Functional role of the anterior olfactory nucleus in sensory information processing
-
Renata Medinaceli Quintela
Renata Medinaceli Quintela is a doctoral researcher at the University of Veterinary Medicine Hannover. She holds a bachelor’s and a master’s degree in biology from the RWTH Aachen University with a focus in neuroscience. Since her master thesis, her research has focused on understanding the functional role of cortical feedback in early sensory information processing.
Daniela Brunert studied biology at the Ruhr University in Bochum and received her Ph.D. from the International Graduate School of Neuroscience (IGSN) working on neuromodulation in the mouse olfactory epithelium. She received her postdoctoral training at the University of Florida under the supervision of Barry Ache, where she received a Feodor Lynen Fellowship, and the University of Utah under the supervision of Matt Wachowiak, where she was awarded the Young Investigator Award of the Association for Chemoreception Sciences for her work on serotonergic modulation of the olfactory bulb. She is a postdoc in the lab of Markus Rothermel since 2014, working on olfactory information processing and the interaction of olfactory bulb and cortex.
 and
Markus Rothermel
and
Markus Rothermel
Markus Rothermel studied biology at the Ruhr University Bochum. His doctoral thesis with Hanns Hatt focused on “trigeminal perception” and was funded by scholarships from the research training group GRK 736. For his postdoctoral time, he joined the laboratory of Matt Wachowiak in the US, focusing on information processing in the rodent olfactory system, a project for which he was awarded a DFG research fellowship. In October 2014, he returned to Germany to establish an Emmy Noether research group at the RWTH Aachen University. In 2018, he became a part of the DFG Research Training Group (GRK 2416), followed by an IZKF (Interdisciplinary Center for Clinical Research) joint research project in 2020. In 2021, he accepted a Professorship position for Cellular Biology at the University of Veterinary Medicine Hanover. His main interests are sensory systems in health and disease.


Abstract
Olfaction, despite being evolutionarily one of the oldest senses, is complex in structure and function. It can distinguish between trillions of odorants, provides orientation, mediates social interactions, and serves as a warning system. Sensory signals from the periphery are first processed in the olfactory bulb (OB) and then distributed to several olfactory cortical structures. Unlike other sensory modalities, this primary sensory information is not relayed via the thalamus. One prominent olfactory cortical region is the anterior olfactory nucleus (AON), a two-layered structure located within the olfactory peduncle. The AON exerts strong reciprocal connections not only to the OB but also to higher brain areas, e.g., the piriform cortex (PCx), thereby serving as a hub for bottom-up and top-down information processing. However, the functional role of the AON is not well-understood. Here, we provide an overview of recent publications investigating the function of AON in olfactory processing and behavior and present a framework for future research on this fascinating archicortical structure.
Zusammenfassung
Der Geruchssinn, obwohl er evolutionär zu den ältesten Sinnen zählt, ist komplex in Struktur und Funktion. Er kann Billionen von Geruchsstoffen unterscheiden, gibt Orientierung, vermittelt soziale Interaktionen und dient als Warnsystem. Sensorische Signale aus der Peripherie werden zuerst im olfaktorischen Bulbus (OB) verarbeitet und dann an mehrere kortikale Strukturen verteilt. Im Gegensatz zu anderen sensorischen Modalitäten werden diese primären sensorischen Informationen nicht über den Thalamus geleitet. Eine besonders interessante kortikale Region ist der vordere olfaktorische Kern (AON), eine zweischichtige Struktur, innerhalb des olfaktorischen Pedunkels. Der AON hat starke wechselseitige Verbindungen zum OB, sowie auch zu höheren Hirnarealen, wie z. B. dem piriformen Kortex (PCx), und dient somit als Drehscheibe der olfaktorischen Informationsverarbeitung. Über die genaue funktionale Rolle des AON ist jedoch nur wenig bekannt. Hier geben wir einen Überblick über aktuelle Veröffentlichungen, die die Funktion des AON bei olfaktorischer Verarbeitung und Geruchsverhalten untersuchen, und einen Ausblick in die zukünftige Forschung an dieser faszinierenden archäkortikalen Struktur.
Introduction and objectives
Olfaction is of utmost importance for animals and can influence many behaviors, from social interactions and reproduction to foraging and predator avoidance. Despite its relevance for survival and well-being, olfaction remains one of the least studied sensory modalities. In contrast with other sensory systems, olfactory information is distributed directly to primary sensory cortices, bypassing the thalamus. The output cells of the olfactory bulb (OB), the first relay station of olfactory information in the brain, differentially project to several cortical areas. However, the exact nature of the received information and how it is processed and modulated within different areas is yet to be fully elucidated.
One prominent olfactory region is the anterior olfactory nucleus (AON), the most rostral portion of the olfactory cortex, located between the OB and the piriform cortex (PCx) in an area known as the olfactory peduncle. The AON exhibits a unique connection pattern with strong bilateral reciprocal interactions to olfactory areas as well as strong input from neuromodulatory and limbic centers. The integration of bottom-up and top-down inputs suggests a prominent role of the AON in olfactory processing. Furthermore, it shows histological changes in the earliest phases of neurodegenerative (Murray et al., 2020; Stevenson et al., 2020) as well as neurodevelopmental disorders (Eltokhi et al., 2021), pointing to the AON’s strong potential for translational research. Still, the AON is one of the least understood structures in olfactory processing. Recent advances in neuronal data acquisition and optogenetic techniques have enabled new studies investigating the AON’s functional role in different aspects of olfactory processing, leading to surprising results. Here, we provide a short overview of the recent literature and identify missing pieces of knowledge on this interesting olfactory brain area.
Anatomy and connectivity
The AON possesses neither a nuclear structure nor a characteristic three-layered cortical organization. The two AON layers are organized similarly and harbor principal cells that physiologically and anatomically resemble piriform neurons. Therefore, the AON is sometimes referred to as the anterior olfactory cortex, although AON remains the most widely used terminology.
Since the anatomy of AON has been reviewed extensively in an excellent review by Brunjes et al. 2005, we will only briefly summarize the most important features. Histologically, the AON consists of two zones, pars externa and pars principalis (Brunjes et al., 2005; Valverde et al., 1989). Pars principalis neurons comprise the majority of the AON cell population (Brunjes et al., 2011) and can be further subdivided into four regions based on their cardinal location, pars dorsalis, pars medialis, pars lateralis, and pars ventralis (also often referred to as pars ventroposterioralis). Pars externa and the subregions of pars principalis strongly diverge in terms of cytoarchitecture and connectivity, arguing that they might serve different olfactory processing functions. Similar to the PCx, the olfactory input to AON pars principalis is not topographically organized. By contrast, the sensory activation of pars externa resembles the topography of the OB, at least along the dorsoventral axis, i.e., dorsal OB regions target dorsally located cells in the AON (Schoenfeld and Macrides, 1984; Scott et al., 1985). On the functional side, pars externa was shown to represent changes in odorant concentration, whereas pars principalis does not (Kay et al., 2011). However, how other olfactory stimulus features are encoded within each subdivision is yet to be investigated, and there is, in general, little information about how the AON responds to olfactory stimuli. One study showed that individual AON neurons can be activated by a wide variety of odorants and odor mixtures (Lei et al., 2006) both from the ipsilateral and the contralateral (via the anterior commissure) nostril (Kikuta et al., 2008). The broader response profile of the AON neurons when compared to OB output cells supports the synaptic convergence of differentially tuned OB projection neurons and the hypothesis that complex odor representations or “gestalts” are created at this level (Haberly, 2001).

AON input-output connections. As part of the olfactory cortex, the AON is directly innervated by mitral and tufted cells, the principal neurons of the OB. The AON projects in a top-down fashion back to the OB. The AON is the only area sending projections to both the ipsi- and contralateral OB side. Within the bottom-up sensory pathway, the AON projects to other olfactory cortical structures, most notably the PCx with whom it also shares reciprocal connections. AON activity can be strongly regulated by neuromodulatory input from the basal forebrain and limbic projections originating from hippocampus and amygdala. AON, anterior olfactory nucleus; AOD, AON pars dorsalis; AOM, AON pars medialis; AOL, AON pars lateralis; AOV, AON pars ventralis; AOE, AON pars externa; PCx, piriform cortex; OB, olfactory bulb; MCL, mitral cell layer; HDB, horizontal limb of the diagonal band of Broca.
Besides the input from the OB, AON’s activity can be further shaped by cortical feedback from the PCx, neuromodulatory input from the basal forebrain (Rothermel and Wachowiak, 2014; Zaborszky et al., 2012), and limbic projections from the hippocampus (Aqrabawi and Kim, 2018a; Cenquizca and Swanson, 2007) and the amygdala (Petrovich et al., 1996; Pardo-Bellver et al., 2012) (Figure 1). Also here, differences between AON subdivisions can be observed, e.g., the ventral hippocampus preferentially targets the AON pars medialis. The AON represents the largest source of cortical glutamatergic feedback to the OB, and different subdivisions display distinct projection patterns. Most AON subdivisions project bilaterally, except for pars externa, sending projections almost exclusively to the contralateral OB. Conversely, pars medialis preferentially targets the ipsilateral side. Feedforward projections to the anterior PCx arise predominantly from pars lateralis, whereas the posterior PCx is mainly innervated by pars medialis (Haberly and Price, 1978). Despite these clear input–output differences of the AON subdivisions, so far, most functional studies have treated the AON as a single entity.
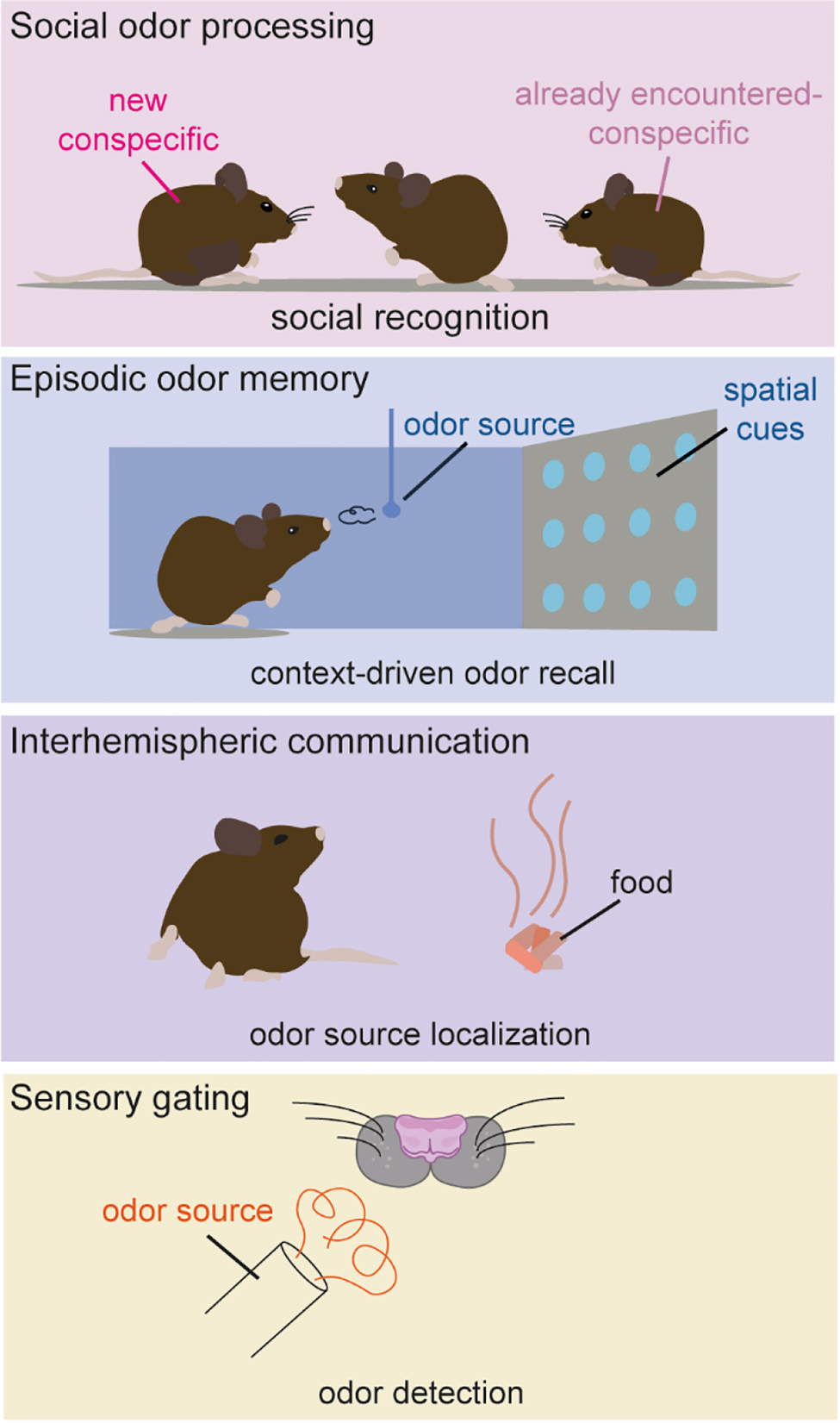
AON modulation of olfactory behaviors. The AON has been implicated in different functions, including regulation of social interactions, recollection of context-dependent odor memories, interhemispheric communication to support odor source localization, and influencing olfactory detection by gating olfactory signal transmission to the cortex.
Functional roles of the AON
In recent years, the AON has been implicated in a variety of functions, mostly reflecting its connectivity within and outside the olfactory system. Here, we briefly introduce behaviorally relevant AON functions, including social odor processing, episodic odor memory, interhemispheric communication, and sensory gating (Figure 2).
Social odor processing
Social behavior, i.e., behavior toward conspecific animals, is mediated predominantly via the accessory olfactory system but several social behaviors could also be attributed to the main olfactory system. One of these behaviors is social recognition, the ability to remember an already encountered animal. Oxytocin and vasopressin are neuropeptides involved in the regulation of socially relevant olfactory behaviors including social recognition (Wacker and Ludwig, 2012). Receptors for both oxytocin and vasopressin have been found in the AON (Tribollet et al., 1988; Vaccari et al., 1998), rendering it a strong candidate for cortical integration of social contexts. Indeed, AON projections to the OB are activated in response to thalamic oxytocin release (Oettl et al., 2016) and are capable of modulating OB output activity. The deletion of AON oxytocin receptors had a detrimental effect on the recognition of conspecifics, supporting that this pathway is both physiologically and behaviorally relevant (Oettl et al., 2016).
Similar to oxytocin, vasopressin signaling in the AON has been implicated in social recognition (Wacker et al., 2011; Wacker and Ludwig, 2019). Whether vasopressin also regulates social processing via AON feedback to the OB remains to be investigated. The presence of vasopressin cells in the OB and the AON has also been reported, and vasopressin OB neurons have also been implicated in mediating social recognition (Suyama et al., 2021; Tobin et al., 2010).
Episodic odor memory
So far, most functional studies treated the AON as a single entity. The study by Aqrabawi et al. in 2016 was the first to focus on a specific AON subdivision (pars medialis), almost exclusively projecting to the ipsilateral OB. The study revealed that the AON is capable of modulating olfactory-dependent behaviors via its OB projections, and that AON activity itself is influenced by limbic input from the ventral hippocampus, demonstrating a hippocampus-AON-OB pathway. Later studies demonstrated that hippocampal projections are not exclusive for pars medialis, but that these innervations are topographically organized, with ventral parts of the hippocampus targeting medial AON portions and dorsal/intermediate areas projecting more laterally in the AON (Aqrabawi and Kim, 2018a). Hippocampal projections were further shown to supply the AON with contextual information necessary for the recollection of odor memories associated with spatiotemporal cues (Aqrabawi and Kim, 2018b; Levinson et al., 2020), proposing the AON as a potential episodic odor memory storage site (Aqrabawi and Kim, 2020). These proposed higher order AON functions are surprising, given that classically the PCx was believed to function as the first real olfactory associative cortex and the site of convergence for olfactory and multimodal information (Haberly, 2001). Only very recently, PCx was shown to also encode spatial information, however, to guide navigation, rather than memory recollection (Poo et al., 2022).
Interhemispheric communication
The AON is the only structure in the olfactory system that sends and receives information from the contralateral hemisphere (Brunjes et al., 2005). These interhemispheric projections might mediate perceptual unity (i.e., odors being perceived as the same, irrespective of the nostril the information is taken up) or direction sensing (i.e., locating odor objects based on differences between the inputs from the two nostrils). Pars externa projecting predominantly to the contralateral OB and the contralateral pars dorsalis (Illig and Eudy, 2009) is thought to interconnect mirror-symmetric OB output neurons, thus building a mechanistic basis for perceptional unity (Grobman et al., 2018). Furthermore, the AON pars externa compares ipsinostril to contranostril odor inputs (Kikuta et al., 2010), enabling odor source localization, and AON activation in awake animals led to a reorientation of the animal’s nose toward an odor source (Esquivelzeta-Rabell et al., 2018). The AONs function in integrating odor and spatiotemporal cues argue for a role in odor orientation and foraging behavior.
Sensory gating
By demonstrating that the activation of pars medialis reduces olfactory detection sensitivity, Aqrabawi et al., 2016, provided first evidence that the AON is capable of gating olfactory signals. Our group reported that the AON activation impaired the trained behavioral response to olfactory stimuli. This strong behavioral inhibition was observed across a wide range of odorants and concentrations. Electrophysiological recordings supported these findings, as AON activation significantly reduced odor-evoked output activity in anesthetized as well as awake mice (Medinaceli Quintela et al., 2020). Thus, the AON seems capable of interfering with olfactory signal transmission to cortex, most likely through the direct activation of local OB inhibitory interneurons and subsequent inhibition of projection neurons (Markopoulos et al., 2012). Since AON projections to the OB are odor-specific (Rothermel and Wachowiak, 2014), their activation may support odor habituation, i.e., the active adaptation to continuously present odorants to favor the detection of novel stimuli (Li, 1990; Li and Hertz, 2000), a process that has been shown to depend on glutamate release onto OB cells (Chaudhury et al., 2010). However, AON projections to the OB are also modulated by input from higher brain areas such as the hippocampus (Aqrabawi et al. 2016) and neuromodulatory inputs from the basal forebrain (Rothermel and Wachowiak, 2014), a region strongly associated with attentional processes. Thus, the basal forebrain-AON-OB pathway might direct attention to relevant stimuli while suppressing the detection of known odorants.
Pathophysiological implications
Olfactory dysfunctions can be observed in neurodevelopmental disorders (Hartig et al., 2021; Tonacci et al., 2017), mental health conditions (Daniels and Vermetten, 2016; Vermetten et al., 2007), and early stages of neurodegenerative diseases. The AON has been linked to Alzheimer’s (Saiz-Sanchez et al., 2010; Saiz-Sanchez et al., 2013) and Parkinson’s disease (Pearce et al., 1995; Ubeda-Banon et al., 2017) in both animal models and postmortem human tissues, where an accumulation of misfolded proteins and cell loss was observed (Pearce et al., 1995). Aggregates were not only restricted to neurons but also found in non-neuronal AON cell types, especially microglia (Stevenson et al., 2020). The AONs contribution to neurodevelopmental disorders and mental health conditions is a hot topic and currently under investigation.
Outlook
We have come a long way from the assumption of the AON being a unimodal feature correlator (Haberly, 2001) to the various higher order functions, as proposed of today. Recent research has implicated the AON in social context, odor memory, directional and interhemispheric sensing, and gating of the OB output. These different functions seem to be at least partially attributable to different AON subsections, e.g., pars externa with a strong connection to interhemispheric information exchange (Grobman et al., 2018) and pars medialis for the integration of contextual information (Aqrabawi and Kim, 2018b). Future studies will help to elucidate these functions further by specifically studying the differences between AON subsections.
Interestingly, many of the so-far proposed AON functions are connected to its projections to the OB, e.g., the role in oxytocin-mediated social recognition and odorant output gating. However, the AON has extensive projections not only to olfactory areas such as the PCx, olfactory tubercle, and lateral entorhinal cortex but also to the hypothalamus, basal forebrain, and striatum (Brunjes et al., 2005). So far, these connections have hardly been examined in a functional context. Doing so will provide further insight into the importance of the AON for behavior.
Increasing our knowledge of the AON is of great importance not only to understand olfactory behavior, but also to reveal its role in pathology including neurodegenerative diseases or neurodevelopmental disorders.
Funding source: Deutsche Forschungsgemeinschaft
Award Identifier / Grant number: 250583768/RO 4046/2-1 and /2-2
Award Identifier / Grant number: 368482240/GRK2416
Award Identifier / Grant number: 445965029/RO 4046/5-1
Funding source: IZKF Aachen
Award Identifier / Grant number: IZKF TN1-7532007
About the authors

Renata Medinaceli Quintela is a doctoral researcher at the University of Veterinary Medicine Hannover. She holds a bachelor’s and a master’s degree in biology from the RWTH Aachen University with a focus in neuroscience. Since her master thesis, her research has focused on understanding the functional role of cortical feedback in early sensory information processing.

Daniela Brunert studied biology at the Ruhr University in Bochum and received her Ph.D. from the International Graduate School of Neuroscience (IGSN) working on neuromodulation in the mouse olfactory epithelium. She received her postdoctoral training at the University of Florida under the supervision of Barry Ache, where she received a Feodor Lynen Fellowship, and the University of Utah under the supervision of Matt Wachowiak, where she was awarded the Young Investigator Award of the Association for Chemoreception Sciences for her work on serotonergic modulation of the olfactory bulb. She is a postdoc in the lab of Markus Rothermel since 2014, working on olfactory information processing and the interaction of olfactory bulb and cortex.

Markus Rothermel studied biology at the Ruhr University Bochum. His doctoral thesis with Hanns Hatt focused on “trigeminal perception” and was funded by scholarships from the research training group GRK 736. For his postdoctoral time, he joined the laboratory of Matt Wachowiak in the US, focusing on information processing in the rodent olfactory system, a project for which he was awarded a DFG research fellowship. In October 2014, he returned to Germany to establish an Emmy Noether research group at the RWTH Aachen University. In 2018, he became a part of the DFG Research Training Group (GRK 2416), followed by an IZKF (Interdisciplinary Center for Clinical Research) joint research project in 2020. In 2021, he accepted a Professorship position for Cellular Biology at the University of Veterinary Medicine Hanover. His main interests are sensory systems in health and disease.
-
Author contributions: All the authors have accepted responsibility for the entire content of this submitted manuscript and approved submission.
-
Research funding: Supported by the Deutsche Forschungsgemeinschaft (DFG, German Research Foundation; 250583768/RO4046/2-1 and /2-2 [MR]; 445965029/RO4046/5-1 [MR]; 368482240/GRK2416 [RMQ, MR]) and the Interdisciplinary Center for Clinical Research within the faculty of Medicine at the RWTH Aachen University Grant IZKF TN1-7 532007 [MR].
-
Conflict of interest statement: The authors declare no conflicts of interest regarding this article.
References
Aqrabawi, A.J., Browne, C.J., Dargaei, Z., Garand, D., Khademullah, C.S., Woodin, M.A. and Kim, J.C. (2016). Top-down modulation of olfactory-guided behaviours by the anterior olfactory nucleus pars medialis and ventral hippocampus. Nat. Commun. 7, 13721.10.1038/ncomms13721Search in Google Scholar PubMed PubMed Central
Aqrabawi, A.J. and Kim, J.C. (2018a). Topographic organization of hippocampal inputs to the anterior olfactory nucleus. Front. Neuroanat. 12, 12, https://doi.org/10.3389/fnana.2018.00012.Search in Google Scholar PubMed PubMed Central
Aqrabawi, A.J. and Kim, J.C. (2018b). Hippocampal projections to the anterior olfactory nucleus differentially convey spatiotemporal information during episodic odour memory. Nat. Commun. 9, 2735, https://doi.org/10.1038/s41467-018-05131-6.Search in Google Scholar PubMed PubMed Central
Aqrabawi, A.J. and Kim, J.C. (2020). Olfactory memory representations are stored in the anterior olfactory nucleus. Nat. Commun. 11, 1246, https://doi.org/10.1038/s41467-020-15032-2.Search in Google Scholar PubMed PubMed Central
Brunjes, P.C., Illig, K.R., and Meyer, E.A. (2005). A field guide to the anterior olfactory nucleus (cortex). Brain Res Brain Res Rev 50, 305–335, https://doi.org/10.1016/j.brainresrev.2005.08.005.Search in Google Scholar PubMed
Brunjes, P.C., Kay, R.B., and Arrivillaga, J.P. (2011). The mouse olfactory peduncle. J. Comp. Neurol. 519, 2870–2886, https://doi.org/10.1002/cne.22662.Search in Google Scholar PubMed PubMed Central
Cenquizca, L.A. and Swanson, L.W. (2007). Spatial organization of direct hippocampal field CA1 axonal projections to the rest of the cerebral cortex. Brain Res. Rev. 56, 1–26, https://doi.org/10.1016/j.brainresrev.2007.05.002.Search in Google Scholar PubMed PubMed Central
Chaudhury, D., Manella, L., Arrellanos, A., Escanilla, O., Cleland, T.A., and Linster, C. (2010). Olfactory bulb habituation to odor stimuli. Behav. Neurosci. 124, 490–499, https://doi.org/10.1037/a0020293.Search in Google Scholar PubMed PubMed Central
Daniels, J.K., and Vermetten, E. (2016). Odor-induced recall of emotional memories in PTSD-Review and new paradigm for research. Exp. Neurol. 284, 168–180, https://doi.org/10.1016/j.expneurol.2016.08.001.Search in Google Scholar PubMed
Eltokhi, A., Gonzalez-Lozano, M.A., Oettl, L.L., Rozov, A., Pitzer, C., Roth, R., Berkel, S., Huser, M., Harten, A., Kelsch, W., et al.. (2021). Imbalanced post- and extrasynaptic SHANK2A functions during development affect social behavior in SHANK2-mediated neuropsychiatric disorders. Mol Psychiatry 26, 6482–6504, https://doi.org/10.1038/s41380-021-01140-y.Search in Google Scholar PubMed PubMed Central
Esquivelzeta-Rabell, J., Mutlu, K., Noutel, J., del Olmo, P.M., and Haesler, S. (2018). Spontaneous rapid odor source localization behavior requires interhemispheric communication. Chem. Senses 43, E34–E35.10.1016/j.cub.2017.04.027Search in Google Scholar PubMed
Grobman, M., Dalal, T., Lavian, H., Shmuel, R., Belelovsky, K., Xu, F.Q., Korngreen, A., and Haddad, R. (2018). A mirror-symmetric excitatory link coordinates odor maps across olfactory bulbs and enables odor perceptual unity. Neuron 99, 800–813, https://doi.org/10.1016/j.neuron.2018.07.012.Search in Google Scholar PubMed
Haberly, L.B. (2001). Parallel-distributed processing in olfactory cortex: new insights from morphological and physiological analysis of neuronal circuitry. Chem. Senses 26, 551–576, https://doi.org/10.1093/chemse/26.5.551.Search in Google Scholar PubMed
Haberly, L.B. and Price, J.L. (1978). Association and commissural fiber systems of the olfactory cortex of the rat. J. Comp. Neurol. 178, 711–740, https://doi.org/10.1002/cne.901780408.Search in Google Scholar PubMed
Hartig, R., Wolf, D., Schmeisser, M.J., and Kelsch, W. (2021). Genetic influences of autism candidate genes on circuit wiring and olfactory decoding. Cell Tissue Res. 383, 581–595, https://doi.org/10.1007/s00441-020-03390-8.Search in Google Scholar PubMed PubMed Central
Illig, K.R. and Eudy, J.D. (2009). Contralateral projections of the rat anterior olfactory nucleus. J. Comp. Neurol. 512, 115–123, https://doi.org/10.1002/cne.21900.Search in Google Scholar PubMed PubMed Central
Kay, R.B., Meyer, E.A., Illig, K.R., and Brunjes, P.C. (2011). Spatial distribution of neural activity in the anterior olfactory nucleus evoked by odor and electrical stimulation. J. Comp. Neurol. 519, 277–289, https://doi.org/10.1002/cne.22519.Search in Google Scholar PubMed PubMed Central
Kikuta, S., Kashiwadani, H., and Mori, K. (2008). Compensatory rapid switching of binasal inputs in the olfactory cortex. J. Neurosci. 28, 11989–11997, https://doi.org/10.1523/jneurosci.3106-08.2008.Search in Google Scholar
Kikuta, S., Sato, K., Kashiwadani, H., Tsunoda, K., Yamasoba, T., and Mori, K. (2010). Neurons in the anterior olfactory nucleus pars externa detect right or left localization of odor sources. Proc Natl Acad Sci U S A. 107, 12363–12368, https://doi.org/10.1073/pnas.1003999107.Search in Google Scholar PubMed PubMed Central
Lei, H., Mooney, R., and Katz, L.C. (2006). Synaptic integration of olfactory information in mouse anterior olfactory nucleus. J. Neurosci. 26, 12023–12032, https://doi.org/10.1523/jneurosci.2598-06.2006.Search in Google Scholar PubMed PubMed Central
Levinson, M., Kolenda, J.P., Alexandrou, G.J., Escanilla, O., Cleland, T.A., Smith, D.M., and Linster, C. (2020). Context-dependent odor learning requires the anterior olfactory nucleus. Behav. Neurosci. 134, 332–343, https://doi.org/10.1037/bne0000371.Search in Google Scholar
Li, Z. (1990). A model of olfactory adaptation and sensitivity enhancement in the olfactory bulb. Biol. Cybern. 62, 349–361, https://doi.org/10.1007/bf00201449.Search in Google Scholar
Li, Z. and Hertz, J. (2000). Odour recognition and segmentation by a model olfactory bulb and cortex. Network 11, 83–102, https://doi.org/10.1088/0954-898x_11_1_305.Search in Google Scholar
Markopoulos, F., Rokni, D., Gire, D.H., and Murthy, V.N. (2012). Functional properties of cortical feedback projections to the olfactory bulb. Neuron 76, 1175–1188, https://doi.org/10.1016/j.neuron.2012.10.028.Search in Google Scholar
Medinaceli Quintela, R., Bauer, J., Wallhorn, L., Le, K., Brunert, D., and Rothermel, M. (2020). Dynamic impairment of olfactory behavior and signaling mediated by an olfactory corticofugal system. J. Neurosci. 40, 7269–7285, https://doi.org/10.1523/jneurosci.2667-19.2020.Search in Google Scholar
Murray, H.C., Dieriks, B.V., Swanson, M.E.V., Anekal, P.V., Turner, C., Faull, R.L.M., Belluscio, L., Koretsky, A., and Curtis, M.A. (2020). The unfolded protein response is activated in the olfactory system in Alzheimer’s disease. Acta Neuropathol Commun 8, 109, https://doi.org/10.1186/s40478-020-00986-7.Search in Google Scholar
Oettl, L.L., Ravi, N., Schneider, M., Scheller, M.F., Schneider, P., Mitre, M., Gouveia, M.D., Froemke, R.C., Chao, M.V., Young, W.S., et al.. (2016). Oxytocin enhances social recognition by modulating cortical control of early olfactory processing. Neuron 90, 609–621, https://doi.org/10.1016/j.neuron.2016.03.033.Search in Google Scholar
Pardo-Bellver, C., Cadiz-Moretti, B., Novejarque, A., Martinez-Garcia, F., and Lanuza, E. (2012). Differential efferent projections of the anterior, posteroventral, and posterodorsal subdivisions of the medial amygdala in mice. Front. Neuroanat. 6, 33, https://doi.org/10.3389/fnana.2012.00033.Search in Google Scholar
Pearce, R.K., Hawkes, C.H., and Daniel, S.E. (1995). The anterior olfactory nucleus in Parkinson’s disease. Mov. Disord. 10, 283–287, https://doi.org/10.1002/mds.870100309.Search in Google Scholar
Petrovich, G.D., Risold, P.Y., and Swanson, L.W. (1996). Organization of projections from the basomedial nucleus of the amygdala: a PHAL study in the rat. J. Comp. Neurol. 374, 387–420, https://doi.org/10.1002/(sici)1096-9861(19961021)374:3<387::aid-cne6>3.0.co;2-y.10.1002/(SICI)1096-9861(19961021)374:3<387::AID-CNE6>3.0.CO;2-YSearch in Google Scholar
Poo, C., Agarwal, G., Bonacchi, N., and Mainen, Z.F. (2022). Spatial maps in piriform cortex during olfactory navigation. Nature 601, 595–599, https://doi.org/10.1038/s41586-021-04242-3.Search in Google Scholar PubMed
Rothermel, M. and Wachowiak, M. (2014). Functional imaging of cortical feedback projections to the olfactory bulb. Front Neural Circuits 8, 73, https://doi.org/10.3389/fncir.2014.00073.Search in Google Scholar PubMed PubMed Central
Saiz-Sanchez, D., De La Rosa-Prieto, C., Ubeda-Banon, I., and Martinez-Marcos, A. (2013). Interneurons and beta-amyloid in the olfactory bulb, anterior olfactory nucleus and olfactory tubercle in APPxPS1 transgenic mice model of Alzheimer’s disease. Anat. Rec. 296, 1413–1423, https://doi.org/10.1002/ar.22750.Search in Google Scholar PubMed
Saiz-Sanchez, D., Ubeda-Banon, I., de la Rosa-Prieto, C., Argandona-Palacios, L., Garcia-Munozguren, S., Insausti, R., and Martinez-Marcos, A. (2010). Somatostatin, tau, and beta-amyloid within the anterior olfactory nucleus in Alzheimer disease. Exp. Neurol. 223, 347–350, https://doi.org/10.1016/j.expneurol.2009.06.010.Search in Google Scholar PubMed
Schoenfeld, T.A. and Macrides, F. (1984). Topographic organization of connections between the main olfactory bulb and pars externa of the anterior olfactory nucleus in the hamster. J. Comp. Neurol. 227, 121–135, https://doi.org/10.1002/cne.902270113.Search in Google Scholar PubMed
Scott, J.W., Ranier, E.C., Pemberton, J.L., Orona, E., and Mouradian, L.E. (1985). Pattern of rat olfactory bulb mitral and tufted cell connections to the anterior olfactory nucleus pars externa. J. Comp. Neurol. 242, 415–424, https://doi.org/10.1002/cne.902420309.Search in Google Scholar PubMed
Stevenson, T.J., Murray, H.C., Turner, C., Faull, R.L.M., Dieriks, B.V., and Curtis, M.A. (2020). alpha-synuclein inclusions are abundant in non-neuronal cells in the anterior olfactory nucleus of the Parkinson’s disease olfactory bulb. Sci. Rep. 10, 6682, https://doi.org/10.1038/s41598-020-63412-x.Search in Google Scholar PubMed PubMed Central
Suyama, H., Egger, V., and Lukas, M. (2021). Top-down acetylcholine signaling via olfactory bulb vasopressin cells contributes to social discrimination in rats. Communications Biology 4, https://doi.org/10.1038/s42003-021-02129-7.Search in Google Scholar PubMed PubMed Central
Tobin, V.A., Hashimoto, H., Wacker, D.W., Takayanagi, Y., Langnaese, K., Caquineau, C., Noack, J., Landgraf, R., Onaka, T., Leng, G., et al.. (2010). An intrinsic vasopressin system in the olfactory bulb is involved in social recognition. Nature 464, 413–U110, https://doi.org/10.1038/nature08826.Search in Google Scholar PubMed PubMed Central
Tonacci, A., Billeci, L., Tartarisco, G., Ruta, L., Muratori, F., Pioggia, G., and Gangemi, S. (2017). [Formula: see text]Olfaction in autism spectrum disorders: a systematic review. Child Neuropsychol. 23, 1–25, https://doi.org/10.1080/09297049.2015.1081678.Search in Google Scholar PubMed
Tribollet, E., Barberis, C., Jard, S., Dubois-Dauphin, M., and Dreifuss, J.J. (1988). Localization and pharmakological characterization of high affinity binding sites for vasopressin and oxytocin in the rat brain by light microscopic autoradiography. Brain Res. 442, 105–118, https://doi.org/10.1016/0006-8993(88)91437-0.Search in Google Scholar
Ubeda-Banon, I., Flores-Cuadrado, A., Saiz-Sanchez, D., and Martinez-Marcos, A. (2017). Differential effects of Parkinson’s disease on interneuron subtypes within the human anterior olfactory nucleus. Front. Neuroanat. 11, 113, https://doi.org/10.3389/fnana.2017.00113.Search in Google Scholar PubMed PubMed Central
Vaccari, C., Lolait, S.J., and Ostrowski, N.L. (1998). Comparative distribution of vasopressin V1b and oxytocin receptor messenger ribonucleic acids in brain. Endocrinology 139, 5015–5033, https://doi.org/10.1210/endo.139.12.6382.Search in Google Scholar PubMed
Valverde, F., Lopez-Mascaraque, L., and De Carlos, J.A. (1989). Structure of the nucleus olfactorius anterior of the hedgehog (Erinaceus europaeus). J. Comp. Neurol. 279, 581–600, https://doi.org/10.1002/cne.902790407.Search in Google Scholar PubMed
Vermetten, E., Schmahl, C., Southwick, S.M., and Bremner, J.D. (2007). Positron tomographic emission study of olfactory induced emotional recall in veterans with and without combat-related posttraumatic stress disorder. Psychopharmacol. Bull. 40, 8–30.Search in Google Scholar
Wacker, D. and Ludwig, M. (2019). The role of vasopressin in olfactory and visual processing. Cell Tissue Res. 375, 201–215, https://doi.org/10.1007/s00441-018-2867-1.Search in Google Scholar PubMed PubMed Central
Wacker, D.W. and Ludwig, M. (2012). Vasopressin, oxytocin, and social odor recognition. Horm. Behav. 61, 259–265, https://doi.org/10.1016/j.yhbeh.2011.08.014.Search in Google Scholar PubMed
Wacker, D.W., Engelmann, M., Tobin, V.A., Meddle, S.L., and Ludwig, M. (2011). Vasopressin and social odor processing in the olfactory bulb and anterior olfactory nucleus. Ann. N. Y. Acad. Sci. 1220, 106–116, https://doi.org/10.1111/j.1749-6632.2010.05885.x.Search in Google Scholar PubMed
Zaborszky, L., van den Pol, A., and Gyengesi, E. (2012) The basal forebrain cholinergic projection system in mice. The Mouse Nervous System. C. Watson, G. Paxinos, and L. Puelles, eds. Amsterdam: (Elsevier), pp. 684–718.10.1016/B978-0-12-369497-3.10028-7Search in Google Scholar
© 2022 the author(s), published by De Gruyter, Berlin/Boston
This work is licensed under the Creative Commons Attribution 4.0 International License.
Articles in the same Issue
- Frontmatter
- Editorial
- Editorial
- Review articles
- Perireceptor events and peripheral modulation of olfactory signals in the olfactory epithelium of vertebrates
- Mammalian social memory relies on neuromodulation in the olfactory bulb
- How the body rules the nose
- Information about space from time: how mammals navigate the odour landscape
- Functional role of the anterior olfactory nucleus in sensory information processing
- How the sense of smell influences cognition throughout life
- Presentation of scientific institutions
- The “MultiSensE” consortium
- Nachrichten aus der Gesellschaft
- Nachrichten aus der Gesellschaft
Articles in the same Issue
- Frontmatter
- Editorial
- Editorial
- Review articles
- Perireceptor events and peripheral modulation of olfactory signals in the olfactory epithelium of vertebrates
- Mammalian social memory relies on neuromodulation in the olfactory bulb
- How the body rules the nose
- Information about space from time: how mammals navigate the odour landscape
- Functional role of the anterior olfactory nucleus in sensory information processing
- How the sense of smell influences cognition throughout life
- Presentation of scientific institutions
- The “MultiSensE” consortium
- Nachrichten aus der Gesellschaft
- Nachrichten aus der Gesellschaft