Die Regulation der hippokampalen Informationsenkodierung durch metabotrope Glutamatrezeptoren
-
Denise Manahan-Vaughan
Denise Manahan-Vaughan ist Neurophysiologin, Neurowissenschaftlerin und Leiterin der Abteilung für Neurophysiologie an der Medizinischen Fakultät der Ruhr-Universität Bochum (www.rub.de/neurophys). Gleichzeitig ist sie auch Direktorin und Studiendekanin der International Graduate School of Neuroscience (www.rub.de/igsn) und Sprecherin des Sonderforschungsbereichs „Integration und Repräsentation Sensorischer Prozesse“ (www.rub.de/sfb874) an der Ruhr-Universität. Nach ihrem ersten Studienabschluss in Naturwissenschaften am Trinity College Dublin, Irland, wurde sie auch dort in Neuropharmakologie promoviert. Anschließend ging sie nach Deutschland an das Leibniz-Institut für Neurobiologie in Magdeburg. Nachdem sie ihre Habilitation in Physiologie an der Otto-von-Guericke-Universität Magdeburg erlangte, ging sie an das Johannes-Müller-Zentrum für Physiologie an der Charité, Berlin und im Jahr 2003 nach Bochum. Ihre Forschungsprojekte beschäftigen sich mit der Beziehung zwischen hippokampaler und kortikaler synaptischer Plastizität und Langzeit-assoziativer und räumlicher Gedächtnisbildung.


Zusammenfassung
Der Hippokampus unterstützt den Erwerb von sowohl räumlichen Repräsentationen als auch von langfristigem räumlichen Gedächtnis. Dies wird ermöglicht durch das Zusammenwirken dreier physiologischer Prozesse, nämlich der Organisation und dem Transfer von Informationen mittels neuronaler Oszillationen, der Erstellung von kontextabhängigen räumlichen Karten mittels Ortszellen und der langfristigen Speicherung räumlicher Lernerfahrung mittels synaptischer Plastizität. Alle drei Prozesse werden durch das glutamaterge System ermöglicht. Die Bindung von Glutamat an ionotrope Glutamatrezeptoren ermöglicht sowohl die schnelle exzitatorische synaptische Transmission (über AMPA-Rezeptoren) als auch die Initiierung einer synaptischen Langzeitspeicherung (über NMDA-Rezeptoren). Aber Glutamat bindet sich auch an metabotrope Glutamat-Rezeptoren (mGlu). Diese Rezeptoren tragen nicht nur zur Stabilität der hippokampalen Enkodierung und der Langlebigkeit der synaptischen Plastizität bei, sie können auch die synaptische Informationsspeicherung unabhängig von der Aktivierung des NMDA-Rezeptors unterstützen und sind für Erwerb und Erhalt des Langzeitgedächtnisses wichtig.
Einführung
Der Hippokampus dient als Drehscheibe für die Erstellung von Gedächtnissen für Ereignisse (in nicht humanen Tieren) (Eichenbaum, 2017) bzw. Episoden (in Menschen) (Horner und Doeller, 2017), und seine herausragende Rolle bei der Ermöglichung räumlicher Kognition unterstützt sehr wahrscheinlich diese Prozesse. Der Hippokampus integriert Sinneserfahrungen in räumliche Repräsentationen und das räumliche Langzeitgedächtnis (Manahan-Vaughan, 2017). Dies geschieht, indem langfristige Veränderungen der synaptischen Effizienz in Form von synaptischer Plastizität (Bliss und Collingridge, 1993; Martin und Buno 2005; Kemp und Manahan-Vaughan, 2007; Manahan-Vaughan, 2017), Netzwerk-Oszillationsaktivität (Buzsaki und Draguhn 2004; Hasselmo 2005), und Bildung von Ortszellen (O’Keefe und Dostovsky 1971; Knierim et al. 1995) ermöglicht werden. Der Neurotransmitter Glutamat ist für diese Prozesse von höchster Bedeutung. Glutamat bindet sich an zwei Kategorien von Neurotransmitter-Rezeptoren. Diese bestehen aus ionotropen und metabotropen Glutamatrezeptoren. Ionotrope Glutamatrezeptoren sind ligandenabhängige Ionenkanäle, bestehend aus α-Amino-3-hydroxy-5-methyl-4-isoxazol-propionsäure – (AMPA), Kainat- und N-Methyl-D-Aspartat (NMDA)-Rezeptoren. Während AMPA-Rezeptoren eine schnelle exzitatorische synaptische Übertragung ermöglichen, sind NMDA-Rezeptoren Schlüsselelemente bei der Induktion synaptischer Plastizitätsprozesse wie der Langzeitpotenzierung (LTP) (Bliss et al., diese Neuroforum-Ausgabe). Metabotrope Glutamatrezeptoren (mGlu) sind G-Protein-gekoppelte Rezeptoren, die über Second-Messenger-Systeme wirken und den intrazellulären Adenylylcyclase- oder Phospholipase C – Spiegel regulieren (Tabelle 1). Diese Rezeptoren spielen eine sehr wichtige Rolle hinsichtlich der Informationsverarbeitung im Hippokampus, die sich auf den langfristigen Erhalt räumlicher Lernerlebnisse bezieht (Mukherjee und Manahan-Vaughan, 2012).
Klassifizierung von mGlu-Rezeptoren und synaptischer Verteilung
mGlu Rezeptor | Subtypen | Hippokampale Expression | Bindung |
Gruppe I | mGlu1, mGlu5 | Überwiegend postsynaptisch | Phospholipase C |
Gruppe II | mGlu2 mGlu3 | Überwiegend präsynaptisch | Adenylyl cyclase |
Gruppe III | mGlu4,mGlu6*, mGlu7,mGlu8 | Überwiegend präsynaptisch | Adenylyl cyclase |
Rezeptoren werden anhand ihrer Signaltransduktionsmechanismen und pharmakologischen Eigenschaften klassifiziert (siehe: Mukherjee und Manahan-Vaughan, 2013 zur Übersicht). Gruppe-I-mGlu-Rezeptoren sind positiv an Phospholipase C gekoppelt und sind hauptsächlich postsynaptisch im Hippokampus lokalisiert. Die Rezeptoren der Gruppen II und III sind negativ an Adenylylcyclase gekoppelt und präsynaptisch lokalisiert (Ohishi et al., 1993; Mukherjee und Manahan-Vaughan, 2012; Goddyn et al., 2015; Tanabe et al., 1993; Okamoto und al., 1994) Corti et al., 1998). * N. B.: mGlu6 wird ausschließlich in der Retina exprimiert (Nomura et al., 1994).
Ortszellen, synaptische Plastizität, neuronale Oszillationen
Ortszellen sind Pyramidenzellen, die hauptsächlich in den CA1- und CA3-Regionen des Hippokampus von Nagern (Grieves und Jeffery, 2017) und Menschen (Ekstrom et al., 2003) vorkommen und hochfrequente Entladungen aufweisen, wenn Versuchstiere einen spezifischen Ort einer Umgebung durchqueren. Der Ort, an dem die Ortszelle feuert, wird als „Ortsfeld“ bezeichnet (Abb. 1). Das Feuerverhalten von Ortszellen ist spezifisch auf den Kontext bezogen, in dem sich das Tier befindet, wobei sensorische Signale aus unterschiedlichen Modalitäten als Basis zur Erstellung einer „räumlichen Karte“ verwendet werden können (Zhang und Manahan-Vaughan, 2015). Während sich Ortsfelder entwickeln und stabilisieren, solange ein Tier durch eine räumliche Umgebung navigiert und sich damit vertraut macht, wird die langfristige Speicherung von räumlicher Erfahrung in Form von Langzeitgedächtnis durch die synaptische Plastizität des Hippokampus ermöglicht: An Nagetieren wurde bewiesen, dass synaptische Plastizität in Form von Langzeitpotenzierung (LTP) und Langzeitdepression (LTD), die länger als 24 Stunden andauert, stark mit der Bildung und Speicherung von räumlichem Gedächtnis verbunden ist. Tatsächlich wurde an Mäusen gezeigt, dass synaptische Plastizität direkt durch räumliche Erfahrung ausgelöst werden kann (Goh und Manahan-Vaughan, 2013a).
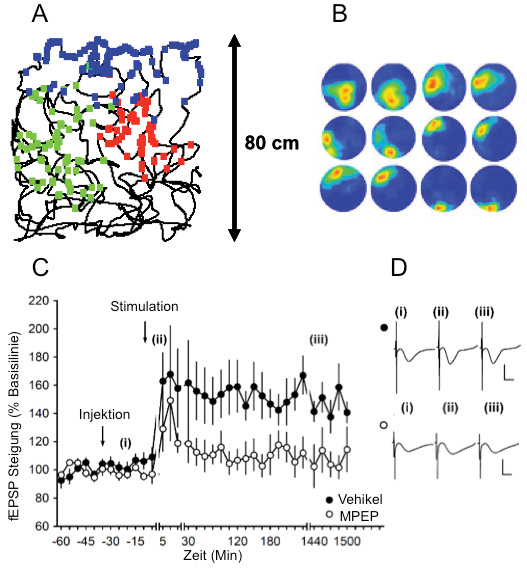
Regulation von hippokampalen Ortsfeldern und synaptischer Plastizität durch mGlu5-Rezeptoren
A: Die dünne schwarze Linie zeigt den Weg, den eine Ratte nahm, als sie eine 80 × 80 cm große quadratische Umgebung erforschte, während mittels Tetroden aus der CA1-Region des Hippokampus abgeleitet wurde.
Die grünen, blauen und roten Punkte zeigen die jeweiligen Regionen der Umgebung, in denen drei verschiedene Ortszellen aufleuchteten, wobei die von einer Farbe bedeckte Region dem Ortsfeld dieser Zelle entspricht.
B: Tiere, die während der Exploration einer quadratischen Umgebung zuerst stabile Ortsfelder zeigten, wurden mit einem mGlu5-Rezeptorantagonisten behandelt und durften dann eine neue (runde) Umgebung erkunden. Die Exposition in der gleichen Umgebung einen Tag später war mit einer Neuverortung der Felder verbunden, was darauf hinweist, dass die Stabilisierung und Konsolidierung der Ortsfelder durch die mGlu5-Antagonisten beeinträchtigt war. Die Beispiele stellen (von links nach rechts) zwei Expositionen in der neuen Umgebung am ersten Tag dar, gefolgt von zwei Expositionen in derselben Umgebung am zweiten Tag. Von oben nach unten: Ortsfelder von Ortszellen, die von drei verschiedenen Tieren aufgezeichnet wurden. (Von Zhang und Manahan-Vaughan, 2014).
C: Antagonismus von mGlu5 verhindert hippokampale LTP, die bei Ratten normalerweise länger als 24 Stunden andauern würde.
Die Hochfrequenzstimulation (HFS) (4 Folgen von 100 Impulsen bei 100 Hz) von MF-CA3-Synapsen löst in Vehikel-injizierten, sich frei bewegenden Tieren eine Langzeitpotenzierung LTP (> 24h) aus. Eine intrazerebrale Applikation des mGlu5-Antagonisten MPEP (1,8 μg) vor der HFS blockiert signifikant die LTP.
D: fEPSP-Beispiele
Darstellung der fEPSPs, die während eines LTP-Experiments hervorgerufen wurden. Die Ableitungen wurden von den MF-CA3 Synapsen gemacht. Oben: Vehikel-behandelte, unten: MPEP-behandelte Tiere (i) vor HFS, (ii) 5 min nach HFS und (iii) 24 h nach HFS. (Aus: Hagena und Manahan-Vaughan, 2015).
Neuronale Oszillationen sind ein intrinsischer Bestandteil funktionaler neuronaler Netzwerke und unterstützen eine Echtzeit- und Langzeit-Informationsenkodierung in Bezug auf räumliche Erfahrung. Eine Veränderung der synaptischen Stärke von hippokampalen Neuronen erfolgt schnell während des Verhaltens der Tiere, wobei neuronale Oszillationen in Theta- und Gamma-Frequenzen eine entscheidende Rolle spielen (siehe: Buzsáki, 2005; Buzsáki und Draguhn, 2004). Daher hängen die hippokampalen Theta- und Gamma-Oszillationen funktionell zusammen und leiten sich von intrinsischen oszillatorischen Eigenschaften der Neuronen und Interneuronen ab, deren rhythmische Aktivierung durch intra- und extrahippokampale Verbindungen gesteuert wird (Bartos et al., 2007). Synaptische Plastizität und neuronale Theta-Gamma-Oszillationen sind voneinander abhängig: Veränderungen der Theta-Gamma-Frequenz-Kopplung während der Induktion von LTP sagen voraus, ob der Induktionsversuch erfolgreich sein wird (Bikbaev und Manahan-Vaughan, 2007, 2008), und die Gabe von Stimuli auf dem Höchst- oder Tiefpunkt von hippokampalen Theta-Wellen führt zur Induktion von LTP bzw. LTD (Hölscher et al, 1997).
Die Beteiligung von mGlu – Rezeptoren an Ortszellen, synaptischer Plastizität und neuronalen Oszillationen
Gruppe I-mGlu Rezeptoren
Obwohl die Beteiligung von NMDA-Rezeptoren an der Entstehung von Ortsfeldern nachgewiesen wurde (Kentros et al., 1998), haben sich nur wenige Studien mit der Frage befasst, inwieweit mGlu-Rezeptoren zur Erzeugung oder Stabilität von Ortsfeldern beitragen. Bei Ratten verhindert der pharmakologische Antagonismus des mGlu5-Rezeptors die Langzeitstabilität von Ortsfeldern und reduziert den Informationsgehalt und die Feuerungsraten von Ortszellen in einer neuartigen Umgebung (Zhang und Manahan-Vaughan 2014, Abb. 1). Diese Erkenntnis schafft eine faszinierende Verbindung zwischen der Bildung räumlicher Karten mittels Ortszellen und der langfristigen Enkodierung räumlicher Lernerfahrung mittels synaptischer Plastizität: Der mGlu5-Rezeptor ist sowohl für persistierende Formen der synaptischen Plastizität als auch für das Langzeitgedächtnis von zentraler Bedeutung (Hagena und Manahan-Vaughan, 2017). Darüber hinaus wird der mGlu5-Rezeptor benötigt für die oben erwähnten, durch Tetanisierung induzierten Veränderungen der Theta- und Gamma-Oszillationen, die wiederum die erfolgreiche Expression von LTP vorhersagen (Bikbaev und Manahan-Vaughan, 2017), sowie für die zellspezifische Plastizität durch neuronale Oszillationen (Zarndadze et al., 2016) Von allen mGlu-Rezeptoren könnte der mGlu5-Rezeptor für hippokampale Enkodierungsprozesse am wichtigsten sein: Die Aktivierung dieses Rezeptors ist für das räumliche Langzeitgedächtnis (reference memory) (Naie und Manahan-Vaughan, 2004; Manahan-Vaughan und Braunewell, 2005), das Wiedererkennungsgedächtnis (recognition memory) (Marszalek-Grabska et al., 2018) und das Extinktionslernen (André et al, 2015) erforderlich. Der mGlu5-Rezeptor unterstützt Verstärkungen von LTP, die auf Grund einer Umweltanreicherung (environmental enrichment) auftreten (Buschler und Manahan-Vaughan, 2017). Die Aktivierung des Rezeptors ist essenziell nötig für die Expression von Formen von LTP und LTD, die durch räumliches Lernen unterstützt werden (Popkirov und Manahan-Vaughan, 2011; Goh und Manahan-Vaughan, 2013b; Hagena und Manahan-Vaughan, 2015), und kann Proteinsyntheseformen der synaptischen Plastizität induzieren, ohne dass der NMDA-Rezeptor aktiviert wird (Huber et al., 2001; Naie und Manahan-Vaughan, 2006).
Sein Gegenstück, der mGlu1-Rezeptor, ist ebenso intrinsisch beteiligt an der hippokampalen synaptischen Plastizität und Gedächtnisbildungsprozessen (Naie und Manahan-Vaughan, 2005), einschließlich der vom Hippokampus abhängigen nicht räumlichen assoziativen Lernformen (Gil-Sanz et al., 2008). Während der mGlu5-Rezeptor die (späte) Proteinsynthesephase von LTP und LTD unterstützt (Balschun und Wetzel, 2002; Naie und Manahan-Vaughan, 2004; Popkirov und Manahan-Vaughan, 2011), fördert der mGlu1-Rezeptor die Induktion von LTP (Neymann und Manahan-Vaughan, 2008; Naie und Manahan-Vaughan, 2005). Der Mechanismus umfasst vermutlich die Begünstigung der Erhöhung der intrazellulären Ca2+-Konzentrationen, die neuronale Depolarisation, die Erhöhung der Häufigkeit spontaner inhibitorischer postsynaptischer Potenziale (Mannaioni et al., 2001) und die Regulierung der NMDA-Rezeptorströme (Skeberdis et al. 2001) sowie deren Zyklisierung (Lan et al., 2001).
Gruppe II mGlu-Rezeptoren
Während mGlu-Rezeptoren der Gruppe I sowohl die hippokamale LTP als auch die LTD regulieren, können mGlu-Rezeptoren der Gruppe II nur direkt an der LTD beteiligt sein. Der Antagonismus dieser Rezeptoren verhindert die Entstehung anhaltender Formen der LTD (Manahan-Vaughan, 1997; Kulla et al., 1999) und verhindert interessanterweise ebenso das räumliche Langzeitgedächtnis (Altinbilek und Manahan-Vaughan, 2009). Die agonistische Aktivierung dieser Rezeptoren kann jedoch LTP verhindern, selbst wenn Ligandendosen verwendet werden, die keine Wirkung auf die basale synaptische Übertragung haben (Kulla et al., 1999). Dies kann jedoch mit der Regulation der Erregbarkeit des Hippokampus zusammenhängen: Gruppe-II-Rezeptoren sind im Hippokampus größtenteils präsynaptisch gelegen (Shigemoto et al., 1997) und dienen überwiegend einer Autorezeptorfunktion (Mukherjee und Manahan-Vaughan, 2013). Obwohl die postsynaptische Exprimierung von Gruppe-II-mGlu-Rezeptoren weniger stark ist (Petralia et al., 1996), können sie jedoch bei der Regulation der CA3-Netzwerkaktivität (Ster et al., 2011) für die Informationsverarbeitung in Zusammenhang mit Theta-Aktivität eine entscheidende Rollen spielen.
Es wird für möglich gehalten, dass der mGlu2-Rezeptor am Wiedererkennungsgedächtnis beteiligt ist (Marszalek-Grabska et al., 2018), und es wurde in transgenen Mäusen gezeigt, dass dieser Rezeptor im räumlichen Arbeitsgedächtnis eine Rolle spielt (de Filippis et al., 2015). Transgene Mäuse, denen sowohl mGlu2- als auch mGlu3-Rezeptoren fehlen, sind ebenfalls bei Aufgaben beeinträchtigt, die das räumliche Arbeitsgedächtnis betreffen (Lyon et al., 2011). Diese Ergebnisse liefern wiederum wertvolle Einsichten in die mögliche Beziehung zwischen LTP, LTD und Komponenten des räumlichen Gedächtnisses: Lernaufgaben bezogen auf die räumliche Lage von Gegenständen in Verbindung mit dem Wiedererkennungsgedächtnis lösen LTD im Hippokampus der Maus aus, wohingegen die Magnitude der LTD als Index für die Fähigkeit des räumlichen Arbeitsgedächtnisses dienen könnte (Nakao et al., 2002).
Gruppe III-mGlu-Rezeptoren
Ähnlich wie bei den mGlu-Rezeptoren der Gruppe II beeinträchtigt der Antagonismus der mGlu-Rezeptoren der Gruppe III die Expression persistierender (> 24 h) LTD, aber nicht der LTP, sowohl in der CA1-Region als auch im Gyrus dentatus (Klausnitzer et al., 2004; Altinbilek und Manahan-Vaughan, 2007). Transgene Mäuse, denen mGlu7-Rezeptoren fehlen, weisen ebenfalls Defizite in der Kurzzeit-Potenzierung auf (Bushell et al., 2002). Der Rezeptor-Antagonismus beeinträchtigt bei einem Versuch im Radial Arm Maze das räumliche Referenzgedächtnis (Altinbilek und Manahan-Vaughan, 2007), und die mGlu-Rezeptoren der Gruppen II und III sind in Zusammenhang mit dem Abruf von kontextabhängigem Angstgedächtnis gebracht worden (Szapiro et al, 2001). Transgene Mäuse, denen mGlu7-Rezeptoren fehlen, sind bei Experimenten im Water Maze bei der Bildung des Referenzgedächtnisses beeinträchtigt, wohingegen transgene Mäuse, denen mGlu4- bzw. mGlu8-Rezeptoren fehlen, keine derartigen Defizite aufwiesen (Goddyn et al., 2015). Daraus kann geschlossen werden, dass mGlu7-Rezeptoren von besonderer Bedeutung für die Informationsverarbeitung im Hippokampus sein können. Damit übereinstimmend wurde gezeigt, dass positive allosterische Modulation von mGlu7-Rezeptoren bei einem Maus-Modell mit Rett-Syndrom die LTP wiederherstellt und die kontextuelle Angstkonditionierung und das Erkennen neuartiger Objekte verbessert.
Fazit
MGlu-Rezeptoren sind nicht nur an sich wichtig für die hippokampale Informationsenkodierung, die unterschiedlichen Untergruppen dieser Rezeptoren übernehmen auch unterschiedliche Funktionen. Dies spiegelt sich wider in der Rolle der Rezeptoren der Gruppe I, die die synaptische Plastizität des Hippokampus bidirektional beeinflussen, die überwiegende Beteiligung der Gruppe II und II-Rezeptoren an der LTD und den Unterschieden im Sinne von Gedächtnisformen, die durch mGlu-Rezeptorgruppen reguliert werden. Es ist klar, dass ein enges Wechselspiel zwischen mGlu-Rezeptoren ein Schlüsselfaktor der hippokampalen Informationsverarbeitung ist: Zum einen wurde gezeigt, dass Gruppe-I-Rezeptoren die Expression von Rezeptoren der Gruppe I und II regulieren (Marszalek-Grabska et al., 2008; Bikbaev et al., 2008). Zum anderen wurde die spezifische Rolle dieser Rezeptoren bei der Ermöglichung der synaptischen Plastizität in synaptischen Unterbereichen gezeigt. Zum Beispiel sind mGlu7-Rezeptoren wichtig für die bidirektionale Plastizität an den Moosfasersynapsen, wobei die Richtungsänderung des Synapsengewichts durch den relativen Aktivierungs- und Expressionszustand des Rezeptors bestimmt wird (Pelkey et al., 2005). Im Gegensatz dazu bestimmen mGlu5-Rezeptoren die Richtungsänderung der synaptischen Stärke bei Moosfasersynapsen verglichen mit Kommisural-/Assoziationsfasern(AC)-CA3-Synapsen, wobei der Antagonismus von mGlu5-Rezeptoren die Moosfaser-LTP (Fig. 1) und die AC-CA3-LTD (Hagena und Manahan-Vaughan, 2015) beeinträchtigt. Zusammengefasst sollte die Bedeutung dieser Rezeptoren für die hippokampale Informationsenkodierung und die langfristige Speicherung räumlicher Lernerlebnisse nicht unterschätzt werden.
Danksagung: Wir danken Heide Brusis und Olga Neumann für ihre Unterstützung. Die Forschungsarbeiten der Autorin zu diesem Thema wurden von der Deutschen Forschungsgemeinschaft (DFG, MA 1843, SFB 874/B1, B3, B10) gefördert.
About the author

Denise Manahan-Vaughan ist Neurophysiologin, Neurowissenschaftlerin und Leiterin der Abteilung für Neurophysiologie an der Medizinischen Fakultät der Ruhr-Universität Bochum (www.rub.de/neurophys). Gleichzeitig ist sie auch Direktorin und Studiendekanin der International Graduate School of Neuroscience (www.rub.de/igsn) und Sprecherin des Sonderforschungsbereichs „Integration und Repräsentation Sensorischer Prozesse“ (www.rub.de/sfb874) an der Ruhr-Universität. Nach ihrem ersten Studienabschluss in Naturwissenschaften am Trinity College Dublin, Irland, wurde sie auch dort in Neuropharmakologie promoviert. Anschließend ging sie nach Deutschland an das Leibniz-Institut für Neurobiologie in Magdeburg. Nachdem sie ihre Habilitation in Physiologie an der Otto-von-Guericke-Universität Magdeburg erlangte, ging sie an das Johannes-Müller-Zentrum für Physiologie an der Charité, Berlin und im Jahr 2003 nach Bochum. Ihre Forschungsprojekte beschäftigen sich mit der Beziehung zwischen hippokampaler und kortikaler synaptischer Plastizität und Langzeit-assoziativer und räumlicher Gedächtnisbildung.
Literatur
André, M. E., Güntürkün, O. und Manahan-Vaughan, D. (2015). The metabotropic glutamate receptor, mGlu5 is required for extinction learning that occurs in the absence of a context change. Hippocampus. 25, 149–158.10.1002/hipo.22359Search in Google Scholar PubMed PubMed Central
Altinbilek, B. und Manahan-Vaughan, D. (2007). Antagonism of group III metabotropic glutamate receptors results in impairment of LTD but not LTP in the hippocampal CA1 region, and prevents long-term spatial memory. Eur. J. Neurosci. 26, 1166–1172.10.1111/j.1460-9568.2007.05742.xSearch in Google Scholar PubMed
Altinbilek, B. und Manahan-Vaughan, D. (2009). A specific role for group II metabotropic glutamate receptors in hippocampal long-term depression and spatial memory. Neuroscience 158, 149–158.10.1016/j.neuroscience.2008.07.045Search in Google Scholar PubMed
Balschun, D. und Wetzel, W. (2002). Inhibition of mGluR5 blocks hippocampal LTP in vivo and spatial learning in rats. Pharmacol. Biochem. Behav. 73, 375–380.10.1016/S0091-3057(02)00847-XSearch in Google Scholar PubMed
Bartos, M., Vida, I. und Jonas, P. (2007). Synaptic mechanisms of synchronized gamma oscillations in inhibitory interneuron networks. Nat. Rev. Neurosci. 8, 45–56.10.1038/nrn2044Search in Google Scholar PubMed
Bikbaev, A. und Manahan-Vaughan, D. (2007). Hippocampal network activity is transiently altered by induction of long-term potentiation in the dentate gyrus of freely behaving rats. 1, 1–7.10.3389/neuro.08.007.2007Search in Google Scholar PubMed PubMed Central
Bikbaev, A. und Manahan-Vaughan, D. (2008). Relationship of hippocampal theta and gamma oscillations to potentiation of synaptic transmission. Front. Neurosci. 2, 56–63.10.3389/neuro.01.010.2008Search in Google Scholar PubMed PubMed Central
Bikbaev, A., Neyman, S., Ngomba, R., Nicoletti, F., Conn, P.J und Manahan-Vaughan, D. (2008). The metabotropic glutamate receptor, mGluR5, mediates the functional interaction between late-LTP, hippocampal network activity, and learning by a mechanism involving regulation of mGluR1 expression. PloS One. 13, e2155.10.1371/annotation/58313075-ff2e-4268-9272-e942aed8d2f6Search in Google Scholar
Bikbaev, A. und Manahan-Vaughan, D. (2017). Metabotropic glutamate receptor, mGlu5, regulates hippocampal synaptic plasticity and is required for tetanisation-triggered changes in theta and gamma oscillations. Neuropharmacology. 115, 20–29.10.1016/j.neuropharm.2016.06.004Search in Google Scholar PubMed
Bliss, T.V. und Collingridge, G.L. (1993). A synaptic model of memory: long-term potentiation in the hippocampus. Nature 361, 31–39.10.1038/361031a0Search in Google Scholar PubMed
Buschler, A. und Manahan-Vaughan, D. (2017). Metabotropic glutamate receptor, mGlu5, mediates enhancements of hippocampal long-term potentiation after environmental enrichment. Neuropharmacology. 115, 42–50.10.1016/j.neuropharm.2016.06.003Search in Google Scholar PubMed
Bushell, T.J., Sansig, G., Collett, V.J., van der Putten, H. und Collingridge, G.L. (2002). Altered short-term synaptic plasticity in mice lacking the metabotropic glutamate receptor mGlu7. Scientific World Journal. 2, 730–737.10.1100/tsw.2002.146Search in Google Scholar PubMed PubMed Central
Buzsáki, G. (2005). Theta rhythm of navigation:link between path integration and landmark navigation, episodic and semantic memory. Hippocampus 15, 827–840.10.1002/hipo.20113Search in Google Scholar PubMed
Buzsaki, G. und Draguhn, A. (2004). Neuronal oscillations in cortical networks. Science. 304, 1926–1929.10.1126/science.1099745Search in Google Scholar PubMed
Corti, C., Aldegheri, L., Somogyi, P., Ferraguti, F (2002) Distribution and synaptic localisation of the metabotropic glutamate receptor 4 (mGluR4) in the rodent CNS. Neuroscience 110, 403e420.10.1016/S0306-4522(01)00591-7Search in Google Scholar PubMed
De Filippis B, Lyon L, Taylor A, Lane T, Burnet PW, Harrison PJ, Bannerman DM (2015) The role of group II metabotropic glutamate receptors in cognition and anxiety: comparative studies in GRM2(-/-), GRM3(-/-) and GRM2/3(-/-) knockout mice. Neuropharmacology. 89, 19–32.10.1016/j.neuropharm.2014.08.010Search in Google Scholar PubMed PubMed Central
Eichenbaum, H. (2017). On the Integration of Space, Time, and Memory. Neuron. 95, 1007–1018.10.1016/j.neuron.2017.06.036Search in Google Scholar PubMed PubMed Central
Ekstrom, A. D., Kahana, M.J., Caplan, J.B., Fields, T.A., Isham, E. A., Newman, E.L. und Fried, I. (2003). Cellular networks underlying human spatial navigation. Nature. 425, 184–188.10.1038/nature01964Search in Google Scholar PubMed
Gil-Sanz, C., Delgado-García, J.M., Fairén, A. und Gruart, A. (2008). Involvement of the mGluR1 receptor in hippocampal synaptic plasticity and associative learning in behaving mice. Cereb. Cortex 18, 1653–63.10.1093/cercor/bhm193Search in Google Scholar PubMed
Gogliotti, R.G., Senter, R.K., Fisher, N.M., Adams, J., Zamorano, R., Walker, A.G., Blobaum, A.L., Engers, D.W., Hopkins, C.R., Daniels, J.S., Jones, C.K., Lindsley, C.W., Xiang, Z., Conn, P.J. und Niswender, C.M. (2017). mGlu7 potentiation rescues cognitive, social, and respiratory phenotypes in a mouse model of Rett syndrome. Sci. Transl. Med. 9, eaai7459.10.1126/scitranslmed.aai7459Search in Google Scholar PubMed PubMed Central
Goh, J. und Manahan-Vaughan, D. (2013a). Endogenous hippocampal LTD that is enabled by spatial object recognition requires activation of NMDA receptors and the metabotropic glutamate receptor, mGlu5. Hippocampus. 23, 129–138.10.1002/hipo.22072Search in Google Scholar PubMed
Goh, J. und Manahan-Vaughan, D. (2013b). Endogenous hippocampal LTD that is enabled by spatial object recognition requires activation of NMDA receptors and the metabotropic glutamate receptor, mGlu5. Hippocampus. 23, 129–138.10.1002/hipo.22072Search in Google Scholar
Grieves, R.M. und Jeffery, K.J. (2017). The representation of space in the brain. Behav. Processes. 135, 113–131.10.1016/j.beproc.2016.12.012Search in Google Scholar PubMed
Hagena, H. und Manahan-Vaughan, D. (2015). mGlu5 acts as a switch for opposing forms of synaptic plasticity at mossy fiber-CA3 and commissural associational-CA3 synapses J. Neurosci. 35, 4999–5006.10.1523/JNEUROSCI.3417-14.2015Search in Google Scholar PubMed PubMed Central
Hagena, H. und Manahan-Vaughan, D. (2017). mGlu5: a metabotropic glutamate receptor at the hub of hippocampal information processing, persistent synaptic plasticity and long-term memory. In: mGlu Receptors. Ngomba, R.T., DiGiovanni, G., Battaglia, G., Nicoletti, F., eds. (Humana Press), pp 79–102. doi:10.1007/978-3-319-56170-7_5.10.1007/978-3-319-56170-7_5Search in Google Scholar
Hasselmo, M.E (2005). What is the function of hippocampal theta rhythm?—linking behavioral data to phasic properties of field potential and unit recording data. Hippocampus. 15, 936–949.10.1002/hipo.20116Search in Google Scholar PubMed
Hölscher, C., Anwyl, R. und Rowan, M. J. (1997). Block of theta-burst-induced long-term potentiation by (1S,3S)-1-aminocyclopentane-1,3-dicarboxylic acid: further evidence against long-term potentiation as a model for learning. Neuroscience 81, 17–22.10.1016/S0306-4522(97)00183-8Search in Google Scholar
Horner, A.J. und Doeller, C.F. (2017). Plasticity of hippocampal memories in humans. Curr. Opin. Neurobiol. 43,102–109.10.1016/j.conb.2017.02.004Search in Google Scholar PubMed PubMed Central
Huber, K.M., Roder, J.C. und Bear, M.F. (2001). Chemical induction of mGluR5- and protein synthesis--dependent long-term depression in hippocampal area CA1. J. Neurophysiol. 86, 321–325.10.1152/jn.2001.86.1.321Search in Google Scholar PubMed
Kemp, A. und Manahan-Vaughan, D. (2007). Hippocampal long-term depression: master or minion in declarative memory processes? Trends Neurosci. 30, 111–118.10.1016/j.tins.2007.01.002Search in Google Scholar PubMed
Kemp, A. und Manahan-Vaughan, D. (2008). The hippocampal CA1 region and dentate gyrus differentiate between environmental and spatial feature encoding through long-term depression. Cereb. Cortex. 18, 968–977.10.1093/cercor/bhm136Search in Google Scholar PubMed
Kentros, C., Hargreaves, E., Hawkins, R.D., Kandel, E.R., Shapiro, M. und Muller, R.V. (1998). Abolition of long-term stability of new hippocampalplace cell maps by NMDA receptor blockade. Science 280, 2121–2126.10.1126/science.280.5372.2121Search in Google Scholar PubMed
Klausnitzer, J., Kulla, A. und Manahan-Vaughan, D. (2004). Role of the group III metabotropic glutamate receptor in LTP, depotentiation and LTD in dentate gyrus of freely moving rats. Neuropharmacology. 46, 160–170.10.1016/j.neuropharm.2003.09.019Search in Google Scholar PubMed
Knierim, J.J., Kudrimoti, H.S. und McNaughton, B.L. (1995). Place cells, head direction cells, and the learning of landmark stability. J. Neurosci. 15, 1648–1659.10.1523/JNEUROSCI.15-03-01648.1995Search in Google Scholar PubMed PubMed Central
Kulla, A., Reymann K.G. und Manahan-Vaughan, D. (1999). Time-dependent induction of depotentiation in the dentate gyrus of freely moving rats: involvement of group 2 metabotropic glutamate receptors. Eur. J. Neurosci. 11, 3864–3872.10.1046/j.1460-9568.1999.00807.xSearch in Google Scholar PubMed
Lan, J.Y., Skeberdis, V. A., Jover, T., Zheng, X., Bennett, M.V. und Zukin, R.S. (2001). Activation of metabotropic glutamate receptor 1 accelerates NMDA receptor trafficking. J. Neurosci. 21, 6058–6068.10.1523/JNEUROSCI.21-16-06058.2001Search in Google Scholar PubMed PubMed Central
Lyon, L., Burnet, P.W., Kew, J.N., Corti, C., Rawlins, J.N., Lane, T., De Filippis, B., Harrison, P.J. und Bannerman, D.M. (2011). Fractionation of spatial memory in GRM2/3 (mGlu2/mGlu3) double knockout mice reveals a role for group II metabotropic glutamate receptors at the interface between arousal and cognition. Neuropsychopharmacology. 36, 2616–2628.10.1038/npp.2011.145Search in Google Scholar PubMed PubMed Central
Manahan-Vaughan, D. (1997). Group 1 and 2 metabotropic glutamate receptors play differential roles in hippocampal long-term depression and long-term potentiation in freely moving rats. J. Neurosci. 17, 3303–3311.10.1523/JNEUROSCI.17-09-03303.1997Search in Google Scholar PubMed PubMed Central
Manahan-Vaughan, D. (2017). Learning-Related Hippocampal Long-Term Potentiation and Long-Term Depression. In: Learning and Memory: A Comprehensive Reference (2nd Edition). Reference Module in Neuroscience and Biobehavioral Psychology, Elsevier, pp 585–609. http://dx.doi.org/10.1016/B978-0-12-809324-5.21104-8.10.1016/B978-0-12-809324-5.21104-8Search in Google Scholar
Manahan-Vaughan, D. und Braunewell, K.H. (2005). The metabotropic glutamate receptor, mGluR5, is a key determinant of good and bad spatial learning performance and hippocampal synaptic plasticity. Cereb. Cortex. 15, 1703–1713.10.1093/cercor/bhi047Search in Google Scholar PubMed
Mannaioni, G., Marino, M.J., Valenti, O., Traynelis, S.F. und Conn, P.J. (2001). Metabotropic glutamate receptors 1 and 5 differentially regulate CA1 pyramidal cell function. J. Neurosci. 21, 5925e5934.10.1523/JNEUROSCI.21-16-05925.2001Search in Google Scholar PubMed PubMed Central
Marszalek-Grabska, M., Gibula-Bruzda, E., Bodzon-Kulakowska, A., Suder, P., Gawel, K., Filarowska, J., Listos, J., Danysz, W. und Kotlinska, J.H. (2018). Effects of the Positive Allosteric Modulator of Metabotropic Glutamate Receptor 5, VU-29, on Impairment of Novel Object Recognition Induced by Acute Ethanol and Ethanol Withdrawal in Rats. Neurotox. Res. doi: 10.1007/s12640-017-9857-z.10.1007/s12640-017-9857-zSearch in Google Scholar PubMed PubMed Central
Martin, E.D. und Buno, W. (2005). Stabilizing effects of extracellular ATP on synaptic efficacy and plasticity in hippocampal pyramidal neurons. Eur. J. Neurosci. 21, 936–944.10.1111/j.1460-9568.2005.03925.xSearch in Google Scholar PubMed
Mukherjee. S. und Manahan-Vaughan, D. (2013). Role of metabotropic glutamate receptors in persistent forms of hippocampal plasticity and learning. Neuropharmacology 66, 65–81.10.1016/j.neuropharm.2012.06.005Search in Google Scholar PubMed
Naie, K. und Manahan-Vaughan, D. (2004). Regulation by metabotropic glutamate receptor 5 of LTP in the dentate gyrus of freely moving rats: relevance for learning and memory formation. Cereb. Cortex 14, 189–198.10.1093/cercor/bhg118Search in Google Scholar PubMed
Naie, K. und Manahan-Vaughan, D. (2005). Pharmacological antagonism of metabotropic glutamate receptor 1 (mGluR1) regulates LTP and spatial reference memory in the dentate gyrus of freely moving rats via NMDA and mGluR-dependent mechanisms. Eur. J. Neurosci. 21, 411–421.10.1111/j.1460-9568.2005.03864.xSearch in Google Scholar PubMed
Naie, K. und Manahan-Vaughan, D. (2006). Investigations of the protein synthesis dependency of mGluR-induced long-term depression in the dentate gyrus of freely moving rats. Neuropharmacology 49 (Suppl. 1), 35–44.10.1016/j.neuropharm.2005.06.001Search in Google Scholar PubMed
Nakao, K., Ikegaya, Y., Yamada, M.K., Nishiyama, N. und Matsuki, N. (2002). Hippocampal long-term depression as an index of spatial working memory. Eur. J. Neurosci. 16, 970–974.10.1046/j.1460-9568.2002.02159.xSearch in Google Scholar PubMed
Neyman, S. und Manahan-Vaughan, D. (2008). Metabotropic glutamate receptor 1 (mGluR1) and 5 (mGluR5) regulate late phases of LTP and LTD in the hippocampal CA1 region in vitro Eur. J. Neurosci. 27, 1345–1352.10.1111/j.1460-9568.2008.06109.xSearch in Google Scholar PubMed PubMed Central
Nomura, A., Shigemoto, R., Nakamura, Y., Okamoto, N., Mizuno, N. und Nakanishi, S. (1994). Developmentally regulated postsynaptic localization of a metabotropic glutamate receptor in rat rod bipolar cells. Cell 77, 361–369.10.1016/0092-8674(94)90151-1Search in Google Scholar PubMed
Ohishi, H., Shigemoto, R., Nakanishi, S. und Mizuno, N. (1993). Distribution of the messenger RNA for a metabotropic glutamate receptor, mGluR2, in the central nervous system of the rat. Neuroscience 53, 1009–1018.10.1016/0306-4522(93)90485-XSearch in Google Scholar
Okamoto, N., Hori, S., Akazawa, C., Hayashi, Y., Shigemoto, R., et al. (1994). Molecular characterization of a new metabotropic glutamate receptor mGluR7 coupled to inhibitory cyclic AMP signal transduction. J. Biol. Chem. 269, 1231–123610.1016/S0021-9258(17)42247-2Search in Google Scholar
O’Keefe, J. und Dostrovsky, J. (1971). The hippocampus as a spatial map. Preliminary evidence from unit activity in the freely-moving rat. Brain Res. 34, 171–175.10.1016/0006-8993(71)90358-1Search in Google Scholar PubMed
Petralia, R.S., Wang, Y.X., Niedzielski, A.S. und Wenthold, R.J. (1996). The metabotropic glutamate receptors, mGluR2 and mGluR3, show unique postsynaptic, presynaptic and glial localizations. Neuroscience. 71, 949–976.10.1016/0306-4522(95)00533-1Search in Google Scholar PubMed
Popkirov, S.G. und Manahan-Vaughan, D. (2011). Involvement of metabotropic glutamate receptor mGluR5 in learning facilitated-plasticity at CA1 synapses. Cereb. Cortex. 21, 501–519.10.1093/cercor/bhq093Search in Google Scholar PubMed PubMed Central
Shigemoto, R., Kinoshita, A., Wada, E., Nomura, S., Ohishi, H., Takada, M., Flor, P.J., Neki, A., Abe, T., Nakanishi, S. und Mizuno, N. (1997). Differential presynaptic localization of metabotropic glutamate receptor subtypes in the rat hippocampus. J. Neurosci. 17, 7503–7522.10.1523/JNEUROSCI.17-19-07503.1997Search in Google Scholar PubMed PubMed Central
Skeberdis, V. A., Lan, J., Opitz, T., Zheng, X., Bennett, M.V. und Zukin, R.S. (2001). mGluR1-mediated potentiation of NMDA receptors involves a rise in intracellular calcium and activation of protein kinase C. Neuropharmacology 40, 856–865.10.1016/S0028-3908(01)00005-3Search in Google Scholar PubMed
Ster, J., Mateos, J.M., Grewe, B.F., Coiret, G., Corti, C., Corsi, M., Helmchen, F. und Gerber, U. (2011). Enhancement of CA3 hippocampal network activity by activation of group II metabotropic glutamate receptors.Proc. Natl. Acad. Sci. U.S. A. 108, 9993–9997.10.1073/pnas.1100548108Search in Google Scholar PubMed PubMed Central
Szapiro, G., Barros, D.M., Ardenghi, P., Vianna, M.R., Choi, H., Silva, T., Medina, J.H. und Izquierdo, I. (2001). Facilitation and inhibition of retrieval in two aversive tasks in rats by intrahippocampal infusion of agonists of specific glutamate metabotropic receptor subtypes. Psychopharmacology (Berl). 156, 397–401.10.1007/s002130100755Search in Google Scholar PubMed
Tanabe, Y., Nomura, A., Masu, M., Shigemoto, R., Mizuno, N. und Nakanishi, S. (1993). Signal transduction, pharmacological properties, and expression patterns of two rat metabotropic glutamate receptors, mGluR3 and mGluR4. J. Neurosci. 13, 1372–1378.10.1523/JNEUROSCI.13-04-01372.1993Search in Google Scholar PubMed PubMed Central
Zarnadze, S., Bäuerle, P., Santos-Torres, J., Böhm, C., Schmitz, D., Geiger, J.R., Dugladze, T. und Gloveli, T. (2016). Cell-specific synaptic plasticity induced by network oscillations. eLife. 24, 5.10.7554/eLife.14912Search in Google Scholar PubMed PubMed Central
Zhang, S. und Manahan-Vaughan, D. (2014). Place field stability requires the metabotropic glutamate receptor, mGlu5. Hippocampus, 24, 1330–1340.10.1002/hipo.22314Search in Google Scholar PubMed PubMed Central
Zhang, S. und Manahan-Vaughan, D. (2015). Spatial olfactory learning facilitates place field formation in the hippocampus. Cereb. Cortex 25, 423–432.10.1093/cercor/bht239Search in Google Scholar PubMed PubMed Central
Zhou, R., Chen, F., Feng, X., Zhou, L., Li, Y. und Chen, L. (2015). Perinatal exposure to low-dose of bisphenol A causes anxiety-like alteration in adrenal axis regulation and behaviors of rat offspring: a potential role for metabotropic glutamate 2/3 receptors. J. Psychiatr. Res. 64, 121–129.10.1016/j.jpsychires.2015.02.018Search in Google Scholar PubMed
Anmerkung
Übersetzung der englischen Version des Artikels online verfügbar unter https://doi.org/10.1515/nf-2018-A007
© 2018 Walter de Gruyter GmbH, Berlin/Boston
Articles in the same Issue
- Titelseiten
- Übersichtsartikel
- Hippokampale Langzeitpotenzierung (LTP) – Geschichte, Gegenwart und Zukunft
- Hippocampal long-term potentiation (LTP) – past, present and future
- Langzeitpotenzierung im Hippokampus: Entdeckung, Mechanismen und Funktion
- Long-term potentiation in the hippocampus: discovery, mechanisms and function
- Die Regulation der hippokampalen Informationsenkodierung durch metabotrope Glutamatrezeptoren
- Regulation of hippocampal information encoding by metabotopic glutamate receptors
- Geclusterte Plastizität bei Langzeitpotenzierung: Wie starke Synapsen bestehen bleiben, um Langzeitgedächtnis aufrechtzuerhalten
- Clustered plasticity in Long-Term Potentiation: How strong synapses persist to maintain long-term memory
- Hippocampale synaptische Plastizität bei neurodegenerativen Erkrankungen: Aβ, Tau und darüber hinaus
- Hippocampal synaptic plasticity in neurodegenerative diseases: Aß, tau and beyond
- Die Analyse synaptischer Plastizität auf Einzelzellebene mit Hilfe der STDP
- Analyzing synaptic plasticity at the single cell level with STDP
- Institutsvorstellung
- Die Neurowissenschaftliche Gesellschaft ist Gründungsmitglied des German Brain Council
- DFG-Graduiertenkolleg 1957 „Adipocyte-Brain Crosstalk“
- Nachrichten
- Jugend forscht – Sonderpreis der Neurowissenschaftlichen Gesellschaft 2018
Articles in the same Issue
- Titelseiten
- Übersichtsartikel
- Hippokampale Langzeitpotenzierung (LTP) – Geschichte, Gegenwart und Zukunft
- Hippocampal long-term potentiation (LTP) – past, present and future
- Langzeitpotenzierung im Hippokampus: Entdeckung, Mechanismen und Funktion
- Long-term potentiation in the hippocampus: discovery, mechanisms and function
- Die Regulation der hippokampalen Informationsenkodierung durch metabotrope Glutamatrezeptoren
- Regulation of hippocampal information encoding by metabotopic glutamate receptors
- Geclusterte Plastizität bei Langzeitpotenzierung: Wie starke Synapsen bestehen bleiben, um Langzeitgedächtnis aufrechtzuerhalten
- Clustered plasticity in Long-Term Potentiation: How strong synapses persist to maintain long-term memory
- Hippocampale synaptische Plastizität bei neurodegenerativen Erkrankungen: Aβ, Tau und darüber hinaus
- Hippocampal synaptic plasticity in neurodegenerative diseases: Aß, tau and beyond
- Die Analyse synaptischer Plastizität auf Einzelzellebene mit Hilfe der STDP
- Analyzing synaptic plasticity at the single cell level with STDP
- Institutsvorstellung
- Die Neurowissenschaftliche Gesellschaft ist Gründungsmitglied des German Brain Council
- DFG-Graduiertenkolleg 1957 „Adipocyte-Brain Crosstalk“
- Nachrichten
- Jugend forscht – Sonderpreis der Neurowissenschaftlichen Gesellschaft 2018