Morphological and molecular characterization of Chnoospora minima (Scytosiphonaceae, Ectocarpales) along Mexican coasts, with the description of C. ramosissima sp. nov.
-
María Luisa Núñez Resendiz
María Luisa Núñez Resendiz is a full-time professor at the Universidad Autónoma Metropolitana, where she received her PhD degree. Currently, she is working on phylogeny, taxonomy and phylogeography of marine algae.

 ,
Kurt M. Dreckmann
,
Kurt M. Dreckmann
Kurt M. Dreckmann is a full-time professor at the Universidad Autónoma Metropolitana, where he received his PhD degree. Over 35 years, his research has focused on ecology, taxonomy, biogeography, and phylogeny of marine algae.
,
Oscar E. Hernández
Oscar E. Hernández is currently doing his PhD degree at Universidad Autónoma Metropolitana, where he is working in biogeography and systematics of marine algal species in Mexico.
,
Carlos Adán Palma-Ortíz
Carlos Adán Palma-Ortíz is currently doing his PhD degree at Universidad Autónoma Metropolitana, where he is working on population genetics, phylogeography and systematics of marine algal species in Mexico.
and
Abel Sentíes
Abel Sentíes is currently a professor-researcher of marine botany at Universidad Autónoma Metropolitana. He has spent over 35 years at the Marine Macroalgae Laboratory, focusing his research on systematics, biogeography, and conservation of seaweeds.

Abstract
Chnoospora is one of the genera in Scytosiphonaceae with many taxonomic changes, comprising only two currently taxonomically accepted species. Chnoospora minima, its type species, has the widest distribution range and is characterized by a simple morphology with cylindrical or flattened thalli and dichotomous branching. On the Mexican coasts, its distribution is disjunct, being recorded on both the Pacific and Atlantic sides. In the present study we aim to characterize morphologically and molecularly the Mexican populations of C. minima. Our results revealed two phylogenetically independent groups, corresponding with different species with sympatric distribution in the tropical Mexican Pacific, C. minima and C. ramosissima sp. nov. Chnoospora ramosissima has the typical morphology of the genus, however, it is distinct in having relatively constant length of the branching segments, that are gradually reducing towards the apical portions, giving the thallus a profusely branched appearance; in addition, cellular measurements also differ between them. In contrast, C. minima, presented five different morphotypes on the Mexican coasts, however, all of them were different from C. ramosissima by having a central branching segment longer than the rest, subsequently reducing towards the apical portions, giving the thallus the appearance of being elongated and little branched.
1 Introduction
The family Scytosiphonaceae Farlow (Ectocarpales, Phaeophyceae) groups 65 species of marine brown algae, distributed in 29 genera, widely recorded in temperate and tropical waters throughout the world (Guiry and Guiry 2024). These species are characterized by a single plastid bearing a large pyrenoid and a heteromorphic life cycle (except for some species of the genera Myelophycus Kjellman and Melanosiphon M.J. Wynne), where a large and parenchymatous erect thallus (gametophyte), whose growth can be subapical or diffuse (Farlow 1881; Santiañez et al. 2018a; Setchell and Gardner 1925; Silberfeld et al. 2011), alternates with a small and pseudoparenchymatous prostrate thallus (sporophyte) (Feldmann 1949; Kogame et al. 1999; Nakamura and Tatewaki 1975; Wynne 1969). However, within the family there is an important problem in defining the phylogenetic relationships of the species (Cho et al. 2006; Lee et al. 2014a; Santiañez et al. 2018b), which is why, with the introduction of molecular markers, numerous taxonomic changes have been proposed, mainly at the generic level (Hoshino et al. 2020; 2021; McDevit and Saunders 2017; Santiañez et al. 2020; Santiañez and Kogame 2019; 2022; Schultz et al. 2015; Vieira et al. 2024).
Chnoospora J. Agardh is one of the genera in which a great number of taxonomic arrangements have been made since its circumscription (Agardh 1847; Nelson and Duffy 1991; Papenfuss 1956; Santiañez et al. 2018b). It was established in 1847 by Agardh based on specimens collected from Venezuela in the Atlantic Ocean and San Agustín, Oaxaca, Mexico in the Pacific Ocean, which he named C. atlantica J. Agardh and C. pacifica J. Agardh, respectively (Agardh 1847). In 1848, he described three new species for the genus, C. pannosa J. Agardh, C. implexa J. Agardh, and C. fastigiata J. Agardh. In the same work, he reduced C. atlantica and C. pacifica to the status of taxonomic varieties of C. fastigiata. However, at that time Agardh never designated a type species for Chnoospora. Chnoospora pacifica and C. fastigiata var. pacifica, were used in the literature until Papenfuss (1956) uncovered the identity and location of the type of Fucus minimus Hering (Nelson and Duffy 1991). This species was briefly described from specimens collected in Port Natal (Durban), South Africa and, because there was no illustration, there was confusion over its identity. In 1956, Papenfus proposed the conspecificity of C. fastigiata with Fucus minimus combined both names as C. minima over C. pacifica, based on Hering’s priority, designating here the specimen from Port Natal, Durban (South Africa) as the lectotype of C. minima. Papenfuss also pointed the similarity of the specimens of C. minima (as F. minima) collected in South Africa by Hering, previously reported by Barton (1898). However, it was not until the work of Huisman (2015), in which he conducted a review of Australian brown algal species that C. minima was designated as the type species for the genus. Subsequently, Santiañez et al. (2018b), based on their observations of specimens from the Philippines, Japan, French Polynesia, as well as comparisons with illustrations of South African specimens by Anderson et al. (2016), pointed out the same similarity between their specimens that Barton and Papenfuss previously reported, also assigning C. minima as the type species for the genus. In their work, Santiañez et al. (2018b) missed reading the work of Huisman with his previous proposal, but it has the taxonomic priority.
When Agardh described Chnoospora implexa (1848), as part of the genus, he also mentioned its dubious identity which needs to be reviewed in detail. In agreement with this observation, Santiañez et al. (2018a) suggested the possibility of segregating C. implexa from the genus, based on genetic and morphological observations. Based on specimens collected in the USA, they establish the phylogenetic and morphological separation of C. implexa, for which they describe the monospecific genus Pseudochnoospora Santiañez, G.Y.Cho et Kogame to accommodate this species.
Currently, the diversity of Chnoospora has been drastically reduced from nine to two taxonomically accepted species, C. bicanaliculata V. Krishnamurthy et Thomas, with Indo-Pacific distribution, and C. minima with global distribution (Guiry and Guiry 2024). Both species are mainly characterized by having erect, branched thalli that are free or inter-adhesive, solid, cylindrical or complanate, with hollows between the medullary cells, a sub-apical growth pattern, axes with acute apices, numerous superficial cryptostomata from which phaeophycean hairs arise, sometimes associated with plurilocular sori covered with a loosened cuticle (Nelson and Duffy 1991; Santiañez and Wynne 2024; Santiañez et al. 2018b).
Chnoospora minima has a wide geographical distribution range, mainly in the tropical regions from the Indo-Pacific Ocean to the Atlantic Ocean (Guiry and Guiry 2024). Particularly in Mexico, it has been widely recorded, on both the coasts of the Mexican Pacific and in the Gulf of Mexico and the Caribbean Sea. In the Mexican Pacific, it has been recorded, from the temperate regions of Baja California to Sonora and Nayarit, and the tropical regions of Jalisco, Colima, Michoacan to Guerrero and Oaxaca (Pedroche and Sentíes 2020; Pedroche et al. 2008). On the Mexican Atlantic side, it has been recorded along the coasts of Veracruz, Tabasco, and Quintana Roo, in the tropical region (García-García et al. 2021). Given its wide and disjunct distribution range on the Pacific and Atlantic Mexican coasts, our objective was to characterize morphologically and molecularly to the populations of C. minima. Additionally, we found a phylogenetically independent group that we have described as C. ramosissima sp. nov.
2 Materials and methods
Thirty-four samples tentatively identified in the field as Chnoospora minima using external morphological characters (such as thallus shape or branching pattern), were collected in different localities and dates along the Mexican Pacific and Atlantic coasts of Guerrero and Michoacan, and Veracruz and Quintana Roo, respectively, as well as three samples from Brazilian coasts (Figure 1), at a depth of 1 m. Of the total samples, 25 were used for molecular analysis (Table 1). Apical sections were preserved in silica gel for DNA extraction and molecular analysis. Samples collected for morphological analysis were preserved in 3 % formaldehyde seawater solution. Fresh specimens were mounted on herbarium sheets and incorporated into the algal collections (Table 1) at Metropolitan Herbarium UAMIZ (Thiers 2021). Specimens from the same collection were incorporated under the same UAMIZ number (Table 1).
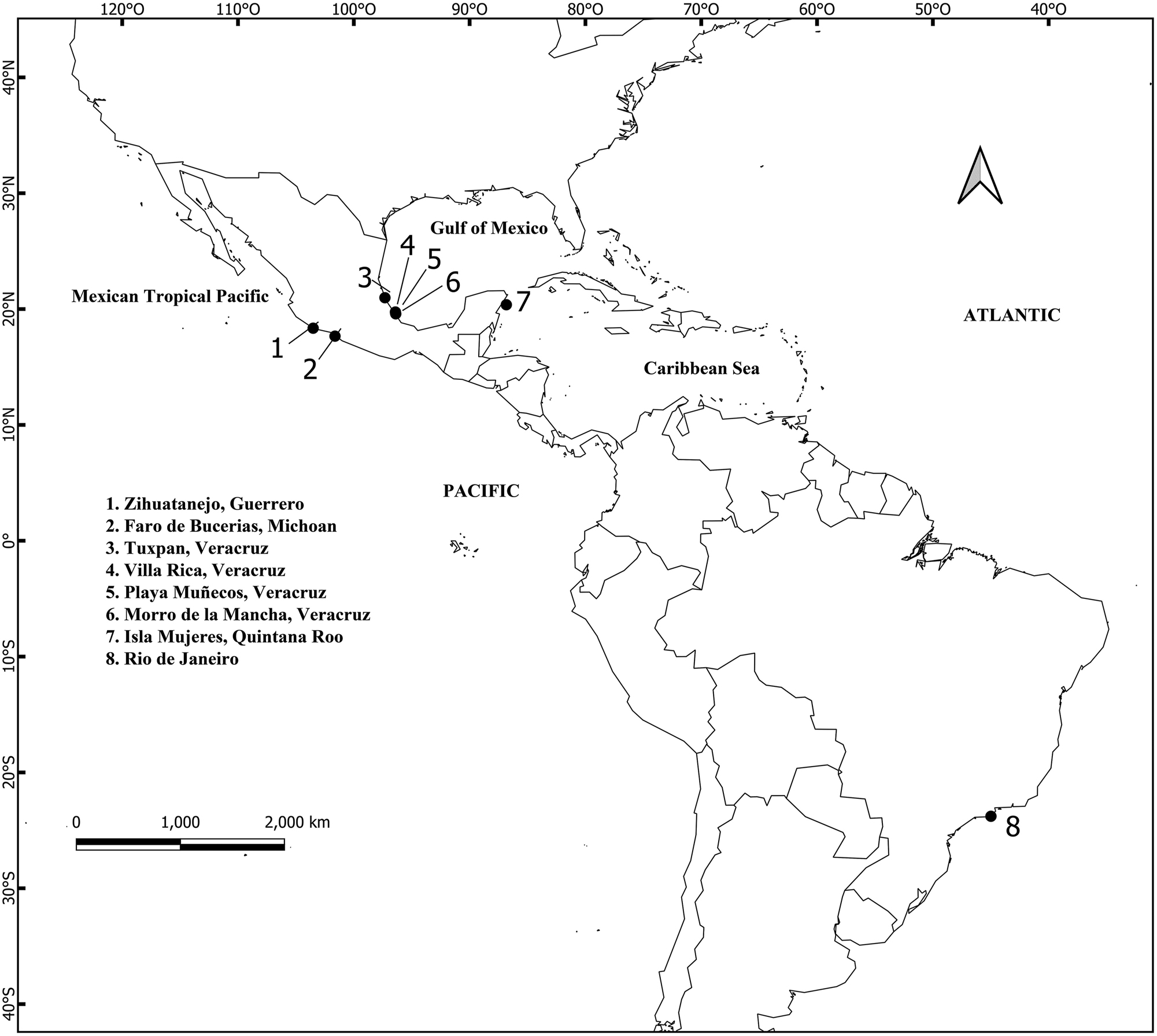
Map of the collection sites for Chnoospora in the Mexican coasts and Brazil. The numbers correspond to site names.
Collection data, voucher number (UAMIZ), field numbers and GenBank accession numbers of specimens of Chnoospora examined in the present study.
| Specimen | Site, GPS coordinates, date, and collector | Voucher no. | Field no. | GenBank accession no. | |
|---|---|---|---|---|---|
| Cox3 | rbcL | ||||
| Chnoospora minima | Villa Rica, Veracruz, Mexico, 15 April 2013, M.L Núñez Resendiz | UAMIZ-1518 | VRCh3 | PQ204662 | PP849834 |
| Playa Muñecos, Veracruz, Mexico, 19° 44.39′ N, 96° 24.24′ W, 17 March 2020, M.L. Núñez Resendiz | UAMIZ-1517 | PMCh1 | PQ204663 | PP849835 | |
| PMCh2 | – | ||||
| Morro de la Mancha, Veracruz, Mexico, 19° 35.22′ N, 96° 22.43′ W, 15 April 2013, M.L. Núñez Resendiz | UAMIZ-1519 | MMCh2 | PQ204666 |
– | |
| MMCh5 | PP849839 | ||||
| Tuxpan, Veracruz, Mexico, 20° 97.31′ N, 97° 30.58′ W, 19 April 2002, A. Sentíes | UAMIZ-1520 | TCh1 | PQ204665 |
– | |
| TCh2 | PP849838 | ||||
| Tch3 | – | ||||
| Isla Mujeres, Quintana Roo, Mexico, 21° 20.28′ N, 86° 71.14′ W, 18 February 2008, A. Sentíes | UAMIZ-1521 | IMCh1 | PQ204664 | PP849836 | |
| IMCh2 | – | ||||
| IMCh3 | – | ||||
| IMCh5 | PP849837 | ||||
| Faro de Bucerías, Michoacan, Mexico, 18° 35.13′ N, 103° 51.14′ W, 19 September 1986, Kurt M. Dreckmann | UAMIZ-23 | FB5 | PQ204667 | PP849840 | |
| UAMIZ-46 | FB12 | PP849841 | |||
| Ixtapa Zihuatanejo, Guerrero, Mexico, 17° 63.86′ N, 101° 55.38′ W, 17 April 1988, Kurt M. Dreckmann | UAMIZ-1153 | IxZ1 | – | ||
| Ixtapa Zihuatanejo, Guerrero, Mexico, 17° 63.86′ N, 101° 55.38′ W, 18 October 2024, ML Núñez Resendiz | UAMIZ-1533 | Zh7-24 | PQ568258 | PQ568254 | |
| Rio de Janeiro, Brazil, −22° 95.32′ S, −43° 15.51 E, 25 July 2024, Abel Sentíes | UAMIZ-1522 | RJB1 | PQ204668 | PQ204670 | |
| RJB2 | PQ204669 | PQ204671 | |||
| RBJ3 | – | – | |||
| C. ramosissima | Ixtapa Zihuatanejo, Guerrero, Mexico, 17° 66.46′ N, 101° 62.41′ W, 19 July 2019, A. Sentíes | UAMIZ-1515, Holotype | IxZCh3 | PQ204659 | PP849832 |
| UAMIZ-1516, Isotype | IxZCh4 | PQ204660 | PP849833 | ||
| IxZCh6 | PQ204661 | – | |||
| Ixtapa Zihuatanejo, Guerrero, Mexico, 17°63.86′N, 101°55.38′W, 18 October 2024, ML Núñez Resendiz | UAMIZ-1531 | Zh1-24 | PQ568256 | PQ568252 | |
| UAMIZ-1532 | Zh3-24 | PQ568257 | PQ568253 | ||
| Faro de Bucerías, Michoacan, Mexico, 18°35.13′N, 103°51.14′W, 19 September 1986, Kurt M. Dreckmann | UAMIZ-44 | FBM1 | PQ568259 | PQ568255 | |
DNA for molecular analysis was extracted from 5 to 10 mg of dried tissue using a Qiagen DNeasy Plant Mini Kit (Qiagen, Valencia, California USA) according to the manufacturer’s protocols. The mitochondrial cox3 region was amplified using the primers CAF4A and CAR4A (Kogame et al. 2005). The chloroplast rbcL region was amplified using the primers rbc-F0, rbc-F3 (Kawai and Sasaki 2004), Ral-R952 (Lim et al. 2007) and PBR-R3 (Kogame et al. 1999). The PCR procedure followed Lim et al. (2007). PCR products were purified with QIAquick Gel Extraction Kit (Qiagen, Valencia, California USA) and sequenced commercially (Genomic Sequencing Laboratory, Pabellón Nacional de la Biodiversidad, UNAM). The same sets of primers were used for sequencing. The sequences generated were assembled and edited using the program Sequencher® version 5.4.5. The final alignment with sequences from GenBank of other species of Scytosiphonaceae (Supplementary Table S1) was performed using Bioedit (Hall 1999). Ectocarpus siliculosus (Dillwyn) Lyngbye, Chordaria flagelliformis (O.F. Müller) C. Agardh, and Pylaiella littoralis (Linnaeus) Kjellman were used as outgroups for the cox3 data set; meanwhile Ectocarpus siliculosus and Cladosiphon okamuranus Tokida were used for the rbcL data set. The outgroup species were selected as they belong to different families within the Ectocarpales. Phylogenetic analyses using maximum likelihood (ML) and Bayesian inference (BI) were performed with codons partitioned. The evolutionary model selected was GTR + I + G (general time reversible + invariable sites + gamma distribution) determined based on the ML ratio test implemented by TOPALi version 2 software (Milne et al. 2009). ML analysis was performed using RAxML software (Stamatakis 2006) with the GTR + I + G model. Support for each branch was obtained from 1,000 bootstrap replications. BI analysis was performed using MrBayes 3.2.2 (Ronquist et al. 2012). Four chains of Markov chain Monte Carlo were used, starting with a random tree, and sampling the data every 500 generations for 6 million generations. Twenty-five percent (25 %) of trees were discarded as burn-in. Pairwise distance values (p distances) were calculated using MEGA X (Kumar et al. 2018).
Microscopic cross-sections were made by hand using a razor blade and mounted in an 80 % Karo®/distilled water solution. Photomicrographs were taken using a Quasar digital camera adapted to a Leica DMLB microscope (Heidelberg, Germany). Photographs were taken with a Nikon D7000 digital camera. Morphological measurements were obtained from micrographs using SigmaScan©Pro automated image analysis software (Jandel Scientific, Sausalito, California).
Data on the distribution of Chnoospora minima were obtained both from specialized databases such as Algaebase (Guiry and Guiry 2024) and Global Biodiversity Information Facility (GBIF), and from specialized literature (García-García et al. 2021; Norris et al. 2017; Pedroche and Sentíes 2020; Pedroche et al. 2008; Wynne 2022). With these data, a global distribution map was generated in the software QGis v.3.28.1.
3 Results
The cox3 alignment consisted of 48 sequences of 466 base pairs (bp), the rbcL alignment consisted of 48 sequences of 1,289 bp in length. In both analyses, topologies resulting from ML and BI analyses did not differ, so only the ML trees with bootstrap and BI values integrated, are shown (Figures 2 and 3).
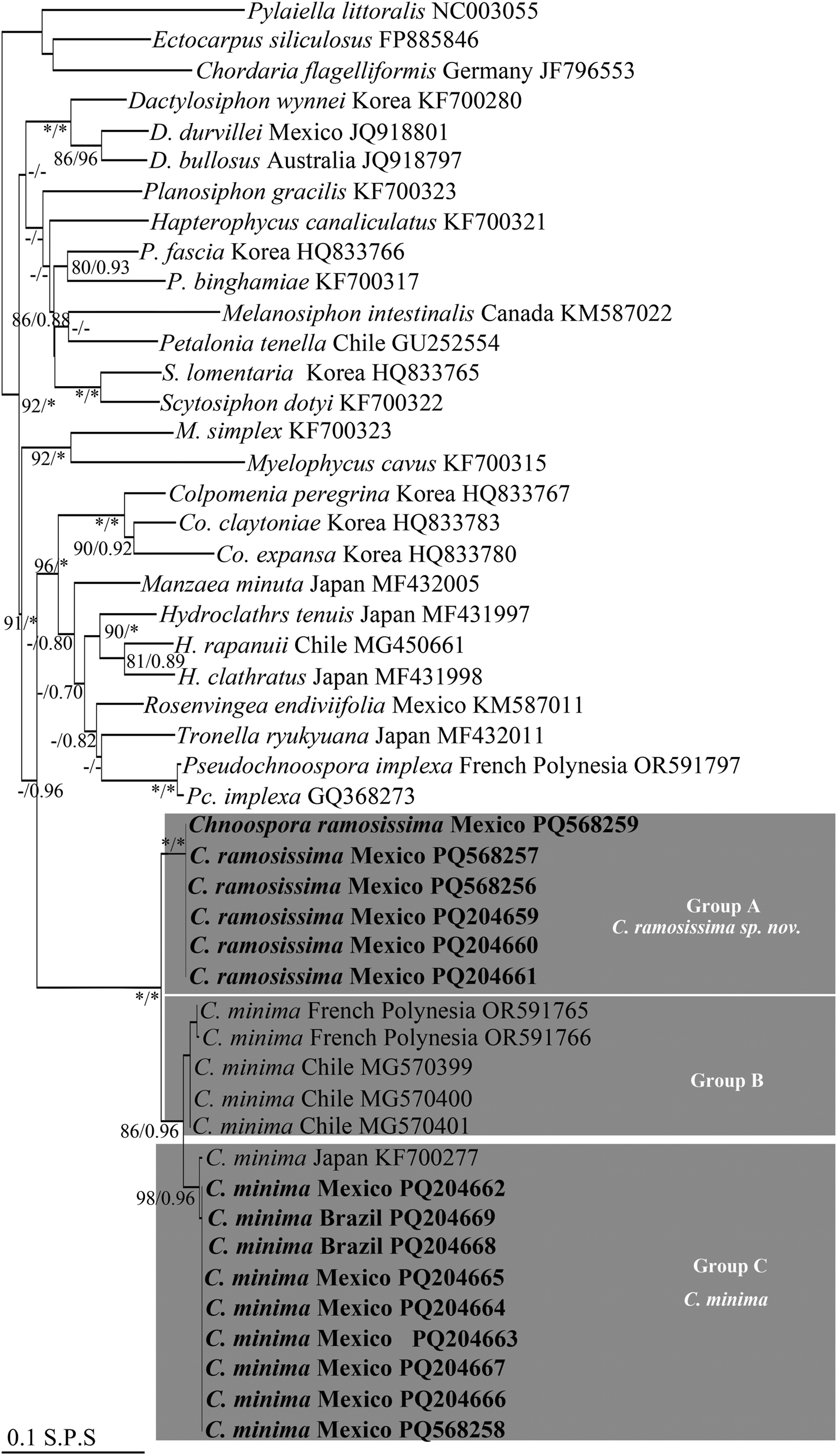
Maximum likelihood (ML) topology based on cox3 sequences data. ML bootstrap (left) followed by Bayesian inference (BI) values (right) on branches. Asterisks indicate full support (ML = 100 %, BI = 1.0 %), hyphens indicate values below 70 %. Group A, Group B, and Group C indicate the three clusters within Chnoospora, presented as C. minima and C. ramosissima sp. nov. Sequences generated in this study are in boldface. S.P.S. = substitutions per site.
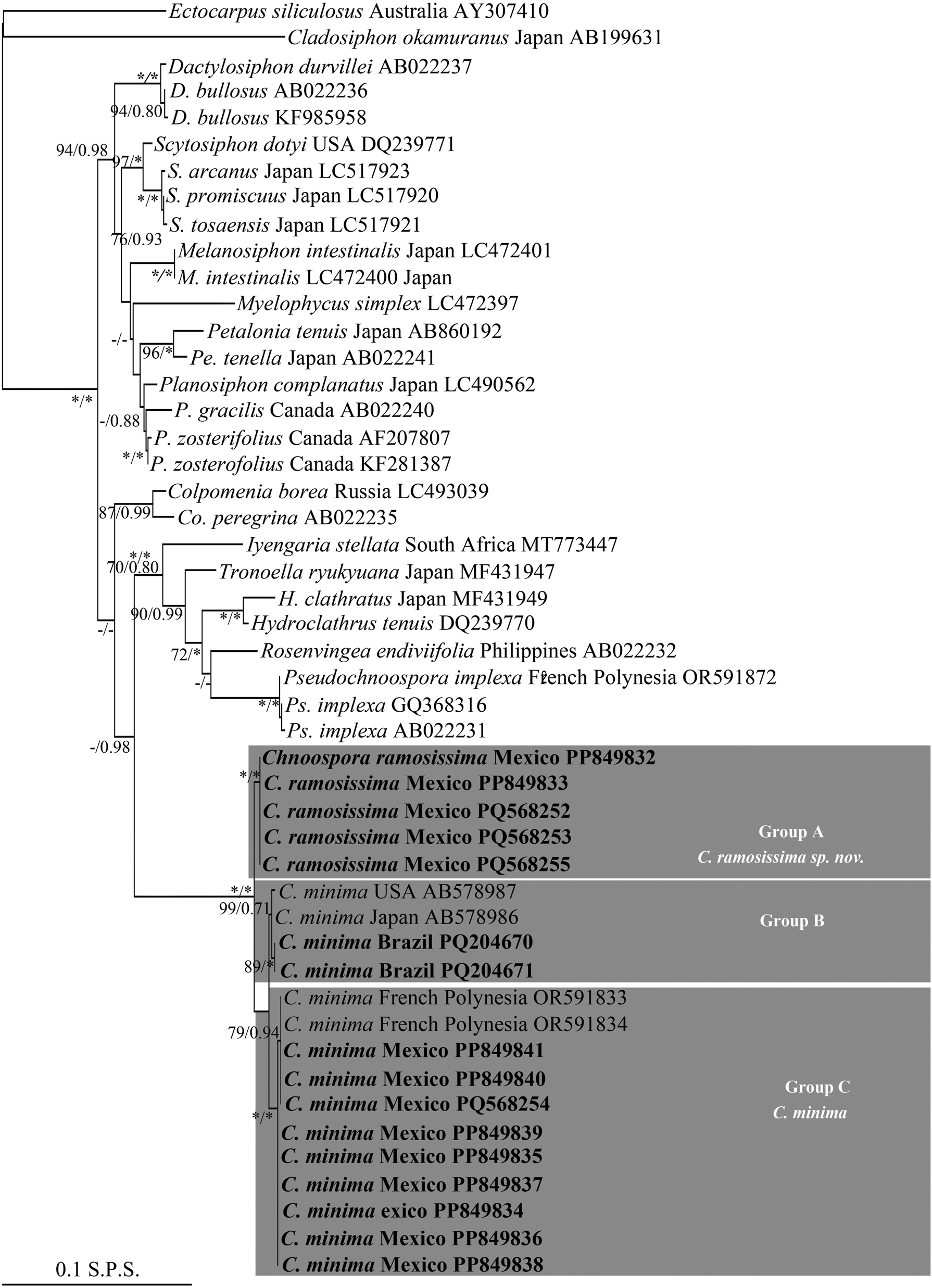
Maximum likelihood (ML) topology based on rbcL sequences data. ML bootstrap (left) followed by Bayesian inference (BI) values (right) on branches. Asterisks indicate full support (ML = 100 %, BI = 1.0 %), hyphens indicate values below 70 %. Group A, Group B, and Group C indicate the three clusters within Chnoospora, presented as C. minima and C. ramosissima sp. nov. Sequences generated in this study are in boldface. S.P.S. = substitutions per site.
With cox3, sequences generated in this study and those of Chnoospora minima from GenBank formed a monophyletic group within Scytosiphonaceae, with the maximum phylogenetic support (Figure 2). In turn, this group was resolved as the sister clade of the genera Colpomenia (Endlicher) Derbès et Solier, Hydroclathrus Bory, Manzaea Santiañez et Kogame, Rosenvingea Børgesen, Tronella Santiañez et Kogame, and Pseudochnoospora Santiañez, G.Y.Cho et Kogame, with low support (ML = 59 %; BI = 0.96) (Figure 2). Within Chnoospora, all the sequences formed three sister subclades (Group A, Group B, and Group C). Group A, with the maximum phylogenetic support, clustered five sequences from Zihuatanjeo, Guerrero and one from Faro de Bucerias, Michoacan, Mexico (Figure 2, Table 1), and was resolved in turn as the sister subclade of Group B and Group C, both more closely related to each other (Figure 2). Group B, with good support (ML = 88 %; BI = 0.92), clustered two sequences from French Polynesia and three from Chile; Group C, also with good support (ML = 98 %; BI = 0.96), clustered four sequences from Veracruz, one from Quintana Roo, Michoacan, and Zihuatanejo, respectively, two from Brazil, and one from Japan (Figure 2, Table 1). With rbcL, these sequences also formed a monophyletic group within Scytosiphonaceae, where the Chnoospora clade was resolved, in turn, as the sister group of the genera Iyengaria Børgesen, Tronoella, Rosenvingea, Hydroclathrus, and Pseudochnoospora, with low support (ML = 50 %; BI = 0.98) (Figure 3). Similarly, sequences of C. minima also formed a major clade, with the maximum phylogenetic support, resolved in turn into three sister subclades (Group A, Group B, and Group C). Group A, with the maximum phylogenetic support, was resolved, in turn, as the sister group of the subclades Group B and Group C, both more closely related between them, and clustered four sequences from Zihuatanejo, Guerrero and one Faro de Bucerias, Michoacan, Mexico; Group B, well supported (ML = 89 %; BI = 1.0), clustered two sequences from Brazil, one from Japan, and one from USA; Group C, with the maximum support, clustered two sequences from Michoacan, two from Quintana Roo, four from Veracruz, and one from Zihuatanejo, Guerrero, with two from French Polynesia (Figure 3, Table 1).
With cox3, the genetic distance values within groups (Table S2) were: Group A: no variation; Group B: ranged from 0.2 to 0.6 %; and Group C: 0.4 % (with a sequence from Japan). The genetic distance values between groups were: Group A and B: ranged from 4.1 % (with sequences from Chile) to 4.3 % (with sequences from French Polynesia); Group A and C: 5.1 %, and Group B and C: ranged from 1.7 (with sequences from Chile) to 2.4 % (with sequences from French Polynesia). With rbcL, the genetic distances values within groups (Table S3) were: Group A: no variation; Group B: ranged from 0.1 % (with sequences from Brazil and Japan) to 0.2 % (with sequences from Brazil and USA); Group C: 0.1 %. The genetic distance values between groups were: Group A and B: ranged from 0.8 % (with a sequence from Japan) to 0.9 % (with sequences from Brazil and USA); Group A and C: ranged from 0.9 % (with sequences from Mexico) to 1 % (with sequences from French Polynesia); Group B and C: ranged from 0.4 % (with sequences from USA and Japan) to 0.5 (with sequences from Brazil).
These genetic distance values between groups support the presence of, at least, two phylogenetic independent lineages within Chnoospora (Figure 2 and 3). Considering the morphological characters, and distribution patterns (Figure 4), as well as the type locality for C. minima in the Indian Ocean, we described Group B and Group C as the true C. minima and Group A as a new lineage that we have named C. ramosissima sp. nov. The close relationship between Group B and Group C is discussed below.

Chnoospora minima. (4) UAMIZ-1519. Morphotype from Morro de la Mancha, Veracruz, showing intermediate branching segments longer than the rest (arrow). Scale bar = 1 cm. (5) UAMIZ-1518. Morphotype from Villa Rica, Veracruz, showing intermediate branching segments longer than the rest (arrows). Scale bar = 1 cm. (6) UAMIZ-1517. Morphotype from Playa Muñecos, Veracruz, showing intermediate branching segments longer than the rest (arrows) and curved angles (arrowheads). Scale bar = 1 cm. (7) UAMIZ-1521. Morphotype from Isla Mujeres, Quintana Roo, showing intermediate branching segments longer than the rest (arrow) and acute angles (arrowhead). Scale bar = 1 cm. (8) UAMIZ-46. Morphotype from Faro de Bucerías, Michoacan, showing intermediate branching segments longer than the rest (arrow). Scale bar = 1 cm. (9) UAMIZ-1522 showing intermediate branching segments longer than the rest (arrows) and branching angles (arrowheads). Scale bar = 7 mm.
3.1 Chnoospora minima (Hering) Papenfuss (Figures 4–15)
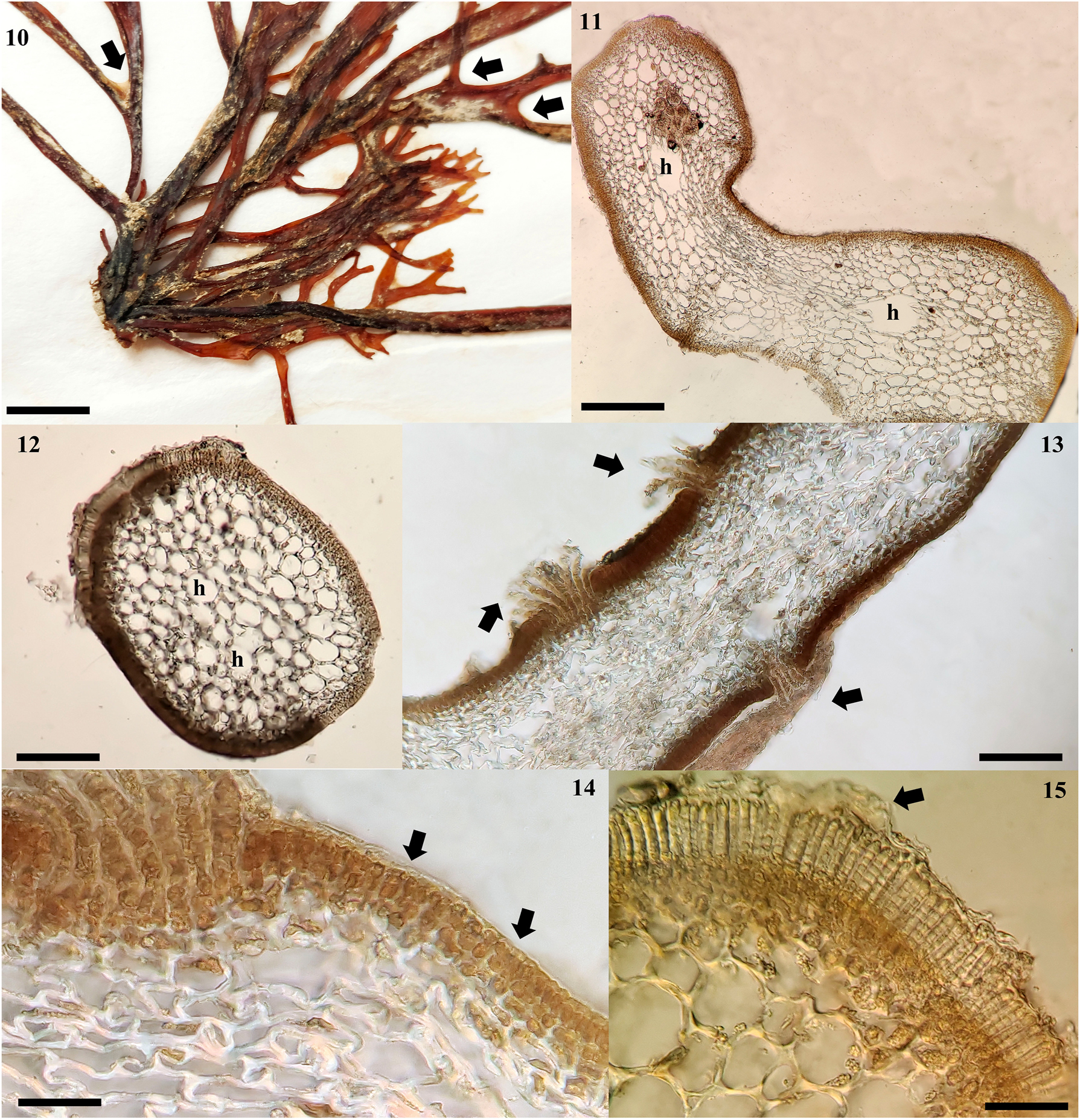
Chnoospora minima. (10) UAMIZ-1517. Specimen from Villa Rica, Veracruz showing a consolidated holdfast. Scale bar = 4 mm. (11) UAMIZ-1518. Cross section through middle part of an erect axis with compressed form showing medullary cells and hollows (h) between them. Scale bar = 150 μm. (12) UAMIZ-23. Cross section through middle part of an erect axis with cylindrical form showing medullary cells with hollows (h) between them. Scale bar = 150 μm. (13) UAMIZ-1517. Cross section of an erect axis showing cortical cryptostomata with numerous long hairs (arrows). Scale bar = 150 μm. (14) UAMIZ-1520. Cross section of an erect axis showing three layers of cortical cells (arrows). Scale bar = 40 μm. (15) UAMIZ-46. Cross section showing uniseriate plurangia covered with a loosened cuticle (arrow). Scale bar = 70 µm.
Type locality: Port Natal (Durban), South Africa (Papenfuss 1956).
Observations: Thalli forming bushy tufts, golden brown in color (Figures 4–9), attached to the substratum by a holdfast, associated or not with the prostrate, crustose phase of its biological cycle (Figure 10). Erect axes, monopodial, terete to compressed (Figures 11 and 12), 3.5–8 (−12) cm in length and variable diameter from the base (400–500 × 1,200–1,300 µm) at the apical portions (850–900 × 1,100−1,300 µm) and median portions (200–250 × 1,300−1,400 µm); branching dichotomously or trichotomous (Figures 4−9), 2–5 times on each axis (Figures 4−9), with acute or blunt apices; branch angle predominantly greater than 50°, curved (Figures 4−9). Segments between branches of variable diameter along the axes, broader at the base of each dichotomy (Figure 4), with variable length, being shorter from the stipe (0.5–0.8 cm) towards the intermediate portions, where the segments elongate notably (2–3.5 cm), reducing their size again towards the apical portions (0.5–1 cm) (Figures 4–9). Cryptostomata on axes and branches, numerous, superficial, from which elongated microscopic hairs arise (Figure 13). In cross section, cortex made up of three layers of pigmented, elongated, rectangular cortical cells, relatively constant in diameter between the apical (2.5 × 5 µm) and middle portions (2.5–3.5 × 5 µm) and larger in the basal portion (3.5–5 × 7.5−8.75 µm) of the axes (Figure 14); medulla made up of 12–15 layers of hemispherical, elongated, not very dense cells, with variable diameter in different sections of each axis: apical portion (7.5) 30–35 × (5) 12.5−20 µm, middle portion (15) 50−70 µm, and basal portion (15) 25–40 × (25) 60−75 µm, reducing progressively in size towards the cortex (Figures 11 and 12), giving the thallus the appearance of being elongated and little branched. Large diameter hollows are observed between the medullary cells in median and basal portions (Figure 11). Sporangia plurilocular elongate, growing uniseriate or biseriate on cortical sori, covered with a loosened cuticle (Figure 15), 60–70 µm in length; mature plurangia formed 12−14 locules, quadrangular cells, 3.5 × 5 µm in diameter.
Distribution: Tropical and subtropical coasts of the world. Atlantic Ocean: Ascencion Island (John et al. 2004), Bahamas (GBIF 2024), Benin (John et al. 2004), Brazil (Taylor 1960), Cape Verde (John et al. 2004), Colombia (Schnetter 1976), Cuba (Suárez et al. 2023), Equatorial Guinea (John et al. 2004), Ghana (John et al. 2004), Ivory Coast (John et al. 2004), Lesser Antilles (Taylor 1960), Liberia (John et al. 2004), Martinique (Delnatte and Wynne 2016), Mexico (García-García et al. 2021), Netherlands Antilles (Taylor 1960), Nigeria (John et al. 2004), Panama (Taylor 1960), Puerto Rico (Ballantine et al. 2021), São Tomé and Príncipe (John et al. 2004), Sierra Leone (John et al. 2004), Togo (John et al. 2004), Trinidad and Tobago (Duncan and Lee Lum 2006) and Venezuela (Taylor 1960). Indian Ocean: Christmas Island (Silva et al. 1996), India (Silva et al. 1996, Yadav et al. 2023), Indonesia (Silva et al. 1996), Mauritius (Silva et al. 1996), Mozambique (Silva et al. 1996), Malaysia (GBIF 2024), Myanmar (Soe-Htun et al. 2021), Red Sea (Einav et al. 2021), Réunion (Silva et al. 1996), Seychelles (Silva et al. 1996), South Africa (Silva et al. 1996) and Sri Lanka (Silva et al. 1996). Pacific Ocean: American Samoa (Littler and Littler 2003), Australia (Kraft 2009), Central Polynesia (Tsuda and Walsh 2013), China (Liu 2008), Colombia (GBIF 2024), Easter Island (Santiañez et al. 2018b), El Salvador (GBIF 2024), Federated States of Micronesia (Lobban and Tsuda 2003), Fiji (South and Skelton 2003), French Polynesia (N’Yeurt and Payri 2006), Guam (Lobban and Tsuda 2003), Hawaiian Islands (Huisman et al. 2007, Sherwood and Guiry 2023), Japan (Yoshida et al. 2015), Mariana Islands (Tsuda 2003), Mexico (Pedroche et al. 2008, Pedroche and Sentíes 2020), New Zealand (Nelson et al. 2021), Papua New Guinea (GBIF 2024), Philippines (Ang et al. 2014), Revillagigedo Islands (Serviere-Zaragoza et al. 2007), Samoan Archipelago (Skelton and South 2007), Solomon Islands (Womersley and Bailey 1970), Taiwan (Lewis and Norris 1987), Thailand (Coppejans et al. 2011), Tuamotu Islands (Taylor 1973) and Viet Nam (Nguyen et al. 2013) (Figure 16).
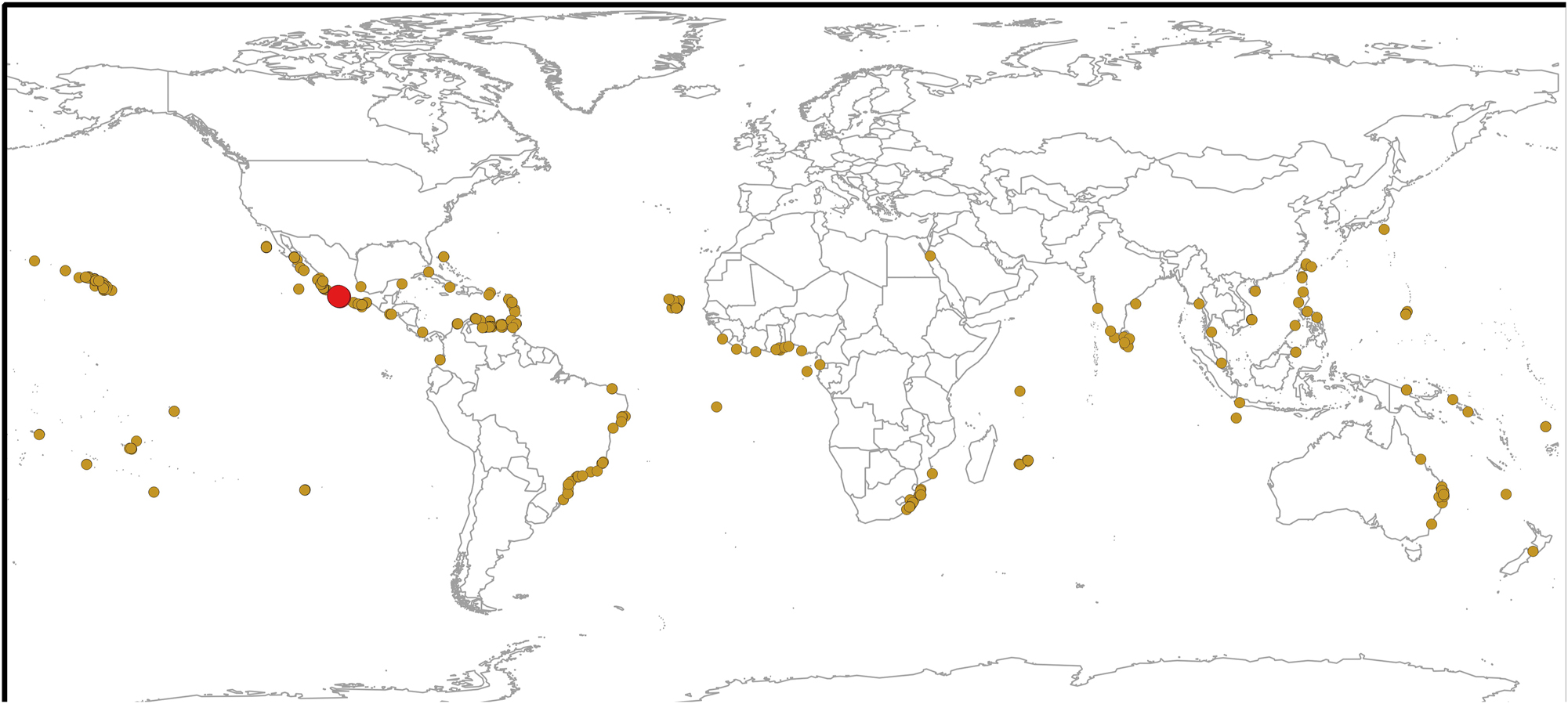
Map of the global distribution of Chnoospora minima (brown circles) and C. ramosissima (red circle).
Habitat: Plants growing in the intertidal zone, on rock walls or rocky platforms, exposed directly to the waves (Littler and Littler 2003; this study).
3.2 Chnoospora ramosissima Núñez-Resendiz, Dreckmann et Sentíes sp. nov. (Figures 17−24)

Chnoospora ramosissima sp. nov. (17) Holotype specimen. UAMIZ-1515. Thallus showing abundant branching with predominantly acute branching angles (arrows). Scale bar = 1 cm. (18) UAMIZ-44. Thalli of Chnoospora ramosissima from Faro de Bucerias, Michoacan, showing one clustered thallus from a single holdfast (arrowhead), and abundant branching with predominantly acute branching angles (arrows). Scale bar = 1 cm. (19) UAMIZ-1515. Cross section through middle part of an erect axis showing medullary cells with hollows (h) between them. Scale bar = 150 μm. (20) UAMIZ-1516. Cross section of an erect axis showing cortical cryptostomata with short hairs (arrow). Scale bar = 50 μm. (21) UAMIZ-1516. Cross section showing cortical layers (arrow). Scale bar = 40 μm. (22) UAMIZ-1515. Cross section showing uniseriate (arrowheads) and biseriate plurangia (arrows). Scale bar = 65 μm. (23) UAMIZ-1516. Cross section showing the development of the plurangia from the cortical cells (arrow), at different stages of maturation. Scale bar = 65 μm. (24) UAMIZ-1515. Detail of the development of biseriate plurangia from cortical cells (arrows). Scale bar = 30 µm.
Description: Thalli forming bushy tufts, yellow to dark, wiry, fixed by a holdfast; erect axes compressed, 6–8 (−10) cm length and 1,000−1,100 µm in diameter, branching profusely, 6–8 (to 12) times, dichotomously, with length progressively reducing from the base (2.6–3 cm) to the tips (0.3−0.5 cm) where the branching is most profuse, acute angle; surfaces with numerous cryptostomata associated with short-microscopic hairs; cortex of 2–4 layers of golden and rectangular cells, diameter variable in apical portions (3.5–45 × 7.5–10 µm), middle portions (5 × 7.5 µm), and basal portions (5 × 7.5–8.5 µm); medulla of 15–18 layers of rounded, large, or irregular cells, compact, diameter slightly variable: 35−50 (−80) µm in apical portions, (25) 40–50 µm × (20) 50–65 µm in middle portions, and (15) 25–40 × 30−50 (−65) µm in basal portions; hollows scattered among the medullary cells, not clearly visible in the basal portions; uniseriate or biseriate sporangia plurilocular without a cuticle; mature plurangia 50–80 µm long, subdivided into 10–16 squarish locules, 5 µm in diameter.
Type locality: Zihuatanejo, Guerrero, Mexico (17° 66.46′ N, 101° 62.41′ W; Figure 1).
Holotype: UAMIZ-1515 (Figure 12), Zihuatanejo, Guerrero, Mexico, A. Sentíes; deposited in UAMIZ.
Isotype: UAMIZ-1516, Zihuatanejo, Guerrero, Mexico, A. Sentíes; deposited in UAMIZ.
Etymology: The specific epithet (ramosissima = with many branches) refers to the presence of numerous branches in the thallus.
Observations: Thalli were forming bushy tufts, yellow brown in color when fresh to dark when dried (Figures 17 and 18), cartilaginous, wiry, non-lubricated, fixed to the substratum by a holdfast, from which an erect monopodial axis arose (Figures 17 and 18). Erect axes were compressed (Figure 19), 6–8 (−10) cm length with constant diameter in the apical (300–350 × 1,000−1,100 µm) and middle (400 × 1,000−1,100 µm) portions of the axes and notably different and wider in the basal portion (1,100 × 1,800 µm), branching profusely, 6–8 (to 12) times, dichotomously, with acute tips; branching angle was predominantly acute and straight, between 45° and less than 90°, slightly curved in older portions (Figure 18). Branching segment diameter was relatively constant along the axes, slightly broader at each dichotomy (Figure 18); the length was progressively reduced from the base to the tips to 2.6–3 cm in the basal segment, 0.8–1 cm in the intermediate segments and 0.3–0.5 cm in the apical portions, giving the thallus the appearance of being profusely branched. Surfaces were smooth, with numerous cryptostomata in axes and branches, associated with short-microscopic hairs (Figure 20).
In cross section, the cortex was made up of three layers of golden cells, rectangular, with different diameters between the apical portion (3.5–5 × 7.55–10 µm), the middle portion (5 × 7.5 µm), and the basal portion (5 × 7.5−8.5 µm; Figure 21); medulla was made up of 15–18 layers of rounded, large, or irregular cells, with thick walls, compact, also with different diameters in the apical portion [10–35 × 35–50 (−80) µm], the middle portion [(25) 40–50 × (20) 50−65 µm], and the basal portion [(15) 25–40 × 30−50 (−65) µm], with constant size from center to the cortex (Figure 14); as has been described for all the species in Scytosiphonaceae, whose species can be hollow or partially hollow, some hollows appear scattered among the medullary cells, some hollows appear scattered among the medullary cells, without an apparent pattern, not clearly seen in basal sections (Figure 19).
Sporangia plurilocular growing in uniseriate or biseriate cortical sori without a loosened cuticle (Figures 22−24); mature plurangia were elongated, 50–80 µm long, subdivided into 10–16 squarish locules, 5 µm in diameter.
Habitat : Plants growing on rock walls or rocky platforms, exposed directly to the waves, in the intertidal zone, between 0.5 and 1 m deep, exposed to direct light during low tide. Present throughout the year although they are more abundant in the months of April to September; growing in close association with species of Ectocarpus Lyngbye and Chaetomorpha Kützing.
Distribution: In the Mexican coasts from Zihuatanejo, Guerrero and Faro de Bucerias, Michoacan.
4 Discussion
From our molecular and morphological results, we can establish that, within the Mexican populations of C. minima, there are two phylogenetically independent entities, C. minima being the most common, with a wide disjunct distribution, and C. ramosissima sp. nov., with sympatric distribution with C. minima in its type locality in Zihuatanejo, Guerrero and Faro de Bucerias, Michoacan, Mexico.
The resulting phylogenies with the cox3 and rbcL markers showed that, within the Chnoospora clade, there are three genetically differentiated sister groups, Group A, basal to Groups B and C, which in turn, are more closely related to each other. In the phylogenies of Santiañez et al. (2018b) and Vieira et al. (2024) with the cox3 marker, the formation of at least two genetically independent groups within C. minima is also observed. In Santiañez et al. (2018b), the sequences of C. minima are clustered into two groups, the first made up of sequences from Chile generated in that study, and the second group made up of a sequence from Japan. Similarly, in Vieira et al. (2024), two groups are also formed, one made up of the sequences from Chile (from Santiañez et al. 2018a, 2018b) along with the sequences generated in their study, and the second group also with the sequence from Japan. In our analysis with cox3, we also have the formation of these same groups in Group B and Group C, in the latter in addition, our sequences from the Mexican Atlantic coasts, Brazil and in the Mexican Pacific, Guerrero and Michoacan were grouped, while Group A only contained sequences from Zihuatanejo in Guerrero and Faro de Bucerias, Michoacan (Figure 2). With the analysis based on rbcL, we observed the same three genetic groups, Group A, which remained independent by grouping the same Zihuatanejo and Faro de Bucerias sequences as in cox3, and Groups B and C that did vary the relationships shown previously (Figure 3). In group B, the sequences from USA and Japan were grouped with those from Brazil generated in the present study, while Group C contained sequences from French Polynesia and the rest of the localities in Mexico. This mixture between the sequences that make up Groups B and C observed between cox3 and rbcL suggests that, although there is genetic differentiation within C. minima, it must correspond to a relatively recent event, since the rbcL marker, which is more conserved than cox3, does not reflect these phylogenetic relationships.
Additionally, the interspecific genetic distance values calculated with cox3 between Groups A and B (4.1−4.3 %) and Groups A and C (5.1 %) coincided with those reported by Boo et al. (2011) and Lee et al. (2014b) between Colpomenia (Endlicher) Derbès et Solier species (4.4−7.69 %; Co. peregrina Sauvageau and Co. claytoniae S.M. Boo, K.M. Lee, G.Y. Cho et W. Nelson) and those reported by Santiañez et al. (2018a) between Hydroclathrus Bory species [4.1−7.4 %; H. clathratus (C. Agardh) M. Howe and H. tenuis C.K. Tseng et Lu Baroen], providing evidence of the taxonomic independence between Group A and Groups B and C. On the other hand, the intraspecific genetic distance values between Groups B and C (1.7−2.4 %) were also consistent with those reported by Lee et al. (2014b) for species of Colpomenia (0−3.7 %; C. peregrina) and Santiañez et al. (2018a) for the genus Hydroclathrus [0−2.6 %; H. tilesii (Endlicher) Santiañez et M.J. Wynne], providing evidence that both groups do not constitute independent lineages from each other, but rather are the same lineage. Regarding the genetic distances calculated with rbcL, the values between Groups A and B (0.8−0.9 %) and A and C (0.9−1 %), were also consistent with the interspecific distance values provided by Kogame et al. (1999) (0.4−0.5 %, for species of Scytosiphon C. Agardh), also supporting the independence between Group A with Groups B and C. On the other hand, the distance calculated with this marker between Groups B and C (0.4−0.5 %), could support their independence according to the values reported among other species of Scytosiphonaceae; however, we previously discussed evidence that rules out this possibility, mainly due to the existing mixture between the sequences that make up these groups between both phylogenies, which suggests that there is no solid differentiation between them.
Given the phylogenetic relationships described in each group and considering the genetic distance values proposed to establish this difference, we recognize only two phylogenetically independent lineages within C. minima, the first corresponding to Group A and the second with Group B/C, which, based on their morphological characteristics and distribution patterns discussed below, we have designated as C. ramosissima sp. nov. (Group A) and the true C. minima (Group B/C). Unfortunately, we do not have sequences of C. bicanaliculata, the third species in the genus, to compare with our results and establish its phylogenetic relationships with the other two, but this must be a future work.
Regarding its distribution, Chnoospora minima has the widest distribution range, since it has been recorded from the coasts of the eastern and western Pacific, Indian Ocean to the Atlantic coasts (Guiry and Guiry 2024). Meanwhile, C. bicanaliculata so far has a distribution restricted to India and surrounding regions (Krishnamurthy and Baluswami 2010; Rao and Gupta 2015; Yadav et al. 2023). Until now, the distribution of C. minima had only been confirmed molecularly from Japan, Hawaii, French Polynesia, and Chile (Santiañez et al. 2018b; Vieira et al. 2024). Particularly, on the tropical Mexican coasts it presents a disjunct distribution pattern (García-García et al. 2021; Norris et al. 2017; Pedroche and Sentíes 2020), while, so far, C. ramosissima is restricted to the Zihuatanejo and Faro de Bucerias, in the tropical Pacific Mexican, where it co-occurs with C. minima. From an analysis of global records and considering its current distribution pattern, we can determine that C. minima could have had an Atlantic origin. In the distribution map (Figure 16), it is observed that most of the records are concentrated in the Western Atlantic portion, being scarce in the African region and the Mediterranean, which makes it possible to infer that the original population came from the southern portion of the western Atlantic coast, where the prevailing current pattern, from south to north, allowed its establishment in the northwest Atlantic region. Likewise, prior to the closure of the Isthmus of Panama, approximately 3 m.y.a (Coates and Obando 1996; Coates and Stallard 2013), populations were able to cross and establish themselves in the eastern and western Pacific, and from there, later, establish themselves in the Indo-Pacific, which is also suggested by the few records in the area. However, detailed studies of population genetics and phylogeography must be carried out, in order to provide evidence on the geological and biogeographic history, not only of C. minima but of the rest of the species in the genus.
Morphologically, C. bicanaliculata, C. minima, and C. ramosissima present the typical morphology of the genus, apparently little variable, consisting of cylindrical to complanate axes and branches and the characteristic branching pattern in one or two planes (Santiañez et al. 2018b). However, to discriminate them, the characters consistently proposed by Santiañez et al. (2018b), Santiañez and Wynne (2024) or Vieira et al. (2024), are the shape of cortical and medullary cells, number of layers, size, and those characters associated with the plurilocular sporangia of the macroscopic thallus (Table 2). From our observations, specimens of C. minima present a predominantly curved angle, mainly towards the apical portions (Figures 4−7), while C. ramossisima presents a predominantly acute angle, slightly curved towards the most mature portions but predominantly straight (Figure 18), similar to that described for C. bicanaliculata (Yadav et al. 2023). However, C. bicanaliculata is a morphologically distinct species in the genus due to the presence of two central grooves or constrictions on the main axes, which form two separate canals clearly distinguishable in cross section (Yadav et al. 2023).
Morphological differences between Chnoospora ramosissima, C. bicanaliculata, and C. minima, including the comparison with specimens of C. minima from different regions where the species has been described.
| Species | C. ramosissima sp. nov. | C. bicanaliculata | C. minima | C. minima | C. minima | C. minima |
|---|---|---|---|---|---|---|
| Thallus: shape; height (cm); diameter (µm) | Compressed; 6–8 (−10); AP: 300–350 × 1,000–1,100, MP: 400 × 1,000–1,100, BP: 1,100–1,800 | Cylindrical to slightly flattened; 7–20; 500–2,000 | Terete to compressed | Subterete to compressed | Cylindrical to flattened; 4–8; 500–2,000 | Terete to compressed; 3.5–8 (−12); AP: 850–900 × 1,100–1,300, MP: 200–250 × 1,400, BP: 400–500 × 1,300–1,400 |
| Branching: pattern; times in the axes | Dichotomously; profusely, six to eight (to 12) | Irregular to subdichotomously | Dichotomously | (Sub-) dichotomously; up to seven | Regularly dichotomously | Dichotomously; two to five on each axis |
| Branching segments: length (cm) | Progressively reducing from the base to the tips, base (2.6–3), intermediate portions (0.8–1), apical portions (0.3–0.5) | Median dichotomy with two distinct grooves on both edges and slightly curved in mature stage | – | – | Apical dichotomy up to 4 mm, usually unequal | Variable length, shorter from the stipe (0.5–0.8), intermediate portions (2–3.5), apical portions (0.5–1) |
| Medullary cells: layers; shape; diameter (µm) | 15–18; rounded, large, or irregular; AP: 10–35 × 35–50 (−80), MP: (20) 50–65, BP: (15) 25–40 × 30–50 (−65); compact | 6–8; elongate to polygonal; 24.5–160 × 19–32 | Irregular, large, circular to ovoid | Rounded larger and narrowly elliptical; 150 × 100 | 6–8; elongate, spherical; 21–85 × 17–28, compact | 6–8; elongate, spherical; AP: 10–35 × 35–50(−80), MP: (20)50–65 × (25) 40–50, BP: (15) 25–40 × 30–50 (−65); not very dense |
| Cortical cells: layers; shape; diameter (µm) | 2−4; rectangular; AP: 3.5–5 × 7.5–10, MP: 5 × 7.5, BP: 5 × 7.5–8.5 | 4–7; spherical; 28–35 | 1–2; cuboidal to rounded | 2–4; oblong to slightly domed; 11–26 | 5–9; spherical to polygonal; 12-30 | 3; elongated to rectangular; AP: 3.5–5 × 7.5–10, MP: 5 × 7.5, BP: 5 × 7.5–8.5 |
| Mature plurangia: length (µm) | Uniseriate or biseriate without a cuticle; 50–80 | Uni- to biseriate sori | – | Biseriate, covered with a loosened cuticle; up to 50 | Uni- to biseriate | Uniseriate or biseriate, covered with a loosened cuticle; 60–70 |
| Locules: number; shape; diameter (µm) | 10–16; squarish; 4.5–5 × 5 | – | – | Squarish to rectangular; 4–5 × 2–5 | Multicellular | 8–12; quadrangular; 5–5.5 |
| Geographic region | Mexico: Guerrero | India | New Zealand | Easter Island (Hawai) | India | Mexico: Pacific, Gulf of Mexico and Caribbean Sea |
| References | This study | Yadav et al. (2023) | Nelson and Duffy (1991) | Santiañez et al. (2018a, 2018b) | Yadav et al. (2023) | This study |
-
AP, apical portions; MP, median portions; BP, basal portions.
The main morphological difference between C. ramosissima with C. minima and C. bicanaliculata is the length of the branching segments and the stipe. C. ramosissima has a stipe longer (2.6–3 cm) than the branching segments (0.3−1 cm) and these are of relatively constant size, progressively reducing to the apical portions (Figure 18), giving the thallus the appearance of being densely branched. In contrast, in both C. minima and C. bicanaliculata, the stipes are shorter (0.5–0.8 cm) than the branching segments along the thallus and, in both cases, a segment considerably longer (2–3.5 cm) than the rest is seen in the central portion of the thallus, reducing its size again towards the apical portions, which gives the thallus its elongated and poorly branched appearance (Figures 4−9). In the photographs of C. minima habits published by Nelson and Duffy (1991), Santiañez et al. (2018b), Taylor (1942), and Yadav et al. (2023), it is also possible to observe this central segment of greater length than the rest in the thallus. Other important characters in the delimitation between the three species are summarized in Table 2, however, the most evident are, at an anatomical level, both the number of cellular layers that make up the medulla and its diameter both in the basal, middle and apical portions of the thallus, as well as among those reported by the different authors, although in these works the level of the thallus at which these characters were observed is not specified. In C. ramosissima, it is possible to observe a dense medulla, composed of 16–18 layers of cells with a constant diameter and considerably smaller than that observed in both, C. bicanaliculata and C. minima, with lax medullae, evident intermedullary hollows and diameters almost double those of C. ramosissima, similar to those shown by Santiañez et al. (2018b) and Yadav et al. (2023). In this regard, we observed that C. ramosissima does not present obvious holes in the basal portions as it does in the middle and apical portions, which also differs with C. minima, where the holes are evident in any region of the thallus, in agreement with Santiañez et al. (2018b). Although the reproductive characters, such as the length and number of locules of the plurangia, that we observed between C. minima and C. ramosissima (Table 2) differ slightly (C. minima have shorter plurangia than those found in C. ramosissima), Santiañez et al. (2018b) report for the first time in C. minima plurangia covered by a loosened cuticle. This feature was confirmed for our specimens of C. minima (Figure 15) but was not observed in C. ramosissima (Figures 22−24).
According to our taxonomic review of Chnoopsora, the proposed synonyms for C. minima of the different related names such as C. atlantica, C. fastigiata, and C. pacifica, were based on the morphological review of these specimens and the coincidence of their characteristics with C. minima, which, despite having a very simple original description without an illustration, was assigned to the type in Durban, South Africa, after a thorough review. From the morphology of the topotype of C. minima (Anderson et al. 2016), it is possible to observe that its characteristics correspond with those that we have identified for the Mexican specimens in the Atlantic and the Pacific, that is, a thallus with a little branched appearance due to the presence of central branching axes that are longer than in the basal and apical portions. However, the figure presented by Taylor (1942: Figure 2) for C. pacifica, is very similar to our specimens of C. ramosissima, but this specimen was collected in Aruba Island, close to South America in the Caribbean region, not to San Agustín or nearby in the Mexican Pacific, therefore, although we have found two phylogenetically differentiated entities within C. minima on the Mexican Pacific coasts, molecular determination of specimens from Oaxaca and Aruba Island specimens are required to resolved its relationship with C. ramosissima or with other species that are currently part of the list of synonyms for C. minima. Given the above, at the moment we have named the new species C. ramosissima, by its profusely branching pattern.
Despite the phenotypic plasticity exhibited by populations of C. minima on the Mexican coasts, with five morphotypes differentiated in the present study (Figures 4−9), the characters described here to discriminate from C. ramosissima were constant among all of them. Among our specimens, the C. minima morphotype most similar externally to C. ramosissima was that from the Caribbean Sea (Figure 7), with predominantly acute angles and branching axes of constant diameter, however, this morphotype also presented a branching segment longer intermediate, like the rest of the morphotypes presented here for C. minima (arrows, Figures 4−9). This plasticity consisted mainly of variations in the number of branches of the thallus, diameters, thickenings at the base of the dichotomies and apices from acute to blunt or rounded (Figures 4−9). In Taylor (1942), two different morphotypes of C. minima are presented, the typical little branched and another highly branched. This second is similar to the one presented by Nelson and Duffy (1991) for New Zealand and Santiañez et al. (2018b) from Easter Island, but all of them, despite presenting dense branching, also present notably elongated central branching segments, typical of the morphology of C. minima that we have observed in all our specimens.
In general, despite their abundance and wide distribution on the Mexican coasts, little is known about the species of Scytosiphonaceae and their phylogenetic relationships, as well as their real diversity or the historical geological processes responsible for their current distribution. From this first approach to the study of C. minima populations on the Mexican coast, questions remain about the real distribution of C. minima and C. ramosissima in other regions of the world, as well as their relationship with C. bicanaliculata, since the information available so far is practically of morphology only, so it is unknown if there is greater cryptic diversity within C. minima or not. However, these studies must be extended to other members of the Scytosiphonacae in Mexico, given their ecological importance and abundance, in order to form a more consolidated integration of their knowledge.
Funding source: Universidad Autónoma Metropolitana
Funding source: Programa para el Desarrollo Profesional Docente de la Secretaría de Educación Pública (PRODEP-SEP)
Award Identifier / Grant number: UAMI-CBS2023-2026
Award Identifier / Grant number: UAMI-CA-117
About the authors

María Luisa Núñez Resendiz is a full-time professor at the Universidad Autónoma Metropolitana, where she received her PhD degree. Currently, she is working on phylogeny, taxonomy and phylogeography of marine algae.

Kurt M. Dreckmann is a full-time professor at the Universidad Autónoma Metropolitana, where he received his PhD degree. Over 35 years, his research has focused on ecology, taxonomy, biogeography, and phylogeny of marine algae.

Oscar E. Hernández is currently doing his PhD degree at Universidad Autónoma Metropolitana, where he is working in biogeography and systematics of marine algal species in Mexico.

Carlos Adán Palma-Ortíz is currently doing his PhD degree at Universidad Autónoma Metropolitana, where he is working on population genetics, phylogeography and systematics of marine algal species in Mexico.

Abel Sentíes is currently a professor-researcher of marine botany at Universidad Autónoma Metropolitana. He has spent over 35 years at the Marine Macroalgae Laboratory, focusing his research on systematics, biogeography, and conservation of seaweeds.
Acknowledgments
The authors would like to thank the anonymous reviewers. CAPO thanks the Consejo Nacional de Ciencia y Tecnología (CONACyT) for the scholarship granted.
-
Research ethics: Not applicable.
-
Informed consent: Not applicable.
-
Author contributions: The authors have accepted responsibility for the entire content of this manuscript and approved its submission. All the authors have actively contributed to this study with sampling, analysis, discussion of the results and writing.
-
Use of Large Language Models, AI and Machine Learning Tools: None declared.
-
Conflict of interest: The authors state no conflict of interest.
-
Research funding: This work was supported by the projects: UAMI-CBS2023-2026 y UAMI-CA-117, Programa para el Desarrollo Profesional Docente de la Secretaría de Educación Pública (PRODEP-SEP) and Universidad Autónoma Metropolitana.
-
Data availability: Not applicable.
References
Agardh, J.G. (1847). Nya alger från Mexico. Öfversigt af Kongl. In Vetenskaps-Adademiens Förhandlingar, Stockholm, Vol. 4, pp. 5–17.Search in Google Scholar
Anderson, R.J., Stegenga, H., and Bolton, J.J. (2016). Seaweeds of the South African South coast. World Wide Web Electronic Publication, University of Cape Town, Available at: http://southafrseaweeds.uct.ac.za (Accessed 28 October 2024).Search in Google Scholar
Ang, P.O.Jr., Leung, S.M., and Choi, M.M. (2014). A verification of reports of marine algal species from the Philippines. Philipp. J. Sci., Special Issue 142: 5–49.Search in Google Scholar
Ballantine, D.L., Norris, J.N., and Ruiz, H. (2021). The marine benthic algal flora of Puerto Rico, I. Ochrophyta: Phaeophyceae, Pelagophyceae, and Xanthophyceae. Smithson. Contrib. Bot. 114: 1–114.10.5479/si.16709413Search in Google Scholar
Boo, S.M., Lee, K.M., Cho, G.Y., and Nelson, W. (2011). Colpomenia claytonii sp. nov. (Scytosiphonaceae, Phaeophyceae) based on morphology and mitochondrial cox 3 sequences. Bot. Mar. 54: 159–167, https://doi.org/10.1515/bot.2011.017.Search in Google Scholar
Cho, Y.G., Kogame, K., and Boo, S.M. (2006). Molecular phylogeny of the family Scytosiphonaceae (Phaeophyceae). Algae 21: 175–183, https://doi.org/10.4490/algae.2006.21.2.175.Search in Google Scholar
Coates, A.G. and Obando, J.A. (1996) The geologic evolution of the Central American Isthmus. In: Jackson, J.B.C., Budd, A.F., and Coates, A.G. (Eds.). Evolution and environment in tropical America. The University of Chicago Press, Chicago, USA, pp. 21–56.Search in Google Scholar
Coates, A.G. and Stallard, R.F. (2013). How old is the Isthmus of Panama? Bull. Mar. Sci. 89: 801–814, https://doi.org/10.5343/bms.2012.1076.Search in Google Scholar
Coppejans, E., Prathep, A., Leliaert, F., Lewmanomont, K., and De Clerck, O. (2011). Seaweeds of Mu Ko Tha Lae Tai (SE Thailand). Methodologies and field guide to the dominant species. Biodiversity Research and Training Program (BRT), Bangkok, Thailand.Search in Google Scholar
Delnatte, C. and Wynne, M.J. (2016). A revised checklist of marine algae and seagrasses of Martinique, French West Indies. Nova Hedwigia 102: 415–440, https://doi.org/10.1127/nova_hedwigia/2016/0364.Search in Google Scholar
Duncan, E.J. and Lee Lum, L.M. (2006). A checklist of the marine macroalgae of the Republic of Trinadad and Tobago. Caribb. Mar. Stud. 7: 1–96.Search in Google Scholar
Einav, R., Guiry, M.D., and Israel, A. (2021). A revised list of seaweeds from the Red Sea (1756–2020). Isr. J. Plant Sci. 67: 1–73, https://doi.org/10.1163/22238980-bja10036.Search in Google Scholar
Farlow, W.G. (1881). The marine algae of New England. Rep. U.S. Fish Commission 1879: 1–120.Search in Google Scholar
Feldmann, J. (1949). L’ordre des Scytosiphonales. Mem. Soc. Hist. Nat. Afr. Nord. 2: 103–115.Search in Google Scholar
García-García, A.M.E., Cabrera-Becerril, E., Núñez-Resendiz, M.L., Dreckmann, K.M., and Sentíes, A. (2021). Actualización taxonómica de las algas pardas (Phaeophyceae, Ochrophyta) marinas bentónicas del Atlántico mexicano. Acta Bot. Mex. 128: e1968, https://doi.org/10.21829/abm128.2021.1968.Search in Google Scholar
GBIF. (2024). Global biodiversity information facility (GBIF) occurrence download. Available at: http://GBIF.org (Accessed 25 May 2024)Search in Google Scholar
Guiry, M.D. and Guiry, G.M. (2024). AlgaeBase. Galway: world-wide electronic publication. National University of Ireland, Available at: https://www.algaebase.org (Accessed 15 May 2024).Search in Google Scholar
Hall, T.A. (1999). BioEdit a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. 41: 95–98.Search in Google Scholar
Hoshino, M., Croce, M.E., Hanyuda, T., and Kogame, K. (2020). Species delimitation of Planosiphon gracilis morphospecies (Scytosiphonaceae, Phaeophyceae) from Japan and the description of Pl. nakamurae sp. nov. Phycologia 59: 116–126, https://doi.org/10.1080/00318884.2019.1709144.Search in Google Scholar
Hoshino, M., Tanaka, A., Kamiya, M., Uwai, S., Hiraoka, M., and Kogame, K. (2021). Systematics, distribution, and sexual compatibility of six Scytosiphon species (Scytosiphonaceae, Phaeophyceae) from Japan and the description of four new species. J. Phycol. 57: 416–434, https://doi.org/10.1111/jpy.13089.Search in Google Scholar PubMed
Huisman, J.M. (2015). Algae of Australia: marine benthic algae of north-western Australia. 1. Green and brown algae. ABRS & CSIRO Publishing, Canberra & Melbourne, pp. 320.Search in Google Scholar
Huisman, J.M., Abbott, I.A., and Smith, C.M. (2007). Hawaiian reef plants. University of Hawai’i Sea Grant College Program, Honolulu, USA.Search in Google Scholar
John, D.M., Prud’homme van Reine, W.F., Lawson, G.W., Kostermans, T.B., and Price, J.H. (2004). A taxonomic and geographical catalogue of the seaweeds of the western coast of Africa and adjacent islands. Nova Hedwigia 127: 1–339.Search in Google Scholar
Kawai, H. and Sasaki, H. (2004). Morphology, life history, and molecular phylogeny of Stschapovia flagellaris (Tilopteridales, Phaeophyceae) and the erection of the family Stschapoviaceae fam. nov. J. Phycol. 40: 1156–1169, https://doi.org/10.1111/j.1529-8817.2004.03153.x.Search in Google Scholar
Kogame, K., Horiguchi, T., and Masuda, M. (1999). Phylogeny of the order Scytosiphonales (Phaeophyceae) based on DNA sequences of rbcL, partial rbcS, and partial LSU rDNA. Phycologia 38: 496–502, https://doi.org/10.2216/i0031-8884-38-6-496.1.Search in Google Scholar
Kogame, K., Uwai, S., Shimada, S., and Masuda, M. (2005). A study of sexual and asexual populations of Scytosiphon lomentaria (Scytosiphonaceae, Phaeophyceae) in Hokkaido, northern Japan, using molecular markers. Eur. J. Phycol. 40: 313–322, https://doi.org/10.1080/09670260500193008.Search in Google Scholar
Kraft, G.T. (2009). Algae of Australia. Marine benthic algae of Lord Howe Island and the southern great Barrier reef. 2. Brown algae. Australian Biological Resources Study and CSIRO Publishing, Canberra & Melbourne, Australia.Search in Google Scholar
Krishnamurthy, V. and Baluswami, M. (2010). Phaeophyceae of India and neighbourhood. Krishnamurthy Institute of Algology, Chennai.Search in Google Scholar
Kumar, S., Stecher, G., Li, M., Kniyaz, C., and Tamura, K. (2018). MEGA X: molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 35: 1547–1549, https://doi.org/10.1093/molbev/msy096.Search in Google Scholar PubMed PubMed Central
Lee, K.M., Hong, D.D., and Boo, S.M. (2014a). Phylogenetic relationships of Rosenvingea (Scytosiphonaceae, Phaeophyceae) from Vietnam based on cox3 and psaA sequences. Algae 29: 389–387, https://doi.org/10.4490/algae.2014.29.4.289.Search in Google Scholar
Lee, K.M., Boo, G.H., Coyer, J.A., Nelson, W.A., Miller, K.A., and Boo, S.M. (2014b). Distribution patterns and introduction pathways of the cosmopolitan brown alga Colpomenia peregrina using mt cox 3 and atp 6 sequences. J. Appl. Phycol. 26: 491–504, https://doi.org/10.1007/s10811-013-0052-1.Search in Google Scholar
Lewis, J.E. and Norris, J.N. (1987). A history and annotated account of the benthic marine algae of Taiwan. Smithson. Contrib. Mar. Sci. 29: 1–38, https://doi.org/10.5479/si.01960768.29.iv.Search in Google Scholar
Lim, P.E., Sakaguci, M., Hayunda, T., Kogame, K., Phang, S.M., and Kawai, H. (2007). Molecular phylogeny of crustose brown seaweeds (Ralfsiales, Phaeophyceae) inferred from rbcL sequences resulting in proposal for Neoralfsiaceae fam. nov. Phycologia 46: 456–466.10.2216/06-90.1Search in Google Scholar
Littler, D.S. and Littler, M.M. (2003). South Pacific reef plants. A diver’s guide to the plant life of the South Pacific Coral Reefs. OffShore Graphics, Inc., Washington, DC, USA.Search in Google Scholar
Liu, R.Y. (2008). Checklist of biota of Chinese seas. Science Press, Academia Sinica, Beijing, China.Search in Google Scholar
Lobban, C.S. and Tsuda, R.T. (2003). Revised checklist of benthic marine macroalgae and seagrasses of Guam and Micronesia. Micronesica 35/36: 54–99.Search in Google Scholar
McDevit, D.C. and Saunders, G.W. (2017). A molecular investigation of Canadian Scytosiphonaceae (Phaeophyceae) including descriptions of Planosiphon gen. nov. and Scytosiphon promiscuus sp. nov. Botany 95: 653–671, https://doi.org/10.1139/cjb-2017-0042.Search in Google Scholar
Milne, I., Lindner, D., Bayer, M., Husmeier, D., McGuire, G., Marshall, D.F., and Wright, F. (2009). TOPALi v2: a rich graphical interface for evolutionary analyses of multiple alignments on HPC clusters and multi-core desktops. Bioinformatics 25: 126–127, https://doi.org/10.1093/bioinformatics/btn575.Search in Google Scholar PubMed PubMed Central
N’Yeurt, A. and Payri, C.E. (2006). Marine algal flora of French Polynesia I. Phaeophyceae (Ochrophyta, brown algae). Cryptogamie, Algol. 27: 111–152.Search in Google Scholar
Nakamura, Y. and Tatewaki, M. (1975). The life history of some species of Scytosiphonales. Sci. Pap. Inst. Algol. Res., Fac. Sci. Hokkaido Univ. 6: 57–93.Search in Google Scholar
Nelson, W.A. and Duffy, C.A. (1991). Chnoospora minima (Phaeophyta) in Port Underwood, Marlborough, a curious new algal record for New Zealand, New Zealand. J. Bot. 29: 341–344, https://doi.org/10.1080/0028825x.1991.10416612.Search in Google Scholar
Nelson, W.A., D’Archino, R., Neill, K.F., and Robinson, N.M. (2021). Introduced marine macroalgae: new perspectives on species recognition and distribution in New Zealand. Bot. Mar. 64: 379–393, https://doi.org/10.1515/bot-2021-0042.Search in Google Scholar
Nguyen, T.V., Le, N.H., Lin, S.-M., Steen, F., and De Clerck, O. (2013). Checklist of the marine macroalgae of Vietnam. Bot. Mar. 56: 207–227.10.1515/bot-2013-0010Search in Google Scholar
Norris, J.N., Aguilar-Rosas, L.E., and Pedroche, F.F. (2017). Conspectus of the benthic marine algae of the Gulf of California: Rhodophyta, Phaeophyceae, and Chlorophyta. Smithson. Contrib. Bot. 106: 1–125, https://doi.org/10.5479/si.1938-2812.106.Search in Google Scholar
Papenfuss, G.F. (1956). Notes on South African marine algae. IV. S. Afr. J. Bot. 22: 65–77, https://doi.org/10.5962/p.392380.Search in Google Scholar
Pedroche, F.F. and Sentíes, A. (2020). Diversidad de macroalgas marinas en México. Una actualización florística y nomenclatural. Cymbella 6: 4–55.Search in Google Scholar
Pedroche, F.F., Silva, P.C., Aguilar-Rosas, L.E., Dreckmann, K.M., and Aguilar-Rosar, R. (2008). Catálogo de las algas marinas bentónicas del Pacífico de México II. Phaeophycota. Universidad Autónoma Metropolitana, Universidad Autónoma de Baja California & University of California Berkeley, Mexicali & Berkeley.Search in Google Scholar
Rao, P.S.N., and Gupta, R.K. (Eds.). (2015). Algae of India. Volume 3. A checklist of Indian marine algae (excluding diatoms & dinoflagellates). Botanical Survey of India Ministry of Environment, Forests and Climate Change Government of India, Salt Lake, Kolkata.Search in Google Scholar
Ronquist, F., Teslenko, M., Van Der Mark, P., Ayres, D.L., Darling, A., Hohna, S., Larget, B., Liu, L., Suchard, M.A., and Huelsenbeck, J.P. (2012). MrBayes 3.2: efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 61: 539–542, https://doi.org/10.1093/sysbio/sys029.Search in Google Scholar PubMed PubMed Central
Santiañez, W.J.E. and Kogame, K. (2019). Proposals to recognize Petalonia tenella comb. nov. and to resurrect Hapterophycus canaliculatus (Scytosiphonaceae, Phaeophyceae). Bot. Mar. 62: 149–153, https://doi.org/10.1515/bot-2018-0077.Search in Google Scholar
Santiañez, W.J.E. and Kogame, K. (2022). Manzaea minuta gen. et comb. nov. (Scytosiphonaceae, Phaeophyceae) from the tropical Northwestern Pacific Ocean. Philipp. J. Syst. Biol. 16: 1–7.10.26757/pjsb2022a16001Search in Google Scholar
Santiañez, W.J.E. and Wynne, J. (2024). Overview of the systematics of the brown algal family Scytosiphonaceae (Ectocarpales, Phaeophyceae). Jeju J. Isl. ScI. 1: 20–33.Search in Google Scholar
Santiañez, W.J.E., Lee, K.M., Uwai, S., Kurihara, A., Geraldino, P.J.L., Ganzon-Fortes, E.T., Boo, S.M., and Kogame, K. (2018a). Untangling nets: elucidating the diversity and phylogeny of the clathrate brown algal genus Hydroclathrus, with the description of a new genus Tronoella (Scytosiphonaceae, Phaeophyceae). Phycologia 57: 61–78, https://doi.org/10.2216/17-68.1.Search in Google Scholar
Santiañez, W.J.E., Macaya, E.C., Lee, K.M., Cho, G.Y., Boo, S.M., and Kogame, K. (2018b). Taxonomic reassessment of the Indo-Pacific Scytosiphonaceae (Phaeophyceae): Hydroclathrus rapanuii sp. nov. Chnoospora minima from Easter Island, with proposal of Dactylosiphon gen. nov. and Pseudochnoospora gen. nov. Bot. Mar. 61: 47–64, https://doi.org/10.1515/bot-2017-0089.Search in Google Scholar
Santiañez, W.J.E., Al-Bader, D., West, J.A., Bolton, J.J., and Kogme, K. (2020). Status, morphology, and phylogenetic relationships of Iyengaria (Scytosiphonaceae, Phaeophyceae), a brown algal genus with a disjunct distribution in the Indian Ocean. Phycol. Res. 68: 323–331, https://doi.org/10.1111/pre.12438.Search in Google Scholar
Schnetter, R. (1976). Marine Algen der karibischen Küsten von Kolumbien. I. Phaeophyceae. Bibl. Phycol. 24: 1–125.Search in Google Scholar
Schultz, N.E., Lane, C.E., Le Gall, L., Gey, D., Bigney, A.R., De Reviers, B., Rousseau, F., Schneider, C.W., Lamouroux, J.V., and Oliveira, E.C. (2015). A barcode analysis of the genus Lobophora (Dictyotales, Phaeophyceae) in the western Atlantic Ocean with four novel species and the epitypification of L. variegata. Eur. J. Phycol. 50: 481–500, https://doi.org/10.1080/09670262.2015.1078500.Search in Google Scholar
Serviere-Zaragoza, E., Riosmena-Rodríguez, R., León-Tejera, H., and González-González, J. (2007). Distribución espacial de macroalgas marinas en las islas Revillagigedo, México. Ciencia Mar. 11: 3–13.Search in Google Scholar
Setchell, W.A. and Gardner, N.L. (1925). The marine algae of the Pacific coast of North America. Part III. Melanophyceae. Univ. Calif. Publ. Bot. 8: 383−898.10.5962/bhl.title.5719Search in Google Scholar
Sherwood, A.R. and Guiry, M.D. (2023). Inventory of the seaweeds and seagrasses of the Hawaiian Islands. Biology 12: 1–71, https://doi.org/10.3390/biology12020215.Search in Google Scholar PubMed PubMed Central
Silberfeld, T., Racault, M.-F.L.P., Fletcher, R.L., Couloux, A., Rousseau, F., and De Reviers, B. (2011). Systematics and evolutionary history of pyrenoidbearing taxa in brown algae (Phaeophyceae). Eur. J. Phycol. 46: 361–377, https://doi.org/10.1080/09670262.2011.628698.Search in Google Scholar
Silva, P.C., Basson, P.W., and Moe, R.L. (1996). Catalogue of the benthic marine algae of the Indian Ocean. Univ. Calif. Publ. Bot. 79: 1–1259.Search in Google Scholar
Skelton, P.A. and South, G.R. (2007). The benthic marine algae of the Samoan Archipelago, south Pacific, with emphasis on the Apia District. Nova Hedwigia Beih. 132: 1–350.Search in Google Scholar
Soe-Htun, U., Kyaw, S.P.P., Wai, M.K., San, J., Khaing, S.M.M., and Aye, C.T.P.P. (2021). A review on the seaweed resources of Myanmar. J. Aquac. Mar. Biol. 10: 152–166.10.15406/jamb.2021.10.00317Search in Google Scholar
South, G.R. and Skelton, P.A. (2003). Catalogue of the marine benthic macroalgae of the Fiji islands, south pacific. Aust. Syst. Bot. 16: 699–758, https://doi.org/10.1071/sb03011.Search in Google Scholar
Stamatakis, A. (2006). RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 22: 2688–2690, https://doi.org/10.1093/bioinformatics/btl446.Search in Google Scholar PubMed
Suárez, A.M., Martínez-Daranas, B., Alfonso Sánchez, Y., Moreira-González, A.R., and Capote, J. (2023). Lista actualizada de las macroalgas marinas cubanas. Acta Bot. Mex. 130: 1–61, https://doi.org/10.21829/abm130.2023.2196.Search in Google Scholar
Taylor, W.R. (1942). Caribbean marine algae of the Allan Hancock Expedition, 1939. Allan Hancock Atlantic expedition, USA.Search in Google Scholar
Taylor, W.R. (1960). Marine algae of the eastern tropical and subtropical coasts of the Americas. The University of Michigan Press, Ann Arbor, USA.Search in Google Scholar
Taylor, W.R. (1973). Marine algae of the Smithsonian-Bredin Expedition to the Society and Tuamoto islands. Pac. Sci. 27: 37–43.Search in Google Scholar
Thiers, B. (2021). Index Herbariorum: a global directory of public herbaria and associated staff. New York Botanical Garden’s. Virtual Herbarium. http://www.nybg.org/bsci/ih/ih.html (consulted August 2021).Search in Google Scholar
Tsuda, R.T. (2003). Checklist and bibliography of the marine benthic algae from the Mariana Islands (Guam and CNMI). Technical Report. University of Guam Marine Laboratory, Vol. 107, pp. 1–49.Search in Google Scholar
Tsuda, R.T. and Walsh, S.K. (2013). Bibliographic checklist of the marine benthic algae of Central Polynesia in the Pacific Ocean (excluding ‘i and French Polynesia). Micronesica 2013–02: 1–91.Search in Google Scholar
Vieira, C., Kim, M.S., and Zubia, M. (2024). French Polynesian Scytosiphonaceae (Ectocarpales, Phaeophyceae): a combined molecular and morphological approach to their diversity and systematics. J. Phycol. 60: 447–464, https://doi.org/10.1111/jpy.13432.Search in Google Scholar PubMed
Womersley, H.B.S. and Bailey, A. (1970). Marine algae of the Solomon Islands. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 259: 257–352.10.1098/rstb.1970.0060Search in Google Scholar
Wynne, M.J. (1969). Life history and systematic studies of some Pacific North American Phaeophyceae (brown algae). Univ. Calif. Publ. Bot. 50: 1−88.Search in Google Scholar
Wynne, M.J. (2022). A checklist of benthic marine algae of the tropical and subtropical Western Atlantic: fifth revision. Nova Hedwigia Beih. 153: 1–178.Search in Google Scholar
Yadav, S.K., Kumar, Y., A.S., and Palanisamy, M. (2023). Taxonomic account of two brown seaweed genera Chnoospora and Pseudochnoospora (Phaeophyta) in India. J. Plant Dev. Sci. 15: 415–423.Search in Google Scholar
Yoshida, T., Suzuki, M., and Yoshinaga, K. (2015). Checklist of marine algae of Japan (revised in 2015). Jpn. J. Phycol. 63: 129–189.Search in Google Scholar
Supplementary Material
This article contains supplementary material (https://doi.org/10.1515/bot-2024-0067).
© 2025 the author(s), published by De Gruyter, Berlin/Boston
This work is licensed under the Creative Commons Attribution 4.0 International License.
Articles in the same Issue
- Frontmatter
- In this issue
- Physiology and Ecology
- Bacteria-released algal growth and morphogenesis factors regenerate axenic calli derived from the macroalga Ulva (Chlorophyta) and change the fatty acid profile
- Annual cycle and ecological interactions of macroalgal epiphytes with Cymodocea nodosa and Ruppia maritima in Epanomi and Vravrona, Greece
- Past and present fucalean diversity in the island of Marettimo, Egadi Islands Marine Protected Area (Central Mediterranean, Italy)
- Taxonomy/Phylogeny and Biogeography
- Morphological and molecular characterization of Chnoospora minima (Scytosiphonaceae, Ectocarpales) along Mexican coasts, with the description of C. ramosissima sp. nov.
- Phylogeography of the marine benthic alga Gracilaria salicornia (Gracilariales, Rhodophyta) in Southeast Asia
- Unveiling a novel species, Pachymeniopsis shinchonai sp. nov., from Korea, with reference to phylogenetic relationships within the Grateloupiaceae (Halymeniales, Rhodophyta)
- First record of the red alga Ceramium pallidum (Ceramiales, Ceramiaceae) for the Mediterranean Sea
- Chemistry and Applications
- Chemical profiling of seaweeds of the Arabian Gulf by liquid chromatography-mass spectrometry and in-silico screening against MPOX
Articles in the same Issue
- Frontmatter
- In this issue
- Physiology and Ecology
- Bacteria-released algal growth and morphogenesis factors regenerate axenic calli derived from the macroalga Ulva (Chlorophyta) and change the fatty acid profile
- Annual cycle and ecological interactions of macroalgal epiphytes with Cymodocea nodosa and Ruppia maritima in Epanomi and Vravrona, Greece
- Past and present fucalean diversity in the island of Marettimo, Egadi Islands Marine Protected Area (Central Mediterranean, Italy)
- Taxonomy/Phylogeny and Biogeography
- Morphological and molecular characterization of Chnoospora minima (Scytosiphonaceae, Ectocarpales) along Mexican coasts, with the description of C. ramosissima sp. nov.
- Phylogeography of the marine benthic alga Gracilaria salicornia (Gracilariales, Rhodophyta) in Southeast Asia
- Unveiling a novel species, Pachymeniopsis shinchonai sp. nov., from Korea, with reference to phylogenetic relationships within the Grateloupiaceae (Halymeniales, Rhodophyta)
- First record of the red alga Ceramium pallidum (Ceramiales, Ceramiaceae) for the Mediterranean Sea
- Chemistry and Applications
- Chemical profiling of seaweeds of the Arabian Gulf by liquid chromatography-mass spectrometry and in-silico screening against MPOX