Identification and growth of Prorocentrum cf. cassubicum (Dinoflagellata: Prorocentraceae) from Bahía de La Paz: first record for the Gulf of California
-
Ana Emilia Ramos-Santiago
Ana Emilia Ramos-Santiago is currently a PhD student in Marine Sciences at IPN-CICIMAR, La Paz, Mexico. Her main research interests are epibenthic marine dinoflagellates, sampling techniques, cultivation, and their identification using microscopy and molecular tools. Currently, she is dedicated to understanding the autecology of epibenthic dinoflagellates species from Bahía de La Paz, Mexico.
 ,
Gema V. Villa-Arce
,
Gema V. Villa-Arce
Gema V. Villa-Arce obtained her degree in Fisheries Engineering at the Universidad Autónoma de Baja California Sur in 2013. During her last year she specialized in marine food chemistry. In 2021 she completed her Bachelor’s thesis, focused on growth curves and identification of dinoflagellate strains of the genus
Prorocentrum (Dinophyceae) from the Bahía de La Paz, B.C.S., Mexico at IPN-CICIMAR.
,
Ignacio Leyva-Valencia
Ignacio Leyva-Valencia is a researcher of the Secretariat of Science, Humanities, Technology, and Innovation, affiliated to IPN-CICIMAR. His main research is focused on the isolation and identification of toxigenic dinoflagellate species using microscopy and molecular biology tools. He also conducts experimental work to determine the cytotoxic and antibacterial activity of dinoflagellate extracts, as well as their effect on early stages of bivalve mollusks.
 ,
Yuri Okolodkov
,
Yuri Okolodkov
Yuri Okolodkov started his career in the Academy of Sciences of the URSS in 1973. In 2001 he moved to Mexico by invitation of CONAHCyT. Since 2005 he has been a researcher at the Universidad Veracruzana where he founded the Laboratory of Marine Botany and Planktology, as well as the algae collection of the institute. One of his main interests is the taxonomy of marine dinoflagellates.
and
Christine J. Band-Schmidt
Christine J. Band-Schmidt is a researcher at IPN-CICIMAR where she is leader of the research team studying harmful marine dinoflagellates. She is a founding member of the Mexican Society for the Study of Harmful Algae (SOMEFAN, A.C.) and the CONAHCyT network of harmful algal blooms (RedFAN). Her research has been focused mainly on the autecology of harmful dinoflagellate species from the Gulf of California, with particular interest in understanding the interactions between phytoplankton species and bacteria-phytoplankton interactions.

Abstract
Prorocentrum includes ∼80 species inhabiting benthic and planktonic marine environments. Some species, such as Prorocentrum cassubicum and P. norrisianum, have similar morphology, which complicates their identification based solely on morphological characters. This study describes the morphometrics of two Prorocentrum strains (PNCETMAR-1 and PNCETMAR-2) isolated from the Ensenada de La Paz, Gulf of California. Sequences of the 28S and ITS regions of rDNA of both strains and the growth curve of strain PNCETMAR-1 were obtained. Strain PNCETMAR-1 was grown in modified GSe medium and f/2 medium at a salinity of 34, at 24 °C and 120–150 μmol m−2 s−1 in a 12 h light: 12 h dark cycle. There was no significant difference in growth between the culture media. In f/2 medium the maximum abundance was 74,312 ± 2,008 cells ml−1, while in GSe medium it reached 69,766 ± 2,154 cells ml−1. Morphometric and molecular analyses indicate that both strains belong to the same species. However, because the morphological and molecular information on P. cassubicum and P. norrisianum is contradictory, in this study they are reported as Prorocentrum cf. cassubicum, contributing to the first report of the occurrence and the growth of this species in the Gulf of California.
1 Introduction
Prorocentrum Ehrenberg is one of the most diverse dinoflagellate taxa, with a wide geographic distribution, containing species that inhabit benthic environments; however, they can also inhabit planktonic environments (Chomérat et al. 2019; Hoppenrath et al. 2023). The genus has about 80 species including ∼29 epibenthic species (Guiry and Guiry 2025; Hoppenrath et al. 2014, 2023). Prorocentrum cells measure from 10 to 100 µm in length. Cells are desmokont, characterized by being laterally compressed and by presenting two lateral thecal plates (right and left) separated by a sagittal suture. The periflagellar area is composed of a developed apical spine (although several species do not have this), a second poorly developed spine (several species have none and, if present, it is a wing), and 7 to 8 platelets that present unique morphological characteristics, which help in the identification of species. However, the periflagellar area is difficult to observe, so it has not been characterized in all species (Chomérat et al. 2019; Han et al. 2016; Hoppenrath et al. 2023). In addition, the periflagellar area has two pores: a flagellar pore from which both flagella emerge, and an accessory pore related to mucus production (Hoppenrath et al. 2014). Other diagnostic features used to differentiate Prorocentrum species are the size and arrangement of pores (often referred to in the literature as “trichocyst pores”) on the thecal surface (Hoppenrath et al. 2014; Verma et al. 2019). Although the group presents a “simple” morphology, some characteristics are shared among several taxa, and this has led to the misidentification of morphologically similar species.
The strains in this study have been identified as Prorocentrum cf. cassubicum. Initially, P. cassubicum was described as Exuviaella cassubica Wołoszyńska. Dodge and Bibby (1973) conducted a comparative study on the cell structures of the genera Prorocentrum and Exuviaella Cienkowski from Plymouth (England), Naples (Italy) and Woods Hole (East Coast of the USA) and proposed transferring E. cassubica to the genus Prorocentrum. Prorocentrum cassubicum was found on macroalgae in Ria de Aveiro, Portugal; the species was abundant in low salinity waters during winter and spring (Hinzmann et al. 2008); P. aff. cassubicum has been reported in high abundance in Brazil (Borsato et al. 2023; Melo et al. 2007). A strain from La Jolla, California, was used to study the phylogeny of Prorocentrum species, in which the species was found to be a sister group to P. lima (Ehrenberg) F. Stein and P. hoffmannianum M.A. Faust in the cox 1 gene-based analysis (Cembella et al. 2021; Murray et al. 2009; Tarazona-Janampa et al. 2020). The first record of P. cassubicum in Mexico based on morphological characteristics was reported by Nava-Ruiz and Valadez-Cruz (2012). The species was found in water samples obtained at a depth of 0.3 m in Laguna Lagartos, Quintana Roo in the Mexican Caribbean.
Prorocentrum norrisianum was isolated and described for the first time from floating detritus in mangrove intertidal zones in the Twin Cays in Belize (Faust 1997). After this report, it was found in Malaysia in samples of macroalgae, dead coral and sand (Al-Has and Mohammad-Noor 2011; Mohammad-Noor et al. 2007), while P. cf. norrisianum has been reported in surface sediments (organic dust) from Martinique Island in the eastern Caribbean Sea (Chomérat et al. 2019).
Currently, only P. cassubicum is in the IOC-UNESCO list of harmful microalgal species; however, it has not been confirmed as producing harmful metabolites (Lundholm et al. 2024). The toxicity of strains of P. norrisianum and P. cf. norrisianum is unknown. This study reports the presence of the P. cf. cassubicum, an almost unknown species in the Gulf of California and the Mexican Pacific, and reviews the available information on P. norrisianum and P. cassubicum to reveal their morphometric and molecular differences.
2 Materials and methods
2.1 Sampling site
Both strains were isolated in August 2018 from the beach in Ensenada de La Paz, a shallow coastal lagoon connected with Bahía de La Paz through an inlet of 1.2 km width (Sandoval and Gomes-Valdes 1997) in Baja California Sur, Mexico (Figure 1). This region has a dry or semi-desertic climate, with an average annual temperature of 24 °C (minimum = 5 °C, maximum = 41.5 °C) and an annual rainfall that fluctuates between 180 and 250 mm (Chávez 1985) with the maximum in September and the minimum from March to June (Carmona et al. 2002). It has a mixed tidal regime with a predominance of semi-diurnal tides (Álvarez-Arellano et al. 1997). The Ensenada de La Paz lagoon is anti-estuarine in character, as salinity increases towards the interior of the lagoon due to the shallowness, high rate of evaporation, low precipitation and absence of freshwater runoff (Espinoza 1977). In particular, the sampling site is between 1 and 2 m deep, with water temperatures that fluctuate from 22 to 28.4 °C, dissolved organic matter from 423.1 to 549.40 μg l−1, pH from 7 to 8.3, and salinity from 27.2 to 40.3 (Cruz-Escalona, V.H. pers comm).

Isolation site of the strains PNCETMAR-1 and PNCETMAR-2 used in this study.
2.2 Sampling method and isolation
Samples were collected with phytoplankton nets of 10- and 20-μm mesh size, using shallow horizontal tows during 10 min at an approximate depth of 0.5 m. This depth was chosen considering that the sampling sites have maximum depths of 2 m. In the laboratory, cells were isolated under a Zeiss AXIO Vert A1 inverted microscope with fine tip capillaries. Subsequently, cells were transferred to 96-well plates with culture medium and were progressively scaled-up to 20–25 ml of media in 50-ml culture tubes.
2.3 Strains and growth conditions
Strains used in this study were provided by the IPN-CICIMAR (Instituto Politécnico Nacional, Centro Interdisciplinario de Ciencias Marinas) dinoflagellate collection (Table 1). The PNCETMAR-1 strain was maintained in modified GSe medium (Blackburn et al. 2001; Bustillos-Guzmán et al. 2015) and the PNCETMAR-2 strain in modified K medium (Keller et al. 1987), both were modified with the addition of vermicompost extract. Strains were maintained at a salinity of 34, at 24 °C ± 1 °C, and 120–150 μmol m−2 s−1 in a 12 h light: 12 h dark cycle. Strains were maintained in 50-ml culture tubes with 20 ml of medium and 1 ml of stock culture (inoculum) with monthly reseeding.
Strain details of Prorocentrum cf. cassubicum from Bahía de La Paz, Gulf of California.
| Strain | Date of isolation | Place of isolation | Isolated by |
|---|---|---|---|
| PNCETMAR-1 | 09/08/2018 | CETMAR beach, Bahía de La Paz, southern Gulf of California | C.J. Band-Schmidt |
| PNCETMAR-2 | 01/07/2019 | CETMAR beach, Bahía de La Paz, southern Gulf of California | A.E. Ramos-Santiago |
2.4 Light microscopy
For morphological identification, in general, cell shape and size, symmetry/asymmetry of the lateral thecal plates, periflagellar platelets, pore types and their distribution pattern on the thecal plates were considered. Light microscopy (bright field) photographs were obtained from live cells (Axio Vert.A1 inverted microscope, Carl Zeiss, Germany). An epifluorescence microscopy was used (Axio Scope.A1 compound microscope, Carl Zeiss, with EC-Neofluar Plan 40×/0.75 and 63×/0.95 objectives, and 6-megapixel Axiocam 506 color camera), applying 0.2 % Calcofluor White M2R stain (Fritz and Triemer 1985) for the theca and DAPI (Sigma; 4′,6-diamidino-2-phenylindole; Kapuscinski 1995) to visualize the position and size of the nucleus.
2.5 Scanning electron microscopy
Two techniques were used for scanning electron microscope (SEM) analysis, from which micrographs were obtained. Samples of 2 ml of strain PNCETMAR-1 were fixed with 4 % formaldehyde and processed as follows: a preliminary wash in distilled water was done to remove the fixative and salts (five times), followed by dehydration in a series of ethanol solutions of increasing concentration (30, 50, 70, 90 and 100 %) and two washes with 100 % ethanol (30 min per step in 1.5-ml Eppendorf tubes, without centrifugation). The samples were mounted directly on a 12.7-mm aluminum specimen stubs with a micropipette, dried at room temperature, and coated by high-resolution sputtering (Polaron SC7640, Quorum Technologies, Newhaven, SXE, UK) with Au-Pd. Observations were made and micrographs were taken with a JEOL JSM-7600F SEM at 5 kV, at a working distance of 15.9–16.7 mm, in the Laboratorio Nacional para el Estudio de Nano y Biomateriales of the Departamento de Física Aplicada del Centro de Investigación y Estudios Avanzados of the Instituto Politécnico Nacional (CINVESTAV-IPN), Unidad Mérida, Yucatán, Mexico.
Samples (1 ml) of the PNCETMAR-2 strain were prefixed with 0.5 ml of 4 % glutaraldehyde and left in darkness for 90 min. Subsequently, post fixation was performed with 2 % osmium tetroxide (OsO4); the samples were left in darkness for 2 h until cells were blackened (indicating complete fixation). Prefixation and postfixation was done at room temperature (∼24 °C). After fixation, several centrifugation washes (Thermo Scientific™ Sorvall™ Legend™ XTR, USA) were performed with cold sterile distilled water to remove fixative residues. Gradual dehydration was performed with ethanol (EtOH) at 10, 20, 30, 40, 40, 50, 60, 70, 80, 90 % and twice at 100 %. At each dehydration step, cells were gently resuspended for 1 min and centrifuged at 800g at 4 °C for 5 min. To the decanted samples, 100–200 μl of hexamethyldisilazane (HMDS) were added, and samples were then placed in 0.30-mm metal sample holders (1 cm2 surface area), and left to dry for 12 h at room temperature. Samples were kept in a desiccator with silica gel until they were sent to the Servicio Académico de Microscopía Electrónica de Barrido (SAMEB) of the Instituto de Ciencias del Mar y Limnología (ICMyL), Universidad Nacional Autónoma de México (UNAM) in Mexico City. The samples were coated with Au in an ionizer (Ion Sputter JEOL-JFC-1100, Japan) for 5 min and examined in the JEOL JMS-6360-LV type SEM at a voltage of 10–20 kV and a working distance of 20 mm. The micrographs were edited with Photoshop CS6 Portable® (v. 13.1.2, 2013). We followed unified terminology for morphological characters of the Prorocentrum species (Hoppenrath et al. 2013; Tillmann et al. 2019).
2.6 DNA extraction, amplification, and sequencing
DNA extraction was performed using the Quick-DNA™ Miniprep plus universal (Zymo Research, Irvine, CA, USA). A mixture of 4 µl of PCR Master mix 5X (Gene and Cell Technologies, USA), 14.8 µl of H2O milli-Q, 2 µl of primers and 2 µl of DNA was used for amplification. For DNA amplification, 1:100 and 1:500-µl dilutions (DNA: H2O milli-Q) were used. Published primers for the 28S and ITS regions of rDNA were used (Table 2). All PCR reactions were performed using an iCycler PCR System (Bio-Rad Laboratories, California, USA). The amplification program for the 28S region (gen D1–D2) of the rDNA consisted of a denaturation step at 95 °C for 5 min, followed by 35 cycles at 95 °C for 60 s, alignment temperature of 52 °C for 60 s, 72 °C for 120 s, and finally an extension at 72 °C for 7 min. The program for the ITS region consisted of a denaturation at 94 °C for 230 s, followed by 35 cycles at 94 °C for 50 s, alignment temperature of 47 °C for 60 s and 72 °C for 80 s, ending with an extension at 72 °C for 10 min. PCR products were purified using Quick-DNATM Miniprep kit (Zymo Research) and sent for sequencing to MCLAB (San Francisco, CA, USA).
Primers for 28S and ITS rDNA used in this study.
| Oligonucleotides | Forward | Reverse | Reference |
|---|---|---|---|
| 28S | ACCCGCTGAATTTAAGCATA | TGGTCCGTGTTTCAAGAGACT | Hosoi-Tanabe et al. (2006) |
| ITS | CCTCGTAACAAGGHTCCGTAGGT | CAGATGCTTAARTTCAGCRGG | Adachi et al. (1996) |
2.7 Sequence alignment and phylogenetic analyses
Sequences were edited with Sequencher v. 4.1.4. BLAST analyses of the consensus sequences of this study were performed with sequences from the GenBank database (https://blast.ncbi.nlm.nih.gov/Blast.cgi). Sequences from the corresponding regions (28S and ITS) were selected for the reconstruction of the phylogenetic trees, and sequences from Karenia brevis (C.C. Davis) Gert Hansen et Moestrup were chosen as an outgroup. Sequence alignment was performed with the program MEGA11 v. 10.0.5 (Tamura et al. 2021) using the multiple alignment algorithm. With JModel test v. 2.1.10 (Darriba et al. 2012), the models that best described the nucleotide substitution rates of the sequences were selected. Mega v. 10.0.5 was used for the maximum likelihood (ML) and maximum parsimony (MP) algorithms, and Mr. Bayes v. 3.2.7a (Huelsenbeck and Ronquist 2001) for Bayesian inference (BI). The construction of the trees was performed with a bootstrap of 1000 replicates for ML and MP, and 3 × 106 generations for BI. The trees were edited with FigTree v. 1.4.4-2006.
2.8 Growth curve
The growth curve of the PNCETMAR-1 strain was determined in GSe (Blackburn et al. 2001) and f/2 media (Guillard and Ryther 1962) modified with the addition of vermicompost extract, pre-adapting the cells to each culture medium for at least three generations. For this, monocultures were grown in 250-ml Erlenmeyer flasks with an initial concentration of 500 cells ml−1 in triplicate. Growth curves were determined by cell counts carried out every second day; for this, 2 ml of each replicate was taken and fixed with acid Lugol (Throndsen 1979). Counts were performed in a Sedgwick-Rafter chamber with a 1-ml capacity in an Axio Vert.A1 inverted microscope in brightfield. The identification of the growth phases (lag, exponential, stationary, and decay phases) were carried out following the criteria of Vonshak and Maske (1982). The growth rate (K) in the exponential phase was determined according to Guillard (1973), where C1 is the cell density (cells ml−1) at the initial time (T1) of the exponential phase, and C2 is the cell density (cells ml−1) at the final time (T2) of the exponential phase.
3 Results
3.1 Morphological observations
Single cells, growing in clusters on the culture tube wall (Figure 2A and B), had one to three pusules comprising a quarter of the cells (Figure 2C–G). The cells are oval to oblong (Figures 2C–H, 4A and B and 5A–E). Generally, a circular pyrenoid is in the center of the cells (diameter of 3.28 ± 0.33 µm, n = 10); in a few cases, two pyrenoids per cell could be observed (Figures 2H–K and 4A–B). Chloroplasts were generally peripheral, with a coloration from green to yellow (Figure 4A–E). Cells with round green bodies, possibly oil droplets, were observed, although in some cases the latter showed autofluorescence, so they could be pigment accumulation bodies (Figures 2L–N and 4C–J). The nucleus is located posteriorly in the cells (Figure 4K–Q). The right thecal plate is convex (slight indentation in the center), while the left thecal plate is more flattened (Figures 4R and S and 5A–F). In the right lateral view, they show an asymmetry with respect to their sagittal axis (Figures 2C and D, 4R and S and 5A–E). Cells of strain PNCETMAR-1 measured 18.79 ± 1.25 µm long and 12.15 ± 1.01 µm deep (n = 30), while strain PNCETMAR-2 measured 18.47 ± 1.53 µm long and 11.83 ± 1.05 µm deep (n = 38). Cells did not show a developed apical spine (Figure 5A–F). The intercalary band (IB) that forms the junction of the lateral thecal plates is thin in non-dividing cells, while in cells showing possible division, the IB is broad, with a smooth, spaced horizontal and transverse striation (Figure 5G and H). In the mid-apical region, a tubular-shaped protuberance may be present, which is an extreme intercalary band along the periflagellar area with a smooth striation on its surface (Figure 5I and J). The periflagellar area has a “V” shaped form consisting of seven visible platelets (Figure 5I and J). The thecal plates do not show a regular pattern of pore distribution, and only one type of pore, although variable in size (Figure 5K–L) with a center mostly devoid of pores (Figure 5C–E), was observed, and a total of 108 fine pores (diameter of 0.10 ± 0.02 µm) were counted on the right thecal plate (Figure 5C–E).
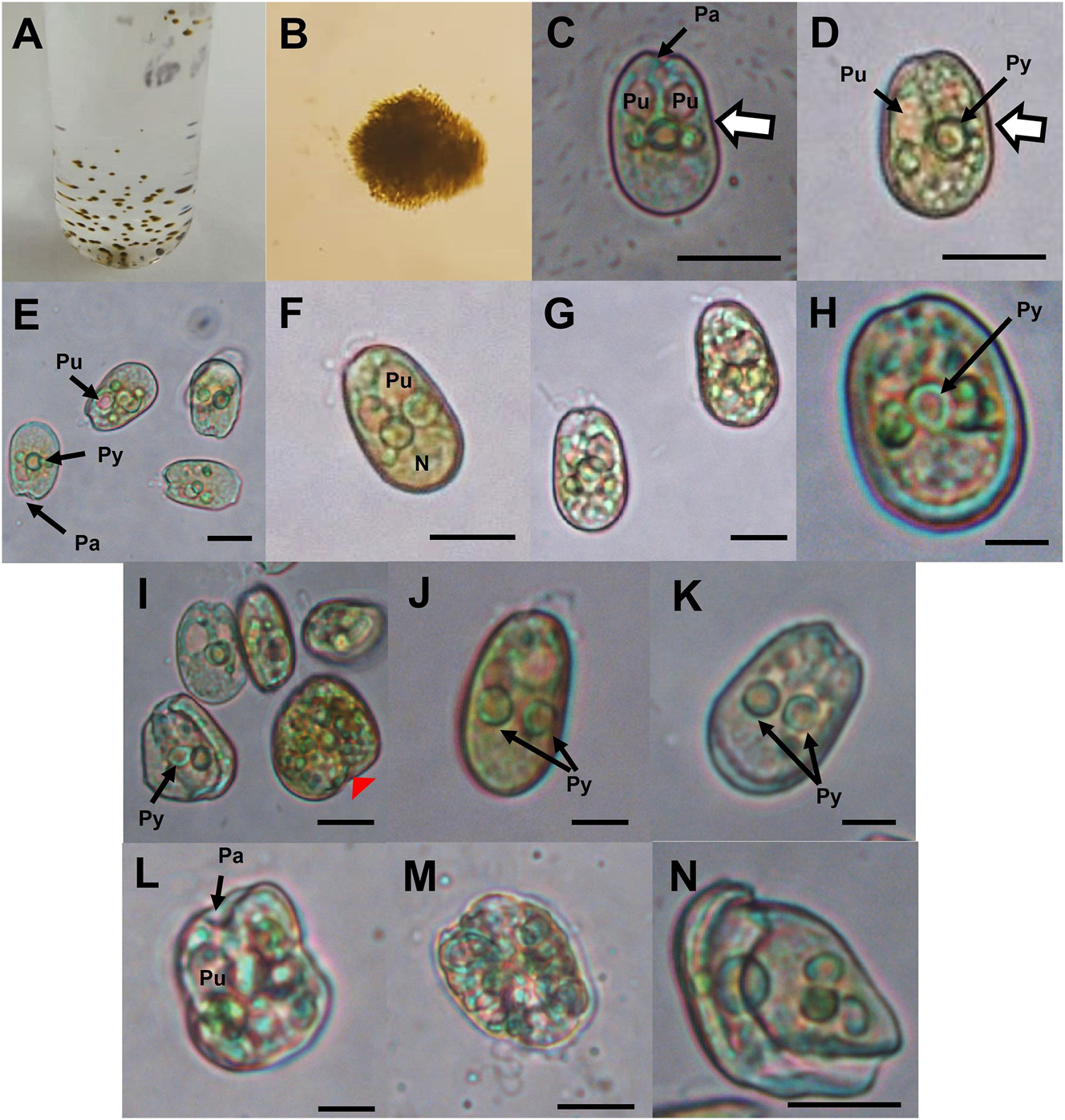
Cells of strain PNCETMAR-2 under a light microscope. (A and B) Cell clusters. (C–H, J, K) General view, showing periflagellar area, flagella, pyrenoid, pusules, chloroplasts, nucleus and concave margin (white arrow with margin). (I, L) Granulated cells in the process of division (red arrowhead). (L–N) Irregular planozygotes. Py, pyrenoid; Pa, periflagellar area; Pu, pusule; N, nucleus. Scale bars: 10 µm in (C–G, I and L–N); 5 µm in (H, J, K).

Cells of strain PNCETMAR-2 under a light microscope. (A–D) Planozygotes initiating cell division; the red arrowhead indicates deformation at cell contact or junction, division planes are indicated by asterisks (*) and a concave margin (white arrow). (E–J) Gametes fused in the central valve zone of the lateral thecal platelets, finishing division in the periflagellar area. (K–L) Three- and four-cell stages (arrows indicate the number of planozygotes). LF, longitudinal flagellum; ab, accumulation bodies; Py, pyrenoid; Pu, pusule; tr, translucent rounded bodies; N, nucleus; Chl, chloroplasts. Scale bars: 10 µm in (A, D, E, F, J and L); 5 µm in (C, B, G, H, I and K).
![Figure 4:
Micrographs of Prorocentrum cf. cassubicum: (A–C, E–G, I, K–Ñ) strain PNCETMAR-1; (D, H, J, O–S) strain PNCETMAR-2. (A–B, I, J, M and P) Cells under phase-contrast microscope; concave margin [broad white arrow in (B)]. (D–H, K, L, N–O and Q–S) Cells under epifluorescence microscope, stained with DAPI, showing the shape and position of the nucleus. (D–H) Epifluorescence images of chloroplasts. (F–H) Accumulation bodies with autofluorescence are shown. (R and S) Cells stained with Calcofluor White M2R; the thecal surface is shown. Pa, periflagellar area; LF, longitudinal flagellum; ab, accumulation bodies; Py, pyrenoid; Pu, pusule; N, nucleus; Chl, chloroplast. Scale bars: 10 µm in (A, M, N, F, P and Q); 5 µm in (C, D, G, H–L, Ñ, O, P–S).](/document/doi/10.1515/bot-2025-0010/asset/graphic/j_bot-2025-0010_fig_004.jpg)
Micrographs of Prorocentrum cf. cassubicum: (A–C, E–G, I, K–Ñ) strain PNCETMAR-1; (D, H, J, O–S) strain PNCETMAR-2. (A–B, I, J, M and P) Cells under phase-contrast microscope; concave margin [broad white arrow in (B)]. (D–H, K, L, N–O and Q–S) Cells under epifluorescence microscope, stained with DAPI, showing the shape and position of the nucleus. (D–H) Epifluorescence images of chloroplasts. (F–H) Accumulation bodies with autofluorescence are shown. (R and S) Cells stained with Calcofluor White M2R; the thecal surface is shown. Pa, periflagellar area; LF, longitudinal flagellum; ab, accumulation bodies; Py, pyrenoid; Pu, pusule; N, nucleus; Chl, chloroplast. Scale bars: 10 µm in (A, M, N, F, P and Q); 5 µm in (C, D, G, H–L, Ñ, O, P–S).

Prorocentrum cf. cassubicum cells in a scanning electron microscope. (A–B, F, G and L) strain PNCETMAR-2. (C–E and H–K) strain PNCETMAR-1. (G) Antapical view of a cell with a broad intercalary band (BI). (H) Cells of different sizes; black arrows indicate horizontal striation; yellow arrows show transverse striation. (I–J) Rounded cell with prolongation of the periflagellar area. (K–L) Detail of thecal surface. IB, intercalary band; RTP, right thecal plate; LTP, left thecal plate; as, posterior tip of the periflagellar area; SS, sagittal suture. Scale bars: 4 µm in (A–H); 1 µm in (I–L).
3.2 Reproductive stages
Different stages of cell division were documented in the PNCETMAR-2 strain. Deformed motile cells have a widened periflagellar zone; granular cells with colorless circular bodies generally marked the beginning of the division process (Figure 3A–D). The possible fusion of gametes starting from the intercalary band zone was observed, showing changes in morphology in the contact zone (Figure 3A). Other reproductive stages were documented in cells with preserved longitudinal flagella showing the division lines (Figure 3B–D). Possible asexual reproductive stages were documented in daughter cells, which separated from the posterior zone ending in the periflagellar area (Figure 3E–J). Dyads, triads and tetrads were also observed (Figure 3J–L). The pusule (one to three per cell) and the pyrenoid were observed in both motile and reproducing cells (generally immotile), the latter being larger.
3.3 Phylogenetic position of Prorocentrum cf. cassubicum
Phylogenetic analysis of the 28S rDNA region, using the Kimura 2-parameter model, grouped the sequence of strain of PNCETMAR-2 into a well-supported clade (bootstrap = 100) with the sequence strain of PNCETMAR-1 (Figure 6). These two sequences showed a degree of genetic divergence of 0.02. Both sequences in this study diverged from the five sequences identified as P. cf. norrisianum from the study of Chomérat et al. (2019), with a genetic distance of 0.22 with respect to the sequences in this study, that were also separated from the P. lima sequences with a genetic distance of 0.29. In contrast, employing the Kimura 2-parameter model, the PNCETMAR-2 (PQ182182) strain sequence of the ITS region clustered with those of P. cassubicum (EU927557) and Prorocentrum sp. (FJ160593) with a 100 % bootstrap support, and genetic distances of 0.030 and 0.032, respectively (Figure 7).

Phylogenetic tree of Prorocentrum from the 28S rDNA region. The phylogenetic tree was obtained using the maximum parsimony (MP), maximum likelihood (MV) and Bayesian inference (BI) methods. The nodes show the bootstrap percentage and posterior probability (MP/MV/BI). The analysis included 19 sequences; dataset based on 660 bp.
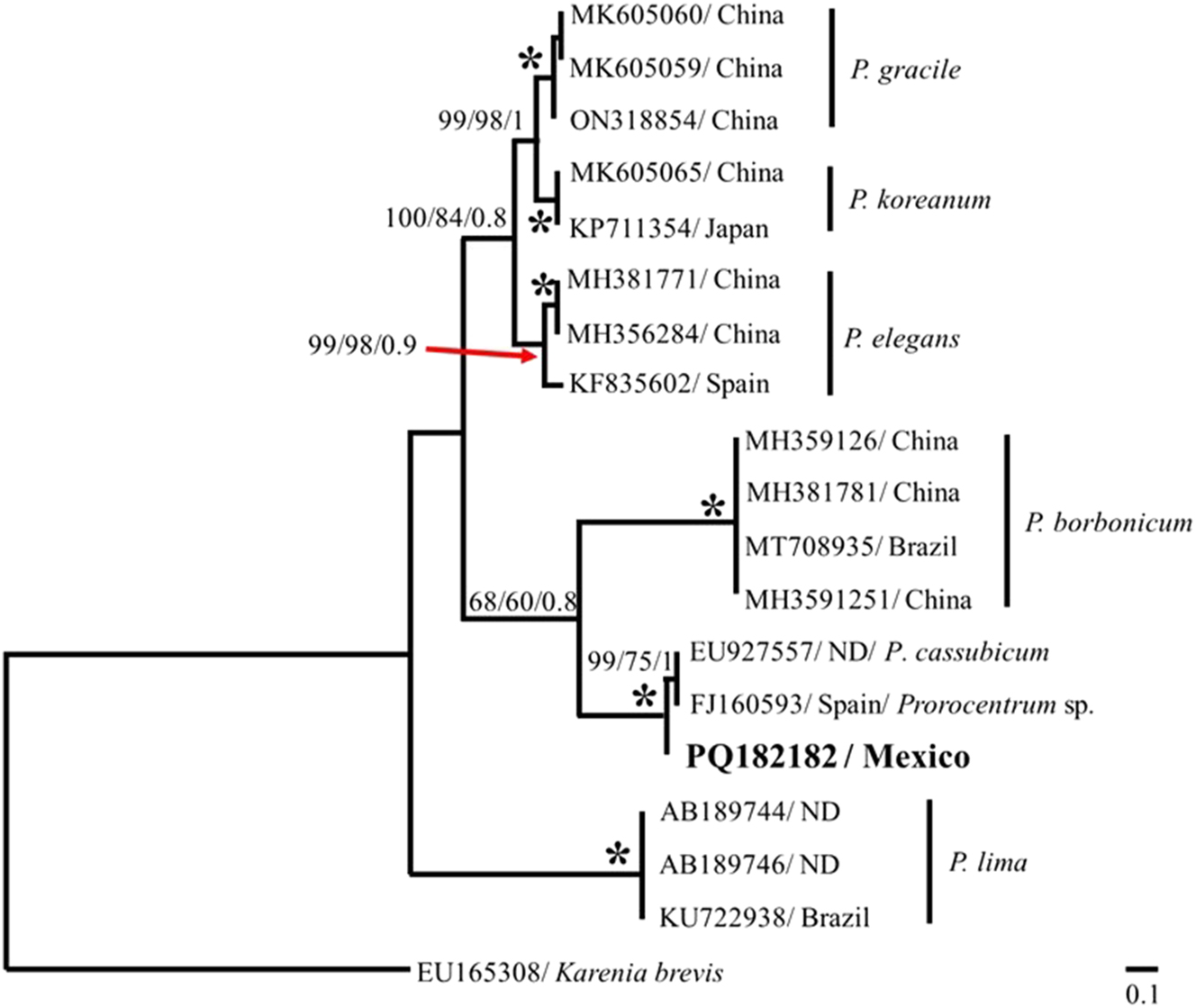
Phylogenetic tree of Prorocentrum from the ITS region of rDNA. The phylogenetic tree was developed using the maximum parsimony (MP), maximum likelihood (ML) and Bayesian inference (BI) methods. The percentage of bootstrap and posterior probability (MP/ML/BI) are shown at the nodes. The analysis included 19 sequences; dataset based on 524 bp. ND, no data.
3.4 Growth curve
The growth curves of strain PNCETMAR-1 showed no significant differences when grown in GSe and f/2 media. In both culture media the adaptation phase lasted 10 days (Figure 8). The exponential phase in f/2 medium ended on day 34 and in GSe medium on day 36, reaching a maximum biomass of 74,312 ± 2,008 cells ml−1 and 69,766 ± 2,154 cells ml−1, respectively (Figure 8). The growth rate in f/2 medium was 0.428 ± 0.002 divisions per day and with GSe medium was 0.464 ± 0.006 divisions per day. This strain is characterized by individual free-swimming, highly motile vegetative cells during the adaptive and exponential phase; they decrease in motility and form clumps during the late exponential phase.

Growth curve of the PNCETMAR-1 strain in GSe ( ) and f/2 (
) and f/2 ( ) medium at 24 °C, salinity of 34 and a 12 h light: 12 h dark cycle. Error bars indicate standard deviations (n = 3).
) medium at 24 °C, salinity of 34 and a 12 h light: 12 h dark cycle. Error bars indicate standard deviations (n = 3).
4 Discussion and conclusions
4.1 Morphology
Prorocentrum norrisianum is from 14 to 25 µm long and from 10 to 16 µm wide (Faust 1997; Hoppenrath et al. 2023; Mohammad-Noor et al. 2007). Strains identified as Prorocentrum cf. norrisianum have larger cells, ranging from 20 to 37 µm long and 20–26 µm wide (Chomérat et al. 2019). One of the characteristics previously considered to differentiate P. norrisianum from P. cassubicum is its size; the cells of the former are smaller (Table 3). However, there is a discrepancy between the reported sizes because there is an overlap in the cell size ranges reported to date (Table 3). Another species often confused with P. norrisianum and P. cassubicum is P. lima; however, the latter is larger, with a length from 30 to 57 µm and depth from 21 to 46 µm (Hoppenrath et al. 2023; Nava-Ruiz and Valadez-Cruz 2012).
Morphometry of Prorocentrum cassubicum and P. norrisianum from different geographical areas.
| Species | Length (µm) | Width (µm) | Origin | Environmental conditions | Remarks | References |
|---|---|---|---|---|---|---|
| Exuviaella cassubica (P. cassubicum) | 22 | 15 | Baltic Sea | Shallow, low salinity | Planktonic, associated with purple bacteria | Wołoszyńska (1928) a |
| P. cassubicum | 30 | – | Woods Hole, USA | – | – | Dodge and Bibby (1973) |
| P. cassubicum | 22–25 | 16 | – | – | – | Dodge (1975) a |
| P. cassubicum | 21–37.5 | 16.8–27.5 | Cancún, Quintana Roo, Mexico | Lagoon, average depth of 1.7 m, 27.3 °C, and average salinity of 8.7 | Samples obtained with a Van Dorn 2 L at a depth of 0.3 m | Nava-Ruiz and Valadez-Cruz (2012) |
| P. cf. cassubicum (PNCETMAR-1) | 26.56–34.79 | 18.56–25.95 | Bahía de La Paz, Mexico | – | – | Villa-Arce (2021) |
| P. cf. cassubicum (PNCETMAR-1)b | 16.76–21.42 | 10.92–14.52 | Bahía de La Paz, Mexico | 1 and 2 m depth, with temperatures from 22 to 28.4 °C, and salinity from 27.2 to 40.3 | Phytoplankton net samples | This study |
| P. cf. cassubicum (PNCETMAR-2) | 14.7–20.3 | 9.8–13.7 | Bahía de La Paz, Mexico | 1 and 2 m depth, with temperatures from 22 to 28.4 °C, and salinity from 27.2 to 40.3 | Phytoplankton net samples | This study |
| P. norrisianum | 20–25 | 13–16 | The Lair at Twin Cays, Belize | 2–3 m depth, 30–36 °C, 30–34 average salinity | Benthic, associated with detritus | Faust (1997) a |
| P. norrisianum | 14–18 | 10–12 | Malaysia | – | Seagrass and macroalgae Padina spp. | Mohammad-Noor et al. (2007) a |
| P. norrisianum | 20–25 | 11–16 | Malaysia | – | Sand samples analyzed | Al-Has and Mohammad-Noor (2011) a |
| P. cf. norrisianum | 31–37 | 20–26 | Caribbean Sea | – | Surface sediment samples | Chomérat et al. (2019) |
| P. norrisianum | 14–25 | 10–16 | – | – | Summary reports | Hoppenrath et al. (2023) a |
-
Measurements that agree with those reported in this study are indicated with a superscript a. The superscript b indicates the revision of strain PNCETMAR-1 (Villa-Arce 2021), in which cell sizes were corrected in the present study.
Another trait that has also been proposed to distinguish between these two species is the shape of the periflagellar platelets: P. cassubicum has curved platelets, while the apical platelets of P. norrisianum are rectangular, vertical or curved (Faust 1997). In summary, due to phenotypic plasticity, the descriptions used for P. norrisianum and P. cassubicum could apply to both species. For example, P. norrisianum is described as an oval species with a length between 20 and 25 µm, with a central pyrenoid (Al-Has and Mohammad-Noor 2011); both features are also reported for P. cassubicum (Dodge and Bibby 1973). Also, cells of P. norrisianum are described as compressed, with the lateral thecal plates ornamented with fine pores and a triangular or V-shaped periflagellar area (Al-Has and Mohammad-Noor 2011); these characteristics are also described in P. cassubicum (Nava-Ruiz and Valadez-Cruz 2012). Therefore, it is recommended to perform an exhaustive analysis in strains that are identified both morphologically and molecularly, in which there are non-contradictory morphometric differences that will help to establish characters to differentiate these two species in future studies.
In Bahía de La Paz, Villa-Arce (2021) reported P. cassubicum with a description of the morphological characteristics and a sequence of the 28S rDNA region of strain PNCETMAR-1. The only difference between the strain of this study (PNCETMAR-2) and strain PNCETMAR-1 was its size (Table 3). Therefore, the biological material used by Villa-Arce (2021) was requested for review, and micrographs were taken to measure the cells, as well as epifluorescence micrographs to observe other characteristics, such as the position of the nucleus and the pigment distribution. This strain presented smaller sizes than those originally reported by the author. Therefore, this study complements the information on both strains to determine if they are distinct species or, given the limited information, to report them as P. cf. cassubicum until more precise, non-contradictory and detailed information on both species is available.
The strains included in this study were isolated from Ensenada de La Paz, a coastal lagoon in the southwest of the Gulf of California. Samples were collected in the water column in a shallow area between 1 and 2 m. These conditions make it difficult to determine whether the isolated cells are benthic or planktonic in nature, since they could have been resuspended in the water column by winds. According to existing reports, P. cassubicum appears to be planktonic, while P. norrisianum is more associated with a benthic habitat (Table 3). Also, precedents indicate that P. cassubicum has been observed in brackish-water environments with a salinity of <10 (Nava-Ruiz and Valadez-Cruz 2012; Wołoszyńska 1928), while P. norrisianum has been recorded in areas with salinity higher than 30 (Faust 1997; Table 3). Originally, P. cassubicum was described from brackish water in Gdansk Bay, the southern Baltic Sea, on the Polish coast. Wołoszyńska (1928) did not indicate the salinity of the sampled water; however, we infer a range between 2 and 12, based on recent studies, although they referred to the period of 1950–2015 in a shallow coastal zone between Hel and Władisławowo (Świątek 2019). Therefore, the difference in the salinity in the two habitats (Gdansk Bay and the southern Gulf of California) is significant and could be considered an argument against the conspecificity of the cells inhabiting the two regions in temperate and subtropical zones, respectively. However, the available information is still limited because not all reports include information on the environment from which they are isolated, and this prevents us from establishing definitive criteria for their ecology and distribution patterns.
In culture, cells with asexual processes and colorless translucent rounded bodies were observed, indicating the initiation of cell division. Cells in pairs, in which two longitudinal flagella were observed, were probably evidence of a sexual process (indicative of gamete union); furthermore, these cells were deformed in the union zone. Deformed gametes in the contact zones have also been reported in Prorocentrum cordatum (Ostenfeld) J.D. Dodge (syn.: Prorocentrum minimum (Pavillard) J. Schiller) (Berdieva et al. 2018).
4.2 Molecular phylogeny
Phylogenetic reconstruction with sequences from the 28S rDNA region grouped the sequence of PNCETMAR-1 and PNCETMAR-2 within the same clade with 100 % bootstrap support. Phylogenetic analyses indicate that both strains belong to the same species. Based on the above, it was possible to amplify the ITS region for strain PNCETMAR-1. The phylogenetic reconstruction of this region showed a clade including this sequence with the sequences of P. cassubicum (EU927557) and Prorocentrum sp. (FJ160593) with bootstrap values of 100 %. However, it is crucial to highlight the lack of sequences of P. norrisianum from the ITS region in the GenBank database for comparison. Although there are seven sequences identified as P. cf. norrisianum for the 28S rDNA region, there are only two sequences of P. cassubicum for the same rDNA region; however, the latter sequences are very short and are not included in this analysis. It is important to mention that the seven sequences of P. cf. norrisianum analyzed in this study come from the strains with cell sizes that agree with the original description for P. cassubicum, showing a slightly larger size than P. norrisianum, and with the sizes reported in this study for P. cf. cassubicum. There are no sequences of P. norrisianum and P. cf. norrisianum for the ITS region for comparison. In addition, the sequence of P. cassubicum (EU927557), used for the phylogenetic reconstruction of the ITS region does not come from published material. The lack of verified sequences may cause a bias in the phylogenetic relationships that have been determined so far; therefore, they should be treated with caution. In addition, the lack of sequence information on P. cassubicum from the area of its first record (Baltic Sea) makes it difficult to determine whether it is one or two distinct species.
Current morphometric information is contradictory for both species, even with the few studies that have been carried out that complement morphological analysis with molecular analysis. It is likely that the limited information in the descriptions or reports of these two species is due to the fact that their precise identification cannot be achieved under light microscopy after sampling, due to their small cell size. This study reports for the first time the presence of P. cf. cassubicum in the region of the Ensenada de La Paz and the Gulf of California.
4.3 Growth curve
Prorocentrum cf. cassubicum grows slowly under the culture conditions tested, taking more than 30 days to reach its maximum biomass. No significant differences in maximum biomass (69,722–74,312 cells ml−1), nor in growth rate (0.43–0.46 divisions day−1) were observed when cultivating strain PCCETMAR-1 in f/2 and GSe media at 24 °C. No growth data are available for other strains of P. cassubicum and P. norrisianum. In general, epibenthic dinoflagellate species exhibit slow growth. Because of its toxicity, one of the most studied species is the P. lima complex (Grigoriyan et al. 2024). Numerous experiments have been carried out in the laboratory with various strains under different conditions where it has been determined that temperature is the main factor affecting their growth, registering growth rates between 0.30 and 0.75 divisions day−1 and maximum biomasses of 600 to 70,000 cells ml−1 (Grigoriyan et al. 2024). Several species of Prorocentrum meet the characteristics of being halotolerant and euryhaline (Glibert 2020). More knowledge of epibenthic species is needed to understand their physiology, life cycles, nutrition, and the environmental factors that regulate their growth.
Funding source: BEIFI
Award Identifier / Grant number: A240310
Funding source: CONAHCyT FOMIX-Yucatán
Award Identifier / Grant number: 2008-108160
Funding source: SIP
Award Identifier / Grant number: 2024-0544
Award Identifier / Grant number: 2025-1372
Award Identifier / Grant number: 2189
Funding source: CONAHCyT PRONAII, PRONACES SSyS
Award Identifier / Grant number: 319104
Funding source: CONAHCyT LAB-2009-01
Award Identifier / Grant number: 123913
Funding source: CONAHCyT
Award Identifier / Grant number: 1151607
About the authors

Ana Emilia Ramos-Santiago is currently a PhD student in Marine Sciences at IPN-CICIMAR, La Paz, Mexico. Her main research interests are epibenthic marine dinoflagellates, sampling techniques, cultivation, and their identification using microscopy and molecular tools. Currently, she is dedicated to understanding the autecology of epibenthic dinoflagellates species from Bahía de La Paz, Mexico.

Gema V. Villa-Arce obtained her degree in Fisheries Engineering at the Universidad Autónoma de Baja California Sur in 2013. During her last year she specialized in marine food chemistry. In 2021 she completed her Bachelor’s thesis, focused on growth curves and identification of dinoflagellate strains of the genus Prorocentrum (Dinophyceae) from the Bahía de La Paz, B.C.S., Mexico at IPN-CICIMAR.

Ignacio Leyva-Valencia is a researcher of the Secretariat of Science, Humanities, Technology, and Innovation, affiliated to IPN-CICIMAR. His main research is focused on the isolation and identification of toxigenic dinoflagellate species using microscopy and molecular biology tools. He also conducts experimental work to determine the cytotoxic and antibacterial activity of dinoflagellate extracts, as well as their effect on early stages of bivalve mollusks.

Yuri Okolodkov started his career in the Academy of Sciences of the URSS in 1973. In 2001 he moved to Mexico by invitation of CONAHCyT. Since 2005 he has been a researcher at the Universidad Veracruzana where he founded the Laboratory of Marine Botany and Planktology, as well as the algae collection of the institute. One of his main interests is the taxonomy of marine dinoflagellates.

Christine J. Band-Schmidt is a researcher at IPN-CICIMAR where she is leader of the research team studying harmful marine dinoflagellates. She is a founding member of the Mexican Society for the Study of Harmful Algae (SOMEFAN, A.C.) and the CONAHCyT network of harmful algal blooms (RedFAN). Her research has been focused mainly on the autecology of harmful dinoflagellate species from the Gulf of California, with particular interest in understanding the interactions between phytoplankton species and bacteria-phytoplankton interactions.
Acknowledgments
The authors thank L.E. Gómez-Lizárraga for the SEM service at UNAM and D.A. Huerta-Quintanilla (CINVESTAV-IPN) for obtaining SEM micrographs, E. Mendez (University of Tennessee, Knoxville, TN, USA) and M.M. Gowing (Seattle, WA, USA) for language edition support, and A.F. Krakhmalnyi (Institute for Evolutionary Ecology of the National Academy of Sciences of Ukraine, Kiev, Ukraine) for translation from Polish.
-
Research ethics: Not applicable.
-
Informed consent: Not applicable.
-
Author contributions: A.E. Ramos-Santiago: conceptualization; research; methodology; data curation; formal analysis; writing – original draft preparation; writing – review & editing. G.V. Villa-Arce: methodology; data curation; formal analysis. I. Leyva-Valencia: funding acquisition; methodology; supervision; review & editing. Y.B. Okolodkov: conceptualization; research; methodology; writing – review & editing; supervision. C.J. Band-Schmidt: conceptualization; funding acquisition; methodology; supervision; writing – review & editing. All authors have accepted responsibility for the entire content of this manuscript and approved its submission.
-
Use of Large Language Models, AI and Machine Learning Tools: None declared.
-
Conflict of interest: The authors state no conflict of interest.
-
Research funding: This study was financed with the projects CONAHCyT FOMIX-Yucatán No. 2008-108160 and CONAHCyT LAB-2009-01 No. 123913 (P. Quintana-Owen from CINVESTAV-IPN), CONAHCyT PRONAII, PRONACES SSyS No. 319104, and the institutional project SIP-2024-0544, SIP 2025-1372 and SIP 2189. A.E.R.S. was recipient to the scholarship 1151607 from CONAHCyT and A240310 from BEIFI. C.J.B.S. is a COFAA and EDI recipient.
-
Data availability: DNA sequences, GenBank accessions. Strain PNCETMAR-1: PQ182180 (28S), PQ182182 (ITS). Strain PNCETMAR-2: PQ182181 (28S).
References
Adachi, M., Sake, Y., and Ishida, Y. (1996). Analysis of Alexandrium (Dinophyceae) species using sequences of the 5.8S ribosomal DNA and internal transcribed spacer regions. J. Phycol. 32: 424–432, https://doi.org/10.1111/j.0022-3646.1996.00424.x.Search in Google Scholar
Al-Has, A. and Mohammad-Noor, N. (2011). Identification of marine sand-dwelling dinoflagellates in Dinawan Island, Sabah. Borneo Sci. 28: 37–45.Search in Google Scholar
Álvarez-Arellano, A.D., Rojas-Soriano, H., and Prieto-Mendoza, J.J. (1997) Geología de la Bahía de La Paz y áreas adyacentes. In: Urbán, J., and Ramírez Rodríguez, M. (Eds.), La Bahía de La Paz, investigación y conservación. Universidad Autónoma de Baja California Sur, Centro Interdisciplinario de Ciencias Marinas, La Paz, Baja California Sur, México, pp. 13–29.Search in Google Scholar
Berdieva, M., Pozdnyakov, I., Matantseva, O., Knyazev, N., and Skarlato, S. (2018). Actin as a cytoskeletal basis for cell architecture and a protein essential for ecdysis in Prorocentrum minimum (Dinophyceae, Prorocentrales). Phycol. Res. 66: 127–136, https://doi.org/10.1111/pre.12214.Search in Google Scholar
Blackburn, S.I., Bolch, C.J.S., Haskard, K.A., and Hallegraeff, G.M. (2001). Reproductive compatibility among four global populations of the toxic dinoflagellate Gymnodinium catenatum (Dinophyceae). Phycologia 40: 78–87, https://doi.org/10.2216/i0031-8884-40-1-78.1.Search in Google Scholar
Borsato, G.T., Salgueiro, F., De’ Carli, G.A.L., Morais, A.M., Goulart, A.S., de Paula, J.C., and Nascimento, S.M. (2023). Taxonomy and abundance of epibenthic Prorocentrum (Dinophyceae) species from the tropical and subtropical Southwest Atlantic Ocean including a review of their global diversity and distribution. Harmful Algae 127: 102470, https://doi.org/10.1016/j.hal.2023.102470.Search in Google Scholar PubMed
Bustillos-Guzmán, J.J., Band-Schmidt, C.J., Durán-Riveroll, L.M., Hernández-Sandoval, F.E., López-Cortés, D.J., Núñez-Vázquez, E.J., Cembella, A., and Krock, B. (2015). Paralytic toxin profile of the marine dinoflagellate Gymnodinium catenatum Graham from the Mexican Pacific as revealed by LC-MS/MS. Food Addit. Contam. Part A: 1–14, https://doi.org/10.1080/19440049.2014.1000978.Search in Google Scholar PubMed
Carmona, R., Ruiz-Campos, G., Castillo-Guerrero, J.A., and Zamora-Orozco, M. (2002). Current distribution status of the white-faced ibis, Plegadis chihi, in the Peninsula of Baja California, Mexico, with special reference to the region of La Paz. Southwest Natl. 47: 476–482, https://doi.org/10.2307/3672509.Search in Google Scholar
Cembella, A.D., Durán-Riveroll, L.M., Tarazona-Janampa, U.I., Okolodkov, Y.B., García-Sandoval, R., Krock, B., Hörstmann, C., and John, U. (2021). Phylogeography and diversity among populations of the toxigenic benthic dinoflagellate Prorocentrum from coastal reef systems in Mexico. Front. Mar. Sci. 8: 716669, https://doi.org/10.3389/fmars.2021.716669.Search in Google Scholar
Chávez, H. (1985). Aspectos biológicos de las lisas (Mugil spp.) de la Bahía de La Paz, B.C.S, México, con referencia especial a juveniles. Investig. Mar. CICIMAR 2: 1–22.Search in Google Scholar
Chomérat, N., Bilien, G., and Zentz, F. (2019). A taxonomical study of benthic Prorocentrum species (Prorocentrales, Dinophyceae) from Anse Dufour (Martinique Island, eastern Caribbean Sea). Mar. Biodivers. 49: 1299–1319, https://doi.org/10.1007/s12526-018-0913-6.Search in Google Scholar
Darriba, D., Taboada, G., Doallo, R., and Posada, D. (2012). jModelTest 2: more models, new heuristics and parallel computing. Nat. Methods 9: 772, https://doi.org/10.1038/nmeth.2109.Search in Google Scholar PubMed PubMed Central
Dodge, J.D. (1975). The Prorocentrales (Dinophyceae). II. Revision of the taxonomy within the genus Prorocentrum. J. Linn. Soc., Bot. London 71: 103–125, https://doi.org/10.1111/j.1095-8339.1975.tb02449.x.Search in Google Scholar
Dodge, J.D. and Bibby, B.T. (1973). The Prorocentrales (Dinophyceae): I. A comparative account of fine structure in the genera Prorocentrum and Exuviaella. Bot. J. Linn. Soc. 67: 175–187, https://doi.org/10.1111/j.1095-8339.1973.tb01737.x.Search in Google Scholar
Espinoza, A.J. (1977). Los principales parámetros de nitratos y silicatos en la Ensenada de La Paz, MS thesis. Centro de Investigaciones Biológicas del Noroeste, La Paz, Baja California Sur, México.Search in Google Scholar
Faust, M.A. (1997). Three new benthic species of Prorocentrum (Dinophyceae) from Belize: P. norrisianum sp. nov., P. tropicalis sp. nov., and P. reticulatum sp. nov. J. Phycol. 33: 851–858, https://doi.org/10.1111/j.0022-3646.1997.00851.x.Search in Google Scholar
Fritz, L. and Triemer, R.E. (1985). A rapid simple technique utilizing Calcofluor White M2R for the visualization of dinoflagellates thecal plates. J. Phycol. 21: 662–664, https://doi.org/10.1111/j.0022-3646.1985.00662.x.Search in Google Scholar
Glibert, P.M. (2020). Harmful algae at the complex nexus of eutrophication and climate change. Harmful Algae 91: 101583, https://doi.org/10.1016/j.hal.2019.03.001.Search in Google Scholar PubMed
Grigoriyan, A., Lorini, M.L., de Souza Lima Figueiredo, M., Vasconcelos Corrêa, A., and Nascimiento, S.M. (2024). Effects of culture conditions on the growth of the benthic dinoflagellates Ostreopsis cf. ovata, Prorocentrum lima and Coolia malayensis (Dinophyceae): a global review. Harmful Algae 132: 102565, https://doi.org/10.1016/j.hal.2023.102565.Search in Google Scholar PubMed
Guillard, R. (1973) Division rates. In: Stein, R.J. (Ed.), Handbook of phycological methods. Cambridge University Press, London, UK, pp. 289–311.Search in Google Scholar
Guillard, R. and Ryther, J.H. (1962). Studies of marine planktonic diatoms. I. Cyclotella nana Hustedt and Detonula confervacea (Cleve) Gran. Can. J. Microbiol. 8: 229–239, https://doi.org/10.1139/m62-029.Search in Google Scholar PubMed
Guiry, M.D. and Guiry, G.M. (2025). AlgaeBase. World-wide electronic publication. National University of Ireland, Galway, Available at: https://www.algaebase.org (Accessed 03 January 2025).Search in Google Scholar
Han, M.-S., Wang, P., Kim, J.H., Cho, S.Y., Park, B.S., Kim, J.H., Katano, T., and Kim, B.H. (2016). Morphological and molecular phylogenetic position of Prorocentrum micans sensu stricto and description of Prorocentrum koreanum sp. nov. from southern coastal waters in Korea and Japan. Protist 167: 32–50, https://doi.org/10.1016/j.protis.2015.12.001.Search in Google Scholar PubMed
Hinzmann, M., Craveiro, S., and Calado, A. (2008) Quantification of epibenthic communities, including toxic dinoflagellates, in different green macroalgal substrates in Ria de Aveiro (Portugal). In: 12th international conference on harmful algae, Copenhagen, Denmark, pp. 190–192.Search in Google Scholar
Hoppenrath, M., Chomérat, N., Horiguchi, T., Schweikert, M., Nagahama, Y., and Murray, S. (2013). Taxonomy and phylogeny of the potentially toxic, benthic Prorocentrum species (Dinophyceae) – a proposal and review. Harmful Algae 27: 1–28.10.1016/j.hal.2013.03.006Search in Google Scholar
Hoppenrath, M., Murray, S.A., Chomérat, N., and Horiguchi, T. (2014). Marine benthic dinoflagellates – unveiling their worldwide biodiversity. Kleine Senckenberg-Reihe, Vol. 54. Schweizerbart’sche Verlagsbuchhandlung (Nägele u. Obermiller), Stuttgart, Germany, p. 276.Search in Google Scholar
Hoppenrath, M., Chomérat, N., Horiguchi, T., Murray, S.A., and Rhodes, L. (2023). Marine benthic dinoflagellates – their relevance for science and society, 2nd ed. E. Schweizerbart’sche Verlagsbuchhandlung (Nägele u. Obermiller) und Senckenberg Gesellschaft für Naturforschung, Stuttgart, Germany, p. 375.Search in Google Scholar
Hosoi-Tanabe, S., Otake, I., and Sako, Y. (2006). Phylogenetic analysis of noxious red tide flagellates Chattonella antiqua, C. marina, C. ovata, and C. verruculosa (Raphidophyceae) based on the rRNA gene family. Fish. Sci. 72: 1200–1208, https://doi.org/10.1111/j.1444-2906.2006.01277.x.Search in Google Scholar
Huelsenbeck, J.P. and Ronquist, F. (2001). MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 17: 754–755, https://doi.org/10.1093/bioinformatics/17.8.754.Search in Google Scholar PubMed
Kapuscinski, J. (1995). DAPI: a DNA-specific fluorescent probe. Biotech. Histochem. 70: 220–233, https://doi.org/10.3109/10520299509108199.Search in Google Scholar PubMed
Keller, M.D., Selvin, R.C., Claus, W., and Guillard, R.R.L. (1987). Media for the culture of oceanic ultraplankton. J. Phycol. 23: 633–638, https://doi.org/10.1111/j.1529-8817.1987.tb04217.x.Search in Google Scholar
Lundholm, N., Churro, C., Escalera, L., Fraga, S., Hoppenrath, M., Iwataki, M., Larsen, J., Mertens, K., Moestrup, Ø., Murray, S., et al.. (2024). IOC-UNESCO taxonomic reference list of harmful micro algae, IOC-UNESCO, Available at: https://www.marinespecies.org/hab.Search in Google Scholar
Melo, S., Bozelli, R.L., and Esteves, F.A. (2007). Temporal and spatial fluctuations of phytoplankton in a tropical coastal lagoon, southeast Brazil. Braz. J. Biol. 67: 475–483, https://doi.org/10.1590/s1519-69842007000300012.Search in Google Scholar PubMed
Mohammad-Noor, N., Daugbjerg, N., Moestrup, Ø., and Anton, A. (2007). Marine epibenthic dinoflagellates from Malaysia: a study of live cultures and preserved samples based on light and scanning electron microscopy. Nord. J. Bot. 24: 629–690.10.1111/j.1756-1051.2004.tb01938.xSearch in Google Scholar
Murray, S., Camilla, L.C.I., Moore, R., Nagahama, Y., and Fukuyo, Y. (2009). Are prorocentroid dinoflagellates monophyletic? A study of 25 species based on nuclear and mitochondrial genes. Protist 160: 245–264, https://doi.org/10.1016/j.protis.2008.12.004.Search in Google Scholar PubMed
Nava-Ruiz, V.M. and Valadez-Cruz, F. (2012). Flora planctónica de laguna Lagartos, Quintana Roo. Rev. Mex. Biodivers. 83: 561–582, https://doi.org/10.7550/rmb.24868.Search in Google Scholar
Sandoval, F.J. and Gomes-Valdes, J. (1997). Tides and tidal currents in Ensenada de La Paz lagoon, Baja California Sur, Mexico. Geofis. Int. 39: 37–47.10.22201/igeof.00167169p.1997.36.1.619Search in Google Scholar
Świątek, M. (2019). Long-term variability of water temperature and salinity at the Polish coast. Bull. Geogr. Phys. Geogr. 16: 115–129, https://doi.org/10.2478/bgeo-2019-0008.Search in Google Scholar
Tamura, K., Stecher, G., and Kumar, S. (2021). MEGA11: molecular evolutionary genetics analysis version 11, molecular biology and evolution. Mol. Biol. Evol. 38: 3022–3027, https://doi.org/10.1093/molbev/msab120.Search in Google Scholar PubMed PubMed Central
Tarazona-Janampa, U., Cembella, A.D., Pelayo-Zárate, M.C., Pajares, S., Márquez-Valdelamar, L.M., Okolodkov, Y.B., Tebben, J., Krock, B., and Durán-Riveroll, L.M. (2020). Associated bacteria and their effects on growth and toxigenicity of the dinoflagellate Prorocentrum lima species complex from epibenthic substrates along Mexican coasts. Front. Mar. Sci. 7: 569, https://doi.org/10.3389/fmars.2020.00569.Search in Google Scholar
Throndsen, J. (1979) Special methods-micromanipulators. In: Stein, R.J. (Ed.), Handbook of phycological methods: culture methods and growth measurements. Cambridge Press, London, UK, pp. 139–144.Search in Google Scholar
Tillmann, U., Hoppenrath, M., and Gottschling, M. (2019). Reliable determination of Prorocentrum micans Ehrenb. (Prorocentrales, Dinophyceae) based on newly collected material from the type locality. Eur. J. Phycol. 54: 417–431, https://doi.org/10.1080/09670262.2019.1579925.Search in Google Scholar
Verma, A., Kazandjian, A., Sarowar, C., Harwood, D.T., Murray, J.S., Pargmann, I., Hoppenrath, M., and Murray, S.A. (2019). Morphology and phylogenetics of benthic Prorocentrum species (Dinophyceae) from tropical northwestern Australia. Toxins 11: 571, https://doi.org/10.3390/toxins11100571.Search in Google Scholar PubMed PubMed Central
Villa-Arce, G.V. (2021). Curvas de crecimiento e identificación de cepas de dinoflagelados del género Prorocentrum (Dinophyceae) procedentes de la Bahía de La Paz, B.C.S., México, Bachelor´s thesis. Universidad Autónoma de Baja California Sur, La Paz, Baja California Sur, México.Search in Google Scholar
Vonshak, A. and Maske, H. (1982) Algae: growth techniques and biomass production. In: Coombs, J., Hall, D.O., Long, S.P., and Scurlock, J.M.O. (Eds.), Techniques in bioproductivity and photosynthesis. Section 3. Pergamon Press, Oxford, UK, pp. 62–77.Search in Google Scholar
Wołoszyńska, J. (1928). Dinoflagellatae polskiego Bałtyku i Błot nad Piaśnicᶏ. Arch. Hydrobiol. Rybactwa 3: 153–278.Search in Google Scholar
© 2025 the author(s), published by De Gruyter, Berlin/Boston
This work is licensed under the Creative Commons Attribution 4.0 International License.
Articles in the same Issue
- Frontmatter
- In this issue
- Physiology and Ecology
- Three-dimensional distribution of nutrients and phytoplankton biomass in a semi-enclosed region of the Gulf of California during different ENSO phases
- Proliferation of Undaria pinnatifida along the Atlantic coast of the Iberian Peninsula
- Study on fatty acid methyl ester (FAME) composition of 19 Brazilian thraustochytrid isolates
- Taxonomy/Phylogeny and Biogeography
- Variations in the bacterial and fungal community structure along the hypoxic gradient of the Arabian Sea oxygen-depleted environment based on eDNA metabarcoding analysis
- Studies of North Carolina marine algae XV. DNA sequencing reveals some different Ulva species compared to historical reports and U. carsoniae sp. nov. (Ulvales, Chlorophyta)
- Identification and growth of Prorocentrum cf. cassubicum (Dinoflagellata: Prorocentraceae) from Bahía de La Paz: first record for the Gulf of California
- A new oomycete pathogen Olpidiopsis dasysiphoniae sp. nov. (Oomycota) infecting the red alga Dasysiphonia japonica (Ceramiales, Delesseriaceae)
- Chemistry and Applications
- Exploring the suitable extractive species in an IMTA: inorganic nutrient removal from mariculture effluents by commercially important marine macroalgae
Articles in the same Issue
- Frontmatter
- In this issue
- Physiology and Ecology
- Three-dimensional distribution of nutrients and phytoplankton biomass in a semi-enclosed region of the Gulf of California during different ENSO phases
- Proliferation of Undaria pinnatifida along the Atlantic coast of the Iberian Peninsula
- Study on fatty acid methyl ester (FAME) composition of 19 Brazilian thraustochytrid isolates
- Taxonomy/Phylogeny and Biogeography
- Variations in the bacterial and fungal community structure along the hypoxic gradient of the Arabian Sea oxygen-depleted environment based on eDNA metabarcoding analysis
- Studies of North Carolina marine algae XV. DNA sequencing reveals some different Ulva species compared to historical reports and U. carsoniae sp. nov. (Ulvales, Chlorophyta)
- Identification and growth of Prorocentrum cf. cassubicum (Dinoflagellata: Prorocentraceae) from Bahía de La Paz: first record for the Gulf of California
- A new oomycete pathogen Olpidiopsis dasysiphoniae sp. nov. (Oomycota) infecting the red alga Dasysiphonia japonica (Ceramiales, Delesseriaceae)
- Chemistry and Applications
- Exploring the suitable extractive species in an IMTA: inorganic nutrient removal from mariculture effluents by commercially important marine macroalgae