Potential of rod, sphere and semi-cube shaped gold nanoparticles to induce cytotoxicity and genotoxicity in human blood lymphocytes in vitro
-
Mona A.M. Abo-Zeid
Mona A.M. Abo-Zeid is an Assistant Professor of Cytogenetics at the Genetics and Cytology Department, and a member of the Cancer Biology Laboratory, Center of Excellence for Advanced Sciences, NRC, Egypt. She is a Fellow of the Molecular Cytogenetics Laboratory, Institut für Humangenetik-Universitätsklinikum, Jena, Germany. Mona has published many articles concerning with discoveries for new cancer treatments including the combination of nano-chemotheraputic drugs with photodynamic therapy by photosensitizers and laser irradiation. She has developed bioassays for developing and screening of new cancer chemopreventive drugs. She supervises MD, PhD and Master’s student theses.
 ,
Thomas Liehr
,
Thomas Liehr
Thomas Liehr is a PhD, Dr. h.c. and invited Professor of the Yerevan-State-University, Armenia. He currently works at Institute of Human Genetics, Jena, Germany and is head of the Center for Molecular Cytogenetics. He is interested in small supernumerary marker chromosomes, uniparental disomy, multicolor FISH, chromosomal heteromorphisms, fragile sites, leukemia-research, interphase architecture and chromosomal evolution. He is involved in the international cooperation with Armenia, Brazil, Croatia, Egypt, Portugal, Russia, Serbia and Thailand. He is the editor of
Molecular Cytogenetics . He is on the committee of the Permanent Working Group Marker Chromosomes of the European Cytogenetic Association (ECA), and is a member of the European Board of Medical Genetics of the European Society of Human Genetics. He has written four books, 50 book chapters and over 500 journal articles.Amira received her Dr. Sc. Hum. degree in human Toxicology in 2002 at Faculty of Medicine, Heidelberg University, Germany. Afterwards, she worked for three years in the Biochemistry Department, NRC, Giza, Egypt. Since 2006, she is the group leader of the Cancer Biology Laboratory, Centre of Excellence for Advanced Sciences, NRC, Giza, Egypt. Amira has a long experience in cancer biology, with a special focus on preclinical trials and the influence of therapeutics of the tumor cells/tumor microenvironment using various cell-dependant technologies and animal models.
Mahmoud Zawrah studied Applied Inorganic Chemistry at the Cairo University from 1990 to 1996, specializing in Material Chemistry for his MSc and PhD. He has been Professor of Material Chemistry and Nanotechnology since April 2007 at the National Research Centre (NRC), Cairo, Egypt. His special fields are nanomaterials for different applications such as medical, environmental and structural materials. He has been awarded several national and international prizes. He has published more than 100 articles in international journals and conferences.
Moneeb A.K. Othman assistant researcher and DAAD scholarship holder to study PhD in Germany in Department of Molecular Cytogenetics, Institute of Human Genetics and Anthropology, FSU Jena, Germany. His PhD study concerns with “Cryptic and complex chromosomal aberrations in Acute Leukemia”. He has published several articles in the field of Leukemia

Abstract
Aim: Gold nanoparticles (GNPs) are intended to be used in nanomedicine. Due to nanotechnology innovation GNPs of variable sizes and in different shapes including rods, spheres, cubes, etc., can easily be produced. The aim of the present studies was to evaluate the cyto-and genotoxicity inducible by different shaped GNPs on normal human peripheral blood lymphocytes.
Methods: Four different shapes of GNPs including big rod GNPs (BR-GNPs, 50 nm), small rod GNPs (SR-GNPs, 30 nm), sphere GNPs (S-GNPs, 15 nm) and semi-cube GNPs (SC-GNPs, 15 nm) were studied. Cultured human blood lymphocytes were treated with different concentrations of these GNPs for 24 h in vitro. Cytotoxicity was evaluated based on the mitotic index (MI), while genotoxicity was studied by an interphase-fluorescence in situ hybridization (I-FISH) assay. The following genes were studied in I-FISH: TP53, CASP3, CASP7, CASP9, TNF and CRP. The induction of cell death was investigated using the apoptotic-necrotic assay.
Results: The lowest concentration of BR-GNPs neither had an effect mitotic activity nor enhanced gain or loss of examined gene signals in a significant manner with I-FSH. Other concentrations of BR-GNPs, SR-GNPs, S-GNPs and SC-GNPs with all concentrations inhibited the mitotic activity of the cells and reduced the cell proliferation highly significantly. The different types of GNPs initiated the duplication of CASP3, CASP7 and CASP9 besides TP53 genes; however they reduced the signals of TNF and CRP genes highly significantly as supported by I-FISH results. The apoptotic-necrotic assay demonstrated that small rod, sphere and semi-cube GNPs enhanced the cell death rate through the apoptotic pathway.
Conclusion: GNPs at high concentration can reduce the cell proliferation and induce DNA damage. Low concentration of rod-shaped GNPs at 50 nm was safe on human lymphocytes. Further research studies are required to optimize the concentration, shape and size of GNPs before using them in nanomedicine.
Introduction
Biotechnology research uses nanoparticles in different approaches including drug therapy (1), antimicrobial agents (2), vectors (3) and fluorescence markers (4). Inertness is the main characteristic of gold. This propriety allowed using gold in human medicine starting from dentistry to implantation as radioactive material in cancer treatments (5, 6). Advanced nano-techniques produce gold in different sizes and shapes, such as rod (7), sphere (8), cube (9) and triangle/prism (10), allowing absorption and scattering of light in the visible and near infrared wavelengths (11). Investigation of cytotoxicity of Gold nanoparticles (GNPs) was previously reported in cancer cell lines. Patra et al. (12) demonstrated that GNPs induced apoptosis in human carcinoma lung cancer cell line (A549) and did not induce effects on baby hamster kidney (BHK21) and human hepatocellular carcinoma (HepG2) cell lines. Connor et al. (13) used GNPs in drug delivery assuming their safety upon cells.
The authors studied the potential of macrophages to uptake GNPs (nanorods and nanospheres). They found that macrophages engulfed nanorods more rapidly than nonospheres. Also, they found that GNPs have the ability to penetrate macrophages and induce micropinocytosis as detected with electron microscope. Also, they inserted polyethylene oxide inside nanorods which in turn changed macrophages phenotypes due to increase the time of uptake of PEO-coated nanorods inside macrophages. This longer time induced inflammation due to accumulation of nanoparticles (14).
Recent reports found that GNPs penetrated cellular membrane of B-lymphocyte cell line (CH12.LX) and interacted with NF-κB signaling pathway, responsible for regulating immune system against infection (15), as investigated with TEM. They illustrated that activation of transcriptional NF-κB signaling pathway led to activate IKKα and IKKβ, degenerate IκBα and promote nuclear RelA activation (16). There are several studies reported functional properties of GNPs on different cells including in vitro and in vivo studies (17–19).
Activation of proteases (caspases) considered one of the main pathways affecting apoptosis (programmed cell death), due to morphological and biochemical changes occur inside cells (20, 21). Conglomerzation as a result from chromatin condensation, DNA fragmentation beside cellular shrinkage and blebbing are the main characteristics to apoptosis (22–24). Cells are considered in early apoptosis once they start the programed cell death, in this stage phosphatidyl serine is expressed on intact cellular membrane surface to facilitate identification by phagocytic cells. As apoptosis proceeds, cells transfer from early apoptosis stage to late apoptosis stage or secondary necrosis which leads to permeability of cellular membrane (25). Exposure of cells to environmental chocks including utmost high or low temperatures, mechanical pressure or chemical toxins can induce cellular membrane punctures leading to primary necrotic cells (26). Some authors consider necrosis directs cells to die independently as a passive victim, but others refer to necrosis as atrophy process resulted from mechanism of cell death (27, 28).
Some caspases possess proteolytic activity leading to protein cleavage at aspartic acid residues, others are activated to recognize the neighboring amino acids, and both direct cells irreversibly to programmed cell death. The main functional caspases include the following groups: CASP2, CASP8, CASP9, and CASP10 as initiators; CASP3, CASP6 and CASP7 as effectors or executioners; CASP1, CASP4 and CASP5 as inflammatory groups (29, 30); CASP11 as regulator to apoptosis and cytokine maturation during septic shock; CASP2 as mediator to endoplasmic-specific apoptosis and cytotoxicity by amyloid-B; CASP13 as specific bovine gene; and CASP14 as mediator in embryonic tissues only (31–34).
Tumor suppressor gene (TP53) is known as the guardian of the genome as it regulates cell cycle and protects genome from mutations (35). The importance of the TP53 gene comes from its p53 protein which encodes to antiproliferative functions by controlling protein-protein interactions, thus mutations in TP53 gene is accompanied with human cancer (36, 37). Tumor Necrosis Factor alpha (TNF) is one of the inflammatory cytokines produced by macrophages or monocytes, and organizes a wide range of signals transduction within cells to initiate apoptosis or necrosis (38). C-reactive protein (CRP) is one of the inflammatory proteins expressed in late inflammation (39, 40). Its expression allows identification of pathogens and dead cells to facilitate their elimination by phagocytes (41).
One of the molecular cytogenetic approaches used to investigate genotoxicity is interphase-fluorescence in situ hybridization (I-FISH). The efficiency of I-FISH comes from its potential to screen numerous targets using multicolor I-FISH technique (42). Also, I-FISH screens chromosome mosaicism and intercellular genomic variances such as numerical (aneuploidy or polyploidy) and structural chromosomes abnormalities (43).
This study was targeting to investigate the potential of GNPs to induce cytotoxicity and genotoxicity in human blood lymphocytes in vitro. Therefore different genetic markers including CASP3, CASP7, CASP9, TP53, TNF and CRP genes were studied to assess the genomic damage and apoptosis/necrosis which could be induced by GNPs of different shapes.
Materials and methods
Preparation of gold nanoparticles (GNPs)
Big rod GNPs (BR-GNPs, 50 nm), small rod GNPs (SR-GNPs, 30 nm), sphere GNPs (S-GNPs, 15 nm) and semi-cube GNPs (SC-GNPs, 15 nm) were synthesized following the modified seed-mediated protocol of Nikoobakht and El-Sayed (44). Firstly, the seed nanoparticles were synthesized by adding 5.0 mL of HAuCl4 (1.0 mM) to 5.0 mL of cetyltrimethylammonium bromide (CTAB, 0.2 M) (0.2 M) followed by the addition of ice-cold NaBH4 (0.6 mL, 10.0 mM). The solution was then stirred for 2 min.
The second step was different and related to each shape as follows:
Preparation of BR-GNPs
In the growth solution, the following has been added in sequence; 10 mL of HAuCl4 (1.0 mM), 10 mL of CTAB (0.2 M), 0.5 mL of AgNO3 (4.0 mM), HCl (160 μL, 37%) and 0.14 mL of ascorbic acid (78.8 mM), then gentle shaking until the solution became colorless; after that 24 μL of the seed solution (prepared in the first step) were injected directly to the growth solution; then the growth solution kept undisturbed for 12 h.
Preparation of SR-GNPs
The procedure was as described before, just AgNO3 solution 4.0 mM, 0.35 mL was added.
Preparation S-GNPs
The procedure was as described before, just no AgNO3 solution was added.
Preparation SC-GNPs
The procedure was as described before, just AgNO3 solution 4.0 mM, 0.25 mL was added.
Cell culture
Human blood lymphocytes were collected from a 35-year-old healthy female volunteer for all the experimental studies. Lymphocytes were routinely cultured in RPMI 1640 media with 2 mM L-glutamine, and were supplemented with 10% fetal bovine serum, penicillin (100 units/mL), and streptomycin (100 μg/mL). Cells were maintained in humidified air containing 5%CO2 at 37°C for 48 h before applying the sterile conditions for the different treatments.
Treatments
Five different experiments were performed. In the first experiment, cells were incubated for 24 h without treatments as a negative control. For the other experiments, lymphocytes were treated for 24 h with BR-GNPs, SR-GNPs, S-GNPs or SC-GNPs at variable concentrations (0.05, 0.1, 0.5 and 1.0 μg/mL) before final incubation of 24 h. Colcemid at a final concentration of 1-μg/mL was added to cultured human blood lymphocytes and incubated at 37°C for 90 min, and then the tubes were centrifuged. The pellets were resuspended in 5 mL 0.075 M KCl and incubated at 37°C for 20 min, then centrifuged. Carnoy’s fixative [methanol:acetic acid (3:1)] was added to the pellets, then the mixture was centrifuged, resuspended, and incubated at –20°C for 5 min fixative; finally the preparation was finalized with a last centrifugation step. The supernatant was discarded, and the cells were diluted in 200 μL of the remaining supernatant and placed on a slide (45).
Mitotic index (MI)
The prepared slides for FISH analysis were used to determine the mitotic index (MI), which was based on the scoring of at least 1000 cells for each sample. The number of dividing cells including prophases and metaphases was recorded. The MI was calculated as number of dividing cells/total number of counted cells.
Interphase-FISH analysis (I-FISH)
For I-FISH purposes, the air dried slides with the target DNA were denatured in 70% formamide/2×SSC at 70–75°C for 2–3 min, transferred to chilled 70% ethanol and then dehydrated through ethanol series. The slides were air dried and used for commencing I-FISH. Specific gene markers were prepared by using specific bacterial artificial chromosomes (BACs) and were directly labeled with the different fluorochromes (Table 1). The specific BACs for genes involved in apoptosis and/or necrosis were used are listed in Table 1. The fluorescently labeled DNA probes were hybridized to the samples. The DNA probes were denatured, applied on denatured target DNA on slides, covered by a 22 mm×22 mm cover slip and sealed with rubber cement. The probes and target DNA were hybridized by incubation of slides at 37°C for 24 h and stained with nuclear counterstain 4,6-diamidino-2-phenylindole (DAPI). Fluorescence microscopy (Axio Imager Z2, Carl Zeiss, Jena, Germany) with MetaSystems Isis software was used to analyze and record all observed signal patterns. At least 100 interphase cells were analyzed for each sample with filters specific for each of the probe fluorophores. ISCN-FISH nomenclature (46) was used to describe I-FISH-results (nuc ish).
List of used bacterial artificial chromosomes (BACs) probes, their identification by and specific gene type, the cytogenetic localization of the probes (cytoband), molecular position according to NCBI36/hg18 and the labeled fluorochrome.
| Gene | Cytoband | BAC-Probe | Position (NCBI36/hg18) | Fluorochrome |
|---|---|---|---|---|
| CASP9 | 1p36.21 | RP11-265F14 | 15,630,693-15,811,097 | SpectrumGreen (SG) |
| CRP | 1q23.2 | RP11-206M24 | 157,948,703-157,951,003 | Diethylaminucoumarin (DEAC) |
| CASP3 | 4q35.1 | RP11-818I13 | 185,323,840-185,545,945 | Texas Red (TR) |
| TNF | 6p21.3 | RP11-732M11 | 32,032,289.-32,218,323 | SpectrumOrange (SO) |
| CASP7 | 10q25.3 | RP11-357H24 | 114,874,396-115,058,203 | SpectrumOrange (SO) |
| TP53 | 17p13.1 | RP11-89D11 | 7,436,454-7,603,767 | SpectrumGreen (SG) |
Apoptosis and necrosis test
To acquire the effect of GNPs on cell death, apoptosis and necrosis were investigated using acridine orange/ethidium bromide staining (47). Human blood lymphocytes were cultured in 6-well plates at a density of 5×105 cells/well and treated with two concentrations (0.1 and 1.0 μg/mL) of different shapes of GNPs and then incubated for 24 h. The cells were stained using the nucleic acid-binding dye mixture of 100 μg/mL acridine orange and 100 μg/mL ethidium bromide in PBS, and were examined by fluorescence microscopy. Viable cells had green fluorescent nuclei with organized structure. The early apoptotic cells had yellow chromatin in the nuclei that were highly condensed or fragmented. Apoptotic cells also exhibited membrane blebbing. The late apoptotic cells had orange chromatin with nuclei that were highly condensed and fragmented. Only cells with yellow, condensed, or fragmented nuclei were registered as apoptotic cells in a blind, nonbiased manner. The necrotic cells had bright orange chromatin in round nuclei (48). For each sample, at least 500 cells/well in the six wells were counted, and the percentage of apoptotic or necrotic cells was determined as total number of apoptotic or necrotic cells/total number of counted cells×100.
Statistical analysis
The statistical analysis with GraphPadInStat software was used. MI and apoptotic-necrotic tests were carried out by One-way ANOVA with post test and values were expressed as means±standard deviation (SD). Data for I-FISH was analyzed using 2χ2 contingency test.
Results
Characterization of GNPs
TEM images (Figure 1A) illustrate the four types of prepared GNPs as with almost sizes BR-GNPs 50 nm length and 12 nm width, SR-GNPs 30 nm length and 12 nm width, S-GNPs 15 nm length and SC-GNPs 15 nm length. It is also noted that the nanoparticles in the gold nanoclusters were nearly individually isolated. Also Figure 1B presents the UV-vis absorption spectra of the prepared gold sols, in which the absorption bands were characteristic of the surface plasmon bands of each GNPs type.
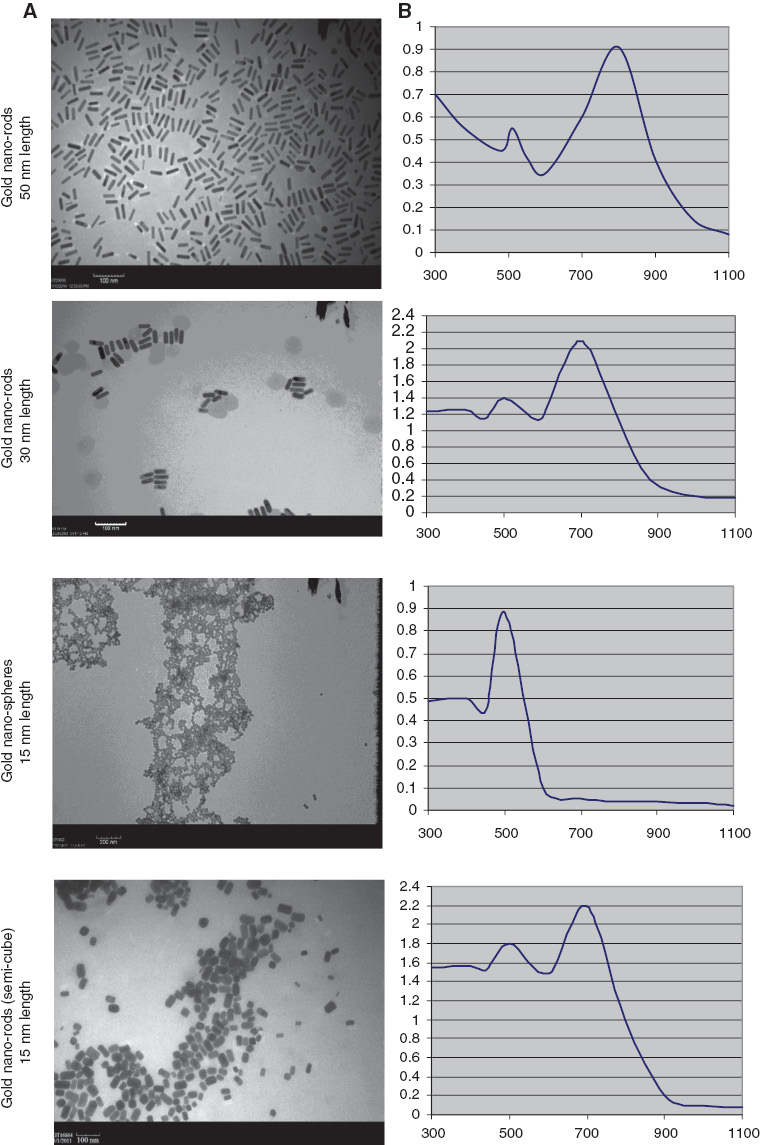
Illustration of (A) the morphology and particle size of prepared gold nanoparticles with TEM and (B) the absorption spectrum of the prepared GNPs solution were measured using a Jasco UV-vis-near-infrared spectrophotometer, V-630.
Mitotic index
Human blood lymphocytes from a healthy female volunteer with normal karyotype were used to carry out these experimental studies (Figure 2A). The effect of different shaped GNPs at variable concentrations on cell division, including divided and non-divided cells, was recorded (Figure 2B, C).
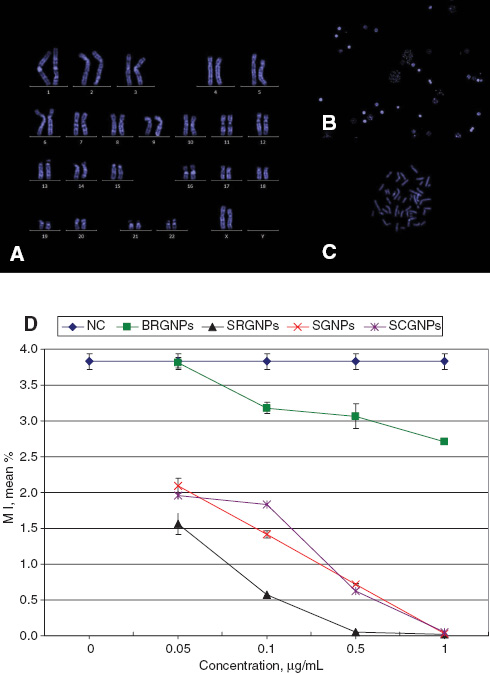
Photos represents (A) karyotype, (B) nucleated and divided cells (20×), (C) metaphase (100×), all with blue DAPI stain. (D) The mean percentage of MI in human blood lymphocytes treated with different concentration of GNPs for 24 h, data represented as Mean±SD, n=3.
BR-GNPs did not affect on MI of lymphocytes in a significant manner when applied at lowest concentrations (0.05 μg/mL). Reduction in cell division was observed to be highly significant (p<0.001) with all other higher concentrations of BR-GNPs (Figure 2D).
For the other GNPs shapes including SR-GNPs, S-GNPs and SC-GNPs, it has been demonstrated that they inhibited the cell proliferation in an extremely significant manner (p<0.001) including the lowest concentrations; also they almost prevented cell division when using the highest concentrations (Figure 2D).
I-FISH
The mode of action of GNPs to induce cyto- and genotoxicity was evaluating by measuring their effects on four genes specific for apoptosis and two specific for necrosis. Each gene exists in two copies per cell in normal cells.
We investigated the genotoxicity of GNPs on lymphocytes by assessing the percentages of deletion or duplication of signals for each gene with different GNPs concentrations. Figure 3 illustrates some typical figures for I-FISH in human blood lymphocytes with differently labeled probes.
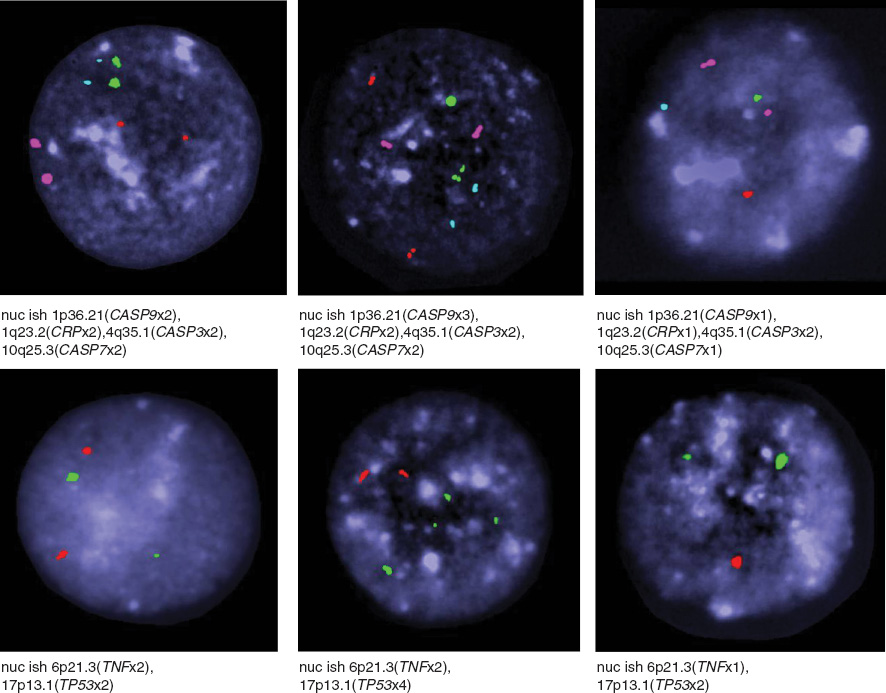
Photographs panel represent examples of variable numbers of signals per nucleus demonstrated with different DNA probes such as CASP3 (pink), CASP7 (red), CASP9 (green), CRP (blue), TP53 (green) and TNF (red), nuclei with DAPI (dark blue) in human blood lymphocytes.
It was demonstrated that all shapes of GNPs did not affect on CASP3, CASP7 and CASP9 genes at low concentration (0.05 μg/mL): neither deletions nor amplifications were observed. Interestingly for CASP3 gene, GNPs initiated highly significantly (p<0.001) duplication of signals which was positively correlated with increasing concentrations. While the percentage of nuclei with loss of signals was reduced significantly with increasing concentrations of GNPs, the proportion of duplication was found to be twice times higher than that of deletions (Table 2). For CASP7 and CASP9 genes, GNPs with all shapes induced in treated cells both duplication and reduction of signals (p<0.001; Table 2).
The number and percentage of loss or gain signals of CASP3, CASP7 and CASP9 DNA probes in human blood lymphocytes after treatment with different shapes of GNPs for 24 h.
| Treatment | Conc., μg/mL | Type of probea | No. of signals/nucleusb | Number and percentage of nuclei with signal-loss | Number and percentage of nuclei with signal-gain | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | No. | % | No. | % | |||
| Control | 0 | CASP3 | 0 | 400 | 0 | 0 | 0 | 0.00 | 0 | 0.00 |
| BR-GNPs | 0.05 | 0 | 398 | 2 | 0 | 0 | 0.00 | 2 | 0.50 | |
| 0.1 | 2 | 374 | 20 | 4 | 2 | 0.50 | 24e | 6.00 | ||
| 0.5 | 8 | 347 | 27 | 18 | 8c | 2.00 | 45e | 11.25 | ||
| 1 | 24 | 310 | 52 | 14 | 24e | 6.00 | 66e | 16.50 | ||
| SR-GNPs | 0.05 | 0 | 390 | 10 | 0 | 0 | 0.00 | 10d | 2.50 | |
| 0.1 | 0 | 372 | 22 | 6 | 0 | 0.00 | 28e | 7.00 | ||
| 0.5 | 18 | 337 | 32 | 13 | 18e | 4.50 | 45e | 11.25 | ||
| 1 | 22 | 310 | 22 | 46 | 22e | 5.50 | 68e | 17.00 | ||
| S-GNPs | 0.05 | 0 | 395 | 1 | 4 | 0 | 0.00 | 5 | 1.25 | |
| 0.1 | 2 | 383 | 4 | 11 | 2 | 0.50 | 15e | 3.75 | ||
| 0.5 | 9 | 322 | 41 | 28 | 9d | 2.25 | 69e | 17.25 | ||
| 1 | 12 | 306 | 33 | 49 | 12d | 3.00 | 82e | 20.50 | ||
| SC-GNPs | 0.05 | 0 | 400 | 0 | 0 | 0 | 0.00 | 0 | 0.00 | |
| 0.1 | 2 | 386 | 5 | 7 | 2 | 0.50 | 12d | 3.00 | ||
| 0.5 | 3 | 350 | 34 | 13 | 3 | 0.75 | 47e | 11.75 | ||
| 1 | 7 | 316 | 41 | 36 | 7c | 1.75 | 77e | 19.25 | ||
| Control | 0 | CASP7 | 2 | 398 | 0 | 0 | 2 | 0.50 | 0 | 0.00 |
| BR-GNPs | 0.05 | 0 | 400 | 0 | 0 | 0 | 0.00 | 0 | 0.00 | |
| 0.1 | 14 | 380 | 6 | 0 | 14d | 3.50 | 6c | 1.50 | ||
| 0.5 | 22 | 366 | 10 | 2 | 22e | 5.50 | 12d | 3.00 | ||
| 1 | 71 | 288 | 32 | 9 | 71e | 17.75 | 41e | 10.25 | ||
| SR-GNPs | 0.05 | 2 | 398 | 0 | 0 | 2 | 0.50 | 0 | 0.00 | |
| 0.1 | 0 | 380 | 7 | 13 | 0 | 0.00 | 20e | 5.00 | ||
| 0.5 | 35 | 330 | 15 | 20 | 35e | 8.75 | 35e | 8.75 | ||
| 1 | 54 | 298 | 24 | 24 | 54e | 13.50 | 48e | 12.00 | ||
| S-GNPs | 0.05 | 4 | 395 | 0 | 1 | 4 | 1.00 | 1 | 0.25 | |
| 0.1 | 11 | 381 | 4 | 4 | 11c | 2.75 | 8c | 2.00 | ||
| 0.5 | 45 | 322 | 17 | 16 | 45e | 11.25 | 33e | 8.25 | ||
| 1 | 57 | 297 | 18 | 28 | 57e | 14.25 | 46e | 11.50 | ||
| SC-GNPs | 0.05 | 4 | 396 | 0 | 0 | 4 | 1.00 | 0 | 0.00 | |
| 0.1 | 11 | 377 | 8 | 4 | 11c | 2.75 | 12d | 3.00 | ||
| 0.5 | 41 | 336 | 16 | 7 | 41e | 10.25 | 23e | 5.75 | ||
| 1 | 52 | 314 | 24 | 10 | 52e | 13.00 | 34e | 8.50 | ||
| Control | 0 | CASP9 | 0 | 400 | 0 | 0 | 0 | 0.00 | 0 | 0.00 |
| BR-GNPs | 0.05 | 5 | 395 | 0 | 0 | 5 | 1.25 | 0 | 0.00 | |
| 0.1 | 16 | 373 | 7 | 4 | 16e | 4.00 | 11d | 2.75 | ||
| 0.5 | 21 | 353 | 22 | 4 | 21e | 5.25 | 26e | 6.50 | ||
| 1 | 72 | 291 | 22 | 15 | 72e | 18.00 | 37e | 9.25 | ||
| SR-GNPs | 0.05 | 2 | 394 | 0 | 4 | 2 | 0.50 | 4 | 1.00 | |
| 0.1 | 4 | 388 | 2 | 6 | 4 | 1.00 | 8c | 2.00 | ||
| 0.5 | 32 | 326 | 25 | 17 | 32e | 8.00 | 42e | 10.50 | ||
| 1 | 34 | 320 | 36 | 10 | 34e | 8.50 | 46e | 11.50 | ||
| S-GNPs | 0.05 | 0 | 400 | 0 | 0 | 0 | 0.00 | 0 | 0.00 | |
| 0.1 | 6 | 386 | 4 | 4 | 6c | 1.50 | 8c | 2.00 | ||
| 0.5 | 28 | 341 | 25 | 6 | 28e | 7.00 | 31e | 7.75 | ||
| 1 | 53 | 300 | 21 | 26 | 53e | 13.25 | 47e | 11.75 | ||
| SC-GNPs | 0.05 | 2 | 394 | 0 | 4 | 2 | 0.50 | 4 | 1.00 | |
| 0.1 | 6 | 379 | 6 | 9 | 6c | 1.50 | 15e | 3.75 | ||
| 0.5 | 27 | 343 | 14 | 16 | 27e | 6.75 | 30e | 7.50 | ||
| 1 | 34 | 319 | 22 | 25 | 34e | 8.50 | 47e | 11.75 | ||
aStandard signals with each probe for normal human blood lymphocytes are two signals. bTotal No. of nuclei (400/experiment). cp<0.05, dp<0.01, ep<0.001 in comparing to non-treated cells (using 2χ2 contingency test).
For TP53, TNF and CRP genes (Table 3), it was demonstrated that BR-GNPs did not stimulate duplication or reduction of signals at the lowest concentration (0.05 μg/mL). With higher concentrations, GNPs evoked significant reduction of signals and these deletions were increased highly significantly and reached significance of p<0.001 when high concentrations of GNPs (1.0 μg/mL) were applied. However, the percentages of cells with duplicated signals were observed to grow with increasing GNP-concentrations, the proportion of signal-loss was higher than that of duplication (Table 3).
The number and percentage of loss or gain signals of TP53, TNF-α and CRP DNA probes in human blood lymphocytes after treatment with different shapes of GNPs for 24 h.
| Treatment | Conc., μg/mL | Type of probea | No. of signals/nucleusb | Number and percentage of nuclei with signal loss | Number and percentage of nuclei with signal-gain | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | 2 | 3 | 4 | No. | % | No. | % | |||
| Control | 0 | TP53 | 10 | 390 | 0 | 0 | 10 | 2.50 | 0 | 0.00 |
| BR-GNPs | 0.05 | 12 | 387 | 0 | 1 | 12 | 3.00 | 1 | 0.25 | |
| 0.1 | 26 | 359 | 14 | 1 | 26d | 6.50 | 15e | 3.75 | ||
| 0.5 | 35 | 334 | 25 | 6 | 35e | 8.75 | 31e | 7.75 | ||
| 1 | 101 | 262 | 28 | 9 | 101e | 25.25 | 37e | 9.25 | ||
| SR-GNPs | 0.05 | 20 | 377 | 3 | 0 | 20 | 5.00 | 3 | 0.75 | |
| 0.1 | 47 | 340 | 11 | 2 | 47e | 11.75 | 13e | 3.25 | ||
| 0.5 | 72 | 311 | 14 | 3 | 72e | 18.00 | 17e | 4.25 | ||
| 1 | 93 | 281 | 22 | 4 | 93e | 23.25 | 26e | 6.50 | ||
| S-GNPs | 0.05 | 19 | 376 | 1 | 4 | 19 | 4.75 | 5 | 1.25 | |
| 0.1 | 30 | 362 | 6 | 2 | 30d | 7.50 | 8c | 2.00 | ||
| 0.5 | 71 | 314 | 12 | 3 | 71e | 17.75 | 15e | 3.75 | ||
| 1 | 86 | 294 | 16 | 4 | 86e | 21.50 | 20e | 5.00 | ||
| SC-GNPs | 0.05 | 22 | 378 | 0 | 0 | 22c | 5.50 | 0 | 0.00 | |
| 0.1 | 57 | 328 | 14 | 1 | 57e | 14.25 | 15e | 3.75 | ||
| 0.5 | 76 | 292 | 28 | 4 | 76e | 19.00 | 32e | 8.00 | ||
| 1 | 79 | 240 | 54 | 27 | 79e | 19.75 | 81e | 20.25 | ||
| Control | 0 | TNF-α | 5 | 393 | 2 | 0 | 5 | 1.25 | 2 | 0.50 |
| BR-GNPs | 0.05 | 10 | 390 | 0 | 0 | 10 | 2.50 | 0 | 0.00 | |
| 0.1 | 27 | 367 | 6 | 0 | 27e | 6.75 | 6 | 1.50 | ||
| 0.5 | 78 | 316 | 6 | 0 | 78e | 19.50 | 6 | 1.50 | ||
| 1 | 166 | 226 | 8 | 0 | 166e | 41.50 | 8 | 2.00 | ||
| SR-GNPs | 0.05 | 22 | 378 | 0 | 0 | 22d | 5.50 | 0 | 0.00 | |
| 0.1 | 55 | 341 | 4 | 0 | 55e | 13.75 | 4 | 1.00 | ||
| 0.5 | 118 | 257 | 22 | 3 | 118e | 29.50 | 25e | 6.25 | ||
| 1 | 153 | 234 | 11 | 2 | 153e | 38.25 | 13d | 3.25 | ||
| S-GNPs | 0.05 | 17 | 379 | 4 | 0 | 17c | 4.25 | 4 | 1.00 | |
| 0.1 | 44 | 344 | 10 | 2 | 44e | 11.00 | 12c | 3.00 | ||
| 0.5 | 89 | 303 | 8 | 0 | 89e | 22.25 | 8 | 2.00 | ||
| 1 | 136 | 258 | 6 | 0 | 136e | 34.00 | 6 | 1.50 | ||
| SC-GNPs | 0.05 | 18 | 382 | 0 | 0 | 18c | 4.50 | 0 | 0.00 | |
| 0.1 | 64 | 332 | 4 | 0 | 64e | 16.00 | 4 | 1.00 | ||
| 0.5 | 97 | 283 | 20 | 0 | 97e | 24.25 | 20e | 5.00 | ||
| 1 | 190 | 188 | 14 | 8 | 190e | 47.50 | 22e | 5.50 | ||
| Control | 0 | CRP | 0 | 400 | 0 | 0 | 0 | 0.00 | 0 | 0.00 |
| BR-GNPs | 0.05 | 5 | 395 | 0 | 0 | 5 | 1.25 | 0 | 0.00 | |
| 0.1 | 18 | 364 | 10 | 8 | 18e | 4.50 | 18e | 4.50 | ||
| 0.5 | 39 | 340 | 19 | 2 | 39e | 9.75 | 21e | 5.25 | ||
| 1 | 158 | 193 | 28 | 21 | 158e | 39.50 | 49e | 12.25 | ||
| SR-GNPs | 0.05 | 4 | 396 | 0 | 0 | 4 | 1.00 | 0 | 0.00 | |
| 0.1 | 0 | 386 | 6 | 8 | 0 | 0.00 | 14e | 3.50 | ||
| 0.5 | 54 | 328 | 12 | 6 | 54e | 13.50 | 18e | 4.50 | ||
| 1 | 121 | 271 | 7 | 1 | 121e | 30.25 | 8c | 2.00 | ||
| S-GNPs | 0.05 | 2 | 398 | 0 | 0 | 2 | 0.50 | 0 | 0.00 | |
| 0.1 | 9 | 378 | 8 | 5 | 9d | 2.25 | 13e | 3.25 | ||
| 0.5 | 116 | 264 | 14 | 6 | 116e | 29.00 | 20e | 5.00 | ||
| 1 | 121 | 246 | 25 | 8 | 121e | 30.25 | 33e | 8.25 | ||
| SC-GNPs | 0.05 | 8 | 392 | 0 | 0 | 8c | 2.00 | 0 | 0.00 | |
| 0.1 | 14 | 382 | 4 | 0 | 14e | 3.50 | 4 | 1.00 | ||
| 0.5 | 107 | 269 | 15 | 9 | 107e | 26.75 | 24e | 6.00 | ||
| 1 | 162 | 205 | 18 | 15 | 162e | 40.50 | 33e | 8.25 | ||
aStandard signals with each probe for normal human blood lymphocytes are two signals. bTotal No. of nuclei (400/experiment). cp<0.05, dp<0.01, ep<0.001 in comparing to non-treated cells (using 2χ2 contingency test).
Apoptosis and necrosis
The effects of different shapes GNPs in inducing apoptotic and/or necrotic effects on lymphocytes were evaluated using ethidium bromide- acridine orange dyes. Figure 4A represents pictures for cells with different colors according to their viability (green), and apoptotic (yellow) or necrotic (orange) cell death.
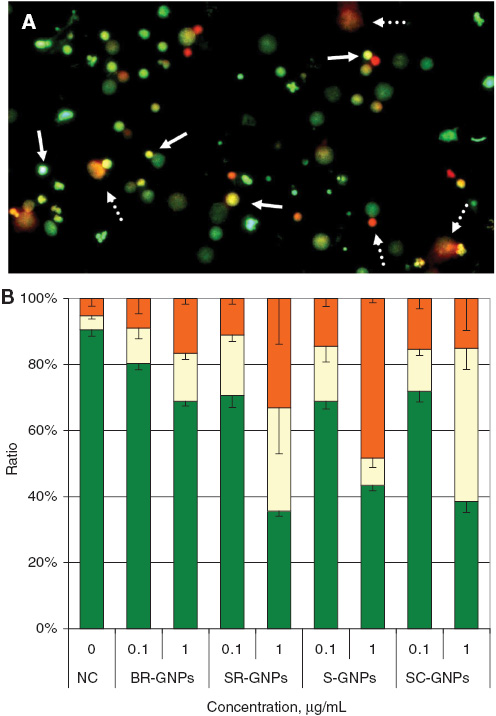
Photograph illustrates the different types of cells using acridine orange/ethidium bromide dye, (A) living cells (green), apoptotic cells (yellow, e.g., solid arrows) and necrotic cells (orange; e.g., dash arrows). (B) Analysis of apoptosis and necrosis, the percentages of the cell populations in the regions of living cells (green), apoptotic (yellow) and necrotic cells (orange) were evaluated for different GNPs concentrations. Values are means±SD, n=3.
Two concentrations were used to evaluate cyto- and genotoxicity of GNPs (0.1 and 1.0 μg/mL). BR-GNPs reduced the ratio of viable cells in comparison to non-treated cells significantly (p<0.01) at concentration 0.1 μg/mL. Still, the higher concentration (1.0 μg/mL) reduced the ratio of cell viability significantly (p<0.001) in comparison to non-treated cells. For SR-GNPs, S-GNPs and SC-GNPs, the cell viability reduced with extremely high significant manner (p<0.001) with both concentrations in comparison to non-treated cells (Figure 4B).
For induction of apoptotic and/or necrotic cell death in the treated lymphocytes, it was observed that BR-GNPs with the two concentrations, and SR-GNPs, S-GNPs and SC-GNPs with the lower concentration did not significantly increase the ratio of apoptotic and/or necrotic cell death. However, SR-GNPs increased the ratios of apoptotic and necrotic cell death extremely significant (p<0.001) in comparison to the ratios of apoptotic and necrotic cell death in non-treated cells. S-GNPs enhanced necrotic cell death path way and the ratio of necrotic cell death increased with strong significance (p<0.001) in comparison to the ratio of necrotic cell death in non-treated cells. SC-GNPs evoked the programmed cell death with apoptotic pathway with the higher concentration and the ratio of apoptotic cell death increased with strong significance (p<0.001) in comparison to the ratio of apoptotic cell death in non-treated cells (Figure 4B).
Discussion
Preparation and investigation of GNPs with different sizes for cancer photothermal therapy is an important mission to detect the suitable size and concentration of GNPs without inducing cyto- and/or genotoxicity.
Our results found that all shapes of GNPs with variable concentrations induced cytotoxicity in human blood lymphocytes. This cytotoxicity represented by mitotic activity inhibition of blood lymphocytes. Only low concentration of BR-GNPs did not induce cytotoxic effects in lymphocytes. These results were in agreement with the previous works which demonstrated that nanorods did not induce significant toxicity in Hela cells (49, 50). However, high concentrations of nanorods induced cytotoxicity in fibroblasts, epithelial cells and melanoma in concentrator dependent manner (51).
The potential of GNPs to induce cytotoxicity in human blood lymphocytes as demonstrated in this research study may be returned back to the small size of GNPs. The cytotoxicity and inhibition of mitotic activity increased with reducing the diameter of GNPs from 50 nm with BR-GNPs to 30 nm with SR-GNPs or 15 nm with S-GNPs and SC-GNPs. The small sizes of GNPs could give them the ability to diffuse freely through the nuclear membrane ad become in direct contact with DNA. Zhao and Nalwa (52) found that small sized GNPs and silica nanoparticles were collected in nuclei, and returned that back to the potential of these small sized nanoparticles to penetrate nuclear membranes and condensate upon DNA.
The evaluation of caspase gene stability with I-FISH assay demonstrated that GNPs enhanced the duplication of CASP3, CASP7 and CASP9 signals in I-FISH highly significantly. Previous works approved that caspases induce radical changes in apoptotic cells due to activating proteolysis proteins which lead to blocking many substrates such as PAK2, rabaptin-5, gelsolin and focal adhesion kinase (53–56). The most effector caspases is CASP3 which concerns mainly with proteolysis substrates and cleaves inactive DNA fragmentation factors to trigger apoptosis (57–59).
Our data suggests that GNPs may induce apoptotic effects upon human blood lymphocytes especially in higher concentrations. The ratio of duplicated signals of caspases to that of deleted signals reached to ∼3-fold with BR-GNPs and SR-GNPs and to about 10 folds with SC-GNPs especially with CASP3 at higher concentrations.
The significant duplication of caspase genes signals (CASP3, CASP7 and CASP9) in concert with GNPs treatments in our experimental I-FISH extremely increased the apoptotic activity. Chen et al. (60) demonstrated that induction of apoptosis on adenomatous polyposis coli is correlated with increasing caspases expression levels. Also, they mentioned that CASP3 is the main executioner caspases, and its activation depends upon initiator Caspases including CASP8, CASP9 and CASP10. Besides, CASP3 responsible for endonuclease caspase-activated DNase (CAD), which degenerates chromosomal DNA forming chromatin condensation, alters cytoskeleton and disintegrates cells into apoptotic bodies (59).
GNPs affected highly significant on TP53 gene copy numbers with the four types BR-GNPs, SR-GNPs, S-GNPs and SC-GNPs. The ratio of TP53 with gained signals and lost signals was observed to be highly significant in comparing to non-treated cells. The main function of TP53 is to regulate cell growth, enhance DNA repair or induce apoptosis (61). Thus, the percentages of gained signals after treatments with the four types of GNPs could be triggered apoptosis. While the high ratio signal-loss might be due to the failure of TP53 gene to regulate cell growth or enhance DNA repair in damaged cells and consequently induction of apoptosis may be the result.
There was a great reduction in the signals of TNF and CRP genes after treatment with all types of GNPs in comparing to non-treated cells. The ratio of lost signals for these genes was about 3-fold higher than the gain signals. It is known that activation of TNF is correlated with induction of inflammation (38), and enhancing CRP leads to rising in response to infection or tissue injury (62). The potential of GNPs to diffuse through cellular membrane without inducing harmful effect goes together well with the absence of any inflammation in the cells or injury during GNP-passing through the nuclear membrane; thus, they just induced DNA damage as observed as duplicated or lost signals of TNF and CRP genes as demonstrated by I-FISH.
Apoptosis is a form of cell self-murdering and needs specific proteolytic enzymes including caspases (63). Our experimental studies demonstrated that the highly significant duplication of caspases signals as observed with I-FISH could trigger the induction of cell suicide in human blood lymphocytes after treatments with variable shaped and sized GNPs. We confirmed these results with apoptotic-necrotic evaluation. In this experiment, we found that SR-GNPs induced both apoptotic and necrotic cell death highly significantly. While, S-GNPs induced necrotic effects and SC-GNPs induced apoptotic cell death highly significantly at higher concentration in treated cells. This apoptotic effect could result from the potential of GNPs to penetrate nuclear membrane and trigger imbalance in apoptotic specific genes. Some studies mentioned that necrosis may be due to generation of (primary) necrotic cells by extreme trauma which lead to increasing cellular membrane permeability (26).
Previous studies suggested that extreme necrosis produced due to obduracy of apoptotic cells, overload of suicide cells, failure of phagocytosis and progression of early apoptotic cells into late apoptotic cells (secondary necrotic cells) at which the cellular membrane permeability increases (64). This suggestion explains the high ratio of necrotic cells observed in our experiments after treating human blood lymphocytes with variable sized and shaped GNPs. The potential of nanoparticles to induce cell cycle arrest was assessed previously. Silver nanoparticles triggered cell cycle arrest in G2/M phase of human glioblastoma cells (U251), besides it induced DNA damage as recorded with the Comet assay and micronucleus test (65). Also, GNPs arrested cell cycle at S phase as investigated with cell cycle analyzer using flow cytometry, and induced apoptosis in multiple myeloma (U266) cell line, and the late apoptotic cells caused DNA fragmentation (66). These findings explain our results which demonstrated that GNPs triggered apoptosis through increasing signals related to apoptosis such as caspases and TP53 genes rather than other genes as observed with I-FISH analysis. Also GNPs affected severely on cell division as observed with mitotic activity have the potential to affect on human genome in G2/M phase. That is why the proportion of cells reaches to metaphase reduced highly significantly as detected with mitotic index in a concentration dependent manner with all GNPs types.
In conclusion, our research study demonstrated that high concentrations of GNPs reduce mitotic activity and inhibit cell proliferation due to a stop of cell division in human blood lymphocytes. GNPs enhance duplication of caspases and TP53 genes which are involved in apoptosis in human blood lymphocytes. The high overload of dying cells due to apoptosis led to progression of early apoptotic cells into late apoptotic cells or secondary necrotic cells as demonstrated with apoptotic-necrotic assays. Further investigations should be done to investigate the mode of cell death with different shapes of GNPs and their direct effects on human genome.
About the authors

Mona A.M. Abo-Zeid is an Assistant Professor of Cytogenetics at the Genetics and Cytology Department, and a member of the Cancer Biology Laboratory, Center of Excellence for Advanced Sciences, NRC, Egypt. She is a Fellow of the Molecular Cytogenetics Laboratory, Institut für Humangenetik-Universitätsklinikum, Jena, Germany. Mona has published many articles concerning with discoveries for new cancer treatments including the combination of nano-chemotheraputic drugs with photodynamic therapy by photosensitizers and laser irradiation. She has developed bioassays for developing and screening of new cancer chemopreventive drugs. She supervises MD, PhD and Master’s student theses.

Thomas Liehr is a PhD, Dr. h.c. and invited Professor of the Yerevan-State-University, Armenia. He currently works at Institute of Human Genetics, Jena, Germany and is head of the Center for Molecular Cytogenetics. He is interested in small supernumerary marker chromosomes, uniparental disomy, multicolor FISH, chromosomal heteromorphisms, fragile sites, leukemia-research, interphase architecture and chromosomal evolution. He is involved in the international cooperation with Armenia, Brazil, Croatia, Egypt, Portugal, Russia, Serbia and Thailand. He is the editor of Molecular Cytogenetics. He is on the committee of the Permanent Working Group Marker Chromosomes of the European Cytogenetic Association (ECA), and is a member of the European Board of Medical Genetics of the European Society of Human Genetics. He has written four books, 50 book chapters and over 500 journal articles.

Amira received her Dr. Sc. Hum. degree in human Toxicology in 2002 at Faculty of Medicine, Heidelberg University, Germany. Afterwards, she worked for three years in the Biochemistry Department, NRC, Giza, Egypt. Since 2006, she is the group leader of the Cancer Biology Laboratory, Centre of Excellence for Advanced Sciences, NRC, Giza, Egypt. Amira has a long experience in cancer biology, with a special focus on preclinical trials and the influence of therapeutics of the tumor cells/tumor microenvironment using various cell-dependant technologies and animal models.

Mahmoud Zawrah studied Applied Inorganic Chemistry at the Cairo University from 1990 to 1996, specializing in Material Chemistry for his MSc and PhD. He has been Professor of Material Chemistry and Nanotechnology since April 2007 at the National Research Centre (NRC), Cairo, Egypt. His special fields are nanomaterials for different applications such as medical, environmental and structural materials. He has been awarded several national and international prizes. He has published more than 100 articles in international journals and conferences.

Moneeb A.K. Othman assistant researcher and DAAD scholarship holder to study PhD in Germany in Department of Molecular Cytogenetics, Institute of Human Genetics and Anthropology, FSU Jena, Germany. His PhD study concerns with “Cryptic and complex chromosomal aberrations in Acute Leukemia”. He has published several articles in the field of Leukemia
Acknowledgments
This work was financially supported by German Academic Exchange Service (DAAD) in cooperation with the Egyptian Ministry of Higher Education (MoHE) (A1294875).
Conflict of interest statement The authors declare no conflict of interest.
References
1. Kreuter J, Gelperina S. Use of nanoparticles for cerebral cancer. Tumori 2008;94:271–7.10.1177/030089160809400220Search in Google Scholar
2. Yoon KY, Hoon BJ, Park JH, Hwang J. Susceptibility constants of escherichia coli and bacillus subtilis to silver and copper nanoparticles. Sci Total Environ 2007;373:572–5.10.1016/j.scitotenv.2006.11.007Search in Google Scholar
3. Tan WB, Jiang S, Zhang Y. Quantum-dot based nanoparticles for targeted silencing of HER2/neu gene via RNA interference. Biomaterials 2007;28:1565–71.10.1016/j.biomaterials.2006.11.018Search in Google Scholar
4. Su J, Zhang J, Liu L, Huang Y, Mason RP. Exploring feasibility of multicolored CdTe quantum dots for in vitro and in vivo fluorescent imaging. J Nanosci Nanotechnol 2008;8:1174–7.10.1166/jnn.2008.18167Search in Google Scholar
5. Möhlen KH, Beller FK. Use of radioactive gold in the treatment of pleural effusions caused by metastatic cancer. J Cancer Res Clin Oncol 1979;94:81–5.10.1007/BF00405352Search in Google Scholar
6. Rosenberg SJ, Loening SA, Hawtrey CE, Narayana AS, Culp DA. Radical prostatectomy with adjuvant radioactive gold for prostatic cancer: a preliminary report. J Urol 1985;133:225–7.10.1016/S0022-5347(17)48892-6Search in Google Scholar
7. Sau TK, Murphy CJ. Seeded high yield synthesis of short Au nanorods in aqueous solution. Langmuir 2004;20:6414–20.10.1021/la049463zSearch in Google Scholar PubMed
8. Horisberger M, Tacchini-Vonlanthen M. Ultrastructural localization of Kunitz inhibitor on this sections of Glycine max (soybean) cv. Maple arrow by the gold method. Histochemistry 1983;77:37–50.10.1007/BF00496634Search in Google Scholar PubMed
9. Kim F, Connor S, Song H, Kuykendall T, Yang PD. Platonic gold nanocrystals. Angew Chem Int Ed 2004;43:3673–7.10.1002/anie.200454216Search in Google Scholar PubMed
10. Shankar SS, Rai A, Ankamwar B, Singh A, Ahmad A, Sastry M. Biological synthesis of triangular gold nanoprisms. Nat Mater 2004;3:482–8.10.1038/nmat1152Search in Google Scholar PubMed
11. Li JL, Gu M. Gold-Nanoparticle-Enhanced Cancer Photothermal Therapy. IEEE J Sel Top Quant Electronics 2010;16:989–96.10.1109/JSTQE.2009.2030340Search in Google Scholar
12. Patra HK, Banerjee S, Chaudhuri U, Lahiri P, Dasgupta AK. Cell selective response to gold nanoparticles. Nanomed Nanotechnol 2007;3:111–9.10.1016/j.nano.2007.03.005Search in Google Scholar
13. Connor EE, Mwamuka J, Gole A, Murphy CJ, Wyatt MD. Gold nanoparticles are taken up by human cells but do not cause acute cytotoxicity. Small 2005;1:325–7.10.1002/smll.200400093Search in Google Scholar
14. Bartneck M, Keul HA, Singh S, Czaja K, Bornemann J, Bockstaller M, et al. Rapid uptake of gold nanorods by primary human blood phagocytes and immunomodulatory effects of surface chemistry. ACS Nano 2010;4:3073–86.10.1021/nn100262hSearch in Google Scholar
15. Hayden MS, West AP, Ghosh S. NF-κB and the immune response. Review. Oncogene 2006;25:6758–80.10.1038/sj.onc.1209943Search in Google Scholar
16. Sharma M, Salisbury RL, Maurer EI, Hussain SM, Sulentic CE. Gold nanoparticles induce transcriptional activity of NF-κB in a B-lymphocyte cell line. Nanoscale 2013;5:3747–56.10.1039/c3nr30071dSearch in Google Scholar
17. Lewinski N, Colvin V, Drezek R. Cytotoxicity of nanoparticles. Small 2008;4:26–49.10.1002/smll.200700595Search in Google Scholar
18. Alkilany A, Murphy C. Toxicity and cellular uptake of gold nanoparticles; what we have learned so far? J. Nanopart Res 2010;12:2313–2333.10.1007/s11051-010-9911-8Search in Google Scholar
19. Khlebtsov N, Dykman L. Biodistribution and toxicity of engineered gold nanoparticles: a review of in vitro and in vivo studies. Chem Soc Rev 2011;40:1647–71.10.1039/C0CS00018CSearch in Google Scholar
20. Nicholson DW, Thornberry NA. Caspases: killer proteases. Trends Biochem Sci 1997;22:299–306.10.1016/S0968-0004(97)01085-2Search in Google Scholar
21. Porter AG, Ng P, Jänicke RU. Death substrates come alive. Bioessays 1997;19:501–7.10.1002/bies.950190609Search in Google Scholar
22. Wyllie AH. Glucocorticoid-induced thymocyte apoptosis is associated with endogenous endonuclease activation. Nature 1980;284:555–6.10.1038/284555a0Search in Google Scholar
23. Wyllie AH, Kerr JF, Currie AR. Cell death: the significance of apoptosis.Int Rev Cytol 1980;68:251–306.10.1016/S0074-7696(08)62312-8Search in Google Scholar
24. Earnshaw WC. Nuclear changes in apoptosis. Curr Opin Cell Biol 1995;7:337–43.10.1016/0955-0674(95)80088-3Search in Google Scholar
25. Gregory CD, Devitt A. The macrophage and the apoptotic cell: an innate immune interaction viewed simplistically? Immunology 2004;113:1–14.10.1111/j.1365-2567.2004.01959.xSearch in Google Scholar PubMed PubMed Central
26. Bianchi ME. DAMPs, PAMPs, alarmins. All we need to know about danger. J Leukoc Biol 2007;81:1–5.10.1189/jlb.0306164Search in Google Scholar PubMed
27. Majno G, Joris I. Apoptosis, oncosis, and necrosis. An overview of cell death. Am J Pathol 1995:146:3–15.Search in Google Scholar
28. Levin S, Bucci TJ, Cohen SM, Fix AS, Hardisty JF, LeGrand EK, et al. The nomenclature of cell death: recommendations of an ad hoc committee of the society of toxicologic pathologists. Toxicol Pathol 1999;27:484–90.10.1177/019262339902700419Search in Google Scholar PubMed
29. Cohen GM. Caspases: the executioners of apoptosis. Biochem J 1997;326:1–16.10.1042/bj3260001Search in Google Scholar PubMed PubMed Central
30. Rai NK, Tripathi K, Sharma D, Shukla VK. 2005. Apoptosis: a basic physiologic process in wound healing. Int J Low Extrem Wounds 4:138–44.10.1177/1534734605280018Search in Google Scholar PubMed
31. Hu S, Snipas SJ, Vincenz C, Salvesen G, Dixit VM. Caspase-14 is a novel developmentally regulated protease. J Biol Chem 1998;273:29648–53.10.1074/jbc.273.45.29648Search in Google Scholar PubMed
32. Nakagawa T, Zhu H, Morishima N, Li E, Xu J, Yankner BA, et al. Caspase-12 mediates endoplasmic-reticulum-specific apoptosis and cytotoxicity by amyloid-beta. Nature 2000;403:98–103.10.1038/47513Search in Google Scholar
33. Koenig U, Eckhart L, Tschachler E. Evidence that Caspase-13 is not a human but a bovine gene. Biochem Biophys Res Commun 2001;285:1150–4.10.1006/bbrc.2001.5315Search in Google Scholar
34. Kang SJ, Wang S, Kuida K, Yuan J. Distinct downstream pathways of Caspase-11 in regulating apoptosis and cytokine maturation during septic shock response. Cell Death Differ 2002;9:1115–25.10.1038/sj.cdd.4401087Search in Google Scholar
35. Matlashewski G, Lamb P, Pim D, Peacock J, Crawford L, Benchimol S. Isolation and characterization of a human p53 cDNA clone: expression of the human p53 gene. Embo J 1984;3:3257–62.10.1002/j.1460-2075.1984.tb02287.xSearch in Google Scholar
36. Petitjean A, Mathe E, Kato S, Ishioka C, Tavtigian SV, Hainaut P, et al. Impact of mutant p53 functional properties on TP53 mutation patterns and tumor phenotype: lessons from recent developments in the IARC TP53 database. Hum Mutat 2007;28:622–9.10.1002/humu.20495Search in Google Scholar
37. Olivier M, Petitjean A, Marcel V, Petre A, Mounawar M, Plymoth A, et al. Recent advances in p53 research: an interdisciplinary perspective. Cancer Gene Ther 2009;16:1–12.10.1038/cgt.2008.69Search in Google Scholar
38. Idriss HT, Naismith JH. TNF alpha and the TNF receptor superfamily: structure-function relationship(s). Microsc Res Tech 2000;50:184–95.10.1002/1097-0029(20000801)50:3<184::AID-JEMT2>3.0.CO;2-HSearch in Google Scholar
39. Blake GJ, Ridker PM. C-reactive protein: a surrogate risk marker or mediator of atherothrombosis? Am J Physiol Regul Integr Comp Physiol 2003;285:R1250–2.10.1152/ajpregu.00227.2003Search in Google Scholar
40. Hirschfield GM, Pepys MB. C-reactive protein and cardiovascular disease: new insights from an old molecule. Qjm 2003;96:793–807.10.1093/qjmed/hcg134Search in Google Scholar
41. Volanakis JE. Human C-reactive protein: expression, structure, and function. Mol Immunol 2001;38:189–97.10.1016/S0161-5890(01)00042-6Search in Google Scholar
42. Liehr T, Claussen U. Multicolor-FISH approaches for the characterization of human chromosomes in clinical genetics and tumor cytogenetics. Curr Genomics 2002;3:213–35.10.2174/1389202023350525Search in Google Scholar
43. Iourov IY, Vorsanova SG, Yurov YB. Intercellular genomic (chromosomal) variations resulting in somatic mosaicism: Mechanisms and consequences. Curr Genomics 2006;7: 435–46.10.2174/138920206779116756Search in Google Scholar
44. Nikoobakht B, El-Sayed MA. Surface-Enhanced raman scattering studies on aggregated gold nanorods. J Phys Chem A 2003;107:3372–8.10.1021/jp026770+Search in Google Scholar
45. Liehr T, Thoma K, Kammler K, Kammler K, Gehring C, Ekici A, et al. Direct preparation of uncultured EDTA-treated or heparinized blood for interphase FISH analysis. Appl Cytogenet 1995;21:185–8.Search in Google Scholar
46. ISCN 2013. An international system for human cytogenetic nomenclature (2013). In: Shaffer LG, McGowan-Jordan J, Schmid M, editors. Recommendations of the international standing committee on human cytogenetic nomenclature. Published in Karger in Collaboration with ‘Cytogenetic and Genome Research 2013.Search in Google Scholar
47. Giuliano M, Lauricella M, Nassallo E, Carabillo M, Nento R, Tesoriere G. 1998. Induction of apoptosis in human retinoblastoma cells by topoisomerase inhibitors. Invest Ophth Vis Sci 39:1300–11.Search in Google Scholar
48. Gohel A, Mccarthy M, Gronowicz G. Estrogen prevents glucocorticoid-induced apoptosis in osteoblasts in vivo and in vitro. Endocrinology 1999;140:5339–47.10.1210/endo.140.11.7135Search in Google Scholar PubMed
49. Hauck TS, Ghazani AA, Chan WC. Assessing the effect of surface chemistry on gold nanorod uptake, toxicity, and gene expression in mammalian cells. Small 2008;4:153–9.10.1002/smll.200700217Search in Google Scholar PubMed
50. Khan JA, Pillai B, Das TK, Singh Y, Maiti S. Molecular effects of uptake of gold nanoparticles in HeLa cells. Chembiochem 2007;8:1237–40.10.1002/cbic.200700165Search in Google Scholar PubMed
51. Pan Y, Neuss S, Leifert A, Fischler M, Wen F, Simon U, et al. Size- dependent cytotoxicity of gold nanoparticles. Small 2007;3:1941–9.10.1002/smll.200700378Search in Google Scholar PubMed
52. Zhao Y, Nalwa HS. Nanotoxicology: interactions of nanomaterials with biological systems. Los Angeles, CA: American Scientific, 2007.Search in Google Scholar
53. Rudel T, Bokoch GM. Membrane and morphological changes in apoptotic cells regulated by caspase-mediated activation of PAK2. Science 1997;276:1571–4.10.1126/science.276.5318.1571Search in Google Scholar PubMed
54. Cosulich SC, Horiuchi H, Zerial M, Clarke PR, Woodman PG. Cleavage of Rabaptin-5 blocks endosome fusion during apoptosis. EMBO J 1997;16:6182–91.10.1093/emboj/16.20.6182Search in Google Scholar PubMed PubMed Central
55. Kothakota S, Azuma T, Reinhard C, Klippel A, Tang J, Chu K, et al. Caspase-3-generated fragmente of gelsolin: effector of morphological change in apoptosis. Science 1997;278:294–8.10.1126/science.278.5336.294Search in Google Scholar PubMed
56. Wen L-P, Fahrni JA, Troie S, Guan J-L, Orth K, Rosen GD. Cleavage of focal adhesion kinase by caspases during apoptosis.J Biol Chem 1997;272:26056–61.10.1074/jbc.272.41.26056Search in Google Scholar
57. Liu X, Zou H, Slaughter C, Wang X. DFF, a heterodimeric protein that functions downstream of caspase-3 to trigger DNA fragmentation during apoptosis. Cell 1997;89:175–84.10.1016/S0092-8674(00)80197-XSearch in Google Scholar
58. Enari M, Sakahira H, Yokoyama H, Okawa K, Iwamatsu A, Nagata S. A caspase-activated DNase that degrades DNA during apoptosis ans its inhibitor ICAD. Nature 1998;391:43–50.10.1038/34112Search in Google Scholar PubMed
59. Sakahira H, Enari M, Nagata S. Cleavage of CAD inhibitor in CAD activation and DNA degradation during apoptosis. Nature 1998;391:96–9.10.1038/34214Search in Google Scholar PubMed
60. Chen T, Yang I, Irby R, Shain KH, Wang HG, Quackenbush J, et al. Regulation of caspase expression and apoptosis by adenomatous polyposis coli. Cancer Res 2003;63:4368–74.Search in Google Scholar
61. Vousden KH, Lu X. Live or let die: the cell’s response to p53, Nat Rev Cancer 2002;2:594–604.10.1038/nrc864Search in Google Scholar PubMed
62. Volanakis JE, Kaplan MH. Specificity of C-reactive protein for choline phosphate residues of pneumococcal C-polysaccharide. Exp Biol Med 1971;136:612–4.10.3181/00379727-136-35323Search in Google Scholar PubMed
63. Guo X-X, Guo Q, Li Y, Lee SK, Wei X-N, Jin Y-H. Ginsenoside Rh2 induces human hepatoma cell apoptosis via bax/bak triggered cytochrome C release and caspase-9/caspase-8 activation. Int J Mol Sci 2012;13:15523–35.10.3390/ijms131215523Search in Google Scholar PubMed PubMed Central
64. Krysko DV, D’Herde K, Vandenabeele P. Clearance of apoptotic and necrotic cells and its immunological consequences. Apoptosis 2006;11:1709–26.10.1007/s10495-006-9527-8Search in Google Scholar PubMed
65. AshaRani PV, Low Kah Mun G, Prakash Hande M, Valiyaveettil S. Cytotoxicity and genotoxicity of silver nanoparticles in human cells. ACS Nano 2009;3:279–90.10.1021/nn800596wSearch in Google Scholar PubMed
66. Patra HK, Dasgupta AK, Sarkar S, Biswas I, Chattopadhyay A. Dual role of nanoparticles as drug carrier and drug. Cancer Nanotechnol 2011;2:37–47.10.1007/s12645-010-0011-3Search in Google Scholar PubMed PubMed Central
©2015 by De Gruyter
Articles in the same Issue
- Frontmatter
- In this issue
- Guest Editorial and News
- Complement activation-related pseudoallergy: insights into a stress reaction to nanomedicines in blood
- What’s up in nanomedicine?
- Special section CARPA Part 1
- Reviews
- The possible role of factor H in complement activation-related pseudoallergy (CARPA): a failed attempt to correlate blood levels of FH with liposome-induced hypersensitivity reactions in patients with autoimmune disease
- Mini Reviews
- Rodent models of complement activation-related pseudoallergy: Inducers, symptoms, inhibitors and reaction mechanisms
- Pulmonary intravascular macrophages: prime suspects as cellular mediators of porcine CARPA
- Regular contributions
- Critical Review
- Towards nano-diagnostics for bacterial infections
- Review
- Effects of silver nanoparticles on human health
- Original article
- Potential of rod, sphere and semi-cube shaped gold nanoparticles to induce cytotoxicity and genotoxicity in human blood lymphocytes in vitro
Articles in the same Issue
- Frontmatter
- In this issue
- Guest Editorial and News
- Complement activation-related pseudoallergy: insights into a stress reaction to nanomedicines in blood
- What’s up in nanomedicine?
- Special section CARPA Part 1
- Reviews
- The possible role of factor H in complement activation-related pseudoallergy (CARPA): a failed attempt to correlate blood levels of FH with liposome-induced hypersensitivity reactions in patients with autoimmune disease
- Mini Reviews
- Rodent models of complement activation-related pseudoallergy: Inducers, symptoms, inhibitors and reaction mechanisms
- Pulmonary intravascular macrophages: prime suspects as cellular mediators of porcine CARPA
- Regular contributions
- Critical Review
- Towards nano-diagnostics for bacterial infections
- Review
- Effects of silver nanoparticles on human health
- Original article
- Potential of rod, sphere and semi-cube shaped gold nanoparticles to induce cytotoxicity and genotoxicity in human blood lymphocytes in vitro