Unterwegs mit Seehunden – Wie deren Sinnessysteme zu Orientierung, Navigation und Futtersuche beitragen
-
Frederike D. Hanke
Frederike D. Hanke studierte Biologie an der Universität Freiburg, wobei sie sich auf Tierphysiologie/Neurobiologie, Ökologie/Evolution, Genetik und Verhaltensbiologie/Psychologie spezialisierte. Ihren Doktortitel erhielt sie dann von der Internationalen Graduiertenschule für Biowissenschaften der Universität Bochum. Während ihrer Promotion sowie auch in der darauffolgenden Postdoktorandenzeit hatte sie die einmalige Gelegenheit, zahlreiche Projekte im In- und Ausland durchzuführen. Kürzlich vollendete sie erfolgreich ihre Habilitation an der Universität Rostock. Während ihrer wissenschaftlichen Karriere erfreute Frederike Hanke sich daran, mit den unterschiedlichsten und spannenden Tieren wie Honigbienen, Rabenvögeln, Fischen und Bären zu arbeiten. Ihre Hauptversuchstiere sind jedoch Robben und Kopffüßer. In ihrer Forschung legt sie den Schwerpunkt auf deren Sinnessysteme, wobei hierbei hauptsächlich das visuelle System Beachtung findet. Frederike Hanke interessiert sich für vielerlei Aspekte des Sehens so z. B. die Anpassungen des visuellen Systems an die artspezifische Lebensweise und –art bis zu Bewegungs- und Polarisationssehen, visuelle Orientierung und Kognition sowie visuelle Neuroanatomie.

 and
Guido Dehnhardt
and
Guido Dehnhardt
Guido Dehnhardt studierte Biologie und Geografie an der Universität Münster. Bereits während seines Studiums fokusierte er sich stark auf Meeressäuger und besuchte deshalb auch das Kewalo Basin Marine Mammal Laboratory, University of Hawaii, und das Naval Ocean Systems Center. Danach verfolgte er weiter seinen Weg im Rahmen einer Doktorarbeit auf dem Gebiet der Meeressäugerforschung. Seinen Doktortitel erhielt er ebenfalls von der Universität Münster. Nach einer Postdoktorandenzeit an der Universität Bielefeld wechselte Guido Dehnhardt an die Universität Bonn. Während der Bonner Zeit hatte er auch die Gelegenheit, Forschungsprojekte an der University of Joensuu, Finnland, und der University of Newfoundland, Kanada, durchzuführen, bevor er sich dann an der Universität Bonn habilitierte. Jahre intensiver Forschungstätigkeit an der Universität Bochum folgten, woraufhin er dann eine Lichtenberg-Professur der VolkswagenStiftung erhielt, die an der Universität Rostock etabliert wurde. In Rostock gründete er das weithin bekannte Marine Science Center, die größte Robbenforschungsstation weltweit. Guido Dehnhardt interessiert sich in seiner Forschung für sensorische und kognitive Fähigkeiten als Grundlage der Orientierung mariner Säuger. Im Speziellen wurde er bekannt durch seine Erforschung der Funktion der Vibrissen als haptisches und hydrodynamisches System.

Zusammenfassung
Robben sind in ihrem Lebensraum, den Küstenregionen und Meeren, wohlorientiert und zeigen sich als erfolgreiche Jäger. Sowohl auf ihren Wanderungen zwischen Ruheplätzen und Futtergründen als auch beim Aufspüren von Beute leisten die Sinnessysteme der Robben gute Arbeit, obwohl sich die Robben und damit ihre Sinnessysteme in ihrem Lebensraum und aufgrund ihrer amphibischen Lebensweise mit einigen Herausforderungen konfrontiert sehen. In diesem Übersichtsartikel werden die bisher vorhandenen Informationen über die Sinnessysteme der Robben und über spezifische Anpassungen der Sinne an Lebensraum und -weise im ersten Kapitel zusammengetragen und im letzten Kapitel kritisch beleuchtet. Hierbei wird der Fokus auf die Erkenntnisse gelegt, die über die Sinne des Seehunds erhalten wurden, da der Seehund durch intensive Erforschung in den letzten Jahrzehnten zum Modellorganismus innerhalb der Robben hinsichtlich der Sinnesorgane wurde. Im zweiten und dritten Kapitel werden dann die sensorischen Grundlagen in Bezug zu Orientierung, Navigation und Futtersuche gesetzt. Dies ermöglicht auch das Aufzeigen von Fragenstellungen, z. B. wie und wo die Informationen einzelner Sinne integriert werden.
Summary
Seals and sea lions are well-oriented in their habitat, the coastal regions and oceans, and are, moreover, successful hunters. During their movements between haul-out places and foraging grounds as well as during foraging, the sensory systems of seals and sea lions provide useful information, although the animals, and thus their sensory systems, face considerable challenges in their habitat and due to their amphibious lifestyle. In this review, in the first chapter, we compiled and later (chapter 4) discuss the information on the senses of seals and sea lions in general and their specific adaptations to habitat and lifestyle in particular. We hereby focus on the senses of harbor seals. Harbor seals turned into a model organism regarding the sensory systems due to intensive sensory research of the last decades. In the second and third chapter, the sensory basics are put into the context of orientation, navigation, and foraging. This allows formulating new research questions, such as where and how the information from different senses is integrated.
Robben besiedeln die Küstengebiete und Meere in weiten Teilen der Welt. An den deutschen Küsten finden sich Seehunde und Kegelrobben. Durch das Anbringen von Sendern auf der Schulterregion der Tiere können deren Bewegungen zwischen Küste und offenem Meer nachvollzogen werden. Die so erhobenen Daten zeigen, dass Seehunde teils bis zu 50 km weit aufs offene Meer hinausschwimmen und auch mehrere Tage unterwegs sein können (Thompson und Miller, 1990). Ein interessantes Bewegungsmuster konnten Forscher bei einigen Kegelrobben im Ärmelkanal beobachten: Die Tiere überquerten den Kanal in knapp zwei Tagen auf direktem Weg (Chevaillier et al., 2014). Ganz allgemein enthüllen diese Studien, dass die Robben in ihrem Lebensraum wohlorientiert sind; sie verlassen ihre Ruheplätze im Küstenbereich, schwimmen hinaus aufs offene Meer, wo sie ihre Beute jagen, und kehren gezielt zu ihren Ruheplätzen zurück. Obwohl diese Bewegungsmuster bereits seit Jahren aufgezeichnet werden, ist das Verständnis der zugrunde liegenden Mechanismen der Orientierung und Navigation sowie der Futtersuche nach wie vor rudimentär. Die genannten Verhaltensleistungen erfordern Informationen über die Umwelt, die die Sinnessysteme liefern können. Aus diesem Grund wurden in den letzten Jahrzehnten die sensorischen Fähigkeiten der Robben intensiv erforscht. In diesem Übersichtsartikel sollen nun die Erkenntnisse dieser sensorischen Experimente zusammengefasst, diskutiert und im Hinblick auf Orientierung, Navigation und Futtersuche zusammengetragen und weitergedacht werden. Hierbei liegt der Fokus auf den Fähigkeiten der Seehunde, da sie durch die Forschung der letzten Jahrzehnte bezüglich der angesprochenen Themen zum Modellorganismus wurden.
1 Die Sinnessystemen der Seehunde
1.1 Visuelles System
Seehunde besitzen sehr große Augen mit einem Durchmesser von ~ 40 mm im erwachsenen Tier (Jamieson und Fisher, 1972; Abb.1a-c). Das menschliche Auge weist im Vergleich hierzu nur einen Durchmesser von ~ 24 mm auf (Augusteyn et al., 2012), es sitzt aber in einem im Vergleich zum Seehundschädel in Länge, Breite und Höhe etwas größeren Schädel. Die Augen der Seehunde zeigen spezifische Anpassungen an die amphibische Lebensweise, an den kontinuierlichen Wechsel zwischen Wasser und Land, sowie an den Wechsel zwischen hohen und niedrigen Umgebungshelligkeiten (Hanke et. al., 2009a). Die Hornhaut (Cornea) des Seehundauges weist einen hohen Grad an Astigmatismus auf (Hanke et al., 2006a; Jamieson, 1971; Abb.1c), d. h., dass die Brechkraft im vertikalen und horizontalen Meridian nicht identisch ist. Dieser Astigmatismus liegt in einer Abflachung der Hornhaut in der Vertikalen begründet. Zusammen mit der vertikalen Spaltpupille (Abb.1a) scheint diese corneale Abflachung das Hauptproblem der amphibischen Lebensweise zu lösen, den optischen Verlust der Brechkraft der Cornea unter Wasser. Bei hoher Umgebungshelligkeit, wenn die Spaltpupille zu einem vertikalen Spalt geschlossen ist (Abb.1a), fällt nur Licht durch die corneale Abflachung ins Auge. Aufgrund der Abflachung wird dieses Licht nur schwach an der Cornea gebrochen, sodass diese optische Situation in Luft der unter Wasser gleicht. Messungen der Sehschärfe bestätigten in der Tat, dass bei hoher Umgebungshelligkeit in Luft eine Sehschärfe vorliegt, die mit 5,6 Zyklen/Grad mit der unter Wasser vergleichbar ist (Hanke und Dehnhardt, 2009; Jamieson und Fisher, 1970; Schusterman und Balliet 1970; Weiffen et al., 2006). Während in Luft die Sehschärfe vor allen Dingen durch die Umgebungshelligkeit bestimmt wird, unterliegt die Sehschärfe unter Wasser auch dem Einfluss der Trübheit; steigt die Trübheit, so nimmt die Sehschärfe deutlich ab (Weiffen et al., 2006).
Die Netzhaut (Retina) der Seehunde wird durch die Stäbchen dominiert (Jamieson und Fisher 1971; Landau und Dawson 1970; Peichl et al., 2001; Peichl und Moutairou, 1998). Die weniger lichtempfindlichen Photorezeptoren, die Zapfen, liegen nur in geringer Zahl vor; bei einer dem Seehund eng verwandten Robbenart, der Ringelrobbe (Phoca hispida), waren ca. 1,5 % der Photorezeptoren Zapfen (Peichl und Moutairou, 1998). Interessanterweise weisen alle Meeressäuger, bis auf die Seekühe (Cohen et al., 1982; Newman und Robinson, 2006), nur einen Zapfentyp auf. Dieser wird von Licht im Grünbereich maximal erregt (Crognale et al., 1998; Fasick und Robinson, 2000; Lavigne und Ronald, 1975; Levenson et al., 2006; Newman und Robinson, 2005). Hieraus resultiert, dass die Seehunde zum zapfenbasierten Farbensehen, wie es der Mensch oder auch viele Säuger besitzen, nicht in der Lage sind. Auch ein Farbensehen, basierend auf Stäbchen und Zapfen unter mesopischen Helligkeiten, wenn sowohl Stäbchen als auch Zapfen aktiv sind, konnte experimentell nicht nachgewiesen werden (Scholtyssek et al., 2015). Demnach kann die Multifokalität der kugelförmigen Linse (Hanke et al., 2008b), d. h., dass die Linse distinkte Bereiche besitzt, die Licht unterschiedlicher Wellenlänge unterschiedlich stark bricht, keine Anpassung an das Farbensehen sein. Eine Multifokallinse könnte alternativ eine Erhöhung der Tiefenschärfe bei geringen Umgebungshelligkeiten, wenn die Pupille kreisrund geweitet ist (Abb. 1b), bewirken.
Der Vorteil einer stäbchendominierten Retina ist, dass sie das Auge sehr lichtempfindlich macht. Dies ist für die Seehunde unter Schwachlichtbedingungen entscheidend; eine Bedingung, die sie erfahren, wenn sie nachts unterwegs sind oder wenn sie in die Tiefe tauchen. Parameter, die zusätzlich die Empfindlichkeit des Auges erhöhen, sind: (a) Das hinter der Retina liegende Tapetum lucidum (Jamieson und Fisher, 1971; Johnson, 1901; Walls, 1942), welches Photonen, die bei der Passage der Retina nicht absorbiert wurden, reflektiert, und somit eine Absorption bei der zweiten Passage der Retina ermöglicht; (b) Die Pupille (Hanke et al., 2009a; Levenson und Schusterman, 1997), die sich von einer kleinen Lochblende über einen vertikalen Spalt (Abb. 1a) kreisrund (Abb. 1b) weiten kann. Hierüber kann während des Tauchens sehr schnell eine Anpassung an die Lichtverhältnisse in der Tiefe stattfinden (Levenson und Schusterman, 1999).
Auf der Ebene der retinalen Ganglienzellen, die Nervenzellen, die die optische Information an höhere Gehirnzentren weiterleiten, findet sich ein visueller Streifen (Abb. 1d), womit Ereignisse entlang einer Horizontalen, etwa der Wasseroberfläche oder des Meeresgrunds, mit erhöhter Auflösung abgebildet werden (Hanke et al., 2009b). Es findet sich jedoch auch eine Stelle schärfsten Sehens, über die den Seehunden eine Auflösung vermittelt wird, die mit der im Verhaltensexperiment bestimmten Auflösung recht gut übereinstimmt (Hanke und Dehnhardt, 2009; Hanke et al., 2009b; Jamieson und Fisher, 1970; Schusterman und Balliet, 1970; Weiffen et al., 2006). Im optischen Nerv, der von den Axonen der Ganglienzellen gebildet wird, finden sich bei Seehunden ∼200.000 Axone (Pütter, 1903; Wohlert et al., 2016). Eine erste Untersuchung des Kreuzungsmusters der optischen Nerven des rechten und linken Auges am optischen Chiasma des Seehunds (Kröger, 2012) legt eine partielle Kreuzung ähnlich der der Klappmütze (Cystophora cristata), ebenfalls eine Hundsrobbenart, nahe (Cystophora cristata; Dohmen et al., 2015).
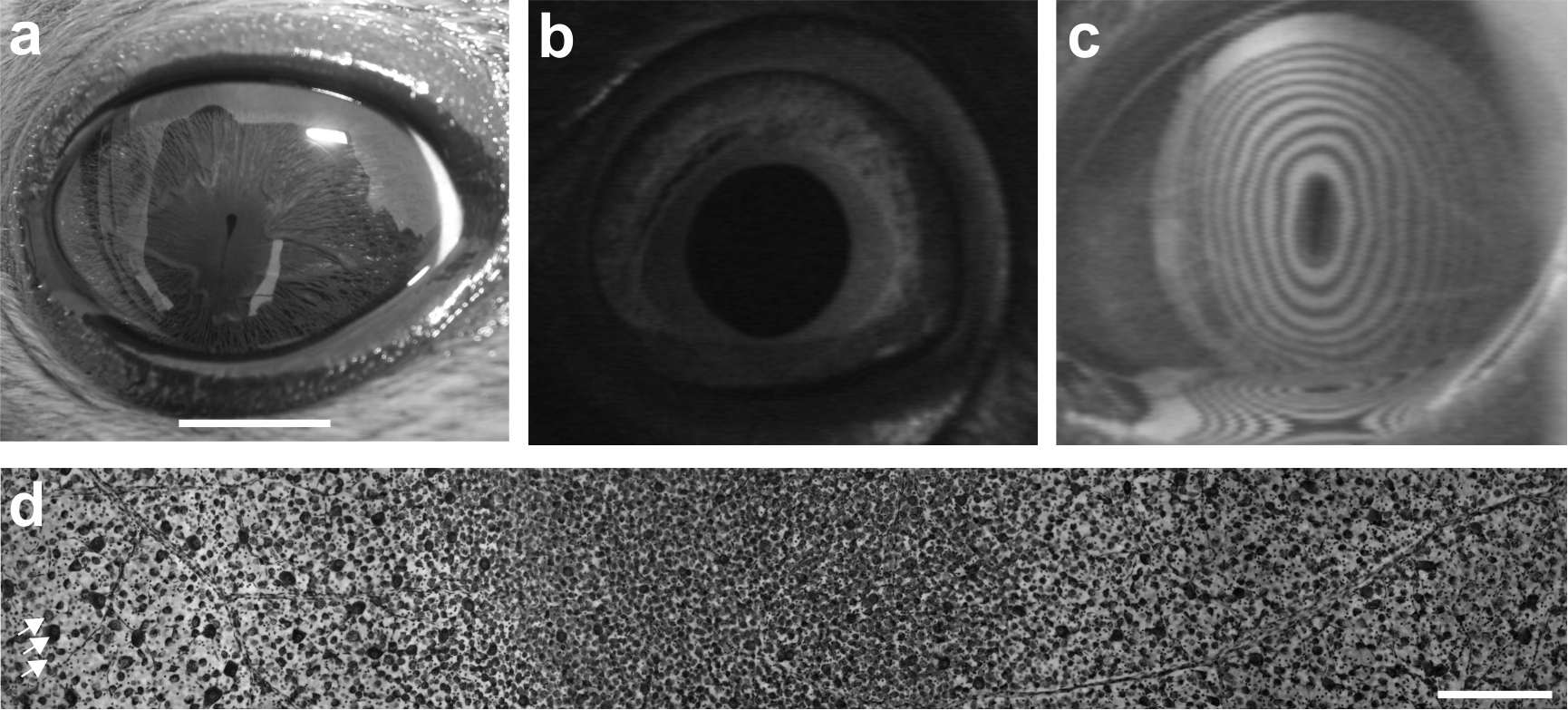
Das visuelle System der Seehunde. a Ein Seehundauge mit geschlossener Pupille bei hohen Umgebungshelligkeiten. Maßstab 10 mm. b Ein Seehundauge mit geweiteter Pupille in Dunkelheit. c Visualisierung des Astigmatismus der Seehundhornhaut mit einer Placido-Scheibe, deren kreisrunde Ringe in der Reflektion aufgrund des starken Astigmatismus ellipsoid verzerrt sind. d Ganglienzellverteilung der Seehundretina. In einem zentralen horizontalen Streifen findet sich eine erhöhte Ganglienzellverteilung, eine hohe Zelldichte (visueller Streifen; in dieser Abbildung mittig zu sehen). Dorsal ist rechts, ventral links, temporal oben, nasal unten. Am linken Rand der Abbildung wurden drei Ganglienzellen mit Pfeilen markiert. Maßstab 250 µm.
Die Augen weisen eine beträchtliche Beweglichkeit auf, welche das visuelle Feld vor allen Dingen nach dorsal stark erweitert (Hanke et al., 2006b). Zusätzlich stabilisieren sie äußere Bewegung (Hanke et al., 2008a). Seehunde haben ein ausgeprägtes Bewegungssehen, sie sind sehr empfindlich für großflächig bewegte Reize (Weiffen et al., 2014), die z. B. auftreten, wenn sich ein Seehund durch eine Partikelwolke hindurch bewegt und damit ein optisches Flussfeld erzeugt. Für Seehunde konnte gezeigt werden, dass sie optischen Fluss wahrnehmen können (Gläser et al., 2014). Dies erfordert ein Umdenken hinsichtlich im Wasser gelöster Partikel, die bisher nur als Sicht einschränkend angesehen wurden. Entgegen dieser Meinung stellt eine Bewegung durch eine Partikelwolke den Tieren eine reiche Quelle an optischer Information, optischen Fluss, zur Verfügung, der für einen Vielzahl an Tätigkeiten genutzt werden kann. Hierzu zählt z. B. das Vermeiden von Kollisionen mit Objekten, eine Fähigkeit die z. B. bei Insekten dokumentiert wurde (Serres und Ruffier, 2017).
1.2 Mechanorezeption
1.2.1 Haptik und Hydrodynamik
Seehunde besitzen Vibrissen im Schnauzenbereich (mystaziale Vibrissen), auf der Nase (rhinale Vibrissen) und oberhalb der Augen (supraorbitale Vibrissen; Abb. 2a). Die Vibrissen der Seehunde, wie allgemein die Vibrissen der meisten Hundsrobben, sind unduliert, d. h. das Haar weist eine Wellenstruktur auf (Hanke et al., 2010; Miersch et al., 2011). Aufgrund dieser Wellenstruktur lösen sich, wenn die Vibrisse durch das Wasser bewegt wird, Wirbel hinter der Vibrisse ab, diese werden jedoch bei Entstehung gleich wieder zerstört. Aus diesem Grund beginnt die Vibrisse nicht zu vibrieren (Hanke et al., 2010; Miersch et al., 2011). Dies ist bei der Detektion von äußeren Ereignissen durchaus von Vorteil; so muss die Eigenbewegung der Vibrisse, sprich des Sensors, nicht herausgerechnet werden, sondern das äußere Ereignis kann direkt gemessen werden.
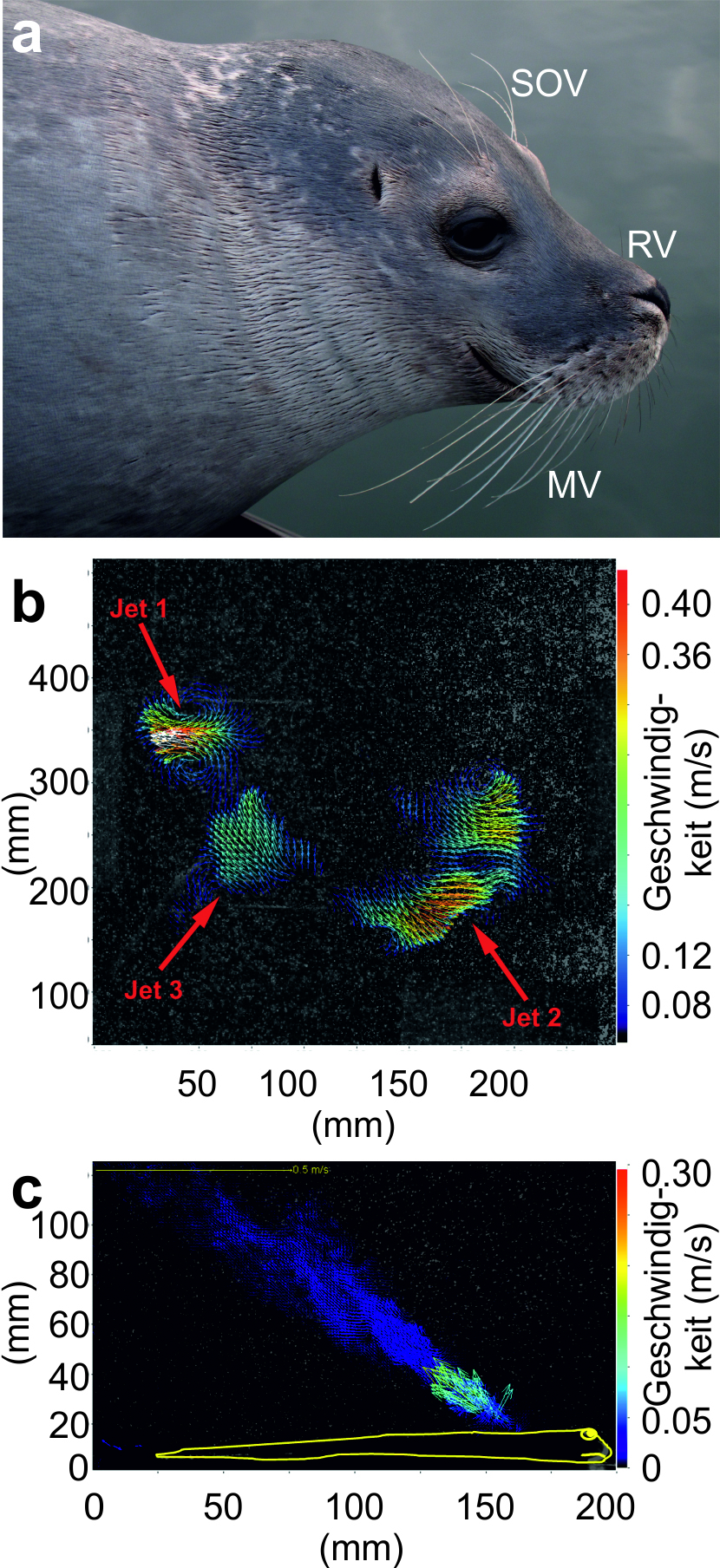
Das Vibrissensystem der Seehunde. a Seehunde besitzen supraorbitale Vibrissen (SOV), ein paar rhinale Vibrissen (RV) und mystaziale Vibrissen (MV). b Fluchtreaktion eines Fisches. Die Wasserbewegungen werden visualisiert, indem die Bewegung von angestrahlten Partikeln im Wasser über die Zeit aufgezeichnet und ausgewertet wird. Die Pfeile geben die Richtung der Wasserbewegung an, die Farbe die Geschwindigkeit in m/s; diese Konventionen gelten auch für Teilabbildung c. Bei dieser Fluchtreaktion, einem C-Start, bei dem der Fisch mit einer C-förmigen Körperbewegung flüchtet, sieht man bei Jet 1 sehr deutlich, dass ein einzelner Wirbelring entsteht, der von den Seehunden wahrgenommen und hinsichtlich seiner Bewegungsrichtung und Größe ausgewertet werden kann. c Atemwasserstrom einer Flunder (Fischsilhouette in gelb). Die durch die Atmung der Flunder entstehenden Wasserbewegungen liegen im Wahrnehmungsbereich der Seehunde
Die Vibrisse ist in einem Follikel in der Haut inseriert. Der Follikel der Robbenvibrisse weist einige Spezifika auf, die als Anpassungen an den aquatischen Lebensraum interpretiert werden (Hyvärinen, et al., 2009; Hyvärinen, 1989; Hyvärinen und Katajisto, 1984): (a) Der Follikel besitzt im Vergleich zu landlebenden Säugetieren (Ebara et al., 2002) einen zusätzlichen cavernösen Blutsinus, der ungefähr 60 % der Länge des Gesamtfollikels ausmacht und den oberen Teil des Follikels ummantelt. Die Region des Follikels, in dem die Mechanorezeptoren zu finden sind, ist dadurch tief in die Haut verlagert. Der Blutsinus heizt auch das Sinnessystem (Mauck et al., 2000), womit das Vibrissensystem und die durch die Vibrissen vermittelten Fähigkeiten weniger anfällig für die Temperatur des umgebenden Mediums sind (Dehnhardt et al., 1998b); (b) Die Follikel der Robben werden von einer sehr hohen Anzahl an Nervenfasern innerviert (siehe z. B. Hyvärinen, 1995; Marshall et al., 2006; McGovern et al., 2015), die die Innervierung der Follikel der landlebenden Säugetiere (Rice et al., 1986) um ein Zehnfaches oder mehr übersteigt; so z. B. werden die Vibrissen der Ringelrobbe von 160.000 Nervenfasern innerviert (Hyvärinen, 1995), die Bartrobben (Erignathus barbatus) dahingegen besitzen 320.000 Nervenfasern zur Innervierung der Vibrissen (Marshall et al., 2006).
Mithilfe ihrer Vibrissen können Seehunde Objekte aktiv ertasten (Haptik; Dehnhardt, et al., 2014; Dehnhardt und Kaminski, 1995; Dehnhardt et al., 1998b; Dehnhardt et al., 1997) sowie auch Wasserbewegungen wahrnehmen und interpretieren (Hydrodynamik; siehe z. B. Dehnhardt und Mauck, 2008; Dehnhardt et al., 1998a; Dehnhardt et al., 2001; Abb. 2b,c). Die Funktion der Vibrissen im Sinne eines haptischen und hydrodynamischen Sensors wird im Kapitel 3. Futtersuche ausführlich erläutert.
1.2.2 Audition
Das wohl auffälligste Merkmal aller Hundsrobben bezüglich des Ohres, allgemein und im Vergleich zu Ohrenrobben, ist das fehlende Außenohr (Abb. 3). Dies wurde vermutlich im Laufe der Evolution zur Erhöhung der Stromlinienförmigkeit des Körpers reduziert. Die sowohl in Luft wie auch unter Wasser nachgewiesene Schalllokalisationsfähigkeit in der Medianebene (Byl, 2017; Byl et al., 2016), also eine Unterscheidung, ob der Schall von oben oder unten bzw. exakt von vorne oder exakt von hinten auf das Ohr getroffen ist, kann dem entsprechend nicht wie z. B. beim Menschen über die Ohrmuschel erklärt werden (Blauert, 1997; Muller und Bovet 1999; Wightman und Kistler, 1997).
Das Ohr der Seehunde weist einige anatomische Veränderungen im Vergleich zu landlebenden Säugern auf (Nummela, 2008; Hemilä et al., 2006; Repenning, 1972; Wartzok und Ketten 1999). (a) Die Mittelohrknochen sind bei den Seehunden größer, zehnmal schwerer und auch dichter als bei Landsäugern mit vergleichbarer Schädelgröße (Nummela, 1995). (b) Im Mittelohr und äußeren Gehörgang ist ein Schwellgewebe vorhanden, welches sich wahrscheinlich beim Tauchen ausdehnt, um die u. U. großen Druckunterschiede während des Tauchens in die Tiefe auszugleichen (Mohl, 1967). (c) Trommelfell und ovales Fenster sind mit 82,9 mm2 und 4,94 mm2 vergrößert (Hemilä, et al., 1995). (d) Der äußere Gehörgang liegt der Schädeloberfläche an und verläuft somit von hinten nach vorne (Boenninghaus, 1903).
Ganz allgemein spiegeln die Erkenntnisse zum Hören der Seehunde die amphibische Lebensweise und zeigen, dass die Seehunde sowohl in Luft als auch unter Wasser gut hören können (Reichmuth et al., 2013). In Luft ist der äußere Gehörgang geöffnet, und der Schall wird über das Mittelohr zum Innenohr weitergeleitet, der allgemeine Weg der Schallweiterleitung der Säuger. Dies ermöglicht den Seehunden ein Hören im Bereich von ∼100 Hz bis ∼30 kHz, wobei der Bereich des besten Hörens bei 2–12 kHz liegt (Reichmuth et al., 2013; Kastak und Schusterman, 1998; Mohl, 1968; Terhune, 1974), und eine gute Schalllokalisationsfähigkeit (Holt et al., 2005; Holt et al., 2004) mit den niedrigsten Schwellenwerten von 4,1 Grad bei 1 kHz und von 3,8 Grad bei 16 kHz. Zur Bestimmung dieser Schwellenwerte musste das Versuchstier bestimmen, ob der Schall von links oder rechts kam; der Schwellenwert wurde dann berechnet als der Winkel (angegeben als Azimuth) der mit einer Leistung von 75 % Richtigwahlen korrespondiert, wobei hier die Richtigwahlen für einen Winkel nach rechts und links zusammengefasst wurden.
Unter Wasser ist das Ohr muskulär geschlossen. In dieser Situation ist der Mechanismus der Schallübertragung aufs Innenohr ungeklärt. Meist wird eine Weiterleitung über die Knochen postuliert; hierzu steht jedoch z. B. die gute Schalllokalisationsfähigkeit der Seehunde unter Wasser im Widerspruch (Bodson et al., 2007; Bodson et al., 2006; Byl et al., 2016). Seehunde hören unter Wasser generell besser als in Luft (Watkins und Wartzok, 1985), der Hörbereich geht bis ca. 100 kHz mit einem Bereich besten Hörens zwischen 4–16 kHz (Reichmuth et al., 2013; Kastak und Schusterman, 1998; Kastelein et al., 2009; Mohl, 1968; Southall et al., 2005; Terhune, 1988; Wolski et al., 2003). Die niedrigsten Hörschwellen von ~ 2,5 – 5,5 Grad aus Schalllokalisationsstudien finden sich zwischen 0,2 – 1 kHz (Bodson et al., 2007; Bodson et al., 2006; Byl et al., 2016).

Die Ohren der Robben. a Das Ohr eines Seehunds, einer Hundsrobbe, ist durch das Fehlen einer Ohrmuschel charakterisiert. b Das Ohr eines Seebärs, einer Ohrenrobbe, besitzt dahingegen eine zipfelförmige Ohrmuschel
1.3 Chemorezeption
Dem Geruchs- und Geschmackssinn der Robben ist bisher recht wenig Aufmerksamkeit geschenkt worden. Allgemein sind die Bedeutung von Olfaktorik und olfaktorische Diskriminationsleistungen bei Ohrenrobben besser als bei den Hundsrobben untersucht (siehe z. B. Kim et al., 2013; Kishida et al., 2007; Laska et al., 2010; Laska et al., 2008; Pitcher et al., 2011). Bezüglich der Hundsrobben und speziell bezüglich der Seehunde gilt, dass sie in Luft, wo die Nase anders als unter Wasser geöffnet ist, riechen können. Die relativ zu terrestrischen Säugern geringere Größe des olfaktorischen Bulbus (Reep et al., 2006; Spitzka, 1890) suggeriert eine geringe Ausprägung des Geruchssinns. Dahingegen unterscheidet sich aber die Siebbeinplatte kaum in der Größe von der terrestrischer Säuger (Pihlström et al., 2005); hieraus könnte eine vergleichbare olfaktorische Sensitivität resultieren. Seehunde sind sehr empfindlich für Dimethylsulfid (DMS; Kowalewsky et al., 2006; Tab. 1), ein Abbauprodukt, welches entsteht, wenn Phytoplankton Zooplankton grast; die Wahrnehmung von DMS wird im Hinblick auf das Auffinden von futterreichen Gegenden diskutiert (siehe Kapitel 2. Orientierung und Navigation). Bei Hundsrobben scheint der Geruch auch eine gewisse Rolle bei der Mutter-Kind-Interaktion (Burton et al., 1975; Kovacs, 1995) oder bei der Paarung (Hardy et al., 1991) zu spielen.
Bezüglich des Geschmacksinns der Robben beschrieb Sonntag (1923) die Anatomie der Zunge u. a. der Seehunde. Er schlussfolgerte, dass Seehunde innerhalb der Flossenfüsser die beste Ausstattung an gustatorischen Organen besitzen. Mit deren Hilfe können die Seehunde Unterschiede im Salzgehalt bestimmen; sie nehmen bei einem Salzgehalt von 30 Promille, der im Lebensraum der Tiere natürlicherweise vorkommt, bereits eine Abweichung von lediglich ≥4 % wahr (Sticken und Dehnhardt, 2000; Tab. 1). Dieser Fähigkeit wird im Hinblick auf Orientierungsleistungen und im Hinblick auf ein Auffinden von Frontensystemen Aufmerksamkeit geschenkt (siehe Kapitel 2. Orientierung und Navigation).
2 Orientierung und Navigation
Orientierung bezeichnet die Fähigkeit eines Individuums, eine Richtung zu erkennen und beizubehalten, während Navigation impliziert, dass das Tier eine Richtung relativ zu einem Ziel bestimmt und diese Richtung dann während der Bewegung auf das Ziel hin beibehält bzw. kontinuierlich anpasst. Für letzteres benötigt es auch Informationen über seine aktuelle Position in Relation zum Ziel.
Das visuelle System könnte auf vielfältige Art und Weise zu Orientierung und Navigation beitragen. Eine Orientierung am Sternenhimmel scheint denkbar, zumal erste Untersuchungen zeigten, dass Seehunde künstliche wie auch natürliche Sterne bis zu einer scheinbaren Helligkeit von 4,4 mag wahrnehmen (Mauck et al., 2005) und einen Stern in Anwesenheit des gesamten Sternenhimmels ähnlich einem Leitstern gezielt anschwimmen können (Mauck et al., 2008; Abb. 4). Während ihnen polarisiertes Licht nicht zugänglich zu sein scheint (Hanke et al., 2013), bleibt es zu untersuchen, ob entweder die Sonne oder der Mond selbst als Orientierungshilfe genutzt werden. Auch eine Orientierung mithilfe von Landmarken, die z. B. in Küstennähe zur Verfügung stehen, gilt es noch experimentell anzugehen.
Ein spannendes Arbeitsgebiet hinsichtlich visueller Navigation bietet die 2014 beschriebene Wahrnehmung von optischem Fluss (Gläser et al., 2014; Abb. 4). Über optischen Fluss könnten Seehunde bestimmen, wie weit sie geschwommen sind. Die Distanzabschätzung ist ein wichtiges Element u. a. der Wegintegration (Etienne und Jeffrey, 2004; Mittelstaedt und Mittelstaedt, 1982). Dieser Navigationsmechanismus würde es den Tieren ermöglichen, auf direktem Weg zum Ausgangspunkt der Reise oder zu einem beliebigen Punkt entlang der Reiseroute zurückzukehren. Hierzu müssten sie über alle bis zum Umkehrpunkt geschwommenen Richtungen und eingeschlagenen Winkel des Reisewegs integrieren. Wegintegration ist hinsichtlich der Meeressäuger sehr vielversprechend, da sie auf Basis von externen Informationen, aber auch allein auf Basis idiothetischer Informationen, also Informationen aus der Eigenbewegung, ablaufen kann, was in dem recht strukturlosen Lebensraum der Tiere von Bedeutung sein könnte.
In diesem Kontext ist anzumerken, dass bisher, was Orientierung und Navigation der Meeressäuger anbelangt, zumeist die klassischen Sinnessysteme berücksichtigt wurden. Jenseits dieser könnten für die Meeressäuger jedoch auch die Parameter Raum und Zeit eine entscheidende Rolle bei den Bewegungen in ihrem Lebensraum spielen. Eine erste Untersuchung offenbarte, dass Seehunde über einen gut ausgeprägten Zeitsinn verfügen und Zeitintervalle von wenigen Millisekunden bis zu einer halben Minute mit hoher Genauigkeit diskriminieren können (Heinrich et al., 2016; Heinrich et al., unpublizierte Daten). Angenommen, dass der Zeitsinn der Seehunde auch bei längeren Zeitintervallen zuverlässige Informationen liefert, wäre es für die Seehunde möglich, abzuschätzen, wie lange oder, unter Berücksichtigung der Schwimmgeschwindigkeit, wie weit sie bereits in eine Richtung geschwommen ist.
Über eine hydrodynamische bzw. auditorische Orientierung wird bisher nur spekuliert. Denkbar ist, dass Seehunde Strömungen unter Wasser nutzen, um ihre Bewegungsrichtung zu bestimmen oder zu modifizieren. Dies setzt voraus, dass diese Strömungen entweder einen zeitweisen oder permanent konstanten Verlauf aufweisen oder systematisch z. B. mit äußeren Parametern variieren. Im Wechselspiel der Gezeiten könnte das mit der Flut einlaufende bzw. mit der Ebbe auslaufende Wasser den Seehunden wichtige hydrodynamische Hinweise zur Orientierung liefern. Die unter Wasser vorherrschende akustische Landschaft könnte Äquivalentes leisten, wie bereits von Norris (1967) für Wale vorgeschlagen wurde.
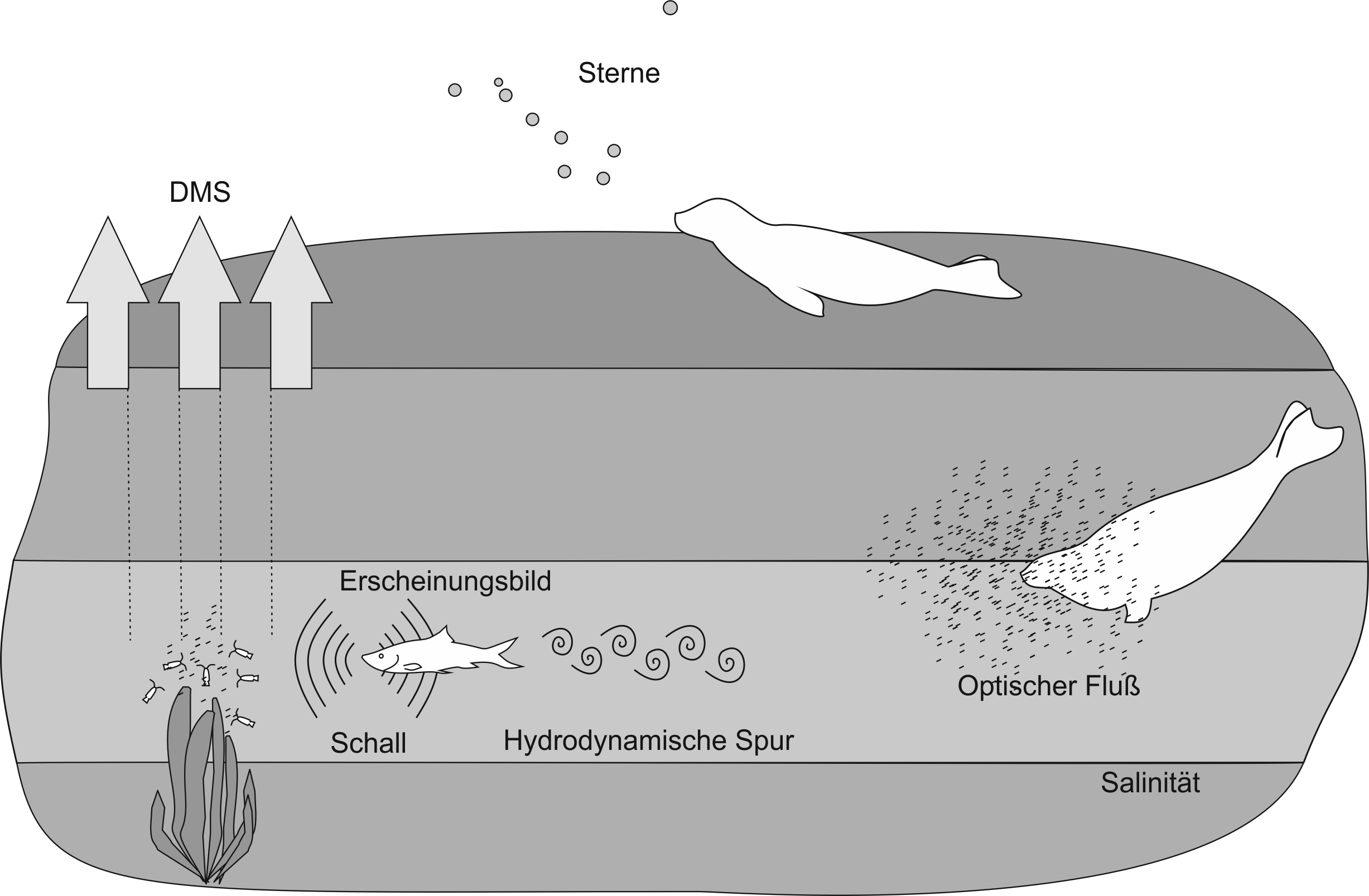
Zusammenfassung der Mechanismen der Orientierung, Navigation und Futtersuche, deren sensorische Grundlage bei Seehunden bereits untersucht wurden. Hierzu zählen (a) die olfaktorische Wahrnehmung von Dimethylsulfat (DMS), welches als Abbauprodukt an Orten entsteht, an denen Zooplankton Phytoplankton grast, (b) die visuelle Wahrnehmung von Sternen, optischem Fluss hervorgerufen durch das Schwimmen durch partikelreiches Wasser oder direkt von Beutetieren, (c) die auditorische Wahrnehmung von Beutetieren, (d) die durch die Vibrissen vermittelte Wahrnehmung hydrodynamischer Spuren und Atemwasserströme hervorgerufen durch die Bewegung und Atmung von Beutetieren sowie (e) die gustatorische Wahrnehmung von Salinität (in dieser Abbildung werden Salinitätsunterschiede in horizontalen Schichten angenommen)
Bezüglich einer chemosensorischen Orientierung wurden bereits sehr plausible Szenarien entwickelt und deren sensorische Grundlage erforscht. Eines der Szenarien basiert auf der Erkenntnis, dass Futtergründe, also Orte reichen Fischvorkommens, durch eine hohe Konzentration von DMS in der Atmosphäre gekennzeichnet sind (Andreae et al., 1994; Bürgermeister et al., 1990). An Orten hoher DMS-Konzentration finden sich entsprechend der Nahrungspyramide auch zahlreiche Fische ein. Da Fische von Seehunden gefressen werden, sind diese Orte wiederum auch für Seehunde interessant. Um fischreiche Gebiete anhand der erhöhten DMS-Konzentration lokalisieren zu können (Abb. 4), gilt es, DMS wahrzunehmen; eine Fähigkeit, die bereits bei Seevögeln eingehend untersucht wurde (für eine Übersicht siehe Nevitt, 2008; Tab. 1). Kowalewsky und Koautoren (2006) zeigten, dass Seehunde diesen Geruchsstoff tatsächlich wahrnehmen können und darüber hinaus, dass Seehunde sehr empfindlich für geringe DMS-Konzentrationen sind. In einem zweiten Schritt müsste nun noch gezeigt werden, dass Seehunde auch DMS lokalisieren können und von DMS-Wolken angezogen werden.
Auch die Salinität charakterisiert teils Orte reichen Nahrungsvorkommens. Speziell an Salinitätsfronten (Abb. 4) charakterisiert durch starke Salinitätsgradienten konnten Areale großer biologischer Produktivität aufgefunden werden (Bowman und Esaias, 1978; Floodgate et al., 1981). Mithilfe ihrer hohen Empfindlichkeit für Salinitätsunterschiede (Sticken und Dehnhardt, 2000; Tab. 1) verfügen Seehunde über die sensorische Grundlage, um diese Gebiete zur Nahrungsaufnahme aufsuchen zu können. Schichten unterschiedlicher Salinität werden von Fischen auch zur Langstreckenorientierung genutzt (Atema, 1988; Westerberg, 1984). Dementsprechend wäre es interessant, die Bewegungsmuster wilder Seehunde mit dem Parameter Salinität zu korrelieren, um einen eventuellen Einfluss der Salinität auf das Verhalten der Seehunde zu offenbaren.
Viele Organismen, wie z. B. Schildkröten, Nacktmulle, Haie und Vögel, orientieren sich am oder navigieren mit dem Erdmagnetfeld (Kalmijn, 1982; Kimchi und Terkel, 2001; Kirschvink et al., 2001; Lohmann und Lohmann, 1996; Mouritsen und Ritz, 2005; Wiltschko und Wiltschko, 2006). Bereits in den 1990er Jahren untersuchte Renouf (1991), ob Seehunde in der Lage sind, Magnetfelder wahrzunehmen, jedoch ohne Erfolg. Das Thema Magnetfeldorientierung wurde dann von Hanke und Koautoren (unpublizierte Daten) aufgegriffen. Obwohl eine Vielzahl an experimentellen Ansätzen mit großen und kleinen Spulen sowie mit Stabmagneten zum Einsatz kam, konnte jedoch auch unsere Forschergruppe keinen Lernerfolg bezüglich der Wahrnehmung von Magnetfeldern oder bezüglich magnetischer Anomalien durch Seehunde verzeichnen. Dementsprechend muss gegenwärtig davon ausgegangen werden, dass Seehunde nicht in der Lage sind, sich am Erdmagnetfeld zu orientieren oder dieses nicht zu Navigationszwecken einsetzen können.
3 Futtersuche
In vielen Fällen werden Seehunde aufs offene Meer hinausschwimmen, um nach Beute zu suchen. Sind die Seehunde in einem fischreichen Gebiet angekommen, so gilt es die Beute aufzuspüren. Bei guten Sichtverhältnissen können die Seehunde ihre Beute direkt sehen und jagen (Kilian et al., 2015). Jagd auf Sicht ist auch bei weniger guten Sichtverhältnissen in direkter Nähe, also kurz vorm Ergreifen der Beute, möglich. Hinweise, dass das visuelle System tatsächlich auf die Nähe eingestellt ist, wurden in einigen Studien zum visuellen System der Seehunde erhalten (Hanke et al., 2009a; Hanke et al., 2011; Scholtyssek et al., 2008; Weiffen et al., 2006).
Wenn die Sicht durch Trübung oder geringe Lichtverhältnisse eingeschränkt ist oder sich die Seehunde noch nicht in der Nähe ihrer Beute befinden, müssen andere Sinnessysteme das Aufspüren der Beute übernehmen. Viele Fische erzeugen Geräusche (Kasumyan, 2008; Wilson et al., 2004), sodass eine Lokalisation der Beute über das auditive System möglich scheint (Abb. 4).
Wenn ein Beutefisch im Freiwasser schwimmt, hinterlässt er eine hydrodynamische Spur, eine Wirbelstraße (Abb. 2b, 4). Seehunde können bei Auftreffen auf eine solche hydrodynamische Spur nicht nur erkennen, dass eine Wasserverwirbelung vorliegt (Dehnhardt et al., 1998a), sondern ihr auch aktiv folgen (Dehnhardt und Mauck, 2008; Dehnhardt et al., 2001). Diese hydrodynamische Spurverfolgung wurde im Experiment mit ferngesteuerten U-Booten (Dehnhardt et al., 2001), einem Artgenossen (Schulte-Pelkum et al., 2007) und künstlichen Fischen (Kilian, 2010) gezeigt. Die Versuchstiere konnten einer frischen Spur folgen. Sie erreichten jedoch auch ihr Ziel, wenn die hydrodynamische Spur über eine halbe Minute alt war, eine Simulation der Situation, in der ein Fisch bereits weitergeschwommen wäre, oder wenn die hydrodynamische Spur Gleitphasen aufwies (Wieskotten et al., 2010b). Eine hydrodynamische Spur scheint eine Vielzahl an weiterführenden Informationen wie die Richtung, in die der Spurgenerator geschwommen ist, sowie seine Größe und Form zu tragen (Wieskotten et al., 2010a; Wieskotten et al., 2011). Selbst wenn nur einzelne Wirbelringe präsentiert werden, die in einer hydrodynamischen Spur enthalten sind oder auch bei Fluchtreaktionen von Fischen entstehen (Niesterok und Hanke, 2013; Tytell und Lauder, 2008; Abb. 2b), können Seehunde Bewegungsrichtung und Größe des Wirbelrings ablesen (Krüger, 2017; Krüger et al., 2018). Dies ermöglicht den Seehunden, bei Auftreffen auf eine solche hydrodynamische Spur Entscheidungen, z. B. hinsichtlich seiner weiteren Fortbewegungsrichtung, zu treffen. Es bleibt zu zeigen, dass Seehunde auch der hydrodynamischen Spur eines echten Fisches und dies unter Echtbedingungen, bei z. B. Vorliegen natürlicher Strömungen, folgen können.
Fische im Freiwasser machen oft nur einen geringen Anteil der Nahrung der Seehunde aus. Mageninhaltsanalysen zeigten, dass sich Seehunde mancherorts fast ausschließlich von Plattfischen ernähren (siehe z. B. Härkönen, 1987; Pierce et al., 1991; Tollit und Thompson, 1996). Plattfische liegen meist vergraben im Sand, an den sie zudem noch farblich angepasst sind. Eine visuelle Detektion erscheint darum fast undenkbar, auch wenn es möglich sein könnte, diese Fische über die kohärente Bewegung des Körpers zu detektieren (Lui et al., 2012; Weiffen et al., 2014). Auch ein elektrorezeptives Aufspüren der Fische wurde bislang ausgeschlossen, da elektrorezeptive Fähigkeiten bei Seehunden noch nicht nachgewiesen werden konnten. Bislang wurde angenommen, dass Seehunde die Plattfische durch direkten Kontakt mit den Vibrissen, also haptisch aufspüren. Haptisch könnte in der Tat die Anwesenheit eines Fisches sowie dessen Form, Größe oder Oberflächenbeschaffenheit bestimmt werden (Dehnhardt et al., 2014; Dehnhardt und Kaminski, 1995; Dehnhardt et al., 1998b; Dehnhardt et al., 1997). Ein direkter Kontakt der Vibrissen mit der Beute trat aber auf Videoaufzeichnungen, die Seehunde bei der Jagd zeigen, nicht zu Tage. Vielmehr schwimmen die Seehunde oftmals in deutlichem Abstand über den Meeresgrund. Dies berücksichtigend hypothetisierten Niesterok und Koautoren (Niesterok et al., 2017a; Niesterok et al., 2017b), dass Seehunde Plattfische über den Wasserstrom, der bei der Atmung der Fische entsteht (Bublitz, 2010; Abb. 2c), lokalisieren können. Diese Hypothese wurde untermauert durch: (a) Vermessungen der Atemwasserströme, die, was die Wassergeschwindigkeiten anbelangt, im Wahrnehmungsbereich der Seehunde liegen. (b) Verhaltensexperimente, in denen Seehunde einen künstlichen Atemwasserstrom mit vergleichbaren Eigenschaften wie dem Atemwasserstrom von Plattfischen in einem großen Areal aufspüren konnten. Somit wäre auch eine hydrodynamische Detektion von benthisch lebenden Fischen denkbar.
4 Diskussion und Ausblick
Die Forschung der letzten Jahrzehnte hat das Verständnis der Sinnessysteme der Seehunde und allgemein der Robben stark verbessert. Dennoch gibt es, wie auch bereits in den vorherigen Kapiteln aufgezeigt, noch viel Forschungsbedarf, um die bestehenden Lücken hinsichtlich der Sinnessysteme selbst und hinsichtlich Mechanismen der Orientierung, Navigation und Futtersuche zu schließen.
Dieser Übersichtsartikel hat sich in großen Teilen mit Erkenntnissen, die mit Seehunden erzielt wurden, befasst. Bevor der Seehund durch systematische Erforschung bezüglich seiner Sinnessysteme zum Modellorganismus wurde, waren nur Einzelinformationen über die Sinnessysteme unterschiedlicher Robbenarten vorhanden. Oftmals wurde dann von einer Robbenart auf die andere geschlossen. Diese Rückschlüsse sind jedoch nur bedingt sinnvoll, da alle Robbenarten sehr spezifische Anpassungen an ihren Lebensraum und ihre Lebensweise zeigen. Nun, da wir jedoch ein etwas detaillierteres Bild der Sinnessysteme einer Robbenart haben, wären Vergleichsstudien mit anderen Robben hinsichtlich ausgewählter Aspekte durchaus interessant. Beispielsweise birgt die unterschiedliche Struktur der Vibrissen innerhalb der Robben, undulierte oder glatte Vibrisse, viel Potenzial für weitere Vergleichsstudien (Gläser et al., 2011; Miersch et al., 2011). Oder auch die Tatsache, dass innerhalb der Robben vollständig und nur teilweise abgeflachte Corneae aufzufinden sind (Hanke et al., 2006a; Dawson et al., 1987), wirft die Frage auf, ob es weitere Ausprägungen gibt oder ob sich hier Ohren- von Hundsrobben systematisch unterscheiden.
Ein vergleichender Blick auf Tierarten, die z. B. den aquatischen Lebensraum teilen oder in einem Lebensraum mit ähnlichen Eigenschaften leben, kann ebenfalls zu spannenden Fragestellungen führen. Bezüglich des letztgenannten Aspekts ist z. B. ein Blick auf die Wüstenameise interessant, die in einem weitestgehend strukturlosen Lebensraum vergleichbar dem offenen Meer vorkommt. Die Wüstenameise betreibt Wegintegration, die unserer Meinung nach auch ein vielversprechender Navigationsmechanismus im Hinblick auf die Meeressäuger ist. In unebenem Terrain dokumentiert die Wüstenameise bei der Berechnung des Heimvektors, der sie nach erfolgreicher Futtersuche auf direktem Wege zu ihrem Nest zurückbringt, jedoch nicht die tatsächlich zurückgelegte Strecke. Die Wüstenameise berücksichtigt nur die Distanz, die mit der Projektion der Wegsegmente auf die Horizontale korrespondiert (Wohlgemuth et al., 2001). Vergleichbares könnte bei Meeressäugern auftreten, bei denen aufgrund von Tauchgängen die tatsächlich von der horizontal zurückgelegten Strecke ebenfalls abweichen kann.
Bisherige Studien zu den Sinnessystemen der Robben wurden Laborexperimenten vergleichbar durchgeführt. Die dadurch sehr kontrollierten Versuchsbedingungen sind essenziell für die Erlangung eines grundlegenden Verständnisses, z. B. der Funktionsweise der Sinnessysteme. Sie bilden aber nur bedingt die Situationen ab, die eine Robbe in ihrem natürlichen Lebensraum vorfindet. In Zukunft wäre es deshalb erstrebenswert, Experimente unter natürlichen Bedingungen, z. B. direkt im Lebensraum der Tiere, durchzuführen. Diese Freilandexperimente hätten zudem den Vorteil, dass Raum kein einschränkender Parameter darstellte; dies ist eine entscheidende Voraussetzung für Orientierungs- oder Navigationsexperimente, die auch u. a. aus diesem Grund noch nicht durchgeführt worden sind.
Während das Verständnis der Leistungen einzelner Sinnessysteme der Robben gewachsen ist, wurde bisher weitestgehend vernachlässigt, dass Sinnessysteme interagieren, sich ergänzen oder ersetzen. Daher sehen wir viel Potenzial in Studien, die Multimodaliät und sensorische Integration zum Thema haben. Sensorische Integration könnte experimentell, z. B. mithilfe der funktionellen Magnetresonanztomografie, angegangen werden. Diese Methode kann auch die Lücke zwischen den Sinnessystemen und dem Zentralnervensystem schließen, ein bislang stark vernachlässigtes Forschungsgebiet, was unseren Modelorganismus anbelangt. Ganz allgemein verspricht der Blick in ein komplexes Zentralnervensystem eines Säugers, welcher sich aber über ~ 30 Mio Jahre an den aquatischen Lebensraum angepasst hat, viele neuartige Erkenntnisse.
Zur Erlangung eines grundlegenden Verständnis der Orientierungs- und Navigationsleistungen der Robben ist es auch essenziell, die Lücke zwischen den sensorischen Fähigkeiten, die mit Labortieren ermittelt wurden, und dem tatsächlichen Verhalten von Wildtieren zu schließen. Beide Disziplinen können sich befruchten: Zum einen können sensorische Fähigkeiten helfen, das Verhalten von Wildtieren zu beschreiben. Zum anderen kann das Verhalten der Wildtiere zu neuen Hypothesen führen, die daraufhin mit Labortieren getestet werden können.
Danksagung
Die Autoren danken der VolkswagenStiftung und der Deutschen Forschungsgemeinschaft, welche die diesem Übersichtartikel hauptsächlich zugrunde liegende Forschung finanziert haben.
About the authors

Frederike D. Hanke studierte Biologie an der Universität Freiburg, wobei sie sich auf Tierphysiologie/Neurobiologie, Ökologie/Evolution, Genetik und Verhaltensbiologie/Psychologie spezialisierte. Ihren Doktortitel erhielt sie dann von der Internationalen Graduiertenschule für Biowissenschaften der Universität Bochum. Während ihrer Promotion sowie auch in der darauffolgenden Postdoktorandenzeit hatte sie die einmalige Gelegenheit, zahlreiche Projekte im In- und Ausland durchzuführen. Kürzlich vollendete sie erfolgreich ihre Habilitation an der Universität Rostock. Während ihrer wissenschaftlichen Karriere erfreute Frederike Hanke sich daran, mit den unterschiedlichsten und spannenden Tieren wie Honigbienen, Rabenvögeln, Fischen und Bären zu arbeiten. Ihre Hauptversuchstiere sind jedoch Robben und Kopffüßer. In ihrer Forschung legt sie den Schwerpunkt auf deren Sinnessysteme, wobei hierbei hauptsächlich das visuelle System Beachtung findet. Frederike Hanke interessiert sich für vielerlei Aspekte des Sehens so z. B. die Anpassungen des visuellen Systems an die artspezifische Lebensweise und –art bis zu Bewegungs- und Polarisationssehen, visuelle Orientierung und Kognition sowie visuelle Neuroanatomie.

Guido Dehnhardt studierte Biologie und Geografie an der Universität Münster. Bereits während seines Studiums fokusierte er sich stark auf Meeressäuger und besuchte deshalb auch das Kewalo Basin Marine Mammal Laboratory, University of Hawaii, und das Naval Ocean Systems Center. Danach verfolgte er weiter seinen Weg im Rahmen einer Doktorarbeit auf dem Gebiet der Meeressäugerforschung. Seinen Doktortitel erhielt er ebenfalls von der Universität Münster. Nach einer Postdoktorandenzeit an der Universität Bielefeld wechselte Guido Dehnhardt an die Universität Bonn. Während der Bonner Zeit hatte er auch die Gelegenheit, Forschungsprojekte an der University of Joensuu, Finnland, und der University of Newfoundland, Kanada, durchzuführen, bevor er sich dann an der Universität Bonn habilitierte. Jahre intensiver Forschungstätigkeit an der Universität Bochum folgten, woraufhin er dann eine Lichtenberg-Professur der VolkswagenStiftung erhielt, die an der Universität Rostock etabliert wurde. In Rostock gründete er das weithin bekannte Marine Science Center, die größte Robbenforschungsstation weltweit. Guido Dehnhardt interessiert sich in seiner Forschung für sensorische und kognitive Fähigkeiten als Grundlage der Orientierung mariner Säuger. Im Speziellen wurde er bekannt durch seine Erforschung der Funktion der Vibrissen als haptisches und hydrodynamisches System.
Literatur
Andreae, T.W., Andreae, M.O. and Schebeske, G. (1994). Biogenic sulfur emissions and areosols over the tropical south Atlantic 1. Dimethylsulfide in seawater and the atmosphere boundary layer. J. Geophys. Res. 99. 22819–22829.10.1029/94JD01837Search in Google Scholar
Atema, J. (1988). Distribution of chemical stimuli. In: Atema, J., Fay, R.R., Popper, A.N., Tavolga, W.N. (eds) Sensory biology of aquatic animals. Springer, Berlin Heidelberg New York, pp 29–56.10.1007/978-1-4612-3714-3_2Search in Google Scholar
Augusteyn, R.C., Nankivil, D., Mohamed, A., Maceo, B., Pierre, F. and Parrel, J.-M. (2012). Human ocular biometry. Exp. Eye Res. 102c. 70–75.10.1016/j.exer.2012.06.009Search in Google Scholar PubMed PubMed Central
Blauert. J. (1997). Spatial hearing: the psychophysics of human sound localization. MIT Press, Cambridge10.7551/mitpress/6391.001.0001Search in Google Scholar
Belitz. H.-D. and Grosch, W. (1982). Lehrbuch der Lebensmittelchemie. Springer Verlag, Berlin Heidelberg New York, pp 260–307.10.1007/978-3-662-08306-2_6Search in Google Scholar
Bodson, A., Miersch, L. and Dehnhardt, G. (2007). Underwater localization of pure tones by harbor seals (Phoca vitulina). J. Acoustic Soc. Am. 122. 2263–2269.10.1121/1.2775424Search in Google Scholar PubMed
Bodson, A., Miersch, L., Mauck, B. und Dehnhardt, G. (2006). Underwater auditory localization by a swimming harbor seal (Phoca vitulina). J. Acoustic Soc. Am. 120. 1550–1557.10.1121/1.2221532Search in Google Scholar PubMed
Boenninghaus, G. (1903). Das Ohr des Zahnwales, zugleich ein Beitrag zur Theorie der Schalleitung. Eine biologische Studie. Zoolog. Abt. Anat. Ont. 19. 1–175.Search in Google Scholar
Bonadonna, F., Caro, S., Jouventin, P. and Nevitt, G.A. (2006). Evidence that blue petrel, Halobaena caerulea, fledglings can detect and orient to dimethyl sulfide. J. Exp. Biol. 209. 2165–2169.10.1242/jeb.02252Search in Google Scholar PubMed
Bowman, M.J. and Esaias, W.E. (1978). Oceanic fronts in coastal processes. Springer, Berlin Heidelberg New York10.1007/978-3-642-66987-3Search in Google Scholar
Bublitz, A. (2010). Wasserbewegungen von stationären Fischen und ihre mögliche Bedeutung für fischfressende Tiere. Diploma thesis, University of RostockSearch in Google Scholar
Bürgermeister, S., Zimmermann, R.L., Georgii, H.W., Bingemer, H.G., Kirst, G.O., Janssen, M. and Ernst, W. (1990). On the biogenic origin of dimethylsulfide: relation between chlorophyll, ATP, organismic DMSP, phytoplankton species, and DMS distribution in Atlantic surface water and atmosphere. J. Geophys. Res. 95. 20607–20615.10.1029/JD095iD12p20607Search in Google Scholar
Burton, R.W., Anderson, S. S. and Summers, C.F. (1975). Perinatal activities in the grey seal (Halichoerus grypus). J. Zool. 177. 197–201.10.1111/j.1469-7998.1975.tb05978.xSearch in Google Scholar
Byl, J.A. (2017). Aerial and underwater sound localization abilities of harbor seals (Phoca vitulina) in the median plane. PhD thesis, University of RostockSearch in Google Scholar
Byl, J.A., Miersch, L., Wieskotten, S. and Dehnhardt, G. (2016). Underwater sound localization of pure tones in the median plane by harbor seals (Phoca vitulina). J. Acoustic Soc. Am. 140. 4490.10.1121/1.4972531Search in Google Scholar PubMed
Chevaillier, D., Karyptchev, M., McConnell, B.J., Moss, S. and Vincent, C. (2014). Can gray seals maintain heading within areas of high tidal current? Preliminary results from numerical modeling and GPS observations. Mar. Mamm. Sci. 30(1). 374–380.10.1111/mms.12024Search in Google Scholar
Cohen, J.L., Tucker, G.S. and Odell, D.K. (1982). The photoreceptor of the West Indian manatee. J. Morphol. 173. 197–202.10.1002/jmor.1051730207Search in Google Scholar PubMed
Crognale, M.A., Levenson, D.H., Ponganis, P.J., Deegan II, J.F. and Jacobs, G.H. (1998). Cone spectral sensitivity in the harbor seal (Phoca vitulina) and implications for color vision. Can. J. Zool. 76. 2114–2118.Search in Google Scholar
Dawson, W.W., Schroeder, J.P. and Sharpe, S. N. (1987). Corneal surface properties of two marine mammal species. Mar. Mamm. Sci. 3(2). 186–197.10.1111/j.1748-7692.1987.tb00161.xSearch in Google Scholar
Dehnhardt, G., Hanke, W., Wieskotten, S., Krüger, Y. and Miersch, L. (2014). Hydrodynamic perception in seals and sea lions. In: Bleckmann, H. (ed) Flow sensing in air and water. Springer, Berlin Heidelberg, pp 147–167.10.1007/978-3-642-41446-6_6Search in Google Scholar
Dehnhardt, G. and Kaminski, A. (1995). Sensitivity of the mystacial vibrissae of harbour seals (Phoca vitulina) for size differences of actively touched objects. J. Exp. Biol. 198. 2317–2323.10.1242/jeb.198.11.2317Search in Google Scholar PubMed
Dehnhardt, G. and Mauck, B. (2008). Mechanoreception in secondarily aquatic vertebrates. In: Thewissen, J.G.M., Nummela, S. (eds) Sensory evolution on the threshold: adaptations in secondarily aquatic vertebrates. University of California Press, Berkeley, Los Angeles, pp 295–314.10.1525/california/9780520252783.003.0018Search in Google Scholar
Dehnhardt, G., Mauck, B. and Bleckmann, H. (1998a). Seal whiskers detect water movements. Nature 394. 235–236.10.1038/28303Search in Google Scholar
Dehnhardt, G., Mauck, B., Hanke, W. and Bleckmann, H. (2001). Hydrodynamic trail-following in harbor seals (Phoca vitulina). Science 293. 102–104.10.1126/science.1060514Search in Google Scholar PubMed
Dehnhardt, G., Mauck, B. and Hyvärinen, H. (1998b). Ambient temperature does not affect tactile sensitivity of mystacial vibrissae in harbour seals. J. Exp. Biol. 201. 3023–3029.10.1242/jeb.201.22.3023Search in Google Scholar PubMed
Dehnhardt, G., Sinder, M. and Sachser, N. (1997). Tactual discrimination of size by means of mystacial vibrissae in harbor seals: in air versus underwater. Zeitschr. Saugetierkd. 62. 40–43.Search in Google Scholar
Dohmen, M. et al. (2015). Understanding fiber mixture by simulation in 3D Polarized Light Imaging. NeuroImage 111. 464–475.10.1016/j.neuroimage.2015.02.020Search in Google Scholar PubMed
Ebara, S., Kumamoto, K., Matsuura, T., Mazurkiewicz, J.E. and Rice, F. (2002). Similarities and differences in the innervation of mystacial vibrissa follicle-sinus complexes in the rat and cat: a confocal microscopic study. J. Comp. Neurol. 449. 103–119.10.1002/cne.10277Search in Google Scholar PubMed
Etienne, A.S. and Jeffrey, K.J. (2004). Path integration in mammals. Hippocampus 14. 180–192.10.1002/hipo.10173Search in Google Scholar PubMed
Fasick, J.I. and Robinson, P.R. (2000). Spectral-tuning mechanisms of marine mammal rhodpsins and correlations with foraging depth. Vis. Neurosci. 17. 781–788.10.1017/S095252380017511XSearch in Google Scholar PubMed
Floodgate, G.D., Fogg, G.E., Jones, D.A., Lochte, K. and Turley, C.M. (1981). Microbiological and zooplankton activity at a front in Liverpool Bay. Nature 290. 133–136.10.1038/290133a0Search in Google Scholar
Friedl, W.A., Nachtigall, P. E., Moore, P.W.B., Chun, N.K.W., Haun, J.E., Hall, R.W. and Richards, J.L. (1990). Taste reception in the Pacific bottlenose dolphin (Tursiops truncatus gilli) and the California sea lion (Zalophus californianus). In: Thomas, J., Kastelein, R. (eds), Sensory abilities of cetaceans. Plenum Press, New York, pp 447–454.10.1007/978-1-4899-0858-2_31Search in Google Scholar
Gläser, N., Mauck, B., Kandil, F., Lappe, M., Dehnhardt, G. and Hanke, F.D. (2014). Harbour seals (Phoca vitulina) can perceive optic flow underwater. PloS One 9: e103555.10.1371/journal.pone.0103555Search in Google Scholar PubMed PubMed Central
Gläser, N., Wieskotten, S., Otter, C., Dehnhardt, G. and Hanke, W. (2011). Hydrodynamic trail following in a California sea lion (Zalophus californianus). J. Com. Physiol. A 197. 141–151.10.1007/s00359-010-0594-5Search in Google Scholar PubMed
Hanke, F.D. and Dehnhardt, G. (2009). Aerial visual acuity in harbor seals (Phoca vitulina) as a function of luminance. J. Comp. Physiol. A 195. 643–650.10.1007/s00359-009-0439-2Search in Google Scholar PubMed
Hanke, F.D., Dehnhardt, G., Schaeffel, F. and Hanke, W. (2006a). Corneal topography, refractive state, and accommodation in harbor seals (Phoca vitulina). Vis. Res. 46. 837–847.10.1016/j.visres.2005.09.019Search in Google Scholar PubMed
Hanke, F.D., Hanke, W., Hoffmann, K.-P, and Dehnhardt, G. (2008a). Optokinetic nystagmus in harbor seals (Phoca vitulina). Vis. Res. 48. 304–315.10.1016/j.visres.2007.11.012Search in Google Scholar PubMed
Hanke, F.D., Hanke, W., Scholtyssek, C. and Dehnhardt, G. (2009a). Basic mechanisms in pinniped vision. Exp. Brain Res. 199. 299–311.10.1007/s00221-009-1793-6Search in Google Scholar PubMed
Hanke, F.D., Kröger, R.H.H., Siebert, U. and Dehnhardt, G. (2008b). Multifocal lenses in a monochromat: the harbour seal. J. Exp. Biol. 211. 3315–3322.10.1242/jeb.018747Search in Google Scholar PubMed
Hanke, F.D., Miersch, L., Warrant, E.J., Mitschke, F.M. and Dehnhardt, G. (2013). Are harbour seals (Phoca vitulina) able to perceive and use polarised light? J. Comp. Physiol. A 199. 509–519.10.1007/s00359-012-0762-xSearch in Google Scholar PubMed
Hanke, F.D., Peichl, L. and Dehnhardt, G. (2009b). Retinal ganglion cell topography in juvenile harbor seals (Phoca vitulina). Brain Behav. Evol. 74. 102–109.10.1159/000235612Search in Google Scholar PubMed
Hanke, F.D., Scholtyssek, C., Hanke, W. and Dehnhardt, G. (2011). Contrast sensitivity in a harbor seal (Phoca vitulina). J. Comp. Physiol. A 197. 203–210.10.1007/s00359-010-0600-ySearch in Google Scholar PubMed
Hanke, W., Römer, R. and Dehnhardt, G. (2006b). Visual fields and eye movements in a harbor seal (Phoca vitulina). Vis. Res. 46. 2804–2814.10.1016/j.visres.2006.02.004Search in Google Scholar PubMed
Hanke, W. et al. (2010). Harbor seal vibrissa morphology suppresses vortex-induced vibrations. J. Exp. Biol. 213. 2665–2672.10.1242/jeb.043216Search in Google Scholar PubMed
Hardy, M.H., Roff, E., Smith, T.G. and Ryg, M. (1991). Facial skin glands of ringed and gray seals, and their possible function as odoriferous organs. Can. J. Zool. 69. 189–200.Search in Google Scholar
Härkönen, T. (1987). Seasonal and regional variations in the feeding habits of the harbour seal, Phoca vitulina, in the Skagerrak and the Kattegat. J. Zool. 213. 535–543.10.1111/j.1469-7998.1987.tb03724.xSearch in Google Scholar
Heinrich, T., Dehnhardt, G. and Hanke, F.D. (2016). Harbor seals (Phoca vitulina) are able to time precisely. Anim. Cogn. 19. 1133–1142.10.1007/s10071-016-1020-3Search in Google Scholar
Hemilä, S., Nummela, S. and Reuter, T., (1995), What middle ear parameters tell about impedence matching and high frequency hearing. Hear. Res. 85. 31–44.10.1016/0378-5955(95)00031-XSearch in Google Scholar
Hemilä, S., Nummela, S., Berta, A. and Reuter, T. (2006). High-frequency hearing in phocid and otariid pinnipeds: an interpretation based on inertial and cochlear constraints (L). J. Acoustic Soc. Am. 120. 3463–3466.10.1121/1.2372712Search in Google Scholar
Holt, M.M., Schusterman, R.J., Kastak, D. and Southall, B.L. (2005). Localization of aerial pure tones by pinnipeds. J. Acoustic Soc. Am. 118. 3921–3926.10.1121/1.2126931Search in Google Scholar
Holt, M.M., Schusterman, R.J., Southall, B.L. and Kastak, D. (2004). Localization of aerial broadband noise by pinnipeds. J. Acoustic Soc. Am. 115. 2339–2345.10.1121/1.1694995Search in Google Scholar
Hyvärinen, H. (1989). Diving in darkness: whiskers as sense organs of the ringed seal (Phoca hispida). J. Zool. 218. 663–678.10.1111/j.1469-7998.1989.tb05008.xSearch in Google Scholar
Hyvärinen, H. (1995). Structure and function of the vibrissae of the ringed seal (Phoca hispida Schr.). In: Kastelein, R.A., Thomas, J.A., Nachtigall, P. E. (eds) Sensory systems of aquatic mammals. De Spil, Woerden, pp 429–445.Search in Google Scholar
Hyvärinen, H. and Katajisto, H. (1984). Functional structure of the vibrissae of the ringed seal (Phoca hispida Schr.). Acta Zool. 171. 27–30.Search in Google Scholar
Hyvärinen, H., Palviainen, A., Strandberg, U. and Holopainen, I.J. (2009). Aquatic environment and differentiation of vibrissae: comparison of sinus hair systems of ringed seal, otter and pole cat. Brain Behav. Evol. 74. 268–279.10.1159/000264662Search in Google Scholar
Jamieson, G.S. (1971). The functional significance of corneal distortion in marine mammals. Can. J. Zool. 49. 421–423.10.1139/z71-063Search in Google Scholar
Jamieson, G.S. and Fisher, H.D. (1970). Visual discrimination in the harbour seal Phoca vitulina, above and below water. Vis. Res. 10. 1175–1180.10.1016/0042-6989(70)90035-0Search in Google Scholar
Jamieson, G.S. and Fisher, H.D. (1971). The retina of the harbour seal, Phoca vitulina. Can. J. Zool. 49. 19–23.10.1139/z71-005Search in Google Scholar
Jamieson, G.S. and Fisher, H.D. (1972). The pinniped eye: a review. In: Harrison, R.J. (ed) Functional anatomy of marine mammals, vol 1. Academic Press, London, New York, pp 245–261.Search in Google Scholar
Johnson, G.L. (1901). Contributions to the comparative anatomy of the mammalian eye, chiefly based on ophthalmoscopic examination. Phil. Trans. R. Soc. B 194. 1–30.Search in Google Scholar
Kalmijn, A.J. (1982). Electric and magnetic field detection in elasmobranch fishes. Science 218. 916–918.10.1126/science.7134985Search in Google Scholar
Kastak, D. and Schusterman, R.J. (1998). Low-frequency amphibious hearing in pinnipeds: methods, measurements, noise, and ecology. J. Acoustic Soc. Am. 103. 2216–2228.10.1121/1.421367Search in Google Scholar
Kastelein, R.A., Wensveen, P.J., Hoek, L., Verboom, W.C. and Terhune, J.M. (2009). Underwater detection of tonal signals between 0.25 and 100kHz by harbor seals (Phoca vitulina). J. Acoustic. Soc. Am. 125. 1222–1229.10.1121/1.3050283Search in Google Scholar
Kasumyan, A.O. (2008). Sounds and sound production in fishes. J. Ichthyol. 48. 981–1030.10.1134/S0032945208110039Search in Google Scholar
Kilian, M. (2010). Hydrodynamische Spurverfolgung beim Seehund (Phoca vitulina). Diploma thesis, University of RostockSearch in Google Scholar
Kilian, M., Dehnhardt, G. and Hanke, F.D. (2015). How harbor seals (Phoca vitulina) pursue schooling herring. Mamm. Biol. 80. 385–389.10.1016/j.mambio.2015.04.004Search in Google Scholar
Kim, S.K., Amundin, M. and Laska, M. (2013). Olfactory discrimination ability of South African fur seals (Arctocephalus pusillus) for enantiomers. J. Comp. Physiol. A 199. 535–544.10.1007/s00359-012-0759-5Search in Google Scholar
Kimchi, T. and Terkel, J. (2001). Magnetic compass orientation in the blind mole rat Spalax ehrenbergi. J. Exp. Biol. 204. 751–758.10.1242/jeb.204.4.751Search in Google Scholar
Kirschvink, J.L., Walker, M.M. and Diebel, C.E. (2001). Magnetite-based magnetoreception. Curr. Opin. Neurobiol. 11. 462–467.10.1016/S0959-4388(00)00235-XSearch in Google Scholar
Kishida, T., Kubota, S., Shirayama, Y. and Fukami, H. (2007). The olfactory receptor gene repertoires in secondary-adapted marine vertebrates: evidence for reduction of the functional proportions in cetaceans. Biol. Lett. 3. 428–430.10.1098/rsbl.2007.0191Search in Google Scholar PubMed PubMed Central
Kovacs, K.M. (1995). Mother-pup reunions in harp seals, Phoca groenlandica – cues for he relocation of pups. Can. J. Zool. 73. 843–849.Search in Google Scholar
Kowalewsky, S., Dambach, M., Mauck, B. and Dehnhardt, G. (2006). High olfactory sensitivity for dimethyl sulphide in harbor seals. Biol. Lett. 2. 106–109.10.1098/rsbl.2005.0380Search in Google Scholar PubMed PubMed Central
Kröger, J. (2012). Application of tracing methods for the staining of selected cranial nerves in mammals. Master thesis, University of RostockSearch in Google Scholar
Krüger, Y. (2017). Perception of single vortex rings by harbour seals (Phoca vitulina). PhD thesis, University of RostockSearch in Google Scholar
Krüger, Y., Hanke, W., Miersch, L. and Dehnhardt, G. (2018). Detection and direction discrimination of single vortext rings by harbour seals (Phoca vitulina). J. Exp. Biol. doi:10.1242/jeb.17075310.1242/jeb.170753Search in Google Scholar
Landau, D. and Dawson, W.W. (1970). The histology of retinas from the pinnipedia. Vis. Res. 10. 691–702.10.1016/0042-6989(70)90016-7Search in Google Scholar
Laska, M., Lord E., Selin, S. and Amundin, M. (2010). Olfactory discrimination of aliphatic odorants in South African fur seals (Arctocephalus pusillus). J. Comp. Psychol. 124. 187–193.10.1037/a0018189Search in Google Scholar
Laska, M., Svelander, M. and Amundin, M. (2008). Successful acquisition of an olfactory discrimintation paradigm by South African fur seals, Arctocephalus pusillus. Physiol. Behav. 93. 1033–1038.10.1016/j.physbeh.2008.01.019Search in Google Scholar
Lavigne, D.M. and Ronald, K. (1975). Pinniped visual pigments. Comp. Biochem. Physiol. 52. 325–329.10.1016/0305-0491(75)90073-5Search in Google Scholar
Leonardos, G., Kendall, D. and Barnard, N. (1969). Odor threshold determination of 53 odorant chemicals. J. Air Pollut. Contr. Assoc. 19. 91–95.10.1080/00022470.1969.10466465Search in Google Scholar
Levenson, D.H., Ponganis, P.J., Crognale, M.A., Deegan II, J.F., Dizon, A. and Jacobs, G.H. (2006). Visual pigments of marine carnivores: pinnipeds, polar bear, and sea otter. J. Comp. Physiol. A 192. 833–843.10.1007/s00359-006-0121-xSearch in Google Scholar PubMed
Levenson, D.H. and Schusterman, R.J. (1997). Pupillometry in seals and sea lions: ecological implications. Can. J. Zool. 75. 2050–2057.10.1139/z97-838Search in Google Scholar
Levenson, D.H. and Schusterman, R.J. (1999). Dark adaptation and visual sensitivity in shallow and deep-diving pinnipeds. Mar. Mam. Sci. 15. 1303–1313.10.1111/j.1748-7692.1999.tb00892.xSearch in Google Scholar
Lohmann, K.J. and Lohmann, C.M.F. (1996). Detection of magnetic field intensity by sea turtles. Nature 380. 59–61.10.1038/380059a0Search in Google Scholar
Lui, L.L., Dobiecki, A.E., Bourne, J.A. and Rosa, M.G.P. (2012). Breaking camouflage: responses of neurons in the middle temporal area to stimuli defined by coherent motion. Eur. J. Neurosci. 36. 2063–2076.10.1111/j.1460-9568.2012.08121.xSearch in Google Scholar PubMed
Marshall, C.D., Amin, H., Kovacs, K.M. and Lydersen, C. (2006). Microstructure and innervation of the vibrissal follicle-sinus complex in the bearded seal, Erignathus barbatus (Pinnipedia: Phocidae). Anat. Rec. 288A. 13–25.10.1002/ar.a.20273Search in Google Scholar PubMed
Mauck, B., Brown, D., Schlosser, W., Schaeffel, F. and Dehnhardt, G. (2005). How a harbour seal sees the night sky. Mar. Mam. Sci. 21. 646–656.10.1111/j.1748-7692.2005.tb01257.xSearch in Google Scholar
Mauck, B., Eysel, U. and Dehnhardt, G. (2000). Selective heating of vibrissal follicles in seals (Phoca vitulina) and dolphins (Sotalia fluviatilis guianensis). J. Exp. Biol. 203. 2125–2131.10.1242/jeb.203.14.2125Search in Google Scholar PubMed
Mauck, B., Gläser, N., Schlosser, W. and Dehnhardt, G. (2008). Harbour seals (Phoca vitulina) can steer by the stars. Anim. Cogn. 11. 715–718.10.1007/s10071-008-0156-1Search in Google Scholar PubMed
McGovern, K.A., Marshall, C.D. and Davis, R.W. (2015). Are vibrissae viable sensory structures for prey capture in Northern elephant seals, Mirounga angustirostris? Anat. Rec. 298. 750–760.10.1002/ar.23061Search in Google Scholar PubMed
Miersch, L. et al. (2011). Flow sensing in pinniped whiskers. Phil. Trans. R. Soc. Lond. B 366. 3077–3084.10.1098/rstb.2011.0155Search in Google Scholar PubMed PubMed Central
Mittelstaedt, H. and Mittelstaedt, M.-L. (1982). Homing by path integration. In: Papi, F., Wallraff, H.G. (eds) Avian Navigation – International Symposium on Avian Navigation (ISAN) held at Tirrenia (Pisa), September 11–14, 1981. Springer, Berlin, pp 290–298.10.1007/978-3-642-68616-0_29Search in Google Scholar
Mohl, B. (1967). Seal ears. Science 157. 99.Search in Google Scholar
Mohl, B. (1968). Auditory sensitive of the common seal in air and water. J. Aud. Res. 8. 27–38.Search in Google Scholar
Mouritsen, H. and Ritz, T. (2005). Magnetoreception and its use in bird navigation. Curr. Opin. Neurobiol. 15. 406–414.10.1016/j.conb.2005.06.003Search in Google Scholar PubMed
Muller, B.S. and Bovet, P. (1999). Role of pinnae and head movements in localizing pure tones. Swiss J. Psychol. 58. 170–179.10.1024//1421-0185.58.3.170Search in Google Scholar
Nevitt, G.A. (2008). Sensory ecology on the high seas: the odor world of the procellariiforms seabirds – review. J. Exp. Biol. 211. 1706–1713.10.1242/jeb.015412Search in Google Scholar
Nevitt, G.A. and Bonadonna, F. (2005). Sensitivity to dimethyl sulphide suggests a mechanism for olfactory navigation by sea birds. Biol. Lett. 1. 303–305.10.1098/rsbl.2005.0350Search in Google Scholar
Newman, L.A. and Robinson, P.R. (2005). Cone visual pigments of aquatic mammals. Vis. Neurosci. 22. 873–879.10.1017/S0952523805226159Search in Google Scholar
Newman, L.A. and Robinson, P.R. (2006). The visual pigments of the West Indian manatee (Trichechus manatus). Vis. Res. 46. 3326–3330.10.1016/j.visres.2006.03.010Search in Google Scholar
Niesterok, B., Dehnhard, G. and Hanke, W. (2017a). Hydrodynamic sensory threshold in harbour seals (Phoca vitulina) for artificial flatfish breathing currents. J. Exp. Biol. 220. 2364–2371.10.1242/jeb.158055Search in Google Scholar
Niesterok, B. and Hanke, W. (2013). Hydrodynamic patterns from fast-starts in teleost fish and their possible relevance to predator-prey interactions. J. Comp. Physiol. A 199. 139–149.10.1007/s00359-012-0775-5Search in Google Scholar
Niesterok, B., Krüger, Y., Wieskotten, S., Dehnhard, G. and Hanke, W. (2017b). Hydrodynamic detection and localization of artificial flatfish breathing currents by harbour seals (Phoca vitulina). J. Exp. Biol. 220. 174–185.10.1242/jeb.148676Search in Google Scholar
Norris, K. (1967). Some observations on the migration and orientation of marine mammals. In: Storm, R.M. (ed) Animal orientation and navigation. Oregon State University Press, Corvallis, OR, pp 101–125Search in Google Scholar
Nummela, S. (1995). Scaling of the mammalian middle ear. Hear Res. 85.18–30.10.1016/0378-5955(95)00030-8Search in Google Scholar
Nummela, S. (2008). Hearing in aquatic mammals. In: Thewissen, J.G.M., Nummela, S. (eds) Sensory evolution on the threshold: adaptations in secondarily aquatic vertebrates. University of California Press, Berkeley Los Angeles, London, pp 211–224.10.1525/california/9780520252783.003.0013Search in Google Scholar
Peichl, L., Behrmann, G. and Kröger, R.H.H. (2001). For whales and seals the ocean is not blue: a visual pigment loss in marine mammals. Eur. J. Neurosci. 13. 1520–1528.10.1046/j.0953-816x.2001.01533.xSearch in Google Scholar PubMed
Peichl, L. and Moutairou, K. (1998). Absence of short-wavelength sensitive cones in the retinae of seals (Carnivora) and African giant rats (Rodentia). Eur. J. Neurosci. 10. 2586–2594.10.1046/j.1460-9568.1998.00265.xSearch in Google Scholar
Pfaffmann, C., Bartoshuk, L.M. and McBurney, D. (1971). Taste psychophysics. In: Beidler, L.M. (ed) Handbook of sensory physiology. (Chemical senses, vol 4: Taste). Springer, Berlin Heidelberg New York, pp 73–99.10.1007/978-3-642-65245-5_5Search in Google Scholar
Pierce, G.J., Thompson, P.M., Miller, A., Diack, J.S.W., Miller, D. and Boyle, P.R. (1991). Seasonal variation in the diet of common seals (Phoca vitulina) in the Moray Firth area of Scotland. J. Zool. 223. 641–652.10.1111/j.1469-7998.1991.tb04393.xSearch in Google Scholar
Pihlström, H., Fortelius, M., Hemilä, R., Forsman, R. and Reuter, T. (2005). Scaling of mammalian ethmoid bones can predict olfactory organ size and performance. Proc. R. Soc. B 272. 957–962.10.1098/rspb.2004.2993Search in Google Scholar PubMed PubMed Central
Pitcher, B.J., Harcourt, R.G., Schaal, B. and Charrier, I. (2011). Social olfaction in marine mammals: wild female Australian sea lions can identify their pup’s scent. Biol. Lett. 7: 60–6210.1098/rsbl.2010.0569Search in Google Scholar PubMed PubMed Central
Pütter, A. (1903). Die Augen der Wassersäugethiere. Zoolog. Abt. Anat. Ont. 17. 99–402.Search in Google Scholar
Reep, R.L., Finlay, B.L. and Darlington, R.B. (2006). The limbic system in mammalian brain evolution. Brain Behav. Evol. 70. 57–70.10.1159/000101491Search in Google Scholar PubMed
Reichmuth, C., Holt, M.M., Mulsow, J., Sills, J.M. and Southall, B.L. (2013). Comparative assessment of amphibious hearing in pinnipeds. J. Comp. Physiol. A 199. 491–507.10.1007/s00359-013-0813-ySearch in Google Scholar PubMed
Renouf, D. (1991). Sensory reception and processing in Phocidae and Otariidae. In: Renouf, D. (ed) Behaviour in Pinnipeds. University Press, Cambridge.10.1007/978-94-011-3100-1_7Search in Google Scholar
Repenning, C.A. (1972). Underwater hearing in seals: functional morphology. In: Harrison, R.J. (ed) Functional anatomy of marine mammals. Academic Press, London, pp 307–331.Search in Google Scholar
Rice, F.L., Mance, A. and Munger, B.L. (1986). A comparative light microscopic analysis of the sensory innervation of the mystacial pad. i. Innervation of vibrissal follicle-sinus complexes. J. Comp. Neurol. 252. 154–174.10.1002/cne.902520203Search in Google Scholar PubMed
Scholtyssek, C., Kelber, A. and Dehnhardt, G. (2008). Brightness discrimination in the harbor seal (Phoca vitulina). Vis. Res. 48. 96–103.10.1016/j.visres.2007.10.012Search in Google Scholar PubMed
Scholtyssek, C., Kelber, A. and Dehnhardt, G. (2015). Why do seals have cones? Behavioral evidence for colorblindness in harbor seals. Anim. Cogn. 18. 551–560.10.1007/s10071-014-0823-3Search in Google Scholar PubMed PubMed Central
Schulte-Pelkum, N., Wieskotten, S., Hanke, W., Dehnhardt, G. and Mauck, B. (2007). Tracking of biogenic hydrodynamic trails in harbour seals (Phoca vitulina). J. Exp. Biol. 210. 781–787.10.1242/jeb.02708Search in Google Scholar PubMed
Schusterman, R.J. and Balliet, R.F. (1970). Visual acuity of the harbour seal and the Stellar sea lion under water. Nature 226. 563–564.10.1038/226563a0Search in Google Scholar PubMed
Serres, J.R. and Ruffier, F. (2017). Optic flow-based collision-free strategies: from insects to robots. Arthropod. Struct. Dev. 46. 703–717.10.1016/j.asd.2017.06.003Search in Google Scholar PubMed
Sonntag, C.F. (1923). The comparative anatomy of the tongues of the mammalia. VIII, Carnivora. Proc. Zool. Soc. Lond. 9. 129–153.10.1111/j.1096-3642.1923.tb02177.xSearch in Google Scholar
Southall, B.L., Schusterman, R.J., Kastak, D. and Reichmuth Kastak, C. (2005). Reliability of underwater hearing thresholds in pinnipeds. J. Acoustic Soc. Am. 6. 243–249.10.1121/1.1985956Search in Google Scholar
Spitzka, E.C. (1890). Remarks on the brain of the seals. Am. Nat. 24. 115–122.10.1086/275074Search in Google Scholar
Sticken, J. and Dehnhardt, G. (2000). Salinity discirmination in harbour seals: a sensory basis for spatial orientation in the marine environment? Naturwissenschaften 87. 499–502.10.1007/s001140050767Search in Google Scholar PubMed
Terhune, J.M. (1974). Directional hearing of a harbour seal in air and water. J. Acoustic Soc. Am. 56. 1862–1865.10.1121/1.1903523Search in Google Scholar PubMed
Terhune, J.M. (1988). Detection thresholds of a harbour seal to repeated underwater high-frequency, short-duration sinusoidal pulses. Can. J. Zool. 66. 1578–1582.10.1139/z88-230Search in Google Scholar
Thompson, P.M. and Miller, D. (1990). Summer foraging activity and movements of radio-tagged common seals (Phoca vitulina) in the Moray Firth, Scotland. J. Appl. Ecol. 27. 492–501.10.2307/2404296Search in Google Scholar
Tollit, D.J. and Thompson, P.M. (1996). Seasonal and between-year variations in the diet of harbour seals in the Moray Firth, Scotland. Can. J. Zool. 74. 1110–1121.10.1139/z96-123Search in Google Scholar
Tytell, E.D. and Lauder, G.V. (2008). Hydrodynamics of the escape response in bluegill sunfish, Lepomis macrochirus. J. Exp. Biol. 211. 3359–3369.10.1242/jeb.020917Search in Google Scholar PubMed PubMed Central
Walls, G.L. (1942). The vertebrate eye and its adaptive radiation. Hafner Press, New York10.1097/00005053-194409000-00057Search in Google Scholar
Wartzok, D. and Ketten, D.R. (1999). Marine mammal sensory systems. In: Reynolds III, J.E., Rommel, S. A. (eds) Biology of Marine Mammals. Smithsonian Institution Press, Washington London, pp 117–175.Search in Google Scholar
Watkins, W.A. and Wartzok, D. (1985). Sensory biophysics of marine mammals. Mar. Mam. Sci. 1. 219–260.10.1111/j.1748-7692.1985.tb00011.xSearch in Google Scholar
Weiffen, M., Mauck, B., Dehnhardt, G. and Hanke, F.D. (2014). Sensitivity of a harbor seal (Phoca vitulina) to coherent visual motion in random dot displays. SpringerPlus 3:688.10.1186/2193-1801-3-688Search in Google Scholar PubMed PubMed Central
Weiffen, M., Möller, B., Mauck, B. and Dehnhardt, G. (2006). Effect of water turbidity on the visual acuity of harbor seals (Phoca vitulina). Vis. Res. 46. 1777–1783.10.1016/j.visres.2005.08.015Search in Google Scholar PubMed
Westerberg, H. (1984). The orientation of fish and the vertical stratification at fine- and micro-structure scales. In: Arnold, G.P., Dodson, J.J., Neill, W.H. (eds) Mechanisms of migration in fishes. Plenum, New York, pp 179–203.10.1007/978-1-4613-2763-9_12Search in Google Scholar
Wieskotten, S., Dehnhardt, G., Mauck, B., Miersch, L. and Hanke, W. (2010a). Hydrodynamic determination of the moving direction of an artificial fin by a harbour seal (Phoca vitulina). J. Exp. Biol. 213. 2194–2200.10.1242/jeb.041699Search in Google Scholar PubMed
Wieskotten, S., Dehnhardt, G., Mauck, B., Miersch, L. and Hanke, W. (2010b). The impact of glide phases on the trackability of hydrodynamic trails in harbour seals (Phoca vitulina). J. Exp. Biol. 213. 3734–3740.10.1242/jeb.047134Search in Google Scholar PubMed
Wieskotten, S., Mauck, B., Miersch, L., Dehnhardt, G. and Hanke, W. (2011). Hydrodynamic discrimination of wakes caused by objects of different size or shape in a harbour seal (Phoca vitulina). J. Exp. Biol. 214. 1922–1930.10.1242/jeb.053926Search in Google Scholar PubMed
Wightman, F.L. and Kistler, D.J. (1997). Monaural sound localization revisited. J. Acoustic Soc. Am. 101. 1050–1063.10.1121/1.418029Search in Google Scholar PubMed
Wilson, B., Batty, R.S. and Dill, L.M. (2004). Pacific and Atlantic herring produce burst pulse sounds. Biol. Lett. 271. S95-S97.10.1098/rsbl.2003.0107Search in Google Scholar PubMed PubMed Central
Wiltschko, R. and Wiltschko, W. (2006). Magnetoreception. BioEssays 28. 157–168.10.1002/bies.20363Search in Google Scholar PubMed
Wohlert, D. et al. (2016). A comparative morphometric analysis of three cranial nerves in two phocids: the hooded seal (Cystophora cristata) and the harbor seal (Phoca vitulina). Anat. Rec. 299. 370–378.10.1002/ar.23298Search in Google Scholar PubMed
Wohlgemuth, S., Ronacher, B. and Wehner, R. (2001). Ant odometry in the third dimension. Nature 411. 795–798.10.1038/35081069Search in Google Scholar PubMed
Wolski, L.F., Anderson, R.C., Bowles, A.E. and Yochem, P.K. (2003). Measuring hearing in the harbor seal (Phoca vitulina): comparison of behavioral and auditory brainstem response techniques. J. Acoustic Soc. Am. 113. 629–637.10.1121/1.1527961Search in Google Scholar PubMed
Anmerkung
Die englische Version des Artikels ist online verfügbar unter https://doi.org/10.1515/nf-2018-A012
Anhang
Übersicht über die chemosensorischen Schwellenwerte der Seehunde im Vergleich mit anderen Arten
Art | Schwellenwert für DMS | Quelle |
Seehund | 13–20 pmol/m-3 | Kowalewsky et al. 2006 |
Blausturmvogel (Halobaena caerulea) | <10 pmol/l | Bonadonna et al. 2006 |
Taubensturmvogel (Pachyptila desolata) | ~3–4nM | Nevitt and Bonadonna 2005 |
Mensch | 0,0026–1 µg/l | z. B. Leonardos et al. 1969 Belitz and Grosch 1982 |
Art | Schwellenwert für Meerwasser/NaCl | Quelle |
Seehund | <4 % bei 30 Promille (beste Unterschiedsschwelle) | Sticken and Dehnhardt 2000 |
Kalifornischer Seelöwe (Zalophus californianus) | 3,6 Promille (absolute Schwelle) | Friedl et al. 1990 |
Pazifischer Grosser Tümmler (Tursiops truncatus gilli) | 1,6 Promille (absolute Schwelle) | Friedl et al. 1990 |
Schwarzmeer Grosser Tümmler (Tursiops truncatus ponticus) | 5,4 Promille (absolute Schwelle) | Kuznetsov 1978 (zitiert nach Friedl et a. 1990) |
Mensch | 18 % bei 20 Promille (beste Unterschiedsschwelle | Sticken and Dehnhardt 2000 |
0,18 Promille (absolute Schwelle) | Pfaffmann et al. 1971 |
© 2018 Walter de Gruyter GmbH, Berlin/Boston
Articles in the same Issue
- Titelseiten
- Übersichtsartikel
- Vom lateralen Rand ins Zentrum des Kortex: Die Entwicklung der menschlichen Inselrinde
- From the lateral edge to the center of the cortex: The development of the human insula
- Neurotransmittergesteuerte Rezeptoren in neuem Licht: Optische Methoden zur Kontrolle physiologischer Funktion
- New light on neurotransmitter-gated receptors: Optical approaches for controlling physiological function
- Schauen Sie, was Sie hören – Wie das Gehirn mit allen seinen Sinnen die Welt wahrnimmt
- See what you hear – How the brain forms representations across the senses
- Unterwegs mit Seehunden – Wie deren Sinnessysteme zu Orientierung, Navigation und Futtersuche beitragen
- On route with harbor seals – how their senses contribute to orientation, navigation and foraging
- Die Blut-Hirn-Schranke und ihre Rolle in der Alzheimer – Krankheit
- The Blood brain-barrier and its role in Alzheimer’s disease
- Nachrichten
- Nachrichten
- Corrigendum
- Corrigendum zu: Langzeitpotenzierung im Hippokampus: Entdeckung, Mechanismen und Funktion
- Corrigendum to: Long-term potentiation in the hippocampus: discovery, mechanisms and function
Articles in the same Issue
- Titelseiten
- Übersichtsartikel
- Vom lateralen Rand ins Zentrum des Kortex: Die Entwicklung der menschlichen Inselrinde
- From the lateral edge to the center of the cortex: The development of the human insula
- Neurotransmittergesteuerte Rezeptoren in neuem Licht: Optische Methoden zur Kontrolle physiologischer Funktion
- New light on neurotransmitter-gated receptors: Optical approaches for controlling physiological function
- Schauen Sie, was Sie hören – Wie das Gehirn mit allen seinen Sinnen die Welt wahrnimmt
- See what you hear – How the brain forms representations across the senses
- Unterwegs mit Seehunden – Wie deren Sinnessysteme zu Orientierung, Navigation und Futtersuche beitragen
- On route with harbor seals – how their senses contribute to orientation, navigation and foraging
- Die Blut-Hirn-Schranke und ihre Rolle in der Alzheimer – Krankheit
- The Blood brain-barrier and its role in Alzheimer’s disease
- Nachrichten
- Nachrichten
- Corrigendum
- Corrigendum zu: Langzeitpotenzierung im Hippokampus: Entdeckung, Mechanismen und Funktion
- Corrigendum to: Long-term potentiation in the hippocampus: discovery, mechanisms and function