Response of Sunflower Cultivars to Deficit Irrigation
-
Ali Reza Safahani Langeroodi
 ,
Behnam Kamkar
,
Behnam Kamkar

Abstract
A 2-year study was conducted to determine the response of four sunflower cultivars (Master, Lakumka, Favorite and Berezanski) to different irrigation regimes: 45 (control), 60, 75 and 90% (I1–I4) of maximum allowable depletion (MAD) of available soil water (ASW). Severe deficit irrigation (I3 and I4) significantly decreased water-use efficiency, radiation use efficiency, yield and yield-related components. The results also showed that water deficit, compared to full-irrigated treatment (I1), increased the activity of all antioxidant enzymes in leaves. Master had the highest seed yield in full and limited irrigation as well as the highest antioxidant enzymes content. Sunflower plants extracted most of the soil moisture from 0 to 50 cm soil layer. Therefore, it is recommended that only the 0–50 cm soil profile be considered for scheduling irrigation. Measured and simulated results revealed a special irrigation regime threshold for each cultivar with respect to seed yield. The response rate to irrigation regime ceased in higher irrigation rates as Ijoin (65, 63, 65 and 70% MAD for Master, Lakumka, Favorite and Berezanski, respectively). Moreover, all estimated Ijoin for all cultivars were higher than the current recommendation of 50% MAD. This shows the ability to sustain the yield of these sunflower cultivars in the study region under deficit irrigation. The seed yield of all four cultivars tended to respond similarly to irrigation in both years, but Master was more efficient than the remaining three cultivars with respect to the response to diverse treatments.
Introduction
The objective of well-regulated deficit irrigation is to save water by subjecting crops to periods of moisture stress with minimal effects on yield while also identifying a particular cultivar under local conditions of climate and soil fertility which would allow irrigation scheduling to maximize crop yield and use scarce water resources most efficiently (Panda et al., 2004). Iran has an arid and a semiarid climate (91% of total area), thus drought stress is considered to be one of its main production problems, especially in shallow soils. Sunflower (Helianthus annuus L.) belongs to the Asteraceae family and is one of the most important oil crops due to its high content of unsaturated fatty acids and lack of cholesterol. Although sunflower is moderately tolerant to water stress, its production is greatly affected by drought (Casadebaig et al., 2008). Crop simulation models that predict plant growth, water use and yield are being used to understand the response of crops to the dynamics of climate – plant – water systems, to evaluate physiological traits for genetic yield improvement and to help make decisions that optimize the use of available resources (Soltani et al., 2006; Kamkar et al., 2011), particularly under conditions of salinity (Rauf et al., 2012). There are no reports in the literature analyzing grain yield response in sunflower for the purpose of irrigation crop modeling. In plants under water deficit conditions, an oxidative brush system may be used while the antioxidant system, including antioxidant enzymes such as glutathione peroxidase (GPX), superoxide dismutase (SOD), peroxidase (POX), catalase (CAT), and others, plays a key role in controlling levels of reactive oxygen species (ROS) (Aroca et al., 2003). The level of damage caused by abiotic stress may be limited by enzymatic and non-enzymatic scavengers of free radicals (Foyer and Noctor, 2000; Aroca et al., 2003; Hasanuzzaman et al., 2012). The specific objectives of this study were as follows:
To study the effect of different irrigation schedules on sunflower root zone soil water status, growth, yield parameters, water-use efficiency and response of antioxidant enzyme contents, including GPX, GR, CAT and SOD;
To recommend an effective irrigation water management strategy for sunflower, grown in semi-arid regions, particularly under conditions of water scarcity.
Materials and methods
Experimental site
The experiment was carried out at the Agricultural Research Station of Kabotar Abad Isfahan, Isfahan Province, Iran (47°56′E and 32°31′N; 1,545 m elevation) during 2011 and 2012 on a sandy loam soil having 0.15% total nitrogen content, 0.40 kg ha−1 phosphorus (Hamm et al., 1970), 5.70 kg ha−1 exchangeable potassium (Rhoades, 1982), 1.5% organic matter (Tiessen and Moir, 1993), and a bulk density of 1.45 and 1.50 g cm−3 in the 0–0.50 and 0.50–1.00 m profile, respectively. The soil pH was 7.2. The average field capacity and permanent wilting point of the root zone soil in the crop field were 15.8 and 6.7%, respectively. Water lodging was not observed in the area and the soil water table was deeper than 90 cm. The experimental site has mild rainy winters and dry summers. An overview of experiment conditions is given in Table 1.
Temperature, precipitation, solar radiation and reference evapotranspiration (ET0) for the growing season in 2 years (2011–2012)
| Month | T max (°C) | T min (°C) | Rainfall (mm) | Radiation(MJ m−2 d−1) | ET0(mm per day) | |||||
| First year | Second year | First year | Second year | First year | Second year | First year | Second year | First year | Second year | |
| June | 35.1 | 36.7 | 16.3 | 17.4 | 26.2 | 22 | 23.9 | 24.2 | 5.5 | 6 |
| July | 38.4 | 39.6 | 18.1 | 19.4 | 36.6 | 24 | 22.7 | 23 | 7 | 7.6 |
| August | 36.2 | 37.7 | 17.5 | 17.7 | 32.2 | 26.1 | 21 | 21.8 | 7.5 | 7.8 |
| September | 32.3 | 34.4 | 11.2 | 14.7 | 30.4 | 25.3 | 18.8 | 19 | 6.9 | 7.4 |
| October | 28.1 | 29.5 | 9.1 | 9.8 | 25.9 | 22 | 14.5 | 14.9 | 4.5 | 4.9 |
Field layout and experimental details
A factorial experiment replicated in triplicate was carried out in a randomized complete block design during two consecutive years (2011 and 2012). Factors included sunflower cultivars (Master, Lakumka, Berezanski and Favorite) and four irrigation regimes, which consisted of irrigation scheduling based on maximum allowable depletion (MAD) of the total available soil water (ASW). Each irrigation regime was based on a predefined level of MAD, which was a fixed percentage of the total ASW. Irrigation water was applied whenever the threshold value of MAD for the particular irrigation treatment was attained. The irrigation treatments were assigned as I1–I4 for 45 (control), 60, 75 and 90% MAD of ASW. Three sunflower cultivars have a low oleic content (Master, Lakumka and Favorite) while one has a high oleic content (Berezanski), all four cultivars being the most common over the past 15 years in central Iran (Ataei, 2004).
The area of each plot was 3.6×5 m (18 m2) consisting of 6 rows, 5 m long and 0.60 m apart. The plots were surrounded by dykes, and a 2-m wide strip was left bare between adjacent plots to prevent water percolation. Plots were hand-sown on the 20th and 22nd June in 2011 and 2012, respectively and thinned to a target density of 9 plants m−2 (0.20 m×0.60 m). Weeds were manually controlled during the experiment. In both experiments, triple super phosphate fertilizer was cast broadly before sowing at a rate of 100 kg ha−1. Urea (150 kg ha−1) was applied at planting (one-third of the application) and side-dressed at the vegetative stage (two-thirds of the application). For all four cultivars, the amount of irrigation applied was identical for all irrigation regimes from the day after sowing (DAS) until the complete establishment of sunflower plants (after 8 leaves had formed; Chimenti and Hall, 1993). After this stage, the plots were irrigated according to the aforementioned irrigation regimes.
For estimating soil water storage, the effective root zone of a sunflower crop was considered as 0–1.00 m (Ataei, 2004). Soil moisture was measured gravimetrically. The amount of water applied after attaining the predefined MAD was calculated according to eq. [1] (Panda et al., 2004):
where Vd, Rz, A, FC and WP are the volume of irrigation water, the effective rooting depth, bulk density, surface area of the plot, root zone water storage in the limits of field capacity and wilting point, respectively. Rz was considered as 0.50 m during 45–50 DAS, and as 1.00 m from 50 DAS to maturity (Ataei, 2004).
Total water use (TWU) was managed by using the Excel-based soil water balance tool, which was expressed in terms of water depletion in the effective root zone Dr,i (mm) at the end of each day based on eq. [2] (Todorovic, 2006):
where Dr,i–1 represents root zone depletion at the end of the previous day i–1 (mm), Pi is the effective precipitation on day i (mm), IRi is the net irrigation supply on day i (mm), CRi is the capillary rise on day i (assumed to be zero), ETc,i is crop evapotranspiration (mm), ROi is the runoff on day i (mm) and DPi is the deep percolation on the same day (mm).
The effective rainfall was estimated using a simple rainfall coefficient approach with a value of 0.8 during the whole season. The application efficiency used for estimating net irrigation supply was assumed to be variable (for the applied furrow irrigation method, between 80 and 90%) depending on the time of irrigation, crop cover, etc. Crop evapotranspiration was calculated from reference evapotranspiration (ET0) through the single crop coefficient approach:
where effects of both crop transpiration and soil evaporation are integrated in a single crop coefficient Kc. Kc values were assumed on the basis of the FAO56 data (Allen et al., 1998) checked for the Isfahan province as 0.35 during the initial growing stage and at the time of harvesting and as 1.15 during the mid-season (the period from flowering to maturity). Daily ET0 was also calculated by the FAO Penman–Monteith equation (Allen et al., 1998).
The management of allowable depletion was assumed to be 0.45 of total available water (p = 0.45) during the whole growing cycle as suggested in FAO56 (Allen et al., 1998). However, when the root zone soil water content drops below the predefined threshold, the crop evapotranspiration was adjusted as:
where ETc,adj is the crop evapotranspiration adjusted for water stress, Ks is a dimensionless reduction coefficient (varies from zero to unit) dependent on ASW content and given by:
where TAW is total available water, RAW is readily available water (45% of total), Dr is the root zone soil water depletion and p is the fraction of total available water (0.45).
In the soil water balance calculation, runoff and capillary rise were assumed to be zero while deep percolation was calculated as the surplus of water in the root zone caused by excessive precipitation and/or irrigation. Accordingly, the amount of net irrigation in different irrigation schedules for 45, 60, 75 and 90% MAD for the first year (2011) was 418, 350, 298 and 200 mm; and for the second year (2007) was 438, 381, 310 and 220 mm. More water was applied in the second year than in the first because rainfall was higher in the former (140 mm) than in the latter (110 mm).
Data collection
Crop measurements (leaf area index, dry biomass) and intercepted solar radiation were performed every 2 weeks. The plants were randomly sampled by collecting five representative plants from each plot. Dry biomass was measured after drying in an oven (75°C), until biomass reached constant weight. The intercepted solar radiation was measured by a light bar (LI-190SA) six times during the growing season at midday (solar time) on days when the sky was clear.
Yield components were recorded on five randomly selected plants from each plot. To measure harvest index (HI) as the ratio of seed weight per mature weight of the above-ground plant parts, five plants plot−1 were oven-dried (at 65°C until constant weight during three consecutive days) and weighed. Seed yield was measured by harvesting 8 m2 in the center of each plot. Water use efficiency (WUE) was calculated as seed yield/TWU (FAO, 2000).
The proline content, superoxide dismutase (SOD), glutathione peroxidase (GPX), glutathione reductase (GR) and catalase (CAT) were measured according to Torrelo and Rice (1986), Beyer and Fridovich (1987), Aebi (1984), Rao (1992) and Rotruck et al. (1973), respectively.
Experimental data were statistically analyzed for variance using the SAS system (SAS Institute, 1999). When analysis of variance showed significant treatment effects, the LSD test was applied to compare the means at p < 0.05. Also, significant responses of the various cultivars to irrigation regimes were investigated by regressing seed yield against MAD (I1-I4) using a segmented quadratic-plateau model as follows (eq. [6]); (Soltani et al., 2006; Kamkar et al., 2011).
where W is seed yield (kg ha−1), I is MAD, Ijoin is the joined point or the rate of MAD at which the plateau finished, and a, b, c, and plateau are model coefficients. The regression plateau was used to estimate the maximum yield while the join point of the regression estimated the rate of MAD at which maximum seed yield was achieved. These nonlinear regression coefficients were estimated using the iterative optimization method by the Solver Add-ins tool of Microsoft Excel (2003) to obtain the least squared difference between measured and simulated values by a model.
Results
Soil water content variation and crop evapotranspiration
The results of soil water balance were obtained on a daily basis and are given by means of the root zone soil water depletion as illustrated in Figure 1 for I1–I4. The intensity of water shortage for different treatments was monitored by means of the Ks coefficient, used to reduce ETc and calculated by eq. [5]. Temporal variation of soil moisture observed in experimental plots at 45% MAD is presented in Figure 1. The amplitude of cyclic variation of soil water was high only in the 0–50 cm soil layer and low in other layer (50–100 cm), indicating that most of the water required by the plant was extracted from the top layer under this schedule (Figure 1). The results also revealed that during the latter stages of growth, when roots were full developed, the plant also extracted some water from the lower layer i.e. 50–100 cm (Figure 1).
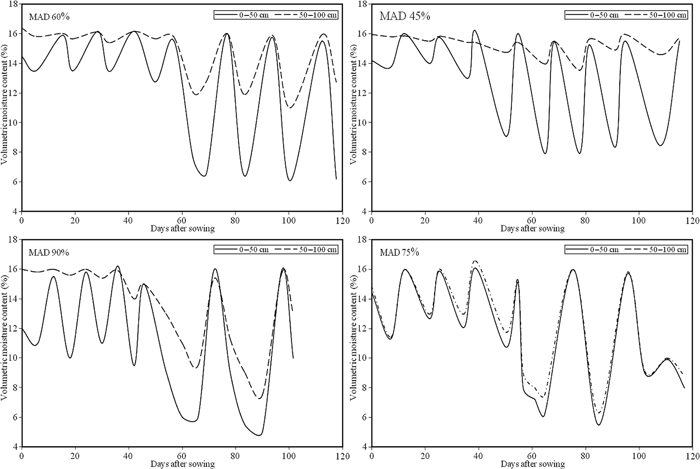
Temporal variation of soil moisture in the root zone of sunflower at: different MAD of available soil water (data averaged over 2 years, 2011–2012)
Under the irrigation schedule with 60% MAD (I2), soil water was extracted from all the layers of the root zone; but most of the extraction was from 0 to 50 cm soil layer (Figure 1). Schedule I2, being a dryer regime than I1, the magnitude of cyclic variation was higher in the 0–50 cm soil layer than in a similar layer in I1. The frequency of irrigation was lower and the volume of water applied was higher under this irrigation schedule than in I1. According to the soil water content, the water deficit started on DAS = 70, reaching maximum depletion on DAS = 100, when Ks (ETc reduction coefficient) was estimated to be 0.6. A high amplitude of cyclic variation was noted under the irrigation schedule with 75% MAD (I3) in the 0–50 cm soil layer of the root zone and low variation was noted in the 50–100 cm soil layer (Figure 1). The frequency of irrigation was lower and the volume of water applied each time was comparatively higher than in I2. In this irrigation schedule, the plant roots penetrated deeper to explore water resources. Considerable soil moisture fluctuation was observed under the 75% MAD (I3) schedule. A high amplitude of cyclic variation was noted in the 0–50 cm soil layer of the root zone and little variation was seen in the 50–100 cm soil layer (Figure 1). Water shortage started on DAS = 53 in which Ks reached 0.76. The fluctuation of soil water was quite high under the 90% MAD (I4) schedule (Figure 1). The 0–50 cm soil layer exhibited higher cyclic variation than the 50–100 cm soil layer. The water was lost through the uppermost soil layer at a faster rate because of the evaporation from the soil surface and transpiration from adult plants. The time span of cyclic variation was higher during I4 than during other treatments. The first indication of water shortage was at 40 DAS, but more intense deficit started on 50 DAS with a Ks value of 0.85.
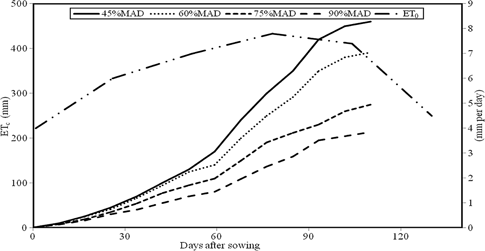
ET0 and cumulative crop evapotranspiration for different irrigation regimes (data averaged over 2 years, 2011–2012)
Crop evapotranspiration (Figure 2) was highest for I1 (460 mm) but it was slightly lower for I2, with a mild water shortage after 70 DAS (400 mm). I3 and I4, with a severe deficit irrigation, had significantly lower crop evapotranspiration, 310 and 213 mm, respectively. After flowering, when crop ET demand was higher and water shortage was more serious, the difference in ETc had become more evident.
Crop growth
The evolution of dry matter (DM) is shown in Figure 3. Greater DM accumulation was observed in the first year due to favorable weather conditions (Table 1). Genotypic differences in above-ground DM occurred among cultivars. Above-ground DM of Lakumka was greater throughout the growing period, followed by Master, Favorite and Berezanski under all irrigation levels (Figure 3). It can be inferred from Figure 3 that DM accumulation in sunflower was positively influenced by the irrigation regimen and followed a sigmoidal pattern. A decrease in total DM by as much as 50% was noticed when irrigation interval increased from 45 to 90% MAD (I1–I4), caused directly by earlier senescence of leaves whose senescence decreased biomass. In addition, not only was early crop growth faster in I1 and I2, but so too did the period of rapid growth last longer in I1 and I2 than in I3 and I4 over both years (Figure 3).
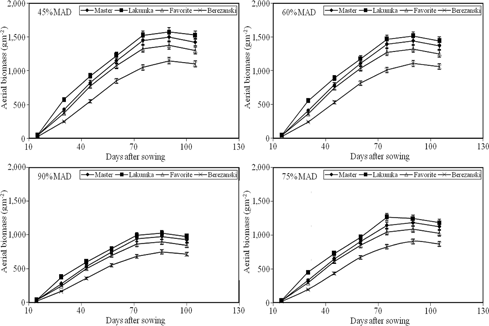
Dry matter accumulation of sunflower as influenced by irrigation regimes and cultivars (data averaged over 2 years, 2011–2012)
Intercepted photosynthesis active radiation and radiation use efficiency
Irrespective of the irrigation regime, the fraction of intercepted radiation (FINT) increased from 15 DAS to 60 DAS and then tended to decline (Figure 4). Maximum FINT values for all irrigation regimes were observed 1 week before anthesis, which is correlated with the highest number of leaves on plants. Maximum FINT values for I1 and I2 were 86% while for I3 and I4 it was lower, 64 and 55%, respectively. After anthesis (about 60 DAS), FINT decreased substantially for all irrigation regimes reaching about 22% for I1 and I2 and 7% for I3 and I4 at harvest. As expected, the greatest values of cumulative IPAR were obtained for I1 and I2 (642.41 and 632.99 MJ m−2, respectively) while related values were 556.4 and 499.8 MJ m−2 for I3 and I4, respectively. Plants irrigated at I1 and I2 produced greater leaf area than plants irrigated at lower levels of irrigation (data not shown) and therefore had greater IPAR.
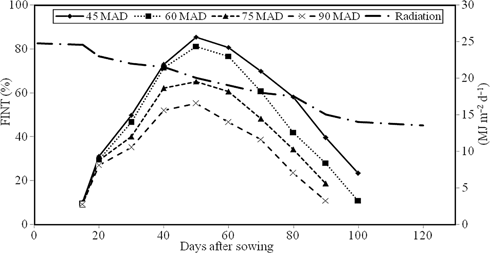
Variations of radiation and fraction of intercepted radiation (FINT) for different irrigation regimes during the sunflower growing season (data averaged over 2 years, 2011–2012)
There was high variation in RUE among treatments with a breakdown of linearity observed at anthesis. In Table 2, the RUE values of pre- and post-anthesis periods, served to distinguish periods determined by regression analysis, are compared. The correlation coefficients for the pre-anthesis period were high and the slopes were similar for all treatments. RUE up to anthesis was 13% lower for I3 and I4 relative to I1. In the post-anthesis period, there was a substantial decrease in biomass in I4 resulting in a negative slope for that treatment. These results show that RUE decreased linearly as the severity of water deficit increased.
The best fitting slope of linear relationship between the biomass and IPAR and the biomass and ETc for different irrigation regimes. Data averaged for 2 years (2011–2012)
| RUE | |||
| Irrigation regime | Slope (g M J−1) | R2 | |
| Pre-anthesis | I1 and I2 | 3.4 | 0.85 |
| I3 and I4 | 2.5 | 0.81 | |
| Post-anthesis | I1 | 1.0 | 0.89 |
| I2 | 0.7 | 0.65 | |
| I3 | 0.3 | 0.71 | |
| I4 | −0.6 | 0.68 | |
| WUE | |||
| Irrigation regime | Slope (kg m−3) | R2 | |
| Initial period | I1 and I2 | 1.37 | 0.99 |
| I3 and I4 | 1.15 | 0.99 | |
| Intensive growth up to anthesis | I1 and I2 | 6.88 | 0.99 |
| I3 and I4 | 5.79 | 0.99 | |
| Post-anthesis | I1 | 1.04 | 0.66 |
| I2 | 0.81 | 0.54 | |
| I3 | 0.39 | 0.59 | |
| I4 | −0.4 | 0.67 |
Biomass and yield water use efficiency
Variation in the relationship between biomass and cumulative ETc during the growing season is presented for different irrigation treatments in Table 2. An evident breakdown of data was observed distinguishing between initial growth stage, intensive crop growth up to anthesis and post-anthesis periods. Consequently, biomass WUE was separately analyzed for each of three above-mentioned stages. The results of biomass WUE values for the three different crop growth periods are presented in Table 2.
The correlation coefficient of regression lines was very high for the first and second stage (from 0.9 to 0.99), while for the third period (post-anthesis) it was much lower. During the initial growth stage, the slopes of regression lines were very similar among irrigation regimes (between 1.15 and 1.37 kg m−3), while in the phase of intensive crop growth, the slope increased (Table 2). The slopes of the post-anthesis period were less consistent. Seasonal biomass WUE for full irrigation treatment (I1) was slightly greater than the values commonly found in the literature. This can be explained by regular irrigations in the vegetative growth stage, which both had an impact on the T/ET ratio and improved WUE. Biomass WUE decreases regularly as the intensity of water shortage increases, similar to RUE.
WUE was established, also referred to yield (YWUE). In this case, the greatest YWUE was observed in I2 (0.71 kg m−3), followed by I1 (0.704 kg m−3), I3 (0.53 kg m−3), and I4 (0.46 kg m−3). The slight increase in WUE in I2 than in I1 was associated with a higher amount of irrigation water and could be due to a greater loss of water by evapotranspiration than the corresponding increase in seed yield. On average, Master exhibited higher WUE (0.64 kg m−3), followed by Favorite (0.61 kg m−3), Berezanski (0.551 kg m−3) and Lakumka (0.551 kg m−3). The difference in WUE can be explained by the fact that cultivars with high WUE also have a high HI.
Analysis of variance for yield, yield components and some antioxidants of sunflower under different irrigation regimes and cultivars in combined over 2 years
| SOV♦ | df | NG | 1,000 SW | BY | HI | SY | CAT | GPX | GR | SOD | Proline |
| Year (Y) | 1 | ns | * | * | ns | * | ns | ns | ns | ns | ns |
| Cultivar (C) | 3 | * | * | * | * | * | ** | ** | ** | ** | * |
| Irrigation (I) | 3 | ** | ** | * | * | ** | ** | ** | ** | ** | * |
| Y × C | 3 | ns | ns | ns | ns | * | ns | ns | ns | ns | ns |
| Y × I | 3 | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| C × I | 9 | ns | * | ns | ns | ns | ** | ** | ** | ** | ns |
| Y × C × I | 9 | ns | ns | ns | ns | ns | ns | ns | ns | ns | ns |
| C.V (%) | 4.5 | 3.2 | 4.9 | 8.3 | 4.2 | 7.1 | 6.6 | 7.8 | 7.4 | 9.2 |
Yield components
The number of grains head−1 is one of the most important yield components in sunflower. Irrigation regime and cultivar significantly affected the number of grains head−1 and 1,000-seed weight (Table 3). In this study, number of grains head−1 decreased by increasing water deficit, with the minimum value (average of 803 grains) observed for I4 (Table 5). The maximum and minimum 1,000-seed weight was obtained at I2 and I4, respectively (Table 4). A significant reduction in the number of filled seeds head−1 resulted from a reduced head diameter and an increase in the unfilled seed percentage with an increase in water shortage (data not shown). Number of grains head−1 was positively correlated with seed yield (r = 0.63, p < 0.01) under all irrigation regimes. Averaged across irrigation regimes, a significant difference in the number of grains head−1 was observed among cultivars (Table 3), as Master and Berezanski achieving the most and fewest grains head−1, respectively (Table 4). 1,000-seed weight was also significantly different between study years (Table 3) and responded differently to growing season and conditions (Table 4). In 2011, rainfall was higher than in 2007, resulting in higher 1,000-seed weight. Among the four cultivars studied, Lakumka had significantly greater 1,000-seed weight (Table 4).
Mean comparison of yield, yield components and proline of sunflower under irrigation regimes and cultivars (2-years average)
| SY•(kg ha−1) | 1,000-SW(g) | BY(kg ha−1) | HI (%) | NG | Proline(μmol g−1) | ||
| Year (Y) | 2011 | 4,448 a | 63.63 a | 13,914 a | 32.18 a | 1,097 a | 38.13 a |
| 2012 | 3,872 b | 60.23 b | 12,195 b | 32.06 a | 1,024 a | 38.40 a | |
| Cultivar (C) | Master | 4,547 a | 57.23 b | 13,780 b | 32.74 a | 1,224 a | 38.47 a |
| Lakumka | 4,410 a | 70.68 a | 14,715 a | 30.90 b | 1,195 a | 35.9 a | |
| Favorite | 4,096 b | 59.00 b | 12,813 c | 32.70 a | 1,111 a | 35.46 a | |
| Berezanski | 3,331 c | 51.67 c | 10,189 d | 32.02 a | 950 b | 30.78 b | |
| Irrigation (I) | I1 | 5,300 a | 67.28 a | 15,657 a | 34.06 a | 1,213 a | 27.07 d |
| I2 | 5,205 a | 68.00 a | 15,190 a | 34.78 a | 1,170 b | 45.87 a | |
| I3 | 3,600 b | 61.64 b | 12,009 b | 30.20 b | 997 c | 38.24 b | |
| I4 | 2,900 c | 54.34 c | 10,029 c | 29.01 c | 803 d | 31.65 c |
Mean comparison of catalase, glutathione reductase, glutathione peroxidaseand superoxide dismutase contents in sunflower different cultivars under irrigation regimes(2-years average)
| Irrigation | Cultivar | GPX (U mg−1 protein) | SOD (U mg−1 protein) | GR (U mg−1 protein) | CAT (U mg−1 protein) |
| I1 | Master | 41.16 a | 79.90 a | 2.19 a | 19.24 a |
| Lakumka | 41.18 a | 78.40 a | 1.88 a | 19.00 a | |
| Favorite | 41.08 a | 77.90 a | 1.93 a | 18.86 a | |
| Berezanski | 40.90 a | 71.70 b | 1.80 a | 18.07 a | |
| I2 | Master | 80.92 a | 145.38 a | 3.25 a | 26.89 a |
| Lakumka | 67.09 b | 143.35 a | 3.11 a | 24.60 b | |
| Favorite | 65.11 b | 133.01 b | 3.01 a | 23.26 b | |
| Berezanski | 56.16 c | 111.13 c | 2.51 b | 21.04 c | |
| I3 | Master | 56.14 a | 99.50 a | 2.84 a | 22.63 a |
| Lakumka | 50.99 b | 95.90 a | 2.61 a | 21.53 a | |
| Favorite | 49.43 b | 94.88 a | 2.46 a | 21.30 a | |
| Berezanski | 43.24 c | 87.32 b | 2.04 b | 20.00 b | |
| I4 | Master | 53.01 a | 88.90 a | 2.50 a | 22.00 a |
| Lakumka | 46.21 b | 85.60 a | 2.31 a | 20.34 b | |
| Favorite | 46.03 b | 82.01 a | 2.30 a | 19.87 bc | |
| Berezanski | 42.85 c | 75.80 b | 1.84 b | 19.45 c |
The effect of irrigation regime and cultivar on HI was also significant (Table 3). The highest HI was observed in I2 (Table 4), but the HI of I1 was not significantly lower than I2, although the values for I3 and I4 were significantly lower (Table 4). As water shortage increases in severity, though, there could be direct effects on HI in many determinate crops. In this study, severe water shortage treatments (I3 and I4) decreased grain yield more than biological yield resulting in a decrease in HI. Only small differences in HI were observed among cultivars (Table 4). Master had greatest HI. However, increasing the supply of water increased the biological yield more prominently than seed yield in Lakumka, as reflected by a lower HI, relative to the high-yielding Master (Table 4).
Antioxidant enzymes and proline
Results from analysis of variance showed that antioxidant enzymes followed similar trends (Tables 3, 5). The content of antioxidant enzymes was affected by water deficit and cultivar as well as the C×I interaction (Table 3). They were increased by increasing water shortage with the highest level being produced under I2 (Table 5). The different content among cultivars was not significant under the well-irrigated condition (I1) (Table 5). Under moderate irrigation deficit, the content of enzymes in cultivars showed a declining order: Master>Lakumka> Favorite>Berezanski. In severe water shortage (I3 and I4), the highest and lowest enzyme content was in Master and Berezanski, respectively. There was a positive and significant relationship between seed yield and GPX, GR and CAT activity under the fully irrigated condition (r=0.78, 0.62 and 0.92; p<0.01, respectively) and under water deficit (r=0.91, 0.73 and 0.69; p<0.01, respectively).
The proline content was affected by water deficit and cultivar (Table 3). As seen in Table 4, there was a significant increase in proline content in stressed leaves. Rewatering decreased the synthesis of proline in all irrigation regimes (data not shown). The proline content among cultivars varied as follows: Master>Lakumka>Favorite>Berezanski (Table 4). There was a positive and significant relationship between seed yield and proline under water deficit (r=0.73, p<0.01), revealing that Master was more drought tolerant than other cultivars because of its high proline content.
Seed yield
As seen in Table 3, there were significant differences between growing season and yield in the first year which was higher than in the second year (Table 4). The higher sunflower yield in 2011 was due to favorable conditions for the growth of sunflower in that year. The analysis of variance on seed yield data showed that the effect of cultivar and irrigation level on this character was significant (p<0.01) (Table 3). Partitioning of the treatment sum of squares revealed that irrigation regime was the major cause of variation in seed yield in these experiments (data not shown), but the interaction effect of these two factors on seed yield was not significant (Table 4). The highest seed yield was possible when irrigation level was high (i.e., I1) (Table 4), which could be attributed to an increase in both grain number and 1,000-grain weight in I1 in both years (Table 4). Seed yield differed significantly among the four cultivars: Master and Lakumka had the highest seed yield (Table 4). The difference in seed yield between the lowest- and highest-yielding cultivars was about 900 kg ha−1.
Deficit irrigation had a pronounced effect on crop yield and yield attributes such as head diameter and 1,000-seed weight, both of which decreased progressively as water deficit increased, and ultimately reduced the seed yield of all four cultivars. However, the drought-sensitive cultivar (i.e. Berezanski) was seriously affected by water shortage and thus seed yield was more reduced under all water shortage conditions compared to the drought-tolerant cultivar (i.e. Master).
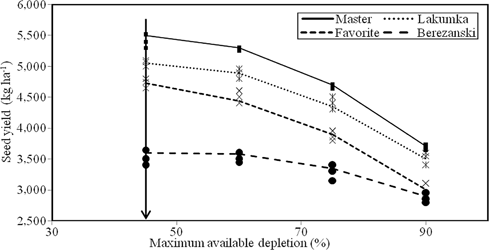
Nonlinear regression output for seed yield produced under different irrigation regimes, averaged data for two experiments, and the vertical solid line indicates the recommended rate of MAD which do irrigation for sunflower production under semiarid conditions in Iran
Seed yield responded to irrigation regime in a curvilinear manner, but the responses were consistent among the four cultivars (Figure 5). The segmented quadratic-plateau model revealed that the reduction in seed yield from I1 to I2 was marginal (i.e. a plateau) due to the marginal effect of moisture stress level between I1 and I2, which was equal to 45–60% MAD (Figure 5). However, the decline in seed yield was more prominent beyond the level of I2, i.e., I3 and I4. This could be due to the fact that a level of soil moisture depletion>60% MAD was sufficient to limit the uptake of water by roots, thereby affecting grain formation. A similar trend was observed during the 2 years. Generally, the response rate to irrigation regime was ceased in higher irrigation rates as Ijoin (65, 63, 65 and 70% MAD for Master, Lakumka, Favorite and Berezanski, respectively; Table 6). Also, all estimated Ijoin for all cultivars were higher than the current recommendation of 50% MAD (Figure 5). This shows that sunflower cultivars could sustain yield in the study region under deficit irrigation (with nearly a 20% decrease in water consumption between recommended irrigation rate and estimated values).
Summary of the nonlinear regression coefficients (a: intercept; b: linear slopet;c: quadratic slopet; Ijoin point, rate of MAD at which the plateau finished; plateau,upper asymptote) for seed yield produced by Master, Lakumka, Favorite and Berezanski,The data were collected from 2 years
| Cultivar | a | b | c | Ijoin | R2 |
| Master | 5,250±175 | 18.632±2.45 | −0.2442±0.06 | 65±6 | 0.98 |
| Lakumka | 5,221±110 | 5.0094±0.912 | −0.1431±0.011 | 63±4 | 0.99 |
| Favorite | 4,920±210 | 3.5285±0.919 | −0.1914±0.065 | 65±8 | 0.97 |
| Berezanski | 3,578±89 | 1.0833±0.194 | −0.0717±0.012 | 70±4 | 0.99 |
The intercept (a value), linear slope (b value), and quadratic slope (c value) coefficients were used to assess the responses of seed yield to irrigation regime for each cultivar (Table 6). The intercept of the regression indicated that maximum yield occurred in most cases when I1 was applied. The inflection point of the regression was used to estimate the water application rate at which maximum seed yield was achieved while the regression plateau was used to estimated maximum yield. There were differences in the linear and quadratic slope coefficients among the four cultivars. Master was the most responsive cultivar to increase water application (highest b value) in terms of seed yield and maximum seed yield was achieved most rapidly (reflected by the more negative quadratic slope coefficient) compared to the other cultivars (Table 6).
Discussion
Our results showed that the activities of antioxidant enzymes increased under mid-irrigation deficit (I2) in comparison to full irrigation while the activities of antioxidant enzymes decreased when sunflower plants suffered extreme water deficit (I3-I4). Previous research showed that enzymatic antioxidant systems, including CAT, GPX, GR and SOD, play an important role in scavenging harmful oxygen species (Singh et al., 2010) and that the activities of these antioxidant enzymes are altered when plants are subjected to stress. Under full irrigation, the content of antioxidant enzymes was often not significantly different among the four cultivars. However, the highest antioxidant enzyme content was observed in Master under water deficit conditions. The correlation results showed that there was a high, positive and significant correlation among yield, CAT and GPX in irrigation deficit. Thus, GPX and CAT content could be considered as a drought tolerance index to select tolerant cultivars under water deficit. Singh et al. (2010) found a significant positive correlation between peroxidase with SOD and CAT. They reported that these antioxidant enzymes could serve as a reliable biochemical marker to identify genotypes with a higher activity of antioxidants such as SOD and CAT. Water deficit stimulated proline accumulation and the extent of proline accumulation was dependent upon the degree of stress applied. It is well established that proline has a protective role in plant growth and productivity by reducing the production of free radicals and/or by scavenging free radicals (Jain et al., 2001). The fact that drought-sensitive cultivar Berezanski tended to accumulate less proline, in contrast to drought-tolerant cultivar Master, under various water deficit conditions, is corroborated by the results of Cechin et al. (2006).
Sridhara and Prasad (2001) showed an increase in total dry matter production in sunflower under optimum irrigation because of the extension of leaf area and its higher durability, providing enough physiological resources to take advantage of received light and therefore produce more DM. The greatest values of accumulated IPAR in I1 and I2, which resulted from higher FINT, could be the main factor affecting dry matter production.
RUE is also a determinant because the fraction of absorbed radiation multiplied by total intercepted radiation multiplied by RUE is equal to dry matter production. Both RUE and WUE slopes were linear and constant until anthesis, which had already been established for several other species (e.g. Sinclair and Muchow, 1999). The change in slope could be explained on the basis of change in the composition of reproductive biomass during seed filling, when oil is the main compound being synthesized (Todorović et al., 2007). Other factors may also explain the decrease in RUE and WUE after anthesis, such as a decrease in leaf nitrogen content, saturation of canopy photosynthesis associated with reduced leaf area, light interception by sunflower heads or increased maintenance respiratory losses due to greater temperatures (Soriano et al., 2004). This change in slope during the crop cycle of sunflower deserves special consideration when modeling crop growth and yield production, especially in those crops having a high energy cost for lipids synthesis during seed filling and for those hybrids having a high early vigor.
Mild deficit irrigation increased the fraction of assimilate allocation to the head, compensating thus the lower number of seeds per head through increased seed weight (Table 4). This consolidates the results of Kram et al. (2007), who reported that the increase in seed weight in stressed sunflower plants was associated with an increase in the fraction of assimilates allocated to the head to the detriment of the translocation to most vegetative organs. The relatively greater fraction of assimilates allocated to the seeds in I2 compared to I1 was associated to a non-significant reduction in seed yield.
The partitioning of dry matter to the head is critical in the process of yield determination in water-stressed sunflower, and the effect of water deficits on the HI of sunflower is complex due to interactions between the timing and intensity of the stress relative to the developmental processes that determine the components of yield (Soriano et al., 2004). Under terminal drought, the HI of grain crops subjected to moderate water deficits is conservative and it is often assumed that HI is a constant fraction of the above-ground biomass under a range of water deficits. Experiments with sunflower in which water deficits were imposed at different growth stages, generated a twofold difference in HI (Soriano et al., 2002), challenging the view that HI may be considered constant over a range of water deficits. Drought during the reproductive stage reduces HI (Soriano et al., 2002). Our results also revealed this trend.
The variability in seed yield in cultivars under water deficit could be attributed to a differential rate of the minimization of water loss that can be achieved either by lowering leaf area, transpiration per unit leaf area (stomatal conductance) or by reducing the energy load of the plant (extinction coefficient) (Soriano et al., 2004), increasing the activity of all antioxidant enzymes and proline.
Deficit irrigation can be used as a tactical measure to reduce irrigation water use when supplies are limited by drought or other factors; it is not known whether it could be used over long time periods. It is imperative to investigate the sustainability of deficit irrigation via long-term experiments and modeling efforts to determine to what extent it can contribute to the permanent reduction of irrigation water use. Overall, seed yield responded to irrigation regimes in a curvilinear manner for all the sunflower cultivars studied. Seed yield increased sharply with increasing water applied rates up to 60% MAD (i.e. I4 to I2), the rate of increase in seed yield plateau as irrigation levels approaching 60% MAD, in which the amount of water was lower than the current recommendation of 45% MAD (Sezena et al., 2011). The magnitude of the responses to water applied rates varied among the cultivars. Increasing the supply of water increased the biological yield more prominently than for seed yield in Lakumka, as reflected by the lower HI (30.9), compared with high-yielding Master (32.74).
However, the WUE of some crops improved significantly due to a reduction in irrigation (Fereres et al., 2007). Zhang et al. (2004) reported WUE values that were higher under deficit conditions than under adequate irrigation, especially when irrigation was applied to critical stages of plant development. A significant reduction in WUE by increasing the irrigation interval more than 60% MAD in this work could be due to a decrease in seed yield by increasing the drought period.
Conclusions
Among studied cultivars, Master had the highest values with respect to above-ground biomass, grain number and grain yield. On the other hand, this cultivar had the highest GPX, GR, CAT and proline content, showing that they could be considered as proper indices to select a cultivar proper for drought-prone environments. Reduced LAI and consequently the fraction of absorbed radiation reduced RUE and HI were all factors affecting the yield in all four cultivars as water shortage severity progressed. Based on our results, deficit irrigation with an approximate threshold of 60% MAD, while considering the 0–50 cm as the target water extraction area, could be considered as a strategic tactic to save water, while maximum WUE is achieved.
Acknowledgments
The authors thank the Research Council of Islamic Azad University, Ahvaz Branch, for providing the necessary facilities. We also thank Dr. Mehdi Karimi, professor of Isfahan University, for his valuable suggestions.
References
Aebi, H., 1984. Catalase in vitro. Methods in Enzymology52: 121–126.10.1016/S0076-6879(84)05016-3Suche in Google Scholar
Allen, R.G., Pereira, L.S., Raes, L.S., Smith, M., 1998. Crop Evapotranspiration-Guidelines for Computing Crop Water Requirements, FAO Irrigation and Drainage Paper 56, Food and Agriculture Organization, Rome.Suche in Google Scholar
Aroca, R., Irigoyen, J.J., Sanchez-Diaz, M., 2003. Drought enhances maize chilling tolerance. II. Photosynthetic traits and protective mechanisms against oxidative stress. Physiologia Plantarum117: 540–549.10.1034/j.1399-3054.2003.00065.xSuche in Google Scholar
Ataei, M., 2004. Effect of row spacing and nitrogen fertilizer on phenology and growth indices of sunflower (Helianthus annuus L.). Ph.D. Thesis, I. A. University Science and Research, Branch-Ahvaz, Iran. (In Persian with English Abstract)Suche in Google Scholar
Beyer, W.F., Fridovich, I., 1987. Assaying for superoxide dismutase activity: some large consequences of minor changes in conditions. Analytical Biochemistry161: 559–566.10.1016/0003-2697(87)90489-1Suche in Google Scholar
Casadebaig, P., Debaeke, P., Lecoeur, J., 2008. Thresholds for leaf expansion and transpiration response to soil water deficit in a range of sunflower genotypes. European Journal of Agronomy28: 646–654.10.1016/j.eja.2008.02.001Suche in Google Scholar
Cechin, I., Rossi, S.C., Oliveira, V.C., Fumis, T.F., 2006. Photosynthetic responses and praline content of mature and young leaves of sunflower plants under water deficit. Photosynthetica44: 143–146.10.1007/s11099-005-0171-2Suche in Google Scholar
Chimenti, C.A., Hall, A.J., 1993. Genetic variation and changes with ontogeny of osmotic adjustment in sunflower (Helianthus annus L.). Euphytica71: 201–210.10.1007/BF00040409Suche in Google Scholar
FAO, 2000. Crops and Drops, Land and Water Development Division, FAO, Rome, Italy, p. 24.Suche in Google Scholar
Fereres, E., Soriano, M.A., 2007. Deficit irrigation for reducing agriculture water use. Journal of Experimental Botany58: 147–159.10.1093/jxb/erl165Suche in Google Scholar PubMed
Foyer, C.H., Noctor, G., 2000. Oxygen processing in photosynthesis: regulation and signalling. New Phytologist146: 359–388.10.1046/j.1469-8137.2000.00667.xSuche in Google Scholar
Hamm, J.W., Radford, F.G., Halstead, E.H., 1970. The simultaneous determination of nitrogen, phosphorous and potassium in sodium bicarbonate extracts of soils. Technicon International Congress. Advances in automatic analysis Vol. 2. Industrial Analysis, Futura Publishing and Co., Mt. Kisco, NY, pp. 65–69.Suche in Google Scholar
Hasanuzzaman, M., Hossain, M.A., Teixeira da Silva, J.A., Fujita, M., 2012. Plant response and tolerance to abiotic oxidative stress: antioxidant defense is a key factor. In: Bandi, V., Shanker, A.K., Shanker, C., Mandapaka, M. (eds.) Crop Stress and Its Management: Perspectives and Strategies, Springer, The Netherlands, pp. 261–315.10.1007/978-94-007-2220-0_8Suche in Google Scholar
Jain, M., Mathur, G., Koul,S., Sarin, N.B., 2001. Amelioration effects of proline on salt stress induced lipid peroxidation in cell lines of groundnut (Arachis hypogaea L.). Plant Cell Reports20: 463.10.1007/s002990100353Suche in Google Scholar
Kamkar, B., Koocheki, A.R., Nassiri Mahallati, M., Teixeira da Silva, J.A., Rezvani Moghaddam, P., Kafi, M., 2011. Fungal diseases and inappropriate sowing dates, the most important reducing factors in cumin fields of Iran, a case study in Khorasan provinces. Crop Protection30: 208–215.10.1016/j.cropro.2010.11.007Suche in Google Scholar
Karam, F., Lahoud, R., Masaad, R., Kabalan, R., Breidi, J., Chalita, C., Rouphael, Y., 2007. Evapotranspiration, seed yield and water use efficiency of drip irrigated sunflower under full and deficit irrigation conditions. Agricultural Water Management90: 213–223.10.1016/j.agwat.2007.03.009Suche in Google Scholar
Panda, R.K., Behera, S.K., Kashyap, P.S., 2004. Effective management of irrigation water for maize under stressed conditions. Agricultural Water Management66: 181–203.10.1016/j.agwat.2003.12.001Suche in Google Scholar
Rao, M.V., 1992. Cellular detoxification mechanisms to determine age dependent injury in tropical plant exposed to SO2. Journal of Plant Physiology140: 7328–7740.10.1016/S0176-1617(11)81031-XSuche in Google Scholar
Rauf, S., Shahzad, A., Teixeira da Silva, J.A., Noorka, I.R., 2012. Biomass partitioning and genetic analyses of salinity tolerance in sunflower (Helianthus annuus L.). Journal of Crop Science and Biotechnology15(3): 53–57.10.1007/s12892-011-0089-0Suche in Google Scholar
Rhoades, J.D., 1982. Soluble salts. In: Page, A. L. et al. (eds.) Methods of Soil Analysis: Part 2: Chemical and Microbiological Properties, Monograph Number 9 (Second Edition). ASA, Madison, WI, pp. 161–179.Suche in Google Scholar
Rotruck, J.T., Pope, A.L., Ganther, H.E, Swanson, A.B, Hafeman, D.G., Hockstra, W.G., 1973. Selenium: Biochemical roles as a component of glutathione peroxidase. Science179: 588–590.10.1126/science.179.4073.588Suche in Google Scholar
SAS Institute, 1999. SAS OnlineDoc. v. 8, SAS Institute, Cary, NC.Suche in Google Scholar
Sezena, S.M., Yazarb, A., Kapurb, B., Tekinb, S., 2011. Comparison of drip and sprinkler irrigation strategies on sunflower seed and oil yield and quality under Mediterranean climatic conditions. Agricultural Water Management98: 1153–1161.10.1016/j.agwat.2011.02.005Suche in Google Scholar
Sinclair, T.R., Muchow, R.C., 1999. Radiation use efficiency. Advances in Agronomy65: 215–265.10.1016/S0065-2113(08)60914-1Suche in Google Scholar
Singh, B.K., Sharma, S.R., Singh, B., 2010. Antioxidant enzymes in cabbage: Variability and inheritance of superoxide dismutase, peroxidase and catalase. Scientia Horticulturae124: 9–13.10.1016/j.scienta.2009.12.011Suche in Google Scholar
Soltani, A., Robertson, M.J., Mohammad-Nejad, Y., Rahemi-Karizaki, A., 2006. Modeling chickpea growth and development: Leaf production and senescence. Field Crops Research99: 14–23.10.1016/j.fcr.2006.02.005Suche in Google Scholar
Soriano, M.A., Orgaz, F., Villalobos, F.J., Fereres, E., 2004. Efficiency of water use of early plantings of sunflower. European Journal of Agronomy21: 465–476.10.1016/j.eja.2004.07.001Suche in Google Scholar
Soriano, M.A., Villalobos, F.J., Fereres, E., 2002. Stress timing effects on sunflower harvest index. In: Villalobos, F.J., Testi, L. (eds.) VII Congress of the European Society for Agronomy. Sevilla, Spain: JA, Consejería de Agricultura y Pesca, pp. 141–142.Suche in Google Scholar
Sridhara, S., Prasad, T.G., 2001. Radiation use efficiency, above ground biomass accumulation, canopy development, leaf area-interception profiles and radiation interception of sunflower (Helianthus annuus L.) as influenced by irrigation regimen. Helia24(35): 101–110.10.1515/helia.2001.24.35.101Suche in Google Scholar
Tiessen, H., Moir, J.O., 1993. Total and organic carbon (wet oxidation-redox titration method). In: Carter, M.R. (ed.) Soil Sampling and Methods of Analysis, Canadian Society of Soil Science, Lewis Publishers, Ann Arbor, MI, pp. 190–191.Suche in Google Scholar
Todorovic, M., 2006. An excel-based tool real time irrigation management at field scale. In: International Symposium on “Water and Land Management for Sustainable Irrigated Agriculture,” Adana, (Turkey), April 4–8.Suche in Google Scholar
Todorović, V., Albrizio, R., Zivotic, L.J., 2007. Deficit irrigation of sunflower under Mediterranean environmental conditions. http://ressources.ciheam.org/om/pdf/b56_1/00800109.pdf. Accessed 23 September 2012.Suche in Google Scholar
Torrelo, W.A., Rice, L.A., 1986. Effects of NaCl stress on proline and cation accumulation in salt sensitive tolerance turfgrass. Plant Soil39: 241–247.10.1007/BF02374226Suche in Google Scholar
Zhang, Y.Q., Kendy, E., Yu, Q., Liu, C.M., Shen, Y.J., Sun, H.Y., 2004. Effect of soil water deficit on evapotranspiration, crop yield, and water use efficiency in the North China Plain. Agricultural Water Management64: 107–122.10.1016/S0378-3774(03)00201-4Suche in Google Scholar
©2014 by Walter de Gruyter Berlin / Boston
This work is licensed under the Creative Commons Attribution-NonCommercial-NoDerivatives 3.0 License.
Artikel in diesem Heft
- Frontmatter
- Research Articles
- Sunflower Genetic Collection at the Vavilov Institute of Plant Industry
- Application of Method Embryo Culture in Combination with Gamma Irradiation and Ultra Sonic (Part II)
- Response of Sunflower Cultivars to Deficit Irrigation
- Hydrothermal Variability and Sunflower Seed Yield in the Humid Tropical Region
- Influence of Subsurface Drip Irrigation System on Growth and Yield of Sunflower (Helianthus annuus L.) in Scarce Rainfall Zone of Andhra Pradesh in Subtropical India
- Heat Tolerance and Adaptability to Drought in Sunflower Can Be Influenced by Pollen Selection
- Use of Protein Spectra in Increasing the Genetic Homogenous Traits of the Parental Lines of Sunflower
- Dynamics of Hybrid Sunflower Disease Resistance
- Quantification of Inulin Content in Selected Accessions of Jerusalem Artichoke (Helianthus tuberosus L.)
- Quality Characteristics of High-Oleic Sunflower Oil Extracted from Some Hybrids Cultivated under Egyptian Conditions
- Obituary
- An Obituary Notice – Prof. Alexandru Viorel Vranceanu, PhD 1927–2014
Artikel in diesem Heft
- Frontmatter
- Research Articles
- Sunflower Genetic Collection at the Vavilov Institute of Plant Industry
- Application of Method Embryo Culture in Combination with Gamma Irradiation and Ultra Sonic (Part II)
- Response of Sunflower Cultivars to Deficit Irrigation
- Hydrothermal Variability and Sunflower Seed Yield in the Humid Tropical Region
- Influence of Subsurface Drip Irrigation System on Growth and Yield of Sunflower (Helianthus annuus L.) in Scarce Rainfall Zone of Andhra Pradesh in Subtropical India
- Heat Tolerance and Adaptability to Drought in Sunflower Can Be Influenced by Pollen Selection
- Use of Protein Spectra in Increasing the Genetic Homogenous Traits of the Parental Lines of Sunflower
- Dynamics of Hybrid Sunflower Disease Resistance
- Quantification of Inulin Content in Selected Accessions of Jerusalem Artichoke (Helianthus tuberosus L.)
- Quality Characteristics of High-Oleic Sunflower Oil Extracted from Some Hybrids Cultivated under Egyptian Conditions
- Obituary
- An Obituary Notice – Prof. Alexandru Viorel Vranceanu, PhD 1927–2014