Pulmonary intravascular macrophages: prime suspects as cellular mediators of porcine CARPA
-
Domokos Csukás
Domokos Csukás graduated as a veterinarian in 2005 at the Faculty of Veterinary Science, Szent István University Budapest, Hungary. After 7 years practice in small animal oriented veterinary hospitals he joined the team of the Department of Surgical Research and Techniques in the Faculty of Medicine of Semmelweis University, Budapest. He has special interests in laboratory animal care, animal model development and immune toxicological tests in the field of nanomedicine. He is working on his PhD thesis focusing on the side effects of nanodrugs, and the role of PIM cells in complement activation related pseudoallergy.

 ,
Rudolf Urbanics
,
Rudolf Urbanics
Rudolf Urbanics MD, PhD, Head of the
in vivo laboratory of Nanomedicine Research and Education Center of Semmelweis University, and SeroScience Ltd., an immunotoxicity CRO, since 2008 in Budapest, Hungary. He obtained MD diploma and the PhD degree at Semmelweis Medical School, Budapest, Hungary. He had various research/collaboration positions at MaxPlanck Institute of Systemphysiology, at University of Pennsylvania, Cerebrovascular Research Center, at Pennsylvania Muscle Institute, in the Knoll AG, working in the field of CNS regulation of blood flow/metabolism, ischemic/hypoxic disorders, stroke and chronic neurodegenerative disease animal models. Urbanics was the Deputy R&D Director and Head of CNS Pharmacology Department at Biorex R&D Co., worked at IVAX/Drug Research Institute Budapest, as Leading researcher in Safety and CNS Pharmacology and later in IVAX/Drug Research Institute, Subsidiary of TEVA as Head of In Vivo Pharmacology Group. Currently, he is working with in vivo models of nano drug - nano carrier induced, complement activation related pseudoallergic reactions (CARPA), clarifying their immune-toxicological and safety hazards.György Wéber received his PhD in medicine in 1992 from University of Pécs, Hungary. He was appointed to professor of surgery in 2003. He is a qualified general and vascular surgeon. In 2010 he founded the Department of Surgical Research and Techniques, Medical Faculty, Semmelweis University, Budapest, Hungary and since then he is the chair of this department. His research is mainly dedicated to nanotechnology and scaffolding in tissue engineering (mesh development for hernia repair etc).
László Rosivall MD, PhD, DSc, Széchenyi and Khwarizmi prizes laureate, the head of International Nephrology Research and Training Center, Department of Pathophysiology, and PhD School of Basic Medical Sciences, Semmelweis University. He pioneered recognizing and characterizing intrarenal renin-angiotensin system. For the first time, he demonstrated the existence of fenestration, nanochannels in endothelium of distal portion of renal afferent arteriole (AA). Using nanotechnology he visualized in vivo the GFR and demonstrated a new mechanism in its regulation. This unique JGA morphology and the high filtration volume in AA is one of the most striking recent observations of renal microcirculation.
Janos Szebeni MD, PhD, DSc, Med. Habil., immunologist, director of the Nanomedicine Research and Education Center at Semmelweis University, Hungary. He is also founder and CEO of a contract research SME “SeroScience”, and full professor of (immune) biology at Miskolc University. He has held various guest professor and scientific positions in Hungary and abroad, mostly in the USA where he lived for 22 years. His research on various themes in hematology, membrane biology and immunology resulted >120 scientific papers (citations: ;4550, H index: 35), 14 book chapters, 2 granted patents, a book entitled “The Complement System: Novel Roles in Health and Disease” (Kluwer, 2004). Three fields stand out where he has been most active: artificial blood, liposomes and the complement system. His original works led to the “CARPA” concept, i.e., that complement activation underlies numerous drug-induced (pseudo)allergic (anaphylactoid) reactions.

Abstract
Pigs provide a highly sensitive and quantitative in vivo model for complement (C) activation-related pseudoallergy (CARPA), a hypersensitivity reaction caused by some state-of-art nanomedicines. In an effort to understand the mechanism of the pigs’ unique sensitivity for CARPA, this review focuses on pulmonary intravascular macrophages (PIMs), which are abundantly present in the lung of pigs. These cells represent a macrophage subpopulation whose unique qualities explain the characteristic symptoms of CARPA in this species, most importantly the rapidly (within minutes) developing pulmonary vasoconstriction, leading to elevation of pulmonary arterial pressure. The unique qualities of PIM cells include the following; 1) they are strongly adhered to the capillary walls via desmosome-like intercellular adhesion plaques, which secure stable and lasting direct exposition of the bulk of these cells to the blood stream; 2) their ruffled surface engaged in intense phagocytic activity ensures efficient binding and phagocytosis of nanoparticles; 3) PIM cells express anaphylatoxin receptors, this way C activation can trigger these cells, 4) they also express pattern recognition molecules on their surface, whose engagement with certain coated nanoparticles may also activate these cells or act in synergy with anaphylatoxins and, finally 5) their high metabolic activity and capability for immediate secretion of vasoactive mediators upon stimulation explain the circulatory blockage and other robust physiological effects that their stimulation may cause. These qualities taken together with reports on liposome uptake by PIM cells during CARPA and the possible presence of these cells in human lung suggests that PIM cells may be a potential therapeutic target against CARPA.
Introduction: the porcine model of CARPA
Complement activation-related pseudoallergy (CARPA) is a common adverse immune effect of i.v. administered drugs and agents, a subclass of type I hypersensitivity reactions (HSRs) which is not mediated by IgE. CARPA can be caused by a variety of i.v.-administered nanomedicines and antibody-based therapeutic or diagnostic agents, including liposomal drugs, antibodies, polymers, etc. (1, 2). The reaction is usually mild and reversible, but in a small percentage of patients the symptoms are severe or even lethal, which lends substantial clinical and regulatory significance to its study in animal models. A subject of previous reviews, CARPA can be induced in many animal species, among which pigs stand out as being the most appropriate species for mimicking the symptoms and other conditions of the reaction in hypersensitive man (3). Accordingly, the porcine CARPA model has been used in many previous studies on liposome and other nanoparticle-induced CARPA wherein the methods applied, the symptoms and their interrelations and many other details of the reaction have been amply described (4–8). One of our previous reviews (3) in this volume highlights the variation of cardiovascular symptoms in porcine CARPA, while the present review focuses solely on the likely role of a particular cell in the lung of pigs: pulmonary intravascular macrophages, or PIM cells. The goal is to summarize the information on PIM cells that point to their central role in CARPA. Since PIM cells can also occur in humans, the review highlights the possibility that their selective suppression might be an effective approach to prevent CARPA.
PIM cells in general
PIMs are resident cells in the pulmonary capillaries of animals in the Artiodactyla order, such as the pig, sheep, cattle, in the horse of the Perissodactyla order, and also in cats (9). They represent a part of the mononuclear phagocyte system (MPS) in the lung of these animals. Just as Kupffer cells in the liver, they play an important role in the removal of blood-born materials, cellular debris, immune cells, bacteria, viruses, endotoxins, etc. The appearance of these cells varies in different species and their origin and life cycle are debated. One theory holds that they differentiate from circulating peripheral blood mononuclear cells in situ, while others believe that they are immigrants from the neighboring interstitium. In some species PIMs appear as a constitutive member of the lung’s MPS, with colonization starting soon after birth (10, 11).
In species where PIMs are not found at birth, their appearance in the lung can be induced by different effects, for example stimulation with lipopolysaccharides (LPS) or infectious agents, or by changes in the general condition of the organism, such as the presence of hepatic cirrhosis, tumors, hematologic diseases or experimental ligation of the bile duct (12, 13). Pulmonary induction of PIM cells in the lung was described in rats, mice, hamsters, rabbits, chicken, dogs and macaques (14–16).
The species that lack PIMs show no pulmonary hypertension in response to CARPAgenic triggers, or need three orders higher doses to indicate the symptoms (17).
The amount and size of PIM cells is in the d=20–80 μm range show wide differences among the species, however, the numbers are difficult to compare directly because of the inconsistent specifications. For example, the amount of PIM cells in the lung of sheep, being the highest, was given as 20% of the endothelial surface, while in pigs it was specified as 14×103/mm3 of lung parenchyma. Horse PIMs are the largest in size (18–20).
Morphology
Figure 1 illustrates the most prominent features of PIM cells; rounded shape, ruffled membrane, presence of pseudopods and adhesion onto the surface.
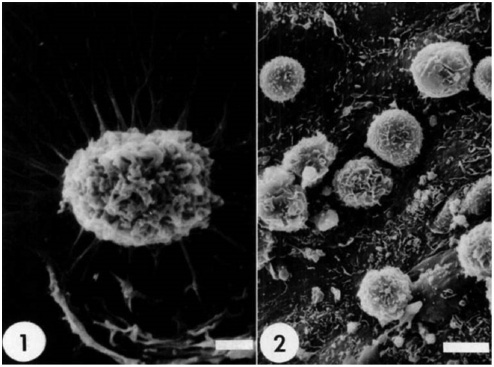
Porcine Pulmonary Intravascular Macrophages (PIM cells).
1) PIMs reaching out with pseudopods attached to the surface of plastic coverslip after 1 h incubation (sizebar 2 μm); 2) PIMs showing rounded shape, ruffled membrane. Picture taken after 8 h incubation on the pulmonary artery endothelium ex vivo, sizebar 5 μm. Modified from Ref. (21) with permission.
Other characteristic features include indented nucleus and a unique glycocalyx embedded in the plasma membrane via lipid and lipoprotein foothills of 50–200 nm is size. This surface plays a key role in receptor–mediated endocytosis or phagocytosis of blood-born materials, endotoxins, tracer particles, liposomes (22–25).
Adherence
PIMs have a significant ability for adherence, which is one of the most important characteristics of these cells. They firmly attach to the capillary endothelium via junction-like intercellular adhesion plaques (ICAPs), as demonstrated for the case of a sheep PIM cells adhered to the pulmonary capillary endothelium (Figure 2). In addition to pulmonary capillaries, in vitro PIM cells adhere via adhesion plaques to pulmonary artery and corneal and aorta segments (21). The intercellular space between the PIM cell membrane and adherence surface is 15–20 nm and electron dense material is present on both sides of the cell membranes with a width of about 30 nm (26).
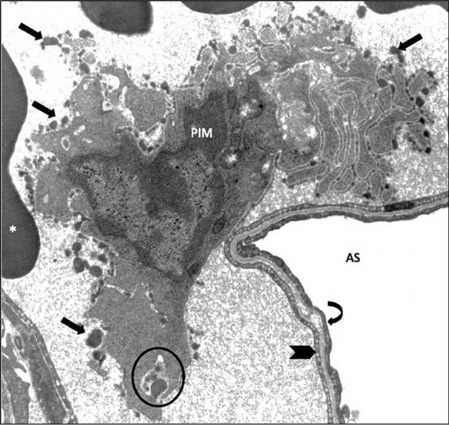
Pulmonary Intravascular Macrophage (PIM) of a sheep, anchored to the pulmonary capillary endothelium.
Unique ruffled globular membrane structure (arrows) is present. AS, alveolar space; *, red blood cells; circle, phagocytosis; thick arrow, capillary wall; curved arrow, aveolar epithelium; magnification, X10,000. Modified from Ref. (25).
As shown in Figure 3, the strong adherence of PIM cells enables their in vitro separation.
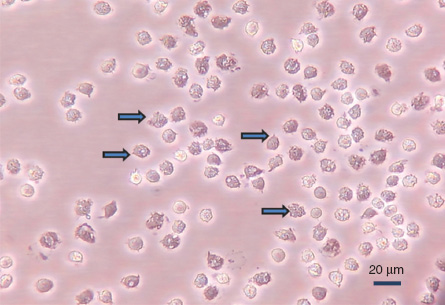
Porcine PIM cells adhered to plastic surface.
The dens, larger cells (marked by arrows) show ruffled membrane structures and multiple vacuoles. Cells were washed out from the capillaries by collagenase and let to adhere to gelatin surface for 30 min at 37°C, essentially as described in Ref. (21). Magnification: 40×, unpublished data.
Function
The primary function of PIM cells is the clearance of infectious or deleterious agents from the blood, such a bacteria, virus or nanoparticles that are recognized as foreign material. Kupffer cells in the liver have similar function, and in different species one or the other cell type dominates. As mentioned, pulmonary PIM cells dominate in pigs, sheep, cattle, horse and cats (9), while hepatic Kupffer cells dominate in rats, rabbit and mice (27, 28). Key difference between PIMs and Kupffer cells is that PIMs response to particle injection with phagocytosis and secretion of vasoconstrictor substances and other mediators (see below), while liver Kupffer cells have huge retention capacity without such secretory capability (27, 28). PIMs have high affinity to endotoxin which can lead to PIM induction, respiratory symptoms, immune reactions, even serious inflammation. Such activation can arise from external and internal LPS, originating in different organs of the body (e.g., intestines).
Phagocytosis
PIM cells are capable for highly effective phagocytosis, for example erythrocytes, fibrin and other extracellular matrix components (29). Constitutive PIMs, like those in the pig, sheep, cattle and horse showed similar kinetics of phagocytic activity (30). Post-phagocytic retention of intravenously administered particles (colloid gold, iron oxide, liposomes, etc.) by PIM cells in the lung in pigs, horses and ruminants is substantial (>40%), while rodent PIM cells had <10% of such activity. Liposome retention by pig PIM cells was found to be over 60%. In swine and ruminants the amount of PIMs and the functional activity of phagocytosis show strong correlation (31–33). Several publications show that PIMs are also able to take up viral particles (e.g., hog cholera, African swine fever) during inflammation (34–36).
Receptors
Table 1 shows the receptors on the surface of PIM cells along with their molecular type and function. These receptors serve the adhesion and secretory function of PIM cells. From the point of view of CARPA the C5aR is especially important, since it is very likely that it plays a key role in channeling the C activation signal to physiological changes during CARPA. Our literature search has not found direct demonstration of C5aR on PIM cells, but the indirect evidence is overwhelming. It includes the omnipresence of C5aR on monocyte/macrophage line cells and multiple demonstration of C5aR on pulmonary alveolar macrophages (36, 45–49). As mentioned, CARPA represents an anaphylactic reaction, a phenomenon whose name reveals its relation to anaphylatoxins.
Receptors on PIM cells.
| Receptor Type | Structure | Function | References |
|---|---|---|---|
| P Integrins | glycosilated- dimeric proteins | – Cell adherence – Intercellular communication – Cell signaling – Regulation of cell shape and motility | (37) |
| Toll-like receptors (TLR 1–9) | dimeric protein | Recognition of – Pathogens – LPS – Lipoproteins – Lipopeptides – β-glucan (Zymosan) | (38–40) |
| Fc-receptor (FcR) | transmembrane tyrosine kinase | Binding of – IgG, – IgA – IgE – Opsonins | (41, 42) |
| Complement-receptors CR1 (CD35) CR3, CR4 | single-chain membrane glycoproteins | CR1 – binds C3b, C4b, – binds immune complexes – Mediates adherence and phagocytosis, – Inhibits C activation via the classic and alternative pathways CR3 (CD11b/18) – binds iC3b – promotes adhesion to the vascular endothelium – binds β-glucan (Zymosan) – C3dg and C3d bind iC3b with low – CR4 receptor binds iC3b – CR3 (CD11b/18) and CR4 (CD11c/18) are both bind to iC3b. CR3 binds iC3b with high and C3dg and C3d with low affinity. – The CR4 receptor binds iC3b but not β-glucan | (43, 44) |
| C5aR (C5R1, CD88)* | Membrane spanning | Binding of anaphylatoxin C5a | (36, 45–49) |
Secretory products and mediators
Following stimulation, PIM cells can secrete a large number of vasoactive and/or inflammatory mediators, listed in Table 2.
PIM cell-derived secretory products and mediators.
| Mediator type | Structure | Function | References |
|---|---|---|---|
| Thromboxane A2 | Eicosanoid | Pulmonary vasoconstriction | (50, 51) |
| Leukotrienes | (52) | ||
| TNF-alpha | Soluble glycoproteins | proinflammatory, activation of neutrophils | (38, 53–57) |
| IL-1beta | proinflammatory, activation of neutrophils | ||
| IL-6 | Proinflammation, leukocyte activation, | ||
| IL-8 | promotion of recruitment of Platelets, and heterophils | ||
| Platelet-derived growth factor (PDGF) | induces cell proliferation, migration and angiogenesis | ||
| Vascular endothelial growth factor (VEGF) | angiogenesis, vascular permeability increase | ||
| Reactive oxygen apecies | oxidative tissue injury, vascular tone mediator | (57) | |
Among the secretory products, thromboxane A2 (TXA2) deserves special attention, as it plays a pivotal role in the hemodynamic changes that we see and measure in the porcine CARPA model. The vasoconstrictive and consequent hypertensive effect of TXA2 has been known for long (58, 59), and the causality between these phenomena obtains spectacular demonstration in the porcine CARPA model inasmuch as the rise of PAP closely follows the rise of TXB2 level in blood (TXB2 is the stable metabolite of TXA2) on the second-to minute scale, with only a few second delay (Figure 4). In addition, indomethacin, an inhibitor of cyclooxygenase which produces TXA2, completely inhibits the rise of PAP and other hemodynamic changes (4), which effect offers a possible therapeutic intervention with CARPA. It should also be noted regarding the efficacy of indomethacin that TXA2 secretion and pulmonary vasoconstriction starts in the lung of pigs within 1–3 min after i.v. administration of liposomes or other C-activating nanoparticles (3). Since indomethacin is an effective inhibitor of CARPA even when it is administered only minutes before triggering the reaction (4), TXA2 must be formed by cyclooxygenase in a very fast reaction during CARPA, rather than being stored in preformed intracellular vesicles and released upon PIM cell activation.
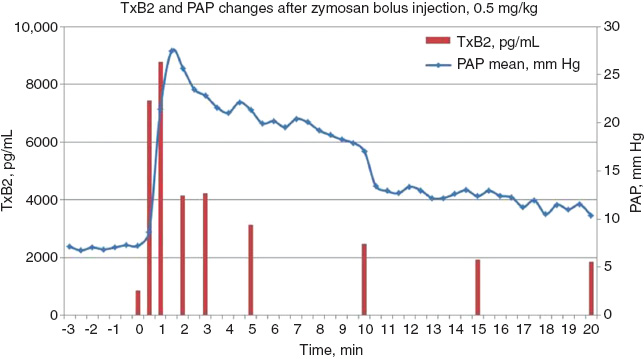
Time correlation between the rises of blood TXB2 and pulmonary arterial pressure during zymosan-induced CARPA in a pig (reaction following 0.5 mg/kg Zymosan i.v. injection).
Zymosan (0.5 mg/kg) was injected i.v. at time 0. At 30 s after the injection TXB2 level exceeds 9× the preinjection value, and clearly precedes the rise of PAP. Original data reproducing a similar experiment wherein the reaction was triggered by liposomes.
In addition to TXB2, PIMs secrete cytokines, oxygen radicals and proteolytic enzymes which contribute to local tissue injury and delayed cardiovascular effects in CARPA and/or inflammation.
Methodical aspects of experimentation with PIMs
Table 3 lists the experimental methods applied for addressing various questions relating to PIM cell structure and function.
Methods applied in PIM cell research.
| Research area | Method | References |
|---|---|---|
| Phagocytic features | tracers used: – monastral blue – radiolabeled molecules – gold colloids – magnetic iron oxide – surfactants – fluorescent molecules | (20, 60) |
| Physiologic function | Depletion by – gadolinium chloride – liposomal clodronate | (49, 61–63) |
| Isolation/culturing | Detachment from endothel by – magnetic field treatment after forced phagocytosis of iron oxide – Enzymatic digestion (collagenase, DNAse) | (21, 32, 64, 65) |
Facts pointing to a causal role of PIM cell stimulation in CARPA
There are many facts and considerations that suggest that PIM cells play a causal role in porcine CARPA. These include the following;
The initial eicosanoid (TXB2) secretion and pulmonary hypertension closely coincide (5);
The kinetics of particle uptake by PIM cells is also in the same time course (in minutes) as the development of pulmonary and other hemodynamic changes (29, 66);
The species that lack PIMs (or have only induced PIMs under normal conditions, i.e., rats) show systemic hemodynamic changes after i.v. bolus injections of reactogenic drugs (Zymosan, Ambisome) only at 2–3 orders of magnitude higher doses than the reactogenic trigger dose in pigs (17);
Depletion of PIM cells eliminates most of the acute abnormal pulmonary hemodynamic changes in endotoxin shock (63) or hyperacute pulmorary xenograft rejection in pigs (49). PIM depletion in sheep led to the loss of pulmonary vaso-responsiveness, whose return coincided with PIM repopulation (67). PIM depletion also attenuated capillary leakage, preserved low pulmonary vascular resistance and decreased the production of thromboxane A2 (49);
In miniature pig reactogenic liposomes caused CARPA and, at the same time, the authors could detect and visualize the presence of liposomes in PIM cells (66).
The double hit theory of CARPA
Porcine CARPA has a feature that is difficult to explain: in the case of certain drugs, only the first dose causes HSR; the second, or third similar, or even greater doses remain ineffective. The phenomen represents self-induced tolerance, or tachyphylaxis. It is observed with PEGylated small unilamellar lispomes, such as Doxil®, certain polymers, but not with higly charged AmBisome, or large multilamellar liposomes. The clinical significance of tachyphylaxis lies in the possibility to tolerize patients against HSR by slow initial administration of the drug in a way the first reaction remains subclinical, and then administering the rest of the dose without adverse event. In fact, the well-known clinical success of low reactogenic administration protocols, applied for antibody-based pharmaceuticals (68–71) and some liposomal drugs, e.g., Doxil® (72) is likely to rests on this principle.
Possible explanations for the phenomenon include the depletion of a reaction meditor (such as natural antibodies), or saturation of a reaction-mediating process (such as C activation or cellular uptake of liposomes) and the “double hit” hypothesis (Figure 5).
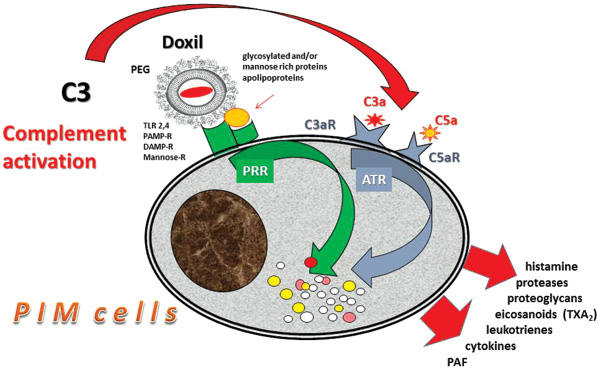
Scheme of the “double hit” hypothesis, explaining tachyphylactic CARPA.
PIM cells are activated both via the C3a/C5a receptor (anaphylatoxin receptors, ATr), and via patter recognizing receptor (PRr)-mediated uptake of Doxil®, illustrating liposomes or other coated nanoparticles that are taken up by these cells. In case of “double hit” simultaneous binding occurs on both receptor types, leading to additive or synergetic stimulation of the cell that entails mediator release and, ultimately, HSR.
According to the “double hit” hypothesis, tachyphylactic CARPA is not only C-activation, and, hence C5aR-dependent, but also depends on cellular uptake of the reactogenic drug by PIM cells, via one or more of their surface receptors (e.g., Fc receptor, Toll-like receptor, or similar pattern recognition receptors). It is only simultaneous occurance of these processes that can trigger PIM cells for release reaction, and if one activation step is too weak or missing, the reaction will fade and then disappear. It is assumed that in the case of tachyphylactic HSRs one or the other activation channel is irreversible, or get downregulated, explaining the lack of reaction upon the second and later repetitive treatments. In case of non-tachyphylactic reactions full secretory response can be achieved from PIMs either by a strong, supra-threshold stimulus of ATr and/or on PRr receptors, or by simultaneous sub-threshold stimuli on both receptor fields. It leads to tachyphylaxis when the stimulus is way under the effective sub-threshold level. The theory, illustrated in Figure 5, equally applies to mast cells and PIM cells (3). Future studies will hopefully reveal more details about the involvement of PIM cells in CARPA and the mechanism of tachyphylaxis.
Outlook with speculations on the mechanism of human CARPA
Man and nonhuman primates lack constitutive PIMs, although in baboons an increased phagocytic activity is shown in the lung by mononuclear cells (15). Macrophages can, nevertheless, accumulate in the intravascular space in human lungs, for example in the so-called hepatopulmonary syndrome, a common complication of hepatic cirrhosis (25, 56, 73). The mechanism and conditions of de novo PIM cell colonization in human lung is still unknown, and in lack of contrary evidence, it is not excluded that a low percentage of healthy people also host PIM cells in their lung. A further speculation might be that the low percentage of man who develops severe CARPA carry somewhere in their circulation intravascular macrophages (perhaps Kupffer cells) which like PIM cells, immediately respond to exposure to particles and/or anaphylatoxins with intense secretion of allergy mediators into the blood. The bottom line is that the study of the role of PIM cells or other macrophages in CARPA will hopefully shed more light on the pathogenesis of this adverse immune reaction to state-of-art (nano)medicines.
About the authors

Domokos Csukás graduated as a veterinarian in 2005 at the Faculty of Veterinary Science, Szent István University Budapest, Hungary. After 7 years practice in small animal oriented veterinary hospitals he joined the team of the Department of Surgical Research and Techniques in the Faculty of Medicine of Semmelweis University, Budapest. He has special interests in laboratory animal care, animal model development and immune toxicological tests in the field of nanomedicine. He is working on his PhD thesis focusing on the side effects of nanodrugs, and the role of PIM cells in complement activation related pseudoallergy.

Rudolf Urbanics MD, PhD, Head of the in vivo laboratory of Nanomedicine Research and Education Center of Semmelweis University, and SeroScience Ltd., an immunotoxicity CRO, since 2008 in Budapest, Hungary. He obtained MD diploma and the PhD degree at Semmelweis Medical School, Budapest, Hungary. He had various research/collaboration positions at MaxPlanck Institute of Systemphysiology, at University of Pennsylvania, Cerebrovascular Research Center, at Pennsylvania Muscle Institute, in the Knoll AG, working in the field of CNS regulation of blood flow/metabolism, ischemic/hypoxic disorders, stroke and chronic neurodegenerative disease animal models. Urbanics was the Deputy R&D Director and Head of CNS Pharmacology Department at Biorex R&D Co., worked at IVAX/Drug Research Institute Budapest, as Leading researcher in Safety and CNS Pharmacology and later in IVAX/Drug Research Institute, Subsidiary of TEVA as Head of In Vivo Pharmacology Group. Currently, he is working with in vivo models of nano drug - nano carrier induced, complement activation related pseudoallergic reactions (CARPA), clarifying their immune-toxicological and safety hazards.

György Wéber received his PhD in medicine in 1992 from University of Pécs, Hungary. He was appointed to professor of surgery in 2003. He is a qualified general and vascular surgeon. In 2010 he founded the Department of Surgical Research and Techniques, Medical Faculty, Semmelweis University, Budapest, Hungary and since then he is the chair of this department. His research is mainly dedicated to nanotechnology and scaffolding in tissue engineering (mesh development for hernia repair etc).

László Rosivall MD, PhD, DSc, Széchenyi and Khwarizmi prizes laureate, the head of International Nephrology Research and Training Center, Department of Pathophysiology, and PhD School of Basic Medical Sciences, Semmelweis University. He pioneered recognizing and characterizing intrarenal renin-angiotensin system. For the first time, he demonstrated the existence of fenestration, nanochannels in endothelium of distal portion of renal afferent arteriole (AA). Using nanotechnology he visualized in vivo the GFR and demonstrated a new mechanism in its regulation. This unique JGA morphology and the high filtration volume in AA is one of the most striking recent observations of renal microcirculation.

Janos Szebeni MD, PhD, DSc, Med. Habil., immunologist, director of the Nanomedicine Research and Education Center at Semmelweis University, Hungary. He is also founder and CEO of a contract research SME “SeroScience”, and full professor of (immune) biology at Miskolc University. He has held various guest professor and scientific positions in Hungary and abroad, mostly in the USA where he lived for 22 years. His research on various themes in hematology, membrane biology and immunology resulted >120 scientific papers (citations: ;4550, H index: 35), 14 book chapters, 2 granted patents, a book entitled “The Complement System: Novel Roles in Health and Disease” (Kluwer, 2004). Three fields stand out where he has been most active: artificial blood, liposomes and the complement system. His original works led to the “CARPA” concept, i.e., that complement activation underlies numerous drug-induced (pseudo)allergic (anaphylactoid) reactions.
References
1. Szebeni J. Complement activation-related pseudoallergy: a new class of drug-induced immune toxicity. Toxicology 2005;216: 106–21.10.1016/j.tox.2005.07.023Suche in Google Scholar
2. Szebeni J. Complement activation-related pseudoallergy: a stress reaction in blood triggered by nanomedicines and biologicals. Mol Immunol 2014;61:163–73.10.1016/j.molimm.2014.06.038Suche in Google Scholar
3. Szebeni J, Bedocs P, Csukas D, Rosivall L, Bunger R, Urbanics R. A porcine model of complement-mediated infusion reactions to drug carrier nanosystems and other medicines. Adv Drug Deliver Rev 2012;64:1706–16.10.1016/j.addr.2012.07.005Suche in Google Scholar
4. Szebeni J, Fontana JL, Wassef NM, Mongan PD, Morse DS, Dobbins DE, et al. Hemodynamic changes induced by liposomes and liposome-encapsulated hemoglobin in pigs: a model for pseudoallergic cardiopulmonary reactions to liposomes. Role of complement and inhibition by soluble CR1 and anti-C5a antibody. Circulation 1999;99:2302–9.10.1161/01.CIR.99.17.2302Suche in Google Scholar
5. Szebeni J, Baranyi L, Savay S, Bodo M, Morse DS, Basta M, et al. Liposome-induced pulmonary hypertension: properties and mechanism of a complement-mediated pseudoallergic reaction. Am J Physiol-Heart C 2000;279:H1319–28.10.1152/ajpheart.2000.279.3.H1319Suche in Google Scholar
6. Szebeni J, Baranyi L, Savay S, Bodo M, Milosevits J, Alving CR, et al. Complement activation-related cardiac anaphylaxis in pigs: role of C5a anaphylatoxin and adenosine in liposome-induced abnormalities in ECG and heart function. Am J PhysiolHeart C 2006;290:H1050–8.10.1152/ajpheart.00622.2005Suche in Google Scholar
7. Szebeni J, Bedőcs P, Urbanics R, Bunger R, Rosivall L, Tóth M, et al. Prevention of infusion reactions to PEGylated liposomal doxorubicin via tachyphylaxis induction by placebo vesicles: a porcine model. J Contr Rel 2012;160:382–7.10.1016/j.jconrel.2012.02.029Suche in Google Scholar
8. Szebeni J, Bedőcs P, Rozsnyay Z, Weiszhár Z, Urbanics R, Rosivall L, et al. Liposome-induced complement activation and related cardiopulmonary distress in pigs: factors promoting reactogenicity of Doxil and AmBisome. Nanomedicine NBM 2012;8:176–84.10.1016/j.nano.2011.06.003Suche in Google Scholar
9. Brain JD, Molina RM, DeCamp MM, Warner AE. Pulmonary intravascular macrophages: their contribution to the mononuclear phagocyte system in 13 species. Am J Physiol Lung Cell Mol Physiol 1999;276:146–54.10.1152/ajplung.1999.276.1.L146Suche in Google Scholar
10. Winkler GC, Cheville NF. Postnatal colonization of porcine lung capillaries by intravascular macrophages: an ultrastructural, morphometric analysis. Microvasc Res 1987;33:224–32.10.1016/0026-2862(87)90019-7Suche in Google Scholar
11. Longworth KE, Albertine KH, Staub NC. Ultrastructural quantification of pulmonary intravascular macrophages in newborn and 2-week-old lambs. Anat Rec 1996;246:238–44.10.1002/(SICI)1097-0185(199610)246:2<238::AID-AR11>3.0.CO;2-PSuche in Google Scholar
12. Chang SW, Ohara N. Chronic biliary obstruction induces pulmonary intravascular phagocytosis and endotoxin sensitivity in rats. J Clin Invest 1994;94:2009–19.10.1172/JCI117554Suche in Google Scholar
13. Singh B, Doane KJ, Niehaus GD. Ultrastructural and cytochemical evaluation of sepsis-induced changes in the rat pulmonary intravascular mononuclear phagocytes. J Anat 1998;192 (Pt 1):13–23.10.1046/j.1469-7580.1998.19210013.xSuche in Google Scholar
14. Warner AE, Brain JD. The cell biology and pathogenic role of pulmonary intravascular macrophages. Am J Physiol 1990;258 (2 Pt 1):L1–12.10.1152/ajplung.1990.258.2.L1Suche in Google Scholar
15. Staub NC. Pulmonary intravascular macrophages. Annu Rev Physiol 1994;56:47–67.10.1146/annurev.ph.56.030194.000403Suche in Google Scholar
16. Miot-Noirault E, Faure L, Guichard Y, Montharu J, Le Pape A. Scintigraphic in vivo assessment of the development of pulmonary intravascular macrophages in liver disease: experimental study in rats with biliary cirrhosis. Chest 2001;120:941–7.10.1378/chest.120.3.941Suche in Google Scholar
17. Dzsi L, Fülöp T, Mészáros T, Szénási G, Urbanics R, Vázsonyi C, et al. Features of complement activation-related pseudoallergy to liposomes with different surface charge and pegylation: comparison of the porcine and rat responses. J Control Release 2014;165:2–10.10.1016/j.jconrel.2014.08.009Suche in Google Scholar
18. Warner AE, Barry BE, Brain JD. Pulmonary intravascular macrophages in sheep. Morphology and function of a novel constituent of the mononuclear phagocyte system. Lab Invest J Tech Method Pathol 1986;55:276–88.Suche in Google Scholar
19. Winkler GC. Pulmonary intravascular macrophages in domestic animal species: review of structural and functional properties. Am J Anat 1988;181:217–34.10.1002/aja.1001810302Suche in Google Scholar
20. Parbhakar OP, Duke T, Townsend HG, Singh B. Immunophenotypic characterization and depletion of pulmonary intravascular macrophages of horses. Vet Res 2004;35:39–51.10.1051/vetres:2003041Suche in Google Scholar
21. Morton D, Bertram TA. Isolation and preliminary in vitro characterization of the porcine pulmonary intravascular macrophage. J Leukocyte Biol 1988;43:403–10.10.1002/jlb.43.5.403Suche in Google Scholar
22. Atwal OS, Singh B, Staempfli H, Minhas K. Presence of pulmonary intravascular macrophages in the equine lung: some structuro-functional properties. Anat Rec 1992;234:530–40.10.1002/ar.1092340408Suche in Google Scholar
23. Singh B, Ireland WP, Minhas K, Atwal OS. Surface coat of sheep pulmonary intravascular macrophages: reconstitution, and implication of a glycosyl-phosphatidylinositol anchor. Anat Rec 1995;243:466–78.10.1002/ar.1092430409Suche in Google Scholar
24. Atwal OS, Minhas KJ, Williams CS. Ultrastructural response of pulmonary intravascular macrophages to exogenous oestrogen in the bovine lung: translocation of the surface-coat and enhanced cell membrane plasticity and angiogenesis. J Anat 2001;198(Pt 5):611–24.10.1046/j.1469-7580.2001.19850611.xSuche in Google Scholar
25. Schneberger D, Aharonson-Raz K, Singh B. Pulmonary intravascular macrophages and lung health: what are we missing? Am J Physiol Lung Cell Mol Physiol 2012;302:L498–503.10.1152/ajplung.00322.2011Suche in Google Scholar
26. Singh B, Atwal OS. Ultrastructural and immunocytochemical study of the pulmonary intravascular macrophages of Escherichia coli lipopolysaccharide-treated sheep. Anat Rec 1997;247:214–24.10.1002/(SICI)1097-0185(199702)247:2<214::AID-AR8>3.0.CO;2-XSuche in Google Scholar
27. Brain JD. Mechanisms, measurement, and significance of lung macrophage function. Environ Health Persp 1992;97:5–10.10.1289/ehp.92975Suche in Google Scholar
28. DeCamp MM, Warner AE, Molina RM, Brain JD. Hepatic versus pulmonary uptake of particles injected into the portal circulation in sheep. Endotoxin escapes hepatic clearance causing pulmonary inflammation. Am Rev Respir Dis 1992;146:224–31.10.1164/ajrccm/146.1.224Suche in Google Scholar
29. Longworth KE. The comparative biology of pulmonary intravascular macrophages. Front BioSci J Virtual Libr 1997;2:d232–41.10.2741/A186Suche in Google Scholar
30. Warner AE, DeCamp MM, Jr., Molina RM, Brain JD. Pulmonary removal of circulating endotoxin results in acute lung injury in sheep. Lab Inves J Tech Method Pathol 1988;59:219–30.Suche in Google Scholar
31. Staub NC. Pulmonary intravascular macrophages. Chest 1988;93(3 Suppl):84s–5s.10.1378/chest.93.3_Supplement.84SSuche in Google Scholar
32. Singh B, Minhas KJ, Atwal OS. Ultracytochemical study of multiple dose effect of monastral blue uptake by equine pulmonary intravascular macrophages (PIMs). J Submicr Cytol Path 1994;26:235–43.Suche in Google Scholar
33. Leifsson PS, Basse A, Jensen HE, Bloch B, Aalbaek B. Pulmonary intravascular macrophages in the pathogenesis of bovine pulmonary lesions caused by Actinomyces pyogenes. J Compar Pathol 1995;112:197–206.10.1016/S0021-9975(05)80061-1Suche in Google Scholar
34. Sierra MA, Carrasco L, Gomez-Villamandos JC, Martin de las Mulas J, Mendez A, Jover A. Pulmonary intravascular macrophages in lungs of pigs inoculated with African swine fever virus of differing virulence. J Compar Pathol 1990;102:323–34.10.1016/S0021-9975(08)80021-7Suche in Google Scholar
35. Carrasco L, Ruiz-Villamor E, Gómez-Villamandos JC, Salguero FJ, Bautista MJ, Maciá M, et al. Classical swine fever: morphological and morphometrical study of pulmonary intravascular macrophages. J Comp Path 2001;125:1–7.10.1053/jcpa.2001.0470Suche in Google Scholar PubMed
36. Aharonson-Raz K, Singh B. Pulmonary intravascular macrophages and endotoxin-induced pulmonary pathophysiology in horses. Can J Vet Res 2010;74:45–9.Suche in Google Scholar
37. Humphries MJ. Integrin structure. Biochem Soc Trans 2000;28:311–39.10.1042/bst0280311Suche in Google Scholar
38. Carrasco L, Nunez A, Salguero FJ, Diaz San Segundo F, Sanchez-Cordon P, Gomez-Villamandos JC, et al. African swine fever: Expression of interleukin-1 alpha and tumour necrosis factor-alpha by pulmonary intravascular macrophages. J Comparat Pathol 2002;126:194–201.10.1053/jcpa.2001.0543Suche in Google Scholar PubMed
39. Basu S, Fenton MJ. Toll-like receptors: function and roles in lung disease. Am J Physiol Lung Cell Mol Physiol 2004;286:L887–92.10.1152/ajplung.00323.2003Suche in Google Scholar PubMed
40. Wassef A, Janardhan K, Pearce JW, Singh B. Toll-like receptor 4 in normal and inflamed lungs and other organs of pig, dog and cattle. Histol Histopathol 2004;19:1201–8.Suche in Google Scholar
41. Ezquerra A, Revilla C, Alvarez B, Perez C, Alonso F, Dominguez J. Porcine myelomonocytic markers and cell populations. Dev Comp Immunol 2009;33:284–98.10.1016/j.dci.2008.06.002Suche in Google Scholar PubMed
42. Swanson JA, Hoppe AD. The coordination of signaling during Fc receptor-mediated phagocytosis. J Leukocyte Biol 2004;76:1093–103.10.1189/jlb.0804439Suche in Google Scholar PubMed
43. Lohmann-Matthes ML, Steinmuller C, Franke-Ullmann G. Pulmonary macrophages. Eur Respir J 1994;7:1678–89.10.1183/09031936.94.07091678Suche in Google Scholar
44. Jacobson AC, Weis JJ, Weis JH. Complement receptors 1 and 2 influence the immune environment in a B cell receptor-independent manner. J Immunol 2008;180:5057–66.10.4049/jimmunol.180.7.5057Suche in Google Scholar PubMed
45. Henson PM, McCarthy K, Larsen GL, Webster RO, Giclas PC, Dreisin RB, et al. Complement fragments, alveolar macrophages, and alveolitis. Am J Pathol 1979;97:93–110.Suche in Google Scholar
46. McCarthy K, Henson PM. Induction of lysosomal enzyme secretion by alveolar macrophages in response to the purified complement fragments C5a and C5a des-arg. J Immunol 1979;123:2511–7.10.4049/jimmunol.123.6.2511Suche in Google Scholar
47. Larsen GL, McCarthy K, Webster RO, Henson J, Henson PM. A differential effect of C5a and C5a des Arg in the induction of pulmonary inflammation. Am J Pathol 1980;100:179–92.Suche in Google Scholar
48. Fine R, Shaw JO, Rogers WR. Effects of C5a on baboon alveolar macrophage migration. Am Rev Respir Dis 1981;123:110–4.Suche in Google Scholar
49. Cantu E, Gaca JG, Palestrant D, Baig K, Lukes DJ, Gibson SE, et al. Depletion of pulmonary intravascular macrophages prevents hyperacute pulmonary xenograft dysfunction. Transplantation 2006;81:1157–64.10.1097/01.tp.0000169758.57679.2aSuche in Google Scholar PubMed
50. Bertram TA, Overby LH, Danilowicz R, Eling TE, Brody AR. Pulmonary intravascular macrophages metabolize arachidonic acid in vitro. Comparison with alveolar macrophages. Am Rev Respir Dis 1988;138:936–44.10.1164/ajrccm/138.4.936Suche in Google Scholar PubMed
51. Siore AM, Parker RE, Stecenko AA, Cuppels C, McKean M, Christman BW, et al. Endotoxin-induced acute lung injury requires interaction with the liver. Am J Physiol Lung Cell Mol Physiol 2005;289:L769–76.10.1152/ajplung.00137.2005Suche in Google Scholar PubMed
52. Bertram TA, Overby LH, Danilowicz R, Eling TE, Brody AR. Pulmonary intravascular macrophages produce prostaglandins and leukotrienes in vitro. Chest 1988;93(3 Suppl):82s–4s.10.1378/chest.93.3_Supplement.82SSuche in Google Scholar
53. Parbhakar OP, Duke T, Townsend HG, Singh B. Depletion of pulmonary intravascular macrophages partially inhibits lipopolysaccharide-induced lung inflammation in horses. Vet Res 2005;36:557–69.10.1051/vetres:2005016Suche in Google Scholar
54. Duke-Novakovski T, Singh-Suri S, Kajikawa O, Caldwell S, Charavaryamath C, Singh B. Immuno-phenotypic and functional characterization of rabbit pulmonary intravascular macrophages. Cell Tissue Res 2013;351:149–60.10.1007/s00441-012-1509-2Suche in Google Scholar PubMed
55. Singh B, Pearce JW, Gamage LN, Janardhan K, Caldwell S. Depletion of pulmonary intravascular macrophages inhibits acute lung inflammation. Am J Physiol Lung Cell Mol Physiol 2004;286:L363–72.10.1152/ajplung.00003.2003Suche in Google Scholar PubMed
56. Thenappan T, Goel A, Marsboom G, Fang YH, Toth PT, Zhang HJ, et al. A central role for CD68(+) macrophages in hepatopulmonary syndrome. Reversal by macrophage depletion. Am J Respir Crit Care Med 2011;183:1080–91.10.1164/rccm.201008-1303OCSuche in Google Scholar PubMed PubMed Central
57. Bhatia M, Moochhala S. Role of inflammatory mediators in the pathophysiology of acute respiratory distress syndrome. J Pathol 2004;202:145–56.10.1002/path.1491Suche in Google Scholar PubMed
58. Warner AE, Molina RM, Brain JD. Uptake of bloodborne bacteria by pulmonary intravascular macrophages and consequent inflammatory responses in sheep. Am Rev Respir Dis 1987;136:683–90.10.1164/ajrccm/136.3.683Suche in Google Scholar PubMed
59. Miyamoto K, Schultz E, Heath T, Mitchel MD, Albertine KH, Staub NC. Pulmonary intravascular macrophages and hemodynamic effects of liposomes in sheep. J Appl Physiol 1988;64:1143–50.10.1152/jappl.1988.64.3.1143Suche in Google Scholar PubMed
60. Staub NC, Sr., Longworth KE, Serikov V, Jerome EH, Elsasser T. Detergent inhibits 70–90% of responses to intravenous endotoxin in awake sheep. J Appl Physiol (1985) 2001;90:1788–97.10.1152/jappl.2001.90.5.1788Suche in Google Scholar
61. Mizgerd JP, Molina RM, Stearns RC, Brain JD, Warner AE. Gadolinium induces macrophage apoptosis. J Leukocyte Biol 1996;59:189–95.10.1002/jlb.59.2.189Suche in Google Scholar
62. Singh B, de la Concha-Bermejillo A. Gadolinium chloride removes pulmonary intravascular macrophages and curtails the degree of ovine lentivirus-induced lymphoid interstitial pneumonia. Int J Exp pathol 1998;79:151–62.Suche in Google Scholar
63. Gaca JG, Palestrant D, Lukes DJ, Olausson M, Parker W, Davis RD, Jr. Prevention of acute lung injury in swine: depletion of pulmonary intravascular macrophages using liposomal clodronate. J Surg Res 2003;112:19–25.10.1016/S0022-4804(03)00142-2Suche in Google Scholar
64. Holt PG, Degebrodt A, Venaille T, O’Leary C, Krska K, Flexman J, et al. Preparation of interstitial lung cells by enzymatic digestion of tissue slices: preliminary characterization by morphology and performance in functional assays. Immunol 1985;54:139–47.Suche in Google Scholar
65. Chitko-McKown CG, Chapes SK, Brown RE, Phillips RM, McKown RD, Blecha F. Porcine alveolar and pulmonary intravascular macrophages: comparison of immune functions. J Leukocyte Biol 1991;50:364–72.10.1002/jlb.50.4.364Suche in Google Scholar PubMed
66. Sakai H, Suzuki Y, Sou K, Kano M. Cardiopulmonary hemodynamic responses to the small injection of hemoglobin vesicles (artificial oxygen carriers) in miniature pigs. J BioMed Mater Res Part A. 2012;100:2668–77.10.1002/jbm.a.34208Suche in Google Scholar PubMed
67. Sone Y, Serikov VB, Staub NC, Sr. Intravascular macrophage depletion attenuates endotoxin lung injury in anesthetized sheep. J Appl Physiol (1985) 1999;87:1354–9.10.1152/jappl.1999.87.4.1354Suche in Google Scholar PubMed
68. Puchner TC, Kugathasan S, Kelly KJ, Binion DG. Successful desensitization and therapeutic use of infliximab in adult and pediatric Crohn’s disease patients with prior anaphylactic reaction. Inflamm Bowel Dis 2001;7:34–7.10.1097/00054725-200102000-00005Suche in Google Scholar PubMed
69. Duburque C, Lelong J, Iacob R, Seddik M, Desreumaux P, Fournier C, et al. Successful induction of tolerance to infliximab in patients with Crohn’s disease and prior severe infusion reactions. Aliment Pharmacol Ther 2006;24:851–8.10.1111/j.1365-2036.2006.03026.xSuche in Google Scholar PubMed
70. Castells MC. Hypersensitivity to antineoplastic agents. CurrPharmDes 2008;14:2892–901.10.2174/138161208786369803Suche in Google Scholar PubMed
71. Castells MC, Tennant NM, Sloane DE, Hsu FI, Barrett NA, Hong DI, et al. Hypersensitivity reactions to chemotherapy: outcomes and safety of rapid desensitization in 413 cases. J Allergy Clin Immunol 2008;122:574–80.10.1016/j.jaci.2008.02.044Suche in Google Scholar PubMed
72. Gabizon AA, Muggia FM. Initial clinical evaluation of pegylated liposomal doxorubicin in solid tumors. In: Woodle MC, G. S, editors. Long-circulating liposomes: old drugs, new therapeutics. Austin, TX: Landes Bioscience, 1998:155–74.Suche in Google Scholar
73. Klingensmith WC, 3rd, Yang SL, Wagner HN, Jr. Lung uptake of Tc-99m sulfur colloid in liver and spleen imaging. J Nucl Med Official Publ Soc Nucl Med 1978;19:31–5.Suche in Google Scholar
©2015 by De Gruyter
Artikel in diesem Heft
- Frontmatter
- In this issue
- Guest Editorial and News
- Complement activation-related pseudoallergy: insights into a stress reaction to nanomedicines in blood
- What’s up in nanomedicine?
- Special section CARPA Part 1
- Reviews
- The possible role of factor H in complement activation-related pseudoallergy (CARPA): a failed attempt to correlate blood levels of FH with liposome-induced hypersensitivity reactions in patients with autoimmune disease
- Mini Reviews
- Rodent models of complement activation-related pseudoallergy: Inducers, symptoms, inhibitors and reaction mechanisms
- Pulmonary intravascular macrophages: prime suspects as cellular mediators of porcine CARPA
- Regular contributions
- Critical Review
- Towards nano-diagnostics for bacterial infections
- Review
- Effects of silver nanoparticles on human health
- Original article
- Potential of rod, sphere and semi-cube shaped gold nanoparticles to induce cytotoxicity and genotoxicity in human blood lymphocytes in vitro
Artikel in diesem Heft
- Frontmatter
- In this issue
- Guest Editorial and News
- Complement activation-related pseudoallergy: insights into a stress reaction to nanomedicines in blood
- What’s up in nanomedicine?
- Special section CARPA Part 1
- Reviews
- The possible role of factor H in complement activation-related pseudoallergy (CARPA): a failed attempt to correlate blood levels of FH with liposome-induced hypersensitivity reactions in patients with autoimmune disease
- Mini Reviews
- Rodent models of complement activation-related pseudoallergy: Inducers, symptoms, inhibitors and reaction mechanisms
- Pulmonary intravascular macrophages: prime suspects as cellular mediators of porcine CARPA
- Regular contributions
- Critical Review
- Towards nano-diagnostics for bacterial infections
- Review
- Effects of silver nanoparticles on human health
- Original article
- Potential of rod, sphere and semi-cube shaped gold nanoparticles to induce cytotoxicity and genotoxicity in human blood lymphocytes in vitro