. clade that also includes U. ohnoi, and U. lactuca.
DNA sequences have confirmed the presence of U. lacinulata in the North Pacific and Eastern Atlantic Oceans, and Mediterranean Sea. The two North Carolina samples in this study, plus three samples from Virginia and North Carolina identified as U. rigida by Melton and Lopez-Bautista (2021) are the first verified records of U. lacinulata in the Western Atlantic. Based on morpho-anatomical characters, the two samples in this study would be identified as U.’curvata’ and U. ‘rotundata’ sensu Kapraun (1984) and Schneider and Searles (1991).
4.9 Ulva sp. NC 9 – Ulva carsoniae sp. nov
This species was represented by a single specimen in this study that was collected at Carrot Island, near Beaufort, NC. Other specimens of this species have been collected from Virginia, Maryland, Delaware and Rhode Island along the United States east coast, and a single specimen is known from Japan’s west coast (Guidone et al. 2013; Melton and Lopez-Bautista 2021; Ogawa et al. 2013). It is resolved in the rbcL and tufA trees within a large clade that includes the types of U. australis, U. expansa, U. fenestrata, U. rigida, and other undetermined species. The single North Carolina collection was a cluster of unbranched blades with fused bases that were growing attached to a small shell. Specimens from Virginia, Maryland and Delaware were reported as having blades with tubular bases that became distromatic with tubular margins, and polygonal cells in surface view with a single pyrenoid (Melton and Lopez-Bautista 2021). The North Carolina specimens were similar in having polygonal shaped cells in surface view, and pyrenoid numbers that were mostly one, but cells in the North Carolina specimens also sometimes had two or three pyrenoids and the blades lacked hollow bases and margins. The central region interior to the surface cells, extending from the terete base of the stipe immediately above the holdfast through the compressed and then flatten base of the blade was filled with a solid mass of rhizoidal extensions of the cell walls. The large size of the Virginia specimen illustrated by Melton and Lopez-Bautista (2021) suggests that the North Carolina specimens were likely young.
5 Conclusions
Nine DNA sequence-based Ulva species were distinguished in this study of 120 specimens from along the North Carolina coast. This is a decrease from the 11 morpho-anatomically based species reported in the most recent treatments of Ulva in North Carolina (Kapraun 1984; Schneider and Searles 1991) and is a product of the inherent morphological plasticity that has been revealed by sequence-based analyses of Ulva species. The analyses of morpho-anatomical characters in these specimens have shown that the accurate identification of North Carolina Ulva species with these characters is not possible, and this is likely the case for all Ulva species worldwide. These findings support the sentiments of Gabrielson et al. (2024) that species names for Ulva specimens that have not been DNA sequenced, and those used in public databases for which there is no confirming type sequence available, may all be incorrect. Consequently, only seven of the nine sequence-based species distinguished in this study could be accurately identified, and an eighth was described herein as a new species. As more Ulva species type specimen sequences become available, the classification of the still unidentified North Carolina species will hopefully be resolved. Understanding diversity at the local level is needed to inform future studies where Ulva species play an economically influential role, both as a nuisance and potential resource.
As a final comment, this study has demonstrated that morpho-anatomical character state inconsistency within Ulva species negates their use for taxonomy and classification, and once again shows the necessity of having DNA sequence data from type specimens to correctly apply species names. The collaboration of researchers and cooperating herbaria to generate DNA sequences from type specimens has proven that this can be done successfully and responsibly. It is here proposed that institutions supporting herbaria provide training for, or hire curatorial staff already knowledgeable in, the removal of the small snippets of type material that could be provided to vetted researchers for the generation of DNA sequences from these materials. This will increase the utility and relevance of herbaria, further promoting their maintenance by supporting institutions.
Funding source: North Carolina Sea Grant, North Carolina State University
Award Identifier / Grant number: project 2020-R/MG-2001
Funding source: CMS DNA Algal Research Fund
Funding source: Red Pond Trust
About the authors

Skye Dibner is a research analyst at the University of North Carolina at Wilmington. She earned her undergraduate degree at Eckerd College, where she completed a thesis on the molecular systematics of holopelagic Sargassum. She conducted her master’s research at UNCW, focusing on Ulva species diversity in North Carolina. Her current work at UNCW’s Shellfish Research Hatchery centers on the molecular genetics of Crassostrea virginica (Eastern Oyster).

Paul W. Gabrielson is an algal systematist. He obtained his PhD at the University of North Carolina, Chapel Hill with Max Hommersand and was a post-doc with Gerald T. Kraft and the late Robert F. Scagel. He retooled from morpho-anatomy to DNA sequencing while teaching at a small, liberal arts college for nine years, before returning to UNC, Chapel Hill. He collaborates with phycologists worldwide sequencing contemporary field-collected specimens, as well as type specimens of seaweeds from the 18th, 19th and early 20th centuries.

D. Wilson Freshwater is researcher at the Center for Marine Science, University of North Carolina, Wilmington. A recent graduate of genomics kindergarten, he has research interests in the molecular phylogeny and taxonomy of marine algae, marine floristics, and the ecology of marine hard bottom communities.
Acknowledgments
The authors thank A. Grogan, M. Lytle, M.L. May, and K.D. Turner, who assisted with the collection of samples; M.B. Ahrens and M. Lytle for assistance with some figures, and J. Jarvis for helpful comments on early versions of the manuscript.
-
Research ethics: All procedures were in accordance with national laws.
-
Informed consent: Not applicable.
-
Author contributions: All authors have accepted responsibility for the entire content of this manuscript and approved its submission. Skye Dibner: field collections, morpho-anatomical analyses, DNA sequencing, phylogenetic analyses, writing; Paul W. Gabrielson: concept, field collections, writing; D. Wilson Freshwater: concept, field collections, morpho-anatomical analyses, DNA sequencing, writing.
-
Use of Large Language Models, AI and Machine Learning Tools: None declared.
-
Conflict of interest: The authors declare no conflicts of interest regarding this article.
-
Research funding: Support for this project was provided by the North Carolina SeaGrant Minigrant program (project 2020-R/MG-2001), the Red Pond Trust and the CMS DNA Algal Research Fund.
-
Data availability: All data used in this study are available within the article, and/or publicly available through the BOLDsystem or GenBank databases.
References
Aziz, K.M.S. and Humm, H.J. (1962). Additions to the algal flora of Beaufort, N. C., and vicinity. J. Elisha Mitchell Sci. Soc. 78: 55–63.Suche in Google Scholar
Blomquist, H.L. and Humm, H.J. (1946). Some marine algae new to Beaufort, North Carolina. J. Elisha Mitchell Sci. Soc. 62: 1–8.Suche in Google Scholar
Blomster, J., Maggs, C.A., and Stanhope, M.J. (1998). Molecular and morphological analysis of Enteromorpha intestinalis and E. compressa (Chlorophyta) in the British Isles. J. Phycol. 34: 319–340, https://doi.org/10.1046/j.1529-8817.1998.340319.x.Suche in Google Scholar
Brodie, J.A., Maggs, C.A., and John, D.M. (2007). Green seaweeds of Britain and Ireland. British Phycological Society, London.Suche in Google Scholar
Burrows, E.M. (1991). Seaweeds of the British Isles, Volume 2 Chlorophyta. Natural History Museum Publications, London, pp. 238.Suche in Google Scholar
Cai, C., Gu, K., Zhao, H., Steinhagen, S., He, P., and Wichard, T. (2021). Screening and verification of extranuclear genetic markers in green tide algae from the Yellow Sea. PLOS One 16: e0250968, https://doi.org/10.1371/journal.pone.0250968.Suche in Google Scholar PubMed PubMed Central
Carneiro, V.A.R., Martins, N.T., Albuquerque Da Silva, S.L., De Barros-Barreto, M.B., Pereira, S.B., and Cassano, V. (2023). Revealing the diversity of the genus Ulva (Ulvales, Chlorophyta) in southeastern Brazil, with a description of Ulva kanagawae sp. nov. Phycologia 62: 1–14, https://doi.org/10.1080/00318884.2023.2243433.Suche in Google Scholar
Charrier, B., Abreu, M.H., Araujo, R., Bruhn, A., Coates, J.C., De Clerck, O., Katsaros, C., Robaina, R.R., and Wichard, T. (2017). Furthering knowledge of seaweed growth and development to facilitate sustainable aquaculture. New Phytol. 216: 967–975, https://doi.org/10.1111/nph.14728.Suche in Google Scholar PubMed
Chase, M.W. and Hills, H.H. (1991). Silica Gel: an ideal material for field preservation of leaf samples for DNA studies. Taxon 40: 215–220, https://doi.org/10.2307/1222975.Suche in Google Scholar
Chávez-Sánchez, T., Piñón-Gimate, A., Melton, J.T., López-Bautista, J.M., and Casas -Valdez, M. (2019). First report, along with nomenclature adjustments, of Ulva ohnoi, U. tepida and U. torta (Ulvaceae, Ulvales, Chlorophyta) from northwestern Mexico. Bot. Mar. 62: 113–123, https://doi.org/10.1515/bot-2018-0007.Suche in Google Scholar
Chávez-Sánchez, T., Piñón-Gimate, A., Serviere-Zaragoza, E., López-Bautista, J.M., and Casa-Valdez, M. (2018). Ulva blooms in the southwestern Gulf of California: reproduction and biomass. Estuar. Coast. Shelf Sci. 200: 202–211, https://doi.org/10.1016/j.ecss.2017.11.007.Suche in Google Scholar
Chávez-Sánchez, T., Piñón-Gimate, A., Serviere-Zaragoza, E., Sánchez-González, A., Hernández-Carmona, G., and Casas-Valdez, M. (2017). Recruitment in Ulva blooms in relation to temperature, salinity and nutrients in a subtropical bay of the Gulf of California. Bot. Mar. 60: 257–270, https://doi.org/10.1515/bot-2016-0066.Suche in Google Scholar
Cui, J., Monotilla, A.P., Zhu, W., Takano, Y., Shimada, S., Ichihara, K., Matsui, T., He, P., and Hiraoka, M. (2018). Taxonomic reassessment of Ulva prolifera (Ulvophyceae, Chlorophyta) based on specimens from the type locality and Yellow Sea green tides. Phycologia 57: 692–704, https://doi.org/10.2216/17-139.1.Suche in Google Scholar
Curtis, M.A. (1867). Geological and natural history survey of North Carolina. Part III. Botany: containing a catalogue of the indigenous and naturalized plants of the state. NC Institution for the Deaf and Dumb and the Blind, Raleigh, NC, pp. 155.10.5962/bhl.title.62310Suche in Google Scholar
Dmytryk, A. and Chojnacka, K. (2018) Algae as fertilizers, biostimulants, and regulators of plant growth. In: Chojnacka, K., Wieczored, P., Schroeder, G., and Michalak, I. (Eds.). Algae biomass: characteristics and applications. Developments in Applied Phycology, Vol. 8. Springer International Publishing, pp. 115–122.10.1007/978-3-319-74703-3_10Suche in Google Scholar
Dominguez, H. and Loret, E.P. (2019). Ulva lactuca, a source of troubles and potential riches. Mar. Drugs 17: 357, https://doi.org/10.3390/md17060357.Suche in Google Scholar PubMed PubMed Central
Edgar, R.C. (2004). MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 32: 1792–1797, https://doi.org/10.1093/nar/gkh340.Suche in Google Scholar PubMed PubMed Central
Fama, P., Wysor, B., Kooistra, W.H.C.F., and Zuccarello, G.C. (2002). Molecular phylogeny of the genus Caulerpa (Caulerpales, Chlorophyta) inferred from cholorplast tufA Gene1. J. Phycol. 38: 1040–1050, https://doi.org/10.1046/j.1529-8817.2002.t01-1-01237.x.Suche in Google Scholar
Fort, A., Mchale, M., Cascella, K., Potin, P., Perrineau, M., Kerrison, P.D., Costa, E., Calado, R., Domingues, M.D.R., Costa Azevedo, I., et al. (2022). Exhaustive reanalysis of barcode sequences from public repositories highlights ongoing misidentifications and impacts taxa diversity and distribution. Mol. Ecol. Resour 22: 86–101, https://doi.org/10.1111/1755-0998.13453.Suche in Google Scholar PubMed
Freshwater, D.W., Tudor, K., O’Shaughnessy, K., and Wysor, B. (2010). DNA barcoding in the red algal order Gelidiales: comparison of COI with rbcL and verification of the “barcoding gap”. Cryptogam. Algol. 31: 435–449.Suche in Google Scholar
Gabrielson, P.W., Smith, A.C., Bruno, J.F., Vision, T.J., and Brandt, M. (2024). Taxonomic assessment of blade-forming Ulva species (Ulvales, Chlorophyta) in the Galápagos Archipelago, Ecuador using DNA sequencing. Bot. Mar. 67: 153–164, https://doi.org/10.1515/bot-2023-0079.Suche in Google Scholar
Gao, G., Zhong, Z., Zhao, X., and Xu, J. (2016). Changes in morphological plasticity of Ulva prolifera under different environmental conditions: a laboratory experiment. Harmful Algae 59: 51–58, https://doi.org/10.1016/j.hal.2016.09.004.Suche in Google Scholar PubMed
Ghaderiardakani, F., Coates, J.C., and Wichard, T. (2017). Bacteria-induced morphogenesis of Ulva intestinalis and Ulva mutabilis (Chlorophyta): a contribution to the lottery theory. FEMS Microbiol. Ecol. 93: fix094, https://doi.org/10.1093/femsec/fix094.Suche in Google Scholar PubMed PubMed Central
Gray, S.F. (1821). A natural arrangement of British plants, according to their relations to each other, as pointed out by Jussieu, De Candolle, Brown, & c. including those cultivated for use; with an introduction to botany, in which the terms newly introduced are explained; illustrated by figures. pp. vol. 1: [i]-xxviii, [1]- 824, pls I-XXI plates. Printed for Baldwin, Cradock & Joy, Paternoster-Row, London.10.5962/bhl.title.43804Suche in Google Scholar
Guidone, M., Carol, T., Wysor, B., O’Kelly, C.J., and Dunton, K. (2013). Molecular and morphological diversity of Narragansett bay (RI, USA) Ulva (Ulvales, Chlorophyta) populations. J. Phycol. 49: 979–995, https://doi.org/10.1111/jpy.12108.Suche in Google Scholar PubMed
Guiry, M.D. and Guiry, G.M. (2025). AlgaeBase. World-wide electronic publication. National University of Ireland, Galway, searched on http://www.algaebase.org (Accessed 28 April 2025).Suche in Google Scholar
Hanyuda, T. and Kawai, H. (2018). Genetic examination of the type specimen of Ulva australis suggests that it was introduced to Australia. Phycol. Res. 66: 238–241, https://doi.org/10.1111/pre.12222.Suche in Google Scholar
Hayden, H.S., Blomster, J., Maggs, C.A., Silva, P.C., Stanhope, M.J., and Waaland, J.R. (2003). Linnaeus was right all along: Ulva and Enteromorpha are not distinct genera. Eur. J. Phycol. 38: 277–294, https://doi.org/10.1080/1364253031000136321.Suche in Google Scholar
Hiraoka, M., Ichihara, K., Zhu, W., Ma, J., and Shimada, S. (2011). Culture and hybridization experiments on an Ulva clade including the Qingdao strain blooming in the Yellow Sea. PLoS One 6: e19371, https://doi.org/10.1371/journal.pone.0019371.Suche in Google Scholar PubMed PubMed Central
Hiraoka, M., Ichihara, K., Zhu, W., Shimada, S., Oka, N., Cui, J., Tsubaki, S., and He, P. (2017). Examination of species delimitation of ambiguous DNA-based Ulva (Ulvophyceae, Chlorophyta) clades by culturing and hybridisation. Phycologia 56: 517–532, https://doi.org/10.2216/16-109.Suche in Google Scholar
Hiraoka, M., Shimada, S., Uenosono, M., and Masuda, M. (2003). A new green-tide-forming alga, Ulva ohnoi Hiraoka et Shimada sp. nov. (Ulvales, Ulvophyceae) from Japan. Phycol. Res 51: 17–29, https://doi.org/10.1111/j.1440-1835.2003.00321.x.Suche in Google Scholar
Hofmann, L.C., Nettleton, J.C., Neefus, C.D., and Mathieson, A.C. (2010). Cryptic diversity of Ulva (Ulvales, Chlorophyta) in the Great Bay estuarine system (Atlantic USA): introduced and indigenous distromatic species. Eur. J. Phycol. 45: 230–239, https://doi.org/10.1080/09670261003746201.Suche in Google Scholar
Hoyt, W.D. (1920). Marine algae of Beaufort, N. C. and adjacent regions. Bull. Bur. of Fish. 36: 367–556, + i–v, 35 pls.10.5962/bhl.title.29440Suche in Google Scholar
Huelsenbeck, J.P. and Ronquist, F. (2001). MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 17: 754–755, https://doi.org/10.1093/bioinformatics/17.8.754.Suche in Google Scholar PubMed
Hughey, J.R., Gabrielson, P.W., Maggs, C.A., Mineur, F., and Miller, K.A. (2021b). Taxonomic revisions based on genetic analysis of type specimens of Ulva conglobata, U. laetevirens, U. pertusa and U. spathulata (Ulvales, Chlorophyta). Phycol. Res. 69: 148–153, https://doi.org/10.1111/pre.12450.Suche in Google Scholar
Hughey, J.R. and Gabrielson, P.W. (2022). DNA analysis of the lectotype specimens of Ulva nematoidea Bory (Ulvaceae, Chlorophyta) determines its synonymy with Ulva lactuca L. Cryptogam. Algol. 43: 117–124, https://doi.org/10.5252/cryptogamie-algologie2022v43a7.Suche in Google Scholar
Hughey, J.R., Gabrielson, P.W., Maggs, C.A., and Mineur, F. (2021a). Genomic analysis of the lectotype specimens of European Ulva rigida and Ulva lacinulata (Ulvaceae, Chlorophyta) reveals the ongoing misapplication of names. Eur. J. Phycol. 57: 143–153, https://doi.org/10.1080/09670262.2021.1914862.Suche in Google Scholar
Hughey, J.R., Maggs, C.A., Mineur, F., Jarvis, C., Miller, K.A., Shabaka, S.H., and Gabrielson, P.W. (2019). Genetic analysis of the Linnaean Ulva lactuca (Ulvales, Chlorophyta) holotype and related type specimens reveals name misapplications, unexpected origins, and new synonymies. J. Phycol. 55: 503–508, https://doi.org/10.1111/jpy.12860.Suche in Google Scholar PubMed
Hughey, J.R., Miller, K.A., and Gabrielson, P.W. (2018). Mitogenome analysis of a green tide forming Ulva from California, USA confirms its identity as Ulva expansa (Ulvaceae, Chlorophyta). Mitochondrial DNA Part B 3: 1302–1303, https://doi.org/10.1080/23802359.2018.1535859.Suche in Google Scholar PubMed PubMed Central
Hughey, J.R., Miller, K.A., and Gabrielson, P.W. (2024). Genetic analysis of Ulva (Ulvaceae, Chlorophyta) type specimens resolves Northeast Pacific blade-forming species. Bot. Mar. 67: 165–179, https://doi.org/10.1515/bot-2023-0072.Suche in Google Scholar
Kang, J.H., Jang, J.E., Kim, J.H., Byeon, S.Y., Kim, S., Choi, S.K., Kang, Y.H., Park, S.R., and Lee, H.J. (2019). Species composition, diversity, and distribution of the genus Ulva along the coast of Jeju Island, Korea based on molecular phylogenetic analysis. PLoS One 14: e0219958, https://doi.org/10.1371/journal.pone.0219958.Suche in Google Scholar PubMed PubMed Central
Kapraun, D.F. (1969). Field and cultural studies of the genera Ulva and Enteromorpha. University of Texas, Austin, pp. 191.Suche in Google Scholar
Kapraun, D.F. (1984). An illustrated guide to the benthic marine algae of coastal North Carolina II. Chlorophyta and Phaeophyta. Bibl. Phycol. 58: 1–173.Suche in Google Scholar
Kraft, L.G., Kraft, G.T., and Waller, R.F. (2010). Investigations into southern Australian Ulva (Ulvophyceae, Chlorophyta) taxonomy and molecular phylogeny indicate both cosmopolitanism and endemic cryptic species. J. Phycol. 46: 1257–1277, https://doi.org/10.1111/j.1529-8817.2010.00909.x.Suche in Google Scholar
Krupnik, N., Paz, G., Douek, J., Lewinsohn, E., Israel, A., Carmel, N., Mineur, F., and Maggs, C.A. (2018). Native, invasive and cryptogenic Ulva species from the Israeli Mediterranean Sea: risk and potential. Med. Mar. Sci. 19: 132–146, https://doi.org/10.12681/mms.2104.Suche in Google Scholar
Kuba, G.M., Carpio-Aguilar, B., Eklund, J., and Freshwater, D.W. (2022). A demonstration of DNA barcoding-based identification of blade-form Ulva (Ulvophyceae, Chlorophyta) species from three sites in the San Juan Islands, Washington, USA. Diversity 14: 899, https://doi.org/10.3390/d14110899.Suche in Google Scholar
Lagourgue, L., Gobin, S., Brisset, M., Vandenberghe, S., Bonneville, C., Jauffrais, T., Van Wynsberge, S., and Payri, C.E. (2022). Ten new species of Ulva (Ulvophyceae, Chlorophyta) discovered in New Caledonia: genetic and morphological diversity, and bloom potential. Eur. J. Phycol. 57: 458–478, https://doi.org/10.1080/09670262.2022.2027023.Suche in Google Scholar
Lee, H.W., Kang, J.C., and Kim, M.S. (2019). Taxonomy of Ulva causing blooms from Jeju Island, Korea with new species, U. pseudo-ohnoi sp. nov. (Ulvales, Chlorophyta). Algae 34: 253–266, https://doi.org/10.4490/algae.2019.34.12.9.Suche in Google Scholar
Link, H.F. (1820). Nees von Esenbeck, C.D. (Ed.). Epistola de algis aquatics in genera disponedis in Horae Physicae Berolmensus. Sumtibus Adolphi Marcus, Bonn, pp. 1–8.Suche in Google Scholar
Linnaeus, C. (1753). Species plantarum, exhibentes plantas rite cognitas, ad genera relatas, cum differentiis specificis, nominibus trivialibus, synonymis selectis, locis natalibus, secundum systema sexuale digestas, Vol. 2 pp. [i], 561–1200, [1–30, index], [i, err.]. Impensis Laurentii Salvii, Holmiae [Stockholm].10.5962/bhl.title.59734Suche in Google Scholar
Liu, F., Shao, J.P., Xu, N., Shan, T.F., Sun, S., Hu, X., and Yang, Q.Y. (2010). Ulva diversity in the Yellow Sea during the large-scale green algal blooms in 2008–2009. Phycol. Res. 58: 270–279.10.1111/j.1440-1835.2010.00586.xSuche in Google Scholar
Maggs, C.A., Bunker, A.R., Bunker, F.S.P.D., Harries, D., Kelly, J., Mineur, F., Blomster, J., Díaz-Tapia, P., Gabrielson, P.W., Hughey, J.R., and Brodie, J. (2024). Updating the Ulvaceae in the green seaweeds of Britain and Ireland. Bot. Mar. 67: 181–203, https://doi.org/10.1093/oxfordjournals.molbev.a026190.Suche in Google Scholar PubMed
Marshall, K., Joint, I., Callow, M.E., and Callow, J.A. (2006). Effect of marine bacterial isolates on the growth and morphology of axenic plantlets of the green alga Ulva linza. Microbial Ecol. 52: 302–310, https://doi.org/10.1007/s00248-006-9060-x.Suche in Google Scholar PubMed
Masakiyo, Y. and Shimada, S. (2014). Species diversity of the genus Ulva (Ulvophyceae, Chlorophyta) in Japanese waters, with special reference to Ulva tepida Masakiyo et S. Shimada sp. nov. Bull. Nat. Mus. Nat. Sci. 40: 1–13.Suche in Google Scholar
Meier, R., Zhang, G., and Ali, F. (2008). The use of mean instead of smallest interspecific distances exaggerates the size of the “Barcoding Gap” and leads to misidentification. Systematic Biol. 57: 809–813, https://doi.org/10.1080/10635150802406343.Suche in Google Scholar PubMed
Melton, J.T., Collado-Vides, L., and Lopez-Bautista, J.M. (2016). Molecular identification and nutrient analysis of the green tide species Ulva ohnoi M.Hiraoka & S.Shimada, 2004 (Ulvophyceae, Chlorophyta), a new report and likely nonnative species in the Gulf of Mexico and Atlantic Florida, USA. Aquat. Invasions 11: 225–237.10.3391/ai.2016.11.3.01Suche in Google Scholar
Melton, J.T. and Lopez-Bautista, J.M. (2021). Diversity of the green macroalgal genus Ulva (Ulvophyceae, Chlorophyta) from the east and gulf coast of the United States based on molecular data. J. Phycol. 57: 551–568, https://doi.org/10.1111/jpy.13120.Suche in Google Scholar PubMed
Norris, J.N. (2010). Marine algae of the northern gulf of California: Chlorophyta and Phaeophyceae. Smithson. Contr. Bot. 94: 1–276, https://doi.org/10.5479/si.0081024x.94.276.Suche in Google Scholar
Ogawa, T., Ohki, K., and Kamiya, M. (2013). Differences of spatial distribution and seasonal succession among Ulva species (Ulvophyceae) across salinity gradients. Phycologia 52: 637–651, https://doi.org/10.2216/13-199.Suche in Google Scholar
Peng, J., Jinfeng, W., Yulin, C., Youxun, L., Hanzhi, L., and Song, Q. (2008). Molecular phylogenetic analysis of attached Ulvaceae species and free-floating Enteromorpha from Qingdao coasts in 2007. Chinese J. Oceanol. Limnol. 26: 276–279, https://doi.org/10.1007/s00343-008-0276-0.Suche in Google Scholar
Ratnasingham, S. and Hebert, P.D. (2007). BOLD: the barcode of life data system. Mol. Ecol. Resour. 7: 355–364, https://doi.org/10.1111/j.1471-8286.2007.01678.x.Suche in Google Scholar PubMed PubMed Central
Rhyne, C.F. (1973). Field and experimental studies of the systematics and ecology of Ulva curvata and Ulva rotundata. Sea Grant publication, Raleigh, N.C., pp. 131, UNC-SG-73-09.Suche in Google Scholar
Rinehart, S., Guidone, M., Ziegler, A., Schollmeier, T., and Thornber, C. (2014). Overwintering strategies of bloom-forming Ulva species in Narragansett bay, Rhode Island, USA. Bot. Mar. 57: 337–341, https://doi.org/10.1515/bot-2013-0122.Suche in Google Scholar
Runca, E., Bernstein, A., Postma, L., and Di Silvo, G. (1996). Control of macroalgae blooms in the Lagoon of Venice. Ocean Coast. Manag. 30: 235–257.10.1016/0964-5691(95)00065-8Suche in Google Scholar
Rybak, A.S. (2018). Species of Ulva (Ulvophyceae, Chlorophyta) as indicators of salinity. Ecol. Indic. 87: 253–261, https://doi.org/10.1016/J.ECOLIND.2017.10.061.Suche in Google Scholar
Saunders, G.W. and Kucera, H. (2010). An evaluation of rbcL, tufA, UPA, LSU and ITS as DNA barcode markers for the marine green macroalgae. Cryptogam. Algol. 31: 487–528.Suche in Google Scholar
Schneider, C.W. and Searles, R.B. (1991). Seaweeds of the southeastern United States: Cape Hatteras to Cape Canaveral. Duke University Press, Durham, pp. 553.10.1515/9780822397984Suche in Google Scholar
Searles, R. and Schneider, C. (1978). A checklist and bibliography of North Carolina seaweeds. Bot. Mar. 21: 99–108, https://doi.org/10.1515/botm.1978.21.2.99.Suche in Google Scholar
Shimada, S., Hiraoka, M., Nabata, S., Iima, M., and Masuda, M. (2003). Molecular phylogenetic analyses of the Japanese Ulva and Enteromorpha (Ulvales, Ulvophyceae), with special reference to the free-floating Ulva. Phycol. Res. 51: 99–108, https://doi.org/10.1046/j.1440-1835.2003.00296.x.Suche in Google Scholar
Spalding, H.L., Conklin, K.Y., Smith, C.M., O’Kelly, C.J., and Sherwood, A.R. (2016). New Ulvaceae (Ulvophyceae, Chlorophyta) from mesophotic ecosystems across the Hawaiian Archipelago. J. Phycol. 52: 40–53, https://doi.org/10.1111/jpy.12375.Suche in Google Scholar PubMed
Spoerner, M., Wichard, T., Bachhuber, T., Stratmann, J., and Oertel, W. (2012). Growth and thallus morphogenesis of Ulva mutabilis (Chlorophyta) depends on a combination of two bacterial species excreting regulatory factors. J. Phycol. 48: 1433–1447, https://doi.org/10.1111/j.1529-8817.2012.01231.x.Suche in Google Scholar PubMed
Stamatakis, R., Ludwig, T., and Meier, H. (2005). RAxML-III: a fast program for maximum likelihood-based inference of large phylogenetic trees. Bioinformatics 21: 456–463, https://doi.org/10.1093/bioinformatics/bti191.Suche in Google Scholar PubMed
Steinhagen, S., Barco, A., Wichard, T., and Weinberger, F. (2019a). Conspecificity of the model organism Ulva mutabilis and Ulva compressa (Ulvophyceae, Chlorophyta). J. Phycol. 55: 25–36, https://doi.org/10.1111/jpy.12804.Suche in Google Scholar PubMed
Steinhagen, S., Karez, R., and Weinberger, F. (2019b). Cryptic, alien and lost species: molecular diversity of Ulva sensu lato along the German coasts of the North and Baltic Seas. Eur. J. Phycol. 54: 466–483, https://doi.org/10.1080/09670262.2019.1597925.Suche in Google Scholar
Steinhagen, S., Weinberger, F., and Karez, R. (2019c). Molecular analysis of Ulva compressa (Chlorophyta, Ulvales) reveals its morphological plasticity, distribution and potential invasiveness on German North Sea and Baltic Sea coasts. Eur. J. Phycol. 54: 102–114, https://doi.org/10.1080/09670262.2018.1513167.Suche in Google Scholar
Tan, I.H., Blomster, J., Hansen, G., Leskinen, E., Maggs, C.A., Mann, D.G., Sluiman, H.J., and Stanhope, M.J. (1999). Molecular phylogenetic evidence for a reversible morphogenetic switch controlling the gross morphology of two common genera of green seaweeds, Ulva and Enteromoprha. Mol. Biol. Evol. 16: 1011–1018, https://doi.org/10.1093/oxfordjournals.molbev.a026190.Suche in Google Scholar PubMed
Taylor, W.R. (1960). Marine algae of the eastern tropical and subtropical coasts of the Americas. The University of Michigan Press, Ann Arbor, pp. 870.Suche in Google Scholar
Tran, L.T., Leliaert, F., Vieira, C., Tran, T.V., Nguyen, T.V., Dam, T.D., and De Clerck, O. (2023). Molecular assessment of Ulva (Ulvales, Chlorophyta) diversity in Vietnam including the new species U. vietnamensis. Phycol. Res. 71: 13–24, https://doi.org/10.1111/pre.12507.Suche in Google Scholar
Turland, N.J., Wiersema, J.H., Barrie, F.R., Greuter, W., Hawksworth, D.L., Herendeen, P.S., Knapp, S., Kusber, W.-H., Li, D.-Z., Marhold, K., et al. (Eds.). (2018) International Code of Nomenclature for algae, fungi, and plants (Shenzhen Code) adopted by the Nineteenth International Botanical Congress Shenzhen, China, July 2017. Regnum Vegetabile 159, Koeltz Botanical Books, Glashütten.10.12705/Code.2018Suche in Google Scholar
Wang, J., Li, N., Jiang, P., Boo, S.M., Lee, W.J., Cui, Y., Lin, H., Zhao, J., Liu, Z., and Qin, S. (2010). Ulva and Enteromorpha (Ulvaceae, Chlorophyta) from two sides of the Yellow Sea: analysis of nuclear rDNA ITS and plastid rbcL sequence data. Chinese J. Oceanol. Limnol. 28: 762–768, https://doi.org/10.1007/s00343-010-9069-3.Suche in Google Scholar
Wassie, T., Niu, K., Xie, C., Wang, H., and Xin, W. (2021). Extraction techniques, biological activities and health benefits of marine algae Enteromorpha prolifera polysaccharide. Front. Nutr. 8, https://doi.org/10.3389/fnut.2021.747928.Suche in Google Scholar PubMed PubMed Central
Wichard, T., Charrier, B., Mineur, F., Bothwell, J.H., De Clerck, O., and Coates, J.C. (2015). The green seaweed Ulva: a model system to study morphogenesis. Front. Plant Sci. 6, https://doi.org/10.3389/fpls.2015.00072.Suche in Google Scholar PubMed PubMed Central
Williams, L.G. (1948). Seasonal alternation of marine floras at Cape Lookout, North Carolina. Am. J. Bot. 35: 683–695, https://doi.org/10.1002/j.1537-2197.1948.tb08137.x.Suche in Google Scholar
Supplementary Material
This article contains supplementary material (https://doi.org/10.1515/bot-2025-0011).
© 2025 Walter de Gruyter GmbH, Berlin/Boston
Artikel in diesem Heft
- Frontmatter
- In this issue
- Physiology and Ecology
- Three-dimensional distribution of nutrients and phytoplankton biomass in a semi-enclosed region of the Gulf of California during different ENSO phases
- Proliferation of Undaria pinnatifida along the Atlantic coast of the Iberian Peninsula
- Study on fatty acid methyl ester (FAME) composition of 19 Brazilian thraustochytrid isolates
- Taxonomy/Phylogeny and Biogeography
- Variations in the bacterial and fungal community structure along the hypoxic gradient of the Arabian Sea oxygen-depleted environment based on eDNA metabarcoding analysis
- Studies of North Carolina marine algae XV. DNA sequencing reveals some different Ulva species compared to historical reports and U. carsoniae sp. nov. (Ulvales, Chlorophyta)
- Identification and growth of Prorocentrum cf. cassubicum (Dinoflagellata: Prorocentraceae) from Bahía de La Paz: first record for the Gulf of California
- A new oomycete pathogen Olpidiopsis dasysiphoniae sp. nov. (Oomycota) infecting the red alga Dasysiphonia japonica (Ceramiales, Delesseriaceae)
- Chemistry and Applications
- Exploring the suitable extractive species in an IMTA: inorganic nutrient removal from mariculture effluents by commercially important marine macroalgae
Artikel in diesem Heft
- Frontmatter
- In this issue
- Physiology and Ecology
- Three-dimensional distribution of nutrients and phytoplankton biomass in a semi-enclosed region of the Gulf of California during different ENSO phases
- Proliferation of Undaria pinnatifida along the Atlantic coast of the Iberian Peninsula
- Study on fatty acid methyl ester (FAME) composition of 19 Brazilian thraustochytrid isolates
- Taxonomy/Phylogeny and Biogeography
- Variations in the bacterial and fungal community structure along the hypoxic gradient of the Arabian Sea oxygen-depleted environment based on eDNA metabarcoding analysis
- Studies of North Carolina marine algae XV. DNA sequencing reveals some different Ulva species compared to historical reports and U. carsoniae sp. nov. (Ulvales, Chlorophyta)
- Identification and growth of Prorocentrum cf. cassubicum (Dinoflagellata: Prorocentraceae) from Bahía de La Paz: first record for the Gulf of California
- A new oomycete pathogen Olpidiopsis dasysiphoniae sp. nov. (Oomycota) infecting the red alga Dasysiphonia japonica (Ceramiales, Delesseriaceae)
- Chemistry and Applications
- Exploring the suitable extractive species in an IMTA: inorganic nutrient removal from mariculture effluents by commercially important marine macroalgae